the effects of cannabidiol on the electrical and
TRANSCRIPT

United Arab Emirates UniversityScholarworks@UAEU
Theses Electronic Theses and Dissertations
10-2014
THE EFFECTS OF CANNABIDIOL ON THEELECTRICAL AND CONTRACTILEPROPERTIES OF CARDIOMYOCYTESRamez Ali Mansour
Follow this and additional works at: https://scholarworks.uaeu.ac.ae/all_theses
Part of the Medicine and Health Sciences Commons
This Thesis is brought to you for free and open access by the Electronic Theses and Dissertations at Scholarworks@UAEU. It has been accepted forinclusion in Theses by an authorized administrator of Scholarworks@UAEU. For more information, please contact [email protected].
Recommended CitationMansour, Ramez Ali, "THE EFFECTS OF CANNABIDIOL ON THE ELECTRICAL AND CONTRACTILE PROPERTIES OFCARDIOMYOCYTES" (2014). Theses. 44.https://scholarworks.uaeu.ac.ae/all_theses/44

United Arab Emirates University
College of Medicine and Health Sciences
Department of Pharmacology and Therapeutics
THE EFFECTS OF CANNABIDIOL ON THE ELECTRICAL AND
CONTRACTILE PROPERTIES OF CARDIOMYOCYTES
Ramez Ali Mansour
This thesis is submitted in partial fulfillment of the requirements for the degree of Master
of Medical Sciences
Under the Supervision of Dr. Murat Oz
October 2014

ii
Declaration of Original Work
I, Ramez Ali Mansour, the undersigned, a graduate student at the United Arab Emirates
University (UAEU) and the author of this thesis entitled “The effects of cannabidiol on
the electrical and contractile properties of cardiomyocytes”, hereby solemnly declare that
this thesis is an original research work that has been done and prepared by me under the
supervision of Dr. Murat Oz, in the College of Medicine and Health Sciences at the
UAEU. This work has not previously formed the basis for the award of any academic
degree, diploma or similar title at this or any other university. The materials borrowed
from other sources and included in my dissertation have been properly cited and
acknowledged.
Student’s Signature___________________ Date___________________

iii
Copyright © 2014 by Ramez Ali Mansour
All Rights Reserved

iv
Approval of the Master Thesis
This Master Thesis is approved by the following Examining Committee Members:
1) Advisor (Committee Chair):
Title:
Department of …
College of …
Signature________________________________ Date___________________________
2) Member:
Title:
Department of …
College of …
Signature________________________________ Date___________________________
3) Member:
Title:
Department of …
College of …
Signature________________________________ Date___________________________
4) Member (External Examiner):
Title:
Department of …
Institution:
Signature________________________________ Date___________________________

v
This Master thesis is accepted by:
Dean of the College of Medicine and Health Science: Professor Dennis Templeton
Signature___________________________________ Date________________
Dean of the College of Graduate Studies: Professor Nagi Wakim
Signature___________________________________ Date________________
Copy ____ of ____

vi
Abstract
In earlier studies, cannabidiol (CBD), a major nonpsychotropic cannabinoid found
in cannabis plant, has been shown to influence cardiovascular functions under various
physiological and pathological conditions. In the present study, the effects of CBD on
contractility and electrical properties of rat ventricular myocytes were investigated. Video
edge detection was used to measure myocyte shortening. Intracellular Ca2+
was measured
in cells loaded with the fluorescent indicator fura-2 AM. CBD (1 µM) caused a
significant decrease in the amplitudes of electrically-evoked myocyte shortening and Ca2+
transients. However, the amplitudes of caffeine-evoked Ca2+
transients and the rate of
recovery of electrically-evoked Ca2+
transients following caffeine application were not
altered. Whole-cell patch-clamp technique was employed to investigate the effect of CBD
on the characteristics of action potentials (APs) and L-type Ca2+
channels. CBD (1 µM)
significantly decreased the duration of APs. Further studies on L-type Ca2+
channels
indicated that CBD inhibits these channels with IC50 of 0.1 µM in a voltage-independent
manner.
Keywords: Cannabidiol, I type calcium channels, ventricular myocyte.

vii
Title and Abstract in Arabic
الكهربية و الانقباضية الخاصة بالخلايا العضلية على الخواص الكانابيديول آثار
القلبية
الملخص
في دراسات سابقة، الكانابيديول، و هو من المؤثرات العقلية الكانابينويدية الرئيسية الموجودة في نبات
ف الظروف الفيسيولوجية و المرضية. الحشيش، وجد أن له تأثيرات على وظائف القلب و الأوعية الدموية تحت مختل
الهدف في هذه الأطروحة هو دراسة اثار الكانابيديول على الانقباضات و الخصائص الكهربية لخلايا الفأر العضلية.
Caو تم قياس انقباض الخلايا العضلية بالاستعانة بطريقة الكشف بالفيديو إيدج. و تم قياس الموجودة داخل الخلايا +2
(.fura-2 AMمؤشر فلوريسنت ) باستخدام
تسبب في انخفاض ملحوظ في سعة انقباض الخلايا (μM 1)الكانابيديول أهم نتائج هذه الدراسة هو أن
Caالعضلية المثارة كهربيا و Ca)العابر +2
2+ transients) غير أن ال .Ca
العابر المثار بالكافيين و معدل +2
Caإنعاش ال الكانابيديول على التالية لعملية إضافة الكافيين لم تتغير. و تم التحقق من تأثير العابر المثار كهربيا +2
L- type Ca( و قنوات ال action potentialsخصائص جهد الفعل )بالاستعانة بتقتية الالتقاط الرقعي لكامل +2
أدى لإنخفاض (μM 1)يول الكانابيد(. بالإضافة إلى ذلك، دلت النتائج أن Whole-cell patch-clampالخلية )
L- type Caملحوظ في المدة الزمنية لجهد الفعل. مزيد من الدراسات على قنوات ال الكانابيديول يمنع بينت أن +2
(.voltage-independent mannerبطريقة الجهد المستقلة ) μM 0.1من IC50هذه القنوات ب

viii
Acknowledgments
I would like to express my deep gratitude to my master thesis advisor, Professor
Murat Oz who has supported me throughout my thesis with his patience and knowledge
whilst allowing me the room to work in my own way. I appreciate his encouragement and
efforts without which this thesis would not have been completed. One simply could not
wish for a better or friendlier supervisor. I am also grateful to Professor Chris Howarth,
Professor Oleg Krishtal and Dr. Rajesh Mohanraj for spending time in reading and
providing useful suggestions about the thesis.
My gratitude also goes to my colleagues to thank them for their assistance to me:
Mr. Anwar, Dr. Nour Alain, Dr. Lina and Khawla. Additionally, I want to acknowledge
some friends who inspirited my efforts to overcome difficulties, so a special thanks to,
Mohammed and Abrar.
I am grateful to my mother, my father and my uncle for all of the sacrifices that
they have made on my behalf. I would not have got to this point without you. Special
thanks go to my brother Ahmed and my sister Yousra for their continuous
encouragement.

ix
Dedication
To my parents, my uncle, my brother Ahmed and my
sister Yousra

x
Table of Contents
Title .................................................................................................................................................. i
Declaration of Original Work .......................................................................................................... ii
Copyright ........................................................................................................................................ iii
Approval of the Master Thesis ....................................................................................................... iv
Abstract .......................................................................................................................................... vi
Title and Abstract in Arabic .......................................................................................................... vii
Acknowledgments ........................................................................................................................ viii
Dedication ...................................................................................................................................... ix
Table of Contents ............................................................................................................................ x
List of Tables ................................................................................................................................. xii
List of Figures .............................................................................................................................. xiii
Glossary ........................................................................................................................................ xiv
Chapter 1: Introduction ................................................................................................................... 1
1.1 Aims and Objectives: ...................................................................................................... 1
1.2 History ............................................................................................................................. 1
1.3 Cannabinoids ................................................................................................................... 6
1.3.1 Phytocannabinoids ................................................................................................... 7
1.3.2 Endocannabinoids .................................................................................................... 8
1.3.3 Synthetic Cannabinoids ........................................................................................... 9
1.4 Cannabidiol ................................................................................................................... 13
1.4.1 Mechanisms of Action of Cannabidiol .................................................................. 13
1.4.2 Clinical Applications ............................................................................................. 20
1.5 Contraction of Cardiac Muscle ...................................................................................... 28
1.5.1 Excitation–Contraction Coupling .......................................................................... 31
1.5.2 Sarcoplasmic Reticulum and Ryanodine Receptors .............................................. 40
1.5.3 The Contractile Machinery .................................................................................... 41
1.5.4 Calcium Sensitivity of Myofilaments .................................................................... 43
Chapter 2: Methods ....................................................................................................................... 44
2.1 Ventricular Myocyte Isolation ....................................................................................... 44
2.2 Measurement of Ventricular Myocyte Shortening ........................................................ 46
2.3 Measurement of Intracellular Ca2+
Concentration ......................................................... 46
2.4 Measurement of Sarcoplasmic Reticulum Ca2+
Content ............................................... 47
2.5 Assessment of Myofilament Sensitivity to Ca2+
............................................................ 48
2.6 Whole-Cell Recordings ................................................................................................. 49
Chapter 3: Results ......................................................................................................................... 53

xi
3.1 Effects of Cannabidiol on Ventricular Myocyte Shortening ......................................... 53
3.2 Effects of Cannabidiol on Intracellular Ca2+
................................................................. 57
3.3 Effect of Cannabidiol on Sarcoplasmic Reticulum Ca2+
Transport ............................... 59
3.4 Effect of Cannabidiol on Myofilament Sensitivity to Ca2+
........................................... 61
3.5 Effects of Cannabidiol on the Action Potentials of Ventricular Myocytes ................... 63
3.6 Effect of Cannabidiol on L-type Ca2+
Currents ............................................................. 65
Chapter 4: Discussion .................................................................................................................... 70
Chapter 5: Limitation and Future Directions ................................................................................. 77
Bibliography .................................................................................................................................. 78

xii
List of Tables
Table 1. Class of Cannabinoids ..................................................................................................... 11
Table 2. Classification of Calcium Channels ................................................................................ 36

xiii
List of Figures
Figure 1. Cannabis Sativa leaves. .................................................................................................... 2
Figure 2. Major Phytocannabinoids ................................................................................................ 2
Figure 3. Major Phytocannabinoids Synthetic Pathways. ............................................................... 5
Figure 4. Heart's electrical system ................................................................................................. 28
Figure 5. Cardiac Myocytes. ......................................................................................................... 29
Figure 6. Ventricular Action Potential Membrane ........................................................................ 30
Figure 7. Cardiac Excitation Contraction Coupling. ..................................................................... 34
Figure 8. Calcium Channel Structure ............................................................................................ 38
Figure 9. Sarcomere Structure. ...................................................................................................... 42
Figure 10. Langendorff Heart ........................................................................................................ 45
Figure 11. Video Edge Detection and Ca2+ Imaging ................................................................... 48
Figure 12. Patch Clamp Setup ....................................................................................................... 50
Figure 13.Microelectrode Puller. ................................................................................................... 51
Figure 14. Glass Electrode Forming Giga Seal with Ventricular Myocyte ................................... 52
Figure 15. Ventricular Myocyte .................................................................................................... 52
Figure 16. Effects of Cannabidiol on Ventricular Myocyte Shortening... ..................................... 55
Figure 17. Concentration-Inhibition Relationship of Cannabidiol Effect.. ................................... 56
Figure 18. Effects of Cannabidiol on Amplitude and Time-Course of Intracellular Ca2+
............. 58
Figure 19. Effect of Cannabidiol on Sarcoplasmic Reticulum Ca2+
Transport ............................. 60
Figure 20. Effect of Cannabidiol on Myofilament Sensitivity to Ca2+
. ......................................... 62
Figure 21. Effect of Cannabidiol on the Excitability of Ventricular Myocytes. ........................... 64
Figure 22. Effect of Cannabidiol on Ca2+
Currents Mediated by L-type Ca2+
Channels ............... 66
Figure 23. The effect of Cannabidiol on Voltage-Current Characteristics ..................................... 66
Figure 24. Steady State Activation and Inactivation of L-type Ca2+
Currents .............................. 68
Figure 25. Effect of Cannabidiol on the Kinetics of L-type Ca2+
Currents ................................... 69

xiv
Glossary
THC Tetrahydrocannabinol
CBN Cannabinol
CBD Cannabidiol
THCA Tetrahydrocannabinolic Acid
CBGA Cannabigerolic Acid
CBG Cannabigerol
CBDA Cannabidiolic Acid
CBCA Cannabichromenic Acid
CBC Cannabichromene
AEA Anandamide
2-AG 2-Arachidonylglycerol
FAAH Fatty Acid Amidehydrolase
MAGL Monoacylglycerol
AJA Ajalemic Acid
TRPV1 Transient Vanniloid Aeceptors 1
PPARγ Peroxisome Proliferator Activated Receptor
VR1 Vannilloid Receptors
PLC Phospholipase C
PKC Phosphokinase C
NADA N-arachidonoyl-Dopamine
NCX Soduim- Calcium Exchanger
APD Action Potential Duration
RyRs Ryanodine Receptors
PKA Protein Kinase A
ADP Adenosine Diphosphate
ATP Adenosine Triphosphate
NT Normal Tyrode
RCL Resting Cell Length
SR Sarcoplasmic Reticulum
TTX Tetrodotoxin
AMP Amplitudes
RU Ratio Unit
SSA Steady-State Activation
SSL Steady-State Inactivation
AP Action Potential
APD Action Potential Duration

Chapter 1: Introduction
1.1 Aims and Objectives:
The aims of this research are to examine the therapeutic potential and mechanism
by which the phytocannabinoid Cannabidiol CBD affects cardiac contractility. This will
be achieved by investigating the effects of CBD on action potential duration, ventricular
shortening, myofilament sensitivity, and L-type calcium current.
1.2 History
Marijuana is a crude preparation of flowering tops, leaves, seeds and stems from
female Indian hemp cannabis sativa plants (Figure 1). It has been used for centuries for
both recreational and therapeutic purposes. Currently cannabis is being investigated for
the treatment of various pathological conditions such as pain, nausea, vomiting and
cancer. It is one of the most extensively studied plants. There were more than 12,000
PubMed articles published in January 2014 using the key word cannabis in relation to the
various biological and pharmacological effects of this drug.

2
Figure 1. Cannabis Sativa Leaves.ScienceDaily, n.d. Web. 30 Apr. 2014
Figure 2. Major Phytocannabinoids

3
The medicinal use of marijuana has a long history of therapeutic use, going back
several thousands of years. It was often used for the same medical conditions it is used to
treat today such as muscle spasms, pain, nausea, vomiting, epilepsy, and glaucoma.
However, use of marijuana is also associated with distorted perception (sights, sounds,
time and touch), panic attacks, trouble with thinking, difficulties in learning and problem
solving, and loss of motor coordination. Due to these adverse effects, the use of
marijuana was banned in the U.S.A and throughout most of the world almost eighty years
ago.
During the second half of the 20th
century, researchers discovered that the
biological actions of marijuana are mediated mainly by a group of chemicals collectively
called cannabinoids. Although cannabis plant extracts contain more than 460 compounds,
based on pharmacological properties and radioligand binding studies, currently about
60 of the compounds found in this plant, are collectively named phytocannabinoids.
Among these phytocannabinoids, Δ9-Tetrahydrocannabinol (THC) is a major chemical
that mediates the psychoactive actions of the cannabis plant (Figure 2). In addition to
THC, other cannabinoids such as Cannabigerol, Cannabichromene, Cannabinol (CBN)
and Cannabidiol (CBD), constitute major non-psychoactive components of the cannabis
plant. These compounds are highly lipophilic and bear significant similarities in their
chemical structure. For example, CBD which is the focus of this study, is a structural
isomer of THC i.e. it has the same chemical composition but the atoms are arranged
differently.

4
While the majority of the adverse psychological effects of marijuana are known to
be mediated by THC, the contribution of other cannabinoids such as CBD and CBN to
the overall influence of marijuana remains unknown. In recent years, various
combinations of these psychoactive and non-psychoactive cannabinoids have been used
in cannabis based treatments. Different extracts of cannabis have been used
therapeutically to relieve, asthma, pain, whooping cough, epilepsy and inflammation.
However, due to the psychoactive effects of cannabis such as anxiety, cognitive
impairment and hallucinations, medicinal use of these compounds was discontinued
(Robson, 2001).
Nevertheless, its therapeutic potential to solve a myriad of medical problems has
attracted considerable research interest. Thus, in recent years research has focused on
the therapeutic potential of the plant to develop new chemicals that retain the therapeutic
benefits without causing psychotropic effects.

5
Figure 3. Illustrating Major Phytocannabinoids Synthetic Pathways.

6
1.3 Cannabinoids
Despite its use for centuries, the active ingredients and chemical constituents
Of the cannabis plant remained largely unknown until the beginning of the 1960s when
the main psychoactive constituent, THC was extracted and identified (Mechoulam &
Gaoni, 1965). In the 1980s, other compounds with chemical structures similar to that of
THC were synthesized by various drug companies in an attempt to discover new drugs to
alleviate pain. Some of these newly synthesized compounds such as CP55,940,
CP55,244, HU-210, JWH-018 displaced radioactively labeled THC in radioligand
binding experiments and lead to the discovery of the first cannabinoid receptor
(CB1) in the brain (Herkenham et al., 1990). In the 1990’s CB1 and CB2 receptors
were cloned, expressed functionally and identified pharmacologically (Pertwee, 1997).
Further studies in this field, indicated that, similar to opioids, there are
endogenous ligands that can bind and activate cannabinoid receptors. These findings
eventually lead to the discovery of “endocannabinoids” i.e. endogenously produced
cannabinoids. Endocannabinoids mainly N-arachidonoylethanolamine (Anandamide;
AEA) and 2-arachidonylglycerol (2-AG) have considerable agonistic effects on the
cannabinoid receptors (Mechoulam et al., 1995), and mimic most of the pharmacological
actions of THC.

7
1.3.1 Phytocannabinoids
As mentioned earlier the most abundant constituents of the cannabis plant are
THC and CBD. Within the plant, phytocannabinoids are synthesized from fatty acid
precursors via a series of transferase and synthase enzymes (Figure 3). The two major
phytocannabinoids, THC and CBD, are derived from a common synthetic precursor,
cannabigerol. From a pharmaco-chemical perspective, whilst THC and CBD have pentyl
side chains, major homologues are Δ9-tetrahydrocannabivarinand cannabidivarin,
respectively, with propyl side chains, derived from cannabigerovarin. Importantly,
despite only small differences in chemical structure, these compounds appear to exhibit
markedly different pharmacological properties (Hill, Williams, Whalley, & Stephens,
2012).
THC has a wide spectrum of pharmacological effects ranging from anti-
inflammatory and analgesic actions to cancer treatment, mainly through the activation of
the cannabinoid receptors (Mechoulam, Parker, & Gallily, 2002). However, CBD has no
agonistic activity on cannabinoid receptors, but it is capable of producing various
pharmacological effects by acting on diverse groups of target proteins ranging from
enzymes and receptors to neurotransmitter transporters and ion channels (Hill et al.,
2012).

8
1.3.2 Endocannabinoids
Endogenous ligands for the cannabinoid receptor are synthesized from
polyunsaturated fatty acids and fatty acid derivatives from glycerol. They mimic the
actions of phytocannabinoids by binding to cannabinoid receptors (Di Marzo, 2008). In
addition to AEA and 2-AG, other endocannabinoids with similar fatty acid-based
chemical structures have also been identified in the last decade (Pertwee et al., 2010).
Some of these endocannabinoids include 2-arachidonylglycerol ether (Noladin ether);
(Hanus et al., 2001), NADA, and virodhamine (Pertwee et al., 2010; Porter et al., 2002).
Endocannabinoids, unlike neurotransmitters, are neither synthesized nor stored in
the neurons. Therefore they are not released by a synaptic process, but produced in
response to cellular activity (Di Marzo et al., 1994). It has been shown that biological
processes such as membrane depolarization and activation of metabotropic receptors
cause increases in intracellular Ca2+
levels and activate enzymes involved in the synthesis
of endocannabinoids (Di Marzo, 2008). Following their synthesis, AEA and 2-AG are
degraded by enzymatic hydrolysis by fatty acid amide hydrolase (FAAH) and
monoacylglycerol lipase (MAGL), respectively (Cravatt & Lichtman, 2002).
Endocannabinoids can also be transported out of the cell by diffusion through cell
membranes or by a mechanism that involves an unidentified transporting protein located
in the plasma membrane (Cravatt & Lichtman, 2002; Glaser, Kaczocha, and Deutsch,
2005).

