the arrangement and function of octopus arm musculature and connective...
TRANSCRIPT

The Arrangement and Function of Octopus ArmMusculature and Connective Tissue
William M. Kier* and Michael P. Stella
Department of Biology, University of North Carolina, Chapel Hill, North Carolina 27599-3280
ABSTRACT The morphology of the musculature andconnective tissues of the arms of Octopus bimaculoideswas analyzed with light microscopy. We also studied O.briareus and O. digueti, which possess relatively moreelongate and less elongate arms, respectively. The mor-phology of the arms was found to be remarkably uniformamong species. The arms consist of a densely packedthree-dimensional arrangement of muscle fibers andconnective tissue fibers surrounding a central axialnerve cord. Three primary muscle fiber orientationswere observed: 1) transverse muscle fibers oriented inplanes perpendicular to the long axis of the arm; 2) lon-gitudinal muscle fibers oriented parallel to the long axis;and 3) oblique muscle fibers arranged in helixes aroundthe arm. The proportion of the arm cross section occu-pied by each of these muscle fiber groups (relative to thetotal cross sectional area of the musculature) remainsconstant along the length of the arm, even though thearm tapers from base to tip. A thin circular muscle layerwraps the arm musculature on the aboral side only.Much of this musculature has its origin and insertion onseveral robust connective tissue sheets including a layersurrounding the axial nerve cord and crossed-fiber con-nective tissue sheets located on the oral and the aboralsides of the arm. An additional thin layer of connectivetissue wraps the arm musculature laterally and alsoserves as a site of origin and insertion of some of themuscle fibers. The fibers of the oral and aboral crossed-fiber connective tissue sheets are arranged oblique tothe long axis of the arm with the same fiber angle as theoblique muscle layers that originate and insert on thesheets. The oblique muscle layers and the crossed-fiberconnective tissue sheets thus form composite right- andleft-handed helical fiber arrays. Analysis of arm mor-phology from the standpoint of biomechanics suggeststhat the transverse musculature is responsible for elon-gation of the arms, the longitudinal musculature is re-sponsible for shortening, and the oblique muscle layersand associated connective tissues create torsion. Armbending may involve unilateral contraction of longitudi-nal muscle bundles in combination with resistance toarm diameter increase due to contraction of the trans-verse musculature or passive stiffness of the arm tis-sues. The arms may also be bent by a combination ofdecrease in diameter due to contraction of the trans-verse musculature and maintenance of constant lengthon one side of the arm by unilateral activity of longitudi-nal muscle bundles. An increase in flexural stiffness ofthe arm may be achieved by cocontraction of the trans-verse and longitudinal muscle. Torsional stiffness maybe increased by simultaneous contraction of both theright- and left-handed oblique muscle layers. J. Morphol.268:831–843, 2007. � 2007 Wiley-Liss, Inc.
KEY WORDS: biomechanics; muscular-hydrostat; muscle;octopus; skeletal support
The eight arms of octopuses are capable of a re-markable diversity and complexity of movementand they serve a wide range of functions. Thearms are used in prey capture, locomotion, manip-ulation, grooming, burying, copulation, defense,chemosensing, and tactile sensing. The adaptabil-ity of the arms to such a wide range of tasks hasattracted considerable attention recently fromrobotics engineers who are using them as a sourceof inspiration for the design and construction of anew class of robotic arms (McMahan et al., 2005,2006; Walker et al., 2005; Jones and Walker,2006a,b). Such robotic arms have significant poten-tial advantages over more conventional ‘‘vertebrate-like’’ robotic arms. For instance, they more readilyadapt to unstructured and cluttered environmentsand they do not require a specific ‘‘end effector’’ foreach type of manipulated object; instead, theentire arm itself can be used for object mani-pulation.
Although the general arrangement of the muscu-lature of octopus arms has been known for sometime (Cuvier, 1817; Colasanti, 1876; Guerin, 1908;Tittel, 1961, 1964; Socastro, 1969; Graziadei, 1971;Browning, 1980; Kier and Smith, 1985; Kier, 1988;Kier and Thompson, 2003), a detailed descriptionof arm morphology is lacking, in particular withrespect to muscle fiber trajectories and thearrangement of connective tissues. The goal of thispaper, therefore, is to provide a precise descriptionof the three-dimensional arrangement and form ofthe musculature and the connective tissues of thearms. This morphology is of particular interestbecause the arms are examples of a group of animalstructures, termed ‘‘muscular-hydrostats,’’ which
Contract grant sponsor: DARPA; Grant number: N66001-03-R-8043.
*Correspondence to: William M. Kier, Department of Biology, CBNo. 3280 Coker Hall, University of North Carolina, Chapel Hill, NC27599-3280. E-mail: [email protected]
Published online 11 July 2007 inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/jmor.10548
JOURNAL OF MORPHOLOGY 268:831–843 (2007)
� 2007 WILEY-LISS, INC.

lack both the fluid-filled cavities that characterizethe hydrostatic skeleton of many invertebrates andthe rigid skeletal elements present in other animals(Kier and Smith, 1985; Smith and Kier, 1989). Thethree-dimensional arrangement of the musculatureof these structures serves both to generate the forcefor movement and to provide skeletal support. Wetherefore analyze the biomechanics of the arms topredict the functional role of the various musclesand connective tissues in support and movement. Toexplore potential variation in arm structure andfunction, we sampled several octopus species repre-senting a range from relatively short to relativelyelongate arms. In addition to the inherent biologicalinterest in these structures, we hope that theinsights gained from the analysis of their structureand function will be helpful in the design and con-struction of robotic arms with greater diversity andcomplexity of movement.
MATERIALS AND METHODSExperimental Animals
The focus of our investigation was the Californian Two-SpotOctopus, Octopus bimaculoides Pickford and McConnaughey,1949. Specimens were obtained from the National ResourceCenter for Cephalopods (Galveston, TX) or from AquaticResearch Consultants (San Pedro, CA) and maintained untiluse in a recirculating artificial seawater system housed in theDepartment of Biology, University of North Carolina, ChapelHill, NC. In addition to the detailed morphological analysis ofarms of this species, arms of the Caribbean Reef Octopus,O. briareus Robson, 1929, and Diguet’s Pygmy Octopus,O. digueti Perrier and Rochebrune, 1894, were also analyzed.Specimens of the latter two species were provided by R. Hanlon,Marine Biological Laboratory, Woods Hole, MA. They wereincluded in this study because they represent species that pos-sess relatively longer (O. briareus) and relatively shorter(O. digueti) arms, compared with O. bimaculoides and withother species in the Octopodidae. In addition to the speciesdescribed above, previously prepared but undescribed histologi-cal material of the arms of the Caribbean Pygmy OctopusO. joubini Robson and the Mediterranean Musky Octopus Ele-done moschata (Lamarck, 1798) was also examined.
Histology
Portions of the arms were removed from specimens that hadbeen anesthetized using 2.5% ethanol in seawater (O’Dor et al.,1990) or a 1:1 mixture of 7.5% MgCl2�6H2O and seawater (Mes-senger et al., 1985) and fixed for 24–48 h in 10% formalin inseawater. Smaller blocks (3–4 mm thickness) were cut from thefixed arm tissue, dehydrated in ethanol, cleared in Citri-Solv(Fisher Scientific, Fair Lawn, NJ), and embedded in ParaplastPlus (MP 568C) (Tyco Healthcare/Kendall, Mansfield, MA). Theblocks were sectioned on a rotary microtome. Serial sections wereobtained in three mutually perpendicular planes. The sectionswere stained with Picro-Ponceau with Weigert iron hematoxylinor with Milligan’s Trichrome (Kier, 1992) and examined withbrightfield and polarized light microscopy.
