the effect of epidural compression on cerebral cortex: a...
TRANSCRIPT

JOURNAL OF NEUROTRAUMAVolume 20, Number 8, 2003© Mary Ann Liebert, Inc.
The Effect of Epidural Compression on Cerebral Cortex: A Rat Model
JENG-RUNG CHEN,1 YUEH-JAN WANG, 2 and GUO-FANG TSENG1
ABSTRACT
We developed a rat model of epidural plastic bead implantation to study the effect of physical com-pression on the cerebral cortex. Epidural implantation of a bead of appropriate size compressed theunderlying sensorimotor cortex without apparent ischemia, since the capillary density of the cortexwas increased. Although the thickness of all layers of the compressed cortex was significantly de-creased, no apparent changes in the number of NADPH-diaphorase reactive neurons, reactive as-trocytes, or microglial cells were observed, nor were apoptotic neurons observed. In fact, the den-sities of the neurons in most cortical layers apparently increased. To determine how epiduralcompression affects neuronal morphology, the dendritic arbors of layer III and V pyramidal neu-rons were evaluated using a fixed tissue intracellular dye injection technique. Neurons in both lay-ers remained pyramidal in shape and their somatic sizes remained unaltered for at least a monthafter compression. On the other hand, their total dendritic length was significantly reduced begin-ning at 3 days post implantation. These analyses showed that apical dendrites were affected soonerthan basal ones. The reduction of dendritic length was associated with a drop in the number of den-dritic branches rather than dendritic trunks, suggesting the trimming of the peripheral part of thedendritic arbor. Detailed analysis showed that dendritic spines on all dendrites were reduced asearly as 3 days following implantation. These results suggest that cortical neurons remodel theirstructures substantially within 3 days after being subjected to epidural compression.
Key words: brain; injury; meningioma; trauma; tumor
767
1Department of Anatomy and Cell Biology, College of Medicine, National Taiwan University, Taipei, Taiwan.2Department of Anatomy, College of Medicine, Tzu-Chi University, Hualien, Taiwan.
INTRODUCTION
PATHOLOGICAL STUDIES show that a brain tumor un-derlying the cerebral cortex can cause compression
and destruction of the nervous tissue via mechanical aswell as chemical factors (Spacek, 1987). On the otherhand, the removal of a meningioma, a tumor that is ex-pected to at least compress adjacent cortical tissue, issometimes thought of as benign and curative. Compres-
sion is likely to account at least in part for the clinicalmanifestations of this abnormality and often the key pur-pose of surgical removal is to relieve this compression.In practice, this compression may be prolonged becauseasymptomatic meningiomas are often observed for a pe-riod of 3–12 months before a definite decision is madeabout the course of treatment. Similar compression ofcortical tissue may also happen following epidural or sub-dural hematoma, hemorrhage in the subarachnoid space,

hemangioma, traumatic head injury, or even in somelarge-scale A-V malformations. Underlying brain tissuemay respond to focal compression with plastic changesover time. Today we have a better understanding of thepathology of meingiomas (O’Rahilly and Muller, 1986;Zülch, 1986) and have seen improvements following theirsurgical removal, which often largely rectifies some oftheir clinical manifestations such as seizure, hemiparesis,visual field loss, aphasia, and other focal symptoms(Reifenberger et al., 1996). However, the effects of thegrowing mass on the underlying cortex at the cellularlevel remain largely unexplored.
In this study, we developed a rat model of focal com-pression on the cerebral cortex by implanting a plasticbead epidurally. This enabled us to study the effect ofphysical compression on brain tissue alone. The im-planted lightweight and inert bead was not expected tosecrete any substance or interact directly with the under-lying cortical tissue, thus enabling us to study the effectof physical pressure with presumably no other compli-cations. The effects of short- and long-term compressionwere studied. To find out how physical compression af-fected cortical function, an intracellular dye injectiontechnique was used to study the soma-dendritic arbors oflayer III and V pyramidal neurons, since they are the prin-cipal output neurons of the cortex. In addition, the den-sities of the dendritic spines on these neurons were alsoexamined, since injury has been reported to be associatedwith alterations in neuronal spine density (Rutledge et al.,1972). Information gathered in the present study is im-portant in leading to a better understanding the patho-physiology of the cerebral cortex opposed by an ex-panding mass and may also provide ideas for improvingclinical decision making.
MATERIALS AND METHODS
Animal and Tissue Preparation
A total of 24 young adult female Wistar rats (CharlesRiver) aged 6–7 weeks (150–175 g) were studied. Ani-mals were housed and cared for according to the guide-lines of the Animal Research Committee of the NationalScience Council. All efforts were made to minimize an-imal suffering during and following surgery. Flatroundish polyethylene beads, 5 mm in diameter and 2.8mm in thickness, were ground down on one of their flatsides to a thickness of 1.5 mm (Fig. 1A). These particleswere used to compress the cortex. For implantation, ratswere anaesthetized with 7% chloral hydrate (4.5 mL/kg,Merck, Darmstadt, Germany) and mounted on a stereo-taxic device; an elliptical hole with a short diameter (~4mm) just smaller and a long diameter (~6 mm) just longer
than the diameter of the flat surface of the bead wasdrilled on the skull over the right sensorimotor cortex(Fig. 1A). The plastic bead was implanted into theepidural space as illustrated in Figure 1B–D and care-fully positioned so that it was lodged in the hole. Thiscould be done easily since the diameter of the bead wasslightly greater than the short axis of the hole (Fig. 1D).With such an arrangement, the bead sealed the hole onthe skull without further manipulation. Animals subjectedto bead implantation were divided into five groups andallowed to survive for 3 days (n � 5), 1 month (n � 5),3 months (n � 2), 6 months (n � 2), or 1 year (n � 4).Sham-operated animals (n � 4) were subjected to thesame surgery without bead implantation. Two additionalunoperated rats served as normal controls. At the end ofthe survival periods the animals were processed for in
CHEN ET AL.
768
FIG. 1. Schematic diagram (not to scale) demonstrating theepidural implantation of a plastic bead. In A, an elliptical holewith a long and short axis of approximately 6 and 4 mm re-spectively was drilled into the skull over the sensorimotor cor-tex of a rat. A plastic bead with a flat surface 5 mm in diame-ter and a thickness of 1.5 mm was used for epidural implantationto compress the cerebral cortex epidurally. After exposing thedura, the bead with its flat surface up was slid obliquely, oneend first, along the short axis of the hole (the direction of thearrow in B and C) and into the hole (B and C). The bead couldbe easily placed since its diameter was slightly shorter than thelong axis of the hole. After the bead was inserted completely,it was pulled gently and slightly in the opposite direction (thearrow in D) so that the other end of the bead could be slid un-derneath the bone at the edge of the other side of the hole (D).The bead stayed lodged in place and sealed the hole completely.This could be easily done since the short axis of the hole wasslightly less than the diameter of the bead.
F1 �

vitro intracellular dye injection as described in an earlierpaper (Wang et al., 2002). Briefly, they were deeplyanaesthetized with chloral hydrate and perfused with 50ml of lukewarm saline, followed by a fixative containing2% paraformaldehyde in 0.1M phosphate buffer (PB), pH7.3, at room temperature for 30 min. Immediately fol-lowing the perfusion, the whole brains were carefully re-moved and sectioned with a vibratome (Technical Prod-ucts International, St. Louis, MO) into 300-�m-thickcoronal slices. Half of the slices containing the com-pressed cortical area were processed by intracellular dyeinjection to reveal the morphology of selective individ-ual cells. The rest were postfixed in 4% paraformalde-hyde in 0.1M PB for 3 days. They were then cryopro-tected and resectioned into 20-�m sections (Tseng et al.,1991) for studying their cytoarchitecture as described be-low.