9
1.3.3 Synthetic Cannabinoids
Following the discovery of cannabinoid receptors there have been attempts to
produce compounds with therapeutic effects such as anti-inflammatory/analgesic and
anti-emetic effects without inducing the psychotropic effects. As a result various
chemicals such as CP55,940, CP55,244,HU-210, JWH-018, and JWH-073 have been
synthesized (Huffman et al., 1998). These compounds are known collectively as synthetic
cannabinoids (Ashton, 2012). Currently, little is known about the biological activities of
these compounds with their diverse chemical structures. However, recent reports indicate
that various combinations of synthetic cannabinoids are employed for recreational
purposes throughout the world in order to avoid legal regulations related to
phytocannabinoids (Burstein and Zurier, 2009). However, the synthetic cannabinoid
Nabilone has recently been approved by the FDA (U.S Food and Drug Administration)
and recommended for the treatment of chemotherapy-induced nausea and vomiting that is
unresponsive to conventional antiemetics (Ware, Daeninck, and Maida, 2008).In addition
nabilone is used for the treatment of anorexia and weight loss in AIDS patients (Burstein
and Zurier, 2009; Pertwee et al., 2010). Nabilone has also been reported to be an effective
pain killer for patients with fibromyalgia (Skrabek, Galimova, Ethans, & Perry, 2008).
Currently the following cannabinoid based medications have been approved by
the FDA and are commercially available (Schicho & Storr, 2011): 1– Sativex® (GW
Pharmaceuticals), for pain and spasticity in patients with multiple sclerosis. Currently,
four different formulations of Sativex® are under investigation, including the high
THC extract (Tetranabinex®), THC:CBD (narrow ratio), THC:CBD (broad ratio)
and the high CBD extract (Nabidiolex®) (Scuderi et al. 2009). currently, three

10
Sativex® delivery systems exist: the oromucosal spray, sublingual tablets and inhaled
dosage forms. In 2005, the oromucosal spray administration of Sativex® was approved
for the treatment of multiple sclerosis symptoms (Perras 2005). To date, Sativex
preparations have been licensed in Canada, the UK and Spain. 2– Marinol® (Solvay
Pharmaceuticals, Belgium), oral capsules containing dronabinol (a synthetic THC), are
recommended as an appetite stimulant and antiemetic. 3– Cesamet® (Valeant
Pharmaceuticals International), is an oral capsule containing nabilone (a synthetic THC
analog), for patients with chemotherapy-induced nausea and vomiting. This drug is
currently licensed in Canada, the USA and the UK.
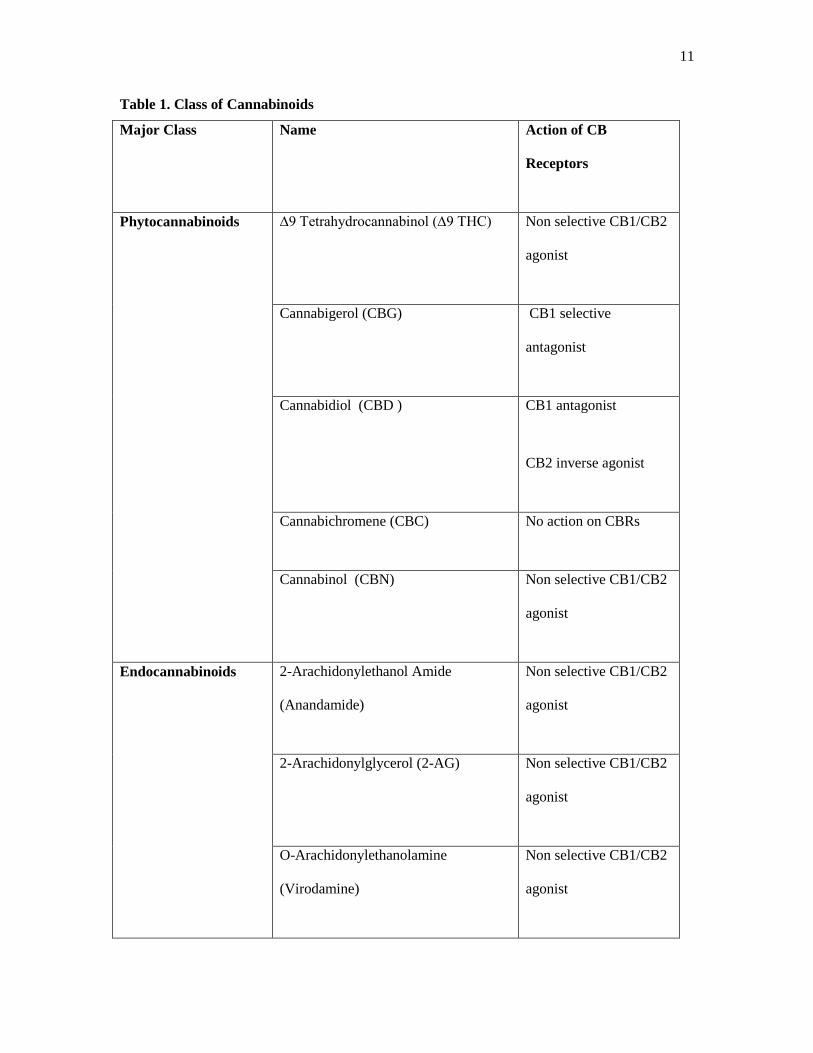
11
Table 1. Class of Cannabinoids
Major Class Name Action of CB
Receptors
Phytocannabinoids ∆9 Tetrahydrocannabinol (∆9 THC) Non selective CB1/CB2
agonist
Cannabigerol (CBG) CB1 selective
antagonist
Cannabidiol (CBD ) CB1 antagonist
CB2 inverse agonist
Cannabichromene (CBC) No action on CBRs
Cannabinol (CBN) Non selective CB1/CB2
agonist
Endocannabinoids 2-Arachidonylethanol Amide
(Anandamide)
Non selective CB1/CB2
agonist
2-Arachidonylglycerol (2-AG) Non selective CB1/CB2
agonist
O-Arachidonylethanolamine
(Virodamine)
Non selective CB1/CB2
agonist

12
Synthetic
Cannabinoids
CP55,940 Non selective CB1/CB2
agonist
WIN55,212 Non selective CB1/CB2
agonist
Sch.336 CB2 selective
Ajulemic acid (AJA) Non selective CB1/CB2
agonist

13
1.4 Cannabidiol
CBD is the main non-psychoactive component of cannabis and constitutes up to
40% of cannabis extracts (Grlic, 1962). It was first isolated in 1940 and its chemical
structure was identified in 1963 (Mechoulam and Hanus, 2002). In recent years, CBD has
been considered to be a promising therapeutic agent for clinical use, since, in addition to
its non-psychotropic properties, it exhibits as well as low toxicity in humans and has a
high therapeutic index and a low teratogenic potential (Rosenkrantz, Fleischman, and
Grant, 1981; Campos, Moreira, Gomes, Del Bel, and Guimaraes, 2012; Scuderi et al.,
2009). An orally-administered liquid containing CBD has received orphan drug status in
the US, for use as a treatment for Dravet Syndrome, a rare-form of epilepsy that is
resistant to treatment with current anti-epileptic drugs (Gloss, Nolan, and Staba, 2014).
Cannabidiol is insoluble in water but soluble in organic solvents, such as DMSO
and alcohol. It is rapidly distributed when administered intravenously, easily passes the
blood-brain barrier and has a prolonged elimination from the body (Samara, Bialer and
Harvey, 1990).
1.4.1 Mechanisms of Action of Cannabidiol
In general, CBD has been shown to act on a wide range of molecular and cellular
target sites. Some of these molecular targets are various types of membrane receptors
such as GPR55 (Ryberg et al., 2007), opioid (Kathmann, Flau, Redmer, Trankle and
Schlicker, 2006) and 5HT1A receptors as well as Transient Vanniloid receptors 1
(TRPV1) (Bisogno et al., 2001), and Peroxisome Proliferator Activated Receptor
(PPARγ) (Bishop-Bailey, 2000). Significantly, CBD, at low µM concentrations acts as an
antagonist for CB1 and CB2 receptors (Pertwee, Ross, Craib and Thomas, 2002). Other

14
proteins that CBD interacts with include voltage-gated and ligand-gated ion channels, and
various types of enzymes (for review see Pertwee, 2008; Izzo et al., 2009; Campos et al.,
2012;Garcia-Arencibia et al., 2007). These cellular and molecular targets are discussed in
detail in the following sections.
a. Actions of cannabidiol on the cannabinoid system:
In earlier clinical studies, CBD, with no detectable activity by itself, was reported
to attenuate some of the effects of THC such as anxiety and tachycardia (Karniol,
Shirakawa, Kasinski, Pfeferman and Carlini, 1974). Similarly, it was found that the
pharmacological actions of cannabinoid receptor agonists can be antagonized by low (nM
range) concentrations of CBD in a mouse’s vas deferens (Pertwee et al., 2002). These
effects of CBD are unlikely to involve direct interaction at cannabinoid receptor CB1 and
CB2 binding sites except at high concentrations of CBD, since CBD appears to have a
relatively low affinity for cannabinoid receptors. Ki values for CBD-induced
displacement of a radiolabelled ligand from cannabinoid CB1 and CB2 receptor binding
sites have been reported to be 4.35 and 2.86 µM, respectively in one study (Showalter,
Compton, Martin and Abood, 1996) and >10 µM in other experiments (Devane, Dysarz,
Johnson, Melvin and Howlett, 1988; Bisogno et al., 2001). Further studies indicated that
CBD behaves as a high-potency antagonist for cannabinoid receptor agonists in a
mouse’s brain tissue and in membranes from CHO cells transfected with cannabinoid
receptors (Thomas et al., 2007). It was also reported that the function of a novel receptor
GPR55 is inhibited by low concentrations of CBD (Ryberg et al., 2007).

15
The interaction of CBD with the cannabinoid system is not limited to cannabinoid
receptors. Tissue levels of endogenously produced cannabinoids are also affected by
CBD. For example, it has been shown that CBD inhibits the uptake and the enzymatic
hydrolysis of AEA (Bisogno et al., 2001). Furthermore, CBD has been shown to cause
activation of cannabinoid receptors indirectly by enhancing tissue concentrations of
endocannabinoids (Fowler, 2004). In functional experiments it has been suggested that
some of the effects of CBD sensitive to CB receptor antagonists are mediated by an
entourage effect of CBD on tissue levels of endocannabinoids (Fowler, 2004).
b. Effects of cannabidiol on other membrane receptors:
CBD has been shown to stimulate Vanilloid Receptors (VR1) with an efficacy
comparable to that of capsaicin, the natural agonist of this receptor (Bisogno et al.,
2001). The stimulation of the VR1 receptor by capsaicin and some of its analogues leads
to fast desensitization, with subsequent analgesic and anti-inflammatory effects. It has
been suggested that, similar to capsaicin, CBD also causes desensitization of VR1 and
enhances the effect of capsaicin on VR1, suggesting that CBD exerts its anti-
inflammatory actions, in part, by desensitization of sensory nociceptors (Bisogno et al.,
2001).CBD has been reported to act as a weak agonist (EC50 = 3.5 µM) on human
TRPV1 receptors inHEK293 cells expressing these receptors (De Petrocellis et al., 2011).
CBD has also been demonstrated to act as an agonist on TRPV2 (Qin et al., 2008) and
TRPA1 receptors, while it acts as an antagonist on the TRPM8 receptor (De Petrocellis et
al., 2008).
It was also shown that at a µM concentration range, CBD displaces the 8-OH-
DPAT, 5-HT1A receptor agonist, from cloned human 5-HT1A receptors expressed in

16
cultured cells obtained from a Chinese hamster ovary (E. B. Russo, Burnett, Hall and
Parker, 2005). In subsequent studies, results of several in vivo experiments have
supported the involvement of 5-HT1A receptors in the effect of CBD (Campos et al.,
2012). At high µM concentrations (10-100 µM), CBD has also been shown to interact
with other G-protein coupled receptors such as 5HT2 and opioid receptors in an allosteric
manner (E. B. Russo et al., 2005; Kathmann, et al., 2006).
c. Effect of cannabidiol on second messengers:
Calcium is considered one of the most important second messengers involved in
the regulation of various cell functions, such as muscle contraction and the release of
neurotransmitters. It has been shown that CBD treatment increases intracellular
concentrations of Ca2+
in cultured hippocampus neurons (Drysdale, Ryan, Pertwee and
Platt, 2006; Ryan, Drysdale, Lafourcade, Pertwee and Platt, 2009). In another study, the
anti-epileptic activity of CBD has been suggested to be due to its bidirectional regulatory
role on intracellular Ca2+
levels (Ryan et al., 2009). In another investigation, it was
proposed that CBD increases intracellular Ca2+
levels under normal physiological
conditions, but decreases Ca2+
levels under highly excitable conditions (Mato, Victoria
Sanchez-Gomez and Matute, 2010; Rao and Kaminski, 2006). This action of CBD was
believed to be due to the regulation of Ca2+
transport in mitochondria which acts as a sink
for intracellular Ca2+
(Ryan et al., 2009).
In addition to its influence on intracellular stores, CBD also acts on active Ca2+
transport proteins such as Ca2+
-ATPase in myocytes (Gilbert, Pertwee and Wyllie, 1977)
and neurons (Gilbert et al., 1977; Ryan et al., 2009). The highly lipophilic nature of CBD
grants it easy access to intracellular targets such as cellular organelles (endoplasmic

17
reticulum and mitochondria) and intracellularly located enzymes that contribute to CBD-
induced alterations in intracellular Ca2+
levels (Collins and Haavik, 1979).In this context,
it is important to note that, the functions of various Ca2+
-activated enzymes such as
phospholipase C (PLC) (Howlett, Scott and Wilken, 1989), phospholipaseA2,
lipoxygenase (Takeda, Usami, Yamamoto and Watanabe, 2009) and Phosphokinase C
(PKC) (Hillard and Auchampach, 1994; White and Tansik, 1980) are also modulated by
low µM concentrations of CBD.
In addition to Ca2+
, CBD acts on the uptake process of several neurotransmitters
(for a review see Pertwee 2008). CBD decreases the uptake of [3H] adenosine in both
murine microglia and macrophages, and binding studies show that CBD binds to the
nucleoside transporter (Carrier, Auchampach and Hillard, 2006). The enhancement of
adenosine signaling through inhibition of its uptake was suggested as providing non-
cannabinoid receptor mechanism by which CBD can decrease inflammation (Carrier et
al., 2006).
d. Cannabidiol effects on voltage-gated ion channels:
Earlier studies of motor neurons showed that CBD depresses the amplitude of
action potential suggesting that it inhibits the function of neuronal voltage-gated Na+
channels (Turkanis and Karler, 1986). However, there has been no further study
investigating the effect of CBD on Na+ channels. Another study indicated that CBD
increased the L-type Ca2+
current recorded in the hippocampal neurons (Drysdale et al.,
2006). Similarly to these Na+ channels, there have been no further studies investigating
the actions of CBD on L-type Ca2+
channels. On the other hand, the effect of CBD on T-
type voltage-gated Ca2+
channels has been investigated in recent years. It has been shown

18
that CBD inhibits different subtypes of CaV3 family channels (Ross, Napier and Connor,
2008).
e. Cannabidiol effects on ligand-gated ion channels:
In HEK-293 cells expressing glycine receptors, CBD showed a positive allosteric
modulatory effect in a low µM concentration range (Ahrens et al., 2009). The EC50
values for the potentiating effects of CBD were 12.3 µM for α1 and 18.1 µM for α1β1
subunit combinations. Direct activation of glycine receptors was also observed at CBD
concentrations above 100 µM. Similarly, glycine-mediated currents recorded in dorsal
horn neurons of rat spinal cord slices were potentiated by CBD applications (Xiong et al.,
2012). Further investigations indicated that mutations of the α1 subunit TM2 serine
residue to isoleucine abolished the co-activation and the direct activation, of the glycine
receptor by CBD (Foadi et al., 2010). In in vivo experiments, it has been shown that
systemic and intrathecal administration of CBD significantly suppressed chronic
inflammatory and neuropathic pain without causing apparent analgesic tolerance in
rodents (Xiong et al., 2012). The analgesic potency of CBD and 11, similarly structured
cannabinoids, is positively correlated with potentiation of the α3-subunitglycine receptor.
In contrast, analgesia induced by these cannabinoids is neither correlated with their
binding affinity for CB1 and CB2 receptors nor with their psychoactive side effects.
Furthermore, NMR analysis revealed a direct interaction between CBD and S296 in the
third transmembrane domain of purified α3 GlyR. More importantly, the CBD-induced
analgesic effect was absent in mice lacking the α3 subunit from the glycine receptor
suggesting that the α3 subunit mediates glycinergic CBD-induced suppression of chronic
pain (Xiong et al., 2012).

19
Another ligand-gated ion channel reported to be modulated by CBD is the α7-
nicotinic cholinergic receptor (Mahgoub et al., 2013). It was shown that the function of
human α7-nicotinic receptors expressed in xenopus oocytes was inhibited by CBD with
an IC50 value of 11.3 µM in a non-competitive manner. Significantly, the 5-HT3
receptor, with structural similarities to the α7-nicotinic receptor was also shown to be
inhibited in xenopus oocytes and HEK-293 cells in an allosteric manner (K. H. Yang et
al., 2010).

20
1.4.2 Clinical Applications
a. Anti-inflammatory actions
As the most abundant non-psychoactive cannabinoid in the plant, the anti-
inflammatory actions of CBD and its analogs have been studied extensively in recent
years (Burstein and Zurier, 2009; Booz, 2011). CBD reduces joint inflammation in
collagen-induced arthritis in mice (Sumariwalla et al., 2004) and carrageenan paw edema
in rats (Costa et al., 2004). In addition, oral administration of CBD (2.5–20 mg/kg)
reduces neuropathic and inflammatory pain in rats. This effect is reversed by vanilloid
but not by CB receptor antagonists (Costa, Trovato, Comelli, Giagnoni and Colleoni,
2007). CBD also reduces intestinal inflammation in mice (Capasso et al., 2008). In
addition to its ability to suppress production of the inflammatory cytokine TNFα, CBD
appears to exert anti-inflammatory pressure by suppressing the fatty acid amidohydrolase
(FAAH), thereby increasing concentrations of the anti-inflammatory endocannabinoid
anandamide. Further, insight into mechanisms whereby CBD exerts therapeutic effects
was provided by experiments which indicated that CBD attenuates inflammation induced
by high glucose levels in diabetic mice (Rajesh et al., 2007). Specifically, CBD treatment
reduces mitochondrial superoxide, inducible nitric oxide synthase, nuclear factor kappa β
activation, and transendothelial migration of monocytes (Burstein and Zurier, 2009;
Booz, 2011).Another potential therapeutic use of CBD may lie in its ability to counter
some undesirable effects of THC (sedation, psychotropic effects, tachycardia), thus
suggesting that if given together with THC, it may allow higher doses of THC (E. Russo
& Guy, 2006). THC and CBD have been administered as an oral mucosal spray to 58
patients with rheumatoid arthritis (Blake, Robson, Ho, Jubb and McCabe, 2006). Treated

21
patients had a significant reduction in pain and an improvement in sleep compared to
patients given a placebo.
b. Neuroprotection
In earlier studies, CBD has been shown to normalize glutamate homeostasis
(Hampson et al., 2000; El-Remessy et al., 2003), reduce oxidative stress (Hampson,
Grimaldi, Axelrod and Wink, 1998; Marsicano, Moosmann, Hermann, Lutz, and Behl,
2002), attenuate glial activation and the occurrence of local inflammatory events (Ruiz-
Valdepenas et al., 2011; Martin-Moreno et al., 2011). It appears that there may be two
key mechanisms underlying the neuroprotective effects of CBD. The first is the
capability of CBD to restore the normal balance between oxidative events and antioxidant
endogenous mechanisms (Fernandez-Ruiz, 2012) that are frequently disrupted in
neurodegenerative disorders, thereby enhancing neuronal survival. The second is CBD as
a neuroprotective compound, and its anti-inflammatory activity. The anti-inflammatory
effects of CBD are related to the control of microglial cell migration and the toxicity
exerted by these cells, i.e.the production of pro-inflammatory mediators (Fernandez-Ruiz,
2012).
The neuroprotective effects of CBD were also evaluated in animal models with
Parkinson’s disease, Huntington’s disease and neonatal ischemia. Pathological changes
induced by 6-hydroxydopamine, a toxic compound that targets catecholaminergic cells,
were significantly reduced by CBD treatment (Lastres-Becker, Molina-Holgado, Ramos,
Mechoulam and Fernandez-Ruiz, 2005; Mechoulam, et al., 2002). It appears that these
reductions after CBD treatment were irreversible, as they did not recover when CBD was
halted (Mechoulam, Peters, Murillo-Rodriguez and Hanus, 2007). CBD was also useful

22
in preventing beta-amyloid–induced neurodegeneration in in vivo and in vitro models of
Alzheimer disease (Fernandez-Ruiz, 2012). Increased production of reactive oxygen
species, lipid peroxidation, DNA fragmentation, and intracellular Ca2+
concentrations
induced by beta-amyloid peptide were significantly reduced after CBD treatment of PC12
cells (Iuvone et al., 2004).
CBD has also been shown to be protective against neuronal damage due to
ischemia (Hampson et al., 2000). In rats subjected to middle cerebral artery occlusion,
infarct size and neurological impairment were reduced by 50-60% by CBD. Similarly,
post-ischemic administration of CBD protected against hyperlocomotion and neuronal
injury following middle cerebral artery occlusion in gerbils (Braida et al., 2003).
Furthermore, it was recently shown that CBD treatment reduced the infarct volume in
brain ischemia, and that this effect was independent of cannabinoid receptor type 1and
transient receptor potential V1, but sensitive to the 5-HT1A receptor antagonist,
WAY100135 (Hayakawa et al., 2004; Mishima et al., 2005) suggesting that activation of
5-HT1A receptors mediate the neuroprotective effects of CBD.
c. Treatment of nausea and vomiting
Cannabis has long been used to treat nausea and vomiting induced by various
drugs and pathological conditions (Parker, Rock and Limebeer, 2011). Several
combinations of phytocannabinoids and synthetic cannabinoids have currently been
approved for use in the treatment of chemotherapy-induced nausea and vomiting (Parker
et al., 2011).The reduction of nausea and vomiting demonstrated by cannabis sativa use
in vivo and in vitro has been attributed to the presence of THC and CBD. Unlike THC,
which exerts its action via CB1 receptors, CBD mimics the anti-nausea and anti-vomiting

23
properties of THC through mechanisms unrelated to CB receptor activation (Mechoulam
et al., 2007). Interestingly, one of the most effective antiemetic drugs used in clinics is a
5HT3 receptor antagonist (Thompson, Chau, Chan and Lummis, 2006). 5HT3 receptors
belong to a superfamily of ligand-gated ion channels that mediate serotonin-induced
rapid depolarizations in intestinal neurons and regulate peristaltic activity of intestinal
smooth muscle (Izzo and Camilleri, 2009). CBD and THC have been shown to inhibit
5HT3receptors (K. H. Yang et al., 2010)and suppress emesis induced by various chemical
and physical stimuli (Parker et al., 2011).

