the hernandez and fernandez genes of drosophila - core · the hernandez and fernandez genes of...
TRANSCRIPT

The hernandez and fernandez genes ofDrosophilaspecify eye and antenna
Magali Suzanne, Carlos Estella, Manuel Calleja, and Ernesto Sa´nchez-Herrero*Centro de Biologıa Molecular Severo Ochoa, Universidad Autonoma de Madrid, Cantoblanco, 28049 Madrid, Spain
Received for publication 17 December 2002, revised 8 April 2003, accepted 16 April 2003
Abstract
The formation of different structures inDrosophila depends on the combined activities of selector genes and signaling pathways. Forinstance, the antenna requires the selector genehomothorax, which distinguishes between the leg and the antenna and can specify distalantenna if expressed ectopically. Similarly, the eye is formed by a group of “eye-specifying” genes, among themeyeless, which can directeye development ectopically. We report here the characterization of thehernandez andfernandez genes, expressed in the antennal and eyeprimordia of the eye–antenna imaginal disc. The predicted proteins encoded by these two genes have 27% common amino acids and includea Pipsqueak domain. Reduced expression of eitherhernandez or fernandez mildly affects antenna and eye development, while theinactivation of both genes partially transforms distal antenna into leg. Ectopic expression of either of the two genes results in two differentphenotypes: it can form distal antenna, activating genes likehomothorax, spineless, andspalt, and it can promote eye development andactivateseyeless. Reciprocally,eyeless can inducehernandez and fernandez expression, andhomothorax and spineless can activate bothhernandez and fernandez when ectopically expressed. The formation of eye by these genes seems to requireNotch signaling, since theinduction of ectopic eyes and the activation ofeyeless by thehernandez gene are suppressed when theNotch function is compromised. Ourresults show that thehernandez and fernandez genes are required for antennal and eye development and are also able to specify eye orantenna ectopically.© 2003 Elsevier Inc. All rights reserved.
Keywords: Drosophila; Antenna; Eye;eyeless; homothorax; spineless; Psq motif
Introduction
One important issue in developmental biology is to un-derstand how specific structures are determined. InDro-sophila melanogaster, this is accomplished in part throughthe activity of a group of genes called selector or selector-like genes, which confer identity to structures as diverse aslegs, eyes, or wings (Mann and Morata, 2000).
Hox (homeotic) genes, which specify structures alongthe anterior–posterior axis of most animals (Gellon andMcGinnis, 1998), are also commonly referred to as selectorgenes (Garcı´a-Bellido, 1975). The functional distinction be-tween Hox and non-Hox selector genes is not straightfor-ward. In general, Hox genes can differentiate between struc-
tures considered homologous (like wings and halteres),whereas non-Hox selector genes can specify structures inregions of the fly in which the homology with the new,ectopic organ, is not evident. Thus,Distal-less (Dll), anon-Hox selector gene, induces leg tissue when expressed inthe wing (Gorfinkiel et al., 1997; Campbell and Tomlinson,1998), and theeyeless (ey) gene makes ectopic eye tissue ifpresent in antennae, legs, or proximal wing (Halder et al.,1995). There are structures inDrosophila which do notrequire Hox information and that can be formed ectopicallyby different non-Hox selector genes. These structures in-clude two head organs, the eye and the antenna, derivedfrom the eye–antenna imaginal disc.
The antennae are ventral structures homologous to legs(Postlethwait and Schneiderman, 1971). Both types of ap-pendages share common positional information determinedby the activity of thehedgehog, decapentaplegic, andwing-
* Corresponding author. Fax:�34-913974799.E-mail address: [email protected] (E. Sa´nchez-Herrero).
R
Available online at www.sciencedirect.com
Developmental Biology 260 (2003) 465–483 www.elsevier.com/locate/ydbio
0012-1606/03/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/S0012-1606(03)00249-5

less (wg) products (Campbell and Tomlinson, 1995; Brooket al., 1996). These molecules determine the distribution ofthe homothorax (hth), Dll, and dachsund (dac) genes, whichgenetically subdivide both appendages along the proximal–distal axis (Dıaz-Benjumea et al., 1994; Lecuit and Cohen,1997; Gorfinkiel et al., 1997; Gonzalez-Crespo et al., 1998;Abu-Shaar and Mann, 1998; Wu and Cohen, 1999; Dong etal., 2001; reviewed in Morata, 2001).
The difference between antennae and legs relies in thedistinct spatial distribution of the Hox gene Antennapedia(Struhl, 1981) and of hth, the selector gene for the antenna.hth and Dll form an antenna in its wild-type location or inother positions if coexpressed (Casares and Mann, 1998;Dong et al., 2000). The spineless (ss) gene is also able tospecify distal antennal development (Duncan et al., 1998),but it seems to mediate Dll and hth function (Duncan et al.,1998; Dong et al., 2002). In the leg disc, where Dll is alsoexpressed, the Antennapedia (Antp) Hox gene prevents hthexpression, thus directing this primordium toward leg de-velopment (Casares and Mann, 1998). Although severalgenes are differently expressed in the antennal primordiumand the leg disc (Dong et al., 2001), only one gene, spalt(sal), is transcribed in the antenna and not in the leg (Wag-ner-Bernholz et al., 1991). However, sal is also expressed inother discs (de Celis et al., 1996; Barrio et al., 1999) and nogene expressed exclusively in the antennal primordium hasyet been described.
The Drosophila eye is a complex organ derived from theeye primordium of the eye–antenna imaginal disc. The eygene was first identified as a “master” or selector gene foreye development, since it is required for the formation of theeye and can induce eye development when ectopically ex-pressed (Halder et al., 1995). Subsequent studies, however,identified other genes, such as twin of eyeless, eyegone, eyesabsent, dachsund, and sine oculis, with similar characteris-tics. Most of them are involved in complex regulatory net-works and protein–protein interactions that determine eyedevelopment (reviewed in Treisman, 1999; Kumar, 2001).Interestingly, ectopic activation of the Notch (N) gene, en-coding a receptor involved in many developmental deci-sions (Artavanis-Tsakonas et al., 1999), induces ectopiceyes in a wild-type head and transforms the eye into antennain an ey mutant background (Kurata et al., 2000). Moreover,recent work suggests that the N and Epidermal growthfactor Receptor (Egfr) signaling pathways are involved inthe distinction between eye and antenna early in develop-ment (Kumar and Moses, 2001). These results suggest thatthere is not a single selector gene for the determination ofthe eye and that signaling pathways are somehow involvedin the regulatory loops that specify this structure.
To identify genes involved in antennal development, wehave used the UAS-yellow method (Calleja et al., 1996) toselect GAL4 lines showing restricted expression patternseither in the leg or the antenna. We have characterized fourof these lines, which are inserted in two related genes, thatwe called hernandez (hern) and fernandez (fer) (Spanish
names for the twin brothers in Tintin comic-books). Arecent report (Emerald et al., 2003) has characterized thesesame genes and given the names of distal antenna (dan) tothe fernandez gene and distal antenna related (danr) to thehernandez one. These genes are expressed in the eye–an-tennal discs and not in the leg or other imaginal discs.Reduced hern or fer expression leads to small defects inantennal and eye development, and when the activity ofboth genes is compromised, there is a partial transformationof the distal antenna into leg. When either hern or fer isexpressed ectopically, they activate hth and ss expressionand transform leg tissue into distal antenna. These genes canalso make ectopic eye tissue and activate ey. Our resultssuggest that the hern and fer genes are involved in antennaand eye specification.
Materials and methods
Isolation of GAL4 lines and genetic strains
The method used to isolate the GAL4 lines has beenpreviously described (Calleja et al., 1996). The UAS-lacZ(Brand and Perrimon, 1993), UAS-y (Calleja et al., 1996),UAS-GFP (Ito et al., 1997), UAS-Necd (Klein et al., 1997),Antp73b (Hazelrigg and Kaufman, 1983), UAS-hth (Pai etal., 1998), Dll3 (Cohen and Jurgens, 1989), UAS-EN-HTH1-430 (Inbal et al., 2001), ssD115.7, UAS-ss (Duncan etal., 1998), and hs-hth (Pai et al., 1998) stocks have beenpreviously described. Ectopic expression was obtained byusing the GAL4/UAS system (Brand and Perrimon, 1993)with the following GAL4 lines: dpp-GAL4 (Staehling-Hampton et al., 1994), Dll-GAL4 (MD23 and EM212 lines;Calleja et al., 1996), GMR-GAL4 (Yamaguchi et al., 1999),and ptc-GAL4 (Hinz et al., 1994). Df slo8 is a deficiencyremoving the genomic region where the hern and fer genesare located (Atkinson et al., 1991).
Isolation of derivatives from the P-element insertions
The MD634 and CES115 P-GAL4 lines were mobilizedto obtain imprecise excisions of the P-element. yw; MD634(or CES115)/TM3, � 2-3 males were individually crossed toyw; TM3/TM6B females, and individual w� males wereisolated and used to establish yw; MD634* (or CES115*)/TM6B stocks (the asterisk marks a possible imprecise ex-cision). The phenotype of MD634* or CES115* homozy-gotes from the stocks was analyzed.
Clonal analysis
Ectopic expression clones (Pignoni and Zipurski, 1997)were induced by giving a 10-min heat shock at 34°C to yhs-flp; act �y�� GAL4 UAS-GFP/ UAS-fer or UAS-hernsecond instar larvae. The act �y�� GAL4 UAS-GFP (Ay-GAL4.25 UAS-GFP S65T) stock has been previously de-
466 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