Histological Procedures for Revealing Cytoarchitecture
Selected sections were stained with Cresyl violet forgeneral evaluation. To find out whether the NO environ-ment in the compressed cortical area was altered, we usedNADPH-diaphorase histochemistry to localize NO-pro-ducing cells in the studied cortical area. Sections wereincubated free-floating in 0.1M PB containing 0.3% Tri-ton X-100, 0.1 mg/mL nitro-blue tetrazolium (Sigma, StLouis, MO), and 1.0 mg/mL �-NADPH (Sigma) at 37°Cfor 20–40 min to reveal NADPH-diaphorase–containingcells. Following the reaction, sections were mounted ontogelatin-coated slides, air-dried, dehydrated, and cover-slipped with Permount (Fisher, Fair Lawn, NJ). To findout whether the microglial cells of the compressed cor-tex were changed, some of the sections were treated in0.5% H2O2 containing 0.5% Triton X-100 in 0.1M phos-phate-buffered saline (PBS), pH 7.3, for 1 h. After beingrinsed three times in PBS, they were incubated in solu-tion containing Griffonia simplicifolia isolectin B4 (GSAI-B4) conjugated with horseradish peroxidase (HRP) (25�g/mL, Sigma) and 0.5% Triton X-100 in 0.05M Trisbuffered saline (TBS), pH 7.4, for 18 h at 4°C (Tseng etal., 1996). To reveal neurons and astrocytes, sectionswere reacted with rabbit polyclonal antibodies to theubiquitous neuronal protein PGP 9.5 (1:1000, Chemicon,Temecula, CA) and mouse monoclonal antibodies to glialfibrillary acidic protein (GFAP, 1:400, Chemicon), re-spectively, for 18 h at 4°C. Biotinylated goat anti-rabbit(1:200) and horse anti-mouse (1:200) immunoglobulinswere used as the secondary antibodies, respectively, andthe sections were incubated for 1 h at room temperature.After being rinsed three times in PBS, they were incu-bated in standard avidin-biotin HRP reagent (Vector,Burlingame, CA) for 1 h at room temperature. Sections
were then reacted in 0.05% 3-3�-diaminobenzidinetetrahydrochloride (DAB, Sigma) in 0.05 M TBS, con-taining 0.01% H2O2. Reacted sections were mounted onsubbed slides, air-dried, and coverslipped in Permount.The fact that GSA I-B4 also labels capillaries in additionto reactive microglial cells (Tseng et al., 1996) enabledus to evaluate the diameters and densities of capillariesin the studied cortical area.
Intracellular Dye Injection andImmunoconversion of the Injected Dye
We used Lucifer yellow (LY, Sigma) as the intracel-lular dye. In order to visually guide the impalement ofselective cells for dye injection, slices were first treatedwith 0.1M PB containing 10�7M 4�, 6-diamidino-2-phenyl-indole (DAPI; Sigma) for 30 min. This made thecell nuclei fluoresce blue under the same fluorescence fil-ter set (390–420, FT 425, LP 450) that visualized LY asyellow. A slice was then placed in a dish on the stage ofa Zeiss fixed-stage, epifluorescence microscope (Axio-scop) and covered with a thin layer of 0.1M PB. An in-tracellular micropipette filled with LY solution (4% inwater) was mounted on a 3-axial hydraulic micromanip-ulator (Narishige, Tokyo, Japan) and a long-working dis-tance objective lens (20�) was used to facilitate the se-lection of neurons in specific layers for dye injection.Constant negative current was used to inject the LY un-til all terminal dendrites fluoresced brightly. The slicewas then removed and postfixed in 4% paraformaldehydein 0.1M PB for 3 days. The slices were then rinsed thor-oughly in 0.1M PB, cryoprotected, and carefully sec-tioned into 60-�m-thick serial sections with a sliding mi-crotome (Wang et al., 1996).
To convert the intracellular dye LY into nonfading ma-terial, sections were preincubated in 1% H2O2 in PB for30–60 min to remove endogenous peroxidase activity.They were then rinsed three times in PBS and incubatedfor 1 h in PBS containing 2% bovine serum albumin(BSA) and 1% Triton X-100. Sections were then treatedin solution containing biotinylated rabbit anti-LY (1:200;Molecular Probes, Eugene, OR) in PBS for 18 h at 4°C.Following subsequent rinses in PBS, sections were incu-bated with standard ABC reagent for 3 h at room tem-perature. They were then reacted at room temperaturewith a solution containing 0.05% DAB and 0.01% H2O2
in 0.05M Tris buffer. Reacted sections were mountedonto slides, dehydrated in alcohol, cleared in xylene, andcoverslipped with Permount.
Data Analysis
To find out how cortical compression affects the mor-phology of cortical output neurons, we reconstructed the
ANATOMICAL FEATURES OF CORTEX UNDER COMPRESSION
769
soma-dendritic trees of layer III and V pyramidal neu-rons through serial sections using a Camera lucida draw-ing tube and a 40� objective lens in two-dimensionalplane. The demarcation between soma and dendrites wastaken as the point where the convex curvature of the somabecame concave (Tseng and Royce, 1986). Dendriteswere divided into basal and apical dendrites followingthe method described for rat cortical pyramidal neurons(Tseng and Prince, 1993). Their lengths were determinedfrom the two-dimensional reconstruction using a PC-based software (Freemax Image-Pro, Media Cybernetics,Silver Spring, MD). Basal and apical dendritic lengthswere the sum of the basal and apical dendritic branches,respectively, while total dendritic length represented thesum of both.
The profuseness of the dendritic arbor of these neu-rons was analyzed following the method of Sholl bycounting dendritic crossings with concentric circlesdrawn at 50-�m step increments in radii starting from the
center of the soma (Tseng and Prince, 1993). To find outwhether cortical compression affects the density of den-dritic spines, we chose to reconstruct in detail represen-tative proximal and distal segments of their basal and api-cal dendrites using a 100� oil-immersion objective lens.For layer III pyramidal neurons, proximal and distal basaldendrites were defined as the segments 25–75 �m and100–150 �m from the soma, respectively. For the rela-tively large layer V pyramidal neurons, proximal and dis-tal basal dendrites were defined as the segments 50–100�m and 150–200 �m from where they originate from thesoma, respectively. On the other hand, the reconstructedproximal and distal apical dendrites were the first or thesecond branch of the apical trunk and the terminal den-drites after the last branch point, respectively, for bothlayer III and V pyramidal neurons. No attempt was madeto correct for tissue shrinkage because its exact extentwas not known. This is unlikely to affect the comparisonbetween groups since all tissue studied was subjected to
CHEN ET AL.
770
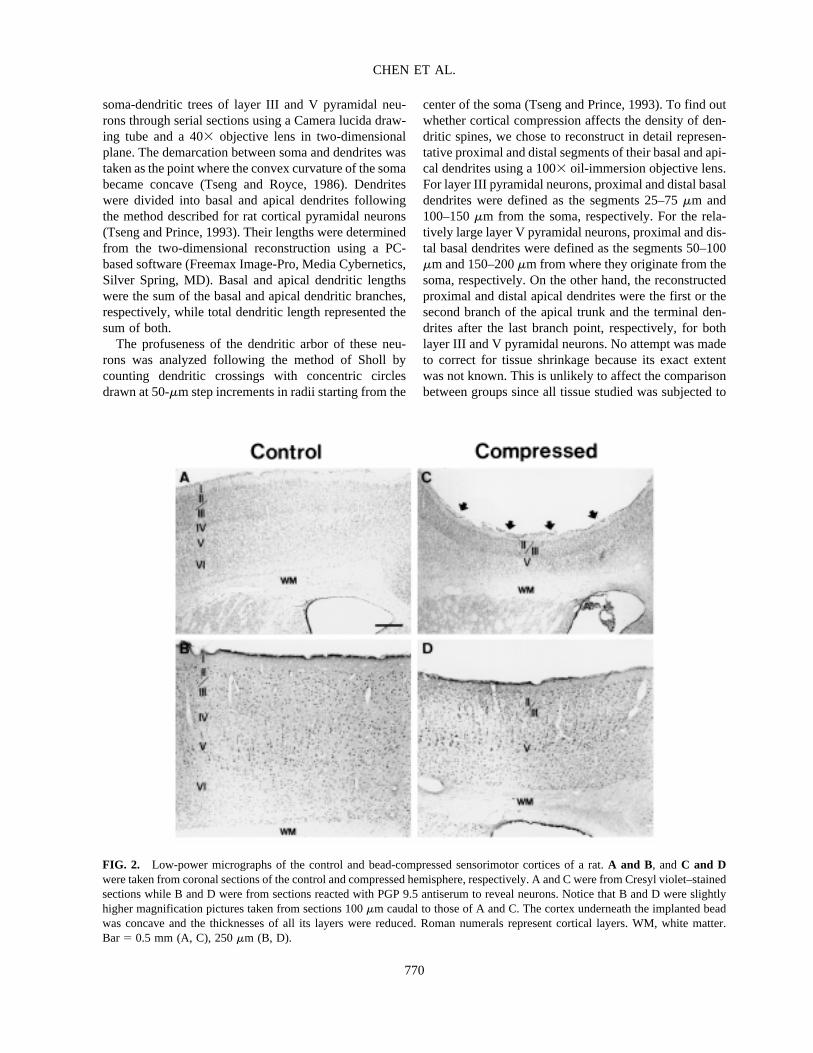
FIG. 2. Low-power micrographs of the control and bead-compressed sensorimotor cortices of a rat. A and B, and C and Dwere taken from coronal sections of the control and compressed hemisphere, respectively. A and C were from Cresyl violet–stainedsections while B and D were from sections reacted with PGP 9.5 antiserum to reveal neurons. Notice that B and D were slightlyhigher magnification pictures taken from sections 100 �m caudal to those of A and C. The cortex underneath the implanted beadwas concave and the thicknesses of all its layers were reduced. Roman numerals represent cortical layers. WM, white matter.Bar � 0.5 mm (A, C), 250 �m (B, D).
similar shrinkage, if any. Data was expressed as mean�SE. Statistical significance between groups was deter-mined using two-tailed Student’s t-test.
