the effect of high-fructose corn syrup on ethanol taking
TRANSCRIPT

The Effect of High-Fructose Corn Syrup on Ethanol Taking, Seeking, and Palatability in Rats
by
Samantha Ayoub
A Thesis Presented to
The University of Guelph
In partial fulfilment of requirements for the degree of
Master of Science in Psychology + Neuroscience
Guelph, Ontario, Canada
© Samantha Ayoub, August, 2018

ii
ABSTRACT
THE EFFECT OF HIGH-FRUCTOSE CORN SYRUP ON ETHANOL TAKING, SEEKING AND PALATABILITY IN RATS
Samantha Ayoub Advisor: University of Guelph, 2018 Dr. Francesco Leri
Many alcoholic beverages contain sweeteners, such as high-fructose corn syrup
(HFCS), which has been implicated in increasing alcohol consumption and encouraging
hazardous drinking patterns. Therefore, four experiments were designed to investigate
whether the addition of HFCS to an ethanol (EtOH) solution could alter its intake,
hedonic taste, and sensitivity to food deprivation stress in Sprague Dawley rats.
Experiments 1 and 2 revealed that HFCS caused a robust increase in EtOH intake
during operant intraoral self-administration (IO SA), however an equally palatable, yet
non-caloric sweetener (saccharin) could not replicate this effect. This suggests the
caloric value of a sweetener mediates the effect of sweeteners on EtOH intake.
Experiments 3 and 4 revealed food deprivation stress increased EtOH consumption
during resumption following extinction, but only when HFCS was added to the solution.
The palatability of 10% EtOH and 25% HFCS was also monitored across IO SA, and
under conditions of food deprivation stress. It was found that the palatability of 10%
EtOH and 25% HFCS increased following acquisition. More interestingly, food
deprivation stress only increased HFCS palatability. Thus, it could be the heightened
palatability of HFCS under stressful conditions that contributes to increased operant
responding for solutions containing HFCS during resumption in rats. These results have
implications for the use of highly palatable and caloric sweeteners in EtOH beverages
because of the impact they may have on excessive EtOH intake.

iii
ACKOWLEDGEMENTS
I would like to thank my advisor, Dr. Francesco Leri, for the opportunity to
complete my thesis in an astounding laboratory. Your mentorship and guidance
throughout this process helped me develop into a better scientific researcher and
academic writer. Your intellect and passion for this field of work is admirable to say the
least.
I would like to thank my thesis committee member, and practicum advisor, Dr.
Linda Parker. It has been my pleasure to work in your lab, even if so briefly.
Additionally, your support and mentorship throughout my thesis was well recognized,
and I cannot thank you enough. Lastly, I would like to thank the members of the Parker
laboratory who helped train me and made me feel welcome; especially Erin Rock.
I would like to thank all my labmates for their continued support and
encouragement throughout the past two years. I could not have asked for a better group
of individuals to work with. In particular, I would like to thank Meenu Minhas for training
me and getting me started up in the Leri laboratory. I will truly miss contacting you when
I am stuck in a rut and am not sure what to do next. I appreciate all your help so much.
Finally, I would like to thank my family and friends. You have all been so positive
and supportive throughout my entire academic career. I love you all so much.

iv
Table of Contents ABSTRACT…………………………………………………………………………………… ii Acknowledgements…………………………………………………………………………… iii List of Figures…………………………………………………………………………………. v General Introduction………………………………………………………………………….. 1 Manuscript………………………………………………………………………………......... 20 Abstract……………………………………………………………............................ 21 Introduction……………………………………………………………………………. 22 Methods………………………………………………………………………………. 25 Statistical analyses…………………………………………………………….......... 32 Results………………………………………………………………………………… 32 Discussion…………………………………………………………………………….. 36 References……………………………………………………………………………. 43 Figure legends……………………………………………………………………….. 53 Figures……………………………………………………………………………….. 56 General Discussion………………………………………………………………………….. 60 Summary of results………………………………………………………………….. 60 Methodological considerations…………………………………………………….. 60 Future directions…………………………………………………………………….. 62 Conclusion…………………………………………………………………………… 64 References…………………………………………………………………………………… 66

v
LIST OF FIGURES Figure 1……………………………………………………………………………………….. 56 Figure 2……………………………………………………………………………………….. 57 Figure 3……………………………………………………………………………………….. 58 Figure 4……………………………………………………………………………………….. 59

1
General rationale and summary of experiments
Heavy episodic drinking is prevalent in youth and has been shown to elevate the
risk of developing an alcohol use disorder later in life (Chassin, Pitts, & Prost, 2002).
Furthermore, youth who display such drinking patterns prior to the age of 18 are less
likely to seek help, and have increased risks for chronic relapsing following treatment
(Hingson, Heeren, & Winter, 2006). It has been suggested that the use of sweeteners in
alcoholic beverages reduces the aversive taste of alcohol in naïve drinkers, and leads to
increased consumption (Mosher & Johnsson, 2005; Lanier, Hayes, & Duffy, 2005;
Romanus, 2000; Copeland, Stevenson, Gates, & Dillon, 2007). In fact, youth that
primarily consume sweetened alcoholic beverages have four times the risk of
experiencing alcohol-related injuries and report a higher frequency of alcohol binges
(Albers, Siegel, Ramirez, Ross, DeJong, & Jernigan, 2015). Although this may seem
intuitively obvious, the mechanisms by which sweeteners increase alcohol intake are
not fully understood. More specifically, sweeteners not only change the taste of alcohol,
but they also support reliable effects on behaviour and the brain. Thus, sweeteners also
have the potential to synergistically alter alcohol’s reinforcing value because they elicit
approach and conditioned behaviour (Yasoshima & Shimura, 2015; Sclafani & Ackroff,
2017; White & Carr, 1985; Agmo & Marroquin, 1997), and maintain high levels of
operant self-administration (Levy et al., 2015; Avena, Long, & Hoebel, 2005). Recently,
our laboratory has explored the behavioural and neurobiological responses to high-
fructose corn syrup (HFCS) in rats (Levy et al., 2014, Levy et al., 2015; Daniels,
Marshall & Leri, 2016; Levy et al., 2018). In comparison to sucrose (which contains 50%
glucose and 50% fructose), HFCS has a higher proportion of fructose (typically, 55%

2
fructose), and has been linked to several adverse health effects including obesity, heart
disease, and poor metabolic health (Meyers, Mourra, & Beeler, 2017; Bray, 2013;
Stanhope et al., 2015). The findings from our lab suggest that HFCS has strong
reinforcing properties, and could possibly be a more potent behavioural reinforcer than
sucrose. HFCS is highly prevalent in our diets, and can be found in many of the
sweetened alcoholic beverages available for purchase (e.g. Palm Bay, Bacardi Silver,
Smirnoff Ice, Mike’s Hard Lemonade). Thus, the current experiments were designed to
investigate the effect of HFCS on ethanol (EtOH) intake, palatability, and persistence of
operant responding in Sprague Dawley rats.
Experiment 1 used an operant intraoral self-administration (IO SA) technique to
explore whether HFCS could alter EtOH intake. It was found that HFCS produced
robust increases in IO SA behaviour and EtOH intake.
In Experiment 2, a similar experimental design was used to assess the effect of an
equally palatable, yet non-caloric sweetener (saccharin; SACC) on EtOH intake. It was
found that SACC increased EtOH intake, however this effect was minimal in comparison
to what was observed by the addition of HFCS. These results suggest that the caloric
value of a sweetener powerfully mediates the effect of sweeteners on EtOH intake in
rats.
Experiment 3 tested the persistence of operant responding following behavioural
extinction in rats trained to self-administer 10% EtOH in 25% HFCS. This experiment
also included a stress manipulation, because stress is known to alter the reinforcing
effects of drugs of abuse and facilitate drug-seeking in many species (Shaham, Erb, &
Stewart, 2000). The stress selected was food deprivation because it has been shown to

3
have potent effects on animal drug-seeking behaviours (Shalev, Yap, & Shaham, 2001;
Carroll, 1985) and because of the relevance to the behaviour being explored (i.e.,
ingestion of calories from both EtOH and HFCS). It was found that food deprivation
stress increased persistent operant responding for EtOH, only when it was mixed in
HFCS.
In Experiment 4, EtOH and HFCS taste reactivity was measured throughout the IO
SA of EtOH + HFCS to assess how the palatability of these solutions are altered over
subsequent exposure periods, and under conditions of food deprivation stress. This
revealed two key findings. First, rats produced more palatable responses to EtOH and
HFCS following IO SA acquisition. Second, food deprivation stress increased palatable
responding to HFCS, but did not alter responding to EtOH. Thus, it could be the
increased palatability of HFCS under stressful conditions that contributes to the
increased operant responding for solutions containing HFCS during resumption
following extinction in rats. In other words, the motivation to consume an EtOH
beverage under normal or stressful conditions can be increased by the addition of
HFCS.
Collectively, these results have implications for the use of highly palatable and
caloric sweeteners in EtOH beverages because of the impact they have on EtOH intake
in naïve drinkers. Below, I will sequentially review the key aspects of my research.
Heavy Episodic Drinking in Youth Increases the Risk of AUD Development
Heavy episodic drinking refers to the consumption of 60 or more grams of ethanol
within a single occasion, occurring at least once per month (World Health Organization,
2014). This quantity of alcohol roughly equates to 5 standard drinks for men, and 4

4
standard drinks for women. Worldwide, heavy episodic drinking patterns are the most
prevelant in young and often underaged drinkers (World Health Organization, 2014). A
nationwide Canadian survey revealed 14.7% of youth aged 15-17 reported these
consumption patterns, while this number increased to 33.4% between respondents
aged 18-19 (Archie, Zangeneh-Kazemi, & Akhtar-Danesh, 2012). Furthermore, the US
National Institute on Alcohol Abuse and Alcoholism (2015) estimates 1.45 million youth
aged 12 to 17 ‘binge’ on alcoholic beverages at least once per month. These statistics
are particularly concerning because young drinkers characterized as ‘bingers’ have an
exceedingly elevated risk for subsequent alcohol use disorders (AUDs) compared to
‘non-binger’ youths (Chassin, Pitts, & Prost, 2002). Furthermore, early onset of alcohol
drinking is correlated with increased risks of subsequent alcohol misuse and associated
problem behaviours in adulthood (Gruber, DiClemente, Anderson, & Lodico, 1996;
Higson, Heeren, & Winter, 2006; DeWit, Adlaf, Offord, & Ogborne, 2000; Hawkins,
Graham, Maguin, Abbott, Hill, & Catalano, 1997). Thus, reducing excessive alcohol
consumption in young, naïve drinkers could be an effective strategy to mitigate the high
prevalence of disordered drinking worldwide.
Alcohol Use Disorders
Alcohol use disorders increase many health risks for the abuser and place a heavy
burden on healthcare systems worldwide. In fact, the World Health Organization (2009)
reported alcohol use as the third leading risk factor contributing to the global burden of
disease. Heavy alcohol consumption is causally linked to liver and heart disease, birth
defects, cancer, and unintentional injuries (Rehm et al., 2009). Additionally, AUDs are
comorbid with a wide range of psychiatric disorders, including, anxiety disorders, major

5
depressive disorders, post-traumatic stress disorder, substance use disorders, and
schizophrenia (Petrakis, Gonzalez, Rosenheck, & Krystal, 2002; Regier et al., 1990).
Despite the negative consequences associated with excessive alcohol consumption, the
prevalence rate for AUDs remains high at approximately 4% worldwide (The World
Health Organization, 2014).
(1) Chronic Relapse
The high occurrence of AUDs is partially attributable to the inability of abusers to
maintain abstinence following alcohol detoxification and in-patient treatment programs.
Studies reveal high relapse rates following both short-term (Moos & Moos, 2006) and
long-term remission periods (Jin, Rourke, Patterson, Taylor, & Grant, 1998), signifying
that a relapse to excessive alcohol consumption can be triggered regardless of long-
term periods of abstinence. For example, Jin et al., (1998) measured relapse rates in
long-term abstainers, defined as individuals with 18-months of stable abstinence, and
found 31% had relapsed prior to an 11-year follow-up period.
The high and frequent occurrence of relapses during recovery has led to the
recognition of AUDs as chronic relapsing disorders. Given that the recovery from an
AUD is a long and sometimes impossible road, preventative measures may be an
effective method to reduce AUD prevalence. That is, preventing disordered drinking in
naïve drinkers may reduce the incidents of AUDs.
(2) Triggers of Relapse
Identifying AUDs as chronic relapsing disorders has drawn attention to the
identification of factors that provoke a relapse, and the neural correlates that underlie
this behaviour. Exposure to alcohol-paired stimuli is one determinant of alcohol relapse

6
following abstinence (Perry, Zbukvic, Kim, & Lawrence, 2014). For example, detoxified
alcoholics experience increased alcohol craving, and physiological responses, when
exposed to alcohol-paired stimuli in a laboratory setting (Garland, Franken, & Howard,
2012; Cooney, et al., 1997). Furthermore, the magnitude of post-treatment reactivity to
alcohol-paired stimuli accurately predicts future relapses (Garland et al., 2012). These
findings suggest alcohol-paired stimuli possess incentive salience, acquired through
repetitive pairings with alcohol’s reinforcing pharmacological properties. That is, through
associative learning, alcohol-paired stimuli become conditioned cues, or conditioned
reinforcers (CRs), capable of eliciting a craving response and motivating behaviour.
Given the taste of alcohol is prominent during consumption, alcohol-paired taste cues
may act as CRs to motivate alcohol consumption behaviour. Indeed, exposure to the
taste of beer significantly increases alcohol craving in heavy drinkers when compared to
other flavoured controls (i.e. Gatorade; Oberlin et al., 2013). Additionally, neuroimaging
studies reveal that the taste of alcohol activates mesocorticolimbic structures in the
brain (Filbey et al., 2008), and increases dopamine (DA) release in these areas (Oberlin
et al., 2013). Given that mesolimbic DA signaling is implicated in reward, motivation
(see Robins & Everitt, 1996 for a review) and goal-directed behaviour (Wise & Rompre,
1989; Horvitz, 2000), these findings suggest alcohol-paired tastes can drive appetitive
behavioural responding for alcohol, and perhaps contribute to relapse following
abstinence.
Exposure to stress is another major determinant of relapses following AUD treatment
(Preston & Epstein, 2011). For example, the occurrence of severe stress following
treatment is correlated with increased relapse risk (Brown, Vik, Patterson, Grant, &

7
Schuckit, 1995; Brown et al., 1990). Brown et al. (1990) found alcoholics who had
relapsed during a three-month treatment follow-up had self-reported significantly more
severe stress during the study period than those who remained abstinent. Additionally,
stress induces alcohol cravings in abstinent alcoholics (Preston & Epstein, 2011). Fox,
Bergquist, Hong, and Sinha, (2007) used a stress imagery technique to induce anxiety
in former alcoholics within a laboratory setting. They found stress imagery significantly
increased alcohol craving, as well as measures of blood pressure, in comparison to a
neutral imagery condition. Similar to cue-induced craving, the magnitude of laboratory
stress-induced alcohol craving can also predict abstinence following treatment (Sinha et
al., 2011; Higley et al., 2011). For example, Higley et al. (2011) demonstrated greater
laboratory stress-induced craving was associated with a significantly shorter duration of
alcohol abstinence, fewer total days of abstinence, and a higher mean number of drinks
per week following treatment.
In sum, both exposure to alcohol-paired stimuli acting as CRs, and stress, have been
implicated in provoking alcohol relapse following AUD treatment. The data presented is
largely derived from human laboratory studies of alcohol-induced craving, or from
correlative measures of CR/stress exposure on subsequent relapse rates. Animal
models of relapse can causally measure the influence of CRs and stress on relapse-like
responding, thereby strengthening the assumptions made regarding CR and stress
exposure on relapse vulnerability. Operant drug self-administration (SA) procedures
model “drug-taking” and “drug-seeking” in animals, and are used to analyze the factors
that contribute to drug addiction, including relapse-like responding (Panlilio & Goldberg,
2007).

