the golgi apparatus of tetrahymena thermophila
TRANSCRIPT

J . Euk. dflrrohml. 40(1). 1993. pp. 1&13 'C 1993 by the .Gmely of i'ro1070@~C~gl515
The Golgi Apparatus of Tetrahymena thermophila SABINE KURZ and ARNO TIEDTKE'
Institute.for General Zoology and Genetics. L'niversiiy of Munster, SchloJplaiz 5 . 0-4400 Munster, Germany
ABSTRACT. Electron microscopic investigations reveal that the Golg apparatus of Tetrahymena therrnophila consists o f numerous tiny dictyosomes, each consisting of one or two cistemae. The dictyosomes are localized predominantly in the cell cortex closely associated with the mitochondria. arranged in meridians alternating with the ciliary meridians. We estimated about 300-400 of thew dictyosomes in the periphery of a cell, a value corresponding to the number of somatic cilia per cell. Cytochernical assays of thiamine pyrophosphatase and acid phosphatase. both marker enzymes of trans Golgi cisternae, resulted in deposits of lead or cerium phosphate in the ouiermost cisternae of the dictyosomes. In addition, cisternae located at the bases of the basal body/parasomal sac arrangements are stained. This indicates that these cistemae may belong to the Golgi apparatus of the cell.
Supplementary key words. Acid phosphatase, cytochemistry, dictyosomes, thiamine pyrophosphatase.
HE Golgi apparatus is the central sorting organelle of eu- T karyotic cells. Proteins destined for the plasma membrane, secretory vesicles, and lysosomes are addressed for and distrib- uted to their sites of destination by this organelle. N o classical Golgi apparatus has been observed in the ciliate Tetruhj~mena. Instead. tiny dictyosomes have been described as Golgi appa- ratus-equivalent organelles [ I . 8). Elliott & Zieg [ 5 ] interpreted stacks of smooth cisternae near the junction area ofconjugating cells as Golgi structures. None of the structures however. has been identified as Golgi apparatus by the use of marker enLymes of this organelle. Tetrah~wzerru thertnophilu possesses at least two independent secretory pathways, one resulting in secretion ofmucocysts [ 141 and another resultingin secretion of lysosomal enzymes [7. 181. Trimming ol' N-linked carbohjdrate moieties of lysosomal enzymes is thought to occur during their passage through the Golgi cisternae [ 1 I . 161. These discoveries prompt- ed us to study the location and distribution of Golgi structures in this cell.
We found several hundred tiny Golgi stacks located predom- inantly around the entire surface of the cell. Most of these dic- tyosomes occur closely associated with cortical mitochondria aligned in rows between the ciliary meridians. llsing cytochem- ical staining of Golgi-specific marker enzymes, we identified the outermost cisternae of these dictyosomes as trans Golgi com- partments.
MATERIALS AND METHODS Cell culture. We used the strains CU 399 (wild type) and a
secretory mutant MS- I , blocked in secretion of acid hydrolases, of 7'. therrnophila in this study. Cells were cultured as previously described [ 10) in 1 O/o proteose peptone medium supplemented with 0. I0/o yeast extract and 0.003% sequestrene.
Cytochemical localization of thiamine pyrophosphatase. Fix- urion. Cells were fixed for 45 min at 4' C in 1.S0/n glutaralde- hyde. 60 mM sodium cacodylate, pH 7.4. After 5 rinses in the same buffer supplemented with 5% sucrose the cells were equil- ibrated in 60 niM Tris-maleate, pH 7.2. containing Soh sucrose.
Enzyme assay. The fixed cells were incubated for 60 min at 37" C in an en/.yme reaction medium which consisted of 2.2 mM thiamine pyrophosphate (TPP) chloride, 5 mM manganese chloride, 3 mM cerium chloride, 60 mM Tris-maleate. pH 7.2, 5% sucrose. For control ofthe enzyme specificity, the incubation was pcrfnrmcd in medium without TPP.
I'os!fiuurion. After washing in 60 mM Tris-maleate, pH 7.2, 5Yo sucrose. and 60 mM sodium cacodylate, pH 6.0, 5% sucrose the cells were postfixed in lo/o OsO,. 60 mM sodium cacodylate pH 7 .7 . 5010 sucrose.
Cytochemical lucalizrrfiun vf acid phosphatase. Fixation. Cells were fixed for 45 niin at 4" t ' i i l 0.4"h glutaraldetiydc 1x1
I To whom correspondence should be addressed.