Morphometrics
For measurements of fiber angles, digital micrographs weretaken of grazing sections of the connective tissue sheets or musclelayers. The angles formed with the longitudinal axis of the arm
by connective tissue fibers or muscle fibers were measured usingmorphometrics software (SigmaScan Pro; Systat Software, PointRichmond, CA). The software was also used to measure the areasof components of the intrinsic muscle mass along the length ofthe arm. For these measurements, an entire arm was fixed andcross-sectional slices of the arm were removed from five equallyspaced intervals covering the entire length of the arm. These tis-sue slices were then processed as described earlier and transversesections of the arm were obtained from each sample. The totalarea of the transverse, longitudinal, and oblique musculature,and the axial nerve cord was measured from each of these trans-verse sections. The relative area of each of these components inthe cross section was calculated by dividing the measured area bythe total area of the intrinsic arm musculature.
RESULTSArm Gross Morphology
The following definitions of section planes andarm orientation will be used in the morphologicaldescriptions below. Given the significant rear-rangement of the arm crown in late embryonicdevelopment (Budelmann et al., 1977) these defini-tions are not necessarily related to the morpho-logical orientation (in general understood as equiv-alent to ‘‘embryological orientation’’) of the cepha-lopod body. The oral surface of the octopus arm isdefined as the surface bearing suckers. If the eightarms of an octopus are brought together (as hap-pens for instance when the animal uses the mantleto swim backwards by jet locomotion) the oral sur-faces of the arms face one another surrounding themouth. Transverse section planes are defined hereas planes perpendicular to the longitudinal axis ofthe arm. Frontal section planes are defined hereas planes parallel to the longitudinal axis and tothe oral surface. The sagittal plane is defined hereto be at the center of the arm in a plane parallelto the longitudinal axis and perpendicular to thefrontal plane.
In the description of arm morphology that fol-lows, sections from the arms of Octopus bimacu-loides are included in all figures. In spite of thedifferences in the relative length of the arms ofthe various octopus species analyzed in this study,the morphology we observed was remarkably uni-form. Thus, although the figures are of only one ofthe species, the description that follows applies toall species studied. Our decision to use materialfrom O. bimaculoides for the illustrations wasbased simply on the quantity and the quality ofthe preparations of this species. This similarity inmorphology was also apparent in the arms of asingle individual, in spite of differences in the rela-tive lengths of the four pairs of arms. It is of inter-est that in a recent parallel study, Hanlon et al.(personal communication) have not observed anydifference in the behavior of the arms among thesespecies even though the differences in arm propor-tions are dramatic.
The musculature of the arm was classified byGraziadei (1965, 1971) into 1) the intrinsic muscu-
832 W.M. KIER AND M.P. STELLA
Journal of Morphology DOI 10.1002/jmor

lature of the suckers, 2) the intrinsic musculatureof the arms, and 3) the acetabulo-brachial muscu-lature that connects the suckers to the arm muscu-lature (Fig. 1A). The intrinsic musculature of thesuckers and the acetabulo-brachial musculaturehas been described in detail elsewhere (Guerin,1908; Tittel, 1964; Kier and Smith, 1990, 2002)and the focus of the current study is therefore onthe intrinsic musculature of the arms.
The arm intrinsic musculature consists of adensely packed, three-dimensional array of musclefibers surrounding a large central axial nerve cordthat extends the length of the arm (Figs. 1B and2). The axial nerve cord includes nerve cell bodieslocated along the lateral sides and oral side,numerous axons of a range of sizes in a tract onthe dorsal surface of the cord, and a central core ofneuropile (see Guerin, 1908; Rossi and Graziadei,1954; Graziadei, 1971; Matzner et al., 2000;Sumbre et al., 2001 for detailed descriptions ofarm neuroanatomy). A thick-walled artery extendslongitudinally down the aboral surface of the axialnerve cord. Four intramuscular nerve cords(Guerin, 1908; Rossi and Graziadei, 1956) extendthe length of the arm at locations towards theperiphery of the dense core of musculature (Fig. 2).
The arm is covered by a simple epithelium andan underlying thick, loose fibrous dermal connec-tive tissue layer that includes blood vessels, scat-tered muscle bundles, chromatophores, leuco-phores, and reflective cells (see Cloney and Brocco,1983; Budelmann et al., 1997).
Connective Tissues of the Arm
A thick connective tissue layer surrounds theaxial nerve cord of the arm and is formed of fibersthat show staining reactions typical of collagen(Fig. 2). When viewed in grazing frontal or para-sagittal sections, the birefringent fibers appear tobe arranged as a feltwork and polarized lightmicroscopy suggests a slight degree of preferredorientation in the circular and longitudinal direc-tions. Many of the muscle fibers of the transversemuscle mass (see later) insert on this layer.
Additional thick crossed-fiber connective tissuesheets are present orally and aborally in the armand serve as the site of origin and insertion formuch of the intrinsic musculature of the arm(Figs. 1B and 2). The fibers of these layers are alsohighly birefringent and exhibit staining reactionstypical of collagen. Frontal sections show the fibersto be in two sets of highly organized parallelarrays, one arranged as a right-hand helix and theother arranged as a left-hand helix (Fig. 3). Thefiber angle (angle that the fibers make with the lon-gitudinal axis) was measured to be 688–758.Although the animals were anesthetized when thearms were removed, the state of elongation or short-ening of the arm is difficult to assess. Some of the
variation in fiber angle reported here may reflectdifferences in the state of elongation or shorteningof the arms since the fiber angle is increased byshortening and decreased by elongation.
A thin layer of connective tissue wraps theintrinsic musculature laterally on each side of thearm and also serves as the site of origin and inser-tion of some of the intrinsic muscle fibers, in par-ticular, transverse muscle fibers extending acrossthe arm.
Intrinsic Musculature of the ArmTransverse muscle mass. The transverse mus-
cle mass is located at the core of the arm, sur-rounding the axial nerve cord (Figs. 1B and 2).The transverse muscle fibers are oriented inplanes perpendicular to the longitudinal axis ofthe arm. They are oriented in an approximatelyorthogonal array, i.e., the fibers of the mass areeither oriented approximately parallel to the sagit-tal plane (and thus extending from oral to aboral)or approximately parallel to the frontal plane (andthus extending from side to side).
The transverse muscle fibers extending approxi-mately parallel to the sagittal plane originate onthe thick crossed-fiber connective tissue sheetslocated on the oral and the aboral side of the armand extend towards the central axis of the arm inlongitudinal sheets between bundles of longitudi-nal muscle (Fig. 4). These longitudinal sheets aretermed ‘‘trabeculae’’ (Graziadei, 1965), but in thislocation they are in the form of layers or plates ofmuscle fibers as opposed to isolated bundles. Manyof the transverse muscle fibers insert on the con-nective tissue layer surrounding the axial nervecord. Some of the transverse muscle fibers pass tothe side of the axial nerve cord and either inserton the side of the connective tissue layer surround-ing the axial nerve cord or extend to insert on thecrossed-fiber connective tissue sheet on the oppo-site side.