RESULTS
Implantation of plastic beads did not seem to affect theanimals; they appeared physically sound, continued togain weight, and were indistinguishable in general fromsham-operated or normal animals. The overall appear-ance of the cortex was indistinguishable between the
sham-operated and the normal animals, and also from thecortical area peripheral to the compressed region of thebead-implanted animals (not shown). Therefore, datafrom sham-operated controls were used as the controlsfor all comparison purposes throughout this paper. In allsurvival groups, the sensorimotor cortex immediately be-neath the bead remained six-layered in structure but hada concave surface (Fig. 2). The thicknesses of the com-pressed cortex and its layers were all significantly re-duced (Table 1). No apparent apoptotic neurons werefound in the affected cortical area. To find out whetherthe thinning was accompanied by neuronal death, we cal-
ANATOMICAL FEATURES OF CORTEX UNDER COMPRESSION
771
TABLE 1. THE THICKNESS OF SENSORIMOTOR CORTICAL LAMINAE (�M)
Control Comp-3D Comp-1M Comp-1Y(n � 46) (n � 35) (n � 39) (n � 28)
Layer I 122� 24 108� 23** 106 � 19** 104 � 23**Layer II~III 450 � 64 275� 39** 251 � 41** 247 � 54**Layer IV 229� 42 166� 38** 160 � 27** 159 � 19**Layer V 305� 48 180� 25** 155 � 26** 177 � 37**Layer VI 544� 82 285� 49** 241 � 36** 261 � 52**Total 1648� 143 1014� 11** 913 � 88** 948 � 123**
Values are means� S.E. for the number of sections (n) analyzed from all animals of each treated group. Control is from sham-operated animals; comp-3D, -1M, and -1Y represent compressed cortex examined 3 days, 1 month and 1 year after the deposition ofbead.
** p � 0.01 between the marked value and its corresponding control, 2-tailed Student’s t-test.
TABLE 2. DENSITIES OF BLOOD VESSELS, GLIAL CELLS, AND NEURONS OF THE SENSORIMOTOR CORTEX
Control Comp-3D Comp-1M
Percentage of blood vessel (%) 6.14� 1.13 8.17� 1.2** 8.19 � 1.6**(n � 35) (n � 29) (n � 32)
Diameter of blood vessel (�m) 12.2� 1.5 13.0� 1.5 12.6� 1.8(n � 56) (n � 42) (n � 52)
Density of astrocyte 5.6� 1.3 5.8� 1.1 6.0� 1.1(n � 35) (n � 30) (n � 31)
Density of microglial cell 2.7� 1.0 3.0� 0.9 2.4� 1.1(n � 29) (n � 25) (n � 26)
Density of layer II/III neurons 5.8� 1.0 7.8� 1.4** 7.7 � 1.6**(n � 42) (n � 35) (n � 41)
Density of layer IV neurons 9.6� 1.3 11.3� 1.1** 10.7 � 1.3*(n � 39) (n � 24) (n � 25)
Density of layer V neurons 3.8� 0.7 4.0� 1.0 4.5� 1.1(n � 40) (n � 29) (n � 28)
Density of layer VI neurons 6.9� 1.1 9.1� 1.6** 9.8 � 1.5**(n � 36) (n � 38) (n � 41)
Values are means� S.E. for the number of areas (n) quantified from representative sections of all animals of each group. Control,comp-3D, and comp-1M represent sham-operated and bead-implanted animals 3 days and 1 month following treatment.
*p � 0.05; **p � 0.01 between the marked and its corresponding control, 2-tailed Student’s t-test. Percentage of blood vesselwas calculated as the total area of capillaries per 100 �m2 of the cortical area. Densities were calculated as the number of cells per2500 �m2 of the cortical area.
F2�
T1�
culated the density of neurons in each layer from sectionsthat were reacted with PGP 9.5 antiserum that labeled theneurons (Table 2). The densities of neurons in layer IIthrough VI were all increased at 3 days and 1 month af-ter being subjected to compression, although the differ-ence in layer V failed to reach statistical significance,
perhaps due to the large variability (Table 2). Like their control counterparts (Fig. 3A), few NADPH-di-aphorase–containing neurons were found in the com-pressed cortices of any of the animals examined (Fig.3B). To find out whether the applied compression af-fected regional cortical blood flow, the diameter and the
CHEN ET AL.
772
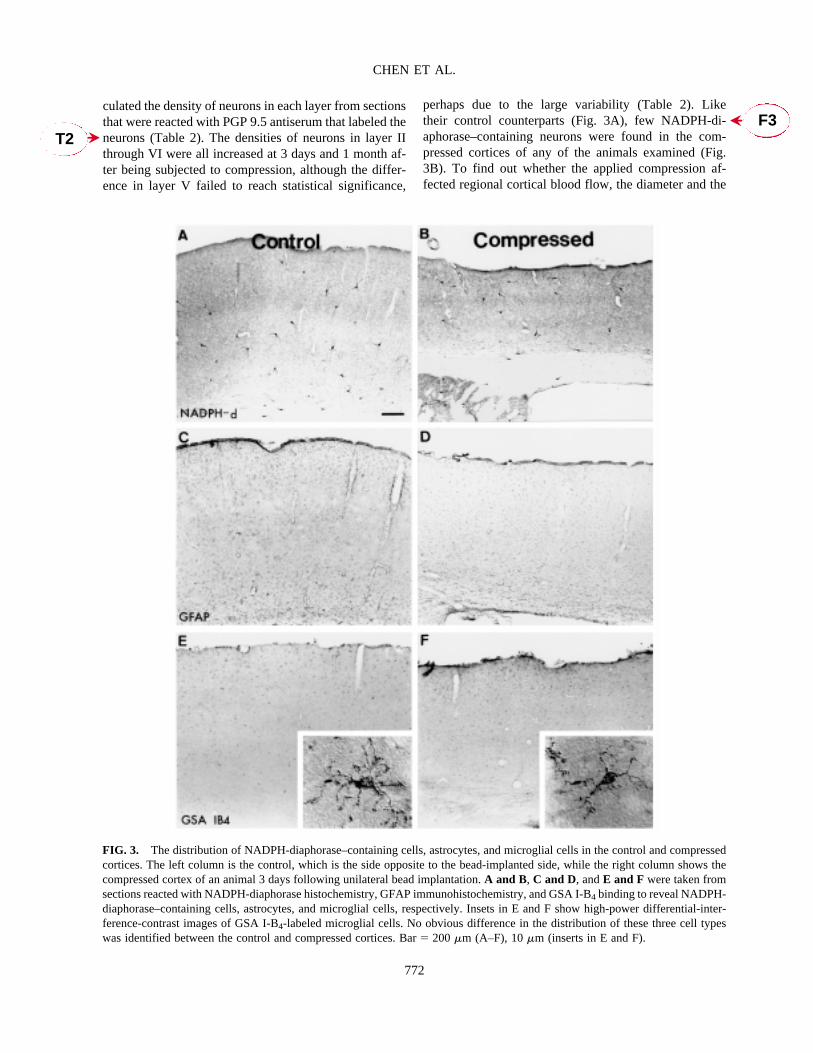
FIG. 3. The distribution of NADPH-diaphorase–containing cells, astrocytes, and microglial cells in the control and compressedcortices. The left column is the control, which is the side opposite to the bead-implanted side, while the right column shows thecompressed cortex of an animal 3 days following unilateral bead implantation. A and B, C and D, and E and F were taken fromsections reacted with NADPH-diaphorase histochemistry, GFAP immunohistochemistry, and GSA I-B4 binding to reveal NADPH-diaphorase–containing cells, astrocytes, and microglial cells, respectively. Insets in E and F show high-power differential-inter-ference-contrast images of GSA I-B4-labeled microglial cells. No obvious difference in the distribution of these three cell typeswas identified between the control and compressed cortices. Bar� 200 �m (A–F), 10 �m (inserts in E and F).