24
d. Cardiovascular effects
i) Anti-arrhythmic role of CBD:
Studies examining the cardioprotective effects of CBD showed that CBD has
antiarrythmic effects after an ischemic reperfusion injury (Walsh, Hepburn, Kane and
Wainwright, 2010). In models of myocardial infarction following ligation of the left
anterior descending coronary artery, in vivo treatment with CBD resulted in a reduction
in infarct size (Rajesh et al., 2010). This finding has been attributed to an immuno-
modulatory effect in CBD, since it was accompanied by a reduction in leukocyte
infiltration and interleukin 6 concentrations. A single dose of CBD given 10 minutes
prior to ischemia or reperfusion resulted in a reduction in infarct size, and a significant
reduction in the incidence of ventricular tachycardia and total number of ventricular
ectopic beats (Stanley, Hind and O'Sullivan, 2013).
The CB1 receptor antagonist, AM251, also demonstrates antiarrhythmic
properties, and treatment with AM251 co-administered with CBD has a synergistic
effect, suggesting that activation of CB1 receptors is proarrhythmic (Walsh et al., 2010).
The synergism observed, which persists when CB1 receptors are blocked prior to CBD
administration, is suggestive of cross talk between CB1 and other CB receptors in the
ischemic heart (Walsh et al., 2010).Furthermore, CBD significantly reduces cardiac
dysfunction in mice with diabetes. CBD treatment has been shown to decrease
myocardial inflammation, reduce oxidative stress as demonstrated by a reduction in
nuclear factor-kB activation, suppress mitogen-activated protein kinase activation, and
reduce the expression of adhesion molecules and tumor necrosis factor (Walsh et al.,
2010).

25
ii) Vascular effects of CBDs:
CBDs exert vasodilative effects as demonstrated by in vivo and in vitro models
(see Stanley, Hind and O'Sullivan, 2013). However, the mechanism and potency of CBD
appear to differ between various experimental models. For example, in some studies,
CBD acts through cyclooxygenase, while other experiment use through CB1 receptor
antagonism and the inhibition of potassium (K+) channel hyperpolarization. The size of
the blood vessel is also a key factor, as maximal responses to anandamide (AEA) are
observed in small resistance vessels (O'Sullivan, Kendall and Randall, 2005).
In rat aorta, time-dependent CBD vasodilatation has been shown to be mediated
through Gi/o protein that is sensitive to pertussis toxin, but is not sensitive to CB1
antagonism or TRPV1 desensitization (O'Sullivan, Sun, Bennett, Randall and Kendall,
2009). Additional differences are observed in other species. The mechanism by which
vasodilatation occurs is also dependent on the type of cannabinoid. For instance, AEA
and N-arachidonoyl-dopamine (NADA) exert similar vasodilatation effects in rat aorta;
however, these effects occur through different mechanisms (Stanley, et al., 2013).
It has also been demonstrated that CBD induces vasodilatation of segments of
human mesenteric artery preconstricted with U46619 and endothelin-1. The authors of
this study determined that mesenteric artery relaxation is endothelium-dependent, and
involves CB1 receptor and TRPV channel activation, NO release and K+ channel
hyperpolarization (Stanley et al., 2013).
iii) Relationship between CBD and peroxisome proliferator-activated receptor
gamma:

26
Nuclear receptors are proteins found within cells that regulate diverse functions,
such as homeostasis, reproduction, development, and metabolism. Peroxisome
proliferator-activated receptor gamma (PPARγ) belongs to a family of nuclear receptors
and plays a crucial role in glucose and lipid homeostasis, in addition to cell proliferation,
cell differentiation, and inflammatory responses (Bishop-Bailey, 2000; Hsueh and
Bruemmer, 2004). There is evidence to suggest that cannabinoids bind to and activate
PPARγ, thereby causing PPAR-mediated responses. THC, which is the major component
of cannabis, causes time-dependent endothelium-dependent and PPARγ-mediated
vasodilatation of rat aorta. In contrast, CBD, which is a weak agonist of PPARγ
receptors, causes increased PPARγ transcriptional activity in PPARγ-overexpressing
HEK293 cells (Hsueh and Bruemmer, 2004).
Since CBD is a weak PPARγ receptor agonist, side effects normally associated with
PPARγ receptor activation, such as weight gain, edema, and increased plasma lipoprotein
can be avoided. Therefore, CBD may prove to have therapeutic potential as a low affinity
agonist (O'Sullivan et al., 2009; Walsh et al., 2010).
iv) Hemodynamic effects of CBD:
Studies on the hemodynamic effects of CBD proved inconclusive. In some studies,
a 16-mm Hg reduction in mean arterial blood pressure was noted, with no change in heart
rate (Walsh et al., 2010). The authors of this study concluded that the effects of CBD are
best seen in models with elevated blood pressure (Walsh et al., 2010). However, other
studies did not demonstrate significant hemodynamic changes (Bergamaschi, Queiroz,
Zuardi and Crippa, 2011).

27
CBD is reported to have anxiolytic properties, and is effective as a treatment for a
fear of public speaking. CBD reduced heart rate and blood pressure responses in Wistar
rats conditioned by fear. This response is believed to be mediated by 5HT1A receptors,
since the effects were inhibited by the 5HT1A receptor antagonist, WAY100635 (Gomes,
Resstel and Guimaraes, 2011; Zuardi, 2008).
v) Vascular protective effects of CBD:
High glucose levels are known to contribute to endothelial dysfunction in patients
with diabetes. High glucose levels promote the inhibition of endothelial NO, decrease the
vasodilatation effects and increase the vasoconstrictor effects of prostanoids, increase
superoxide production, and increase ROS production (Vanhoutte, Shimokawa, Tang and
Feletou, 2009). The aforementioned effects of high glucose were reduced when cells
were co-incubated with CBD (Rajesh et al., 2007). Key elements of atherosclerosis, such
as monocyte adhesion and transendothelial lmigration are reduced by CBD. Neither CB1
nor CB2 receptors appear to be responsible for mediating these effects of CBD (Rajesh et
al., 2007). Treatment with CBD may also protect against diabetic retinopathy. CBD
prevented vascular hyperpermeability at the blood-retinal barrier (BRB), and protected
the retina against oxidative damage, inflammation, and increased levels of cell adhesion
molecules in an in vivo model of diabetic retinopathy (El-Remessy et al., 2006).

28
1.5 Contraction of Cardiac Muscle
Contraction of cardiac muscle cells or cardiomyocytes shows certain characteristics
specific to this muscle. For example, contraction of cardiomyocytes is not initiated by
neurons as in skeletal muscle but by electrical excitation originating from the heart’s own
pacemaker, the sinoatrial node, which generates spontaneous and periodic action
potentials. When an action potential is initiated in one cell, current flows through the gap
junctions and depolarizes neighboring cells. If depolarization causes the membrane
potential to be more positive than the threshold, self-propagating action potentials occur
in the neighboring cells as well. Thus, the generation of an action potential is just as
critical for initiating contraction in cardiac muscle as it is in skeletal muscle, but it is
triggered by the sinoatrial node and the specialized conduction system of the heart
(Figure 4).
Figure 4. Anatomy and Function of the Heart's
Electrical System. Johns Hopkins Medicine,
Baltimore, Maryland.

29
Separate tubular structures, the transverse tubules (T tubules), cross the cell. In the
cardiac myocyte, the T tubule crossings occur at the Z-line, in contrast to the A-I junction
in skeletal muscle. The lumen of the T tubule is continuous with the extracellular fluid
surrounding the cell and, as in skeletal muscles, the action potential is propagated down
the T tubule (Ferrantini et al., 2013). Adjacent cardiac myocytes are joined end-to-end at
structures known as intercalated disks. These always occur at a Z-line (Figure 5). At these
points, the cell membranes form a number of parallel folds and are tightly held together
by desmosomes. This results in strong cell-to-cell cohesion, thus allowing the contraction
of one myocyte to be transmitted axially to the next. Gap junctions exist between cardiac
muscle cells, providing low resistance pathways for the spread of excitation from one cell
to another.
Figure 5. Adjacent cardiac myocytes are joined end-to-end at
structures known as intercalated disks.
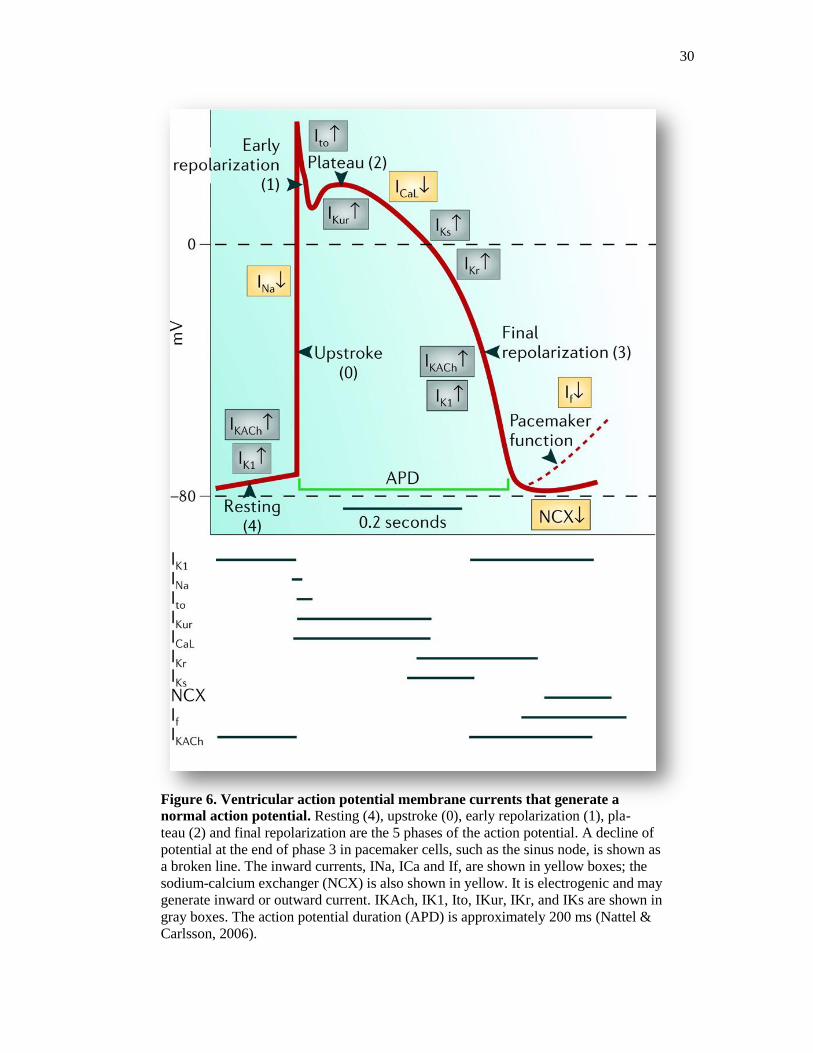
30
Figure 6. Ventricular action potential membrane currents that generate a
normal action potential. Resting (4), upstroke (0), early repolarization (1), pla-
teau (2) and final repolarization are the 5 phases of the action potential. A decline of
potential at the end of phase 3 in pacemaker cells, such as the sinus node, is shown as
a broken line. The inward currents, INa, ICa and If, are shown in yellow boxes; the
sodium-calcium exchanger (NCX) is also shown in yellow. It is electrogenic and may
generate inward or outward current. IKAch, IK1, Ito, IKur, IKr, and IKs are shown in
gray boxes. The action potential duration (APD) is approximately 200 ms (Nattel &
Carlsson, 2006).

31
1.5.1 Excitation–Contraction Coupling
a. The ventricular action potential
The cardiac action potential represents changes in the membrane potential due to
the movement of ions across the cell membrane through voltage-gated ion channels,
pumps, and exchangers, and has distinct characteristics in different regions of the heart.
In ventricular cardiomyocytes (Amin, Tan and Wilde, 2010). According to kinetic
properties, the cardiac action potential is divided into 4 distinct phases (Figure 6). The
first phase of the action potential is characterized by the activation of fast Na+ channels,
which open briefly to produce an influx of positive Na+ ions causing rapid depolarization
of the cell membrane and an upstroke of the action potential. This initial phase is called
phase 0. Following phase 0, a brief repolarization is observed at the peak of the action
potential and is the consequence of both fast Na+ channel inactivation and the activation
of initial repolarizing currents. This brief repolarization phase is called phase 1.
Following phase 1, a long lasting plateau of depolarization (100-200 ms), named phase 2,
is observed. Depolarizing currents, mainly late Na+ and L-type Ca
2+ channels stay open
and continue to offset repolarizing currents during the plateau phase. Finally, L-type Ca2+
channels begin to inactivate and several cardiac K+ channels such as delayed-rectifier and
inward-rectifier K+ channels begin to open and cause the repolarization of the membrane
to resting membrane potential. This repolarization is called phase 3. The ventricular
action potential is regenerative, and continues to excite ventricular cells if the stimulus
exceeds the critical threshold for depolarization as determined by availability of Na+
current. Phase 4 is the resting membrane potential, and describes the membrane potential
when the cardiomyocyte is not being stimulated. In a great majority of contracting

32
cardiomyocytes phase 4 has a low slope (almost a horizontal line). However, in
pacemaking cells, phase 4 is unstable (phase 4 - is the pacemaker potential). In
pacemaker cells (such as the Sinoatrial node), the membrane slowly depolarizes during
phase 4, until it reaches a threshold potential (around -40mV) or until it is depolarized by
an electrical impulse coming from another cell. The reason for this pacemaker potential is
an increased inward current of sodium (Na+) through voltage-dependent channels, but
also an increased inward Ca2+
current and a decrease in the K+ outward current. These
Na+ channels, in cardiac pacemaker cells, contrary to what usually happens in other cells,
open when the voltage is more negative.
b. Calcium-induced calcium release
The action potential propagates along the sarcolemma, invades the transverse
tubules and causes release of Ca2+
from internal Ca2+
stores such as sarcoplasmic
reticulum and leads eventually to the initiation of muscle contraction. This sequence of
events, beginning with action potential generation and resulting in muscle contraction is
known as excitation-contraction coupling (Figure 7). Initially, depolarization caused by
the action potential activates voltage-gated L-type Ca2+
channels which are concentrated
in the Tubules (Shepherd and McDonough, 1998). Activation of these channels induces a
conformational change in the ion channel structure and causes a subsequent influx of
Ca2+
into the cell. This influx of Ca2+
is known as sparklets (Navedo and Santana, 2013).
The presence of sparklets is not sufficient to cause activation of the contractile
machinery. However, it is sufficient to initiate the release of furtherCa2+
from intracellular
stores by interacting with ryanodine receptors (RyRs) on the sarcoplasmic reticulum. The
local amplification of intracellular Ca2+
mediated by RyR activation is also known as a

33
Ca2+
spark (Fearnley, Roderick and Bootman, 2011), and is essential for causing a Ca2+
transient that activates the contractile myofilaments resulting in muscle contraction. In
general, cytosolic Ca2+
influx in ventricular cells occurs mainly through two distinct
sources. About 8 to 23% of Ca2+
enters extracellularly through the opening of L-type
Ca2+
channels and 77% to 92% of total Ca2+
employed in muscle contraction is released
from the sarcoplasmic reticulum through RyRs, although some inter-species differences
do exist (Bers, 2008). The contribution of NCX and mitochondria to cytosolic Ca2+
is
usually not more than 1% (Bers, 2000). Following its release, Ca2+
is pumped back into
the sarcoplasmic reticulum by a Ca2+
-ATPase pump and also removed from the cell via
the Na+/Ca
2+ exchanger located at the plasma membrane (Wang, Song, Lakatta and
Cheng, 2001).In cardiac muscle, the activity of a Ca2+
-ATPase in the sarcoplasmic
reticulum is inhibited by the regulatory protein phospholamban. When phospholamban is
phosphorylated by cAMP-dependent protein kinase, its ability to inhibit the Ca2+
-ATPase
is lost. Thus, activators of cAMP-dependent protein kinase, such as the neurotransmitter
epinephrine, may enhance the rate of cardiac myocyte relaxation.

34
Figure 7. Cardiac excitation contraction coupling. Inset shows the time course of an action
potential, Ca2+
transient and contraction measured in a rabbit ventricular myocyte at 37 °C. NCX,
Na+/Ca
2+ exchange; ATP, ATPase; PLB, phospholamban; SR, sarcoplasmic reticulum (Bers -
2002).
Calcium Channels
Due to the importance of Ca2+
in various vital cellular events such as muscle
contraction, neurotransmitter release, and exocytosis; the regulation of Ca2+
entrance
through voltage-gated Ca2+
channels has been studied in detail (Catterall, Perez-Reyes,
Snutch and Striessnig, 2005). According to pharmacological and biophysical
characteristics three different types of Ca2+
channels have been characterized. These
different types of Ca2+
channels are summarized in Table 2.

35
In the heart, both T-type and L-type Ca2+
channels have been shown to be
expressed (Zhou and January, 1998). However, T-type Ca2+
channels appear to be
localized mainly in cells involved in heart rhythm and automaticity such as pacemaker
and atrial cells, while L-type Ca2+
channels are localized mainly in ventricular myocytes.
As mentioned above, due to their slow activation kinetics, L-type Ca2+
channels require
long depolarizations lasting 100-200 ms. After the activation process, L-type Ca2+
channels are inactivated by various factors which include time, voltage, cytosolic Ca2+
concentration and calmodulin (Bers, 2000).

36
Table 2. Classification of calcium channels
Ca2+
current
type
α1
subunits
Specific
blocker
Principal physiological functions Inherited diseases
L Cav 1.1 DHPs Excitation-contraction coupling in
skeletal muscle, regulation of
transcription
Hypokalemic periodic
paralysis
Cav1.2 DHPs Excitation contraction coupling in
cardiac and smooth muscle, endocrine
secretion, neuronal Ca2+
transients in
cell bodies and dendrites, regulation of
enzyme activity, regulation of
transcription
Timothy syndrome: cardiac
arrhythmia with
developmental abnormalities
and autism spectrum
disorders
Cav1.3 DHPs Visual transduction Stationary night blindness
N Cav2.1 ω-CTx-GVIA Neurotransmitter release, Dendritic
Ca2+
transients
P/Q Cav2.2 ω-Agatoxin Neurotransmitter release, Dendritic
Ca2+
transients
Familial hemiplegic
migraine, cerebellar ataxia
R Cav2.3 SNX-482 Neurotransmitter release, Dendritic
Ca2+
transients
T Cav3.1 None Pacemaking and repetitive firing
Cav3.2 Pacemaking and repetitive firing Absence seizures
Cav3.3
Abbreviations: DHP, dihydropyridine; Ѡ-CTx-GVIA, Ѡ –conotoxin GVIA from the cone snail
Conusgeographus; SNX-482, a synthetic version of a peptide toxin from the tarantula
Hysterocratesgigas (Catterall, 2011).
As shown in Figure 8, the L-type Ca2+
channel consists of 4 subunits: the major
pore-forming 1 subunit and a variety of accessory subunits. Theα1 subunit: Formed of 4
homologous domains (I–IV), each consisting of 6 transmembrane (TM) helices joined by
intracellular links. The loops between TM5 and TM6form the channel pore, while TM4
contains arginine or lysine residues that form the voltage sensor, which is thought to
move or twist as the membrane potential changes.
Ion selectivity is governed by a glutamate residue preserved in the pore-lining
loop of all 4 domains (Dolphin, 2006; J. Yang, Ellinor, Sather, Zhang and Tsien, 1993).
Two Ca2+
ions enter the cell at a time, are regulated by 2 glutamate residues and flow
down the Ca2+
concentration gradient (Heinemann, Terlau, Stuhmer, Imoto and Numa,
1992; J. Yang et al., 1993). Lanthanum, cobalt and cadmium inhibit the α1 subunit

37
(Dolphin, 2009). Expression of the α1 subunit without the auxiliary subunits, results in
low L-type Ca2+
channel expression levels, and abnormal voltage and kinetic properties
in the channel. The β subunit is located on the intracellular side of the channel. It interacts
with the subunit through the -interaction domain (AID), a guanylate cyclase–like
region located between domains I and II (Catterall, 2011; Pragnell et al., 1994). The AID
has a well-conserved gene sequence and also occurs in N-type and P/Q-type
Ca2+
channels (Richards, Butcher and Dolphin, 2004).The β subunit alters channel
expression, voltage dependence, and gating kinetics. Four β subunits have been cloned,
and all except for β2a hyperpolarize activation and steady-state inactivation of Ca2+
(Dolphin, 2003). The α2δ subunit is an accessory subunit consisting of the
transmembrane domain, and 2, an extracellular domain. Four 2 isoforms have been
identified. Co-expression of α2δ subunits increases Ca2+
channel expression and function.
The subunit is a 30 to 40kD inhibitory protein consisting of 4 TM helices (Takahashi,
Seagar, Jones, Reber and Catterall, 1987).