8
Measuring Drug-Intake in Animals: Operant Self-Administration Models
The fundamental assumptions used by the operant SA model date back to the early
works of Thorndike (1898) who proposed behaviours that produce favourable outcomes
are more likely to be repeated than behaviours producing non-favourable outcomes.
Using this theoretical framework, operant SA procedures allow an animal to perform a
behavioural response for the administration of drug reinforcement; thus, if the drug is
indeed a reinforcing substance, the operant response which led to its administration
should continue to be repeated.
In rodents, operant drug SA models are conducted in operant conditioning chambers
that are typically equipped with an active and inactive lever (O’Conner, Chapman,
Butler, & Mead, 2011). Responding on the active lever (a lever-press) results in the
delivery of drug reinforcement, thereby producing a measure of drug-reinforced
responding, or “drug-taking”. A lever-press on the inactive lever results in no
programmed consequences, and can therefore be used as a control measure of non-
reinforced operant responding. Measures of drug intake (number of drug infusions) or
operant responding (active versus inactive lever responding) are used as indications of
SA behavior, and help researchers make inferences about the drug’s abuse potential.
(1) Extinction-Reinstatement Procedures
Extinction-reinstatement procedures stem from operant drug SA models, and
measure “drug-seeking” in animals (Panlilio & Goldberg, 2007; Shaham, Shalev, Lu,
deWitt, & Stewart, 2002). Following initial acquisition of SA, extinction of the drug-taking
behaviour is accomplished by removing response-contingent drug reinforcement. As a
result, responding under extinction conditions declines rapidly as the behaviour is no

9
longer being reinforced (Katner, Magalong, & Weiss, 1999). Thus, any behavioural
responding in the absence of drug reinforcement is a measure of “drug-seeking” rather
than drug-taking. Once drug-seeking is extinguished, a reinstatement test can be used
to delineate the factors that provoke a reinstatement of the extinguished responding (i.e.
drug-seeking) for that drug. Therefore, extinction-reinstatement procedures can
measure relapse-like responding following a period of abstinence and are invaluable
tools in drug addiction research (Panlilio & Goldberg, 2007; Martin & Weiss, 2013).
In corroboration with the human literature on relapse vulnerability, exposure to
alcohol-paired stimuli reinstates extinguished alcohol-seeking in rats. For example, re-
exposure to drug-taking environments potently reinstates previously extinguished drug-
taking behaviour (see Crombag, Bossert, Kova, & Shaham, 2008 for a review).
Chaudhri, Sahuque, Cone and Janak (2008) trained rats to self-administer alcohol in an
environmental context distinguished by distinctive visual, olfactory and tactile stimuli.
Subsequently, alcohol-taking behaviour was extinguished in a separate and
perceptually altered context. During reinstatement, animals were given a small oral
alcohol prime (i.e. re-exposure to oral alcohol) in the drug-taking context, or in the
extinction context. Reinstated responding for alcohol was observed when rats were
primed in the alcohol-taking context but not in the extinction context; thus,
demonstrating alcohol-paired contextual stimuli modulate behavioural responding for
alcohol after periods of abstinence.
The taste of alcohol, or flavours paired with alcohol, also hold conditioned reinforcing
effects that affect alcohol-seeking behavior in rats. For example, oral alcohol primes
(Bienkowski, Koros, Kostowski, & Bogucka-Bonikowska, 2000; Maccioni et al., 2007)

10
and alcohol-paired taste cues (Knight et al., 2016) reinstate responding for alcohol
following behavioural extinction. Knight et al. (2016) paired a distinct flavour with alcohol
during SA acquisition to create an excitatory conditioned stimulus (CS+). Therefore, the
CS+ signaled the presence of alcohol reinforcement while rats acquired and stabilized
alcohol-taking behaviour. Subsequently, a different flavour was paired with water during
an extinction phase to create an inhibitory conditioned stimulus (CS-), which served to
signal the absence of alcohol reinforcement. During a reinstatement session, rats were
placed back into the conditioning chambers and allowed to self-administer either the
CS+, CS- or a neutral taste stimulus (CS0) in water. Lever responding was significantly
elevated in the CS+ group when compared to extinction baseline responding,
demonstrating a flavour previously paired with alcohol reinstated extinguished alcohol-
seeking behaviour in rats. Moreover, there was no reinstatement of lever responding
found in the CS- or CS0 group. Taken together, these studies support the human
literature that suggests the conditioned reinforcing properties of alcohol-paired cues,
such as taste, provoke appetitive behavioural responding for alcohol following periods of
abstinence.
Exposure to stress also reinstates alcohol-seeking behaviour in rats using the
extinction-reinstatement model. Lê, Harding, Juzutsch, Funk and Shaham (2005)
induced a stressed-state in rats by exposing animals to 10-minutes of intermittent
footshock (0.8 mA). Footshock stress immediately preceding a reinstatement test
potently increased alcohol-seeking behaviours following extinction. Moreover, stress
may interact with the effect of conditioned cue exposure to potentiate alcohol-seeking
behaviours. For example, Liu and Weiss (2002) demonstrated a brief exposure to

11
footshock stress exacerbated alcohol-seeking behaviours elicited by conditioned cue
exposure in comparison to alcohol-seeking behaviours elicited by conditioned cue
exposure alone.
(2) Extinction-Reacquisition Procedures
Extinction-reacquisition procedures also stem from SA models to measure the
persistence of drug-taking following the extinction of this behaviour. During reacquisition
testing, response reinforcement is restored, making it possible to explore the capacity
for an animal to relearn extinguished operant responding. Typically, the reacquisition of
extinguished operant drug-reinforced responding, otherwise called resumption, is more
rapid in comparison to the original acquisition (Bouton, Winterbauer, & Todd, 2012;
Willcocks & McNally, 2013). This suggests that the extinction of a reinforced behaviour
does not equate with the unlearning of previously reinforced responding. Moreover,
extinction-reacquisition procedures also support the notion that exposure to CRs and/or
stress exacerbates relapse-like responding. For example, Yu, Chen and Sharp (2014)
demonstrated exposure to restraint stress during a period of abstinence exacerbated
the reacquisition of nicotine self-administration when animals were reintroduced to the
operant chambers.
In sum, the same factors that have been implicated to provoke relapse in human
populations (i.e. alcohol-paired cues, stress) also reinstate extinguished alcohol-seeking
and alcohol-taking behaviours in rats using animal models of relapse. This signifies the
translational efficacy of using operant alcohol SA procedures in animals to make
inferences about human alcohol relapse behaviour. Furthermore, by producing alcohol-
taking and alcohol-seeking behaviour in animal models, the neurobiology of this

12
responding can be investigated by pharmacological manipulation of different
neurotransmitter systems. Understanding the neural mechanisms underlying alcohol-
taking and alcohol-seeking behaviour will assist in the development of therapeutic
treatment options for AUD patients who experience chronic relapses following
treatment.
Food Deprivation Stress
Food deprivation increases behavioural and neurobiological biomarkers of stress in
humans and animals. For example, the magnitude of dietary restriction is associated
with increased urinary cortisol excretion (McLean, Barr, & Prior, 2001) and elevated
salivary cortisol levels (Anderson, Shapiro, Lundgren, Spataro, & Frye, 2002) in healthy
adults. Rodent studies corroborate these findings by showing food deprivation increases
corticosterone levels (Nowland, Hugunin, & Rogers, 2011; Dietze, Lees, Fink, Brosda &
Voigt, 2016). Thus food deprivation elicits a stress response that could produce reward-
seeking behaviour in humans and animals.
In humans, being in a food deprived state is associated with increases in motivation
to seek rewarding stimuli. For example, Leeman, O’Malley, White, and McKee (2010)
found that daily smokers who participated in a 12-hour food deprivation period smoked
more cigarettes during the study period and reported stronger cravings for nicotine than
participants who were satiated. Moreover, studies support that food deprivation
increases a healthy individual’s persistence of behavioural responding for rewarding
stimuli (Hogenkamp, Shechter, St-Onge, Sclafani, & Kissileff, 2017; Bulik & Brinded,
1994). For example, food deprivation has been found to increase the amount of
behavioural responding for sweetened beverages (Hogenkamp, Shechter, St-Onge,

13
Sclafani, & Kissileff, 2017; Bulik & Brinded, 1994) and cigarettes (Bulik & Brinded,
1994).
In animal models, food deprivation stress reliably increases drug-taking and
drug-seeking behaviours. For example, 24- and 48-hour food deprivation has been
shown to potently increase heroin-seeking and cocaine-seeking in rodents (Shalev,
Yap, & Shaham, 2001; D’Cunha, Sedki, Macri, Casola, & Shalev, 2013; Tobin,
Newman, Quinn, Shalev, 2009; Carroll, 1985; Highfield, Mead, Grimm, Rocha, &
Shaham, 2002). Furthermore, animal researchers frequently use food deprivation as a
reliable method to increase the intake of various drugs of abuse (Carr, 2007).
The Self-Administration of Alcohol in Rats
Rats will self-administer alcohol intravenously (IV; Smith & Davis, 1974; Gass &
Olive, 2007), intragastrically (IG; Waller, McBride, Gatto, Lumeng, & Li, 1984; Fidler &
Clews, 2006) or orally (Chaudhri et al., 2008). IV and IG self-administration are useful
tools in studies of drug addiction, however their use in alcohol-related research is limited
by the lack of face-validity provided when applying their findings to the prototypical oral
consumption patterns found in humans. Additionally, IG and IV methods by-pass
important orosensory aspects of alcohol consumption (such as smell and taste), which
as previously mentioned, have been demonstrated to modulate alcohol craving in
humans (Oberlin et al., 2013) and alcohol-seeking behaviours in rats (Katner,
Magalong, & Weiss, 1999; Maccionia et al., 2007; Knight et al., 2016).
Although oral alcohol SA in rats can be established, there are procedural
complications with the existing system which delivers alcohol reinforcement using a fluid
receptacle (Fl-R). More specifically, a dipper-cup raises a small quantity (0.1 ml) of an

14
alcohol solution from a fluid reservoir, into a trough within the operant conditioning
chamber (Chaudhri, et al., 2008; Maccioni et al., 2007). The dipper-cup presents the
alcohol solution for a predetermined time interval, and then recedes back into the Fl-R
until the next reinforcement is delivered. Therefore, this approach requires an animal to
perform the operant response, and then move to the Fl-R to consume the alcohol
reinforcer. As such, the Fl-R delivery approach poses several caveats for measuring
operant alcohol SA responding.
One limitation of the Fl-R delivery approach is the delay experienced between the
operant response and delivery of reinforcement; a variable known to retard SA
acquisition (Williams, 1976; Black, Belluzzi, & Stein, 1985). Williams (1976) measured
the influence of varying response-reinforcement delays on the responding for grain in
food-deprived pigeons. It was demonstrated that even at the shortest delay (3-secs)
pigeons displayed a drastic reduction in responding when compared to a non-delayed
baseline. Additionally, Black, Belluzzi and Stein (1985) found delays as minimal as 1-
sec significantly impaired the acquisition of reinforcing brain self-stimulation in rats.
Thus, the delay experienced between the performance of an operant response and the
delivery of an oral alcohol reinforcer into a Fl-R may produce deficits in alcohol SA
acquisition, and lower behavioural responding.
Another limitation of the Fl-R approach is the inability to accurately measure alcohol
intake on a trial-by-trial basis, limiting the interpretation of consumption data. That is,
following each alcohol presentation the dipper-cup recedes back into the fluid reservoir
without measuring whether the liquid was consumed. The amount of alcohol intake can
be calculated by measuring the remaining liquid in the fluid reservoir at the end of each

15
session; however, trial-by-trial consumption is left undetermined. Additionally, without
trial-by-trial consumption data it is unknown whether each operant response is being
reinforced, thereby devaluing the capacity for SA behaviour to be a reliable measure of
alcohol-reinforced responding. Furthermore, unreinforced responding can lead to an
extinction of the operant response (Katner, Magalong, & Weiss, 1999), hence disturbing
the acquisition and maintenance of SA behaviour.
An additional shortcoming of the Fl-R technique is the variability in timing of liquid
prime deliveries. Oral alcohol primes are often delivered during
reinstatement/reacquisition procedures, as they have been demonstrated to reinstate
extinguished alcohol-seeking in rats (Bienkowski et al., 2000; Maccioni et al., 2007).
Using the Fl-R technique, an effective oral prime delivery is contingent upon the
animal’s approach response to the Fl-R. However, a rat must be responsive to the Fl-R
to receive the administered prime. If rats differ in their receptiveness to the Fl-R, the
timing of obtaining the oral prime could vary between-subjects. It has been
demonstrated that as many as 15 dipper-cup presentations are required to reinstate
extinguished responding for alcohol in rats (Bienkowski et al., 2000), an effect perhaps
attributable to inconsistencies in prime delivery between-subjects.
A Novel Technique Approach to the Oral Self-Administration of Alcohol in Rats.
Recently, a novel intraoral (IO) delivery technique was developed to administer liquid
reinforcement directly into the oral cavity of a rat during operant SA sessions (Levy et
al., 2014). By surgically implanting an oral cannula into the cheek, the IO delivery
technique overcomes many of the caveats posed by the Fl-R approach. First, IO
administrations will strengthen the response-reinforcement contingency by rapidly

16
delivering alcohol reinforcement upon the behavioural response. Second, in IO SA rats
directly consume each liquid reinforcer, providing consumption data for each trial. Third,
IO SA permits passive IO liquid infusions, allowing for a priming technique that does not
require movement toward the Fl-R, thereby decreasing between-subject variability.
In addition to strengthening the measurement of operant alcohol SA responding, IO
SA allows alcohol’s taste properties to be more efficiently investigated during
reinstatement and reacquisition procedures. That is, rather than the presentation of a
liquid prime in a Fl-R (visual + orosensory cues), IO primes are delivered directly into
the oral cavity to permit the analyses of taste and orosensory stimulation selectively.
The Reinforcing Properties of Sweeteners
Sweet substances have substantial and reliable effects on behavior. For example,
people describe the taste of sweeteners as pleasant (Thai et al., 2011; Frank et al.,
2008) and show preference for beverages that have been sweetened (Tatzer, Schubert,
Timischl, & Simbruner, 1985; Ventura & Mennella, 2011). In addition, individuals display
the motivation to consume sweetened beverages by exerting effort to gain access to its
administration (Schebendach et al., 2017; Guttman, 1954). The liking and preference for
sweeteners may be linked to activity in cortical regions that process reward functions
(Frank et al., 2008). Indeed, a variety of sweeteners increase the release of dopamine
into the nucleus accumbens (NAc; Rada, Avena & Hoebel, 2005; Wheeler et al., 2011;
Hajnal, Smith, & Norgren, 2004), a brain region implicated in attributing incentive
salience to rewarding stimuli (Wyvell & Berridge, 2000; Steinberg et al., 2014).
Although individuals show difficulty in discriminating between caloric and non-caloric
sweetness (Frank et al., 2008), caloric sweeteners are known to be more potent

17
reinforcers in terms of their behavioural and neurobiological effects (Domingos et al.,
2011; White & Carr, 1985). Some have even argued that caloric sweeteners may have
“addictive-like” properties, similar to drugs of abuse (Levy et al., 2015; Lenoir, Serre,
Cantin, & Ahmed, 2007; Madsen & Ahmed, 2014).
(1) High fructose corn syrup
High-fructose corn syrup (HFCS) is highly prevalent in our diets (Barlow, McKee,
Basu, & Stuckler, 2017; Bray, Nielsen, & Popkin, 2004), and has been linked to several
adverse health effects including obesity, heart disease, and poor metabolic health
(Meyers, Mourra, & Beeler, 2017; Bray, 2013; Stanhope et al., 2015). In comparison to
sucrose (which contains 50% glucose and 50% fructose), HFCS has a higher proportion
of fructose (typically, 55% fructose), and this is sufficient to engender different
behaviours during operant intraoral self-administration, as well as different patterns of
gene expression in the brain (Levy et al., 2015). Despite the recognition that the
reinforcing properties of HFCS differs from that of sucrose, (Ackroff & Sclafani, 2011;
Levy et al., 2015) research investigating the effect of HFCS on behaviour and brain is
quite limited.
Recently, this laboratory has explored behavioral and neurobiological responses
to high-fructose corn syrup in laboratory rats (HFCS; Levy et al., 2014; Levy et al., 2015;
Levy et al., 2018; Daniels, Marshall, & Leri, 2016). In Levy et al. (2015), rats were
trained to intraorally self-administer isocaloric solutions of 25% HFCS and 20% sucrose
under multiple schedules of reinforcement. It was found that rats self-administered
much more sucrose than HFCS during IO SA. Following IO SA, mRNA for dopamine
receptor 2 and mu-opiod receptor genes were quantified in brain regions that are

18
implicated in addictive behaviours. Results indicated that although rats self-
administered much more sucrose than HFCS, only HFCS led to similar neural
adaptations that are present following repeated exposure to drugs of abuse. Thus, Levy
et al. (2015) concluded that HFCS may be a more potent reinforcer than sucrose in
terms of the behavioural and neural adaptations it produces following exposure.
Sweeteners (HFCS) in alcoholic beverages It has been suggested that the use of sweeteners in alcoholic beverages reduces
the aversive taste of alcohol, and leads to increased consumption in naïve drinkers
(Mosher & Johnsson, 2005; Lanier et al., 2005; Romanus, 2000; Copeland, Stevenson,
Gates, & Dillon, 2007). Indeed, youth that primarily consume sweetened alcoholic
beverages report a higher frequency of binge drinking episodes (Albers, Siegel,
Ramirez, Ross, DeJong, & Jernigan, 2015). Furthermore, it is well established that
animals consume greater quantities of alcohol when it is mixed in a sweet solution. In
fact, rats fail to acquire the self-administration of EtOH without the use of sucrose-
pairings, sucrose-fading or other lengthy initiation procedures (i.e. intermittent access
schedules, food/water deprivation; Samson, 1986; Samson, Pfeffer, & Tolliver, 1988).
Although this may seem intuitively obvious, the mechanisms by which
sweeteners increase alcohol intake are not fully understood. That is, sweeteners not
only change the taste of alcohol, but they also support reliable effects on behaviour and
the brain. Thus, sweeteners also have the potential to synergistically alter alcohol’s
reinforcing value. For example, in addition to increasing alcohol intake in animals, sweet
substances are also capable of eliciting approach behaviour (Yasoshima & Shimura,
2015), conditioning neutral stimuli (Sclafani & Ackroff, 2017; White & Carr, 1985; Agmo

19
& Marroquin, 1997), and maintaining high levels of operant self-administration (Levy et
al., 2015; Avena, Long, & Hoebel, 2005).