T2
�
F3�
density of capillaries revealed by GSA I-B4 binding werealso compared. There was no difference between themean diameters of the capillaries of the compressed andcontrol cortical area; however, the compressed corticalarea had a significantly higher density of capillaries, atleast at the two time points (3 days and 1 month afterbead implantation) that we examined. Regarding neu-roglial cells, neither the densities of the GFAP-labeledastrocytes (Fig. 3C, D) nor those of the GSA I-B4 labeledreactive microglial cells (Fig. 3E, F) were altered in thecompressed cortical areas of animals examined at 3 daysand 1 month following bead implantation. The effect ofcompression was quite reproducible in this model.
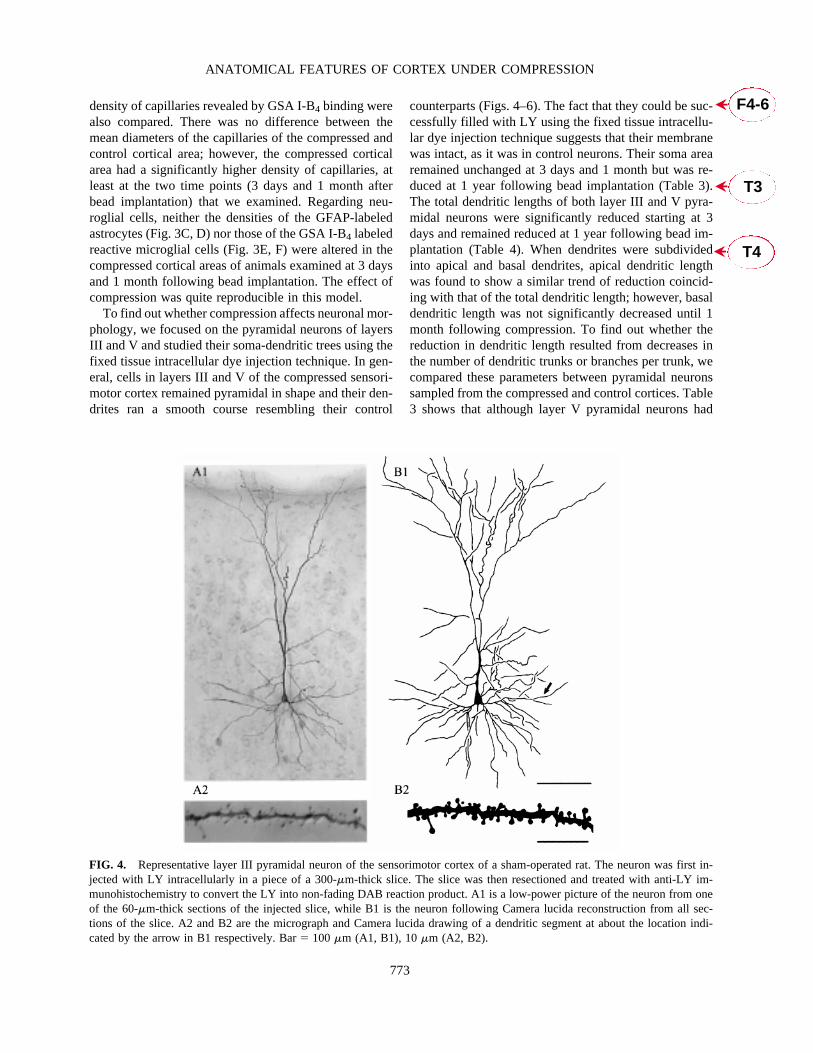
To find out whether compression affects neuronal mor-phology, we focused on the pyramidal neurons of layersIII and V and studied their soma-dendritic trees using thefixed tissue intracellular dye injection technique. In gen-eral, cells in layers III and V of the compressed sensori-motor cortex remained pyramidal in shape and their den-drites ran a smooth course resembling their control
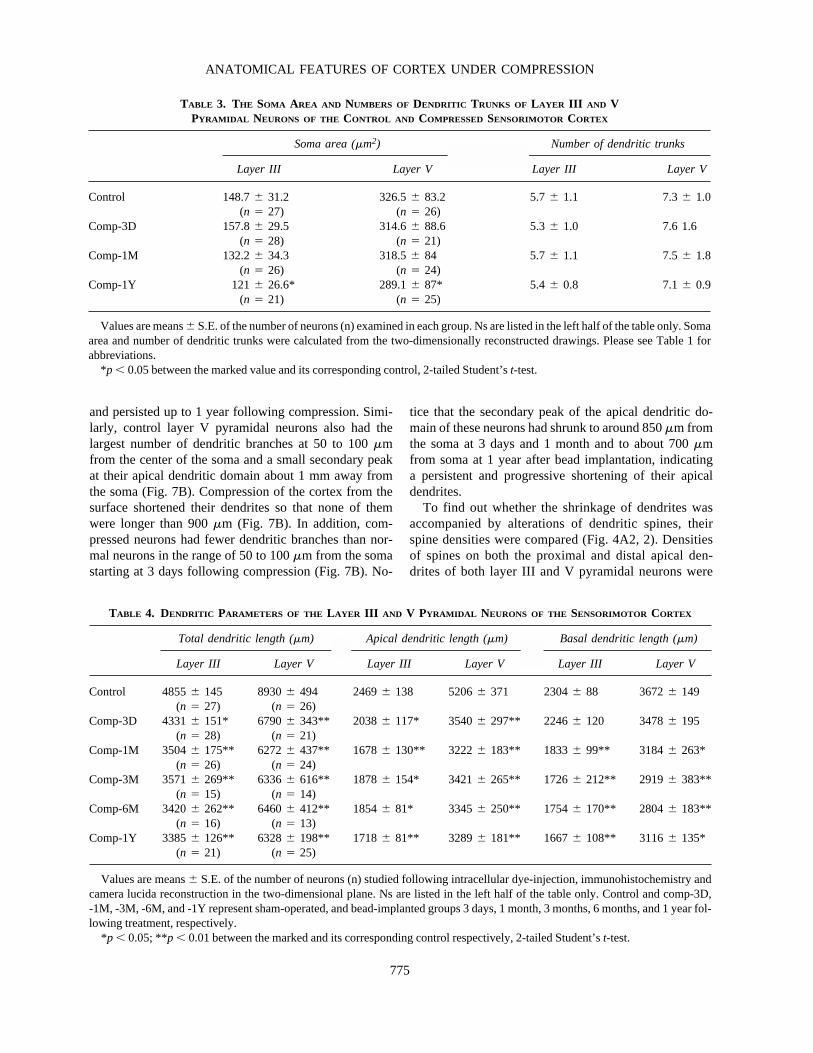
counterparts (Figs. 4–6). The fact that they could be suc-cessfully filled with LY using the fixed tissue intracellu-lar dye injection technique suggests that their membranewas intact, as it was in control neurons. Their soma arearemained unchanged at 3 days and 1 month but was re-duced at 1 year following bead implantation (Table 3).The total dendritic lengths of both layer III and V pyra-midal neurons were significantly reduced starting at 3days and remained reduced at 1 year following bead im-plantation (Table 4). When dendrites were subdividedinto apical and basal dendrites, apical dendritic lengthwas found to show a similar trend of reduction coincid-ing with that of the total dendritic length; however, basaldendritic length was not significantly decreased until 1month following compression. To find out whether thereduction in dendritic length resulted from decreases inthe number of dendritic trunks or branches per trunk, wecompared these parameters between pyramidal neuronssampled from the compressed and control cortices. Table3 shows that although layer V pyramidal neurons had
ANATOMICAL FEATURES OF CORTEX UNDER COMPRESSION
773
FIG. 4. Representative layer III pyramidal neuron of the sensorimotor cortex of a sham-operated rat. The neuron was first in-jected with LY intracellularly in a piece of a 300-�m-thick slice. The slice was then resectioned and treated with anti-LY im-munohistochemistry to convert the LY into non-fading DAB reaction product. A1 is a low-power picture of the neuron from oneof the 60-�m-thick sections of the injected slice, while B1 is the neuron following Camera lucida reconstruction from all sec-tions of the slice. A2 and B2 are the micrograph and Camera lucida drawing of a dendritic segment at about the location indi-cated by the arrow in B1 respectively. Bar� 100 �m (A1, B1), 10 �m (A2, B2).