Fig. 1. Expression of the hern and fer genes and of the P-GAL4 elements inserted in them. (A) Antenna of a y fly. II and III stand for second and third antennalsegments, respectively; ar, arista. (B, C) Antennae of y; MD634/UAS-y� (B) and y; AC116/UAS-y� (C) flies. The MD634 insertion is located close to thehern gene and the AC116 insertion close to the fer gene. Note that the third antennal segment, the arista, and a few bristles of the second antennal segmentare y� in both cases. Symbols as in (A). (D, E) lacZ expression in third instar eye–antennal imaginal discs of MD634/UAS-lacZ (D) and AC116/UAS-lacZ(E) larvae, showing stronger signal in a ring at the position of the third antennal segment and in the central region of the antennal primordium (a), with weakerlacZ levels in between. There is also expression in the eye primordium, mostly in its posterior region. The morphogenetic furrow is indicated by an arrow.In this and following figures, the dorsal part of the antennal disc is up. (F–H) RNA expression of the hern (F) and fer (G) genes in the eye–antennal disc,resembling the expression driven by the GAL4 lines in the antennal primordium (compare with D and E, respectively); in the eye primordium, strong signalis observed anterior to the morphogenetic furrow (indicated by an arrow), while the posterior, differentiated eye (posterior to the morphogenetic furrow) onlyshows weak expression. Detail of the signal in the eye is shown in (H). A and P stand for regions anterior and posterior to the morphogenetic furrow (arrow),respectively.
467M. Suzanne et al. / Developmental Biology 260 (2003) 465–483


scribed (Ito et al., 1997). The clones were identified by theGFP expression.
Molecular techniques, pUAS constructs
The P-Gal4 lines were localized by inverse PCR (http://www.fruitfly.org/about/methods/inverse.pcr.html). The re-maining molecular techniques were done according to stan-dard protocols (Sambrook et al., 1989). The molecularstructure of the genes and the reconstruction of the openreading frame (ORF) were done according to the annotationof the Drosophila genome (Adams et al., 2000). ThepUAST-fer construct was done by cloning first a partiallydigested EcoRI/XhoI 3.1-kb fragment from the LD40883EST into the Bluescript vector; this new vector was thendigested with XhoI and NotI and inserted into the pUASTvector. To make the pUAST-hern construct, we first ampli-fied a 1.4 kb genomic fragment by PCR (Unit Expand LongTemplate PCR system; Roche) with the 5�-ttcgaggagtgca-caccggtggtc-3� upper primer and the 5�-ctactgttcggcgtcgct-ggacccg-3� lower primer, and cloned it in a modified Blue-script vector, digested with EcoRV and then treated withTaq Polymerase (Unit Expand Long Template PCR System;Roche) to add dTTP in its 3� end; then, we cloned a XbaI–XhoI fragment from this construct in the pUAST vector.
PCR analysis of the derivatives
To analyze the ferI49-1 deletion, we amplified by PCR agenomic fragment in the region of the P-element insertionwith 5�-ggcggctggcaaggagctgcag-3� and 5�-gccttcttccagc-gaaggcggg-3� primers, which hybridize to sequences lo-cated at both sides of the insertion. In wild-type flies, thefragment amplified was about 3.5 kb long, and in ferI49-1
homozygous flies, about 1 kb long.
RNA interference
The GAL4-inducible constructs for RNA interferencewere made as follows: for the hern gene, a 524-bp fragmentfrom the hern exon was amplified by PCR with the 5�-tgaggatccggatgagcacgcgcggc-3� upper primer and the 5�-gcaggtaccaggacatcggcagtctgtggg-3� lower primer; for the
fer gene, a 605-bp fragment from the second exon of the fertranscript was amplified by PCR with the 5�-acaggatccatgc-catccagaggatccacg-3� upper primer and the 5� cgcggtaccgg-gattcagctgggccagc-3� lower primer. Each PCR product wasindependently cloned as a BamHI–KpnI fragment in thepHIBS vector (Nagel et al., 2002), to make the pHIBS-hernor pHIBS-fer constructs. The BamHI–SacI fragments frompHIBS-hern or pHIBS-fer were subcloned in the Bluescriptvector, generating the BS-INT-hern and BS-INT-fer con-structs, respectively. Finally, SalI–KpnI fragments frompHIBS-hern and BS-INT-hern (or pHIBS-fer and BS-INT-fer) constructs were cloned together in the pUAS vector atits KpnI site, thus forming the final RNAi constructs, pUAS-ihern (ihern) and pUAS-ifer (ifer). The RNAi constructs(and the UAS-constructs) were injected into y w1118 em-bryos by standard procedures.
In situ hybridization
In situ hybridization was performed according to Az-piazu and Frasch (1993). The DNA probes of the hern andfer genes were synthesized by random priming with theBoehringer Mannheim kit, using as templates a 1361-bpfragment from the coding region of the hern gene and a1192-bp fragment from the second exon of the fer gene,both PCR-amplified with the following primers: 5�-ctgttcg-gcgtcgctggaccc-3� and 5�-gtgcacaccggtggtcgcatccgc-3� forthe hern gene, and 5�-cgccaagctgttcgacaacggcct-3� and 5�-gcccgacggtgtggacgagggtgt-3� for the fer gene. The hernRNA probe, used to detect inactivation of hern by RNAi,was synthesized from a hern cDNA cloned in the Bluescriptvector. This construct was digested with BamHI so that theprobe was specific for the 3� end of the gene (not includedin ihern). The ss probe was synthesized from a ss cDNAcloned in the Bluescript vector and provided by Ian Duncan.
Immunostaining and histochemical staining of Drosophilaimaginal discs
The antibody staining was done according to Azpiazuand Morata (2000). The antibodies used are: guinea piganti-Hth (Azpiazu and Morata, 2002), mouse anti-Dac(mAbdac2-3; Hybridoma Bank, The University of Iowa),
Fig. 2. Location of the P-GAL4 elements, molecular characterization of the ferI49-1 mutation, molecular structure and sequence of the hern and fer genes,and comparison of the Psq domains of the proteins encoded by them. (A) DNA map showing the DNA region containing the hern/danr and fer/dantranscription units, their transcript structure according to the annotation of the Drosophila genomic sequence (Adams et al., 2000), and the location of theP-GAL4 transposable elements. Solid red boxes represent exons, the lines joining them indicate the introns, and the arrows indicate the direction oftranscription. There are two predicted transcription units between the hern and fer transcription units (CG13652 and CG13661), shown as small black squareswhich represent predicted exons. The distance between the insertion points of two GAL4 lines and the predicted transcription start site of the two genes isalso indicated. The extension of the ferI49-1 deletion is shown at the top of the figure, and the ihern and ifer constructs used for RNA interference at the bottom.(B) Aligned protein sequence of the two genes obtained from the conceptual translation of the hern and fer transcription units. Residues in blue indicatesimilar amino acids, and those in red are identical ones. The open box encloses a highly conserved 69-amino-acid domain, and the 48-amino-acid Pipsqueak(Psq) motif is underlined. (C) Alignment of the Psq motifs of the hern and fer genes, as well as that of a third predicted gene (CG13496 transcription unit),which share a high similarity in this domain with the other two (see Siegmund and Lehmann, 2002). Color of the amino acids indicates similarity and identityas in (B).
469M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