60 mM sodium cacodylate pH 7.4 and then rinsed 5 times in 50 mM sodium acetate pH 5.0, 5'10 sucrose.
Enzyme assay. The cells were incubated for 30 min at 30" C in a medium containing 5 parts of 12% Pb (NO,), and 1 part of 3% sn glycerol 2-phosphate, 50 mM sodium acetate, pH 5.0, 5% sucrose. For the control, cells were incubated in medium without the enzymc substrate P-glycerophosphate or preincu- bated in medium containing 10 mM NaF, known to inhibit acid phosphatase activities in Tefruhymma. Non-specific precipi- tates were dissolved by a short rinse with 2% acetic acid.
Postfiwtion. Stained cells were postfixed in 2.5% glutaral- dehyde followed by I % OsO,: 60 mM sodium cacodylate pH 7.4, 5% sucrose.
Embedding. Stained and fixed cells were first contrasted ''en bloc" with 1% uranylacetate then dehydrated and embedded in Epon.
RESULTS Structure and location of the Golgi apparatus. The Golgi
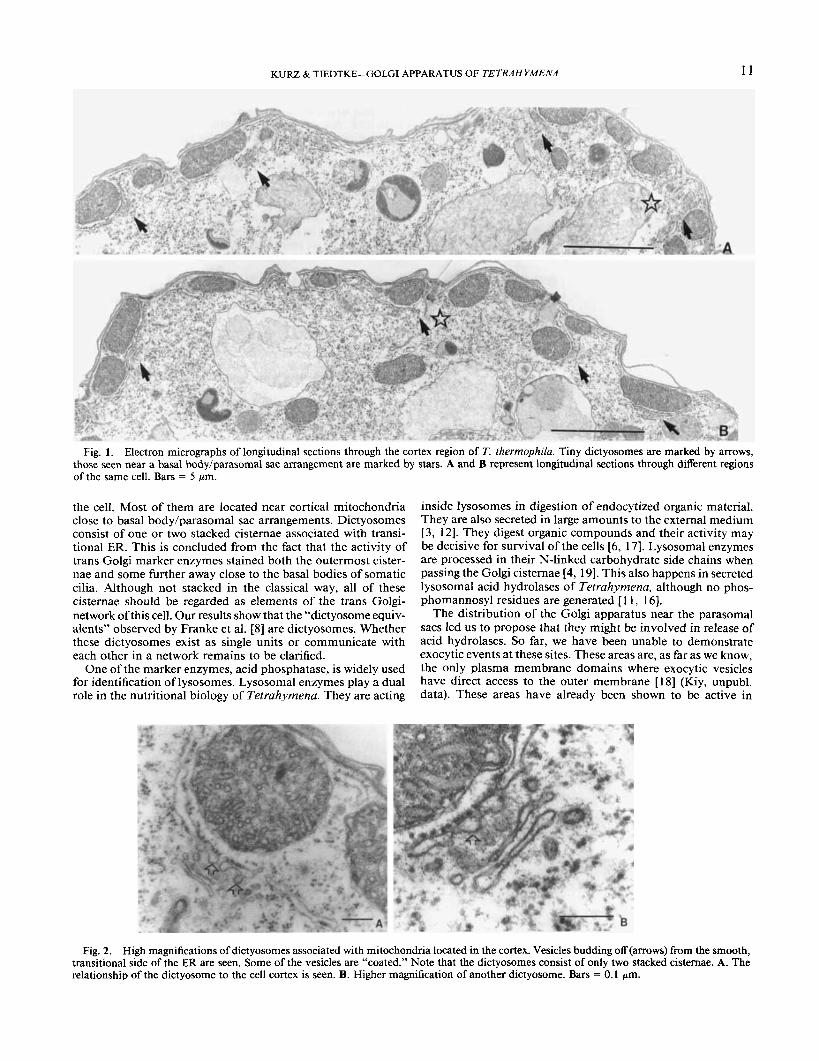
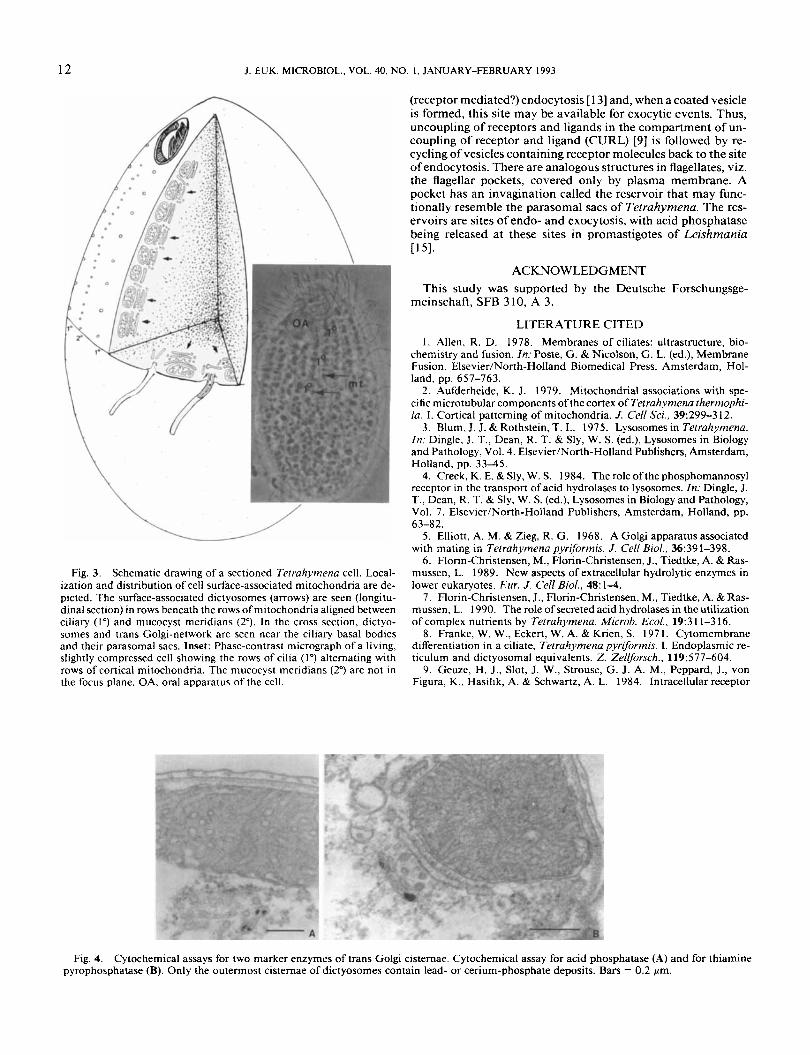
apparatus of Tefrahyrnena consists of hundreds of tiny dictyo- somes predominantly localized in the cortex region of the cell (Fig. I , arrows). The dictyosonies are composed of one or two cisternae which are surrounded by small vesicles. Vesicles pinching off from the smooth transitional membrane of rough endoplasmic reticulum (ER) closely associated with the surface of mitochondria of the cell cortex are regularly observed (Fig. 2). Most mitochondria in Tetruhytnena are aligned in rows be- tween the primary, ciliary meridians and the secondary merid- ians formed by rows of mucocysts in the cell cortex [2] (Fig. 3. inset). From serial sections through cortices ofcells we estimated almost as many tiny dictyosomes as somatic cilia, i.e. about 300-400 per cell. This is depicted in the schematic drawing in Fig. 3.