Transverse muscle fibers extending from side toside parallel to the frontal plane originate on thethin layer of connective tissue surrounding theexternal oblique muscle layers. They extendthrough the oblique muscle layers and longitudinalmusculature laterally and some insert on the con-nective tissue layer surrounding the axial nervecord. Others pass oral and, especially, aboral tothe axial nerve cord and extend to insert on theconnective tissue surrounding the external obliquemuscle layer on the opposite side of the arm (Fig.5). They form trabeculae in the form of parallelsheets of muscle fibers as they extend between thefibers of the longitudinal muscle bundles, and bun-dles of fibers as they extend between the fibers ofthe oblique muscle layers (Fig. 6).
Longitudinal musculature. Longitudinal mus-cle fibers surround the central core of transverse
OCTOPUS ARM STRUCTURE AND FUNCTION 833
Journal of Morphology DOI 10.1002/jmor

musculature and are oriented parallel to the longi-tudinal axis of the arm. The fibers extend longitudi-nally as bundles between the trabeculae of thetransverse muscle mass. Longitudinal muscle fibersare distributed orally, aborally, and laterally relativeto the transverse muscle mass so that essentially
the entire outer perimeter of the arm includes longi-tudinal muscle fibers, although the cross-sectionalarea of longitudinal muscle fibers is larger in theaboral quadrant of the arm, compared with the lat-eral and oral quadrants. A layer of longitudinalfibers is situated between the median and external
Fig. 1. A: Transverse section of the arm of Octopus bimaculoides showing the three major subdivisions of the arm musculature:the intrinsic arm musculature (IA); the intrinsic sucker musculature (IS); and the acetabulo-brachial musculature (AB) connectingthe arm and sucker musculature. Scale bar: 0.5 mm. Brightfield microscopy of 10-lm-thick paraffin section stained with Picro-Ponceau and hematoxylin. B: Diagram of the arm of Octopus showing three-dimensional arrangement of muscle fibers and connec-tive tissue fibers. AN, axial nerve cord; AR, artery; CM, circumferential muscle layer; CT, connective tissue; DCT, dermal connec-tive tissue; EP, epidermis; IN, intramuscular nerve; LM, longitudinal muscle fibers; OME; external oblique muscle layer; OMI,internal oblique muscle layer; OMM, median oblique muscle layer; SU, sucker; TM, transverse muscle fibers; TR, trabeculae; V,vein. [Reproduced with permission from Kier, The Mollusca, Form and Function (Trueman ER, Clarke MR, editors), 1988, Vol. 11,pp 211–252, �Academic Press].
834 W.M. KIER AND M.P. STELLA
Journal of Morphology DOI 10.1002/jmor

oblique muscles and has a crescentic shape in crosssection (Figs. 1B, 2, and 5). It, too, is further subdi-vided by trabeculae of transverse muscle fibers.
Oblique musculature. In addition to the longi-tudinal and transverse musculature, three sets ofobliquely oriented muscle layers are present in theoctopus arm (Figs. 1B, 2, and 5). The names of theoblique muscles used here follow Graziadei (1965).A pair of external oblique muscle layers is the mostsuperficial, occupying a lateral position and enclos-ing the remainder of the intrinsic musculature. Apair of median oblique muscle layers is locatedcloser to the central axis of the arm and is sepa-rated from the external oblique muscle pair by thecrescent-shaped longitudinal muscle massesdescribed earlier. The internal oblique musclelayers are the most central and are located on eachside of the central core of transverse musculature.
The handedness of a given oblique muscle layer(external, median, or internal) is opposite to thehandedness of the other member of the pair on theopposite side of the arm. For instance, if the exter-nal oblique muscle layer on one side of the arm iswrapped as a right-hand helix, the externaloblique muscle layer on the opposite side is left-handed. In addition, on one side of the arm, thehandedness of the external and internal oblique
muscle layers are the same but that of the medianoblique muscle layer is opposite (Fig. 7). The exter-nal and median oblique muscle layers have theirorigin and insertion on the oral and aboralcrossed-fiber connective tissue sheets and thus arerestricted to the sides of the arm (Figs. 1B and 2).The fiber angles of the median and externaloblique muscle layers are similar to those of theoral and aboral crossed-fiber connective sheets towhich they attach [538–728 for external obliquemuscle (mean ¼ 638, SD ¼ 58); 528–698 for medianoblique muscle (mean ¼ 618, SD ¼ 58); 688–758 forconnective tissue sheets (mean ¼ 748, SD ¼ 38)].The fibers of the internal oblique muscle layersappear to interdigitate with the transverse andlongitudinal musculature and do not show as dis-tinct an origin and insertion as is observed for theexternal and median oblique muscle layers. Thefiber angle of the internal oblique muscle layersappears to be lower than that of the other obliquemuscle layers and fibrous connective tissue layersand showed more variation between species [Octo-pus briareus: 398–458 (mean ¼ 428, SD ¼ 28); O.digueti, 478–658 (mean ¼ 568, SD ¼ 68)].
Circular muscle layer. A thin layer of musclefibers arranged circumferentially is also present.This layer is thickest on the aboral side and covers
Fig. 2. Transverse section of the armof Octopus bimaculoides showing themajor components of the intrinsic armmusculature. Connective tissues arestained red and muscle tissue is stainedbrown in this preparation. AN, axialnerve cord; ACT, aboral crossed-fiberconnective tissue sheet; AR, artery; CT,connective tissue; IN, intramuscularnerve; LM, longitudinal muscle fibers;OCT, oral crossed-fiber connective tissuesheet; OME, external oblique musclelayer; OMI, internal oblique musclelayer; OMM, median oblique musclelayer; TM, transverse muscle fibers; TR,trabeculae. Scale bar: 200 lm. Bright-field microscopy of 10-lm-thick paraffinsection (same preparation as Fig. 1B)stained with Picro-Ponceau and hema-toxylin.
OCTOPUS ARM STRUCTURE AND FUNCTION 835
Journal of Morphology DOI 10.1002/jmor

the aboral crossed-fiber connective tissue layer(Fig. 4). It extends down each side of the arm as athin layer (Fig. 5), wrapping the external obliquemuscle layers, and then inserts on the oralcrossed-fiber connective tissue layer. The interfacebetween the circular muscle layer and the externaloblique muscle layer is difficult to resolve in trans-verse sections because there is relatively little con-nective tissue separating the two components. Ingrazing parasagittal sections, however, the differ-ence in muscle fiber orientation makes the inter-face between the two layers quite obvious (Fig. 7).