20
THE EFFECTS OF HIGH-FRUCTOSE CORN SYRUP ON ETHANOL TAKING, SEEKING AND PALATABILITY IN MALE RATS. Samantha Ayoub, Meenu Minhas, Erin Rock, Linda Parker and Francesco Leri Department of Psychology and Collaborative Neuroscience, University of Guelph, Ontario, Canada, N1G 2W1 Corresponding author:
Francesco Leri, PhD
Professor
Department of Psychology
University of Guelph
Guelph, Ontario N1G 2W1
Canada
E-mail: [email protected]
519-824-4120 Ext: 58264
Acknowledgements
This research was funded by grants from the Natural Sciences and Engineering
Research Council (NSERC).

21
Abstract
Many alcoholic beverages contain sweeteners, such as high-fructose corn syrup
(HFCS), which has been implicated in increasing alcohol consumption and encouraging
hazardous drinking patterns. Therefore, four experiments were designed to investigate
whether the addition of HFCS to an ethanol (EtOH) solution could alter its intake,
hedonic taste, and sensitivity to food deprivation stress in Sprague Dawley rats.
Experiments 1 and 2 revealed that HFCS caused a robust increase in EtOH intake
during operant intraoral self-administration, however an equally palatable, yet non-
caloric sweetener (saccharin) could not replicate this effect. This suggests the caloric
value of a sweetener mediates the effect of sweeteners on EtOH intake. Experiments 3
and 4 revealed food deprivation stress increased EtOH consumption during resumption
following extinction, but only when HFCS was added to the solution. The palatability of
10% EtOH and 25% HFCS was also monitored across IO SA, and under conditions of
food deprivation stress. It was found that the palatability of 10% EtOH and 25% HFCS
increased following acquisition. More interestingly, food deprivation stress only
increased HFCS palatability. Thus, it could be the heightened palatability of HFCS
under stressful conditions that contributes to increased operant responding for solutions
containing HFCS during resumption in rats. These results have implications for the use
of highly palatable and caloric sweeteners in EtOH beverages because of the impact
they may have on excessive EtOH intake.
Key words
Operant self-administration, intraoral, alcohol intake, high-fructose corn syrup

22
Introduction Heavy episodic drinking refers to the consumption of 60 or more grams of alcohol
within a single occasion, occurring at least once per month (World Health Organization,
2014). This quantity of alcohol roughly equates to 5 standard drinks for men, and 4
standard drinks for women and is estimated to occur amongst 16% of drinkers
worldwide (World Health Organization, 2014). This statistic is concerning given that
heavy alcohol consumption is linked to numerous adverse health effects including, liver
and heart disease, birth defects, cancer, and unintentional injuries (Rehm et al., 2009).
In fact, the World Health Organization (2009) reported alcohol use as the third leading
risk factor contributing to the global burden of disease. In addition to the adverse health
effects associated with binge episodes (Massey & Arteel, 2012; Bala, Marcos, Gattu,
Catalano, & Szabo, 2014; Mathurin & Deltenre, 2009), the chronic consumption of large
quantities of alcohol increases an individual’s risk for subsequent disordered drinking
patterns and associated problem behaviours (Chassin, Pitts, & Prost, 2002; Hingson,
Heeren, & Winter, 2006).
Although there are several factors that facilitate the overconsumption of alcohol,
a key element appears to be the addition of sweeteners to alcoholic beverages (Mosher
& Johnsson, 2005; Copeland, Stevenson, Gates, & Dillon, 2007; Metzner & Kraus,
2008; Albers, Siegel, Ramirez, Ross, DeJong, & Jernigan, 2015). For example,
individuals that primarily consume sweetened alcoholic beverages have four times the
risk of experiencing alcohol-related injuries and report a higher frequency of heavy
episodic drinking (Albers et al., 2015). Furthermore, animal models show that
sweeteners increase the amount of alcohol self-administered in both home-cage and

23
operant self-administration procedures (Samson, Pfeffer, & Tolliver, 1988). In fact, most
species fail to acquire the self-administration of EtOH without the use of sucrose-fading
or other lengthy initiation procedures (i.e. intermittent access schedules, food/water
deprivation; Samson, Pfeffer, & Tolliver, 1988).
Sweet substances are known to have substantial and reliable effects on
behavior. For example, people describe the taste of sweeteners as pleasant (Thai et al.,
2011; Frank et al., 2008) and show preference for beverages that have been sweetened
(Tatzer, Schubert, Timischl, & Simbruner, 1985; Ventura & Mennella, 2011). In addition,
individuals display the motivation to consume sweetened beverages by exerting effort to
gain access to its administration (Schebendach et al., 2017; Guttman, 1954). The liking
and preference for sweeteners may be linked to activity in cortical regions that process
reward functions (Frank et al., 2008). Indeed, a variety of sweeteners increase the
release of dopamine into the nucleus accumbens (NAc; Rada, Avena & Hoebel, 2005;
Wheeler et al., 2011; Hajnal, Smith, & Norgren, 2004), a brain region implicated in
attributing incentive salience to rewarding stimuli (Wyvell & Berridge, 2000; Steinberg et
al., 2014). Although individuals show difficulty in discriminating between caloric and
non-caloric sweetness (Frank et al., 2008), caloric sweeteners are known to be more
potent reinforcers in terms of their behavioural and neurobiological effects (Domingos et
al., 2011; White & Carr, 1985). Some have even argued that caloric sweeteners may
have “addictive-like” properties (Levy et al., 2015; Lenoir, Serre, Cantin, Ahmed, 2007;
Madsen & Ahmed, 2014).
Recently, this laboratory has explored the behavioral and neurobiological
responses to high-fructose corn syrup in laboratory rats (HFCS; Levy et al., 2014; Levy

24
et al., 2015; Levy et al., 2018; Daniels, Marshall, & Leri, 2016). This sweetener was
selected for investigation because its consumption is highly prevalent in our diets
(Barlow, McKee, Basu, & Stuckler, 2017; Bray, 2013), and linked to several adverse
health effects including obesity, heart disease, and poor metabolic health (Meyers,
Mourra, & Beeler, 2017; Bray, 2013; Stanhope et al., 2015). In comparison to sucrose
(which contains 50% glucose and 50% fructose), HFCS has a higher proportion of
fructose (typically, 55% fructose), and this is sufficient to engender different behaviours
during operant intraoral self-administration in rats, as well as different patterns of gene
expression in the brain (Levy et al., 2015).
The overall aim of this study was to explore the impact of HFCS on operant
intraoral self-administration of ethanol (EtOH) in outbred male rats. This is a relevant
question because HFCS is used as sweetener in many alcoholic beverages available
for purchase (i.e. Smirnoff Ice, Mike’s Hard Lemonade, Bacardi Silver). It is generally
known that sweeteners change the taste of EtOH (Blednov et al., 2008) and facilitate
consumption in various species (Tolliver, Sadeghi, & Samson, 1988; Mandillo, Titchen,
& Miczek, 1998; Lanier, Hayes, & Duffy, 2004), but how HFCS interacts with EtOH’s
reinforcing effect is currently unknown.
Therefore, Experiment 1 was designed to establish whether HFCS can modify
operant intraoral self-administration (IO SA) of alcohol (EtOH; 5%, 10% and 20%
alc/vol) in outbred male rats. Experiment 2 was designed to contrast the effects of
HFCS and saccharin (SACC; a non-caloric sweetener) on EtOH IO SA. Experiment 3
tested the persistence of operant responding following extinction conditions in animals
trained to IO SA EtOH & HFCS. This experiment also included a stress manipulation,

25
which is known to alter the reinforcing effects of drugs of abuse and facilitate drug-
seeking in many species (Shaham, Erb, & Stewart, 2000). We selected food deprivation
stress (Shalev, Marinelli, Baumann, Piazza, & Shaham, 2003; Sinha, 2008) because of
the relevance to the behaviour explored (i.e., ingestion of calories from both EtOH and
HFCS). Finally, Experiment 4 expanded the findings of Experiment 3 by also
investigating how changes in EtOH and HFCS palatability, assessed by taste reactivity
(Grill & Norgren, 1978), may play a role in persistent drug-taking, drug-seeking and the
effect of food deprivation stress.
Methods
Subjects
Male Sprague Dawley rats were acquired at 175-200 grams (Charles River
Laboratories, Saint-Constant, Quebec) for all experiments. Animals were single-housed
and located in a colony room maintained on a 12-hour light/dark cycle (7:00/19:00h).
Standard rat chow and water were provided ad-libitum, unless indicated otherwise
below. Rats were acclimated to the colony room and handled daily for one week prior to
any experimental conditions. All procedures were reviewed and approved by The
University of Guelph Animal Care Committee and adhere to sanctions put forth by the
Canadian Council on Animal Care.
Apparatus
Self-administration chambers. Plexiglas operant conditioning chambers (model
ENV-008CT, Med Associates, Georgia, VT) were used for all SA procedures. Each
conditioning chamber was located within a larger sound- attenuating partition, with a
built-in fan to mask noise and provide constant ventilation. An overhead houselight (28-

26
V) was situated on the back wall of each chamber. On the front wall, a retractable lever
(8-cm above floor level) was situated just below a cue-light (28-V; 11-cm above floor
level). The retractable lever served as an active lever during SA sessions, and was
linked to a liquid infusion pump (Razel Scientific Instruments, Stamford, Connecticut).
On the opposite side of the front wall, there was a non-retractable lever (8-cm above
floor level) that served as an inactive lever during SA sessions. Stainless steel bars
provided the foundation of the conditioning chambers (30.5-cm x 24.1-cm x 21.0-cm). A
MED-PC interface controlled all conditioning chambers through MED-PC compatible
software.
Taste reactivity chambers. Taste reactivity chambers were constructed of clear
Plexiglas (22.5 x 26.0 x 20.0 cm) that sat atop a table with a clear glass top. A mirror
was placed beneath the chamber, tilted at a 45 angle to facilitate adequate viewing of
the rat’s ventral surface. Polyethylene tubing connected the IO cannula to an infusion
pump (KDS100; KD Scientific Inc., USA) which was set to deliver liquid for two minutes
at a rate of 0.5 ml/min. All taste reactivity sessions were recorded with a video camera
for experimenter scoring.
Surgery
IO cannulas were surgically implanted following a week of acclimation to the
colony room. The materials and procedures used for IO cannulation surgery have been
previously described in detail (Levy et al., 2014). Briefly, cannulas were constructed by
flanging polyethylene-90 tubing (approximately 80-mm in length) using a soldering iron.
Rats were then anesthetized using isoflurane, and cannulas were surgically implanted.
Using a 15-G stainless steel needle, cannulas were woven subcutaneously through the

27
skin at the back of the neck to protrude intraorally from the left cheek. A head-cap was
constructed using dental cement to uphold the bolt used to secure IO tubing during SA
sessions. Rats were given a week to recover following all surgical procedures.
General Procedures
Operant self-administration: Rats were food-restricted for all IO SA sessions by
allowing unlimited access to standard rat chow for only three hours daily. Feeding
occurred immediately following IO SA sessions. All IO SA sessions began 4-hrs into the
rat’s dark (active) cycle and ran for 180-min duration. Cannulas were flushed at the
beginning of every session with reverse osmosis water to ensure there were no
obstructions in the delivery line.
Rats were placed into the conditioning chambers and allowed to habituate for 5-
min prior to each session. Illumination of the house light signaled the beginning of a
session, with the active lever extending out after a 10-sec delay. Subsequently,
responding on the active lever resulted in a 0.1-ml IO infusion of liquid reinforcement on
a fixed-ratio 1 (FR1) or fixed-ratio 3 (FR3) schedule of reinforcement, and the
presentation of a cue-light (27.5-secs). During this time, responding on the active lever
was recorded but did not lead to additional reinforcement. Responding on the inactive
lever had no programmed consequences under any experimental conditions as it
served as a baseline of non-reinforced operant responding.
For the first three sessions of each IO SA experiment the lever-press response
was shaped by experimenter-induced priming. During priming sessions, rats were
allowed 15-min to perform an active lever-press response. If no response occurred
during this time, the experimenter held the rat up to the active lever and made the

28
animal press down to receive liquid reinforcement This was repeated every 15-min, up
to six times per session.
Extinction, cue-induced reinstatement and resumption: Once IO SA was stable
on a FR3 schedule of reinforcement (see below), rats received one 180-min extinction
session per day, for five days. Previous IO SA studies in our laboratory indicated that
after 5 days of extinction, responding on the active and inactive levers is comparable.
To maintain the oral sensation of liquid delivery during extinction, rats responded on the
active lever for a 0.1-ml IO delivery of reverse osmosis water. There was no
presentation of a cue-light during IO water delivery.
Cue-reinstatement testing was conducted during a 180-minute session following
the last day of extinction. During reinstatement, rats could respond on the active lever
for a 0.1 ml IO delivery of reverse osmosis water and the illumination of the cue light.
Resumption tests were conducted in an identical manner, but rats could respond
on the active lever for a 0.1 ml IO delivery of EtOH, HFCS or EtOH + HFCS and the
presentation of a cue-light.
Extinction, reinstatement and resumption sessions were tested on a FR3
schedule of reinforcement. Active and inactive lever responding, and number of
infusions earned were recorded.
Experimental procedures
Experiment 1: IO SA of EtOH and combined EtOH + HFCS.
Rats (n=8) self-administered 5% EtOH on a FR1 schedule of reinforcement for
16 consecutive 180-min sessions. The response requirement was increased to a FR3
schedule of reinforcement for the next four sessions (17-20), and then reduced back to

29
an FR1 schedule for four sessions (21-24). This alternation in schedule was performed
to ascertain baseline level of EtOH self-administration. On session 25, 5% EtOH was
mixed in a 25% HFCS solution and animals self-administered the mixed solution on a
FR1 schedule. The FR3 schedule was then reintroduced for 8 additional sessions (26-
33). Following this period, the concentration of EtOH in 25% HFCS was varied in blocks
of four sessions to complete a dose-response curve. That is, rats self-administered a
10% EtOH-HFCS solution for four sessions (34 – 37), then a 20% EtOH-HFCS solution
for four sessions (38 – 41), and then a 5% EtOH-HFCS solution for another four
sessions (42 – 45). To conclude the experiment, HFCS was removed and rats reverted
to self-administering 5% EtOH for four sessions (45-49).
Experiment 2: IO SA of EtOH and combined EtOH + SACC
Rats (n=14) self-administered a 5% EtOH solution mixed in 0.1% saccharin for
10 180-min sessions. Next, the schedule of reinforcement was increased to a FR3
schedule for seven more sessions (11-17). Subsequently, a dose-response curve was
performed as in Experiment 1: the concentration of EtOH was successively increased
(5%, 10%, 20%, 5%) in the 0.1% saccharin solution (sessions 17-32). Following the
dose-response determination, rats received a single session (33) in which they could
respond for 25% HFCS. This session tested whether this group of rats could emit more
responses for a more effective reinforcer (Levy et al. 2016).
Experiment 3: Effect of food deprivation stress on IO SA acquisition, extinction and
resumption of EtOH + HFCS
Rats (n=28) self-administered 10% EtOH in 25% HFCS on an FR1 schedule for
10 180-min sessions (1-10), and then the schedule of reinforcement was increased to