470 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

mouse anti-Dll (Duncan et al., 1998), rabbit anti-Sal (Kuhn-lein et al., 1994), mouse anti-Wg (Brook and Cohen, 1996),rat anti-Elav (Elav-9F8A9; Hybridoma Bank, The Univer-sity of Iowa), mouse anti-Dan (Emerald et al., 2003), rabbitanti-Ey (U. Walldorf, unpublished), and rabbit anti-�-galac-tosidase (Cappel). Secondary antibodies are coupled toRed-X, FITC, and Cy5 fluorochromes (Jackson Immuno-Research) The staining was analyzed in a Zeiss confocalmicroscope. Histochemical staining to detect �-galactosi-dase expression was done according to Ashburner (1989).
Heat-shock experiments.
hs-hth; MD634/UAS-GFP or hs-hth; CES115/UAS-GFPthird instar larvae were given a 15-min heat shock at 37°Cin a water bath. Discs were dissected out of the larvae andfixed 1 h after the heat-shock.
Results
The hern and fer genes are similarly expressed in theantenna and eye primordia
From a collection of P-GAL4 lines that drive expressionin restricted areas of the adult cuticle (Calleja et al., 1996),we selected four lines which specifically direct a similarexpression of the yellow (y) gene: the third antennal seg-ment, the arista, and a few bristles in the second antennalsegment express the y gene (Fig. 1A–C). These lines, whencrossed to a line carrying an UAS-lacZ reporter construct,show �-galactosidase expression in three regions of theeye–antennal disc: in the third antennal segment, in thearista, and within the eye, in the differentiated eye and in aregion just anterior to the morphogenetic furrow (Fig. 1Dand E).
We located by inverse PCR the insertion site of thesefour lines (Fig. 2A): two of them (CES75 and MD634) arein the 5� upstream region of the predicted CG13651 tran-scription unit and the other two (AC116 and CES115) are inthe 5� upstream region of the CG11849 transcription unit,according to the annotated Drosophila genome (Adams et
al., 2000). The two transcription units are located in thechromosomal position 96C2-4, in the same orientation andnear adjacent, separated by about 45 kb. The CG13651transcription unit is 1.259 kb long, bears no introns, andproduces a 1259-bp mRNA, whereas the CG11849 tran-scription unit is 7.175 kb long, has 3 exons, and makes amRNA of 2232 bp (Fig. 2A). There is an EST for theCG11849 unit, indicating that this predicted gene is tran-scribed.
The CG13651 and CG11849 transcription units encodepredicted proteins of 419 and 743 amino acids, respectively,that have 27% identity and 37% similarity between them(Fig. 2B). Both proteins contain similar 69-amino-acid re-gions, located in their N-terminal part, with 78% identityand 88% similarity between them (Fig. 2B). This region in-cludes a previously characterized 48-amino-acid Pipsqueak(Psq) motif, present in several proteins in Drosophila andother species, including human (Horowitz and Berg, 1996;Lehmann et al., 1998; Siegmund and Lehmann, 2002). ThePsq motif is a helix–turn–helix DNA-binding domain sim-ilar to the DNA-binding domain of prokaryotic recombi-nases (Lehmann et al., 1998), which, in turn, also showsimilarities with the homeodomain (Affolter et al., 1991).The CG13496 predicted Drosophila gene codes for a pro-tein with a Psq domain very similar to those of CG13651and CG11849 genes (Siegmund and Lehmann, 2002) (Fig.2C). The CG13496 transcription unit is not expressed in theeye–antennal imaginal disc like the CG13651 and CG11849genes (data not shown).
In situ hybridization experiments show that, among allthe imaginal discs, the CG13651 and CG11849 transcrip-tion units are only and equally expressed in the eye–anten-nal disc. Their expression is very similar to that observedwith the P-GAL4 lines (Fig. 1F and G, compare with Fig.1D and E), except that, in the eye primordium, the RNAsignal in the differentiated eye is very weak (Fig. 1F–H).The same antennal and eye expression persists in earlypupal stages (not shown). Therefore, these genes present asimilar sequence, expression in the eye–antennal disc, reg-ulation, and function (see below). We have called thesegenes hernandez (hern, CG13651) and fernandez (fer,CG11849), and the two of them, the Tintin genes.
Fig. 3. Comparison of MD634-GAL4 expression with those of the hth, Dll, sal, and dac genes in the antennal primordium, and regulation of MD634-GAL4expression by Antp. (A) GFP (green) expression in the antennal primordium of a MD634 UAS-GFP larva. The strong MD634 expression overlaps with thatof hth (B, red) in the proximal part of the third antennal segment. Merged image in (C). (D) Expression of Dll (D, blue) in the same antennal primordium.(E) Expression of Dll (blue) and MD634 (green), showing overlap in the presumptive third antennal segment and the arista. (F) Merged image of GFP, hthand Dll expression. (G) GFP expression (green) in an antennal disc of a MD634 UAS-GFP larva. This disc is from an older larva than that shown in (A).(H) sal (red) and (J) dac (blue) expression in the same disc. (I, K, L) Merged images of MD634 UAS-GFP and sal (I), MD634 UAS-GFP and dac (K), andMD634 UAS-GFP, sal, and dac (L). Note that, in the third antennal segment, the distal limit of strong GFP expression in the third antennal segment coincideswith that of sal (I) and the proximal one with that of dac (K). (M) Scheme of the expression of the hern and fer transcription units in relation to Dll, hth,dac, and sal, after Dong et al. (2001). a1, a2, and a3 refer to the first, second, and third antennal segments, respectively. a4 and a5 represent the base of thearista, and ar stands for arista. The light pink color indicates weak hern and fer expression. It is possible that hern and fer expression extends into a few cellsof a2. (N) Antennae of an Antp73b/� adult, transformed into legs except in the most proximal and more distal regions (arrows, aristae). (O) GFP (green)expression in an Antp73b/MD634 UAS-GFP larva. Note the patchy green expression, which is almost absent in the primordium of the third antennal segment(arrow; compare with A). The expression in the center of the primordium (arrowhead) is almost unchanged, which corresponds with the normal aristae in(N).
471M. Suzanne et al. / Developmental Biology 260 (2003) 465–483
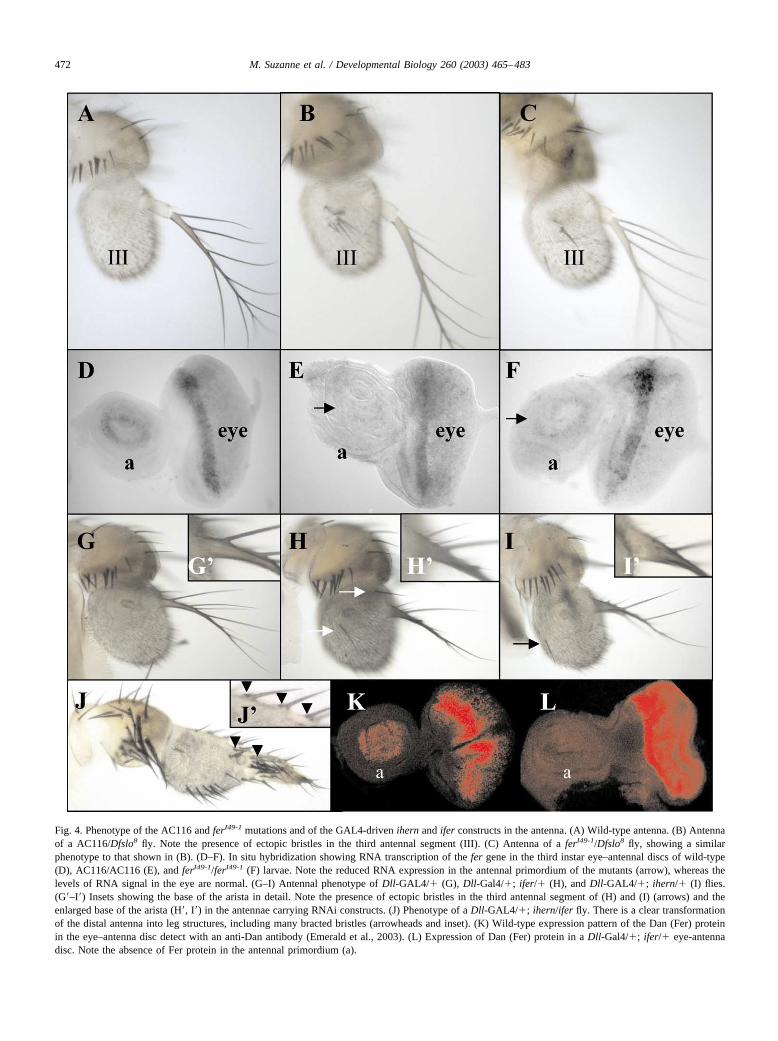
Fig. 4. Phenotype of the AC116 and ferI49-1 mutations and of the GAL4-driven ihern and ifer constructs in the antenna. (A) Wild-type antenna. (B) Antennaof a AC116/Dfslo8 fly. Note the presence of ectopic bristles in the third antennal segment (III). (C) Antenna of a ferI49-1/Dfslo8 fly, showing a similarphenotype to that shown in (B). (D–F). In situ hybridization showing RNA transcription of the fer gene in the third instar eye–antennal discs of wild-type(D), AC116/AC116 (E), and ferI49-1/ferI49-1 (F) larvae. Note the reduced RNA expression in the antennal primordium of the mutants (arrow), whereas thelevels of RNA signal in the eye are normal. (G–I) Antennal phenotype of Dll-GAL4/� (G), Dll-Gal4/�; ifer/� (H), and Dll-GAL4/�; ihern/� (I) flies.(G�–I�) Insets showing the base of the arista in detail. Note the presence of ectopic bristles in the third antennal segment of (H) and (I) (arrows) and theenlarged base of the arista (H�, I�) in the antennae carrying RNAi constructs. (J) Phenotype of a Dll-GAL4/�; ihern/ifer fly. There is a clear transformationof the distal antenna into leg structures, including many bracted bristles (arrowheads and inset). (K) Wild-type expression pattern of the Dan (Fer) proteinin the eye–antenna disc detect with an anti-Dan antibody (Emerald et al., 2003). (L) Expression of Dan (Fer) protein in a Dll-Gal4/�; ifer/� eye-antennadisc. Note the absence of Fer protein in the antennal primordium (a).
472 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