Arm Taper and Morphology
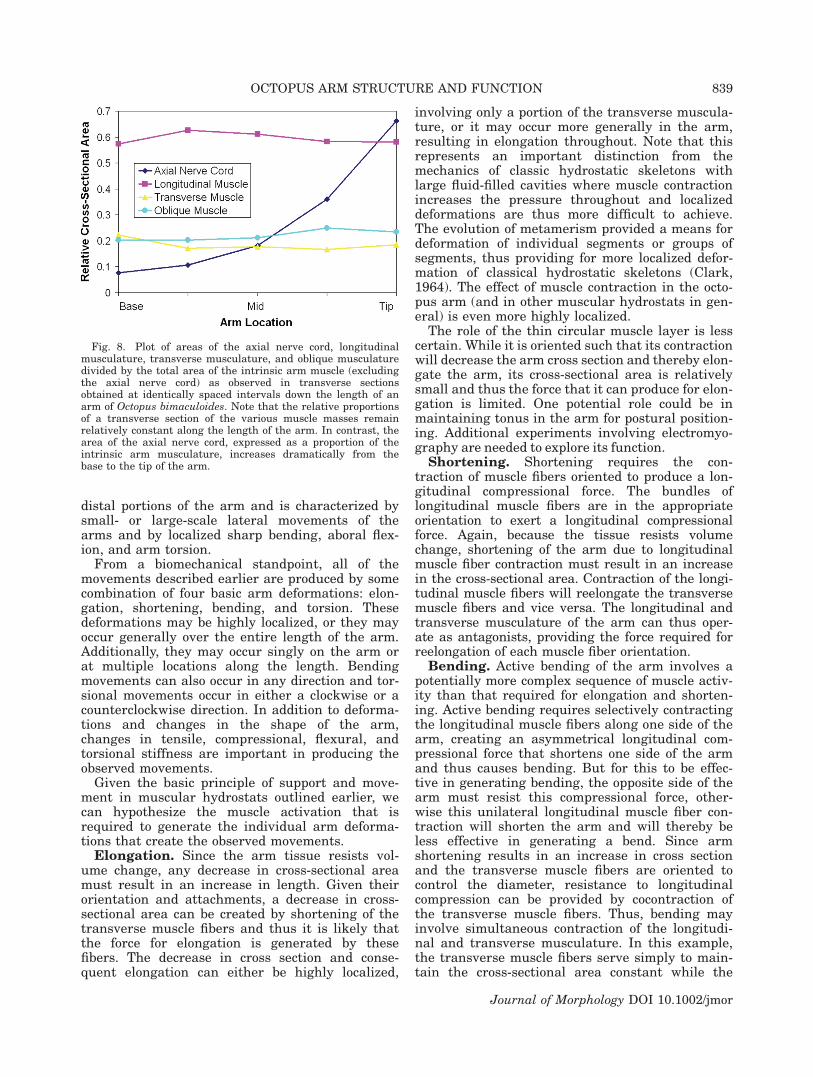
By sampling a series of locations along the arm ofOctopus bimaculoides, we examined potential differ-ences in the relative proportions of the variousmusclemasses as the arm tapers. Although the total cross-sectional area of the intrinsic arm musculature andaxial nerve cord of the sample from the base of the
arm was 144 times the total cross-sectional area ofthe sample from the tip, the area that the longitudi-nal, transverse, and oblique musculature occupies inthe cross section (expressed relative to the total cross-sectional area of the intrinsic musculature at a givenlocation) was remarkably similar (Fig. 8). The longi-tudinal musculature occupies �0.56 of the total areaof the intrinsicmusculature in a cross section, and theoblique and transverse musculature occupy �0.21and 0.17, respectively (mean of measurements takenat each location along the length). Note that theseproportions are expressed relative to the total cross-sectional area of the intrinsic musculature and thusexclude the area occupied by the axial nerve cord.
Although the relative proportions of the variousmuscle masses were similar along the length, therelative proportion of the cross section occupied bythe axial nerve cord (expressed relative to the totalcross-sectional area of the intrinsic muscle) wasalmost an order of magnitude greater at the tipcompared with the base. For instance, the axialnerve cord represented an area of 0.59 at the tip ofthe arm and only 0.07 at the base.
DISCUSSIONPrinciples of Support and MuscularAntagonism
Cephalopod arms and tentacles, many mamma-lian and lizard tongues, and the trunk of the ele-phant are members of a unique class of soft tissuestructures, termed ‘‘muscular hydrostats’’ (Kierand Smith, 1985; Smith and Kier, 1989). Thesestructures consist primarily of a dense, three-dimensional arrangement of muscle fibers andthus lack the characteristics of other forms of skel-etal support observed in animals. Specifically, theylack the rigid internal or external skeletal elementsseen in arthropods, echinoderms, and vertebrates, aswell as the fluid-filled cavities enclosed by a con-tainer reinforced with connective tissue (typicallythe body wall) that characterize the hydrostatic skel-eton of the diverse vermiform animals (Chapman,1950, 1958, 1975; Clark, 1964, 1981; Gutman, 1981;Wainwright, 1970, 1982).
The basic principle of support and movement inmuscular hydrostats is straightforward. Muscle tis-sue, like most tissues lacking gas-filled spaces, ischaracterized by a high bulk modulus and thus itresists volume change. Because the volume of thestructure is approximately constant, any decreasein a given dimension must result in an increase inanother dimension. In muscular hydrostats, themuscle fibers are typically arranged in multiple ori-entations so that all three dimensions can beactively controlled. Thus, by selectively activatingspecific muscle fiber orientations, these structurescan create a wide diversity and complexity of move-ments. This fundamental principle serves as thefoundation for the mechanical analysis and the pre-
Fig. 3. A grazing frontal section of the oral crossed-fiber con-nective tissue sheet of the arm of Octopus bimaculoides. Thefibers of this sheet are arranged as a highly ordered fiber array.The white lines are drawn parallel with the right- and left-handed fibers of the sheet. Longitudinal muscle fibers can beseen beneath the crossed-fiber connective tissue sheet. Scalebar: 100 lm. Polarized light microscopy with first order redfilter of 10-lm-thick paraffin section stained with Milligan’strichrome.
836 W.M. KIER AND M.P. STELLA
Journal of Morphology DOI 10.1002/jmor

Fig. 4. Transverse section of the arm of Octopus bimaculoides aboral to the axial nerve cord showing the details of the muscula-ture and the connective tissue arrangement. Connective tissues are stained red and muscle tissue is stained brown in this prepara-tion. Fibers of the transverse musculature originate on the aboral connective tissue sheet and extend through the longitudinal muscu-lature, dividing it into elongate bundles. Many of the transverse muscle fibers are visible as they extend through the transverse mus-cle mass to insert on the connective tissue surrounding the axial nerve cord. ACT, aboral crossed-fiber connective tissue sheet; AR,artery; CM, circular muscle layer; CT, connective tissue; IN, intramuscular nerve cord; LM, longitudinal muscle fibers; OME, externaloblique muscle layer; OMM, median oblique muscle layer; TM, transverse muscle fibers: TR, trabeculae. Scale bar: 100 lm. Bright-field microscopy of 10-lm-thick paraffin section (same preparation as Figs. 1B and 2) stained with Picro-Ponceau and hematoxylin.
Fig. 5. Transverse section of the armof Octopus bimaculoides lateral to theaxial nerve cord showing the details ofthe musculature and connective tissuearrangement. Connective tissues arestained red and muscle tissue is stainedbrown in this preparation. The fibers ofthe transverse muscle mass are visibleas they extend to the periphery of thearm, penetrating the surroundingoblique and longitudinal musculature.AN, axial nerve cord; AR, artery; CM,circular muscle layer; CT, connective tis-sue; IN, intramuscular nerve cord; LM,longitudinal muscle fibers; OME, exter-nal oblique muscle layer; OMI, internaloblique muscle layer; OMM, medianoblique muscle layer; TM, transversemuscle fibers; TR, trabeculae. Scale bar:100 lm. Brightfield microscopy of 10-lm-thick paraffin section (same preparationas Figs. 1B, 2, and 4) stained with Picro-Ponceau and hematoxylin.
Journal of Morphology DOI 10.1002/jmor

diction of the functional role of the various musclegroups below.