30
an FR3 schedule for 7 sessions (11-17). The concentration of EtOH was selected based
on the results of Experiments 1-2 which determined 10% EtOH + 25% HFCS led to the
highest g/kg alcohol intake (0.91 g/kg). Following 5 180-min extinction sessions, rats
were randomly assigned to food restriction (n=14) or food deprivation (n=14) conditions.
Food restricted rats continued their typical food restriction regimen of 3-hours of
free access to standard rat chow, and the test of cue-induced reinstatement was
followed by two tests of resumption 10% EtOH and 25% HFCS intake, which occurred
on two counterbalanced sessions.
Food deprived rats did not receive any food after their last extinction session, but
they were fed after the test of cue-induced reinstatement. To control for food deprivation
stress carry over effects, they received one more extinction session followed by EtOH -
only or HFCS-only resumption test, and they were fed immediately after the test. This
was repeated once more to counterbalance for the substance self-administered.
Experiment 4: Effect of food deprivation on resumption of EtOH + HFCS IO SA
responding-modulation by palatability of components.
A within-subjects design was used to investigate the effect of food deprivation
stress on 10% EtOH + 25% HFCS intake during IO SA resumption testing. Additionally,
a taste reactivity (TR) test was given at four different time points throughout the SA
period to assess possible changes in palatability of 10% EtOH and 25% HFCS under
pre-acquisition, post-acquisition, food restriction and food deprivation conditions. The
timeline of procedural events that occurred during Experiment 4 is represented in Figure
4A.
The day preceding the first TR test, rats were habituated to TR chambers by

31
intraorally infusing reverse osmosis water at a flow rate of 0.5 ml/min for two minutes.
During TR tests rats received 10% EtOH and 25% HFCS separately over a two-min
exposure period. The appropriate liquid was dispensed at the same flow rate of 0.5
ml/min, resulting in 1 ml of the appropriate solution dispensed over a two-min period.
EtOH and HFCS solutions were presented to rats an hour following an IO SA session
and these solutions were tested in a counterbalanced order across rats. All test
sessions were captured on video and tapes were scored for the mean number of
hedonic reactions of tongue protrusions (protrusions of the tongue from the mouth) and
mouth movements (summed). Additionally, tapes were scored for the mean number of
aversive reactions of gaping (wide triangular shaped open mouth exposing lower
incisors) and the avoidance reaction of passive dripping (passive dripping of liquid from
the mouth to the floor). The aversive reactions of chin rubbing (rubbing chin along the
floor of the cage) and paw treading (movement of forepaws back and forth) were also
assessed, but these behaviors were not seen so they are not discussed further.
Rats (n=18) self-administered 10% EtOH + 25% HFCS on a FR1 schedule of
reinforcement for 10 consecutive 180-min sessions. The response requirement was
increased to a FR3 schedule of reinforcement for the next seven sessions (11-17). On
session 25, rats received five extinction sessions (18-22), followed by tests of cue-
induced reinstatement and resumption of 10% EtOH + 25% HFCS IO SA in both a food
deprived and food restricted state. Extinction sessions were interposed between each
test day as described above.
Drugs
Ethanol solutions (5%, 10%, 20%) were prepared by diluting ethyl alcohol (95%

32
vol; Commercial Alcohols, Tiverton, Ontario) in reverse osmosis water. High-fructose
corn syrup (25%; HFCS-55 formula; Natures Flavors, CA, USA) and saccharin (0.1%;
sodium salt hydrate 99+ %, ACROS Organics, New Jersey) solutions were also
prepared by dilution in reverse osmosis water. Solutions were freshly prepared every
three days to prevent evaporation.
Statistical analyses
One factor analyses of variance (ANOVA) and t-tests with independent or
repeated subjects were conducted as appropriate. Significant main effects or
interactions were further analysed by multiple comparisons using the Student-Newman
Keuls method (α=0.05). All analyses were performed using SigmaStat (v. 12.5).
Results
Experiment 1: IO SA of EtOH and combined EtOH + HFCS.
Rats demonstrated higher IO SA of 5% EtOH + 25% HFCS than 5% EtOH
alone. Figure 1A represents the mean (SEM) infusions rats earned across 25 days of IO
SA acquisition for 5% EtOH (sessions 1-24) and 5% EtOH + 25% HFCS (session 25).
An ANOVA revealed a main effect of session [F(7, 24) = 20.934, p < 001]. Multiple
comparisons indicated that the number of infusions rats earned on session 25 was
significantly higher than sessions 1 – 24, p < 0.001. No other significant differences
were found.
In a dose-response assessment, when 20% EtOH was added to HFCS, rats
showed suppressed IO SA. Figure 1B represents the mean (SEM) infusions rats earned
across sessions 26 – 45, during which the concentration of EtOH in 25% HFCS was
varied to create a dose-response (intake) curve. Blocks of four sessions were used to

33
allow IO SA behaviour to stabilize and the last session of which each concentration was
presented were compared (sessions 33, 37, 41 and 45). An ANOVA revealed a main
effect of session [F (3, 15) = 19.872, p < 0.001]. Multiple comparisons indicated rats
earned significantly less infusions on session 41 (20% EtOH) than on sessions 33 (5%
EtOH; p < 0.001), 37 (10% EtOH; p < 0.001) and 45 (5% EtOH; p < 0.001). No other
significant differences were found.
Rats displayed enhanced IO SA of 5% EtOH following training with 5% EtOH +
25% HFCS. Figure 1C represents the mean (SEM) infusions rats earned on the last day
of acquisition for 5% EtOH prior to HFCS pairing (FR3; session 20) compared to the
four sessions after HFCS was removed (FR3; sessions 46-49). An ANOVA revealed a
significant effect of session [F (4, 16) = 4.675, p = 0.11]. Multiple comparisons indicated
rats consumed less 5% EtOH on session 20 than sessions 46 (p < 0.01), 47 (p < 0.05),
and 49 (p < 0.05). No other significant differences were found.
Experiment 2. IO SA of EtOH and combined EtOH + SACC.
Saccharin was much less effective than HFCS in enhancing IO SA of EtOH .
Figure 2A represents the mean (SEM) infusions rats earned across 16 days of IO SA
acquisition for 5% EtOH in 0.1% SACC. An ANOVA revealed a main effect of session [F
(13, 208) = 2.038, p = 0.012]. Multiple comparisons indicated that infusions earned by
rats did not differ between session 1 and session 17. In other words, although infusions
earned varied across sessions, there was not a significant increase in 5% EtOH in 0.1%
SACC intake between the first and last day of IO SA. Figure 2B represents the mean
(SEM) number of infusions rats earned on the last session of Experiment 1 (session 49;
FR3; 5% EtOH) and the last session of Experiment 2 (session 30; FR3; 5% EtOH in

34
0.1% SACC). An independent samples t-test revealed rats earned more infusions when
EtOH was mixed in 0.1% SACC, [t(20)= -2.68, p=0.01]. Figure 2C represents the mean
(SEM) number of infusions rats earned across sessions 14 to 29 during which the
concentration of EtOH in 0.1% SACC was varied to create a dose-response (intake)
curve. Blocks of four sessions were used to allow IO SA behaviour to stabilize and the
last session of which each concentration was presented were compared (sessions 17,
21, 25, and 29). An ANOVA revealed main effect of session [F (3, 39) = 3.741, p <
0.019]. Multiple comparisons indicated rats earned significantly fewer infusions on
session 25 (20% EtOH) than on session 17 (5% EtOH; p < 0.001). No other significant
differences were found. Figure 2D represents the mean (SEM) number of infusions rats
earned during the IO SA dose-response (intake) curve for EtOH in SACC (Experiment
2) and EtOH in HFCS (Experiment 1). An ANOVA revealed a solution by concentration
interaction [F (3, 72) = 8.324, p < 0.001], a main effect of solution [F (1, 72) = 127.387, p
< 0.001] and a main effect of concentration [F (3, 72) = 16.392, p < 0.001]. Multiple
comparisons on marginal means indicated that higher concentrations of EtOH
significantly reduced intake and that animals consumed significantly more solution when
HFCS was added to EtOH than when SACC was added.
Experiment 3. Effect of food deprivation stress on cue-induced and EtOH or HFCS-
induced resumption of responding.
Rats displayed enhanced responding for EtOH + HFCS across acquistion
sessions. Following extinction, food deprivation stress enhanced cue-induced and
HFCS (but not EtOH)-induced resumption of responding. Figure 3A represents the
mean (SEM) infusions rats earned across 17 days of IO SA of 10% EtOH in 25% HFCS.

35
An ANOVA across sessions revealed a main effect of session [F (1, 27) = 4.690, p <
0.05]. Multiple comparisons indicated rats significantly increased their intake from FR1-1
to FR1-10 (p < 0.001) and from FR3-1 to FR3-7. Furthermore, the number of infusions
earned did not significantly differ across the last four sessions of each schedule of
reinforcement. Figure 3B represents the mean (SEM) active lever responses rats made
on the last day of IO SA acquisition (Baseline; B) and the following 5 extinction
sessions. An ANOVA revealed a main effect of session [F (5, 162) = 100.178, p <
0.001]. Multiple comparisons indicated response rates were lower across all extinction
sessions when compared to the baseline responding during IO SA acquisition.
Furthermore, extinction sessions 2 – 5 produced significantly lower lever responding
when compared to the first extinction session. No other significant differences were
found. Figure 3C represents the mean (SEM) active lever responses made on the test
of cue reinstatement in food restricted and food deprived rats. A t-test revealed food
restricted rats had lower levels of responding compared to food deprived rats when
tested for cue-induced reinstatement, [t(26)= -2.166, p < 0.05]. Figure 3D represents the
mean (SEM) infusions rats earned on the tests of EtOH and HFCS resumption.
Independent samples t-tests revealed food deprivation stress increased HFCS intake
during resumption, [t(26)= 4.068, p=0.001] but had no effect on EtOH intake [t(26)=
0.148, p=0.884].
Experiment 4: Effect of food deprivation on resumption of EtOH + HFCS IO SA
responding-modulation by palatability of components.
Food deprivation stress enhanced EtOH+HFCS resumption of responding
following extinction. Both EtOH and HFCS individually elicited enhanced hedonic

36
responses in the TR test following acquisition training, and food deprivation stress
during resumption testing enhanced hedonic responses of HFCS, but not EtOH. Figure
4B represents the mean (SEM) active lever responses made during EtOH + HFCS
resumption following extinction across food restricted and food deprived conditions. A
paired samples t-test revealed that food deprived rats consumed more EtOH + HFCS
during resumption than rats under food restriction conditions [t(17)= -2.213, p < 0.05].
Figure 4C represents the mean (SEM) hedonic taste responses elicited by EtOH or
HFCS during baseline testing compared to post-acquisition taste reactivity testing. A
paired samples t-test revealed hedonic taste reactions were increased during post-
acquisition compared to baseline TR testing for EtOH [t(19)= -5.098, p < 0.001] and
HFCS [t(19)= -2.311, p < 0.05]. There was no effect on aversive taste reactions to EtOH
and HFCS (data not shown). Figure 4D represents the mean (SEM) hedonic taste
responses elicited by EtOH or HFCS during taste reactivity testing for food restricted
and food deprived rats. Paired samples t-tests revealed that food deprivation stress
increased HFCS palatability [t(19) = -3.733, p < 0.001] but not EtOH palatability [t(19)=
1.027, p =0.318].
Discussion
Young, inexperienced and often under-aged drinkers report a high frequency of
heavy episodic drinking patterns (Archie, Kazemi, & Akhtar-Danesh, 2012) and adding
sweeteners to alcohol may increase the risk of problematic drinking in this population.
Therefore, we designed four experiments in naïve Sprague Dawley rats to investigate
whether the addition of a caloric sweetener (HFCS) to an EtOH solution could alter its
intake, hedonic taste, and sensitivity (intake under) to food deprivation stress. In

37
Experiment 1 (to study the effect of HFCS on EtOH intake) rats were trained to
intraorally self-administer 5% EtOH under multiple schedules of reinforcement. The
addition of 25% HFCS caused a robust increase in EtOH intake during IO SA.
Additionally, when HFCS was removed and rats reverted to self-administering the 5%
EtOH solution, their intake was elevated from levels seen before the HFCS pairings.
Using similar methods, Experiment 2 revealed an equally palatable, yet non-caloric
sweetener (SACC) did not increase EtOH intake to the degree that was observed with
HFCS. In Experiments 3 and 4 (to study the effect of HFCS on EtOH’s sensitivity to food
deprivation stress) rats were trained to self-administer 10% EtOH in 25% HFCS. This
concentration of EtOH was chosen based on results from Experiment 1. Following
acquisition, rats underwent behavioural extinction to test the persistence of operant
responding during cue-induced reinstatement and resumption (of 10% EtOH, 25%
HFCS, and 10% EtOH + 25% HFCS) under food deprivation stress. Food deprivation
stress increased 25% HFCS and 10% EtOH + 25% HFCS intake, but did not alter the
intake of 10% EtOH during resumption. Finally, the changes in palatability of 10% EtOH
and 25% HFCS were assessed using taste reactivity at multiple time points throughout
the IO SA of 10% EtOH + 25% HFCS. Rats elicited more hedonic orofacial responses to
10% EtOH and 25% HFCS following IO SA of 10% EtOH + 25% HFCS. Furthermore,
food deprivation stress increased hedonic orofacial responses to the 25% HFCS
solution, but did not alter 10% EtOH palatability. Thus, it could be the increased
palatability of HFCS under food deprivation conditions that contributes to the increased
motivation to consume HFCS during operant IO SA resumption in rats. These results

38
have implications for the use of highly palatable and caloric sweeteners in EtOH
beverages because of the impact they may have on excessive EtOH intake.
When rats could self-administer 5% EtOH during IO SA, intake was minimal.
However, when the same concentration of EtOH was sweetened with 25% HFCS, a
robust increase in EtOH intake was observed (see Figure 1A). Following this, a dose-
response (intake) curve was produced by varying the concentration of EtOH within
HFCS. Rats maintained the number of infusions earned when the concentration of
EtOH was increased from 5% to 10%, however a further increase to 20% EtOH reduced
the amount of solution consumed (see Figure 1B.). In other words, the 10% EtOH +
25% HFCS solution produced the highest levels of EtOH intake during IO SA. Finally,
when rats reverted to self-administering 5% EtOH, their intake was elevated from levels
seen before the HFCS pairings (see Figure 1C.). It is important to note that although
SACC increased the intake of EtOH during IO SA, this effect was miniscule in
comparison to what was observed when HFCS was used to sweeten the EtOH solution
(see Figure 2).
Caloric sweeteners such as sucrose and glucose increase EtOH intake during
SA in rodents (Samson, Pfeffer, & Tolliver, 1988); however, to our knowledge, we are
the first to demonstrate this effect using HFCS. HFCS was selected as its use is rapidly
increasing throughout North America, yet its effects on behaviour and the brain are still
not well understood. Additionally, HFCS is found in many of the alcoholic beverages
available for purchase (i.e. Bacardi Silver, Palm Bay, Mike’s Hard Lemonade). In
addition to increasing EtOH intake, caloric sweeteners are also capable of eliciting
approach behaviour (Yasoshima & Shimura, 2015) conditioning neutral stimuli (Sclafani

39
& Ackroff, 2017; White & Carr, 1985; Agmo & Marroquin, 1997), and maintaining high
levels of operant self-administration (Levy et al., 2015; Avena, Long, & Hoebel, 2005).
That is, caloric sweeteners are known to have strong behavioural reinforcing properties.
Therefore, it is not surprising that rats increased their EtOH intake when HFCS was
mixed into the EtOH solution. We suggest this increase was mostly caused by caloric
reinforcement, because an equally palatable yet non-caloric sweetener did not cause
such a drastic increase in IO SA behaviour. Although non-caloric sweeteners hold
similar hedonic taste properties as do caloric sweeteners (Frank et al., 2008), it is
known that their behavioural reinforcing properties are minimal in comparison
(Domingos et al., 2011; White & Carr, 1985).
Interestingly, our study also suggests that HFCS may have indirect effects on
EtOH intake. That is, once HFCS was removed from EtOH following multiple pairings,
EtOH intake was elevated in comparison to initial IO SA levels. While we understand
that many factors could have contributed to this effect (increased palatability of EtOH,
tolerance) it could be that HFCS conditioned or altered EtOH’s reinforcing value. After
all, caloric sweeteners are known to modify the reinforcing value of neutral stimuli
(Sclafani & Ackroff, 2017; Sclafani, Glass, Margolskee, & Glendinning, 2010; Fedorchak
& Bolles, 1987; White & Carr, 1985; Agmo & Marroquin, 1997; Delamater, Sclafani, &
Bodnar, 2000). To test this, future research could employ methods that are more
suitable for measuring the reinforcing value of stimuli, such as, progressive ratio operant
testing or choice procedures (Reilly, 1999; Sclafani & Ackroff, 2003; Butler, Irons,
Bassett, & Correia, 2018).