38
Figure 8. Calcium channel subunits. Structure of calcium channels showing the five subunits α,
β, α2δ and (Figure modified from Catterall, 2011).
Secretion of catecholamines from the adrenal medulla and the autonomic nervous
system stimulates β-adrenergic receptors via adenylcyclase, cAMP, and protein kinase A
(PKA) to produce positive chronotropic, inotropic and lusitropic effects. Cardiac L-type
Ca2+
channels are modulated through the β-adrenergic/cAMP signaling pathway; α1 and β
subunits are phosphorylated by PKA (De Jongh et al., 1996; Haase, Bartel, Karczewski,
Morano& Krause, 1996; Hell et al., 1993; Puri, Gerhardstein, Zhao, Ladner and Hosey,
1997). β-adrenergic receptor activation causes a 2-fold increase in the L-type Ca2+
current
by increasing the number of channels and their activation probability. The latter is
mediated through PKA-mediated phosphorylation of the Cav1.2 (1C-subunit) channel
protein and/or associated proteins (Catterall, 2010). As opposed to the adrenergic system,
reciprocal control over Ca2+
entry is provided by ACh, acting through muscarinic Ach

39
receptors, raising intracellular cGMP concentrations and causing cGMP-dependent
phosphorylation of the L-type Ca2+
channels. In turn, the cGMP-dependent
phosphorylation of L-type Ca2+
channels, at sites distinct from those phosphorylated by
the cAMP-dependent kinase, causes a decrease in Ca2+
influx during the cardiac action
potential and thus a decrease in the force of contraction.

40
1.5.2 Sarcoplasmic Reticulum and Ryanodine Receptors
The sarcoplasmic reticulum has many roles in the regulation of muscle
contraction and intracellular Ca2+
homeostasis. The release of Ca2+
from this organelle
causes increased Ca2+
levels to activate various Ca2+
dependent enzymes such as
calmodulin and adenyl cyclase (Fearnley et al., 2011). It is also the site of phospholipid
generation and it communicates with other intracellular organelles, such as the
mitochondria. However, it’s most important function is the regulation of Ca2+
homeostasis. It is well known that cytosolic Ca2+
levels during resting conditions are in
the nM range, while extracellular Ca2+
concentrations are much higher (μM range). As
mentioned above, following an action potential, cytosolic Ca2+
concentration increases
and activates ryanodine receptors, which amplify the Ca2+
signal and stimulate Ca2+
-
induced Ca2+
release. Several isoforms of ryanodine receptors (RyR) have been identified:
RyR1 is expressed in skeletal muscle, RyR2 in cardiac muscle and the brain, and RyR3 in
the brain (Lanner, Georgiou, Joshi and Hamilton, 2010). Although the ryanodine receptor
is a protein of thr SR membrane, 80% of this protein is located in the cytoplasm. The
intracellular position of ryanodine differs from one cell type to another (Zucchi and
Ronca-Testoni, 1997). It can be directly attached (as in skeletal muscles) or detached (as
in cardiac muscles) to the L-type Ca2+
channel. In skeletal muscle, direct interaction of
the ryanodine receptor with the L-type Ca2+
channel causes fast excitation-contraction
coupling and rapid initiation of skeletal muscle contraction (within a few minutes). In
cardiac muscle on the other hand, excitation-contraction is a relatively slow process
(within hundreds of minutes). Function of the ryanodine receptor is also regulated by
several ligands, including Ca2+
, calmodulin and caffeine (Fearnley et al., 2011).

41
1.5.3 The Contractile Machinery
Myofilaments occupy about half of the cell volume in mammalian ventricular
myocytes. As with skeletal muscle, cardiac myocytes contain the contractile proteins
actin (thin filaments) and myosin (thick filaments) together with the regulatory proteins
troponin and tropomyosin. In their structural configuration, the thin filament comprises
actin with a spiraling backbone of tropomyosin, along with troponin complexes (C, I and
T) that occur at certain intervals (Figure 9). Each actin filament consists of a site where
myosin can bind, but these sites are blocked by tropomyosin when cytosolic Ca2+
concentration is low. When cytosolic Ca2+
concentration increases due to Ca2+
release,
Ca2+
binds to troponin C causing tropomyosin to shift, thereby exposing the myosin
binding sites. Adenosine diphosphate (ADP) and phosphate, present on the myosin head
since the previous movement cycle, are released when myosin attaches to myosin binding
sites on the actin filaments. The energy generated in the head is used to initiate upstroke
and sliding, until the actin–myosin complex is liberated by adenosine triphosphate (ATP)
binding the myosin head. ATP is hydrolyzed to ADP and phosphate, releasing energy
that is stored in the myosin head to be used in the subsequent stroke.
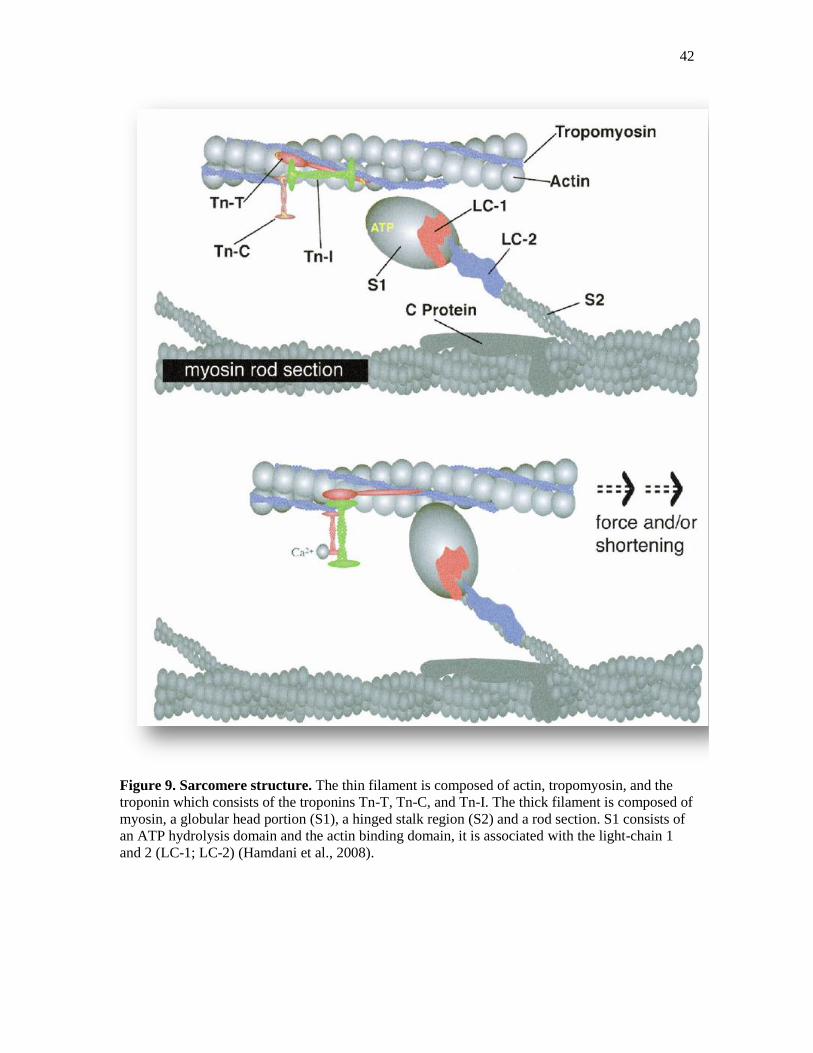
42
Figure 9. Sarcomere structure. The thin filament is composed of actin, tropomyosin, and the
troponin which consists of the troponins Tn-T, Tn-C, and Tn-I. The thick filament is composed of
myosin, a globular head portion (S1), a hinged stalk region (S2) and a rod section. S1 consists of
an ATP hydrolysis domain and the actin binding domain, it is associated with the light-chain 1
and 2 (LC-1; LC-2) (Hamdani et al., 2008).

43
1.5.4 Calcium Sensitivity of Myofilaments
The term myofilament Ca2+
sensitivity came into use after it was discovered that
the force of contraction is not only dependent on the free cytosolic Ca2+
concentration,
but also on the affinity of troponin C for Ca2+
. Biochemical and physiological
experiments have shown that several factors affect myofilamentCa2+
sensitivity,
including pH, temperature, ionic strength and troponin I phosphorylation (Ruegg, 2003).
Furthermore, these factors can be categorized by whether they affect thick or thin
filaments (Bers, 2001). Factors that affect thick filaments tend to affect the relationship
between intracellular Ca2+
concentration and the extent of myosin light chain
phosphorylation. Whereas factors that affect thin filaments, affect phosphorylation of
myosin light chain and force generation due to contractile proteins (Bers, 2001).

44
Chapter 2: Methods
2.1 Ventricular Myocyte Isolation
Ventricular myocytes were isolated from adult male Wistar rats (264 ± 19 g)
according to previously described techniques. This study was carried out in accordance
with the recommendations in the Guide for the Care and Use of Laboratory Animals from
the National Institutes of Health. The protocol was approved by the Committee on the
Ethics of Animal Experiments at the United Arab Emirates University. In brief, the
animals were euthanized using a guillotine and their hearts were removed rapidly and
mounted for retrograde perfusion according to the Langendorff method (Figure 10). The
hearts were perfused at a constant flow of 8 ml g heart-1
min-1
and at 36–37 °C with a
solution containing (mM): 130 NaCl, 5.4 KCl, 1.4 MgCl2, 0.75 CaCl2, 0.4 NaH2PO4, 5
HEPES, 10 glucose, 20 taurine and 10 creatine set to pH 7.3 with NaOH. When the heart
had stabilized perfusion was continued for 4 minutes with a Ca2+
-free isolation solution
containing 0.1 mM EGTA, and then for 6 minutes with cell isolation solution containing
0.05 mM Ca2+
, 0.75 mg/ml collagenase (type 1; Worthington Biochemical Corp., USA)
and 0.075 mg/ml protease (type X1V; Sigma, Germany). Ventricles were excised from
the heart, minced and gently shaken in collagenase-containing isolation solution
supplemented with 1% BSA. Cells were filtered from this solution at 4 minutes intervals
and resuspended in an isolation solution containing 0.75 mM Ca2+
.

45
Figure 10. Langendorff Heart
Solution
Chamber
Heating
Coil
Water jacketed
sugar chamber
Peristaltic
pump

46
2.2 Measurement of Ventricular Myocyte Shortening
Ventricular myocytes were allowed to settle on the glass bottom of a Perspex
chamber mounted on the stage of an inverted microscope (Axiovert 35, Zeiss, Germany).
Myocytes were superfused (3–5 ml/min) with normal Tyrode (NT) containing (mM): 130
NaCl, 5 KCl, 1 MgCl2, 10 glucose, 5 HEPES, 20 taurine, 10 creatine and 0.75 CaCl2 (pH
7.4). Shortening of myocytes was recorded using a video edge detection system (VED-
114, Crystal Biotech, USA) according to previously described techniques (Howarth,
Qureshi and White, 2002). Resting cell length (RCL) time to peak (TPK), time to half
(THALF) relaxation and amplitude of shortening (expressed as a percentage of resting
cell length) were measured in electrically stimulated (1 Hz) myocytes maintained at 35–
36 °C. Data was acquired and analyzed with signal averager software v 6.37 (Cambridge
Electronic Design, UK). Experimental solutions were prepared from stock immediately
prior to each experiment.
2.3 Measurement of Intracellular Ca2+
Concentration
Myocytes were loaded with the fluorescent indicator fura-2 AM (F-1221,
Molecular Probes, USA) as described (Howarth et al., 2002). In brief, 6.25 μl of a 1 mM
stock solution of fura-2 AM (dissolved in dimethyl sulphoxide) was added to 2.5 ml of
cells to give a final fura-2 concentration of 2.5 μM. Myocytes were gently shaken for
10 minutes at 24 °C (room temperature). After loading, myocytes were centrifuged,
washed with NT to remove extracellular fura-2 and then left for 30 min to ensure
complete hydrolysis of the intracellular ester. Intracellular Ca2+
was measured according
to previously described techniques (Howarth et al., 2011). Myocytes were alternately
illuminated by 340 and 380 nm light using a monochromator (Cairn Research, UK)

47
which changed the excitation light every 2 minutes. The resulting fluorescence emitted at
510 nm was recorded by a photomultiplier tube and the ratio of the fluorescence emitted
at the two excitation wavelengths (340/380 ratio) was calculated to provide an index of
intracellular Ca2+
concentration. Resting fura-2 ratio, time to peak (TPK), time to half
(THALF) decay of the Ca2+
transient, and the amplitude of the Ca2+
transient were
measured in electrically stimulated (1 Hz) myocytes maintained at 35-36 . Data were
acquired and analyzed with signal averager software v 6.37 Cambridge Electronic
Design, UK)
2.4 Measurement of Sarcoplasmic Reticulum Ca2+
Content
Sarcoplasmic reticulum (SR) Ca2+
release was assessed using previously described
techniques (Howarth et al., 2002). After establishing a steady state of Ca2+
transients in
electrically stimulated (1 Hz) myocytes maintained at 35–36 °C and loaded with fura-2,
stimulation was paused for a period of 5 seconds (Figure 11). Caffeine (20 mM) was then
applied for 10 seconds using a solution switching device customized for rapid solution
exchange. Electrical stimulation was resumed and the Ca2+
transients were allowed to
recover to a steady state. SR-releasable Ca2+
was assessed by measuring the area under
the curve of the caffeine-evoked Ca2+
transient. Fractional release of SR Ca2+
was
assessed by comparing the amplitude of the electrically evoked steady state
Ca2+
transients with that of the caffeine-evoked Ca2+
transient and refilling of the SR was
assessed by measuring the rate of recovery of electrically evoked Ca2+
transients
following the application of caffeine.

48
2.5 Assessment of Myofilament Sensitivity to Ca2+
In some cells shortening and fura-2 ratio were recorded simultaneously.
Myofilament sensitivity to Ca2+
was assessed from phase-plane diagrams of fura-2 ratio
versus cell length by measuring the gradient of the fura-2-cell length trajectory during
late relaxation of the twitch contraction. The position of the trajectory reflects the relative
myofilament response to Ca2+
and hence, can be used as a measure of myofilament
sensitivity to Ca2+
(Spurgeon et al., 1992; Howarth et al., 2002).
Figure 11. The recording system used for video edge detection and Ca2+ imaging
experiments: The system includes an anti-vibration table (A), a Faraday cage (B), an
inverted microscope (C), a cell superfusion system (D) with a temperature control
system (E), a video edge motion detector (F), a microfluorescence photometry
system (G) and an electrical stimulator (H).

49
2.6 Whole-Cell Recordings
Whole-cell recordings were performed in isolated ventricular myocytes. The
myocytes were placed in the recording chamber (0.4 mL) mounted on the stage of an
inverted microscope (CK2, Olympus, Tokyo, Japan) (Figure 12). After settling to the
bottom of the chamber, cells were superfused with an external solution for 10 min at a
rate of 2–3 mL.min-1
at room temperature. The external solution contained (in mM) 120
NaCl, 1 EGTA, 10 MgCl2, 10 Glucose, 10 HEPES, 1 NaH2PO4 and 5 KCl.
Transmembrane currents were recorded with an axopatch amplifier 200 B (Axon
Instruments, Inc., Foster City, CA, USA). The whole-cell bath solution contained (in
mM): 95 NaCl, 50 TEACl, 2 MgCl2, 2 CaCl2, 10 HEPES and 10 Glucose (adjusted to
pH 7.35 with NaOH). Glass microelectrodes were made using a microelectrode puller
(Model P-97, Sutter Instrument Co.,Novato, CA, USA) (Figure 13) by two-stage pulling,
and had a resistance of 2.0–4.0 MΩ when filled with electrode internal solution
containing (in mM) 140 CsCl, 10 TEACl, 2 MgCl2, 2 HEPES 1 MgATP and 10 EGTA
(adjusted to pH 7.25 with CsOH). Only rod shaped cells with clear cross-striations were
used for the experiments. Figure 14 shows typical ventricular myocytes with patch
pipette in solution. After gigaseal formation (1-10 GΩ) by suction through a pipette, the
membrane was ruptured with a gentle suction to obtain the whole-cell voltage clamp
configuration. Membrane capacitance and series resistance were compensated after
membrane rupture to minimize the duration of capacitive current. The experiment was
conducted under room Temperature (22-24 ºC). To record L-type Ca2+
currents, the
external solution was changed to the Na+-free solution in which Na
+ was replaced by
equimolar TEA (TEA–Cl). The Na+
current was also inactivated at the holding potential -
50 mV. K+currents were suppressed by substituting K
+ by Cs
+. Computer-generated

50
voltage pulses were programmed using the pCLAMP 10.0 software (Axon Instruments).
Cells with stable calcium currents were then tested using different drugs
Figure 12. Patch clamp Setup
Power
supply for
micropump
ss
Light Source
Axopatch
Amplifier
200 B
Micropumps
Solution
Reservoirs
Manipulator

51
Figure 13. Microelectrode puller (model P-97, Sutter Instrument)

52
Figure 14. Glass electrode forming giga seal with ventricular
myocyte
Figure 15. Ventricular myocyte

53
Chapter 3: Results
3.1 Effects of Cannabidiol on Ventricular Myocyte Shortening
In Normal Tyrode (NT) solution, amplitudes of myocyte shortening in response to
electrical stimulation (1 Hz) gradually decreased to 85-80 % of controls during
experiments lasting up to 20 minutes. No further running down of the shortening
amplitude was observed in the NT solution containing 0.02 % DMSO. And, unless it
stated otherwise, DMSO (in the concentration used in CBD containing solution) was also
included routinely in the control NT solutions during the shortening and Ca2+
transient
experiments.
In initial control experiments and in line with earlier studies conducted under
similar conditions it has been shown that 0.01 % DMSO did not significantly affect
contractile parameters and Ca2+
current in rat ventricular myocytes(Sun et al., 2010). The
application of control solution (NT) containing dimethyl sulfoxide at the highest
concentration used in contractility studies (0.02 % v/v; used as vehicle for 1 μM CBD)
caused 11-13 % inhibition of the shortening (n=7-10; P<0.05 compared to a 0 time
point).
In the first set of experiments, we tested the effect of CBD on the contractility of
acutely isolated rat ventricular myocytes. Figure16A shows typical records of shortening
in myocytes superfused with either NT (in the absence of CBD, NT contained 0.02 %
DMSO in all experiments) or NT + 1 µM CBD and during washout with NT. The time
frame of the effect of CBD on amplitude of shortening is shown in Figure 16B. The
effect of CBD reached a steady level within 5 minutes of CBD application. Increasing the
incubation time to 10 minutes did not cause any further change in the amplitude of
shortening. Recovery from CBD inhibition was incomplete during the 5-10 minutes

54
washout time. In summary, the amplitude of shortening measured after 5 minutes bath
application of CBD was significantly reduced (paired t-test; n=17; P<0.05) by up to 46.4
± 3.1 % in the controls (Figure 16C). The effect of CBD increased in a concentration-
dependent manner. The concentration-response relationship normalized to maximal CBD
inhibition indicated that CBD suppresses shortening of amplitudes with an IC50 of 0.6
µM (Figure17). Among other contraction parameters measured, resting cell length
(RCL), time to peak and time to half relaxation were not significantly altered
(n=17; P>0.05) by 10 minutes superfusion with CBD.