hern and fer expression patterns in the antennalprimordium
To delimit more precisely the expression of the hern andfer genes in the antennal primordium, we carried out doublestaining of the MD634 and CES115 GAL4 lines (revealinghern and fer transcription, respectively) with genes ex-pressed in restricted areas of mature eye–antennal discs, likeDll, hth, sal, and dac (Wagner-Bernholz et al., 1991; Dıaz-Benjumea et al., 1994; Mardon et al., 1994; Gorfinkiel et al.,1997; Rieckhof et al., 1997; Pai et al., 1998; Barrio et al.,1999; Dong et al., 2001). Both lines gave the same results.Within the third antennal segment, the hern and fer expres-sion is included within the Dll and hth domains (Fig. 3A–F).The hern and fer proximal limit of expression seems tocoincide with that of dac, and their distal limit of strongexpression in the third antennal segment with that of sal(Fig. 3G–L). We cannot exclude that the GAL4 lines driveexpression in a few cells from the second antennal segment.A scheme of the results obtained is shown in Fig. 3M.
Since hern and fer are not expressed in the leg discs, theymay be part of the mechanism that distinguishes legs fromantennae. The Antennapedia (Antp) Hox gene, which isexpressed in the leg but not in the antennal discs (Wirz etal., 1986; Condie et al., 1991), prevents hth expression andantenna formation in the leg primordia (Struhl, 1981;Casares and Mann, 1998). As expected, in Antp73b/� flies,which show a strong transformation of antennae into legs(Hazelrigg and Kaufman, 1983; Fig. 3N), the expressiondriven by the MD634 GAL4 line is clearly reduced (Fig.3O, compare with Fig. 3A or G).
Inactivation of the hern and fer genes partially transformsdistal antenna into leg
To characterize the function of hern and fer in normaldevelopment, we studied the phenotype of flies withouthern or fer activity. One of the P-GAL4 lines, AC116 (seeFig. 2), is mutant for the antennal function of the fer gene.Homozygous AC116 third instar larvae show no fer tran-scription in the antennal primordium but present normal ferexpression in the eye primordium (Fig. 4E, compare withthe wild-type expression in Fig. 4D). AC116/Df adults showone or more bristles in the third antennal segment, normallydevoid of them (Fig. 4B, compare with the wild-type an-tenna shown in Fig. 4A). To obtain more mutations in thehern and fer genes, we mobilized the MD634 and CES115P-GAL4 lines (the two closer to the hern and fer transcrip-tion units) to isolate imprecise excisions of the transposons.We isolated one w� derivative of the CES115 insertion (102analyzed), ferI49-1, which, in hemizygosis, shows a pheno-type very similar to that described for the AC116/Df adults(Fig. 4C). PCR analysis of the mutation revealed that it is asmall deletion of about 2.5 kb in the 5� upstream region ofthe fer transcription unit (Fig. 2A). Larvae homozygous forthe ferI49-1 mutation present reduced expression of the fer
RNA in the antennal primordium of mature eye–antennaldiscs, whereas the expression in the eye primordium isnormal (Fig. 4F). We conclude that the AC116 insertion andthe ferI49-1 deletion may have affected an antennal regula-tory region. This regulatory region would control only thefer gene, since in AC116 and ferI49-1 homozygous larvae,we cannot detect any change in hern expression. We havenot obtained any mutation for the hern gene (279 w� de-rivatives analyzed), although the MD634 line used for thispurpose is closer to the origin of transcription of the herngene than the CES115 line is to the transcription unit of thefer gene (see Fig. 2).
The weak phenotype of the AC116 and ferI49-1 mutationssuggested that there may be a partial functional redundancybetween hern and fer, and that the inactivation of both genesmay result in a stronger phenotype. To check this, to ascer-tain the phenotype of hern inactivation, and to study theeffect of the absence of Tintin products in the eye, we useddouble-stranded (ds)-mediated RNA interference (RNAi;Fire et al., 1998; Kennerdell and Carthew, 1998; Sharp,2001) to inactivate hern and fer functions. The inactivationin the antennal primordium of either the hern or the fergenes with a Dll-GAL4 (MD23) driver causes a phenotypesimilar to that described for the AC116/Df and ferI49-1/Dfadults: there is one or more bristles in the third antennalsegment and the base of the arista is slightly enlarged (Fig.4H and I, compare with the wild-type in Fig. 4G). However,when the ds-hern and ds-fer RNAs are induced together bythe Dll-GAL4 driver, a clear transformation of part of thedistal antenna into leg is observed: the third antennal seg-ment and the proximal arista are substantially enlarged andcovered with bristles; those in the base of the arista bearbracts, indicating a transformation into leg (Fig. 4J). Wechecked that the RNAi technique is working by doing in situhybridization with a hern probe not included in ihern or bystaining eye–antennal imaginal discs with a specific anti-body against the Fer product (Dan protein; Emerald andCohen, 2003). hern RNA expression is strongly reduced oreliminated in dpp-GAL4/�; ihern/� antennal discs (notshown) and no Fer protein is detected in Dll-Gal4/�; ifer/�antennal primordia (Fig. 4K and L). These results indicatethat both hern and fer are required to develop part of theantenna as opposed to leg and that these two genes arepartially redundant in this function.
The hern and fer genes produce antennal tissue andactivate hth, ss, and sal when ectopically expressed
We have shown that hern and fer are specifically ex-pressed in the eye–antennal disc and that both genes areneeded for normal antennal development. To test whetherthose genes are sufficient to induce eye or antennal devel-opment, we expressed them ectopically using the GAL4/UAS system (Brand and Perrimon, 1993). When either thehern or the fer genes are misexpressed in the leg discs withdpp-GAL4 or Dll-GAL4 (EM212) drivers, distal legs are
473M. Suzanne et al. / Developmental Biology 260 (2003) 465–483
474 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

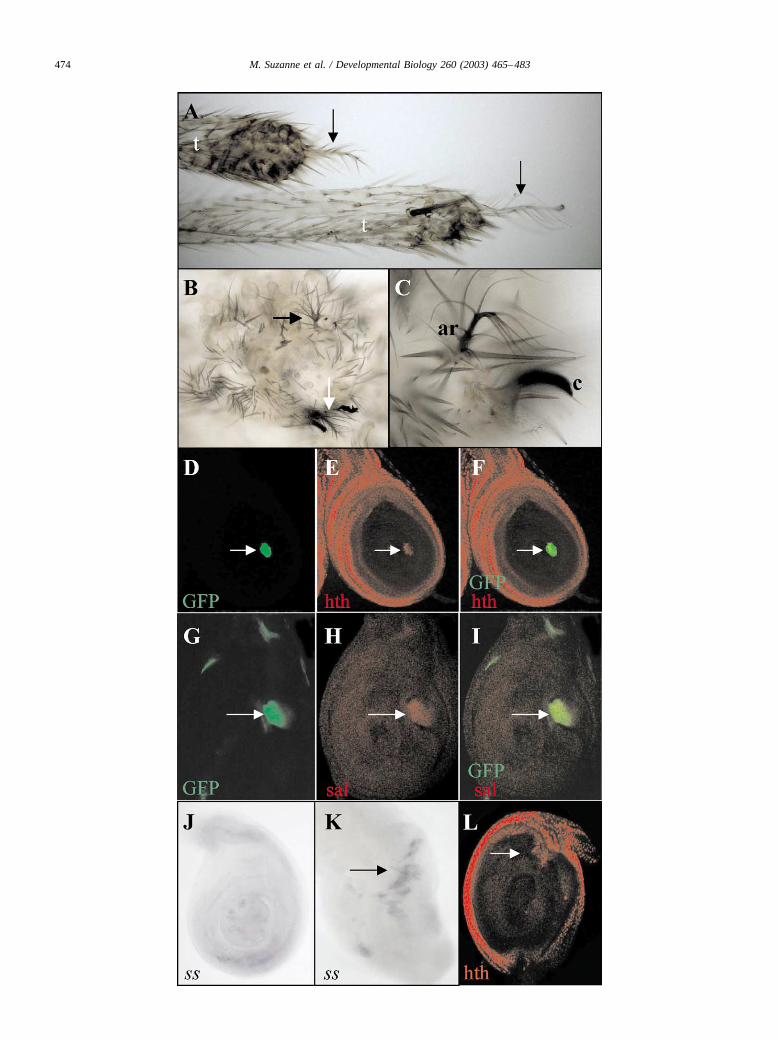
transformed to aristae (Fig. 5A–C). These transforma-tions are accompanied by the ectopic expression of hth,sal, and ss, three genes expressed in the antennal primor-dium but not in the distal region of mature wild-type legdisc (Wagner-Bernholz et al., 1991; Rieckhof et al.,1997; Pai et al., 1998; Casares and Mann, 1998; Duncanet al., 1998; Dong et al., 2002). Clones expressing eitherthe hern or the fer genes in the leg or wing disc havesmooth borders and frequently activate the sal and hthgenes cell-autonomously (Fig. 5D–I), but do not affectDll expression (not shown). In dpp-GAL4/UAS-fer orptc-GAL4/UAS-hern leg (or wing) discs, the expressionof ss is also activated (Fig. 5K, compare with the wild-type ss expression in Fig. 5J). Curiously, although ss isdownstream of hth in the antenna and leg (Dong et al.,2002), ectopic ss in the leg disc can also activate hth ina few cells (Fig. 5L).
Dll, hth, and ss are required for hern and fer expressionin the antenna
The hth or ss genes, together with Dll, are sufficient todevelop ectopic distal antennae when expressed in dif-ferent regions of the adult (Casares and Mann, 1998;Duncan et al., 1998; Dong et al., 2000). We have shownabove that the hern or fer genes are also able to elicit thistransformation in the leg and that they activate hth and ss.Conversely, when we induce high levels of the Hth or Ss
products in the leg discs, we find ectopic expression ofthe hern and fer genes (Fig. 6A–C; and data not shown).To study the interactions between these genes in normaldevelopment, we analyzed the relationship between Dll,hth, ss, and hern/fer in the antennal primordium. A re-duction of Hth activity using a dominant negative form ofhth (UAS-EN-HTH1-430; Inbal et al., 2001) results in adecreased activity of the MD634 and AC116 GAL4 lines,which reveal hern and fer expression, respectively (Fig.7B; and data not shown, compare with the wild-typeexpression in Fig. 7A). Similarly, in antennal discs of aDll strong hypomorph or a ss null mutation, the expres-sion of hern and fer disappears (Fig. 7C and D; and datanot shown, compare with the wild-type expression in Fig.1G or 4D). These results suggests that hth, Dll, and ss arerequired to maintain hern and fer expression in the an-tenna. By contrast, high levels of hern or fer may reducehth expression. In dpp-GAL4/UAS-fer or dpp-GAL4/UAS-hern larvae, the expression of hth (and sal) in thethird antennal segment is eliminated or strongly reduceddorsally (where levels of hern and fer are high) and doesnot change or is ectopically activated ventrally (wherelevels of hern and fer are low; Fig. 7E and F; and data notshown). Similarly, fer-expressing clones are able todownregulate hth expression in the antennal primordium(Fig. 7G and H). These results suggest that levels of hernand fer expression may be important for a normal anten-nal development.
Fig. 6. Hth and ss activate the hern and fer genes ectopically. (A, B) Leg disc of a MD634 UAS-GFP/ hs-hth larva, after heat shock. There are scattered cellsshowing ectopic hth expression (in red, A) and also ectopic activation of GFP expression in small patches (in green, B). (C) dpp-GAL4/UAS-ss leg disc,showing ectopic expression of the fer transcript.
Fig. 5. The ectopic expression of either the hern or fer genes transforms leg into antenna and activates the ss, hth, and sal genes. (A) Distal legs of a Dll-GAL4(EM212)/UAS-hern fly, showing transformation to aristae (arrows); t, tibia. (B, C) Transformation of distal legs to aristae (arrows in B) in a dpp-GAL4/UAS-fer fly. A detail of an ectopic arista (ar) is shown in (C), close to a claw (c). (D–F) Clone expressing the hern gene in the leg disc, marked by the GFPmarker (D, in green), which activates ectopically hth (red in E). Merged image in (F). (G–I) A similar clone, marked with GFP (G), also activatescell-autonomously the gene sal (red in H). Merged image in (I). (J) Wild-type third instar leg disc hybridized with a ss probe, showing almost no expressionin the central region (Duncan et al., 1998). (K) dpp-GAL4/UAS-fer leg disc, showing abnormal development and ectopic ss transcription (arrow). (L)dpp-GAL4/UAS-ss leg disc. A few cells of the dorsal part of the disc activate hth ectopically (arrow).
475M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