40
Once the effect of HFCS on EtOH consumption was determined, our next
objective was to assess how HFCS could alter EtOH’s sensitivity to food deprivation
stress. Food deprivation stress has been shown to elevate corticosterone levels in rats
(McGhee, Jefferson, & Kimball, 2009) and facilitate the reinstatement of operant self-
administration for drugs of abuse (Shalev, Marinelli, Baumann, Piazza, & Shaham 2003;
Highfield et al, 2002; Mantsch, Baker, Funk, Le & Shaham, 2016). Food deprivation
stress was also chosen based on its relevance to the behaviour being explored (i.e.,
ingestion of caloric substances). First, rats acquired IO SA of 10% EtOH + 25% HFCS
(see Figure 3A) then this behaviour was extinguished (see Figure 3B). The absence of
the cue-light during extinction conditions is important to establish that this stimulus only
represents sweetened EtOH reinforcement. The use of water during extinction
conditions is also crucial to maintain the orosensory stimulation that is experienced
during IO SA acquisition. During cue-induced reinstatement food deprivation stress
increased responding for water reinforcement (see Figure 3C). During resumption
testing, food deprivation stress did not increase 10% EtOH intake, however it increased
the intake of 25% HFCS. Finally, Experiment 4 revealed that food deprivation stress
increased the intake of 10% EtOH + 25% HFCS (see Figure 4A) and the palatability of
25% HFCS, but not 10% EtOH.
In this study, the lack of an effect of food deprivation stress on EtOH intake
during resumption was surprising. This is because stress has long been implicated in
exacerbating relapse-like responding for EtOH in humans. For example, the experience
of a stressor in abstinent alcoholics is associated with increased EtOH cravings (Clay et
al., 2018; Fox, Bergquist, Hong, & Sinha, 2007). Additionally, the occurrence of severe

41
stress following treatment is correlated with increased relapse in recovering alcoholics
(Brown, Vik, Patterson, Grant, & Schuckit, 1995; Brown et al., 1990). Finally, exposure
to acute stress increases alcohol consumption in heavy drinkers (Thomas, Bacon,
Randall, Brady, & Ronald, 2011; McGrath, Jones, & Field, 2016). In rodents however,
the effect of stress on EtOH consumption is inconsistent (Pohorecky, 1990) with some
researchers reporting decreases (Sprague & Maickel, 1994; Vengeliene, Siegmund,
Singer, Sinclair, Li, & Spanagel, 2003) while others report increases (Bahi, 2013;
Gomez, Lewis, & Luine, 2012; Vengeliene, et al., 2003). Interestingly, food deprivation
stress increased EtOH intake only when HFCS was mixed in the solution. These results
imply that HFCS increases EtOH’s sensitivity to the effect of food deprivation stress.
Thus, HFCS alters EtOH intake under normal and stressful conditions and this should
be considered when employing efforts to reduce excessive EtOH intake.
The last objective of this study was to assess how the palatability of 10% EtOH
and 25% HFCS is altered throughout the IO SA of 10% EtOH in 25% HFCS. More
specifically, taste reactivity was used to test the palatability of each solution pre-
acquisition (naïve responding), post-acquisition (experienced responding) and under
conditions of food deprivation stress (see Figure 4A). The number of palatable orofacial
responses to EtOH and HFCS increased following IO SA of 10% EtOH in 25% HFCS
(see Figure 4 C.). Interestingly, the number of aversive orofacial responses (i.e. gaping)
did not change for either solution following IO SA experience (data not shown). While
rats displayed no aversive responses to the taste of 25% HFCS, gaping occurred during
pre- and post-acquisition for the 10% EtOH solution (data not shown). During

42
resumption, food deprivation stress increased palatable orofacial responses to 25%
HFCS, but did not alter EtOH palatability (see Figure 4D).
This data suggests that the IO SA of 10% EtOH in 25% HFCS alters the hedonic
palatability of EtOH, without altering its aversive taste properties. In contrast, Kiefer,
Bice, and Badia-Elder (1994) have shown no change in hedonic taste, but a decrease in
aversive taste responses following continuous home-cage access to a variety of EtOH
concentrations. The lack of difference between aversive taste responses during pre-
and post-acquisition may be explained by the use of 25% HFCS during EtOH exposure.
That is, our rats never experienced EtOH by itself during the exposure period as it was
always mixed in 25% HFCS. The effect of food deprivation on EtOH and HFCS
palatability may be the most interesting finding from the taste reactivity data because it
corroborates with the self-administration results from Experiments 3 and 4. Food
deprivation stress had no effect on EtOH intake under resumption testing, nor did it
affect palatability. On the other hand, food deprivation stress significantly increased
HFCS intake under resumption and increased its palatability. We suggest that the
increased palatability of HFCS under stressful conditions may contribute to the
increased motivation to consume HFCS during operant responding.
These data imply the use of HFCS in alcoholic beverages encourages EtOH
consumption, increases its sensitivity to food deprivation stress, and increases its
hedonic taste. Efforts aimed at reducing the harmful consumption of EtOH in naïve
drinkers should include removal of HFCS from the list of ingredients.

43
References
Agmo, A., & Marroquin, E. (1997). Role of gustatory and postingestive actions of
sweeteners in the generation of positive affect as evaluation by place preference
conditioning. Appetite, 29(3), 269-289.
Albers, A. B., Siegel, M., Ramirez, R. L., Ross, C., DeJong, W., & Jernigan, D. H.
(2015). Flavored alcoholic beverage use, risky drinking behaviors, and adverse
outcomes among underage drinkers: Results from the ABRAND study. American
Journal of Public Health, 105(4), 810-815.
Anderson, P., de Bruijn, A., Angus, K., Gordon, R., & Hastings, G. (2009). Impact of
alcohol advertising and media exposure on adolescent alcohol use: A systematic
review of longitudinal studies. Alcohol and Alcoholism, 44(3), 229-243.
Archie, S., Zangeneh Kazemi, A., & Akhtar-Danesh, N. (2012). Concurrent binge
drinking and depression among canadian youth: Prevalence, patterns, and
suicidality. Alcohol, 46(2), 165-72.
Avena, N. M., Long, K. A., & Hoebel, B. G. (2005). Sugar-dependent rats show
enhanced responding for sugar after abstinence: Evidence of a sugar deprivation
effect. Physiology & Behavior, 84(3), 359-362.
Bahi, A. (2013). Increased anxiety, voluntary alcohol consumption and ethanol-induced
place preference in mice following chronic psychosocial stress. Stress, 16(4),
441-451.
Bala, S., Marcos, M., Gattu, A., Catalano, D., & Szabo, G. (2014). Acute binge drinking
increases serum endotoxin and bacterial DNA levels in healthy individuals. PLoS
One, 9(5).

44
Barlow, P., McKee, Basu, & Stuckler, (2017). Impact of the north american free trade
agreement on high-fructose corn syrup supply in canada: A natural experiment
using synthetic control methods. Canadian Medical Association Journal, 189(26),
E881-E887.
Blednov, Y. A., Walker, D., Martinez, M., Levine, M., Damak, S., & Margolskee, R. F.
(2008). Perception of sweet taste is important for voluntary alcohol consumption
in mice. Genes, Brain, and Behavior, 7(1), 1-13.
Bray, G. A. (2013). Energy and fructose from beverages sweetened with sugar or high-
fructose corn syrup pose a health risk for some people. Advances in
Nutrition, 4(2), 220-225.
Brown, S. A., Vik, P. W., McQuaid, J. R., Patterson, T. L., Irwin, M. R., & Grant, I.
(1990). Severity of psychosocial stress and outcome of alcoholism treatment. Journal
of Abnormal Psychology, 99(4), 344-348.
Brown, S. A., Vik, P. W., Patterson, T. L., Grant, I., & Schuckit, M. A. (1995). Stress,
vulnerability and adult alcohol relapse. Journal of Studies on Alcohol, 56(5), 538-
545.
Butler, L. H., Irons, J. G., Bassett, D. T., & Correia, C. J. (2018). Using the multiple-
choice procedure to measure the relative reinforcing efficacy of gambling: Initial
validity evidence among college students. Journal of Gambling Studies, 34(2),
513-520.
Chassin, L., Pitts, S. C., & Prost, J. (2002). Binge drinking trajectories from adolescence
to emerging adulthood in a high-risk sample: Predictors and substance abuse
outcomes. Journal of Consulting and Clinical Psychology, 70(1), 67-78.

45
Clay, J. M., Adams, C., Archer, P., English, M., Hyde, A., Stafford, L. D., & Parker, M.
O. (2018). Psychosocial stress increases craving for alcohol in social drinkers:
Effects of risk-taking. Drug and Alcohol Dependence, 185, 192-197.
Copeland, J., Stevenson, R. J., Gates, P., & Dillon, P. (2007). Young australians and
alcohol: The acceptabllity of ready-to-drink (RTD) alcoholic beverages among 12-
30-year-olds. Addiction, 102(11), 1740-1746.
Daniels, S., Marshall, P., & Leri, F. (2016). Alterations of naltrexone-induced
conditioned place avoidance by pre-exposure to high fructose corn syrup or
heroin in sprague-dawley rats. Psychopharmacology, 233(3), 425-433.
Delamater, A. R., Sclafani, A., & Bodnar, R. J. (2000). Pharmacology of sucrose-
reinforced place-preference conditioning: Effects of naltrexone. Pharmacology
Biochemistry and Behavior, 65(4), 697-704
Domingos, A. I., Vaynshteyn, J., Voss, H. U., Ren, X., Gradinaru, V., Zang, F., . . .
Friedman, J. (2011). Leptin regulates the reward value of nutrient. Nature
Neuroscience, 14(12), 1562-8.
Fedorchak, P. M., & Bolles, R. C. (1987). Hunger enhances the expression of calorie-
but not taste-mediated conditioned flavor preferences. Journal of Experimental
Psychology.Animal Behavior Processes, 13(1), 73-79.
Frank, G. K. W., Oberndorfer, T. A., Simmons, A. N., Paulus, M. P., Fudge, J. L., Yang,
T. T., & Kaye, W. H. (2008). Sucrose activates human taste pathways differently
from artificial sweetener. NeuroImage, 39(4), 1559-1569.
Fox, H. C., Bergquist, K. L., Hong, K., & Sinha, R. (2007). Stress-induced and alcohol
cue-induced craving in recently abstinent alcohol-dependent individuals. Alcoholism,

46
Clinical and Experimental Research, 31(3), 395-403.
Gomez, J. L., Lewis, M. J., & Luine, V. N. (2012). The interaction of chronic restraint
stress and voluntary alcohol intake: Effects on spatial memory in male rats.
Alcohol, 46(5), 499-504.
Grill, H. J., & Norgren, R. (1978). The taste reactivity test. I. mimetic responses to
gustatory stimuli in neurologically normal rats. Brain Research, 143(2), 263-279.
Guttman, N. (1954). Equal-reinforcement values for sucrose and glucose solutions
compared with equal-sweetness values. Journal of Comparative and
Physiological Psychology, 47(5), 358-361.
Hajnal, A., Smith, G. P., & Norgren, R. (2004). Oral sucrose stimulation increases
accumbens dopamine in the rat. American Journal of Physiology. Regulatory,
Integrative and Comparative Physiology, 286(1), R31-R37.
Highfield, D. A., Mead, A. N., Grimm, J. W., Rocha, B. A., & Shaham, Y. (2002).
Reinstatement of cocaine seeking in 129X1/SvJ mice: Effects of cocaine priming,
cocaine cues and food deprivation. Psychopharmacology, 161(4), 417-424.
Hingson, R. W., Heeren, T., & Winter, M. R. (2006). Age of alcohol-dependence onset:
Associations with severity of dependence and seeking treatment.
Pediatrics, 118(3), e755-e763.
Kiefer, S. W., Bice, P. J., & Badia-Elder, N. (1994). Alterations in taste reactivity to
alcohol in rats given continuous alcohol access followed by abstinence.
Alcoholism, Clinical and Experimental Research, 18(3), 555-559.

47
Lanier, S. A., Hayes, J. E., & Duffy, V. B. (2005). Sweet and bitter tastes of alcoholic
beverages mediate alcohol intake in of-age undergraduates. Physiology &
Behavior, 83(5), 821-831.
Lenoir, M., Serre, F., Cantin, L., & Ahmed, S. H. (2007). Intense sweetness surpasses
cocaine reward. PloS One, 2(8), 1.
Levy, A., Limebeer, C. L., Ferdinand, J., Shillingford, U., Parker, L. A., & Leri, F. (2014).
A novel procedure for evaluating the reinforcing properties of tastants in
laboratory rats: Operant intraoral self-administration. Journal of Visualized
Experiments, 84, 1.
Levy, A., Daniels, S., Hudson, R., Horman, T., Flynn, A., Zhou, Y., & Leri, F. (2018).
Bupropion and naltrexone combination alters high fructose corn syrup self-
administration and gene expression in rats. Neuropharmacology, 135, 547-554.
Levy, A., Marshall, P., Zhou, Y., Kreek, M. J., Kent, K., Daniels, S., . . . Leri, F. (2015).
Fructose: Glucose ratios-A study of sugar self-administration and associated
neural and physiological responses in the rat. Nutrients, 7(5), 3869-3890.
Madsen, H. B., & Ahmed, S. H. (2015). Drug versus sweet reward: Greater attraction to
and preference for sweet versus drug cues. Addiction Biology, 20(3), 433-444.
Mandillo, S., Titchen, K., & Miczek, K. A. (1998). Ethanol drinking in socially housed
squirrel monkeys. Behavioural Pharmacology, 9(4), 363-367.
Mathurin, P., & Deltenre, P. (2009). Effect of binge drinking on the liver: An alarming
public health issue? Gut, 58(5), 613-617.

48
Mantsch, J. R., Baker, D. A., Funk, D., Lê, A.,D., & Shaham, Y. (2016). Stress-induced
reinstatement of drug seeking: 20 years of progress.
Neuropsychopharmacology, 41(1), 335-356.
Massey, V. L., & Arteel, G. E. (2012). Acute alcohol-induced liver injury. Frontiers in
Physiology, 3, 193.
McGhee, N. K., Jefferson, L. S., & Kimball, S. R. (2009). Elevated corticosterone
associated with food deprivation upregulates expression in rat skeletal muscle of
the mTORC1 repressor, REDD1. Journal of Nutrition, 139(5), 828-834.
McGrath, E., Jones, A., & Field, M. (2016). Acute stress increases ad-libitum alcohol
consumption in heavy drinkers, but not through impaired inhibitory
control.Psychopharmacology, 233(7), 1227-1234.
Metzner, C., & Kraus, L. (2008). The impact of alcopops on adolescent drinking: A
literature review. Alcohol and Alcoholism, 43(2), 230-239.
Meyers, A. M., Mourra, D., & Beeler, J. A. (2017). High fructose corn syrup induces
metabolic dysregulation and altered dopamine signaling in the absence of
obesity. PLoS One, 12(12).
Mosher, J. F., & Johnsson, D. (2005). Flavored alcoholic beverages: An international
marketing campaign that targets youth. Journal of Public Health Policy, 26(3),
326-342.
Pohorecky, L. A. (1990). Interaction of ethanol and stress: Research with experimental
animals -- an update. Alcohol Alcoholism., 25(2-3), 263-276.
Rada, P., Avena, N. M., & Hoebel, B. G. (2005). Daily bingeing on sugar repeatedly
releases dopamine in the accumbens shell. Neuroscience, 134(3), 737-744.

49
Reilly, S. (1999). Reinforcement value of gustatory stimuli determined by progressive
ratio performance. Pharmacology, Biochemistry, and Behavior, 63(2), 301-311.
Samson, H. H., Pfeffer, A. O., & Tolliver, G. A. (1988). Oral ethanol self-administration
in rats: Models of alcohol-seeking behavior. Alcoholism, Clinical and
Experimental Research, 12(5), 591-598.
Sclafani, A., & Ackroff, K. (2017). Flavor preferences conditioned by nutritive and non-
nutritive sweeteners in mice. Physiology & Behavior, 173, 188-199.
Sclafani, A., & Ackroff, K. (2003). Reinforcement value of sucrose measured by
progressive ratio operant licking in the rat. Physiology & Behavior, 79(4-5), 663-
670.
Sclafani, A., Bodnar, R. J., & Delamater, A. R. (1998). Pharmacology of food
conditioned preferences. Appetite, 31(3), 406-p. 406.
Sclafani, A., Glass, D. S., Margolskee, R. F., & Glendinning, J. I. (2010). Gut T1R3
sweet taste receptors do not mediate sucrose-conditioned flavor preferences in
mice. American Journal of Physiology. Regulatory, Integrative and Comparative
Physiology, 299(6), R1643-R1650.
Schebendach, J., Klein, D. A., Mayer, L. E. S., Attia, E., Devlin, M. J., Foltin, R. W., &
Walsh, B. T. (2017). Assessment of the motivation to use artificial sweetener
among individuals with an eating disorder. Appetite, 109, 131-136.
Shaham, Y., Erb, S., & Stewart, J. (2000). Stress-induced relapse to heroin and cocaine
seeking in rats: A review. Brain Research Reviews, 33(1), 13-33.