Summary of Octopus Arm Movements andBiomechanics
As part of an ongoing collaborative project todesign and build robotic arms inspired by octopusarms (Walker et al., 2005), Hanlon et al. (personalcommunication) are analyzing the behaviors of anumber of octopus species with the goal of catego-rizing the movements of the arms. They haveobserved that the majority of arm movementsappear to serve to place the suckers on objects inthe environment. They have identified three gen-eral categories of arm movement. The first cate-gory, termed ‘‘Reach,’’ is defined as an increase indistance between the proximal and distal portionsof the arm. Reaches can be executed in severalways. One common form has been studied in con-siderable detail (Gutfreund et al., 1996, 1998;Yekutieli et al., 2005a,b) and involves a propagat-ing wave of stiffening and straightening of thearm from the base to the tip, forming a passivebend at the transition between the stiffened baseand relaxed terminal portion of the arm. Alsoobserved are a progressive uncurling and straight-ening of a spirally curled arm and elongation ofthe entire arm in a straight configuration. Thesecond category, termed ‘‘Pull,’’ is defined as adecrease in the distance between the proximal and
distal portions of the arm. Pulls can be createdusing many forms of movement including curlingof the arm from tip to base, overall shortening ofthe arm in a straight configuration, and localizedbending of the arm at ‘‘pseudo joints,’’ termed a‘‘fetching’’ movement, so that the arm does notchange length and behaves as if it were articulated(Sumbre et al., 2005, 2006). The third category,termed ‘‘Search/Grope/Explore’’ involves no signifi-cant change in distance between the proximal and
Fig. 6. Grazing parasagittal section of the arm of Octopusbimaculoides showing arrangement of transverse muscle fibersas they penetrate the oblique and longitudinal musculature.The section is slightly oblique to the long axis of the arm andthe right side of the section cuts deeper into the arm muscula-ture. The transverse muscle fibers are grouped as a plate orlayer as they extend between the longitudinal fibers and arearranged as a series of bundles as they pass through theoblique muscle layers. LM, longitudinal muscle fibers; OMM,median oblique muscle layer; TM, transverse muscle fibers.Scale bar: 50 lm. Brightfield microscopy of 10-lm-thick paraffinsection stained with Milligan’s Trichrome.
Fig. 7. Grazing parasagittal sections of the arm of Octopusbimaculoides showing the arrangement of the oblique musclelayers on one side of the arm. The section shown in (B) isdeeper in the arm musculature than the section shown in (A)and both are from the same side of the arm. A: The thin circu-lar muscle layer surrounding the external oblique muscle layeris apparent because of the difference in fiber orientationbetween the two layers. Note that the handedness of the exter-nal oblique muscle layer is opposite to that of the medianoblique muscle layer, which the section grazes in the center ofthe micrograph. The lines are drawn parallel to the fibers of theexternal oblique and internal oblique muscle layers. B: The sec-tion is deeper on the right side of the micrograph and grazesthe transverse muscle mass, which is adjacent to the internaloblique muscle layer visible on the left side of the micrograph.The line is drawn parallel to the muscle fibers of the internaloblique muscle layer. Note that the handedness of the internaloblique muscle layer is the same as that of the external obliquemuscle layer on the same side of the arm shown in (A). CM, cir-cular muscle layer; LM, longitudinal muscle layer; OME, exter-nal oblique muscle layer; OMI, internal oblique muscle layer;OMM, median oblique muscle layer; TM, transverse musclefibers. Scale bar: 100 lm. Brightfield microscopy of 10-lm-thickparaffin section stained with Milligan’s Trichrome.
838 W.M. KIER AND M.P. STELLA
Journal of Morphology DOI 10.1002/jmor
distal portions of the arm and is characterized bysmall- or large-scale lateral movements of thearms and by localized sharp bending, aboral flex-ion, and arm torsion.
From a biomechanical standpoint, all of themovements described earlier are produced by somecombination of four basic arm deformations: elon-gation, shortening, bending, and torsion. Thesedeformations may be highly localized, or they mayoccur generally over the entire length of the arm.Additionally, they may occur singly on the arm orat multiple locations along the length. Bendingmovements can also occur in any direction and tor-sional movements occur in either a clockwise or acounterclockwise direction. In addition to deforma-tions and changes in the shape of the arm,changes in tensile, compressional, flexural, andtorsional stiffness are important in producing theobserved movements.
Given the basic principle of support and move-ment in muscular hydrostats outlined earlier, wecan hypothesize the muscle activation that isrequired to generate the individual arm deforma-tions that create the observed movements.
Elongation. Since the arm tissue resists vol-ume change, any decrease in cross-sectional areamust result in an increase in length. Given theirorientation and attachments, a decrease in cross-sectional area can be created by shortening of thetransverse muscle fibers and thus it is likely thatthe force for elongation is generated by thesefibers. The decrease in cross section and conse-quent elongation can either be highly localized,
involving only a portion of the transverse muscula-ture, or it may occur more generally in the arm,resulting in elongation throughout. Note that thisrepresents an important distinction from themechanics of classic hydrostatic skeletons withlarge fluid-filled cavities where muscle contractionincreases the pressure throughout and localizeddeformations are thus more difficult to achieve.The evolution of metamerism provided a means fordeformation of individual segments or groups ofsegments, thus providing for more localized defor-mation of classical hydrostatic skeletons (Clark,1964). The effect of muscle contraction in the octo-pus arm (and in other muscular hydrostats in gen-eral) is even more highly localized.
The role of the thin circular muscle layer is lesscertain. While it is oriented such that its contractionwill decrease the arm cross section and thereby elon-gate the arm, its cross-sectional area is relativelysmall and thus the force that it can produce for elon-gation is limited. One potential role could be inmaintaining tonus in the arm for postural position-ing. Additional experiments involving electromyo-graphy are needed to explore its function.
Shortening. Shortening requires the con-traction of muscle fibers oriented to produce a lon-gitudinal compressional force. The bundles oflongitudinal muscle fibers are in the appropriateorientation to exert a longitudinal compressionalforce. Again, because the tissue resists volumechange, shortening of the arm due to longitudinalmuscle fiber contraction must result in an increasein the cross-sectional area. Contraction of the longi-tudinal muscle fibers will reelongate the transversemuscle fibers and vice versa. The longitudinal andtransverse musculature of the arm can thus oper-ate as antagonists, providing the force required forreelongation of each muscle fiber orientation.
Bending. Active bending of the arm involves apotentially more complex sequence of muscle activ-ity than that required for elongation and shorten-ing. Active bending requires selectively contractingthe longitudinal muscle fibers along one side of thearm, creating an asymmetrical longitudinal com-pressional force that shortens one side of the armand thus causes bending. But for this to be effec-tive in generating bending, the opposite side of thearm must resist this compressional force, other-wise this unilateral longitudinal muscle fiber con-traction will shorten the arm and will thereby beless effective in generating a bend. Since armshortening results in an increase in cross sectionand the transverse muscle fibers are oriented tocontrol the diameter, resistance to longitudinalcompression can be provided by cocontraction ofthe transverse muscle fibers. Thus, bending mayinvolve simultaneous contraction of the longitudi-nal and transverse musculature. In this example,the transverse muscle fibers serve simply to main-tain the cross-sectional area constant while the
Fig. 8. Plot of areas of the axial nerve cord, longitudinalmusculature, transverse musculature, and oblique musculaturedivided by the total area of the intrinsic arm muscle (excludingthe axial nerve cord) as observed in transverse sectionsobtained at identically spaced intervals down the length of anarm of Octopus bimaculoides. Note that the relative proportionsof a transverse section of the various muscle masses remainrelatively constant along the length of the arm. In contrast, thearea of the axial nerve cord, expressed as a proportion of theintrinsic arm musculature, increases dramatically from thebase to the tip of the arm.