The hern and fer genes are required for normal eyedevelopment and form eye tissue and activate ey whenectopically expressed
To study the possible role of the hern and fer genes ineye development, we looked to the eye phenotype when
either the hern or fer genes are inactivated by RNAi andalso express them ectopically. Expression of ds-hern ords-fer RNA in the eye primordium with a GMR-GAL4driver causes a slightly rough eye, with some bristles irreg-ularly positioned (Fig. 8A–D). Curiously, the phenotype isnot increased if the ds-hern and ds-fer RNAs are induced inthe same fly. Misexpression experiments also suggest thatboth hern and fer are involved in eye development. Thus,the expression of either hern or fer with different GAL4drivers causes the appearance of ectopic eye tissue in thethird antennal segment or rostral membrane (Fig. 8E–G).These transformations are accompanied by the ectopic ex-pression of ey (Fig. 8H and I), although this effect may alsoindicate the maintenance of a previous ey expression. Con-versely, the misexpression of ey activates the hern and fergenes ectopically (Fig. 8J; and data not shown). Both hernand fer also activate embryonic lethal abnormal vision(elav), a marker of neuronal differentiation (Robinow et al.,1988), when ectopically expressed (Fig. 8I; and data notshown). The analysis of clones expressing the fer gene inthe leg, eye–antennal, or wing discs shows that elav activa-tion is strictly nonautonomous, and only occurs in somecells adjacent to some of these clones (Fig. 8K–N).
The formation of the morphogenetic furrow in the eye islimited laterally by wg signaling (Ma and Moses, 1995;Treisman and Rubin, 1995). We have observed that hernand fer expression within the eye primordium includes themore lateral wg-expressing regions (Fig. 9A). Interestingly,we find that both hern and fer activate wg transcriptionwhen ectopically expressed. In ptc-GAL4/UAS-hern ordpp-GAL4/UAS-fer flies, the wings show several alter-ations, including the appearance of marginal bristles in themiddle of the wing blade (Fig. 9B and C). This phenotypeis characteristic of ectopic wg signaling (Dıaz-Benjumeaand Cohen, 1995), and in fact, wg is ectopically expressedin the wing discs of these larvae (Fig. 9D; and data notshown). Clones expressing the fer genes in the eye–antenna,leg, or wing discs also show induction of wg, mostly withinbut also outside the clone (Fig. 9E, F, and H). We havepreviously described that the elav gene is also inducednonautonomously by these clones. We note, however, thatcells ectopically expressing elav do not coincide with thoseexpressing wg (Fig. 9G and H) and that this reproduces thewild-type situation in the eye.
Interaction of hern and fer genes with N signaling
Signaling pathways can modify the activity of selectorgenes and are needed for proper organ formation. N signal-ing, for instance, is needed for eye formation (Go et al.,1998; Kurata et al., 2000; Kumar and Moses, 2001) and canactivate ey when ectopically activated (Kurata et al., 2000).Moreover, N has been recently implicated in the decision ofmaking eye or antenna, directing eye development, andsuppressing antenna formation (Kurata et al., 2000; Kumarand Moses, 2001). Therefore, we investigated whether N
Fig. 7. Regulatory activations between Dll, ss, hth, and hern/fer genes inthe antennal disc. (A) Antennal primordium of a MD634 UAS-GFP larva,showing GFP expression (in green). (B) Similar antennal disc of a MD634UAS-GFP/UAS-EN-HTH1-430 larva. The GFP signal is now much reducedcompared with that in (A). (C) Dll3 antennal disc hybridized with a ferprobe, showing there is no fer transcription in the antennal primordium (a),but normal expression in the eye. (D) Antennal primordium in a ssD115.7
mutant larva. The expression of fer in the antenna (a) also disappears. (E,F) Antennal primordium of a dpp-GAL4/UAS-fer third instar larva, show-ing ectopic activation of hth (red in E) and sal (green in F) in the ventraldistal part of the third antennal segment (arrowheads) and repression ofboth genes in the dorsal part (arrows). (G, H) Antennal primordium of athird instar imaginal disc carrying clones marked with GFP (in green in H,arrows) which ectopically activate the fer gene. These clones cell-auton-omously repress hth expression (arrows; hth marked in red in G and H).
476 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

signaling could alter the ey and elav expression induced bythe Tintin genes. As shown in Fig. 9I–K, the coexpression ofthe hern gene and a dominant negative form of the Notchreceptor (Klein et al., 1997) substantially reduces ey andeliminates elav ectopic signals (compare with Fig. 8H andI). Accordingly, no ectopic eyes are formed in this geneticcombination. This indicates that the effect of hern on eyexpression and eye formation requires N signaling.
Discussion
We have isolated and characterized two adjacent genes,hern and fer, which share several characteristics: first, theyencode similar protein sequences, including a Psq motif;second, they are expressed in the eye–antennal imaginaldiscs, and not in other discs, in a very similar or identicalpattern; third, they are needed for normal antenna and eyedevelopment; and fourth, both genes can make eyes andantennae when ectopically expressed. Our genetic analysisindicates that the specific role of hern and fer is to contrib-ute to antenna and eye development and that hern and fergenes are partially redundant, at least as to their function inthe antenna. The role of these same genes in the antenna hasbeen recently characterized (Emerald et al., 2003). Theseauthors call these genes distal antenna (dan; the fer gene)and distal antenna-related (danr; the hern gene; see Fig. 2)and they have reached similar conclusions. Particularly, adeficiency for both genes results in an antennal transforma-tion similar to what we see by inactivating both hern and fergenes by RNAi (Emerald et al., 2003).
The Psq motif is present in at least 14 Drosophila pro-teins, including those encoded by the pipsqueak, Bric a bracI and II, Piefke, E93, tyrosine-kinase related, and ribbongenes (Baehrecke and Thummel, 1995; Weber et al., 1995;Horowitz and Berg, 1996; Lehmann et al., 1998; Haller etal., 1987; Bradley and Andrew, 2001; Shim et al., 2001;Siegmund and Lehmann, 2002; Couderc et al., 2002). Basedon sequence similarity and other characteristics, proteinsincluding Psq domains in distinct species can be classifiedinto different groups (Siegmund and Lehmann, 2002). ThePsq motif of the hern and fer genes (as well as that of theCG13496 predicted gene) shows higher similarity to thoseincluded in the products of the human centromere protein Band the human predicted protein CAB66474 (Siegmund andLehmann, 2002). This motif is a helix–turn–helix DNA-binding domain (Lehmann et al., 1998), structurally similarto the DNA-binding domain of the Hin and other recombi-nases (Lehmann et al., 1998; Siegmund and Lehmann,2002). This suggests that hern and fer function as transcrip-tion regulators.
hern and fer specify distal antenna
The concept of selector genes in Drosophila has evolvedfrom a precise and restricted definition (Garcıa-Bellido,
1975) to a more loose interpretation (Mann and Morata,2000). Selector, or selector-like genes, are now consideredas those required to make a particular structure and able toform it in different positions when the gene is expressedectopically. hern and fer fit this definition as selector genesfor the distal antenna. They also can make ectopic eyes,although their requirement for eye development is not soevident as that for antenna formation.
The differentiation of legs or antennae depends on theactivity of the hth and Antp genes (Struhl, 1981; Casares andMann, 1998). The ss gene, however, is also able to trans-form distal leg (and also maxillary palp and rostral mem-brane) into distal antenna (Duncan et al., 1998), and theabsence of ss, like that of hth, transforms antenna into leg(Struhl, 1982; Burgess and Duncan, 1990; Casares andMann, 1998; Pai et al., 1998). Although ss seems to bedownstream of Dll and hth in antenna specification (Duncanet al., 1998; Dong et al., 2001; I. Duncan, cited in Dong etal., 2002), we have shown that ectopic ss can activate hth insome cells of the leg disc. Similarly, misexpression of ss inthe rostral membrane induces Dll expression (Duncan et al.,1998). It seems, therefore, that ss can trigger an antennalgenetic program when misexpressed in certain places.
The hern and fer genes are transcribed, at the late thirdinstar stage, in the third antennal segment and the arista.When the activity of these two genes is impaired, the distalantenna is partially transformed into a leg. This transforma-tion is barely detected when only one of the genes is notactive, implying that they perform a partially redundantrole. In fact, both the expression and protein sequence ofHern and Fer are very similar. When either the hern or fergenes are ectopically expressed in the leg disc, they directdistal antennal development and activate antennal genes likehth, ss, and sal. Therefore, the fer and hern genes are bothrequired and sufficient to make part of the distal antenna.
Interactions of Dll, hth, and ss with hern and fer
Four different genes, hth, ss, hern, and fer, are able toform distal antenna, together with Dll, when ectopicallyexpressed (Casares and Mann, 1998; Duncan et al., 1998;Dong et al., 2000; Emerald et al., 2003; this report). Theirmutual regulation seems to differ when misexpressed in theleg disc or when normally expressed in the antennal pri-mordium. In the leg disc, hern or fer activates hth and ssand, reciprocally, hth and ss induce hern and fer expression.Moreover, even ss can promote hth transcription, althoughjust in a few cells. Taken together, our results suggest thatthe four genes can form distal antenna by activating eachother’s transcription when ectopically expressed (Fig. 10).
In the third antennal segment, Dll, hth, and ss are re-quired to activate hern/fer expression. Since ss is down-stream of Dll and hth in the antenna (Duncan et al., 1998;Dong et al., 2002), the activation of hern/fer by Dll and hthcould be mediated by ss. We note, however, that the levelsof hern and fer may modulate hth expression. Moderately
477M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