50
Shalev, U., Marinelli, M., Baumann, M. H., Piazza, P., & Shaham, Y. (2003). The role of
corticosterone in food deprivation-induced reinstatement of cocaine seeking in
the rat. Psychopharmacology, 168(1-2), 170-176.
Sinha, R. (2008). Chronic stress, drug use, and vulnerability to addiction. Annals of the
New York Academy of Sciences, 1141(1), 105-130.
Sprague, J. E., & Maickel, R. P. (1994). Effects of stress and ebiratide (hoe-427) on
free-choice ethanol consumption: Comparison of lewis and sprague-dawley
rats. Life Sciences, 55(11), 873-878.
Stanhope, K. L., Medici, V., Bremer, A. A., Lee, V., Lam, H. D., Nunez, M. V., . . . Havel,
P. J. (2015). A dose-response study of consuming high-fructose corn syrup-
sweetened beverages on lipid/lipoprotein risk factors for cardiovascular disease
in young adults. American Journal of Clinical Nutrition, 101(6), 1144-1154.
Steinberg, E. E., Boivin, J. R., Saunders, B. T., Witten, I. B., Deisseroth, K., & Janak, P.
H. (2014). Positive reinforcement mediated by midbrain dopamine neurons
requires D1 and D2 receptor activation in the nucleus accumbens. PLoS
One, 9(4).
Tatzer, E., Schubert, M. T., Timischl, W., & Simbruner, G. (1985). Discrimination of
taste and preference for sweet in premature babies. Early Human
Development, 12(1), 23-30.
Thai, P., Tan, E., Tan, W., Tey, T., Kaur, H., & Say, Y. (2011). Sweetness intensity
perception and pleasantness ratings of sucrose, aspartame solutions and cola
among multi-ethnic malaysian subjects. Food Quality and Preference, 22(3), 281-
289.

51
Thomas, S. E., Bacon, A. K., Randall, P. K., Brady, K. T., & See, R. E. (2011). An acute
psychosocial stressor increases drinking in non-treatment-seeking
alcoholics. Psychopharmacology, 218(1), 19-28.
Tolliver, G. A., Sadeghi, K. G., & Samson, H. H. (1988). Ethanol preference following
the sucrose-fading initiation procedure. Alcohol, 5(1), 9-13.
Vengeliene, V., Siegmund, S., Singer, M. V., Sinclair, J. D., Li, T., & Spanagel, R.
(2003). A comparative study on alcohol-preferring rat lines: Effects of deprivation
and stress phases on voluntary alcohol intake. Alcoholism, Clinical and
Experimental Research, 27(7), 1048-1054.
Ventura, A. K., & Mennella, J. A. (2011). Innate and learned preferences for sweet taste
during childhood. Current Opinion in Clinical Nutrition and Metabolic Care, 14(4),
379-384.
Wheeler, R. A., Aragona, B. J., Fuhrmann, K. A., Jones, J. L., Day, J. J., Cacciapaglia,
F., . . . Carelli, R. M. (2011). Cocaine cues drive opposing context-dependent
shifts in reward processing and emotional state. Biological Psychiatry, 69(11),
1067-1074.
White, N. M., & Carr, G. D. (1985). The conditioned place preference is affected by two
independent reinforcement processes. Pharmacology, Biochemistry, and
Behavior, 23(1), 37-42.
World Health Organization. (2009). Global health risks: mortality and burden of disease
attributable to selected major risks. Geneva, World Health Organization.
World Health Organization. (2014). Global status report on alcohol and health. Geneva,
World Health Organization.

52
Wyvell, C. L., & Berridge, K. C. (2000). Intra-accumbens amphetamine increases the
conditioned incentive salience of sucrose reward: Enhancement of reward
"wanting" without enhanced "liking" or response reinforcement. Journal of
Neuroscience, 20(21), 8122-8130.
Yasoshima, Y., & Shimura, T. (2015). A mouse model for binge-like sucrose
overconsumption: Contribution of enhanced motivation for sweetener
consumption. Physiology & Behavior, 138, 154-164.

53
Figure Legends
Figure 1 A. Timeline of procedural events that occurred during Experiment 1. Rats
could self-administer 5% EtOH under multiple schedules of reinforcement, then 25%
HFCS was added to the solution. Following this, a dose-response curve was created by
varying the concentration of EtOH within HFCS, then rats reverted to self-administering
5% EtOH. B. Mean (SEM) infusions earned across 25 days of IO SA of 5% EtOH
(sessions 1 -24) and 5% EtOH + 25% HFCS (session 25). The labels below the x-axis
refer to the schedule of reinforcement. The *** indicates a significance increase from all
IO SA sessions of 5% EtOH, p <0.001. C. Mean (SEM) infusions earned across
sessions 26 to 45 during which the concentration of EtOH in the 25% HFCS solution
was manipulated to create a dose-response (intake) curve: 5%, 10%, 20%, 5%. Animals
were tested on a FR3 schedule of reinforcement. The *** indicates a significance
decrease from sessions 33 (5%), 37 (10%) and 45 (5%), p <0.001. D. Mean (SEM)
infusions rats earned on the last IO SA acquisition day of 5% EtOH prior to the addition
of HFCS (session 20), compared to the four sessions following HFCS removal
(sessions 46 – 49). The * indicates a significant increase from Pre-HFCS (session 20), p
<0.05. The ** indicates a significant increase from Pre-HFCS, p< 0.01.
Figure 2 A. Timeline of procedural events that occurred during Experiment 2. Rats
could self-administer 5% EtOH in 0.1% SACC. Following this, a dose-response curve
was created by varying the concentration of EtOH within SACC. On the final session,
rats could self-administer 25% HFCS. B. Mean (SEM) infusions earned across 17
sessions of IO SA of 5% EtOH in 0.1% SACC. The labels below the x-axis refer to the
schedule of reinforcement. C. Mean (SEM) infusions earned during acquisition for 5%

54
EtOH (data taken from Experiment 1; session 20) compared to 5% EtOH + 0.1% SACC
(data taken from Experiment 2; session 17). D. Mean (SEM) infusions earned across
sessions 14 - 29, during which the concentration of EtOH in 0.1% SACC solution was
manipulated to create a dose-response (intake) curve: 5%, 10%, 20%, 5%. The ***
indicates a significance difference from session 17 (5% EtOH). E. Mean (SEM)
infusions rats earned on the last day each concentration (5%, 10%, 20%, 5%) of EtOH
was presented in the SACC solution and in the HFCS solution (Experiment 1) during the
dose-response (intake) curve. The *** indicates a significance difference from Exp 1 to
Exp 2 solutions.
Figure 3 A. Timeline of procedural events that occurred during Experiment 3. Rats
could self-administer 10% EtOH in HFCS during acquisition, then this behaviour was
extinguished by replacing sweetened EtOH reinforcement with reverse osmosis water
deliveries. Following extinction, a cue-reinstatement test was used to test drug-seeking
for the drug-paired cue light. Following this, two resumption tests were used to test the
resumption of 10% EtOH and 25% HFCS (across two counterbalanced sessions) under
conditions of food restriction and food deprivation. B. Mean (SEM) infusions earned
across 17 days of IO SA of 10% EtOH in 25% HFCS. The labels below the x-axis refer
to the schedule of reinforcement. The * indicates a significant difference from session 1.
C. Mean (SEM) active lever responses made on the last day of IO SA acquisition
(Baseline; B) and the following five extinction sessions. The * indicates a significant
decrease from Baseline, p < 0.001. The # indicates a significant decrease from Ext1, p
<0.001. D. Mean (SEM) active lever responses made on the tests of cue reinstatement
between food restricted and food deprived rats. The * indicates a significant increase in

55
food restricted relative to food deprived rats. E. Mean (SEM) infusions earned during the
tests of EtOH and HFCS resumption across food restricted and food deprived rats. The
** indicates a significant difference from extinction responding within stressed rats, p <
0.01. The # indicates a significant difference from cue-reinstatement responding within
non-stressed animals.
Figure 4 A. Timeline of procedural events that occurred during Experiment 4. Each
taste reactivity (TR) session tested EtOH and HFCS palatability separately over a two-
min liquid exposure. Please see Figure 3 A for a more detailed explaoination of the IO
SA protocol used. B. Mean (SEM) infusions earned during the test of 10% EtOH + 25%
HFCS resumption across food restricted and food deprived rats. The * indicates a
significant difference from food restriction conditions, p < 0.05. C. Mean (SEM) hedonic
taste responses elicited to EtOH and HFCS during baseline (pre-acquisition) and post-
acquisition taste reactivity testing. The * indicates a significant increase from baseline
responding, p < 0.05. The *** indicates a significant increase from baseline responding,
p < 0.001. D. Mean (SEM) hedonic taste responses elicited to EtOH and HFCS across
food restricted and food deprived rats. The *** indicates a significant increase from food
restriction conditions, p < 0.001.

56
Figure 1.
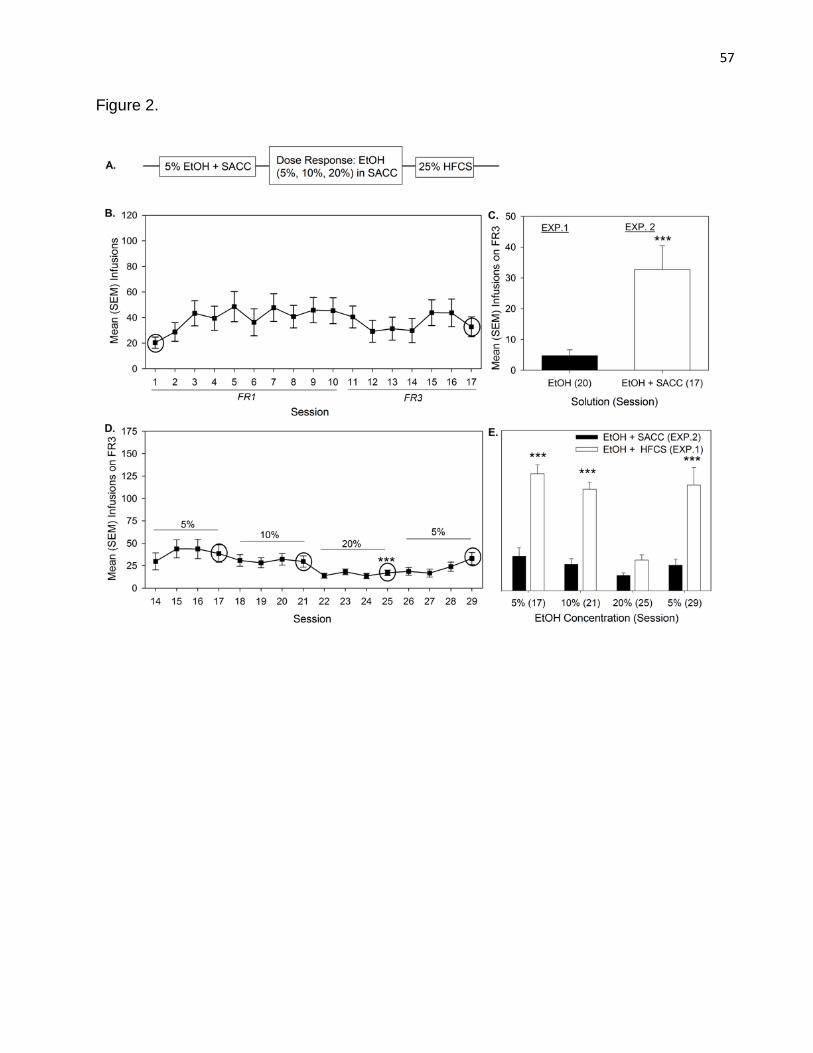
57
Figure 2.

58
Figure 3.

59
Figure 4.

60
General Discussion
Summary of results
Excessive alcohol consumption by young, naive drinkers is prevelant and has
been associated with an increased risk for developing alcohol use disorders later in life
(Chassin, Pitts, & Prost, 2002; Albers et al., 2015). Furthermore, the addition of
sweeteners to alcoholic beverages have been implicated in increasing alcohol intake of
naïve drinkers (Albers et al., 2015). Thus, the current study sought to investigate
whether the addition of HFCS to an EtOH solution could alter EtOH’s intake, hedonic
taste, and sensitivity to food deprivation stress in naïve rats. HFCS caused a drastic
increase in EtOH intake across multiple concentrations; an effect that was not observed
by the addition of a non-caloric sweetener (SACC). HFCS also altered rat’s sensitivity to
the effect of food deprivation stress on EtOH consumption. More specifically, food
deprivation stress increased EtOH intake only when HFCS was mixed in the solution. In
terms of EtOH and HFCS palatability, our study revealed two important findings. First,
multiple exposures to 10% EtOH in 25% HFCS increased the palatability of 10% EtOH
and 25% HFCS. Second, food deprivation stress increased the palatability of HFCS, but
did not alter the palatability of EtOH. Taken together with the results from self-
administration, we suggest that the increased palatability of HFCS under food
deprivation stress may contribute to the increased motivation to seek HFCS under
stressful operant conditions.
Methodological limitations
One methodological limitation to consider when interpreting the results of the
presented study is the selective use of male rats throughout all experiments. Male rats

61
were chosen because our laboratory has already established the consumption patterns
of HFCS in the home-cage and during operant IO SA. This limits the generalizability of
our findings since differences in alcohol intake and alcohol preferences have been
reported across male and female rodents. Generally, females tend to consume larger
quantities, and demonstrate a higher preference for alcohol in comparison to males
(Lancaster & Spiegel, 1992; Lancaster, Brown, Coker, Elliot, & Wren, 1996). In
corroboration with these behavioural findings, female rats show increased
responsiveness to mesolimbic dopamine stimulation by EtOH exposure (Blanchard,
Steindorf, Wang, & Glick, 1993). Human research has also indicated that women reach
a higher peak blood alcohol concentration (BAC) than men after consuming equivalent
amounts of EtOH (Sutker, Tabakoff, Goist, & Randall, 1983). These findings have been
suggested to reflect the differences in EtOH pharmacokinetics between the two sexes
(Taylor, Dolhert, Friedman, Mumenthaler, Yesavage, 1996; Mumenthaler, Taylor,
O’Hara, Yesavage, 1999).
In addition to differences in alcohol intake and preference, the effect of stress on
EtOH intake might also differ between female and male rodents. For example, restraint
stress has been shown to increase homecage EtOH consumption in male mice, while
decreasing consumption in females (Chester, Gustavo, Demaria, & Finegan, 2006).
Taken together, these results highlight the importance of using both males and females
while investigating rodent models of EtOH intake.
Although our laboratory has frequently studied the reinforcing properties of
HFCS, the current experiments represent our first attempt to assess ethanol
reinforcement. As such, our laboratory was not adequately prepared to measure the

62
blood alcohol concentration (BAC) of rats throughout IO SA. During operant SA of
EtOH in rats, BACs are frequently assessed in addition to the total amount of EtOH
consumed (Doherty & Gonzales, 2015; Carnicella, Yowell, & Ron, 2011). Using BACs
as a biomarker of EtOH consumption is necessary to provide a measure of EtOH
exposure that accounts for other factors that have been shown to affect EtOH
consumption in animals (i.e. patterns of drinking, metabolism, sex differences). Thus,
our experiments could have drawn stronger conclusions about the effects of HFCS on
EtOH consumption if BACs were presented along with consumption data. However,
when the consumption patterns of animals responding for 10% EtOH in 25% HFCS
were calculated in ten-minute time bins (Experiment 1), rats consumed 0.91 g/kg within
the first 30 minutes of each session. Briones and Woods (2013) found that home-cage
self-administration of approximately 0.75 g/kg EtOH in rats produced BACs greater than
0.08 g%, which constitutes as a binge consumption pattern according to the National
institute of Alcohol Abuse and Alcoholism (2015). Therefore, although the current study
lacked measures of BACs throughout the IO SA of EtOH + HFCS, we are confident that
animals consumed quantities of EtOH that produced recognized levels of binge
intoxication.
Future directions
This study could be expanded upon by future research. As previously mentioned,
one limitation of the presented experiments is the inability to draw conclusions about
how HFCS changes the relative reinforcing value of EtOH. A particularly useful way to
measure the reinforcing value of a stimulus while using an operant self-administration
model is by applying a progressive-ratio schedule of reinforcement. When a