F4-6�
T4�
T3�
more numerous dendritic trunks than layer III cells, nei-ther cell type showed reduction in its dendritic trunks af-ter being subjected to cortical compression.
The profuseness of the dendritic arbors of these pyra-midal neurons is analyzed and plotted in Figure 6. In con-trol cortex, the number of dendritic branches of layer IIIpyramidal neurons peaked at 50–100 �m from the cen-
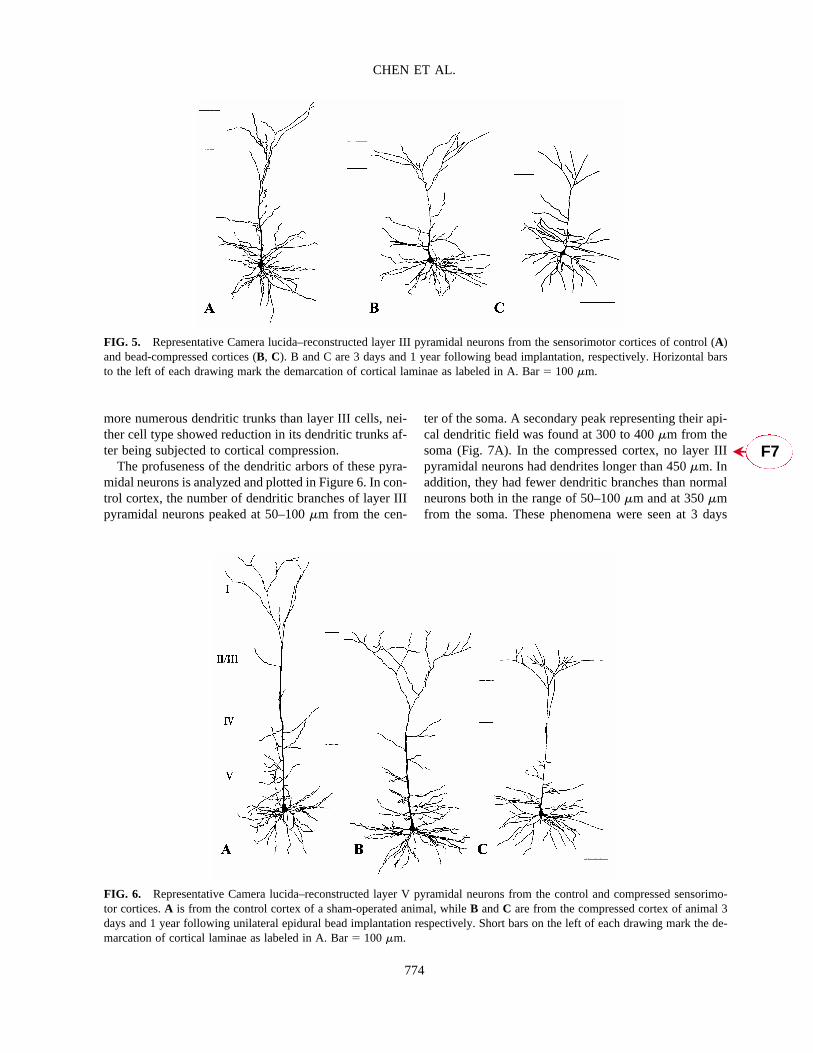
ter of the soma. A secondary peak representing their api-cal dendritic field was found at 300 to 400 �m from thesoma (Fig. 7A). In the compressed cortex, no layer IIIpyramidal neurons had dendrites longer than 450 �m. Inaddition, they had fewer dendritic branches than normalneurons both in the range of 50–100 �m and at 350 �mfrom the soma. These phenomena were seen at 3 days
CHEN ET AL.
774
FIG. 5. Representative Camera lucida–reconstructed layer III pyramidal neurons from the sensorimotor cortices of control (A)and bead-compressed cortices (B, C). B and C are 3 days and 1 year following bead implantation, respectively. Horizontal barsto the left of each drawing mark the demarcation of cortical laminae as labeled in A. Bar� 100 �m.
FIG. 6. Representative Camera lucida–reconstructed layer V pyramidal neurons from the control and compressed sensorimo-tor cortices. A is from the control cortex of a sham-operated animal, while B and C are from the compressed cortex of animal 3days and 1 year following unilateral epidural bead implantation respectively. Short bars on the left of each drawing mark the de-marcation of cortical laminae as labeled in A. Bar� 100 �m.
F7�
and persisted up to 1 year following compression. Simi-larly, control layer V pyramidal neurons also had thelargest number of dendritic branches at 50 to 100 �mfrom the center of the soma and a small secondary peakat their apical dendritic domain about 1 mm away fromthe soma (Fig. 7B). Compression of the cortex from thesurface shortened their dendrites so that none of themwere longer than 900 �m (Fig. 7B). In addition, com-pressed neurons had fewer dendritic branches than nor-mal neurons in the range of 50 to 100 �m from the somastarting at 3 days following compression (Fig. 7B). No-
tice that the secondary peak of the apical dendritic do-main of these neurons had shrunk to around 850 �m fromthe soma at 3 days and 1 month and to about 700 �mfrom soma at 1 year after bead implantation, indicatinga persistent and progressive shortening of their apicaldendrites.
To find out whether the shrinkage of dendrites wasaccompanied by alterations of dendritic spines, theirspine densities were compared (Fig. 4A2, 2). Densitiesof spines on both the proximal and distal apical den-drites of both layer III and V pyramidal neurons were
ANATOMICAL FEATURES OF CORTEX UNDER COMPRESSION
775
TABLE 3. THE SOMA AREA AND NUMBERS OF DENDRITIC TRUNKS OF LAYER III AND V PYRAMIDAL NEURONS OF THE CONTROL AND COMPRESSED SENSORIMOTOR CORTEX
Soma area (�m2) Number of dendritic trunks
Layer III Layer V Layer III Layer V
Control 148.7� 31.2 326.5� 83.2 5.7� 1.1 7.3� 1.0(n � 27) (n � 26)
Comp-3D 157.8� 29.5 314.6� 88.6 5.3� 1.0 7.6 1.6(n � 28) (n � 21)
Comp-1M 132.2� 34.3 318.5� 84 5.7� 1.1 7.5� 1.8(n � 26) (n � 24)
Comp-1Y 121� 26.6* 289.1� 87* 5.4 � 0.8 7.1� 0.9(n � 21) (n � 25)
Values are means� S.E. of the number of neurons (n) examined in each group. Ns are listed in the left half of the table only. Somaarea and number of dendritic trunks were calculated from the two-dimensionally reconstructed drawings. Please see Table 1 for abbreviations.
*p � 0.05 between the marked value and its corresponding control, 2-tailed Student’s t-test.
TABLE 4. DENDRITIC PARAMETERS OF THE LAYER III AND V PYRAMIDAL NEURONS OF THE SENSORIMOTOR CORTEX
Total dendritic length (�m) Apical dendritic length (�m) Basal dendritic length (�m)
Layer III Layer V Layer III Layer V Layer III Layer V
Control 4855� 145 8930� 494 2469� 138 5206� 371 2304� 88 3672� 149(n � 27) (n � 26)
Comp-3D 4331� 151* 6790� 343** 2038 � 117* 3540� 297** 2246 � 120 3478� 195(n � 28) (n � 21)
Comp-1M 3504� 175** 6272 � 437** 1678 � 130** 3222 � 183** 1833 � 99** 3184 � 263*(n � 26) (n � 24)
Comp-3M 3571� 269** 6336 � 616** 1878 � 154* 3421� 265** 1726 � 212** 2919 � 383**(n � 15) (n � 14)
Comp-6M 3420� 262** 6460 � 412** 1854 � 81* 3345� 250** 1754 � 170** 2804 � 183**(n � 16) (n � 13)
Comp-1Y 3385� 126** 6328 � 198** 1718 � 81** 3289 � 181** 1667 � 108** 3116 � 135*(n � 21) (n � 25)
Values are means� S.E. of the number of neurons (n) studied following intracellular dye-injection, immunohistochemistry andcamera lucida reconstruction in the two-dimensional plane. Ns are listed in the left half of the table only. Control and comp-3D, -1M, -3M, -6M, and -1Y represent sham-operated, and bead-implanted groups 3 days, 1 month, 3 months, 6 months, and 1 year fol-lowing treatment, respectively.