OCTOPUS ARM STRUCTURE AND FUNCTION 839
Journal of Morphology DOI 10.1002/jmor

longitudinal muscle fibers on one side of the armshorten. Resistance to longitudinal compressionmight also be provided by passive stiffness fromtransversely or circularly arranged connective tis-sues or the passive stiffness of the musculature, aslong as they are capable of resisting the increasein diameter that results from the longitudinal com-pression. The limitation of a passive mechanismsuch as this, however, is that bending could beproduced at only one arm length since passivecomponents are fixed in length. The potentialinvolvement of transverse muscle fibers in bend-ing, as described earlier, would allow bending tooccur at a range of arm lengths as the arm elon-gates and shortens.
Bending can also occur in situations where thetransverse muscle fibers shorten while selectedlongitudinal muscle fibers maintain a constantlength (Kier and Smith, 1985). In this case, con-tractile activity in the longitudinal muscle fibersmust occur along one side of the arm to resist elon-gation of that side, and shortening of the trans-verse muscle fibers elongates the opposite side,creating a bend. Note that this mechanism and themechanism outlined earlier actually represent end-points in a continuum of relative shortening of thetransverse and longitudinal musculature in bend-ing. Localized bends of even smaller radius of cur-vature can be produced by unilateral shortening ofthe longitudinal musculature, decreasing thelength of one side of the arm, simultaneous withshortening of the transverse musculature, increas-ing the length of the opposite side of the arm. Asbefore, the extent of the bending (gentle vs. ab-rupt) depends on how much of the arm muscula-ture is recruited; highly localized bending, asobserved in the bending pulls or ‘‘fetch’’ behaviors(Sumbre et al., 2005, 2006), might involve thismore localized longitudinal and transverse musclefiber activity.
Forceful bending of the arm is aided by place-ment of the longitudinal muscle bundles as far aspossible from the neutral plane of the arm (theneutral plane of a beam in bending is usually inthe center and is where all bending stresses arezero). Bending in any plane requires that the lon-gitudinal muscle bundles be situated around theentire periphery of the cross section. As seen inFigure 2, both of these conditions are met in thearm as the longitudinal muscle is indeed situatedaway from the central axis of the arm and it isalso present around the entire periphery. As notedearlier, the cross-sectional area of the longitudinalmuscle fibers in the aboral quadrant of the arm isgreater than that of the other quadrants, suggestingthat aboral bending movements of the arm may bemore forceful than bending in other directions.
Stiffening. An increase in flexural stiffness ofthe arm can be caused by cocontraction of thetransverse and longitudinal musculature. This pat-
tern of activation is commonly observed and is animportant component of the reaching behaviorthat has been the subject of an elegant series ofexperimental and theoretical studies by Hochner,Flash, and coworkers (Gutfreund et al., 1996,1998; Yekutieli et al., 2005a,b). Based on thesestudies it was suggested that during reaching, awave of cocontraction of the transverse and longi-tudinal musculature passes from the base to thetip of the arm. As this stiffening propagates fromthe base of the arm outward it forms a passivebend at the transition between the stiffened baseand relaxed terminal portion of the arm, whichtrails behind the advancing wave of cocontraction.A similar wave of stiffening is observed in otheroctopus behaviors and may represent an importantstereotyped component of many octopus arm move-ments (Gutfreund et al., 1996, 1998).
In some arm behaviors, a significant portion ofthe arm, particularly at the base, may be stiffenedand the arm ‘‘swept’’ by rotation of the base in amanner analogous to rotation around a vertebrateshoulder joint. Such a behavior probably involvescocontraction of the transverse and longitudinalmusculature of the intrinsic muscle mass of thearm to stiffen it, while contraction of the muscula-ture of the web at the base of the arms couldcause, or at least enhance, the sweeping motion.The fiber trajectories of this web musculature havebeen well described by Guerin (1908) and includemuscle fibers that, depending on activation, couldrotate the arm bases aborally, orally, and laterally.
Torsion. Torsion or twisting of a muscular hydro-stat results from shortening of helically arrangedmuscle fibers (Kier, 1982; Kier and Smith, 1985).The arrangement and distribution of the obliquemuscle layers and their associated connective tis-sues provides just such a helical array. Becausethe oblique muscle layers have their origin andinsertion on the oral and aboral connective tissuesheets, and because their fiber angles are similar,a composite helical system is formed. Thus, theforce produced by an oblique muscle fiber is trans-mitted to a connective tissue fiber and then to anoblique fiber on the opposite side of the arm, andthen to another connective tissue fiber, and so on,along a helical path down the arm.
The arrangement of oblique muscle fibersobserved in this study suggests that a nonsymmet-rical pattern of activation occurs on opposite sidesof the arm during torsional movements. Asdescribed earlier, for all three pairs of oblique mus-cle layers, the handedness of the helix of one mem-ber of the pair is opposite to that of its pair on theother side of the arm. To generate a torsionalforce, there must be a continuous helical force tra-jectory down the length of the arm. The handed-ness of this helical trajectory determines the direc-tion of torsion. For instance, clockwise torsion ofthe tip of the arm relative to the base (when
840 W.M. KIER AND M.P. STELLA
Journal of Morphology DOI 10.1002/jmor

viewed from base to tip) requires the contractionof a left-handed helical muscle system. If theexternal oblique muscle layer on one side of thearm is left-handed, then it must be coactivatedwith the median oblique muscle layer on the oppo-site side of the arm in order to form the appropri-ate helical force trajectory. We thus predict a non-symmetrical pattern of activation of the obliquemuscle layers during torsional movements. Simul-taneous contraction of the oblique muscle layerpairs on each side of the arm (e.g., both left andright median oblique muscle layers) is predicted tooccur when an increase in torsional stiffness of thearm is required, perhaps in combination with coac-tivation of the transverse and longitudinal muscu-lature for increased flexural stiffness.
The torsional moment of the oblique musclelayers is dependent on the product of the forcethey produce (which is a product of the inherentpeak tension of the muscle fibers and their physio-logical cross-sectional area) and their distancefrom the neutral axis of the arm (the neutral axisis usually located at the center of a beam in tor-sion and is not subject to shear stress). It is thusinteresting that the external and median obliquemuscle layers are located away from the neutralaxis, thereby providing a relatively large momentarm for application of torsional forces. In thisregard the functional role of the internal obliquemuscle layers is uncertain, given their locationnearer to the central transverse musculature.While they possess the same handedness as theexternal oblique muscle layers on the same side ofthe arm and thus might augment the force ofexternal oblique muscle fiber contraction in tor-sion, their smaller torsional moment suggests thatthey possibly serve some other function in thearm. In addition to their more central location,they appear to possess also a lower fiber anglethan that of the median and external oblique mus-cle layers, but the significance of this difference isunclear. It is worth noting that the arms and thetentacular clubs of decapod cephalopods (squid andcuttlefish) include oblique muscle layers similar tothe external and median layers of octopus arms,but they lack the inner oblique muscle layersobserved in the octopus arm (Kier, 1982, 1988;Kier and Thompson, 2003). The general pattern ofmovement of squid arms and tentacular clubs issimilar to that of octopus arms. Octopus armselongate and shorten to a greater extent, but thesignificance of the inner oblique muscle layer inthis difference is unclear. In this regard, futurestudies involving electromyographic recordingsfrom specific arm muscle groups during movementand force generation would be of great interest.