63
progressive-ratio schedule is applied, the amount of effort required to earn
reinforcement successively increases across each trial, until responding for the stimulus
ceases and a ‘breakpoint’ has been achieved (Jones & Comer, 2013; Panlilio &
Goldberg, 2007). Given that breakpoints quantify an organism’s persistence/desire to
achieve reinforcement, with higher breakpoints associated with higher persistence, they
can be used to indicate the relative reinforcing value of different stimuli. Furthermore,
the application of a progressive ratio schedule also stabilizes possible confounds
between solutions used during operant self-administration such as, levels of satiety or
intoxication.
Choice procedures are another method that can be employed to delineate
changes in a stimulus’ reinforcing value; in fact, some researchers argue choice
procedures are the only method by which changes in the reinforcing value of a stimulus
can be determined (Panlilio & Goldberg, 2007). During choice testing, participants are
required to choose between a fixed magnitude of the particular reinforcer of interest, or
a scaling amount of an alternative reinforcer. The point at which the participant stops
choosing the reinforcer of interest over the alternative reinforcer is then used to
compare the relative reinforcing value of the reinforcer of interest (in comparison to the
alternative).
Although the use of progressive ratio or choice testing could allow us to make
stronger inferences about changes in the reinforcing efficacy of EtOH, both procedures
would be complicated to employ as rats do not self-administer reliable amount of EtOH
without the use of sucrose-fading or lengthy intitiation procedures (as reviewed by
Samson, 1986; Samson, Pfeffer, & Tolliver, 1988). Ultimetly the use of sucrose-fading

64
would confound the variable of interest; that is, changes in EtOH reinforcing efficacy as
a result of HFCS pairings. Furthermore, alternate initiation procedures could take
upwards of 6-weeks, and do not lead to strong responding for EtOH alone. Therefore,
although our lab has considered alternate ways to investigate the changes in EtOH’s
reinforcing value, there are multiple limitations in using these methods to measure a
reinforcer that is not reliably self-administered on its own (e.x. EtOH).
Our laboratory is also interested in examining the role of arginine vasopressin in
EtOH intake, palatability and persistence of responding under conditions of food
deprivation stress. Arginine vasopressin (AVP) is synthesized in the supraoptic nucleus
of the hypothalamus and acts as a key regulator of the hypothalamic-pituitary-adrenal
(HPA) axis, the body’s central stress response system (Aguilera & Rabadan-Diehl,
2000; Tanoue et al., 2004). More specifically, following exposure to stress, AVP binds to
V1b receptors (V1bR) in the anterior pituitary to promote the secretion of
adrenocorticotropic hormone (ACTH) into the bloodstream. ACTH subsequently travels
down to the adrenal cortex to promote the synthesis and release of glucocorticoids (see
Beurel & Nemeroff, 2014 for a review). Correspondingly, V1bR antagonists decrease
multiple anxiogenic behaviours in animal models (Griebel et al., 2002; Stemmelin,
Lukovic, Salome, & Griebel., 2005).
Given that AVP modulates anxiogenic responding, it may contribute to stress-
induced relapse. Indeed, AVP secretion is heightened during periods of drug withdrawal
in humans and animals. For example, Eisenhofer, Lambie, Whiteside, and Johnson
(1985) measured AVP levels in alcoholic patients during a drug detoxification program.
Patients displaying withdrawal symptoms had significantly elevated urine and plasma

65
AVP levels in comparison to patients without withdrawal symptomology, or healthy
controls. Furthermore, rats show heightened AVP mRNA gene expression during
periods of acute heroin and cocaine withdrawal (Zhou, Leri, Cummins, Hoeschele, &
Kreek, 2008; Zhou et al., 2005).
There are few studies that have investigated the effect of V1bR antagonists on
relapse-like responding in animals. Zhou, Leri, Cummins, Hoeschele, and Kreek (2008)
found that the administration of a V1bR antagonist (1 mg/kg and 30 mg/kg;
SSR149415) decreased stress induced relapse-like responding for heroin in rats.
Interestingly, the function of AVP in modulating relapse-like responding for alcohol is
unknown. It would be interesting to investigate whether the V1bR antagonist
SSR149415 would decrease stress-induced increases in resumption responding for
EtOH + HFCS and HFCS.
Conclusions
In conclusion, results from Experiment 1 and Experiment 2 suggest that the
addition of a sweet and calorically dense sweetener (HFCS) drastically elevated EtOH
intake in rats, in comparison to an equally palatable, yet non-caloric sweetener.
Furthermore, when HFCS was removed from the EtOH solution, rats demonstrated
increased EtOH intake levels then what was observed prior to the HFCS pairings.
Finally, Experiment 3 and Experiment 4 demonstrated that food deprivation stress only
increased EtOH intake when HFCS was mixed into the solution. Furthermore, results
from taste reactivity measures suggest that food deprivation stress causes increase in
HFCS palatability, which may explain the observed self-administration data. Taken
together, our results suggest that HFCS increases EtOH intake under normal and

66
stressful conditions, and should be reconsidered as an acceptable addition to alcoholic
beverages.

67
References Ackroff, K., & Sclafani, A. (2011). Flavor preferences conditioned by post-oral infusion of
monosodium glutamate in rats. Physiology & Behavior, 104(3), 488-494.
Albers, A. B., Siegel, M., Ramirez, R. L., Ross, C., DeJong, W., & Jernigan, D. H.
(2015). Flavored alcoholic beverage use, risky drinking behaviors, and adverse
outcomes among underage drinkers: Results from the ABRAND study. American
Journal of Public Health, 105(4), 810-815.
Agmo, A., & Marroquin, E. (1997). Role of gustatory and postingestive actions of
sweeteners in the generation of positive affect as evaluation by place preference
conditioning. Appetite, 29(3), 269-289.
Aguilera, G., & Rabadan-Diehl, C. (2000). Vasopressinergic regulation of the
hypothalamic-pituitary-adrenal axis: Implications for stress
adaptation. Regulatory Peptides, 96(1-2), 23-29.
Anderson, D. A., Shapiro, J. R., Lundgren, J. D., Spataro, L. E., & Frye, C. A. (2002).
Self-reported dietary restraint is associated with elevated levels of salivary
cortisol. Appetite, 38(1), 13-17.
Archie, S., Zangeneh-Kazemi, A., & Akhtar-Danesh, N. (2012). Concurrent binge
drinking and depression among canadian youth: Prevalence, patterns, and
suicidality. Alcohol, 46(2), 165-72.
Avena, N. M., Long, K. A., & Hoebel, B. G. (2005). Sugar-dependent rats show
enhanced responding for sugar after abstinence: Evidence of a sugar deprivation
effect. Physiology & Behavior, 84(3), 359-362.

68
Barlow, P., McKee, Basu, & Stuckler, (2017). Impact of the north american free trade
agreement on high-fructose corn syrup supply in canada: A natural experiment
using synthetic control methods. Canadian Medical Association.Journal, 189(26),
E881-E887.
Beurel, E., & Nemeroff, C. B. (2014). Interaction of stress, corticotropin-releasing factor,
arginine vasopressin and behaviour. Current Topics in Behavioral
Neurosciences, 18, 67-80.
Bienkowski, P., Koros, E., Kostowski, W., & Bogucka-Bonikowska, A. (2000).
Reinstatement of ethanol seeking in rats: Behavioral analysis. Pharmacology
Biochemistry and Behavior, 66(1), 123-128.
Black, J., Belluzzi, J. D., & Stein, L. (1985). Reinforcement delay of one second
severely impairs acquisition of brain self-stimulation. Brain Research, 359(1-2),
113-119.
Blanchard, B. A., Steindorf, S., Wang, S., & Glick, S. D. (1993). Sex differences in
ethanol-induced dopamine release in nucleus accumbens and in ethanol
consumption in rats. Alcoholism, Clinical and Experimental Research, 17(5), 968-
973.
Bouton, M. E., Winterbauer, N. E., & Todd, T. P. (2012). Relapse processes after the
extinction of instrumental learning: Renewal, resurgence, and reacquisition.
Behavioural Processes, 90(1), 130-141.
Bray, G. A., Nielsen, S. J., & Popkin, B. M. (2004). Consumption of high-fructose corn
syrup in beverages may play a role in the epidemic of obesity. The American
Journal of Clinical Nutrition, 79(4), 537-543.

69
Bray, G. A. (2013). Energy and fructose from beverages sweetened with sugar or high-
fructose corn syrup pose a health risk for some people. Advances in
Nutrition, 4(2), 220-225.
Briones, T. L., & Woods, J. (2013). Chronic binge-like alcohol consumption in
adolescence causes depression-like symptoms possibly mediated by the effects
of BDNF on neurogenesis. Neuroscience, 254, 324-334.
Brown, S. A., Vik, P. W., McQuaid, J. R., Patterson, T. L., Irwin, M. R., & Grant, I.
(1990). Severity of psychosocial stress and outcome of alcoholism
treatment. Journal of Abnormal Psychology, 99(4), 344-348.
Brown, S. A., Vik, P. W., Patterson, T. L., Grant, I., & Schuckit, M. A. (1995). Stress,
vulnerability and adult alcohol relapse. Journal of Studies on Alcohol, 56(5), 538-
545.
Bulik, C. M., & Brinded, E. C. (1993). The effect of food deprivation on alcohol
consumption in bulimic and control women. Addiction, 88(11), 1545-1551.
Carnicella, S., Yowell, Q. V., & Ron, D. (2011). Regulation of operant oral ethanol self-
administration: A dose-response curve study in rats. Alcoholism, Clinical and
Experimental Research, 35(1), 116-125.
Carr, K. D. (2007). Chronic food restriction: Enhancing effects on drug reward and
striatal cell signaling. Physiology & Behavior, 91(5), 459-472.
Carroll, M. E. (1985). The role of food deprivation in the maintenance and reinstatement
of cocaine-seeking behavior in rats. Drug and Alcohol Dependence, 16(2), 95-
109.

70
Chassin, L., Pitts, S. C., & Prost, J. (2002). Binge drinking trajectories from adolescence
to emerging adulthood in a high-risk sample: Predictors and substance abuse
outcomes. Journal of Consulting and Clinical Psychology, 70(1), 67-78.
Chaudhri, N., Sahuque, L. L., Cone, J. J., & Janak, P. H. (2008). Reinstated ethanol-
seeking in rats is modulated by environmental context and requires the nucleus
accumbens core. European Journal of Neuroscience, 28(11), 2288-2298.
Chester, J. A., Gustavo de, P. B., DeMaria, A., & Finegan, A. (2006). Different effects of
stress on alcohol drinking behaviour in male and female mice selectively bred for
high alcohol preference. Alcohol and Alcoholism: International Journal of the
Medical Council on Alcoholism, 41(1), 44.
Cooney, N. L., Litt, M. D., Morse, P. A., Bauer, L. O., & Gaupp, L. (1997). Alcohol cue
reactivity, negative-mood reactivity, and relapse in treated alcoholic men. Journal
of Abnormal Psychology, 106(2), 243-250.
Copeland, J., Stevenson, R. J., Gates, P., & Dillon, P. (2007). Young australians and
alcohol: The acceptabllity of ready-to-drink (RTD) alcoholic beverages among 12-
30-year-olds. Addiction, 102(11), 1740-1746.
Crombag, H. S., Bossert, J. M., Koya, E., & Shaham, Y. (2008). Context-induced
relapse to drug seeking: a review. Philosophical Transactions of the Royal
Society of London. Series B, Biological Sciences, 363(1507), 3233-3243.
Daniels, S., Marshall, P., & Leri, F. (2016). Alterations of naltrexone-induced
conditioned place avoidance by pre-exposure to high fructose corn syrup or
heroin in sprague-dawley rats. Psychopharmacology, 233(3), 425-433.

71
D'Cunha, T.,M., Sedki, F., Macri, J., Casola, C., & Shalev, U. (2013). The effects of
chronic food restriction on cue-induced heroin seeking in abstinent male rats.
Psychopharmacology, 225(1), 241-250.
DeWit, D. J., Adlaf, E. M., Offord, D. R., & Ogborne, A. C. (2000). Age at first alcohol
use: A risk factor for the development of alcohol disorders. The American Journal
of Psychiatry, 157(5), 745-750.
Dietze, S., Lees, K. R., Fink, H., Brosda, J., & Voigt, J. (2016). Food deprivation, body
weight loss and anxiety-related behavior in rats. Animals, 6(1), 4.
Doherty, J. M., & Gonzales, R. A. (2015). Operant self-administration of sweetened
ethanol and time course of blood ethanol levels in adolescent and adult male
long-evans rats. Alcoholism, Clinical and Experimental Research, 39(3), 485-
495.
Domingos, A. I., Vaynshteyn, J., Voss, H. U., Ren, X., Gradinaru, V., Zang, F., . . .
Friedman, J. (2011). Leptin regulates the reward value of nutrient. Nature
Neuroscience, 14(12), 1562-8.
Eisenhofer, G., Lambie, D. G., Whiteside, E. A., & Johnson, R. H. (1985). Vasopressin
concentrations during alcohol withdrawal. British Journal of Addiction, 80(2), 195-
199.
Fidler, T. L., Clews, T. W., & Cunningham, C. L. (2006). Reestablishing an intragastric
ethanol self-infusion model in rats. Alcoholism, Clinical and Experimental
Research, 30(3), 414-428.

72
Filbey, F. M., Claus, E., Audette, A. R., Niculescu, M., Banich, M. T., Tanabe, J., . . .
Hutchison, K. E. (2008). Exposure to the taste of alcohol elicits activation of the
mesocorticolimbic neurocircuitry. Neuropsychopharmacology, 33(6), 1391-1401.
Fox, H. C., Bergquist, K. L., Hong, K., & Sinha, R. (2007). Stress-induced and alcohol
cue-induced craving in recently abstinent alcohol-dependent
individuals. Alcoholism, Clinical and Experimental Research, 31(3), 395-403.
Frank, G. K. W., Oberndorfer, T. A., Simmons, A. N., Paulus, M. P., Fudge, J. L., Yang,
T. T., & Kaye, W. H. (2008). Sucrose activates human taste pathways differently
from artificial sweetener. NeuroImage, 39(4), 1559-1569.
Garland, E. L., Franken, I. H. A., & Howard, M. O. (2012). Cue-elicited heart rate
variability and attentional bias predict alcohol relapse following treatment.
Psychopharmacology, 222(1), 17-26.
Gass, J. T., & Olive, M. F. (2007). Reinstatement of ethanol-seeking behavior following
intravenous self-administration in wistar rats. Alcoholism, Clinical and
Experimental Research, 31(9), 1441-1445.
Griebel, G., Simiand, J., Serradeil-Le Gal, C., Wagnon, J., Pascal, M., Scatton, B., . . .
Soubrie, P. (2002). Anxiolytic- and antidepressant-like effects of the non-peptide
vasopressin V sub(1b) receptor antagonist, SSR149415, suggest an innovative
approach for the treatment of stress-related disorders. Proceedings of the
National Academy of Sciences, 99(9), 6370-6375.
Gruber, E., DiClemente, R. J., Anderson, M. M., & Lodico, M. (1996). Early drinking
onset and its association with alcohol use and problem behavior in late
adolescence. Preventive Medicine, 25(3), 293-300.