Cytochemical localization of the trans Golgi marker enzymes acid phosphatase and thiamine pyrophosphatase. Acid phos- phatase activity staining was used to identify the trans cisternae of dictyosomes. The deposits of lead phosphate are found in the outermost cisternae of the dictyosomes (Fig. 4a). We confirmed this detection by cytochemical staining of thiamine pyrophos- phatase activity, an additional indicator enzyme of trans Golgi cisternae. Deposits of reaction products of this enzyme were found in the same cisternae as those containing acid phosphatase (Fig. 4b). Not only the outer cisternae of the dictyosonies but also cisternae underneath the basal bodies were stained (Fig. 5) in both marker enzyme assays for trans Golgi cisternae indi- cating that these cisternae may belong to the trans Golgi-net- work.
The Golgi apparatus of 7: thertnrphilu differs from the clas- sical Golgi apparatus of polar cells. It is composed of several hundred tiny dictyosomes located below the entire surface of
10
KURZ & TIEDTKE-GOLGI APPARATUS OF TETRAHYMENA 11
Fig. 1. Electron micrographs of longitudinal sections through the cortex region of T. thermophilu. Tiny dictyosomes are marked by arrows, those seen near a basal body/parasomal sac arrangement are marked by stars. A and B represent longitudinal sections through different regions of the same cell. Bars = 5 pm.
the cell. Most of them are located near cortical mitochondria close to basal body/parasomal sac arrangements. Dictyosomes consist of one or two stacked cisternae associated with transi- tional ER. This is concluded from the fact that the activity of trans Golgi marker enzymes stained both the outermost cister- nae and some further away close to the basal bodies of somatic cilia. Although not stacked in the classical way, all of these cisternae should be regarded as elements of the trans Golgi- network of this cell. Our results show that the “dictyosome equiv- alents” observed by Franke et al. [S] are dictyosomes. Whether these dictyosomes exist as single units or communicate with each other in a network remains to be clarified.