Although the transverse musculature is likely toplay a major role in arm elongation, there is thepossibility that some force for elongation may begenerated by the external and median oblique
muscles. Oblique muscle fibers with a fiber angleequal to 548440 generate torsional forces only.Oblique muscle fibers with a fiber angle less than548440 generate force for shortening in addition totorsion, while those with a fiber angle greaterthan 548440 generate force for elongation in addi-tion to torsion (Kier and Smith, 1985). Because thefiber angle of the oblique muscle fibers in therelaxed octopus arms was measured to be greaterthan 548440, a component of the force creating tor-sion will also generate a force for elongation of thearm. As the arm extends, however, the fiber anglewill decrease and consequently the component ofthis force that elongates the arm will decrease aswell. If sufficient elongation of the arm occurs sothat the fiber angle falls below 548440, contractionof the oblique muscle layers will then generateforce for shortening the arms in addition to gener-ating a torsional force (Kier and Smith, 1985).
Implications of Octopus Arm Structure andFunction for Motor Control
Compared with more conventional hydrostaticskeletons that include large fluid-filled cavities, oc-topus arms and muscular hydrostats in generalhave the potential to generate highly localizedmovements and deformations by selectively con-tracting small groups of muscle fibers. Experimen-tal studies on the neuromuscular system suggestthat octopus arms do indeed have the neuromuscu-lar control required for localized movement; themotor units of the transverse and longitudinalmusculature of the arm are small and there is lit-tle evidence for electrical coupling between themuscle fibers (Matzner et al., 2000). Such localizeddeformations are essential for the production ofthe complex and diverse movements observed inthe octopus arm and in other muscular hydrostats.
The problem with such a system, however, is thepotential complexity of motor control that isrequired. Deformations in any direction can poten-tially occur at any location, or at multiple loca-tions, along the length of the arm. Recent studieson the neuromuscular control of octopus arms sug-gest that various mechanisms exist that may sim-plify arm control. For instance, Gutfreund et al.(1996) have studied the reaching behavior(described earlier) in Octopus vulgaris as the armis extended toward a target. Their analysesrevealed that certain features of the movement arestereotyped and they suggest that this simplifiesthe motor control. Indeed, in a later study, Sumbreet al. (2001) showed that arm extensions with typi-cal kinematics could be produced by arms isolatedfrom the brain, suggesting that a motor programfor arm extension is present in the neural circuitryof the arm, thereby simplifying its motor control.Recently, Sumbre et al. (2005, 2006) have analyzed
OCTOPUS ARM STRUCTURE AND FUNCTION 841
Journal of Morphology DOI 10.1002/jmor

a pulling behavior in O. vulgaris that they term‘‘fetch.’’ The movement is used to transfer prey tothe mouth and involves stiffening of the arm andthe formation of a highly localized bend that formsa ‘‘joint’’ at half the distance from the mouth to thepoint where the prey is held by the suckers. Theysuggest that this strategy provides an optimalsolution for point to point transfer and alsoreduces the number of variables that must be con-trolled, in this case only three degrees of freedom,again simplifying the motor control.
The remarkable similarity in the relative propor-tions of the cross section of the various musclemasses along the length of the tapering arm isalso of interest from the standpoint of neuromus-cular control. Such constancy in the relative size,and thus the force that the various components ofthe musculature can produce along the length ofthe arm, might also be important in simplifyingthe control of arm movement. This arrangementallows a given motor command involving somesubset of the arm musculature to elicit similararm deformations at any position along the lengthof the arm.
Implications of Octopus Arm Structure forMechanics
The arrangement and relative proportions of themusculature of the arms have several interestingmechanical implications. As described earlier,many of the transverse muscle fibers insert on thedense connective tissue sheet surrounding theaxial nerve cord. When the transverse muscula-ture contracts to generate elongation, provide sup-port for bending, or to stiffen the arm, the intrinsicmusculature will be pressurized. Will this pressur-ization compress the axial nerve cord? A prelimi-nary mechanical analysis suggests that the bal-ance of forces is such that the connective tissuesurrounding the axial nerve cord will be placed intension. It is perhaps easiest to envision this if thesystem is imagined as a cylinder within a cylinder,both consisting of flexible tension-resisting mem-branes. In between is a fluid of high bulk modulusand the two flexible membranes are connected bymuscle fibers that pressurize the fluid when theycontract. (Alternatively, it is perhaps easier tothink about the muscle fibers as tension-resistingcables and then pressurize the fluid in the spaceindependently.) In this situation, the total out-wardly radial force (per unit length of arm) will begreater than the inwardly radial force becausewhile the pressure will be the same, the total areaof the outer membrane subjected to that pressureis greater. Thus, even during the generation oflarge forces in the arm, it is likely that the axialnerve cord will not be subjected to the pressuresthat are present in the surrounding musculature.An analogous situation occurs in the trunks of ele-
phants, which are also muscular hydrostats,because the radial muscle of the trunk inserts ondense connective tissues surrounding the nasalpassages. Pressurization due to contraction of themusculature of the trunk also places the connec-tive tissues in tension and keeps the nasal pas-sages open (Kier and Smith, 1985). In the octopusarm it is unclear whether pressurization of theaxial nerve cord is potentially damaging, given thehigh bulk modulus of tissue and the lack of gas-filled cavities. The arrangement might, however,reduce the potential shear stresses on the axialnerve cord, which could indeed be harmful.
CONCLUSIONS
The arms of octopus show remarkable diversityand complexity of movement and thus represent aparticularly interesting example of the dynamicmusculoskeletal system provided by muscularhydrostatic support. A group of muscle fibers inthe arm may actively shorten or it may generateforce to limit elongation and thereby supportmovement of the arm as a whole. In addition, thecontraction of muscle fibers of a given orientationmay be highly localized or it may be more general-ized, with resulting localized versus general defor-mations or stiffening. Such diversity of movementand support is not possible in more conventionalskeletal support systems such as those with rigidskeletal elements where movement is restricted tojoints or most hydrostatic skeletons where musclecontraction increases the pressure throughout. Thepotential limitation of such a system, however, isthe complexity of neuromuscular control that is re-quired. In this regard, recent analyses (Gutfreundet al., 1996; Sumbre et al., 2001, 2005, 2006) thathave identified mechanisms of simplifying the con-trol are of particular interest and importance forour understanding of these appendages.
ACKNOWLEDGMENTS
We thank J. Thomas and S. Guarda for assis-tance with histology and T. Uyeno for assistancewith animal maintenance. We thank J. Hebrankand C. Rahn for helpful insights into arm mechan-ics. We also thank R. Hanlon and coworkers,Marine Resources Center, Marine Biological Labo-ratory, Woods Hole, MA 02543, for informationconcerning their ongoing analysis of octopus armbehaviors and for several specimens examined inthis study. R. Hanlon, B. Hochner, and T. Uyenoprovided helpful suggestions on the manuscript.MPS was supported by a Summer UndergraduateResearch Fellowship from the University of NorthCarolina at Chapel Hill Office of UndergraduateResearch.
842 W.M. KIER AND M.P. STELLA
Journal of Morphology DOI 10.1002/jmor

LITERATURE CITED
Browning J. 1980. The vasculature of Octopus arms: A scanningelectron microscope study of corrosion casts. Zoomorph 96:243–253.
Budelmann BU, Schipp R, Boletzky Sv. 1997. Cephalopoda. In:Harrison FW, Kohn AJ, editors. Microscopic Anatomy ofInvertebrates. Mollusca II, 6A. New York: Wiley. pp 119–414.
Chapman G. 1950. Of the movement of worms. J Exp Biol27:29–39.