73
Guttman, N. (1954). Equal-reinforcement values for sucrose and glucose solutions
compared with equal-sweetness values. Journal of Comparative and
Physiological Psychology, 47(5), 358-361.
Hajnal, A., Smith, G. P., & Norgren, R. (2004). Oral sucrose stimulation increases
accumbens dopamine in the rat. American Journal of Physiology.Regulatory,
Integrative and Comparative Physiology, 286(1), R31-R37.
Hawkins, J. D., Graham, J. W., Maguin, E., Abbott, R., Hill, K. G., & Catalano, R. F.
(1997). Exploring the effects of age of alcohol use initiation and psychosocial risk
factors on subsequent alcohol misuse. Journal of Studies on Alcohol, 58(3), 280-
290.
Higley, A. E., Crane, N. A., Spadoni, A. D., Quello, S. B., Goodell, V., & Mason, B. J.
(2011). Craving in response to stress induction in a human laboratory paradigm
predicts treatment outcome in alcohol-dependent
individuals. Psychopharmacology, 218(1), 121-129.
Highfield, D. A., Mead, A. N., Grimm, J. W., Rocha, B. A., & Shaham, Y. (2002).
Reinstatement of cocaine seeking in 129X1/SvJ mice: Effects of cocaine priming,
cocaine cues and food deprivation. Psychopharmacology, 161(4), 417-424.
Hingson, R. W., Heeren, T., & Winter, M. R. (2006). Age of alcohol-dependence onset:
Associations with severity of dependence and seeking treatment.
Pediatrics, 118(3), e755-e763.
Hogenkamp, P. S., Shechter, A., St-Onge, M., Sclafani, A., & Kissileff, H. R. (2017). A
sipometer for measuring motivation to consume and reward value of foods and

74
beverages in humans: Description and proof of principle. Physiology &
Behavior, 171, 216-227.
Horvitz, J. C. (2000). Mesolimbocortical and nigrostriatal dopamine responses to salient
non-reward events. Neuroscience, 96(4), 651-656.
Jin, H., Rourke, S. B., Patterson, T. L., Taylor, M. J., & Grant, I. (1998). Predictors of
relapse in long-term abstinent alcoholics. Journal of Studies on Alcohol, 59(6),
640-646.
Katner, S. N., Magalong, J. G., & Weiss, F. (1999). Reinstatement of alcohol-seeking
behavior by drug-associated discriminative stimuli after prolonged extinction in
the rat. Neuropsychopharmacology, 20(5), 471-479.
Knight, C. P., Hauser, S. R., Deehan, G. A., Toalston, J. E., McBride, W. J., & Rodd, Z.
A. (2016). Oral conditioned cues can enhance or inhibit ethanol (EtOH)-seeking
and EtOH-relapse drinking by alcohol-preferring (P) rats. Alcoholism, Clinical and
Experimental Research, 40(4), 906-915.
Lancaster, F. E., Brown, T. D., Coker, K. L., Elliott, J. A., & Wren, S. B. (1996). Sex
differences in alcohol preference and drinking patterns emerge during the early
postpubertal period. Alcoholism, Clinical and Experimental Research, 20(6),
1043-1049.
Lancaster, F. E., & Spiegel, K. S. (1992). Sex differences in pattern of drinking.Alcohol
(Fayetteville, N.Y.), 9(5), 415-420.
Lanier, S. A., Hayes, J. E., & Duffy, V. B. (2005). Sweet and bitter tastes of alcoholic
beverages mediate alcohol intake in of-age undergraduates. Physiology &
Behavior, 83(5), 821-831.

75
Le, A. D., Harding, S., Juzytsch, W., Funk, D., & Shaham, Y. (2005). Role of alpha-2
adrenoceptors in stress-induced reinstatement of alcohol seeking and alcohol
self-administration in rats. Psychopharmacology, 179(2), 366-373.
Leeman, R. F., O'Malley, S.,S., White, M. A., & McKee, S. A. (2010). Nicotine and food
deprivation decrease the ability to resist smoking. Psychopharmacology, 212(1),
25-32.
Lenoir, M., Serre, F., Cantin, L., & Ahmed, S. H. (2007). Intense sweetness surpasses
cocaine reward. PloS One, 2(8), 1.
Levy, A., Limebeer, C. L., Ferdinand, J., Shillingford, U., Parker, L. A., & Leri, F. (2014).
A novel procedure for evaluating the reinforcing properties of tastants in
laboratory rats: Operant intraoral self-administration. Journal of Visualized
Experiments, 84, 1.
Levy, A., Marshall, P., Zhou, Y., Kreek, M. J., Kent, K., Daniels, S., . . . Leri, F. (2015).
Fructose: Glucose ratios-A study of sugar self-administration and associated
neural and physiological responses in the rat. Nutrients, 7(5), 3869-3890.
Levy, A., Daniels, S., Hudson, R., Horman, T., Flynn, A., Zhou, Y., & Leri, F. (2018).
Bupropion and naltrexone combination alters high fructose corn syrup self-
administration and gene expression in rats. Neuropharmacology, 135, 547-554.
Liu, X., & Weiss, F. (2002). Additive effect of stress and drug cues on reinstatement of
ethanol seeking: Exacerbation by history of dependence and role of concurrent
activation of corticotropin-releasing factor and opioid mechanisms. The Journal of
Neuroscience, 22(18), 7856-7861.

76
Maccioni, P., Orru, A., Korkosz, A., Gessa, G. L., Carai, M. A. M., Colombo, G., &
Bienkowski, P. (2007). Cue-induced reinstatement of ethanol seeking in
sardinian alcohol-preferring rats. Alcohol, 41(1), 31-39.
Madsen, H. B., & Ahmed, S. H. (2015). Drug versus sweet reward: Greater attraction to
and preference for sweet versus drug cues. Addiction Biology, 20(3), 433-444.
Martin, R., & Weiss, F. (2013). Modeling relapse in animals. Current Topics in
Behavioral Neurosciences, 13, 403-432.
Meyers, A. M., Mourra, D., & Beeler, J. A. (2017). High fructose corn syrup induces
metabolic dysregulation and altered dopamine signaling in the absence of
obesity. PLoS One, 12(12).
McLean, J. A., Barr, S. I., & Prior, J. C. (2001). Cognitive dietary restraint is associated
with higher urinary cortisol excretion in healthy premenopausal women. The
American Journal of Clinical Nutrition, 73(1), 7-12.
Moos, R. H., & Moos, B. S. (2006). Rates and predictors of relapse after natural and
treated remission from alcohol use disorders. Addiction, 101(2), 212-222.
Mosher, J. F., & Johnsson, D. (2005). Flavored alcoholic beverages: An international
marketing campaign that targets youth. Journal of Public Health Policy, 26(3),
326-342.
Mumenthaler, M. S., Taylor, J. L., O'Hara, R., Fisch, H. U., & Yesavage, J. A. (1999).
Effects of menstrual cycle and female sex steroids on ethanol
pharmacokinetics.Alcoholism, Clinical and Experimental Research, 23(2), 250-
255.

77
Nowland, M. H., Hugunin, K. M. S., & Rogers, K. L. (2011). Effects of short-term fasting
in male sprague-dawley rats. Comparative Medicine, 61(2), 138-144.
Oberlin, B. G., Dzemidzic, M., Tran, S. M., Soeurt, C. M., Albrecht, D. S., Yoder, K. K.,
& Kareken, D. A. (2013). Beer flavor provokes striatal dopamine release in male
drinkers: Mediation by family history of
alcoholism. Neuropsychopharmacology, 38(9), 1617-24.
O'Connor, E.,C., Chapman, K., Butler, P., & Mead, A. N. (2011). The predictive validity
of the rat self-administration model for abuse liability. Neuroscience &
Biobehavioral Reviews, 35(3), 912-938.
Panlilio, L. V., & Goldberg, S. R. (2007). Self-administration of drugs in animals and
humans as a model and an investigative tool. Addiction, 102(12), 1863-1870.
Perry, C. J., Zbukvic, I., Kim, J. H., & Lawrence, A. J. (2014). Role of cues and contexts
on drug-seeking behaviour. British Journal of Pharmacology, 171(20), 4636-
4672.
Petrakis, I. L., Gonzalez, G., Rosenheck, R., & Krystal, J. H. (2002). Comorbidity of
alcoholism and psychiatric disorders: An overview. Alcohol Research and
Health, 26(2), 81-89.
Preston, K. L., & Epstein, D. H. (2011). Stress in the daily lives of cocaine and heroin
users: Relationship to mood, craving, relapse triggers, and cocaine use.
Psychopharmacology, 218(1), 29-37.
Rada, P., Avena, N. M., & Hoebel, B. G. (2005). Daily bingeing on sugar repeatedly
releases dopamine in the accumbens shell. Neuroscience, 134(3), 737-744.

78
Regier, D. A., Farmer, M. E., Rae, D. S., Locke, B. Z., Keith, S. J., Judd, L. L., &
Goodwin, F. K. (1990). Comorbidity of mental disorders with alcohol and other
drug abuse: results from the epidemiologic catchment area (ECA) study. Journal
of the American Medical Association, 264(19), 2511-2518.
Rehm, J., Mathers, C., Popova, S., Thavorncharoensap, M., Teerawattananon, Y., &
Patra, J. (2009). Alcohol and global health 1: Global burden of disease and injury
and economic cost attributable to alcohol use and alcohol-use disorders. The
Lancet, 373(9682), 2223-2233.
Robbins, T. W., & Everitt, B. J. (1996). Neurobehavioural mechanisms of reward and
motivation. Current Opinion in Neurobiology, 6(2), 228-236.
Romanus, G. (2000). Alcopops in sweden--a supply side initiative. Addiction, 95 Suppl
4, S609-S619.
Samson, H. H. (1986). Initiation of ethanol reinforcement using a sucrose-substitution
procedure in food- and water-sated rats. Alcoholism, Clinical and Experimental
Research, 10(4), 436-442.
Samson, H. H., Pfeffer, A. O., & Tolliver, G. A. (1988). Oral ethanol self-administration
in rats: Models of alcohol-seeking behavior. Alcoholism, Clinical and
Experimental Research, 12(5), 591-598.
Schebendach, J., Klein, D. A., Mayer, L. E. S., Attia, E., Devlin, M. J., Foltin, R. W., &
Walsh, B. T. (2017). Assessment of the motivation to use artificial sweetener
among individuals with an eating disorder. Appetite, 109, 131-136.

79
Schulteis, G., Hyytiä, P., Heinrichs, S. C., & Koob, G. F. (1996). Effects of chronic
ethanol exposure on oral self-administration of ethanol or saccharin by wistar
rats. Alcoholism, Clinical and Experimental Research, 20(1), 164-171.
Sclafani, A., & Ackroff, K. (2017). Flavor preferences conditioned by nutritive and non-
nutritive sweeteners in mice. Physiology & Behavior, 173, 188-199.
Shaham, Y., Erb, S., & Stewart, J. (2000). Stress-induced relapse to heroin and cocaine
seeking in rats: A review. Brain Research Reviews, 33(1), 13-33.
Shaham, Y., Shalev, U., Lu, L., de Witt, H., Stewart, J. (2002). The reinstatement model
of drug relapse: history, methodology and major findings. Psychopharmacology,
168 (1), 3-20.
Shalev, U., Yap, J., & Shaham, Y. (2001). Leptin attenuates acute food deprivation-
induced relapse to heroin seeking. The Journal of Neuroscience: The Official
Journal of the Society for Neuroscience, 21(4), 1.
Sinha, R. (2008). Chronic stress, drug use, and vulnerability to addiction. Annals of the
New York Academy of Sciences, 1141(1), 105-130.
Sinha, R., Fox, H. C., Hong, K. A., Hansen, J., Tuit, K., & Kreek, M. J. (2011). Effects of
adrenal sensitivity, stress- and cue-induced craving, and anxiety on subsequent
alcohol relapse and treatment outcomes. Archives of General Psychiatry, 68(9),
942-952.
Smith, S. G., & Davis, W. M. (1974). Intravenous alcohol self-administration in the rat.
Pharmacological Research Communications, 6(4), 379-402.
Stanhope, K. L., Medici, V., Bremer, A. A., Lee, V., Lam, H. D., Nunez, M. V., . . . Havel,
P. J. (2015). A dose-response study of consuming high-fructose corn syrup-

80
sweetened beverages on lipid/lipoprotein risk factors for cardiovascular disease
in young adults. American Journal of Clinical Nutrition, 101(6), 1144-1154.
Steinberg, E. E., Boivin, J. R., Saunders, B. T., Witten, I. B., Deisseroth, K., & Janak, P.
H. (2014). Positive reinforcement mediated by midbrain dopamine neurons
requires D1 and D2 receptor activation in the nucleus accumbens. PLoS
One, 9(4).
Stemmelin, J., Lukovic, L., Salome, N., & Griebel, G. (2005). Evidence that the lateral
septum is involved in the antidepressant-like effects of the vasopressin V1b
receptor antagonist, SSR149415. Neuropsychopharmacology, 30(1), 35-42.
Substance Abuse and Mental Health Services Administration. (2015). National Survey
on Drug Use and Health (NSDUH). Table 2.20A: Alcohol Use, Binge, Alcohol
Use, and Heavy Alcohol Use in Past Month, by Detailed Age Category: Numbers
in Thousands. Rockville, MD.
Sutker, P. B., Tabakoff, B., Goist, K. C., & Randall, C. L. (1983). Acute alcohol
intoxication, mood states and alcohol metabolism in women and men.
Pharmacology, Biochemistry, and Behavior, 18 Suppl 1, 349-354.
Tatzer, E., Schubert, M. T., Timischl, W., & Simbruner, G. (1985). Discrimination of
taste and preference for sweet in premature babies. Early Human
Development, 12(1), 23-30.
Tanoue, A., Ito, S., Honda, K., Oshikawa, S., Kitagawa, Y., Koshimizu, T., . . .
Tsujimoto, G. (2004). The vasopressin V1b receptor critically regulates
hypothalamic-pituitary-adrenal axis activity under both stress and resting
conditions. The Journal of Clinical Investigation, 113(2), 302-309.

81
Taylor, J. L., Dolhert, N., Friedman, L., Mumenthaler, M., & Yesavage, J. A. (1996).
Alcohol elimination and simulator performance of male and female aviators: A
preliminary report. Aviation, Space and Environmental Medicine, 67(5), 407-413.
Thai, P., Tan, E., Tan, W., Tey, T., Kaur, H., & Say, Y. (2011). Sweetness intensity
perception and pleasantness ratings of sucrose, aspartame solutions and cola
among multi-ethnic malaysian subjects. Food Quality and Preference, 22(3), 281-
289.
Thorndike, E. L. (1898). Animal intelligence: An experimental study of the associative
processes in animals. Psychological Monographs: General and Applied, 2(4), i-
109.
Tobin, S., Newman, A. H., Quinn, T., & Shalev, U. (2009). A role for dopamine D1-like
receptors in acute food deprivation-induced reinstatement of heroin seeking in
rats. The International Journal of Neuropsychopharmacology, 12(2), 217-26.
Ventura, A. K., & Mennella, J. A. (2011). Innate and learned preferences for sweet taste
during childhood. Current Opinion in Clinical Nutrition and Metabolic Care, 14(4),
379-384.
Waller, M. B., McBride, W. J., Gatto, G. J., Lumeng, L., & Li, T. K. (1984). Intragastric
self-infusion of ethanol by ethanol-preferring and -nonpreferring lines of rats.
Science, 225(4657), 78-80.
Wheeler, R. A., Aragona, B. J., Fuhrmann, K. A., Jones, J. L., Day, J. J., Cacciapaglia,
F., . . . Carelli, R. M. (2011). Cocaine cues drive opposing context-dependent
shifts in reward processing and emotional state. Biological Psychiatry, 69(11),
1067-1074.

82
White, N. M., & Carr, G. D. (1985). The conditioned place preference is affected by two
independent reinforcement processes. Pharmacology, Biochemistry, and
Behavior, 23(1), 37-42.
Wise, R. A., & Rompre, P. P. (1989). Brain dopamine and reward. Annual Review of
Psychology, 40, 191-225.
Willcocks, A. L., & McNally, G. P. (2013). The role of medial prefrontal cortex in
extinction and reinstatement of alcohol-seeking in rats. The European Journal of
Neuroscience, 37(2), 259-268.
Williams, B. A. (1976). The effects of unsignalled delayed reinforcement. Journal of the
Experimental Analysis of Behavior, 26(3), 441-449.
World Health Organization. (2009). Global health risks: mortality and burden of disease
attributable to selected major risks. Geneva, World Health Organization.
World Health Organization. (2014). Global status report on alcohol and health. Geneva,
World Health Organization.
Wyvell, C. L., & Berridge, K. C. (2000). Intra-accumbens amphetamine increases the
conditioned incentive salience of sucrose reward: Enhancement of reward
"wanting" without enhanced "liking" or response reinforcement. Journal of
Neuroscience, 20(21), 8122-8130.
Yasoshima, Y., & Shimura, T. (2015). A mouse model for binge-like sucrose
overconsumption: Contribution of enhanced motivation for sweetener
consumption. Physiology & Behavior, 138, 154-164.

83
Yu, G., Chen, H., & Sharp, B. M. (2014). Amplified reacquisition of nicotine self-
administration in rats by repeated stress during abstinence.
Psychopharmacology, 231(16), 3189-3195.
Zhou, Y., Leri, F., Cummins, E., Hoeschele, M., & Kreek, M. J. (2008). Involvement of
arginine vasopressin and V1b receptor in heroin withdrawal and heroin seeking
precipitated by stress and by heroin. Neuropsychopharmacology, 33(2), 226-236.
Zhou, Y., Bendor, J. T., Yuferov, V., Schlussman, S. D., Ho, A., & Kreek, M. J. (2005).
Amygdalar vasopressin mRNA increases in acute cocaine withdrawal: Evidence
for opioid receptor modulation. Neuroscience, 134(4), 1391-1397.