55
Figure 16. Effects of cannabidiol on ventricular myocyte shortening. (A) Typical records of
shortening in an electrically stimulated (1 Hz) ventricular myocytes superfused with either NT
(containing the vehicle, 0.02% DMSO) or NT + 1 µM CBD and during washout with NT. (B)
Time course of the mean amplitudes (AMP) of shortening expressed as a percentage of control
values in vehicle (NT + 0.02% DMSO), and in presence of CBD (1 µM). Myocytes were
maintained at 35-36 °C and superfused with CBD for 10 minutes. (C) Data shown as means ±
S.E.M., n = 6-8 cells. * indicates statistically significant difference at the level of P< 0.05.
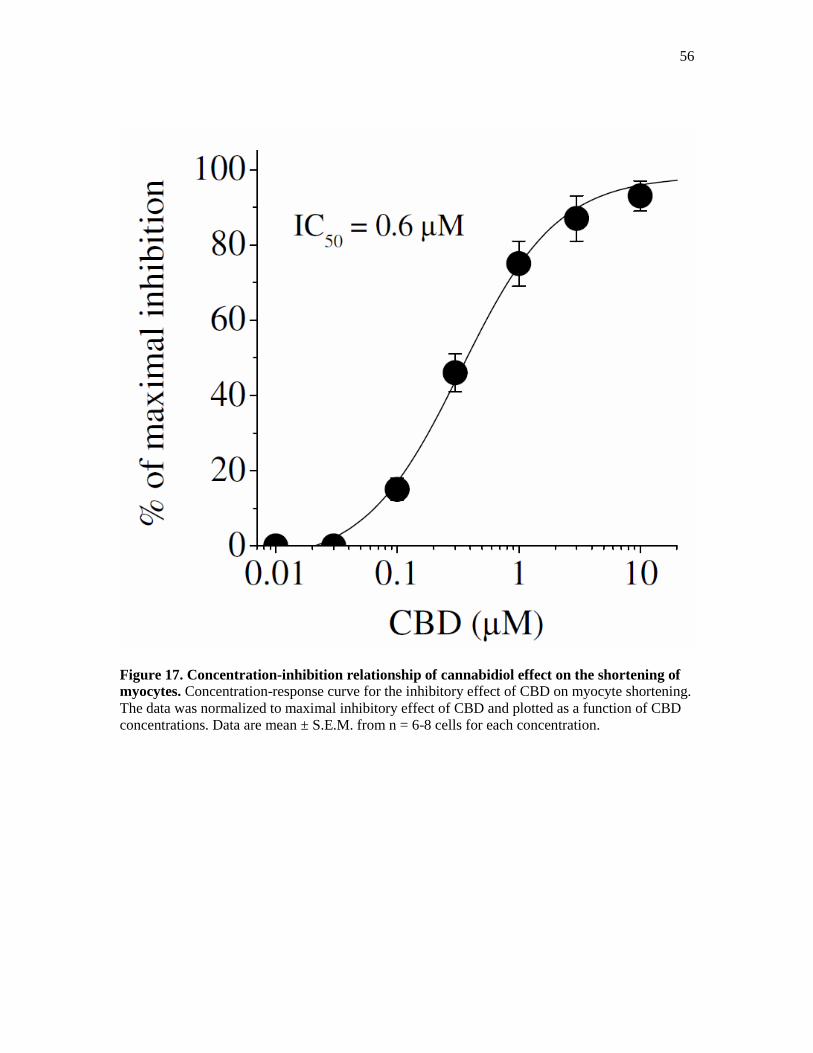
56
Figure 17. Concentration-inhibition relationship of cannabidiol effect on the shortening of
myocytes. Concentration-response curve for the inhibitory effect of CBD on myocyte shortening.
The data was normalized to maximal inhibitory effect of CBD and plotted as a function of CBD
concentrations. Data are mean ± S.E.M. from n = 6-8 cells for each concentration.

57
3.2 Effects of Cannabidiol on Intracellular Ca2+
In the second set of experiments we investigated the effects of 5 minutes bath
application of 1 µM CBD on the resting intracellular Ca2+
levels, and on the amplitudes
and kinetics of Ca2+
transients elicited by electrical-field stimulation. Typical records of
Ca2+
transients in a myocytes superfused with NT (containing 0.02 % DMSO), NT + 1
µM CBD and during washout with NT are shown in Figure18A. The effects of 1 µM
CBD on resting fura-2 ratio, TPK Ca2+
transient, THALF decay of the Ca2+
transient and
AMP of Ca2+
transients are shown in Figure 18B-E, respectively. Although, CBD has
been shown to alter intracellular Ca2+
levels in various types of cells (see Pertwee,
2008;Pertwee, et al., 2010), application of 1 µM CBD for 5 to 10 minutes did not cause a
significant alteration in resting fura-2 ratio, TPK Ca2+
transients, or THALF decay of the
Ca2+
transient (paired t-test; n=11-14 cells, P>0.05). However, the AMP of the Ca2+
transient was significantly reduced by 1 µM CBD (0.337 ± 0.043 fura-2 ratio units)
compared to 0.488 ± 0.056 fura-2 ratio units (paired t-test, n =11 cells) in the control
experiments.
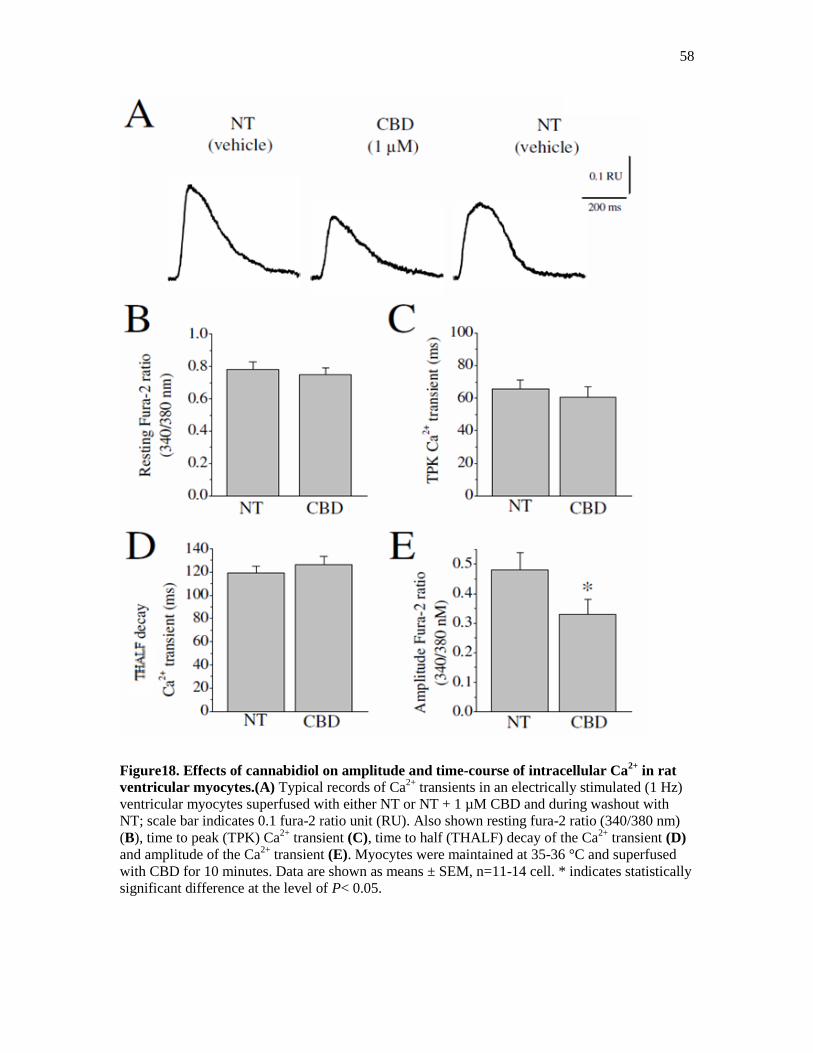
58
Figure18. Effects of cannabidiol on amplitude and time-course of intracellular Ca2+
in rat
ventricular myocytes.(A) Typical records of Ca2+
transients in an electrically stimulated (1 Hz)
ventricular myocytes superfused with either NT or NT + 1 µM CBD and during washout with
NT; scale bar indicates 0.1 fura-2 ratio unit (RU). Also shown resting fura-2 ratio (340/380 nm)
(B), time to peak (TPK) Ca2+
transient (C), time to half (THALF) decay of the Ca2+
transient (D)
and amplitude of the Ca2+
transient (E). Myocytes were maintained at 35-36 °C and superfused
with CBD for 10 minutes. Data are shown as means ± SEM, n=11-14 cell. * indicates statistically
significant difference at the level of P< 0.05.

59
3.3 Effect of Cannabidiol on Sarcoplasmic Reticulum Ca2+
Transport
The effect of CBD on sarcoplasmic reticulum Ca2+
transport was tested.
Figure19A shows a typical record illustrating the protocol used in these experiments.
Initially, the myocyte was electrically stimulated at 1Hz. Electrical stimulation was then
turned off for 5 seconds. Caffeine was then applied for 10 seconds using a rapid solution-
exchanger device. Electrical stimulation was then restarted, and the recovery of
intracellular Ca2+
was recorded during a 60 seconds period. The SR Ca2+
content was
assessed by measuring caffeine-evoked Ca2+
release (under the caffeine-evoked Ca2+
transient) and fractional release of Ca2+
by comparing the amplitude of the electrically
evoked steady-state Ca2+
transients with that of the caffeine-evoked Ca2+
transient in the
presence of either NT alone or NT with 1 µM CBD. Maximal amplitudes and the rates of
Ca2+
release by 20 mM caffeine remained unaltered after a 10 minute bath application of
1 µM CBD (Figure 19B) and (Figure 19C; paired t-test; n = 7 cells, P>0.05). Fractional
release of SR Ca2+
was not significantly altered in 1 µM CBD compared to NT (0.78 ±
0.04 in CBD versus 0.81 ± 0.07 in controls; paired t-test; n =8 cells; Figure 19D). The
recovery of the Ca2+
transients during electrical stimulation following application of
caffeine (Figure19E) was also not significantly altered in myocytes exposed to 1 µM
CBD myocytes compared to the control cells (paired t-test; n = 8 cells, P>0.05).
NT AN
0
0.2
0.4
0.6
0.8
Am
pli
tud
e o
f c
aff
ein
e-e
vo
ked
Ca
2+ t
ran
sie
nt
(Fu
ra 2
ra
tio
un
its)
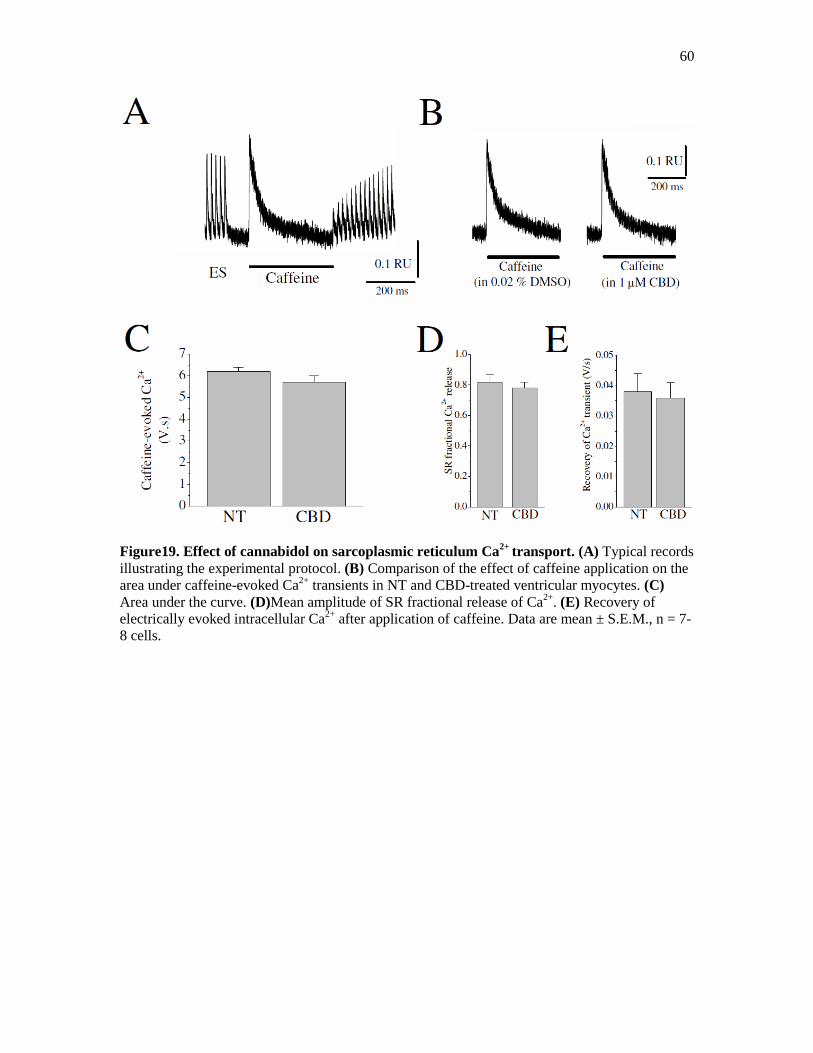
60
Figure19. Effect of cannabidol on sarcoplasmic reticulum Ca2+
transport. (A) Typical records
illustrating the experimental protocol. (B) Comparison of the effect of caffeine application on the
area under caffeine-evoked Ca2+
transients in NT and CBD-treated ventricular myocytes. (C)
Area under the curve. (D)Mean amplitude of SR fractional release of Ca2+
. (E) Recovery of
electrically evoked intracellular Ca2+
after application of caffeine. Data are mean ± S.E.M., n = 7-
8 cells.

61
3.4 Effect of Cannabidiol on Myofilament Sensitivity to Ca2+
The effects of CBD on myofilament sensitivity to Ca2+
were also investigated.
These experiments tested whether CBD decreases the mechanical response by altering
the affinity of the contractile machinery of the ventricular myocytes to intracellular Ca2+
.
Atypical record of myocyte shortening, fura-2 ratio and phase-plane diagrams of
fura-2 ratio versus cell length in myocytes exposed to NT is shown in Figure 20A. The
gradient of the trajectory reflects the relative myofilament response to Ca2+
and hence,
has been used as a measure of myofilament sensitivity to Ca2+
(Spurgeon, et al., 1992;
Howarth, et al., 2002). The gradients of the fura-2-cell length trajectory during late
relaxation of the twitch contraction measured during the periods 500-600 ms (Figure
20B), 500-700 ms (Figure20C), and 500–800 ms (Figure20D) were not significantly
altered in CBD compared to NT suggesting that myofilament sensitivity to Ca2+
is not
reduced by CBD (CBD-treatment was compared to NT containing 0.02 % DMSO, paired
t-test; n = 17 cells; P>0.05).
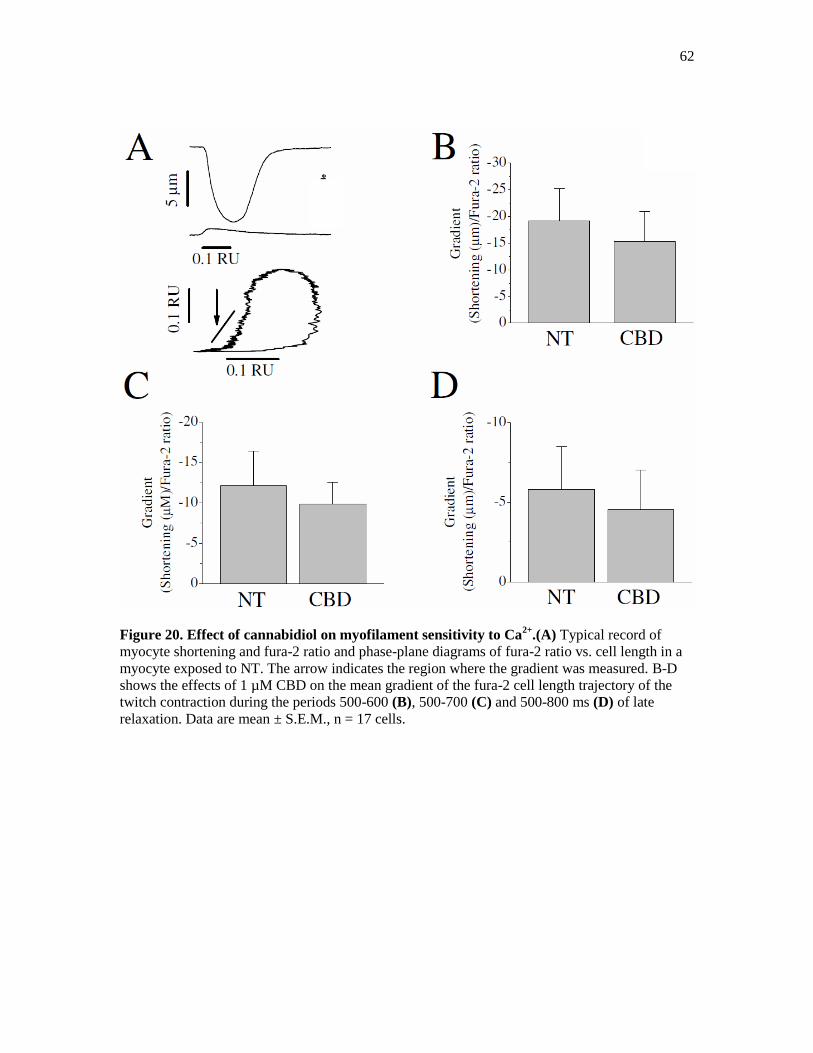
62
Figure 20. Effect of cannabidiol on myofilament sensitivity to Ca2+
.(A) Typical record of
myocyte shortening and fura-2 ratio and phase-plane diagrams of fura-2 ratio vs. cell length in a
myocyte exposed to NT. The arrow indicates the region where the gradient was measured. B-D
shows the effects of 1 µM CBD on the mean gradient of the fura-2 cell length trajectory of the
twitch contraction during the periods 500-600 (B), 500-700 (C) and 500-800 ms (D) of late
relaxation. Data are mean ± S.E.M., n = 17 cells.

63
3.5 Effects of Cannabidiol on the Action Potentials of Ventricular Myocytes
In this set of experiments, patch-clamped ventricular myocyte were exposed to
CBD while continuously monitoring their Vrest and APs in current clamp mode. The
generation of APs was stimulated by 0.9-1 nA depolarizing current pulses of 4 ms
durations. Since the intracellular pipette solution did not contain Ca2+
-chelating agents,
the generation of each AP was accompanied by a myocyte contraction. Therefore, current
pulses were applied at a frequency of 0.2 Hz. During a typical experiment the following
protocols were employed: first, whole-cell configuration was established and a 4 to 5
minute dialysis of the myocytes with a pipette solution was allowed to ensure an
equilibrium of conditions between the pipette solution and the intracellular milieu. After
achieving stable recordings of baseline electrical activity (Vrest and AP parameters),
myocytes were exposed to CBD for 5 to 10 minutes and was subsequently washed out.
The resting membrane potentials (mean ± SEM) were -77.4 ± 1.9 and -79.2 ± 1.7
mV in the control and after CBD treatment (n=9) myocytes, respectively (Figure 21A,
inset and Figure 21B). Maximal amplitudes of AP and the rising rate of AP (maximal
velocity; V/s, Figure 21C and Figure 21D) were not significantly altered in the presence
of CBD. CBD (1 µM) consistently shortened the duration of AP (measured at 60 % of
repolarization, APD60) (Figure 21A inset and Figure 21E). Changes in AP shortening in
response to CBD (1 µM) applications were noticeable within 20 to 30 seconds
(Figure21A). Recoveries were usually partial and required a longer time.
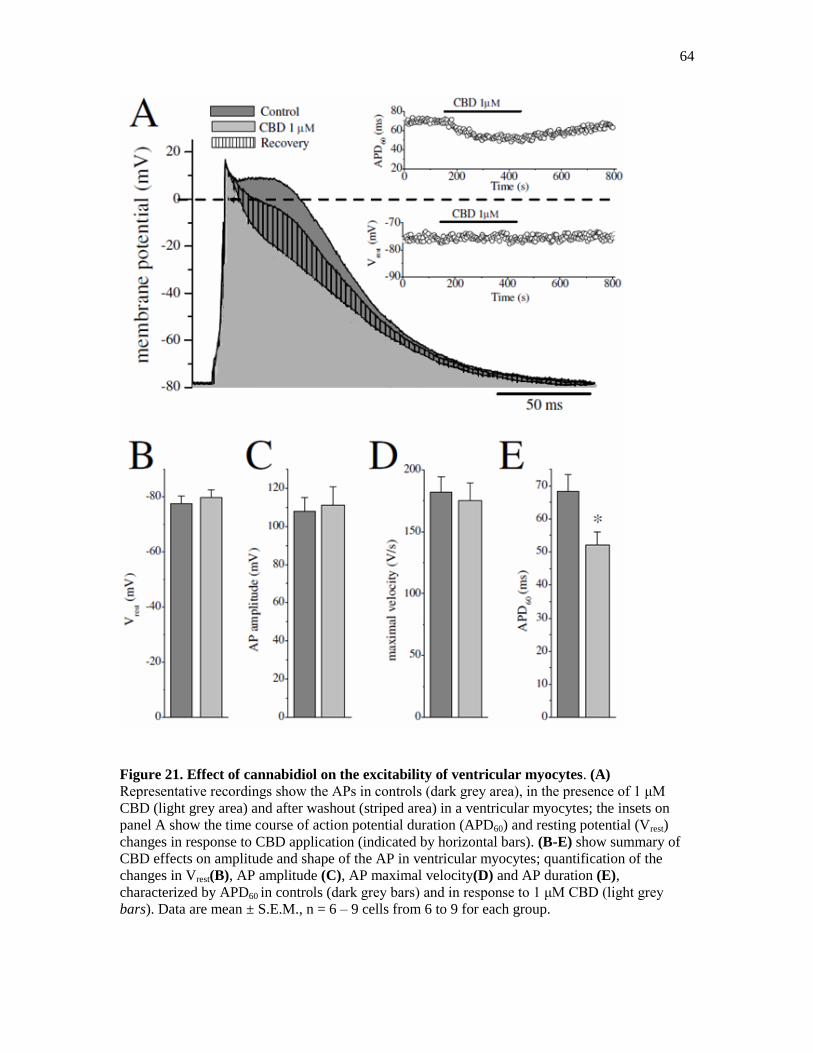
64
Figure 21. Effect of cannabidiol on the excitability of ventricular myocytes. (A)
Representative recordings show the APs in controls (dark grey area), in the presence of 1 μM
CBD (light grey area) and after washout (striped area) in a ventricular myocytes; the insets on
panel A show the time course of action potential duration (APD60) and resting potential (Vrest)
changes in response to CBD application (indicated by horizontal bars). (B-E) show summary of
CBD effects on amplitude and shape of the AP in ventricular myocytes; quantification of the
changes in Vrest(B), AP amplitude (C), AP maximal velocity(D) and AP duration (E),
characterized by APD60 in controls (dark grey bars) and in response to 1 μM CBD (light grey
bars). Data are mean ± S.E.M., n = 6 – 9 cells from 6 to 9 for each group.