One of the marker enzymes, acid phosphatase, is widely used for identification of lysosomes. Lysosomal enzymes play a dual role in the nutritional biology of Tetrahymena. They are acting
inside lysosomes in digestion of endocytized organic material. They are also secreted in large amounts to the external medium [3, 121. They digest organic compounds and their activity may be decisive for survival of the cells [6, 171. Lysosomal enzymes are processed in their N-linked carbohydrate side chains when passing the Golgi cisternae [4, 191. This also happens in secreted lysosomal acid hydrolases of Tetrahymena, although no phos- phomannosyl residues are generated [ 1 1, 161.
The distribution of the Golgi apparatus near the parasomal sacs led us to propose that they might be involved in release of acid hydrolases. So far, we have been unable to demonstrate exocytic events at these sites. These areas are, as far as we know, the only plasma membrane domains where exocytic vesicles have direct access to the outer membrane [ IS] (Kiy, unpubl. data). These areas have already been shown to be active in
Fig. 2. High magnifications of dictyosomes associated with mitochondria located in the cortex. Vesicles budding off (arrows) from the smooth, transitional side of the ER are seen. Some of the vesicles are “coated.” Note that the dictyosomes consist of only two stacked cisternae. A. The relationship of the dictyosome to the cell cortex is seen. B. Higher magnification of another dictyosome. Bars = 0.1 pm.
12
n J. EUK. MICROBIOL., VOL. 40, NO. I , JANUARY-FEBRUARY 1993
Fig. 3. Schematic drawing of a sectioned Tefrahymena cell. Local- ization and distribution of cell surface-associated mitochondria are de- picted. The surface-associated dictyosomes (arrows) are seen (longitu- dinal section) in rows beneath the rows of mitochondria aligned between ciliary ( I " ) and mucocyst meridians (2"). In the cross section, dictyo- s o m a and trans Golgi-network are seen near the ciliary basal bodies and their parasomal sacs. Inset: Phase-contrast micrograph of a living, slightly compressed cell showing the rows of cilia (I") alternating with rows of cortical mitochondria. The mucocyst meridians (2") are not in the focus plane. OA, oral apparatus of the cell.
(receptor mediated?) endocytosis [ 131 and, when a coated vesicle is formed, this site may be available for exocytic events. Thus, uncoupling of receptors and ligands in the compartment of un- coupling of receptor and ligand (CURL) [9] is followed by re- cycling of vesicles containing receptor molecules back to the site of endocytosis. There are analogous structures in flagellates, viz. the flagellar pockets, covered only by plasma membrane. A pockct has an invagination called the reservoir that may func- tionally resemble the parasomal sacs of Tetrahymena. The res- ervoirs are sites of endo- and exocytosis, with acid phosphatasc being released at these sites in promastigotes of Leishmania
ACKNOWLEDGMENT
~ 5 1 .
This study was supported by the Deutsche Forschungsge- meinschaft, SFB 310, A 3.
LITERATURE CITED 1. Allen. R. D. 1978. Membranes of ciliates: ultrastructure, bio-
chemistry and fusion. In: Poste, G. & Nicolson, G. L. (ed.), Membrane Fusion. Elsevier/North-Holland Biomedical Press, Amsterdam, Hol- land, pp. 657-763.
2. Aufderheide, K. J. 1979. Mitochondria1 associations with spe- cific microtubular components ofthe cortex of Teirahymena thermophi- la. I . Cortical patterning of mitochondria. J. Cell Sci., 39:299-312.
3. Blum, J . J . & Rothstein, T. L. 1975. Lysosomes in Tefrahymcna. In: Dingle. J. T., Dean, R. T. & Sly, W. S. (ed.), Lysosomes in Biology and Pathology, Vol. 4. Elsevier/North-Holland Publishers, Amsterdam, Holland, pp. 33115.
4. Creek, K. E. & Sly, W. S. 1984. The role of the phosphomannosyl receptor in the transport of acid hydrolases to lysosomes. In: Dingle, J. T., Dean, R. T. & Sly, W. S. (ed.), Lysosomes in Biology and Pathology, Vol. 7. Elsevier/North-Holland Publishers, Amsterdam, Holland, pp.