478 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

increased levels of fer can activate hth in dpp-GAL4/UAS-fer discs but, when the levels of hern or fer in the antennaare highly increased, the transcription of hth is prevented.These results suggest that the total amount of hern and ferexpression may be regulated in the antennal primordium.Accordingly, in clones mutant for danr (hern), the expres-sion of dan (fer) is upregulated (Emerald et al., 2003). Alsosupporting the conclusion that levels of hern and fer have tobe regulated, we have found that, in ey-GAL4/UAS-hern orey-GAL4/UAS-fer flies, where levels of either hern or ferare highly increased in the eye–antennal disc, both the eyeand the antenna disappear (not shown).
hern and fer specify eye development
Several eye-specifying genes have been identified, andthey fulfill two conditions: they are required to make the eyeand they can form ectopic eyes when expressed in differentparts of the body (Treisman, 1999; Kumar, 2001). The hernand fer genes probably form part of this network of “eye-specification” genes: first, they are expressed in the eyeprimordium, with higher levels of expression anterior to themorphogenetic furrow; second, they activate ey and elavand make ectopic eyes when expressed ectopically; finally,ey also activates the hern and fer genes when ectopicallyexpressed. A recent report has also identified the hern andfer genes as downstream of ey in eye ectopic formation(Michaut et al., 2003). We observe, however, that the inac-tivation of both hern and fer genes by RNAi with the GAL4driver does not grossly affect eye development, like mutantsin the eye-specification genes do (Treisman, 1999; Kumar,2001). The nonautonomous induction of elav when hern orfer are ectopically expressed reproduces the wild-type situ-ation, in which high levels of hern and fer are observedadjacent to the differentiating, elav-expressing, photorecep-tor cells. Another similarity of hern and fer with some of the“eye-specification” genes is that we have obtained ectopiceye tissue in the antennae. The eye-specification genes eyaand dac also form eyes predominantly, when ectopicallyexpressed, in this same position (Bonini et al., 1997; Pig-noni et al., 1997; Shen and Mardon, 1997). This is perhapsdue to ey being expressed in the antennal primordium in lateembryos (Quiring et al., 1994; Kumar and Moses, 2001),thus providing a favorable genetic context for eye forma-tion. In accordance, when we express either the hern or the
fer gene ectopically, we only detect ectopic ey expression inthe antennal disc. We also obtain eyes in the rostral mem-brane when ectopically expressing the fer gene. This may bedue to the absence of hth, since high levels of either hern orfer repress hth and removal of this gene in the rostralmembrane forms ectopic eyes (Pai et al., 1998; Pichaud andCasares, 2000).
hern and fer genes and the decision to make eye orantenna
The hern and fer genes can form ectopic aristae and eyetissue, but only in a limited number of regions of the adultcuticle. This is similar to what happens with other genesmaking ectopic antennae (hth, ss) or eye (eye-specificationgenes) (Halder et al., 1995; Bonini et al., 1997; Chen et al.,1997; Pignoni et al., 1997; Shen and Mardon, 1997; Casaresand Mann, 1998; Duncan et al., 1998; Czerny et al., 1999;Dong et al., 2000; Seimiya and Gehring, 2000). This is dueto the particular developmental context of the region wherethe genes are ectopically activated (Chen et al., 1999; Donget al., 2000).
We have observed transformations of third antennal seg-ment, where hern and fer are normally transcribed, to eyetissue, in Dll-GAL4/UAS-hern or dpp-Gal4/UAS-fer flies.This suggests that the levels of Hern and Fer products maybe important in inducing or maintaining ey expression anddistinguishing eye from antenna. Accordingly, when Hernor Fer products are increased in the antennal primordium,the expression of hth, an inhibitor of eye development (Paiet al., 1998), is eliminated. We also note that, in the wild-type eye–antennal discs, hern and fer show higher levels ofexpression in the eye primordium than in the antennal one,where these genes are coexpressed with hth. However, theamount of Tintin products is not the only factor in thisdistinction, since, for instance, in Dll-GAL4/UAS-hern eye-antennal discs, the area of ectopic ey transcription in theantenna is smaller than the area of hern overexpression. Theactivity of other genes will probably contribute to the for-mation of either eye or antenna. Thus, the ectopic expres-sion of either hern or fer induces wg, an inhibitor of mor-phogenetic furrow formation (Ma and Moses, 1995;Treisman and Rubin, 1995), and this probably limits theplaces where the eye can develop.
Two recent models have been proposed to explain the
Fig. 8. The inactivation of hern and fer genes alters eye development and the ectopic expression of the hern or fer genes transforms parts of the head intoeye tissue and activates ey and elav. (A) Wild-type eye. (B) Slightly rough eye in a GMR-GAL4; ifer/� fly. (C) Detail of eye bristles in a wild-type eye.(D) Similar detail in a GMR-GAL4; ifer/� fly. Note that the bristles are irregularly positioned (arrows). (E, F) Antennae of a Dll-GAL4 (MD23)/UAS-hernfly: ectopic eye tissue appears in the third antennal segment (arrows in E). A detail of the transformation is shown in (F). (G) A similar ectopic eye (arrow)is obtained when the fer gene is expressed under dpp-GAL4 control. Symbols in (E–G) are as in Fig. 1. (H, I) Eye–antennal disc of a Dll-GAL4/UAS-hernlarva, showing ectopic expression of elav (red in I, arrows) and ey (H, I, green; arrow in H). Note that, as in wild-type discs, ey and elav expression do notcoincide. The asterisks in (H) and (I) mark the normal ey and elav expression. a, antennal primordium. (J) Leg disc of a dpp-GAL4/UAS-ey larva, showingectopic activation of the fer gene. (K) Eye–antennal imaginal disc with clones ectopically expressing the fer gene, marked with GFP (green), and activatingelav (in red). (L–N) Detail of the clone boxed in (K), showing the nonautonomous induction of elav (red, M) in one side of the clone expressing fer (greenin L). (N) Merged image.
479M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