65
3.6 Effect of Cannabidiol on L-type Ca2+
Currents
We also investigated the effect of CBD (1 µM) on L-type Ca2+
currents. Figure
22A shows a typical record of L-type Ca2+
currents elicited by applying a single 300 ms
voltage pulse to +10 mV from a holding potential of -50 mV in rat ventricular myocyte
before and after a 10 minute superfusion with 1µM CBD. The time frame of the effect of
CBD on the density of L-type Ca2+
currents is shown in Figure 22B. The application of a
vehicle (0.02% DMSO) for 10 minutes caused a 10-15% inhibition of the current density
of L-type Ca2+
currents in experiments lasting up to 15 to 20 min. The effects of CBD
were also investigated on the biophysical properties of L-type Ca2+
currents in rat
ventricular myocytes. L-type Ca2+
currents were recorded in the presence of intracellular
Cs+ and extracellular TEA
+ to suppress K
+ currents while retaining 95 mM Na
+ in the
extracellular solution. The elimination of contaminating Na+ current during the recording
of L-type Ca2+
currents was achieved by applying voltage step-pulses from relatively
depolarized Vh of -50 mV, which produced steady-state inactivation of sodium channels
(Voitychuk, et al., 2012). As evident from the original recordings and I-V relationships
(Figure23A), L-type Ca2+
currents started to appear at Vm = -30 mV, reached a
maximum at around Vm = +10 mV, and decreased at higher voltages approaching to zero
at about Vm = +60 mV (Figure23B). The maximal amplitudes of L-type Ca2+
currents
were suppressed in the presence of CBD (1 µM).
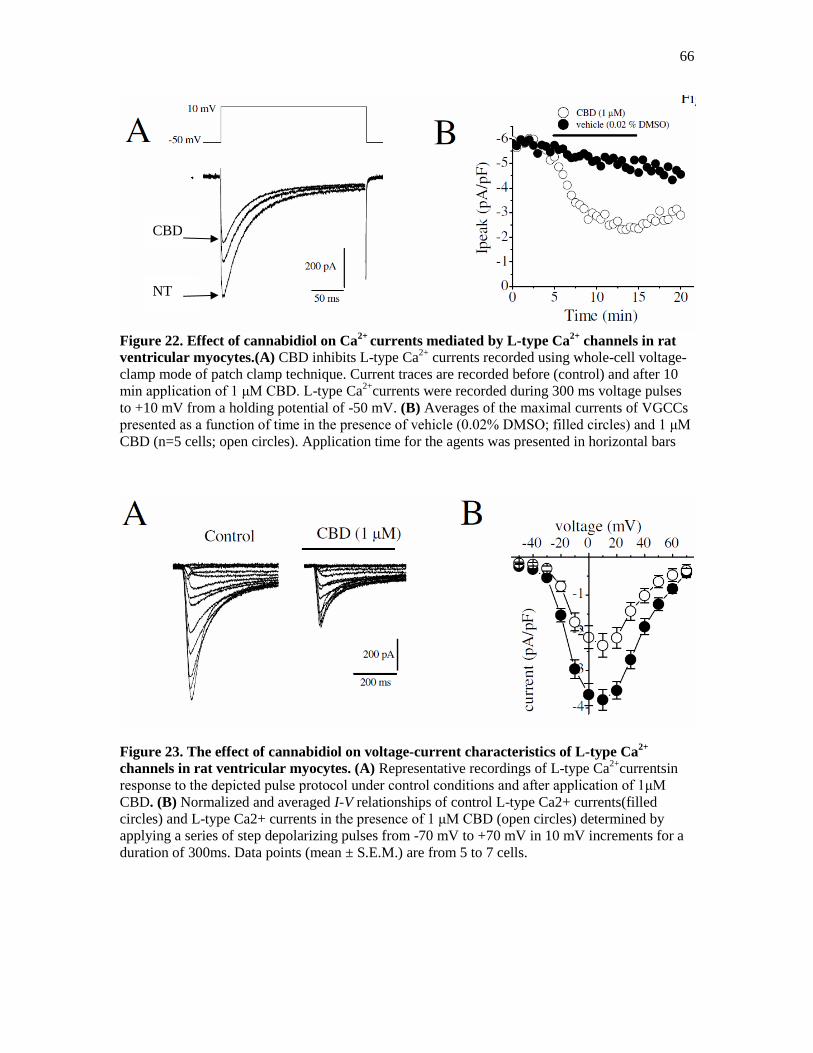
66
Figure 22. Effect of cannabidiol on Ca2+
currents mediated by L-type Ca2+
channels in rat
ventricular myocytes.(A) CBD inhibits L-type Ca2+
currents recorded using whole-cell voltage-
clamp mode of patch clamp technique. Current traces are recorded before (control) and after 10
min application of 1 μM CBD. L-type Ca2+
currents were recorded during 300 ms voltage pulses
to +10 mV from a holding potential of -50 mV. (B) Averages of the maximal currents of VGCCs
presented as a function of time in the presence of vehicle (0.02% DMSO; filled circles) and 1 μM
CBD (n=5 cells; open circles). Application time for the agents was presented in horizontal bars
Figure 23. The effect of cannabidiol on voltage-current characteristics of L-type Ca2+
channels in rat ventricular myocytes. (A) Representative recordings of L-type Ca2+
currentsin
response to the depicted pulse protocol under control conditions and after application of 1μM
CBD. (B) Normalized and averaged I-V relationships of control L-type Ca2+ currents(filled
circles) and L-type Ca2+ currents in the presence of 1 μM CBD (open circles) determined by
applying a series of step depolarizing pulses from -70 mV to +70 mV in 10 mV increments for a
duration of 300ms. Data points (mean ± S.E.M.) are from 5 to 7 cells.
NT
CBD

67
CBD did not change the steady-state activation curve of the L-type Ca2+
current
(V1/2 values in the control experiment and in the presence of CBD, they are -19.1 ± 0.4
mV and -17.6 ± 0.3 mV, respectively; n=6, Paired t-test, P>0.05) (Figure 24A). However,
there was a hyperpolarizing shift of L-type Ca2+
currents in steady-state inactivation by
9.6 mV (i.e., from control value V1/2= -10.2 ± 0.9 mV to V1/2= -19.8 ± 1.1 mV in the
presence of CBD) (Figure 24B). However, there was little effect on the slopes of the
curves (k = 7.7 ± 0.5 mV and k = -4.5 ± 0.4 mV for the control activation and
inactivation, respectively, versus k = 7.9 ± 0.3 mV and k = -4.2 ± 0.3 mV for the CBD-
modified activation and inactivation, correspondingly).
In line with earlier reports (Soldatov, et al., 1998), kinetic analysis of L-type
Ca2+
currents were titled to a double-exponential function with fast (τf) and slow (τs)
inactivation time constants. Comparison of L-type Ca2+
currents in the absence and
presence of CBD revealed noticeable acceleration of the current's inactivation kinetics by
CBD. Quantification of the time constants of L-type Ca2+
currents inactivation showed
that CBD (1 μM) significantly reduced τi in the range of Vm -30 mV and +10 mV (Figure
25).
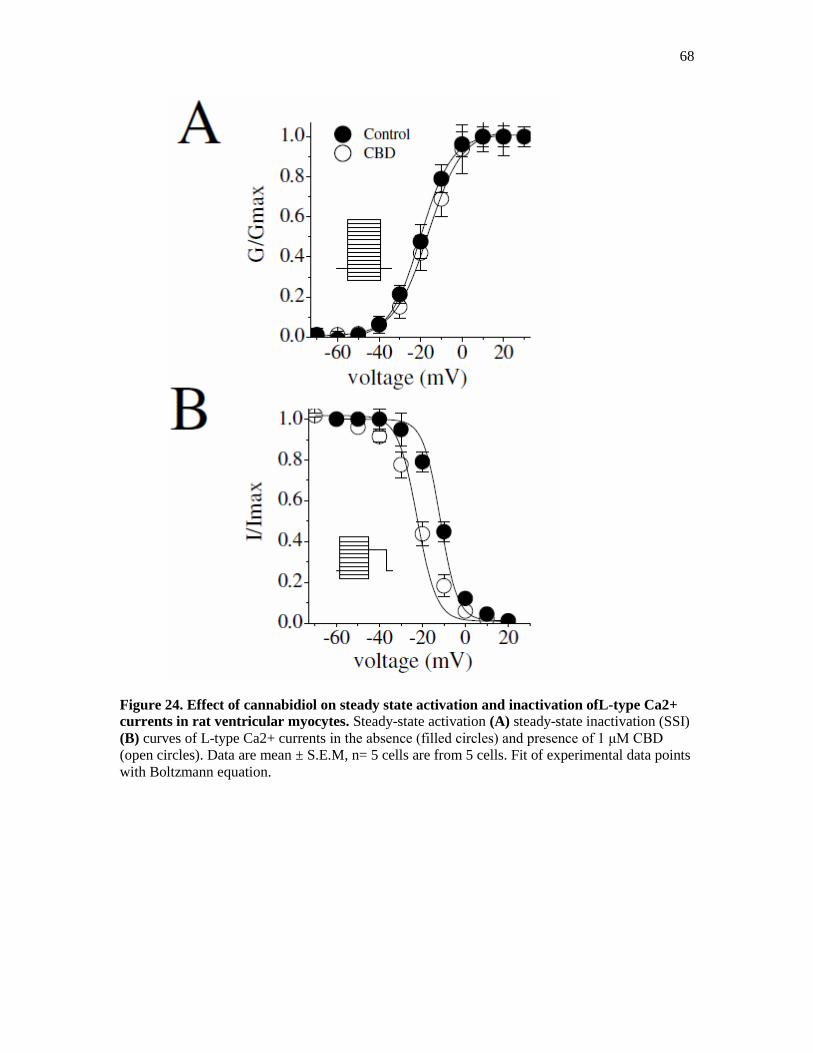
68
Figure 24. Effect of cannabidiol on steady state activation and inactivation ofL-type Ca2+
currents in rat ventricular myocytes. Steady-state activation (A) steady-state inactivation (SSI)
(B) curves of L-type Ca2+ currents in the absence (filled circles) and presence of 1 μM CBD
(open circles). Data are mean ± S.E.M, n= 5 cells are from 5 cells. Fit of experimental data points
with Boltzmann equation.

69
Figure25. Effect of cannabidiol on the kinetics ofL-type Ca2+currents in rat ventricular
myocytes. Voltage-dependent fast (triangles) and slow (circles) inactivation time constants (τi) of
L-type Ca2+ currents under control conditions (filled circles, and triangles) and in the presence of
1 μM CBD (open circles and triangles). Data are mean ± S.E.M., n= 5-6 cells.

70
Chapter 4: Discussion
Our results show that CBD has a negative inotropic effect on the contractility of rat
ventricular myocytes. The negative inotropic effect of CBD appears to be mainly due to
impaired Ca2+
signaling in cardiomyocytes. Furthermore, the results indicate that the
impaired Ca2+
signaling emanates from the inhibitory effects of CBD on the function of L
type Ca2+
channel.
Actions of CBD on the cardiovascular system influences multiple organ systems
and involves complex cellular and molecular mechanisms (see Baranowska-Kuczko,
MacLean, Kozlowska and Malinowska, 2012; Stanley et al., 2013). CBD has been shown
to have various effects on muscular structures, neuronal and endothelial cells and alter the
activities of several receptors and ion channels directly (see Hejazi et al., 2006; Pertwee
2008; 2010). Furthermore, other factors such as the activation of autonomic reflexes, the
presence of endothelial cells and fatty-acid based metabolic products have been reported
to contribute to the complexity of CBD actions on the heart (Pertwee 2008; Stanley et al.,
2013). On the other hand, acutely dissociated ventricular myocytes have several
advantages over in-vivo and traditional in-vitro conditions, since it excludes some of the
factors mentioned above and also the influences of reflex pathways, autonomic nerve
endings, neurotransmitter uptake systems, gap-junction connections and coronary
perfusion status.
In our experiments, using video edge detection techniques in individual myocytes,
it was found that CBD causes a significant reduction in the maximal amplitude of
shortening (expressed as a percentage of resting cell length) without altering the time
frame (such as TPK shortening and THALF relaxation) of myocytes shortening. Our

71
findings from myocyte shortening and intracellular Ca2+
measurement experiments are in
agreement with earlier studies reporting that CBD exerts a negative-inotropic effect in
isolated-cardiac muscle preparations (Smiley, Karler and Turkanis, 1976; Nahas and
Trouve, 1985). Furthermore, results from our study suggest that a direct interaction of
CBD with ventricular myocytes, rather than the actions of CBD on nerve endings and
neurotransmitter uptake systems are mainly responsible for the negative inotropic effects
of CBD in the heart.
The negative-inotropic actions of CBD might be attributed to the impaired release
of Ca2+
from the SR. In fact, CBD and other cannabinoids have been reported to
modulate the ryanodine sensitive intracellular Ca2+
stores in neurons (Drsydale et al.,
2008) glia (Mato et al., 2010; Pertwee et al., 2010). However, the amplitude and kinetics
of caffeine-induced Ca2+
release from intracellular Ca2+
stores were not changed by CBD
suggesting that ryanodine-sensitive intracellular Ca2+
stores are not involved in the
negative-inotropic effects of CBD observed in this study.
Cannabidiol can alter the levels of second messengers such as cAMP, cGMP and
protein kinase C which are known to be involved in tuning the Ca2+
sensitivity of the
contractile proteins. In an earlier study of cardiac muscle membranes, cAMP levels have
been shown to be unaltered in the presence of CBD (Hillard, Pounds, Boyer and Bloom,
1990), Furthermore, the sensitivity of contractile proteins to intracellular Ca2+
remained
unchanged in the presence of CBD suggesting that phosphorylation and de-
phosphorylation of the contractile proteins did not play a significant role in the negative-
inotropic action of CBD. Collectively, these results suggest that the effects of CBD on
myocyte contractility are not related to changes in intracellular Ca2+
release machinery or

72
sensitivity of myofilaments to Ca2+
. In addition, in the presence of CBD, resting levels of
intracellular Ca2+
, and cell length of ventricular myocytes, remained unaltered suggesting
that CBD does not significantly affect Ca2+
homeostasis under resting conditions.
During excitation-contraction coupling, alterations in the amplitudes and kinetics
of the cardiac action potential (AP) are closely associated with corresponding changes in
the contractility of myocytes. In our study, cardiac APs were recorded using the whole
cell patch clamp technique. It was found that the amplitudes and dV/dtmax of APs were
not altered in the presence of CBD suggesting that voltage-gated sodium channels are not
affected by CBD. For this reason, the effect of CBD on the functional properties of
voltage-gated sodium channels was not investigated any further. However, the decrease
of AP duration, and suppression of the second phase of the AP was evident in whole cell
recordings from cardiomyocytes. Since the plateau (phase 2) of AP is mainly due the
influx of Ca2+
through L-type voltage-gated Ca2+
channels, these results suggest that
CBD can act on L-type Ca2+
channels. Indeed, whole cell recordings of Ca2+
currents in
cardiomyocytes, in agreement with the recordings of APs, indicated that CBD inhibits
potently the function of L-type voltage-gated Ca2+
channels with an IC50 value of 0.6 µM.
An analysis of current kinetics indicated that the inactivation of Ca2+
currents can double
the exponential decay and CBD can decrease the amplitudes of the Ca2+
currents by
accelerating both fast and slow components of the inactivation process. The effects of
CBD on the gating characteristics of Ca2+
channels were further investigated by studying
the activation and inactivation curves in the absence and presence CBD. Although half
activation voltage and the slope of the activation curve were not altered, there was a

73
hyperpolarizing-shift of the inactivation curve further suggesting that CBD changes the
voltage-dependent inactivation process.
Collectively, these results suggest that during excitation-contraction coupling,
shortening of action potential due to the inhibition of L-type Ca2+
channels decreases
Ca2+
-induced Ca2+
release from SR and causes the negative-inotropic effect of CBD
observed in our experiments and reported in earlier studies. In line with these findings,
although caffeine induced contractures and myofilament sensitivity to Ca2+
remained
unchanged, electrically-induced Ca2+
transients were significantly depressed by CBD;
further suggesting that Ca2+
-induced Ca2+
release was impaired in the presence of CBD.
In earlier studies CBD has been reported to inhibit the uptake of dopamine and other
neurotransmitters (Poddar and Dewey, 1980; Pandolfo, et al., 2011). However, an uptake
mechanism is not likely to be involved in our studies on isolated ventricular myocytes.
Furthermore, inhibition of uptake mechanisms for dopamine and/or catecholamines such
as epinephrine and norepinephrine would lead to well-established positive inotropic
actions, rather than a negative inotropic effect as observed in the present study.
Although CBD does not activate known cannabinoid receptors, some of its
pharmacological actions have been shown to be reversed by cannabinoid receptor
antagonists (Pertwee, 2008). Recent studies have shown that the antagonist sensitivity of
CBD effects is due to the inhibition of fatty acid amide hydrolase activity, an enzyme that
inactivates endogenous cannabinoids and related to subsequent increases in tissue
endocannabinoids levels (Pertwee, 2010). However, both amplitudes of myocyte
shortening and the L-type Ca2+
currents were not altered by URB597, a potent inhibitor

74
of fatty acid amide hydrolase (Al Kury et al., 2014), suggesting that the activity of
increased fatty acid amide hydrolase was not involved in the actions observed in CBD.
The interaction between L-type Ca2+
channels and CBD has not been studied in
detail. In an earlier study, 45
Ca2+
-uptake in brain synaptosomes had been shown to be
inhibited by CBD (Harris and Stokes, 1982). Similarly, CBD induced increases in
intracellular Ca2+
levels were found to be sensitive to blockers of L-type Ca2+
channels
(Drysdale et al., 2006). In agreement with these results, contractions of smooth muscles
elicited by high-K+-containing solutions were also significantly inhibited by CBD
(Cluny, Naylor, Whittle and Javid, 2011). The interaction between T-type Ca2+
channels
and CBD was investigated in a recent study, on human CaV3 channels stably expressed
in human embryonic kidney 293 cells and T-type channels in mouse sensory neurons
(Ross et al., 2008). At moderately hyperpolarized potentials, CBD inhibited peak CaV3.1
and CaV3.2 currents with IC50 values of 1 µM but was less potent on CaV3.3 channels.
CBD inhibited sensory neuron T-type channels by about 45% at 1 µM. However, in
recordings made from a holding potential of -70 mV, 100 nM CBD inhibited more than
50% of the peak CaV3.1 current. CBD produced a significant hyperpolarizing shift in the
steady state inactivation potentials for each of the CaV3 channels (Ross et al., 2008). In
this respect our results indicate for the first time that CBD inhibits the function of L-type
Ca2+
channels in ventricular myocytes.
In clinical studies, it has been shown that acute CBD intake does not cause a
significant change in blood pressure and heart rate (Stanley, 2013a). However, several
earlier studies indicate that increases in blood pressure and heart rate during stressful
conditions are markedly attenuated by CBD (Gomes, et al., 2013; Granjeiro, Gomes,

75
Guimaraes, Correa and Resstel, 2011; Resstel, Joca, Moreira, Correa and Guimaraes,
2006; Resstel, et al., 2009). In fact, in in vivo studies, it has been demonstrated that
increased heart rate and blood pressure, one of the most consistent effects of cannabis
intoxication, are significantly decreased by CBD (Nahas and Trouve, 1985). These
studies suggest that CBD can exert negative inotropic and chronotropic actions under
certain clinical conditions such as stress and cannabis intoxication.
CBD, in the concentration range used in this study (0.1-10 µM), has been shown
to modulate functions of various receptors and ion channels such as Ca2+
channels (Ross
et al., 2008), 5-HT3 (Yang et al., 2010), and nicotinic receptors (Mahgoub et al., 2013).
The peak plasma concentrations of CBD in healthy volunteers following administration
of SativexTM
(1:1 ratio of ∆9-THC and CBD) was reported to be between 0.01µM to
0.05µM (Massi, et al., 2013). In earlier clinical studies, mean CBD levels of 0.036 μM
were determined in blood analysis following a 6-week oral treatment with CBD at doses
of 10 mg/kg/day (Consroe et al., 1991). However, CBD is a highly lipophilic compound
and easily passes biological membranes and accumulates in tissues and it can reach to
several fold higher tissue concentrations than in blood. Collectively, these findings
suggest that CBD, at relatively high concentrations, can suppress the function of the
cardiovascular system. In fact, cardiac failure due to depressed heart contractility has
been suggested to be the main cause of mortality through cannabis intoxication
(Bergamaschi et al., 2011). Thus it is likely that some of the effects of the cannabis plant
are mediated by CBD during cannabis intoxication.