5. Elliott, A. M. & Zieg, R. G. 1968. A Golgi apparatus associated with mating in Teirahymena pyriformis. 1. Cell Biol., 36:39 1-398.
6. Florin-Christensen, M., Florin-Christensen, J., Tiedtke, A. & Ras- mussen, L. 1989. New aspects of extracellular hydrolytic enzymes in lower eukaryotes. Eur. J. Cell Biol., 48: 1-4.
7. Florin-Christensen, J., Florin-Christensen, M., Tiedtke, A. & Ras- mussen, L. 1990. The role of secreted acid hydrolases in the utilization of complex nutrients by Tetrahymena. Microb. Ecol., 19:3 1 1-3 16.
8. Franke, W. W., Eckert, W. A. & Krien, S. 1971. Cytomembrane differentiation in a ciliate, Tetrahyymena pyriformis. I. Endoplasmic re- ticulum and dictyosomal equivalents. 2. Zellforsch.. 119:577-604.
9. Geuze, H. J., Slot, J. W., Strouse, G. J. A. M., Peppard, J., von Figura. K., Hasilik, A. & Schwartz, A. L. 1984. Intracellular receptor
63-82.
Fig. 4. Cytochemical assays for two marker enzymes of trans Golgi cisternae. Cytochemical assay for acid phosphatase (A) and for thiamine pyrophosphatase (B). Only the outermost cisternae of dictyosomes contain lead- or cerium-phosphate deposits. Bats = 0.2 pm.

KURZ & TIEDTKE-GOLGI APPARATUS OF TETRAHYMENA 13
Fig. 5. Cytochemical assays for two marker enzymes of trans Golgi-network cisternae. A, B. Cisternae of trans Golgi-network at the bases of basal body/parasomal sacs (ps) and of a dictyosome (dy) are stained positively for thiamine pyrophosphatase (dark deposits of cerium phosphate). C, D. Sub-surface cisternae, presumably belonging to the trans Golgi-network, are stained positively for acid phosphatase activity. Bars = 2 pm.
sorting during endocytosis: comparative immunoelectron microscopy of multiple receptors in rat liver. Cell, 37: 195-204.
10. Hunseler, P., Scheidgen-Kleyboldt, G., Tiedtke, A. 1987. Iso- lation and characterization of a mutant of Tetrahymena thermophila blocked in secretion of lysosomal enzymes. J. Cell Sci.. 88:47-55.
1 1. Hiinseler, P., Tiedtke, A. & von Figura, K. 1988. Biosynthesis of secreted 8-hexosaminidase in Tetrahymena thermophila. A compar- ison of the wild type with a secretory mutant. Biochem. J., 252:837- 842.
12. Miiller, M. 1972. Secretion of acid hydrolases and its intracel- Mar source in Tetrahymena pyriformis. J. Cell Biol., 52:478487.
13. Nilsson, J. R. & van Deurs, B. 1983. Coated pits and pinocytosis in Tetrahymena. J. Cell Sci., 63:209-222.
14. Onas, E., Hacks, M. & Satir, B. H. 1983. Isolation and ultra- structural characterization of secretory mutants of Tetrahymena ther- mophila. J. Cell Sci., 64:49-67.
15. Stierhof, Y.-D., Schwarz, H., Menz, B., Russell, D. B., Quinten, M. & Overath, P. 199 1 . Monoclonal antibodies to Leishmania mex- icana promastigote antigens. 11. Cellular localization of antigens in pro- mastigotes and infected macrophages. J. Cell Sci., 99:18 1-186.
16. Taniguchi, T. Mizuochi, T., Banno, Y., Nozawa, Y. & Kobata, A. 1985. Carbohydrates of lysosomal enzymes secreted by Tetrahy- menu pyriformis. J. Biol. Chem., 260: 13941-1 3945.
17. Tiedtke, A. & Rasmussen, L. 1988. Lysosomal enzymes in extracellular digestion in the unicellular eukaryote Tetrahymena. J. Cell. Ph ysiol., 136: 5 54-5 5 6.
18. Tiedtke, A. & Rasmussen, L. 1989. Constitutive secretion of acid hydrolases in Tetrahymena thermophila. J. Protzool., 36:378-382.
19. von Figura, K. & Hasilik, A. 1986. Lysosomal enzymes and their receptors. Ann. Rev. Biochem., 55: 167-1 93.
Received 11-25-91, 7-29-92; accepted 7-31-92