Fig. 9. Ectopic expression of the hern or fer genes activates wg, and N signaling is required for eye formation by the hern gene. (A) Eye primordium of awg-lacZ larva hybridized with a probe of the fer gene, showing coexpression of wg and fer in the lateral regions (squares); detail of this overlap in the inset.(B) Wing of a dpp-GAL4/UAS-fer fly showing the ectopic triple row of bristles in the middle of the wing (arrow), a partial duplication of the wing blade(arrowhead), and a greatly enlarged hinge. (C) Detail of the ectopic triple row appearing in the middle of the wing blade. (D) Wing disc of the same genotypestained with an anti-Wg antibody (red). There is ectopic wg expression (arrow) in the dpp domain in addition to the normal wg signal (arrowhead). (E–H)Clone ectopically expressing the fer gene in the wing disc (in green, marked with GFP in E). This expression causes induction of wg expression (red in F)mostly within the clone, whereas elav induction is strictly cell-nonautonomous (marked in blue in G). (H) Merged image. (I–K) Eye–antennal disc of aDll-GAL4/UAS-Necd UAS-hern larva. The ectopic expressions of elav (red in I) and ey (green in J, arrow) are substantially reduced when compared withthose observed in Dll-GAL4/UAS-hern larvae (Fig. 8H and I). The asterisks in (I) and (J) indicates normal elav and ey expression. Merged image in (K).
480 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

specification of eye and antenna within the eye–antennaldisc. Both models suggest that the activation of the Nsignaling pathway is a key element in this process. Kurata etal. (2000) suggest that N signaling activates both ey and Dllin the eye and antennal primordia; subsequently, ey re-presses Dll in the eye and perhaps the hth and extradenticlegenes repress ey in the antenna. In this way, the exclusiveexpression of ey (in the eye) and Dll and hth (in the antenna)determine eye and antenna identity, respectively. Kumarand Moses (2001) propose that the N and Egfr signalingpathways (together with the hedgehog and wg genes) areinstrumental in the decisions to make eye or antenna (Ku-mar and Moses, 2001). According to these authors, N sig-naling promotes eye development and prevents formation ofthe antenna, whereas Egfr signaling does the opposite. Wehave found that ectopic expression of either hern or fer inthe antenna induces ectopic eyes and activates ey and elav,but that the coexpression of hern and an N dominant-negative protein does not result in ectopic eyes and almosteliminates ey and elav activation. This suggests that Nfunction impinges on hern activity to form ectopic eyes(Fig. 10). As in other cases, the combined activity of sig-naling pathways and selector genes determine the specifi-cation of different structures.
Acknowledgments
We thank G. Morata for his help and encouragementthroughout the work and for the GAL4 lines; S. Cohen forantibodies, stocks, and for communicating results beforepublication; J.F. de Celis and G. Morata for comments onthe manuscript; N. Azpiazu for the gift of the ss probe; A.Cantarero for providing the AC116 line; R. Gonzalez for
preparation of the adult cuticles; and N. Azpiazu, R. Barrio,S. Cohen, D. Cribbs, I. Duncan, Gines Morata, A. Preiss,A. Salzberg, R. Schuh, H. Sun, and U. Walldorf for theantibodies, probes, and stocks. M.S. is recipient of an Eu-ropean Marie Curie fellowship and M.C. is recipient of aRamon y Cajal contract from the Spanish Ministerio deCiencia y Tecnologıa. This work has been supported bygrants from the Direccion General de Investigacion Cientı-fica y Tecnica (N° PB98-0510) and the Comunidad Au-tonoma de Madrid (N° 08.9/0003/98 and 08.1/0031/2001.1)(to E.S.), and an Institutional Grant from the FundacionRamon Areces to the Centro de Biologıa Molecular SeveroOchoa.
References
Abu-Shaar, M., Mann, R., 1998. Generation of multiple antagonistic do-mains along the proximodistal axis during Drosophila leg develop-ment. Development 125, 3821–3830.
Adams, M.D., Celniker, S.E., Holt, R.A., Evans, C.A., Gocayne, J.D.,Amanatides, P.G., Scherer, S.E., Li, P.W., et al., 2000. The genomesequence of Drosophila melanogaster. Science 287, 2185–2195.
Affolter, M., Percival-Smith, A., Muller, M., Billeter, M., Qian, Y.Q.,Otting, G., Wutrich, K., Gehring, W.J., 1991. Similarities between thehomeodomain and the Hin recombinase DNA-binding domain. Cell 64,879–880.
Artavanis-Tsakonas, S., Rand, M.D., Lake, R.J., 1999. Notch signaling:cell fate control and signal integration in development. Science 284,770–776.
Ashburner, M., 1989. Drosophila: A Laboratory Manual. Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY.
Atkinson, N.S., Robertson, G.A., Ganetzky, B., 1991. A component ofcalcium-activated potassium channels encoded by the Drosophila slolocus. Science 253, 551–555.
Azpiazu, N., Frasch, M., 1993. tinman and bagpipe: two homeo box genesthat determine cell fates in the dorsal mesoderm of Drosophila. GenesDev. 7, 1325–1340.
Fig. 10. Model of activity of the hern and fer genes. When expressed ectopically in the leg disc, hth, ss, hern, and fer activate one another (Dong et al., 2002;and this report), although activation of hth by ss is limited to a few cells. In the eye–antennal disc, the N and Egfr signaling pathways (Kumar and Moses,2001) determine whether eye or antennal development follows. In the antennal primordium, hth, together with Dll, activates hern/fer through ss. In the eyeprimordium, hern and fer expression probably depends on the eye-specification gene group.
481M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

Azpiazu, N., Morata, G., 2000. Function and regulation of homothorax inthe wing imaginal disc of Drosohila. Development 127, 2685–2693.
Azpiazu, N., Morata, G., 2002. Distinct functions of homothorax in legdevelopment in Drosophila. Mech. Dev. 119, 55–67.
Baehrecke, E.H., Thummel, C.S., 1995. The Drosophila E93 gene from the93F early puff displays stage- and tissue-specific regulation by 20-hydroxyecdysone. Dev. Biol. 171, 85–97.
Barrio, R., de Celis, J.F., Bolshakov, S., Kafatos, F.C., 1999. Identificationof regulatory regions driving the expression of the Drosophila spaltcomplex at different developmental stages. Dev. Biol. 215, 33–47.
Bonini, N.M., Bui, Q.T., Gray-Board, G.L., Warrick, J.M., 1997. TheDrosophila eyes absent gene directs ectopic eye formation in a pathwayconserved between fly and vertebrates. Development 124, 4819–4826.
Bradley, P.L., Andrew, D.J., 2001. ribbon encodes a novel BTB/POZprotein required for directed cell migration in Drosophila melano-gaster. Development 128, 3001–3015.
Brand, A., Perrimon, N., 1993. Targeted gene expression as a means ofaltering cell fates and generating dominant phenotypes. Development118, 401–415.
Brook, W.J., Cohen, S.M., 1996. Antagonistic interactions between wing-less and decapentaplegic responsible for dorsal–ventral pattern in theDrosophila leg. Science 273, 1373–1377.
Brook, W.J., Dıaz-Benjumea, F.J., Cohen, S.M., 1996. Organizing spatialpattern in limb development. Annu. Rev. Cell Biol. 12, 161–180.
Burgess, E.A., Duncan, I., 1990. Direct control of antennal identity by thespineless-aristapedia gene of Drosophila. Mol. Gen. Genet. 221, 347–352.
Calleja, M., Moreno, E., Pelaz, S., Morata, G., 1996. Visualization of geneexpression in living adult Drosophila. Science 274, 252–255.
Campbell, G., Tomlinson, A., 1995. Initiation of the proximodistal axis ininsect legs. Development 121, 619–628.
Campbell, G., Tomlinson, A., 1998. The roles of the homeobox genesaristaless and Distal-less in patterning the legs and wings of Drosoph-ila. Development 125, 4483–4493.
Casares, F., Mann, R.S., 1998. Control of antennal versus leg developmentin Drosophila. Nature 392, 723–726.
Chen, R., Amoui, M., Zhang, Z., Mardon, G., 1997. Dachsund and eyesabsent proteins form a complex and function synergistically to induceectopic eye development in Drosophila. Cell 91, 893–904.
Chen, R., Halder, G., Zhang, Z., Mardon, G., 1999. Signaling by theTGF-� homolog decapentaplegic functions reiteratively within thenetwork of genes controlling retinal cell fate determination in Drosoph-ila. Development 126, 935–943.
Cohen, S.M., Jurgens, G., 1989. Proximal–distal pattern formation inDrosophila: graded requirements for Distal-less gene activity duringlimb development. Roux’s Arch. Dev. Biol. 198, 157–169.
Condie, L.M., Mustard, J.A., Brown, D.L., 1991. Generation of anti-Antennapedia monoclonal antibodies and Antennapedia protein expres-sion in imaginal discs. Dros. Inf. Serv. 70, 52–54.
Couderc, J.-L., Godt, D., Zollman, S., Chen, J., Li, M., Tiong, S., Cramton,S.E., Sahut-Barnola, I., Laski, F., 2002. The bric a brac locus consistsof two paralogous genes encoding BTB/POZ domain proteins and actsas a homeotic and morphogenetic regulator of imaginal development inDrosophila. Development 129, 2419–2433.
Czerny, T., Halder, G., Kloter, U., Souabni, A., Gehring, W.J., Busslinger,M., 1999. twin of eyeless, a second Pax-6 gene of Drosophila, actsupstream of eyeless in the control of eye development. Mol. Cell. 3,297–307.
de Celis, J.F., Barrio, R., Kafatos, F.C., 1996. A gene complex actingdownstream of dpp in Drosophila wing morphogenesis. Nature 381,421–424.
Dıaz-Benjumea, F., Cohen, B., Cohen, S.M., 1994. Cell interaction be-tween compartments establishes the proximal–distal axis of Drosoph-ila legs. Nature 372, 175–178.
Dıaz-Benjumea, F.J., Cohen, S.M., 1995. Serrate signals through Notch toestablish a Wingless-dependent organizer at the dorsal/ventral compart-ment boundary of the Drosophila wing. Development 121, 4215–4225.
Dong, P.D.S., Chu, J., Panganiban, G., 2000. Co-expression of the ho-meobox genes Distal-less and homothorax determines Drosophila an-tennal identity. Development 127, 209–216.
Dong, P.D.S., Chu, J., Panganiban, G., 2001. Proximodistal domain spec-ification and interactions in developing Drosophila appendages. De-velopment 128, 2365–2372.
Dong, P.D.S., Scholz Dicks, J., Panganiban, G., 2002. Distal-less andhomothorax regulate multiple targets to pattern the Drosophila antenna.Development 129, 1967–1974.
Duncan, D.M., Burgess, E., Duncan, I., 1998. Control of distal antennalidentity and tarsal development in Drosophila by spineless-aristapedia,a homolog of the mammalian dioxin receptor. Genes Dev. 12, 1290–1303.
Emerald, B.S., Curtiss, J., Mlodzik, M., Cohen, S.M., 2003. distal antennaand distal antenna related encode nuclear proteins containingpipsqueak motifs involved in antenna development in Drosophila.Development 130, 1171–1180.
Fire, A., Xu, S., Montgomery, M.K., Kostas, S.A., Driver, S.E., Mello,C.C., 1998. Potent and specific genetic interference by double-strandedRNA in Caenorhabditis elegans. Nature 391, 806–811.
Garcıa-Bellido, A., 1975. Genetic control of wing disc development inDrosophila. Ciba Found. Symp. 29, 161–182.
Gellon, G., McGinnis, W., 1998. Shaping animal body plans in develop-ment and evolution by modulation of Hox expression patterns. Bioes-says 20, 116–125.
Go, M.J., Eastman, S., Artavanis-Tsakonas, S., 1998. Cell proliferationcontrol by Notch signaling in Drosophila development. Development125, 2031.
Gonzalez-Crespo, S., Abu-Shaar, M., Torres, M., Martinez-A, C., Mann,R.S., Morata, G., 1998. Antagonism between extradenticle functionand Hedgehog signalling in the developing limb. Nature 394, 196–200.
Gorfinkiel, N., Morata, G., Guerrero, I., 1997. The homeobox gene Distal-less induces ventral appendage development in Drosophila. Genes Dev11, 2259–2271.
Halder, G., Callaerts, P., Gehring, W.J., 1995. Induction of ectopic eyes bytargeted expression of the eyeless gene in Drosophila. Science 267,1788–1792.
Haller, J., Cote, S., Bronner, G., Jackle, H., 1987. Dorsal and neuralexpression of a tyrosine kinase-related Drosophila gene during embry-onic development. Genes Dev. 1, 862–867.
Hazelrigg, T., Kaufman, T.C., 1983. Revertants of dominant mutationsassociated with the Antennapedia gene complex of Drosophila mela-nogaster, in: Malacinski, G., Klein, W. (Eds.), Molecular Aspects ofEarly Development, Plenum Press, New York, pp. 189–218.
Hinz, U., Giebel, B., Campos-Ortega, J.A., 1994. The basic helix–loop–helix domain of Drosophila lethal of scute protein is sufficient forproneural function and activates neurogenic genes. Cell 14, 77–87.
Horowitz, H., Berg, C.A., 1996. The Drosophila pipsqueak gene encodesa nuclear BTB-domain-containing protein required early in oogenesis.Development 122, 1859–1871.
Inbal, A., Halachmi, N., Dibner, C., Frank, D., Salzberg, A., 2001. Geneticevidence for the transcriptional-activating function of Homothoraxduring adult fly development. Development 128, 3405–3413.
Ito, K., Awano, W., Suzuki, K., Hiromi, Y., Yamamoto, D., 1997. TheDrosophila mushroom body is a quadruple structure of clonal unitseach of which contains a virtually identical set of neurons and glialcells. Development 124, 761–771.
Kennerdell, J.R., Carthew, R.W., 1998. Use of ds-RNA-mediated geneticinterference to demonstrate that frizzled and frizzled 2 act in the Wing-less pathway. Cell 95, 1017–1026.
Klein, T., Brennan, K., Martınez-Arias, A., 1997. An intrinsic dominantnegative activity of Serrate that is modulated during wing developmentin Drosophila. Dev. Biol. 186, 123–134.
Kuhnlein, R.P., Frommer, G., Friedrich, M., Gonzalez-Gaitan, M., Weber,A., Wagner-Bernholz, J.F., Gehring, W.J., Jackle, H., Schuh, R., 1994.spalt encodes an evolutionarily conserved zinc finger protein of novel
482 M. Suzanne et al. / Developmental Biology 260 (2003) 465–483