*p � 0.05; **p � 0.01 between the marked and its corresponding control respectively, 2-tailed Student’s t-test.

reduced starting at 3 days following compression (Table5). The spine densities of the basal dendrites, measuredat both 25–75 �m and at 100–150 �m from the somaof both neurons, were also reduced starting at 3 daysfollowing compression (Table 5). Although both layerIII and V pyramidal cells retracted their dendritic spinesstarting at 3 days following compression, modifications
on both the proximal and distal apical and basal den-drites of layer V pyramidal neurons appeared to progressgradually as the duration of compression lengthened,and might not have peaked even at 1 year followingcompression (Table 5). On the other hand, dendriticspine densities of layer III pyramidal neurons, with theexception of those on the proximal basal dendrites, hadalready reached their maximal reduction 1 month fol-lowing compression.
DISCUSSION
General Effect of Mechanical Compression
In this study, we implanted plastic beads epidurallyover the sensorimotor cortex of rats to study the effect ofphysical compression on the cerebral cortex. Althoughan implanted bead is obviously harder than many clini-cal sources of compression such as blood clot or tumormass, the intact dura and the underlying fluid-filled sub-arachnoid space in our view serve to cushion hardness orfriction that may occur during pulsation or movement.Therefore, we believe that this is a suitable model to eval-uate the effect of physical compression alone with nocomplications from chemical or weight factors, since thebead is made of inert and lightweight material. In ex-ploratory experiments, we tried particles of the same di-mension but of different thickness (1, 1.5, and 2 mm) asthe source of epidural compression on rats. We found thatimplantation of a 1-mm-thick particle resulted in no ap-parent change on the underlying cortex or any of its cor-tical layers. However, if the particle was 2-mm thick, theunderlying, but not peripheral, cerebral cortex becamenecrotic with apparent cell death beginning 3 days afterimplantation, and the cortex of the compressed area virtually disappeared in animals surviving 6 months following implantation. Particles 1.5 mm in thicknesscaused a thinning of the compressed cortex without af-fecting the surrounding cortical areas or the cortex of theopposite side. The intactness of the cortical areas pe-ripheral to the compression suggests that the implanta-tion did not cause a severe increase in the overall in-tracranial pressure, which would be expected to thin ordamage all cortical areas nonspecifically. In addition, re-sults from our ongoing experiments show that the com-pressed cortex regains thickness following the removalof the implanted bead, which argues that thinning of theaffected cortex is due to focal pressure exerted by the im-planted bead on the underlying cortical area specifically(unpublished results). The thinned compressed cortexthat we have examined thus far has remained viable bymorphological and in vitro electrophysiological standards(unpublished results) for as long as 1 year following bead
CHEN ET AL.
776
FIG. 7. Numbers of dendritic branches of layer III (A) andV (B) pyramidal neurons of the sensorimotor cortex. Data fromcontrol (sham-operated) neuron and those 3 days (3D), 1 month(1M), and 1 year (1Y) after epidural bead implantation wereplotted. Dendritic crossings on the concentric circles drawnfrom the center of soma at 50-�m increments in radii were plot-ted. Layer III pyramidal neurons of the compressed cortex hadfewer dendrites in the ranges of 50–100 �m and 300–400 �mfrom the center of the soma. Similarly, layer V pyramidal neu-ron also had reduced dendritic branches in the range of 50–100�m from the center of the soma after being subjected to thecompression. In addition, the apical dendritic trees of layer Vpyramidal neurons of the compressed cortex were significantlyshortened. Numbers of layer III and V pyramidal neurons an-alyzed were 25, 26, 26, 20 and 23, 20, 24, 23, respectively, forthe control, comp-3D, Comp-1M, and Comp-1Y, respectively.Vertical bar represents standard error of the mean. *p � 0.05,** p � 0.01 between the compressed and its corresponding con-trol, two-tailed Student’s t-test.
T5
�

implantation. Therefore, the size and especially the thick-ness of the epidural mass determine the outcome of thecortex under compression much like what has been re-ported in human patients (Zülch, 1986; Hardman, 1997).
In this series of experiments aimed at studying the plas-ticity of cerebral cortex under physical pressure overtime, we decided to use 1.5-mm-thick particles since thecompressed cortex remained viable and thus its plastic-ity could be studied. Based on the response of the cortexto the implantation of particles of different sizes, we believe that there is a fine threshold as to whether anepidural mass compressing on the cerebral cortex can af-fect cortical function. Looking at the mechanical pres-sure aspect alone, a small mass may have a negligible ef-fect on the underlying cortex. However, once the massreaches a certain thickness, it starts to cause morpholog-ical alterations of the cortex in a matter of days. A largeror fast-expanding mass is likely to cause permanent de-structive damage to the underlying cortex. On the otherhand, a mass underlying the cerebral cortex is likely toaffect the nearby cortex right away, since there is no sub-arachnoid space and cerebrospinal fluid to cushion thecompression; cortical neurons are likely to respond to thecompression quickly with alterations of their morphol-ogy, physiology, and/or even cell death. Additional in-fluences from factors released by the expanding massmay intensify this effect in clinical conditions.
In this study we demonstrated that the thicknesses ofall cortical layers directly underneath the epidurally im-planted bead were reduced; however, the degrees of theirshrinkage were different. The thicknesses of layers IIthrough VI were decreased by 40% to 55%, comparableto the shrinkage of the total cortical thickness, althoughlayer IV was reduced somewhat less, around 30%. On
the contrary, layer I was reduced in thickness by onlyaround 10% at the beginning and reached a maximum of15% reduction even at 1 year following bead implanta-tion. The resistance of layer I to compression from thesurface might be related to the fact that the layer consistsmainly of nerve fibers.
Despite this shrinkage, we found no signs of cell deathin the compressed cortical area of all the survival groupsthat we examined. In fact, the densities of neurons in lay-ers II through VI of the compressed cortex were all in-creased. Neuronal densities in layers II/III and VI wereincreased by 30–40%, which was comparable to the de-gree of compaction in these layers. The apparent absenceof neuronal loss is consistent with our finding that thedensity of capillaries in the affected cortical area was notdecreased, since ischemia would have resulted in pro-found cell death (Snyder et al., 1998). These data arguethat epidural implantation of beads of the size we adoptedis unlikely to obstruct the blood flow of the compressedcortical area, although we did not measure the cerebralblood flow of the affected area in our animals. The ab-sence of an increase in reactive microglial cells, astro-cytes, and intense NADPH-diaphorase–containing neu-rons, which would have occurred if there were any tissuedamage following insufficient blood perfusion, also sup-ports this argument. On the other hand, trial implantationof 2-mm-thick beads, which led to the necrosis of the un-derlying cortex, was likely to have compromised localcerebral blood flow.