76
In addition to cardiac contractility, the electrical activity of heart has also been
suggested to be affected by CBD. In an earlier study, CBD has been shown to have
beneficial actions in ischemia-induced cardiac arrhythmias (Walsh et al., 2010). The
shortening of the AP duration by CBD observed in our study can be beneficial or
harmful, depending on the underlying pathology. Thus, during acute ischemia, in which
the duration of the cardiac APD is already shortened, a further decrease should be
proarrhythmic (Den Ruijter et al., 2007). However, shortening of AP duration should be
beneficial in preventing those arrhythmias caused by triggered activities observed in
conditions such as heart failure (Den Ruijter et al., 2007). In conclusion, the results
indicate for the first time, that CBD inhibits myocyte contractility by acting on L-type
Ca2+
channels.

77
Chapter 5: Limitation and Future Directions
The scope of this research is mainly concerned with the effects of CBD on healthy
ventricular cells, although endocannabinoids are stress induced neurotransmitters. In
addition to the effects of CBD on L- type calcium channels its effect on NCX should also
be established due to their influence on cytosolic calcium levels and the viability of
ventricular cells in the presence of CBD.
Future research may also include expressing calcium channels onto HEK-293 cells,
this allows us to exclude or include the indirect effects of CBD on intracellular signaling
that can affect calcium channels. Another method is to express the calcium channel each
subunit at a time and find where CBD exerts it effects most effectively and thereby
establish the mechanism by which CBD exerts there effects.

78
Bibliography
Ahrens, J., Demir, R., Leuwer, M., de la Roche, J., Krampfl, K., Foadi, N., . . . Haeseler, G.
(2009). The nonpsychotropic cannabinoid cannabidiol modulates and directly activates
alpha-1 and alpha-1-Beta glycine receptor function. Pharmacology, 83(4), 217-222. doi:
10.1159/000201556
Amin, A. S., Tan, H. L., & Wilde, A. A. (2010). Cardiac ion channels in health and disease.
Heart Rhythm, 7(1), 117-126. doi: 10.1016/j.hrthm.2009.08.005
Ashton, J. C. (2012). Synthetic cannabinoids as drugs of abuse. Curr Drug Abuse Rev, 5(2), 158-
168.
Baranowska-Kuczko, M., MacLean, M. R., Kozlowska, H., & Malinowska, B. (2012).
Endothelium-dependent mechanisms of the vasodilatory effect of the endocannabinoid,
anandamide, in the rat pulmonary artery. Pharmacol Res, 66(3), 251-259. doi:
10.1016/j.phrs.2012.05.004
Bergamaschi, M. M., Queiroz, R. H., Zuardi, A. W., & Crippa, J. A. (2011). Safety and side
effects of cannabidiol, a Cannabis sativa constituent. Curr Drug Saf, 6(4), 237-249.
Bers, D. M. (2000). Calcium fluxes involved in control of cardiac myocyte contraction. Circ Res,
87(4), 275-281.
Bers, D. M. (2008). Calcium cycling and signaling in cardiac myocytes. Annu Rev Physiol, 70,
23-49. doi: 10.1146/annurev.physiol.70.113006.100455
Bishop-Bailey, D. (2000). Peroxisome proliferator-activated receptors in the cardiovascular
system. Br J Pharmacol, 129(5), 823-834. doi: 10.1038/sj.bjp.0703149
Bisogno, T., Hanus, L., De Petrocellis, L., Tchilibon, S., Ponde, D. E., Brandi, I., . . . Di Marzo,
V. (2001). Molecular targets for cannabidiol and its synthetic analogues: effect on
vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of
anandamide. Br J Pharmacol, 134(4), 845-852. doi: 10.1038/sj.bjp.0704327
Blake, D. R., Robson, P., Ho, M., Jubb, R. W., & McCabe, C. S. (2006). Preliminary assessment
of the efficacy, tolerability and safety of a cannabis-based medicine (Sativex) in the
treatment of pain caused by rheumatoid arthritis. Rheumatology (Oxford), 45(1), 50-52.
doi: 10.1093/rheumatology/kei183
Booz, G. W. (2011). Cannabidiol as an emergent therapeutic strategy for lessening the impact of
inflammation on oxidative stress. Free Radic Biol Med, 51(5), 1054-1061. doi:
10.1016/j.freeradbiomed.2011.01.007

79
Braida, D., Pegorini, S., Arcidiacono, M. V., Consalez, G. G., Croci, L., & Sala, M. (2003). Post-
ischemic treatment with cannabidiol prevents electroencephalographic flattening,
hyperlocomotion and neuronal injury in gerbils. Neurosci Lett, 346(1-2), 61-64.
Burstein, S. H., & Zurier, R. B. (2009). Cannabinoids, endocannabinoids, and related analogs in
inflammation. Aaps j, 11(1), 109-119. doi: 10.1208/s12248-009-9084-5
Campos, A. C., Moreira, F. A., Gomes, F. V., Del Bel, E. A., & Guimaraes, F. S. (2012).
Multiple mechanisms involved in the large-spectrum therapeutic potential of cannabidiol
in psychiatric disorders. Philos Trans R Soc Lond B Biol Sci, 367(1607), 3364-3378. doi:
10.1098/rstb.2011.0389
Capasso, R., Borrelli, F., Aviello, G., Romano, B., Scalisi, C., Capasso, F., & Izzo, A. A. (2008).
Cannabidiol, extracted from Cannabis sativa, selectively inhibits inflammatory
hypermotility in mice. Br J Pharmacol, 154(5), 1001-1008. doi: 10.1038/bjp.2008.177
Carrier, E. J., Auchampach, J. A., & Hillard, C. J. (2006). Inhibition of an equilibrative
nucleoside transporter by cannabidiol: a mechanism of cannabinoid immunosuppression.
Proc Natl Acad Sci U S A, 103(20), 7895-7900. doi: 10.1073/pnas.0511232103
Catterall, W. A. (2010). Signaling complexes of voltage-gated sodium and calcium channels.
Neurosci Lett, 486(2), 107-116. doi: 10.1016/j.neulet.2010.08.085
Catterall, W. A. (2011). Voltage-gated calcium channels. Cold Spring Harb Perspect Biol, 3(8),
a003947. doi: 10.1101/cshperspect.a003947
Catterall, W. A., Perez-Reyes, E., Snutch, T. P., & Striessnig, J. (2005). International Union of
Pharmacology. XLVIII. Nomenclature and structure-function relationships of voltage-
gated calcium channels. Pharmacol Rev, 57(4), 411-425. doi: 10.1124/pr.57.4.5
Cluny, N. L., Naylor, R. J., Whittle, B. A., & Javid, F. A. (2011). The effects of cannabidiolic
acid and cannabidiol on contractility of the gastrointestinal tract of Suncus murinus. Arch
Pharm Res, 34(9), 1509-1517. doi: 10.1007/s12272-011-0913-6
Collins, F. G., & Haavik, C. O. (1979). Effects of cannabinoids on cardiac microsomal CaATPase
activity and calcium uptake. Biochem Pharmacol, 28(15), 2303-2306.
Costa, B., Colleoni, M., Conti, S., Parolaro, D., Franke, C., Trovato, A. E., & Giagnoni, G.
(2004). Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of
cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn
Schmiedebergs Arch Pharmacol, 369(3), 294-299. doi: 10.1007/s00210-004-0871-3
Costa, B., Trovato, A. E., Comelli, F., Giagnoni, G., & Colleoni, M. (2007). The non-
psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in
rat chronic inflammatory and neuropathic pain. Eur J Pharmacol, 556(1-3), 75-83. doi:
10.1016/j.ejphar.2006.11.006

80
Cravatt, B. F., & Lichtman, A. H. (2002). The enzymatic inactivation of the fatty acid amide class
of signaling lipids. Chem Phys Lipids, 121(1-2), 135-148.
De Jongh, K. S., Murphy, B. J., Colvin, A. A., Hell, J. W., Takahashi, M., & Catterall, W. A.
(1996). Specific phosphorylation of a site in the full-length form of the alpha 1 subunit of
the cardiac L-type calcium channel by adenosine 3',5'-cyclic monophosphate-dependent
protein kinase. Biochemistry, 35(32), 10392-10402. doi: 10.1021/bi953023c
De Petrocellis, L., Ligresti, A., Moriello, A. S., Allara, M., Bisogno, T., Petrosino, S., . . . Di
Marzo, V. (2011). Effects of cannabinoids and cannabinoid-enriched Cannabis extracts
on TRP channels and endocannabinoid metabolic enzymes. Br J Pharmacol, 163(7),
1479-1494. doi: 10.1111/j.1476-5381.2010.01166.x
De Petrocellis, L., Vellani, V., Schiano-Moriello, A., Marini, P., Magherini, P. C., Orlando, P., &
Di Marzo, V. (2008). Plant-derived cannabinoids modulate the activity of transient
receptor potential channels of ankyrin type-1 and melastatin type-8. J Pharmacol Exp
Ther, 325(3), 1007-1015. doi: 10.1124/jpet.107.134809
Den Ruijter, H. M., Berecki, G., Opthof, T., Verkerk, A. O., Zock, P. L., & Coronel, R. (2007).
Pro- and antiarrhythmic properties of a diet rich in fish oil. Cardiovasc Res, 73(2), 316-
325. doi: 10.1016/j.cardiores.2006.06.014
Devane, W. A., Dysarz, F. A., 3rd, Johnson, M. R., Melvin, L. S., & Howlett, A. C. (1988).
Determination and characterization of a cannabinoid receptor in rat brain. Mol
Pharmacol, 34(5), 605-613.
Di Marzo, V. (2008). Endocannabinoids: synthesis and degradation. Rev Physiol Biochem
Pharmacol, 160, 1-24. doi: 10.1007/112_0505
Di Marzo, V., Fontana, A., Cadas, H., Schinelli, S., Cimino, G., Schwartz, J. C., & Piomelli, D.
(1994). Formation and inactivation of endogenous cannabinoid anandamide in central
neurons. Nature, 372(6507), 686-691. doi: 10.1038/372686a0
Dolphin, A. C. (2003). G protein modulation of voltage-gated calcium channels. Pharmacol Rev,
55(4), 607-627. doi: 10.1124/pr.55.4.3
Dolphin, A. C. (2006). A short history of voltage-gated calcium channels. Br J Pharmacol, 147
Suppl 1, S56-62. doi: 10.1038/sj.bjp.0706442
Dolphin, A. C. (2009). Calcium channel diversity: multiple roles of calcium channel subunits.
Curr Opin Neurobiol, 19(3), 237-244. doi: 10.1016/j.conb.2009.06.006
Drysdale, A. J., Ryan, D., Pertwee, R. G., & Platt, B. (2006). Cannabidiol-induced intracellular
Ca2+ elevations in hippocampal cells. Neuropharmacology, 50(5), 621-631. doi:
10.1016/j.neuropharm.2005.11.008

81
El-Remessy, A. B., Al-Shabrawey, M., Khalifa, Y., Tsai, N. T., Caldwell, R. B., & Liou, G. I.
(2006). Neuroprotective and blood-retinal barrier-preserving effects of cannabidiol in
experimental diabetes. Am J Pathol, 168(1), 235-244. doi: 10.2353/ajpath.2006.050500
El-Remessy, A. B., Khalil, I. E., Matragoon, S., Abou-Mohamed, G., Tsai, N. J., Roon, P., . . .
Liou, G. I. (2003). Neuroprotective effect of (-)Delta9-tetrahydrocannabinol and
cannabidiol in N-methyl-D-aspartate-induced retinal neurotoxicity: involvement of
peroxynitrite. Am J Pathol, 163(5), 1997-2008.
Fearnley, C. J., Roderick, H. L., & Bootman, M. D. (2011). Calcium signaling in cardiac
myocytes. Cold Spring Harb Perspect Biol, 3(11), a004242. doi:
10.1101/cshperspect.a004242
Fernandez-Ruiz, J. (2012). [Cannabinoid drugs for neurological diseases: what is behind?]. Rev
Neurol, 54(10), 613-628.
Ferrantini, C., Crocini, C., Coppini, R., Vanzi, F., Tesi, C., Cerbai, E., . . . Sacconi, L. (2013). The
transverse-axial tubular system of cardiomyocytes. Cell Mol Life Sci, 70(24), 4695-4710.
doi: 10.1007/s00018-013-1410-5
Foadi, N., Leuwer, M., Demir, R., Dengler, R., Buchholz, V., de la Roche, J., . . . Ahrens, J.
(2010). Lack of positive allosteric modulation of mutated alpha(1)S267I glycine
receptors by cannabinoids. Naunyn Schmiedebergs Arch Pharmacol, 381(5), 477-482.
doi: 10.1007/s00210-010-0506-9
Fowler, C. J. (2004). Oleamide: a member of the endocannabinoid family? Br J Pharmacol,
141(2), 195-196. doi: 10.1038/sj.bjp.0705608
Garcia-Arencibia, M., Gonzalez, S., de Lago, E., Ramos, J. A., Mechoulam, R., & Fernandez-
Ruiz, J. (2007). Evaluation of the neuroprotective effect of cannabinoids in a rat model of
Parkinson's disease: importance of antioxidant and cannabinoid receptor-independent
properties. Brain Res, 1134(1), 162-170. doi: 10.1016/j.brainres.2006.11.063
Gilbert, J. C., Pertwee, R. G., & Wyllie, M. G. (1977). Effects of delta9-tetrahydrocannabinol and
cannabidiol on a Mg2+-ATPase of synaptic vesicles prepared from rat cerebral cortex. Br
J Pharmacol, 59(4), 599-601.
Glaser, S. T., Kaczocha, M., & Deutsch, D. G. (2005). Anandamide transport: a critical review.
Life Sci, 77(14), 1584-1604. doi: 10.1016/j.lfs.2005.05.007
Gloss, D., Nolan, S. J., & Staba, R. (2014). The role of high-frequency oscillations in epilepsy
surgery planning. Cochrane Database Syst Rev, 1, Cd010235. doi:
10.1002/14651858.CD010235.pub2
Gomes, F. V., Alves, F. H., Guimaraes, F. S., Correa, F. M., Resstel, L. B., & Crestani, C. C.
(2013). Cannabidiol administration into the bed nucleus of the stria terminalis alters

82
cardiovascular responses induced by acute restraint stress through 5-HT(1)A receptor.
Eur Neuropsychopharmacol, 23(9), 1096-1104. doi: 10.1016/j.euroneuro.2012.09.007
Gomes, F. V., Resstel, L. B., & Guimaraes, F. S. (2011). The anxiolytic-like effects of
cannabidiol injected into the bed nucleus of the stria terminalis are mediated by 5-HT1A
receptors. Psychopharmacology (Berl), 213(2-3), 465-473. doi: 10.1007/s00213-010-
2036-z
Granjeiro, E. M., Gomes, F. V., Guimaraes, F. S., Correa, F. M., & Resstel, L. B. (2011). Effects
of intracisternal administration of cannabidiol on the cardiovascular and behavioral
responses to acute restraint stress. Pharmacol Biochem Behav, 99(4), 743-748. doi:
10.1016/j.pbb.2011.06.027
Grlic, Ljubiša. "United Nations Office on Drugs and Crime." UNODC. N.p., 1 Jan. 1962. Web.
01 May 2014.
Haase, H., Bartel, S., Karczewski, P., Morano, I., & Krause, E. G. (1996). In-vivo
phosphorylation of the cardiac L-type calcium channel beta-subunit in response to
catecholamines. Mol Cell Biochem, 163-164, 99-106.
Hamdani, N., Kooij, V., van Dijk, S., Merkus, D., Paulus, W. J., Remedios, C. D., . . . van der
Velden, J. (2008). Sarcomeric dysfunction in heart failure. Cardiovasc Res, 77(4), 649-
658. doi: 10.1093/cvr/cvm079
Hampson, A. J., Grimaldi, M., Axelrod, J., & Wink, D. (1998). Cannabidiol and (-)Delta9-
tetrahydrocannabinol are neuroprotective antioxidants. Proc Natl Acad Sci U S A, 95(14),
8268-8273.
Hampson, A. J., Grimaldi, M., Lolic, M., Wink, D., Rosenthal, R., & Axelrod, J. (2000).
Neuroprotective antioxidants from marijuana. Ann N Y Acad Sci, 899, 274-282.
Hanus, L., Abu-Lafi, S., Fride, E., Breuer, A., Vogel, Z., Shalev, D. E., . . . Mechoulam, R.
(2001). 2-arachidonyl glyceryl ether, an endogenous agonist of the cannabinoid CB1
receptor. Proc Natl Acad Sci U S A, 98(7), 3662-3665. doi: 10.1073/pnas.061029898
Hayakawa, K., Mishima, K., Abe, K., Hasebe, N., Takamatsu, F., Yasuda, H., . . . Fujiwara, M.
(2004). Cannabidiol prevents infarction via the non-CB1 cannabinoid receptor
mechanism. Neuroreport, 15(15), 2381-2385.
Heinemann, S. H., Terlau, H., Stuhmer, W., Imoto, K., & Numa, S. (1992). Calcium channel
characteristics conferred on the sodium channel by single mutations. Nature, 356(6368),
441-443. doi: 10.1038/356441a0
Hejazi, N., Zhou, C., Oz, M., Sun, H., Ye, J. H., & Zhang, L. (2006). Delta9-
tetrahydrocannabinol and endogenous cannabinoid anandamide directly potentiate the

83
function of glycine receptors. Mol Pharmacol, 69(3), 991-997. doi:
10.1124/mol.105.019174
Hell, J. W., Yokoyama, C. T., Wong, S. T., Warner, C., Snutch, T. P., & Catterall, W. A. (1993).
Differential phosphorylation of two size forms of the neuronal class C L-type calcium
channel alpha 1 subunit. J Biol Chem, 268(26), 19451-19457.
Herkenham, M., Lynn, A. B., Little, M. D., Johnson, M. R., Melvin, L. S., de Costa, B. R., &
Rice, K. C. (1990). Cannabinoid receptor localization in brain. Proc Natl Acad Sci U S A,
87(5), 1932-1936.
Hill, A. J., Williams, C. M., Whalley, B. J., & Stephens, G. J. (2012). Phytocannabinoids as novel
therapeutic agents in CNS disorders. Pharmacol Ther, 133(1), 79-97. doi:
10.1016/j.pharmthera.2011.09.002
Hillard, C. J., & Auchampach, J. A. (1994). In vitro activation of brain protein kinase C by the
cannabinoids. Biochim Biophys Acta, 1220(2), 163-170.
Hillard, C. J., Pounds, J. J., Boyer, D. R., & Bloom, A. S. (1990). Studies of the role of membrane
lipid order in the effects of delta 9-tetrahydrocannabinol on adenylate cyclase activation
in heart. J Pharmacol Exp Ther, 252(3), 1075-1082.
Howarth, F. C., Qureshi, M. A., Hassan, Z., Al Kury, L. T., Isaev, D., Parekh, K., . . . Adeghate,
E. (2011). Changing pattern of gene expression is associated with ventricular myocyte
dysfunction and altered mechanisms of Ca2+ signalling in young type 2 Zucker diabetic
fatty rat heart. Exp Physiol, 96(3), 325-337. doi: 10.1113/expphysiol.2010.055574
Howarth, F. C., Qureshi, M. A., & White, E. (2002). Effects of hyperosmotic shrinking on
ventricular myocyte shortening and intracellular Ca(2+) in streptozotocin-induced
diabetic rats. Pflugers Arch, 444(3), 446-451. doi: 10.1007/s00424-002-0830-0
Howlett, A. C., Scott, D. K., & Wilken, G. H. (1989). Regulation of adenylate cyclase by
cannabinoid drugs. Insights based on thermodynamic studies. Biochem Pharmacol,
38(19), 3297-3304.
Hsueh, W. A., & Bruemmer, D. (2004). Peroxisome proliferator-activated receptor gamma:
implications for cardiovascular disease. Hypertension, 43(2), 297-305. doi:
10.1161/01.HYP.0000113626.76571.5b
Huffman, J. W., Liddle, J., Duncan, S. G., Jr., Yu, S., Martin, B. R., & Wiley, J. L. (1998).
Synthesis and pharmacology of the isomeric methylheptyl-delta8-tetrahydrocannabinols.
Bioorg Med Chem, 6(12), 2383-2396.
Iuvone, T., Esposito, G., Esposito, R., Santamaria, R., Di Rosa, M., & Izzo, A. A. (2004).
Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis

84
sativa, on beta-amyloid-induced toxicity in PC12 cells. J Neurochem, 89(1), 134-141.
doi: 10.1111/j.1471-4159.2003.02327.x
Izzo, A. A., & Camilleri, M. (2009). Cannabinoids in intestinal inflammation and cancer.
Pharmacol Res, 60(2), 117-125. doi: 10.1016/j.phrs.2009.03.008
Karniol, I. G., Shirakawa, I., Kasinski, N., Pfeferman, A., & Carlini, E. A. (1974). Cannabidiol
interferes with the effects of delta 9 - tetrahydrocannabinol in man. Eur J Pharmacol,
28(1), 172-177.
Kathmann, M., Flau, K., Redmer, A., Trankle, C., & Schlicker, E. (2006). Cannabidiol is an
allosteric modulator at mu- and delta-opioid receptors. Naunyn Schmiedebergs Arch
Pharmacol, 372(5), 354-361. doi: 10.1007/s00210-006-0033-x
Lanner, J. T., Georgiou, D. K., Joshi, A. D., & Hamilton, S. L. (2010). Ryanodine receptors:
structure, expression, molecular details, and function in calcium release. Cold Spring
Harb Perspect Biol, 2(11), a003996. doi: 10.1101/cshperspect.a003996
Lastres-Becker, I., Molina-Holgado, F., Ramos, J. A., Mechoulam, R., & Fernandez-Ruiz, J.
(2005). Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in
vivo and in vitro: relevance to Parkinson's disease. Neurobiol Dis, 19(1-2), 96-107. doi:
10.1016/j.nbd.2004.11.009
Ma, Y., Zhang, L., Zhao, Y. F., Zang, W. J., & Chen, C. (2010). Effects of GH secretagogues on
contractility and Ca2+ homeostasis of isolated adult rat ventricular myocytes.
Endocrinology, 151(9), 4446-4454. doi: 10.1210/en.2009-1432
Mahgoub, M., Keun-Hang, S. Y., Sydorenko, V., Ashoor, A., Kabbani, N., Al Kury, L., . . . Oz,
M. (2013). Effects of cannabidiol on the function of alpha7-nicotinic acetylcholine
receptors. Eur J Pharmacol, 720(1-3), 310-319. doi: 10.1016/j.ejphar.2013.10.011
Marsicano, G., Moosmann, B., Hermann, H., Lutz, B., & Behl, C. (2002). Neuroprotective
properties of cannabinoids against oxidative stress: role of the cannabinoid receptor CB1.
J Neurochem, 80(3), 448-456.
Martin-Moreno, A. M., Reigada, D., Ramirez, B. G., Mechoulam, R., Innamorato, N., Cuadrado,
A., & de Ceballos, M. L. (2011). Cannabidiol and other cannabinoids reduce microglial
activation in vitro and in vivo: relevance to Alzheimer's disease. Mol Pharmacol, 79(6),
964-973. doi: 10.1124/mol.111.071290
Mato, S., Victoria Sanchez-Gomez, M., & Matute, C. (2010). Cannabidiol induces intracellular
calcium elevation and cytotoxicity in oligodendrocytes. Glia, 58(14), 1739-1747. doi:
10.1002/glia.21044

85
Mechoulam, R., Ben-Shabat, S., Hanus, L., Ligumsky, M., Kaminski, N. E., Schatz, A. R., . . . et
al. (1995). Identification of an endogenous 2-monoglyceride, present in canine gut, that
binds to cannabinoid receptors. Biochem Pharmacol, 50(1), 83-90.
Mechoulam, R., & Gaoni, Y. (1965). A TOTAL SYNTHESIS OF DL-DELTA-1-
TETRAHYDROCANNABINOL, THE ACTIVE CONSTITUENT OF HASHISH. J Am
Chem Soc, 87, 3273-3275.
Mechoulam, R., & Hanus, L. (2002). Cannabidiol: an overview of some chemical and
pharmacological aspects. Part I: chemical aspects. Chem Phys Lipids, 121(1-2), 35-43.
Mechoulam, R., Parker, L. A., & Gallily, R. (2002). Cannabidiol: an overview of some
pharmacological aspects. J Clin Pharmacol, 42(11 Suppl), 11s-19s.
Mechoulam, R., Peters, M., Murillo-Rodriguez, E., & Hanus, L. O. (2007). Cannabidiol--recent
advances. Chem Biodivers, 4(8), 1678-1692. doi: 10.1002/cbdv.200790147
Mishima, K., Hayakawa, K., Abe, K., Ikeda, T., Egashira, N., Iwasaki, K., & Fujiwara, M.
(2005). Cannabidiol prevents cerebral infarction via a serotonergic 5-
hydroxytryptamine1A receptor-dependent mechanism. Stroke, 36(5), 1077-1082. doi:
10.1161/01.str.0000163083.59201.34
Nahas, G., & Trouve, R. (1985). Effects and interactions of natural cannabinoids on the isolated
heart. Proc Soc Exp Biol Med, 180(2), 312-316.
Nattel, S., & Carlsson, L. (2006). Innovative approaches to anti-arrhythmic drug therapy. Nat Rev
Drug Discov, 5(12), 1034-1049. doi: 10.1038/nrd2112
Navedo, M. F., & Santana, L. F. (2013). CaV1.2 sparklets in heart and vascular smooth muscle. J
Mol Cell Cardiol, 58, 67-76. doi: 10.1016/j.yjmcc.2012.11.018
O'Sullivan, S. E., Kendall, D. A., & Randall, M. D. (2005). Vascular effects of delta 9-
tetrahydrocannabinol (THC), anandamide and N-arachidonoyldopamine (NADA) in the
rat isolated aorta. Eur J Pharmacol, 507(1-3), 211-221. doi: 10.1016/j.ejphar.2004.11.056
O'Sullivan, S. E., Sun, Y., Bennett, A. J., Randall, M. D., & Kendall, D. A. (2009). Time-
dependent vascular actions of cannabidiol in the rat aorta. Eur J Pharmacol, 612(1-3),
61-68. doi: 10.1016/j.ejphar.2009.03.010
Pandolfo, P., Silveirinha, V., dos Santos-Rodrigues, A., Venance, L., Ledent, C., Takahashi, R.
N., . . . Kofalvi, A. (2011). Cannabinoids inhibit the synaptic uptake of adenosine and
dopamine in the rat and mouse striatum. Eur J Pharmacol, 655(1-3), 38-45. doi:
10.1016/j.ejphar.2011.01.013

86
Parker, L. A., Rock, E. M., & Limebeer, C. L. (2011). Regulation of nausea and vomiting by
cannabinoids. Br J Pharmacol, 163(7), 1411-1422. doi: 10.1111/j.1476-
5381.2010.01176.x
Pertwee, R. G. (1997). Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol Ther,
74(2), 129-180.
Pertwee, R. G. (2008). The diverse CB1 and CB2 receptor pharmacology of three plant
cannabinoids: delta9-tetrahydrocannabinol, cannabidiol and delta9-
tetrahydrocannabivarin. Br J Pharmacol, 153(2), 199-215. doi: 10.1038/sj.bjp.0707442
Pertwee, R. G., Howlett, A. C., Abood, M. E., Alexander, S. P., Di Marzo, V., Elphick, M. R., . . .
Ross, R. A. (2010). International Union of Basic and Clinical Pharmacology. LXXIX.
Cannabinoid receptors and their ligands: beyond CB(1) and CB(2). Pharmacol Rev,
62(4), 588-631. doi: 10.1124/pr.110.003004
Pertwee, R. G., Ross, R. A., Craib, S. J., & Thomas, A. (2002). (-)-Cannabidiol antagonizes
cannabinoid receptor agonists and noradrenaline in the mouse vas deferens. Eur J
Pharmacol, 456(1-3), 99-106.
Poddar, M. K., & Dewey, W. L. (1980). Effects of cannabinoids on catecholamine uptake and
release in hypothalamic and striatal synaptosomes. J Pharmacol Exp Ther, 214(1), 63-67.
Porter, A. C., Sauer, J. M., Knierman, M. D., Becker, G. W., Berna, M. J., Bao, J., . . . Felder, C.
C. (2002). Characterization of a novel endocannabinoid, virodhamine, with antagonist
activity at the CB1 receptor. J Pharmacol Exp Ther, 301(3), 1020-1024.
Pragnell, M., De Waard, M., Mori, Y., Tanabe, T., Snutch, T. P., & Campbell, K. P. (1994).
Calcium channel beta-subunit binds to a conserved motif in the I-II cytoplasmic linker of
the alpha 1-subunit. Nature, 368(6466), 67-70. doi: 10.1038/368067a0
Puri, T. S., Gerhardstein, B. L., Zhao, X. L., Ladner, M. B., & Hosey, M. M. (1997). Differential
effects of subunit interactions on protein kinase A- and C-mediated phosphorylation of L-
type calcium channels. Biochemistry, 36(31), 9605-9615. doi: 10.1021/bi970500d
Qin, N., Neeper, M. P., Liu, Y., Hutchinson, T. L., Lubin, M. L., & Flores, C. M. (2008). TRPV2
is activated by cannabidiol and mediates CGRP release in cultured rat dorsal root
ganglion neurons. J Neurosci, 28(24), 6231-6238. doi: 10.1523/jneurosci.0504-08.2008
Rajesh, M., Mukhopadhyay, P., Batkai, S., Hasko, G., Liaudet, L., Drel, V. R., . . . Pacher, P.
(2007). Cannabidiol attenuates high glucose-induced endothelial cell inflammatory
response and barrier disruption. Am J Physiol Heart Circ Physiol, 293(1), H610-619. doi:
10.1152/ajpheart.00236.2007

87
Rao, G. K., & Kaminski, N. E. (2006). Cannabinoid-mediated elevation of intracellular calcium:
a structure-activity relationship. J Pharmacol Exp Ther, 317(2), 820-829. doi:
10.1124/jpet.105.100503
Resstel, L. B., Joca, S. R., Moreira, F. A., Correa, F. M., & Guimaraes, F. S. (2006). Effects of
cannabidiol and diazepam on behavioral and cardiovascular responses induced by
contextual conditioned fear in rats. Behav Brain Res, 172(2), 294-298. doi:
10.1016/j.bbr.2006.05.016
Resstel, L. B., Tavares, R. F., Lisboa, S. F., Joca, S. R., Correa, F. M., & Guimaraes, F. S. (2009).
5-HT1A receptors are involved in the cannabidiol-induced attenuation of behavioural and
cardiovascular responses to acute restraint stress in rats. Br J Pharmacol, 156(1), 181-
188. doi: 10.1111/j.1476-5381.2008.00046.x
Richards, M. W., Butcher, A. J., & Dolphin, A. C. (2004). Ca2+ channel beta-subunits: structural
insights AID our understanding. Trends Pharmacol Sci, 25(12), 626-632. doi:
10.1016/j.tips.2004.10.008
Robson, P. (2001). Therapeutic aspects of cannabis and cannabinoids. Br J Psychiatry, 178, 107-
115.
Rosenkrantz, H., Fleischman, R. W., & Grant, R. J. (1981). Toxicity of short-term administration
of cannabinoids to rhesus monkeys. Toxicol Appl Pharmacol, 58(1), 118-131.
Ross, H. R., Napier, I., & Connor, M. (2008). Inhibition of recombinant human T-type calcium
channels by Delta9-tetrahydrocannabinol and cannabidiol. J Biol Chem, 283(23), 16124-
16134. doi: 10.1074/jbc.M707104200
Ruiz-Valdepenas, L., Martinez-Orgado, J. A., Benito, C., Millan, A., Tolon, R. M., & Romero, J.
(2011). Cannabidiol reduces lipopolysaccharide-induced vascular changes and
inflammation in the mouse brain: an intravital microscopy study. J Neuroinflammation,
8(1), 5. doi: 10.1186/1742-2094-8-5
Russo, E., & Guy, G. W. (2006). A tale of two cannabinoids: the therapeutic rationale for
combining tetrahydrocannabinol and cannabidiol. Med Hypotheses, 66(2), 234-246. doi:
10.1016/j.mehy.2005.08.026
Russo, E. B., Burnett, A., Hall, B., & Parker, K. K. (2005). Agonistic properties of cannabidiol at
5-HT1a receptors. Neurochem Res, 30(8), 1037-1043. doi: 10.1007/s11064-005-6978-1
Ryan, D., Drysdale, A. J., Lafourcade, C., Pertwee, R. G., & Platt, B. (2009). Cannabidiol targets
mitochondria to regulate intracellular Ca2+ levels. J Neurosci, 29(7), 2053-2063. doi:
10.1523/jneurosci.4212-08.2009

88
Ryberg, E., Larsson, N., Sjogren, S., Hjorth, S., Hermansson, N. O., Leonova, J., . . . Greasley, P.
J. (2007). The orphan receptor GPR55 is a novel cannabinoid receptor. Br J Pharmacol,
152(7), 1092-1101. doi: 10.1038/sj.bjp.0707460
Samara, E., Bialer, M., & Harvey, D. J. (1990). Identification of urinary metabolites of
cannabidiol in the dog. Drug Metab Dispos, 18(5), 571-579.
Schicho, R., & Storr, M. (2011). Alternative targets within the endocannabinoid system for future
treatment of gastrointestinal diseases. Can J Gastroenterol, 25(7), 377-383.
Scuderi, C., Filippis, D. D., Iuvone, T., Blasio, A., Steardo, A., & Esposito, G. (2009).
Cannabidiol in medicine: a review of its therapeutic potential in CNS disorders.
Phytother Res, 23(5), 597-602. doi: 10.1002/ptr.2625
Shepherd, N., & McDonough, H. B. (1998). Ionic diffusion in transverse tubules of cardiac
ventricular myocytes. Am J Physiol, 275(3 Pt 2), H852-860.
Showalter, V. M., Compton, D. R., Martin, B. R., & Abood, M. E. (1996). Evaluation of binding
in a transfected cell line expressing a peripheral cannabinoid receptor (CB2):
identification of cannabinoid receptor subtype selective ligands. J Pharmacol Exp Ther,
278(3), 989-999.
Skrabek, R. Q., Galimova, L., Ethans, K., & Perry, D. (2008). Nabilone for the treatment of pain
in fibromyalgia. J Pain, 9(2), 164-173. doi: 10.1016/j.jpain.2007.09.002
Smiley, K. A., Karler, R., & Turkanis, S. A. (1976). Effects of cannabinoids on the perfused rat
heart. Res Commun Chem Pathol Pharmacol, 14(4), 659-675.
Spurgeon, H. A., duBell, W. H., Stern, M. D., Sollott, S. J., Ziman, B. D., Silverman, H. S., . . .
Lakatta, E. G. (1992). Cytosolic calcium and myofilaments in single rat cardiac myocytes
achieve a dynamic equilibrium during twitch relaxation. J Physiol, 447, 83-102.
Stanley, C. P., Hind, W. H., & O'Sullivan, S. E. (2013). Is the cardiovascular system a therapeutic
target for cannabidiol? Br J Clin Pharmacol, 75(2), 313-322. doi: 10.1111/j.1365-
2125.2012.04351.x
Sumariwalla, P. F., Gallily, R., Tchilibon, S., Fride, E., Mechoulam, R., & Feldmann, M. (2004).
A novel synthetic, nonpsychoactive cannabinoid acid (HU-320) with antiinflammatory
properties in murine collagen-induced arthritis. Arthritis Rheum, 50(3), 985-998. doi:
10.1002/art.20050
Takahashi, M., Seagar, M. J., Jones, J. F., Reber, B. F., & Catterall, W. A. (1987). Subunit
structure of dihydropyridine-sensitive calcium channels from skeletal muscle. Proc Natl
Acad Sci U S A, 84(15), 5478-5482.

89
Takeda, S., Usami, N., Yamamoto, I., & Watanabe, K. (2009). Cannabidiol-2',6'-dimethyl ether, a
cannabidiol derivative, is a highly potent and selective 15-lipoxygenase inhibitor. Drug
Metab Dispos, 37(8), 1733-1737. doi: 10.1124/dmd.109.026930
Thomas, A., Baillie, G. L., Phillips, A. M., Razdan, R. K., Ross, R. A., & Pertwee, R. G. (2007).
Cannabidiol displays unexpectedly high potency as an antagonist of CB1 and CB2
receptor agonists in vitro. Br J Pharmacol, 150(5), 613-623. doi: 10.1038/sj.bjp.0707133
Thompson, A. J., Chau, P. L., Chan, S. L., & Lummis, S. C. (2006). Unbinding pathways of an
agonist and an antagonist from the 5-HT3 receptor. Biophys J, 90(6), 1979-1991. doi:
10.1529/biophysj.105.069385
Turkanis, S. A., & Karler, R. (1986). Cannabidiol-caused depression of spinal motoneuron
responses in cats. Pharmacol Biochem Behav, 25(1), 89-94.
Vanhoutte, P. M., Shimokawa, H., Tang, E. H., & Feletou, M. (2009). Endothelial dysfunction
and vascular disease. Acta Physiol (Oxf), 196(2), 193-222. doi: 10.1111/j.1748-
1716.2009.01964.x
Walsh, S. K., Hepburn, C. Y., Kane, K. A., & Wainwright, C. L. (2010). Acute administration of
cannabidiol in vivo suppresses ischaemia-induced cardiac arrhythmias and reduces
infarct size when given at reperfusion. Br J Pharmacol, 160(5), 1234-1242. doi:
10.1111/j.1476-5381.2010.00755.x
Wang, S. Q., Song, L. S., Lakatta, E. G., & Cheng, H. (2001). Ca2+ signalling between single L-
type Ca2+ channels and ryanodine receptors in heart cells. Nature, 410(6828), 592-596.
doi: 10.1038/35069083
Ware, M. A., Daeninck, P., & Maida, V. (2008). A review of nabilone in the treatment of
chemotherapy-induced nausea and vomiting. Ther Clin Risk Manag, 4(1), 99-107.
White, H. L., & Tansik, R. L. (1980). Effects of delta 9-tetrahydrocannabinol and cannabidiol on
phospholipase and other enzymes regulating arachidonate metabolism. Prostaglandins
Med, 4(6), 409-417.
Xiong, W., Cui, T., Cheng, K., Yang, F., Chen, S. R., Willenbring, D., . . . Zhang, L. (2012).
Cannabinoids suppress inflammatory and neuropathic pain by targeting alpha3 glycine
receptors. J Exp Med, 209(6), 1121-1134. doi: 10.1084/jem.20120242
Yang, J., Ellinor, P. T., Sather, W. A., Zhang, J. F., & Tsien, R. W. (1993). Molecular
determinants of Ca2+ selectivity and ion permeation in L-type Ca2+ channels. Nature,
366(6451), 158-161. doi: 10.1038/366158a0
Yang, K. H., Galadari, S., Isaev, D., Petroianu, G., Shippenberg, T. S., & Oz, M. (2010). The
nonpsychoactive cannabinoid cannabidiol inhibits 5-hydroxytryptamine3A receptor-

90
mediated currents in Xenopus laevis oocytes. J Pharmacol Exp Ther, 333(2), 547-554.
doi: 10.1124/jpet.109.162594
Zhou, Z., & January, C. T. (1998). Both T- and L-type Ca2+ channels can contribute to
excitation-contraction coupling in cardiac Purkinje cells. Biophys J, 74(4), 1830-1839.
doi: 10.1016/s0006-3495(98)77893-2
Zuardi, A. W. (2008). Cannabidiol: from an inactive cannabinoid to a drug with wide spectrum of
action. Rev Bras Psiquiatr, 30(3), 271-280.
Zucchi, R., & Ronca-Testoni, S. (1997). The sarcoplasmic reticulum Ca2+ channel/ryanodine
receptor: modulation by endogenous effectors, drugs and disease states. Pharmacol Rev,
49(1), 1-51.