Chapman G. 1958. The hydrostatic skeleton in the inverte-brates. Biol Rev Camb Philos Soc 33:338–371.
Chapman G. 1975. Versatility of hydraulic systems. J Exp Zool194:249–270.
Clark RB. 1964. Dynamics in Metazoan Evolution: The Originof the Coelom and Segments. London: Oxford UniversityPress. 313 p.
Clark RB. 1981. Locomotion and the phylogeny of the metazoa.Boll Zool 48:11–28.
Cloney RA, Brocco SL. 1983. Chromatophore organs, reflectorcells, iridocytes and leucophores in cephalopods. Am Zool23:581–592.
Colasanti G. 1876. Anatomische und Physiologische Untersu-chungen uber den Arm der Kephalopoden. Arch Anat PhysiolWissenchaftliche Med 9:480–500.
Cuvier G. 1817. Memoires pour servir a l’histoire et al’anatomie des mollusques. Memoires sur les Cephalopodes etsur leur anatomie. Paris: Deterville. 54 p.
Graziadei P. 1965. Muscle receptors in cephalopods. Proc R SocB 161:392–402.
Graziadei P. 1971. The nervous system of the arms. In: YoungJZ, editor. The Anatomy of the Nervous System of Octopusvulgaris. Oxford: Clarendon Press. pp 45–61.
Guerin J. 1908. Contribution a l’etude des systemes cutane,musculaire et nerveux de l’appareil tentaculaire des cephalo-podes. Arch Zool Exp Gen 38:1–178.
Gutfreund Y, Flash T, Yarom Y, Fiorito G, Segev I, Hochner B.1996. Organization of octopus arm movements: A model sys-tem for studying the control of flexible arms. J Neurosci16:7297–7307.
Gutfreund Y, Flash T, Fiorito G, Hochner B. 1998. Patterns ofarm muscle activation involved in octopus reaching move-ments. J Neurosci 18:5976–5987.
Gutmann WF. 1981. Relationships between invertebrate phylabased on functional-mechanical analysis of the hydrostaticskeleton. Am Zool 21:63–81.
Jones BA, Walker ID. 2006a. Kinematics for multisection con-tinuum robots. IEEE Trans Robot 22:43–55.
Jones BA, Walker ID. 2006b. Practical kinematics for real-timeimplementation of continuum robots. IEEE Trans Robot 22:1087–1099.
Kier WM. 1982. The functional morphology of the musculatureof squid (Loliginidae) arms and tentacles. J Morphol 172:179–192.
Kier WM. 1988. The arrangement and function of molluscanmuscle. In: Trueman ER, Clarke MR, editors. The Mollusca,Form and Function, Vol. 11. New York: Academic Press.pp 211–252.
Kier WM. 1992. Hydrostatic skeletons and muscular hydrostats.In: Biewener AA, editor. Biomechanics (Structures and Sys-tems): A Practical Approach. Oxford: IRL Press at OxfordUniversity Press. pp 205–231.
Kier WM, Smith AM. 1990. The morphology and mechanics ofoctopus suckers. Biol Bull 178:126–136.
Kier WM, Smith AM. 2002. The structure and adhesive mecha-nism of octopus suckers. Integr Comp Biol 42:1146–1153.
Kier WM, Smith KK. 1985. Tongues, tentacles and trunks: Thebiomechanics of movement in muscular-hydrostats. J LinnSoc Lond (Zool) 83:307–324.
Kier WM, Thompson JT. 2003. Muscle arrangement, functionand specialization in recent coleoids. Berliner Palaobiolog-ische Abhandlungen 03:141–162.
Matzner H, Gutfreund Y, Hochner B. 2000. Neuromuscular sys-tem of the flexible arm of the octopus: Physiological charac-terization. J Neurophysiol 83:1315–1328.
McMahan W, Jones B, Walker ID. 2005. Robotic manipulatorsinspired by cephalopod limbs. J Eng Des (Eng Des Pract)1P:01P2.
McMahan W, Pritts M, Chitrakaran V, Dienno D, Grissom M,Jones B, Csencsits M, Rahn CD, Dawson D, Walker ID. 2006.Field trials and testing of ‘‘OCTARM’’ continuum robots, Pro-ceedings of IEEE International Conference on Robotics andAutomation, Orlando, FL, pp 2336–2341.
Messenger JB, Nixon M, Ryan KP. 1985. Magnesium chlorideas an anaesthetic for cephalopods. Comp Biochem Physiol82C:203–205.
O’Dor RK, Portner HO, Shadwick RE. 1990. Squid as eliteathletes: Locomotory, respiratory, and circulatory integration.In: Gilbert DL, Adelman WJ, Arnold JM, editors. Squid asExperimental Animals. New York: Plenum Press. pp 481–503.
Rossi F, Graziadei P. 1954. Nouvelles recherches sur le systemenerveux du bras des cephalopodes avec des methodes specifi-ques pour le tissu nerveux. Acta Anat 22:202–215.
Rossi F, Graziadei P. 1956. Nouvelles contributions a la connais-sance du systeme nerveux du tentacule des cephalopodes.Partie III. Acta Anat 28:206–216.
Smith KK, Kier WM. 1989. Trunks, tongues and tentacles:Moving with skeletons of muscle. Am Sci 77:28–35.
Socastro ME. 1969. Observaciones sobre el significado estruc-tural y funcional de la musculatura braquial de los Cefalopo-dos. Bol R Soc Esp Hist Nat (Biol) 67:181–191.
Sumbre G, Gutfreund Y, Fiorito G, Flash T, Hochner B. 2001.Control of octopus arm extension by a peripheral motor pro-gram. Science 293:1845–1848.
Sumbre G, Fiorito G, Flash T, Hochner B. 2005. Motor controlof flexible octopus arms. Nature 433:595–596.
Sumbre G, Fiorito G, Flash T, Hochner B. 2006. Octopuses usea human-like strategy to control precise point-to-point armmovements. Curr Biol 16:767–772.
Tittel K. 1961. Der funktionelle Aufbau des Tintenfischarmesim Vergleich mit dem Muskelkorper der Saugerzungen. Verhd Anat Ges 57:264–275.
Tittel K. 1964. Saugnapf-, epi- und hypofasciale Armmuskulaturder Kephalopoden—ein Beitrag zur funktionellen Anatomiefreibeweglicher Skelettmuskelkorper. Gegenbaurs MorpholJahr 106:90–115.
Wainwright SA. 1970. Design in hydraulic organisms. Natur-wissenschaften 57:321–326.
Wainwright SA. 1982. Structural systems: Hydrostats andframeworks. In: Taylor CR, Johansen K, Bolis LA, editors.Companion to Animal Physiology. New York: Cambridge Uni-versity Press. pp 325–338.
Walker ID, Dawson DM, Flash T, Grasso FW, Hanlon RT, HochnerB, KierWM, Pagano CC, Rahn CD, Zhang QM. 2005. Continuumrobot arms inspired by cephalopods. Proc SPIE 5804:303–314.
Yekutieli Y, Sagiv-Zohar R, Aharonov R, Engel Y, Hochner B,Flash T. 2005a. Dynamic model of the octopus arm: I. Biome-chanics of the octopus reaching movement. J Neurophysiol94:1443–1458.
Yekutieli Y, Sagiv-Zohar R, Hochner B, Flash T. 2005b.Dynamic model of the octopus arm. II. Control of reachingmovements. J Neurophysiol 94:1459–1468.
OCTOPUS ARM STRUCTURE AND FUNCTION 843
Journal of Morphology DOI 10.1002/jmor