The Effect of Mechanical Compression onPyramidal Neurons
Since most pyramidal neurons have apical dendritesdirected toward and reaching the pial surface, they are
ANATOMICAL FEATURES OF CORTEX UNDER COMPRESSION
777
TABLE 5. DENDRITIC SPINE DENSITIES OF LAYER III AND V PYRAMIDAL NEURONS OF THE SENSORIMOTOR CORTEX
Layer III pyramidal neuron Layer V pyramidal neuron
Basal dendrite Apical dendrite Basal dendrite Apical dendrite
Proximal Distal Proximal Distal Proximal Distal Proximal Distal
Control 8.7� 1.3 9.7� 1.3 9.1� 1.2 10.2� 1.5 10.2� 1.1 11.4� 1.6 10.6� 0.9 11.7� 1.1(n � 65) (n � 50) (n � 60) (n � 56) (n � 50) (n � 38) (n � 45) (n � 35)
Comp-3D 7.2� 1.6** 7.6 � 1.9** 7 � 1.6** 7.5 � 1.8** 8.8 � 1.3** 9.4 � 1.8** 9.0 � 1.4** 9.5 � 1.6**(n � 53) (n � 42) (n � 51) (n � 45) (n � 42) (n � 28) (n � 36) (n � 25)
Comp-1M 6.9� 1.4** 7.4 � 1.8** 6.7 � 1.4** 6.4 � 1.6** 7.7 � 1.6** 8.6 � 1.7** 8.4 � 1.4** 9.0 � 1.4**(n � 42) (n � 40) (n � 41) (n � 35) (n � 32) (n � 20) (n � 34) (n � 21)
Comp-1Y 5.5� 1.0** 6.7 � 1.1** 6.6 � 1.0** 7.1 � 0.7** 5.8 � 0.9 6.7� 1.0** 6.5 � 1.0** 7.8 � 1.0**(n � 35) (n � 31) (n � 36) (n � 30) (n � 31) (n � 24) (n � 36) (n � 26)
Values are means� S.E. of the numbers of spines per 10 �m of dendrites for the number of dendritic segments (n) analyzed ineach group. Please see Table 1 for abbreviations.
** p � 0.01 between the marked value and its corresponding control, 2-tailed Student’s t-test.

likely to be the neurons most affected by compressionexerted from the cortical surface. In addition, pyramidalneurons, especially those of layers III and V, are the out-put neurons of the cortex, and thus changes in them couldcause alterations in cortical function and output. In lightof these considerations, we concentrated on studying themorphology of pyramidal neurons of these two key lay-ers.
We used the fixed tissue intracellular dye injectiontechnique (Wang et al., 1996) to reveal their dendritic ar-bors for two reasons. First, since neuronal membranefixed with half the usual strength of 2% paraformalde-hyde fixative still retains the injected dye LY, the den-dritic arbors of selected individual neurons can be re-vealed with negligible background. Second, this partiallyfixed condition enabled us to conduct intracellular dyeinjection of selected cells at room temperature for a muchlonger period, and therefore more cells could be sampledin each slice (Wang et al., 2002). This also meant thatwe could collect sufficient dye-filled cells from the slicesof a limited number of animals of each group, thus min-imizing the error that might be introduced by samplingneurons from a large number of animals. Our key find-ing was that pyramidal neurons reduced their dendriticlength dramatically starting as early as 3 days, reachedmaximal reduction at 1 month, and maintained the samereduced level to at least 1 year following bead implanta-tion.
In this study, the method we used to calculate dendriticlength took the curviness of dendrites into account.Therefore, the reduction in their length probably reflectsa recycling of dendritic membrane instead of an increasein the tortuosity of these dendrites, since no dendrite ofneurons sampled from the compressed cortex appearedto be more tortuous than those of control neurons. Whenapical and basal dendrites were analyzed separately,epidural compression appeared to significantly shortenapical dendritic length starting at 3 days and reached themaximum level at 1 month following bead implantation;however, the effect on basal dendrites seemed to bemilder, proceeded slowly, and was likely to still be de-creasing 1 year following the onset of the compressionespecially when layer III pyramidal cells were consid-ered. These results might reflect the fact that apical den-drites laid closer to the source of compression than basaldendrites. Analysis with Sholl’s concentric shells (Tsengand Prince, 1996) showed that the reduction in dendriticlength was accompanied by a decrease in dendriticbranches while the number of dendritic trunks was notaltered. Therefore, the reduction in dendritic length islikely to be a consequence of the retraction of distal den-drites and/or shortening of dendrites due to membrane re-cycling. Similar reductions in the dendritic arbor of ax-
otomized spinal motor neurons (Brännström et al., 1992)and rubrospinal neurons (Tseng and Hu, 1996; Wang etal., 2002) have been attributed to the loss of distal den-dritic arbors. Regarding the mechanism involved in thereduction in dendritic length, results from our ongoingexperiments show that the apical dendritic shafts of thesepyramidal neurons become twisted or kinked starting at1 day following epidural bead implantation, and thesephenomena disappear by 3 days following implantation(unpublished results). This suggests that focal pressurelimiting the thickness of the compressed cortex is at leastone of the factors inducing the shortening of the dendritesof cortical neurons during the early phase of compres-sion.
Another important finding of the present study is thatthe dendritic spines on these pyramidal neurons of thecompressed cortex were significantly reduced. Earlierstudies showed that dendritic spines are dynamic struc-tures even under normal conditions (Horner, 1993; Segalet al., 2000) and they can change forms and densities withenvironments (Brandon and Coss, 1982; Horner, 1992).Long-term potentiation and enriched environment can in-crease dendritic spine densities (Globus et al., 1973). Onthe other hand, reduction of dendritic spines can resultfrom aging (Feldman and Dowd, 1975), injuries such asdeafferentation (Rutledge et al., 1972) and hypoxia (Kim,1975; Pokorny and Trojan, 1983), and pathological con-ditions such as dementia (Ferrer et al., 1990), temporallobe epilepsy (Olney et al., 1983; Paul and Scheibel,1986), and mental retardation states (Marin-Padilla,1972, 1974, 1976). In addition, earlier morphologicalstudy on the epitumorous human cerebral cortex havedocumented the loss of dendritic spines on neurons nearthe tumor mass (Spacek, 1987). In the literature, alter-ations of dendritic spine density have been shown to oc-cur very fast; the spine density of rat hippocampal den-tate granule cells was increased within 6 h followingwater maze training (O’malley et al., 2000), while its re-duction was shown to occur within 3 days following acutestatus epilepticus induced by pilocarpine (Isokawa,2000).
In the present study, we found that the spine densitieson both apical and basal dendrites were already signifi-cantly decreased by 15–20% at 3 days following beadimplantation, irrespective of how far away they werefrom the source of compression. This suggests a quickonset of spine retraction, which must have occurred ear-lier. In addition to an instantaneous decrease, retractionof dendritic spines progressed continuously, and spineloss increased to 20–30% at 1 month and to 30–40% at1 year after implantation. Since the onset of the spine lossappeared to be earlier than the first time point that weexamined in the present study, the issue of whether spine
CHEN ET AL.
778

retraction on apical dendrites starts earlier than on basaldendrites remains to be explored. In this study, althoughwe found that dendrites could have reduced part of theirlength by retracting their distal parts following compres-sion from the cortical surface, the extent of spine retrac-tion on the distal dendritic segments of either basal orapical dendrites was not different from that on their prox-imal counterparts.
Recent study on live cells shows that spine densitiescan be maintained by AMPA receptor activation even atthe spontaneous synaptic vesicle release level (McKin-ney et al., 1999; Andersen and Soleng, 1999). This sug-gests that dendritic spines are maintained by their func-tion but not the locality of where they reside on thedendrites. Our finding that the degree of spine loss on theproximal and distal portions of any given dendrite wasabout the same suggests that the function of spines allover the dendrites of these cells could have been com-promised. This could have been caused by the disabilityof either the release of glutamate-containing synapticvesicles by presynaptic terminals or the function of post-synaptic AMPA receptors associated with the synapsesof these spines.
The gradual loss of dendritic spines after a quick on-set of spine reduction suggests that some synapses mightlose their activity at different time points following beadimplantation. Compression-induced conduction block ofnerve fibers (Golan and Grossman, 1992) traveling to-ward these neurons could be one explanation. With thisscenario, nerve fibers closer to the cortical surface, es-pecially those in layer I or synapses on apical dendrites,would be expected to be affected more seriously thanthose on basal dendrites. On the other hand, the effect ofcompression might have reached the threshold needed tosilence a large number of synapses both in the superfi-cial and deep layers of the underlying cortex, so thatspines in the deep or superficial parts were all affectedsimilarly and/or simultaneously. The possibility thatspine loss could have resulted from the degeneration ofsome nerve fibers in the compressed cortex followingbead implantation seems to be unlikely since no appar-ent increase in microglial cells was identified in the com-pressed cortical area.
In conclusion, we developed a rat model of epiduralcortical compression that appears not to interfere with lo-cal cerebral blood flow or cerebrospinal fluid circulation.The model in no way simulates meningioma or hematomafor many factors such as alteration of blood-brain bar-rier, vascular compression, and secretion of edemato-genic factors, which could all be involved in many clin-ical cases. It nevertheless allows us to isolate the effectof the mechanical pressure exerted by a mass compress-ing on cerebral cortex. Our model shows that epidural
mechanical compression by itself resulted in structuralalteration of the underlying cerebral cortex in a matter ofdays. In response to the compression, layer III and Vpyramidal neurons, the major output neurons of the cere-bral cortex, trimmed their dendritic length and dendriticspines. This is likely to occur at a time when corticalfunction is compromised, as evidenced in human patientssubjected to early phase meningioma or light hematoma(Zülch, 1986; Hardman, 1997), although it didn’t seemto affect our animals behaviorally, at least at the grosslevel. It will be of profound interest to discover whetherthe cortex regains its thickness and pyramidal neurons re-grow their dendrites and dendritic spines following theremoval of the implanted bead, because surgical removalof an epidural mass often lead to rather marked recoveryof the compromised functions in human patients (Kolb,1995).
ACKNOWLEDGMENTS
The study was supported by grants from the NationalScience Council of Taiwan NSC-91-2320-B002-112 andNSC 91-2320-B-320-013 to G.-F.T. and Y.-J.W.
REFERENCES
ANDERSEN, P., and SOLENG, A.F. (1999). A thorny ques-tion: how does activity maintain dendritic spines? NatureNeurosci. 2, 5–7.
BRANDON J.G., and COSS, R.G. (1982). Rapid dendriticspine stem shortening during one-trial learning: the honey-bee’s first orientation flight. Brain Res. 252,51–61.
BRÄNNSTRÖM, T., HAVTON, L., and KELLERTH, J.O.(1992). Change in size and dendritic arborization patterns ofadult cat spinal alpha-motoneurons following permanent ax-otomy. J. Comp. Neurol. 318,439–451.
FELDMAN, M.L., and DOWD, C. (1975). Loss of dendriticspines in aging cerebral cortex. Anat. Embryol. 148,279–301.
FERRER, I., GUIONNET, N., CRUZ-SANCHEZ, F., et al.(1990). Neuronal alterations in patients with dementia: aGolgi study on biopsy samples. Neurosci. Lett. 114,11–16.
GLOBUS, A., ROSENZWEIG, M.R., BENNETT, E.L., et al.(1973). Effects of differential experience on dendritic spinecounts in rat cerebral cortex. J. Comp. Physiol. Psychol.82,175–181.
GOLAN, H., and GROSSMAN Y. (1992) Synaptic transmis-sion at high pressure: effects of [Ca2�]o. Comp. Biochem.Physiol., A. Comp. Physiol. 103,113–118.
HARDMAN, J.M. (1997). Cerebrospinal trauma, in: Textbook
ANATOMICAL FEATURES OF CORTEX UNDER COMPRESSION
779

of Neuropathology. R.L. Davis and D.M. Roberton (eds),Williams & Wilkins: Baltimore, pps. 1179–1231.
HORNER, C.H. (1993). Plasticity of the dendritic spine. Prog.Neurobiol. 41, 281–321.
ISOKAWA, M. (2000). Remodeling dendritic spines of dentategranule cells in temporal lobe epilepsy patients and the ratpilocarpine model. Epilepsia 41 (Suppl. 6),S14–S17.
KIM, S.U. (1975). Brain hypoxia studied in mouse central ner-vous system cultures. I. Sequential cellular changes. Lab. In-vestigat. 33, 658–669.
KOLB, B. (1995). Is there really recovery from brain damage?,in: Brain plasticity and Behavior. Lawrence Erlbaum Asso-ciates: New Jersey, pps. 37–66.
MARIN-PADILLA, M. (1972). Structural abnormalities of thecerebral cortex in human chromosomal aberrations. A Golgistudy. Brain Res. 44, 625–629.
MARIN-PADILLA, M. (1974). Structural organization of thecerebral cortex (motor area) in human chromosomal aberra-tions. A Golgi study. I. D1(13–15) trisomy, Patau syndrome.Brain Res. 60, 375–391.
MARIN-PADILLA, M. (1976). Pyramidal cell abnormalities inthe motor cortex of a child with Down’s syndrome. A Golgistudy. J. Comp. Neurol. 167,63–82.
MCKINNEY, R.A., CAPOGNA, M., DÜRR, R., et al. (1999).Miniature synaptic events maintain dendritic spines viaAMPA receptor activation. Nature Neurosci.2, 44–49.
OLNEY, J.W., DE GUBAREFF, T., and SLOVITER, R.S. (1983).“Epileptic” brain damage in rats induced by sustained electricalstimulation of the perforant path. II. Ultrastructural analysis ofacute hippocampal pathology. Brain Res. Bull. 10,699–712.
O’MALLEY, A., O’CONNELL, C., MURPHY, K.J., et al.(2000) Transient spine density increases in the mid-molecu-lar layer of hippocampal dentate gyrus accompany consoli-dation of a spatial learning task in the rodent. Neuroscience99, 229–232.
O’RAHILLY, R., and MULLER, F. (1986). The meninges inhuman development. J. Neuropathol. Exp. Neurol. 45,588–607.
PAUL, L.A., and SCHEIBEL, A.B. (1986). Structural sub-strates of epilepsy. Adv. Neurol. 44, 775–786.
POKORNY, J., and TROJAN, S. (1983). Chronic changes inthe receptive field of the pyramidal cells of the rat hip-pocampus after intermittent postnatal hypoxia. Physiol. Bo-hemoslov. 32, 393–402.
REIFENBERGER, G., BOSTROM, J., BETTAG, M., et al.(1996). Primary glioblastroma multiforme of the oculomotornerve. J. Neurosurg. 84, 1062–1066.
RUTLEDGE, L.T., DUNCAN, J., and CANT, N. (1972).Long-term status of pyramidal cell axon collateral and api-
cal dendritic spines in denervated cortex. Brain Res. 41,249–262.
SEGAL, M., KORKOTIAN, E., and MURPHY, D.D. (2000).Dendritic spine formation and pruning: common cellularmechanisms? Trend Neurosci. 23, 53–57.
SNYDER, S.H., JAFFREY, S.R., and ZAKHARY, R. (1998).Nitric oxide and carbon monoxide: parallel roles as neuralmessengers. Brain Res. Rev. 26, 167–175.
SPACEK, J. (1987). Ultrastructural pathology of dendriticspines in epitumorous human cerebral cortex. Acta Neu-ropathol. 73, 77–85.
TSENG, G.-F., and HU, M.-E. (1996). Axotomy induces re-traction of the dendritic arbor of adult rat rubrospinal neu-rons. Acta Anat. 155,184–193.
TSENG, G.-F., and PRINCE, D.A. (1993). Heterogeneity of ratcorticospinal neurons. J. Comp. Neurol.335,92–108.
TSENG, G.-F., and PRINCE, D.A. (1996). Structural and func-tional alterations in rat corticospinal neurons following axo-tomy. J. Neurophysiol. 75, 248–267.
TSENG, G.-F., and ROYCE, G.J. (1986). A Golgi and ultra-structural analysis of the centromedian nucleus of the cat. J.Comp. Neurol. 245,359–378.
TSENG, G.-F., PARADA, I., and PRINCE, D.A. (1991). Dou-ble-labelling with rhodamine beads and biocytin: a techniquefor studying corticospinal and other projection neurons invitro. J. Neurosci. Methods 37, 121–131.
TSENG, G.-F., WANG, Y.-J., and LAI, Q.-C. (1996). Per-ineuronal microglial reactivity following proximal and distalaxotomy of rat rubrospinal neurons. Brain Res. 715,32–44.
WANG, Y.-J, LIU, C.-L., and TSENG, G.-F. (1996). Com-partmentation of calbindin and parvalbumin in different partsof rat rubrospinal neurons. Neuroscience 74, 427–434.
WANG, Y.-J., CHEN, J.-R., and TSENG, G.-F. (2002). Fatesof soma and dendrites of cord-projection central neurons fol-lowing proximal and distal spinal axotomy: an intracellulardye injection study. J. Neurotrauma 19, 1487–1502.
ZÜLCH, K.J. (1986). Tumor and brain, in: Brain Tumors: TheirBiology and Pathology,3rd ed. Springer-Verlag: Berlin, pps.154–189.
Address reprint requests to:Guo-Fang Tseng, Ph.D.
Department of Anatomy and Cell BiologyCollege of Medicine
National Taiwan University, No.1, Section 1Jen-Ai Rd.
Taipei, Taiwan, 100
E-mail: [email protected]
CHEN ET AL.
780