structure which provides homeotic gene function in the head and tailregion of the Drosophila embryo. EMBO J. 13, 168–179.
Kumar, J.P., 2001. Signaling pathways in Drosophila and vertebrate retinaldevelopment. Nat. Rev. 2, 846–857.
Kumar, J.P., Moses, K., 2001. EGF receptor and Notch signaling actupstream of eyeless/Pax6 to control eye specification. Cell 104, 687–697.
Kurata, S., Go, M.J., Artavanis-Tsakonas, S., Gehring, W.J., 2000. Notchsignaling and the determination of appendage identity. Proc. Natl.Acad. Sci. USA 97, 2117–2122.
Lecuit, T., Cohen, S.M., 1997. Proximal–distal axis formation in theDrosophila leg. Nature 388, 139–145.
Lehmann, M., Siegmund, T., Lintermann, K.-G., Korger, G., 1998. ThePipsqueak protein of Drosophila melanogaster binds to GAGA se-quences through a novel DNA-binding domain. J. Biol. Chem. 273,28504–28509.
Ma, C., Moses, K., 1995. wingless and patched are negative regulators ofthe morphogenetic furrow and can affect tissue polarity in the devel-oping Drosophila compound eye. Development 121, 2279–2289.
Mann, R.S., Morata, G., 2000. The developmental and molecular biologyof genes that subdivide the body of Drosophila. Annu. Rev. Coll Dev.Biol. 16, 243–271.
Mardon, G., Solomon, N.M., Rubin, G.M., 1994. dachsund encodes anuclear protein required for normal eye and leg development in Dro-sophila. Development 120, 3473–3486.
Michaut, L., Flister, S., Neeb, M., White, K.P., Certa, U., Gehring, W.J.,2003. Analysis of the eye developmental pathway in Drosophila usingDNA microarrays. Proc. Natl. Acad. Sci. USA 100, 4024–4029.
Morata, G., 2001. How Drosophila appendages develop. Nat. Rev. Mol.Cell Biol. 2, 89–97.
Nagel, A.C., Maier, D., Preiss, A., 2002. Green fluorescent protein as aconvenient and versatile marker for studies on functional genomics inDrosophila. Dev. Genes Evol. 212, 93–98.
Pai, C.-Y., Kuo, T., Jaw, T., Kurant, E., Chen, C., Bessarab, D., Salzberg,A., Sun, Y., 1998. The homothorax homeoprotein activates the nuclearlocalization of another homeoprotein, Extradenticle, and suppresseseye development in Drosophila.. Genes Dev. 12, 435–436.
Pichaud, F., Casares, F., 2000. Homothorax and iroquois-C genes arerequired for the establishment of territories within the developing eyedisc. Mech. Dev. 96, 15–25.
Pignoni, F., Hu, B., Zavitz, K.H., Xiao, J., Garrity, P.A., Zipurski, S.L.,1997. The eye specification proteins so and eya form a complex andregulate multiple steps in Drosophila eye development. Cell 91, 881–892.
Pignoni, F., Zipurski, S., 1997. Induction of Drosophila eye developmentby Decapentaplegic. Development 124, 271–278.
Postlethwait, J.H., Schneiderman, H.A., 1971. Pattern formation and de-termination in the antenna of the homeotic mutant Antennapedia ofDrosophila melanogaster. Dev. Biol. 25, 606–640.
Quiring, R., Walldorf, U., Kloter, U., Gehring, W.J., 1994. Homology ofthe eyeless gene of Drosophila to the Small eye gene in mice andAniridia in humans. Science 265, 785–789.
Rieckhof, G., Casares, F., Ryoo, H.D., Abu-Shaar, M., Mann, R.S., 1997.Nuclear translocation of Extradenticle requires homothorax, which
encodes an Extradenticle-related homeodomain protein. Cell 91, 171–183.
Robinow, S., Campos, A.R., Yao, K.M., White, K., 1988. The elav geneproduct of Drosophila, required in neurons, has three RNP consensusmotif. Science 242, 1570–1572.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: ALaboratory Manual. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
Seimiya, M., Gehring, W.J., 2000. The Drosophila homeobox gene optix iscapable of inducing ectopic eyes by an eyeless-independent mecha-nism. Development 127, 1879–1886.
Sharp, P.A., 2001. RNA interference 2001. Genes Dev. 15, 485–490.Shen, W., Mardon, G., 1997. Ectopic eye development in Drosophila
induced by directed dachsund expression. Development 124, 45–52.Shim, K., Blake, K.J., Jack, J., Krasnow, M.A., 2001. The Drosophila
ribbon gene encodes a nuclear BTB domain protein that promotes epithe-lial migration and morphogenesis. Development 128, 4923–4933.
Siegmund, T., Lehmann, M., 2002. The Drosophila Pipsqueak proteindefines a new family of helix–turn–helix DNA-binding proteins. Dev.Genes Evol. 212, 152–157.
Staehling-Hampton, K., Jackson, P.D., Clark, M.J., Brand, A.H., Hoff-mann, F.M., 1994. Specificity of bone morphogenetic protein-relatedfactors: cell fate and gene expression changes in Drosophila embryosinduced by decapentaplegic but not 60A. Cell Growth Differ. 5, 585–593.
Struhl, G., 1981. A homeotic mutation transforming leg to antenna inDrosophila. Nature 292, 635–638.
Struhl, G., 1982. Spineless-aristapedia: a homeotic gene that does notcontrol the development of specific compartments in Drosophila. Ge-netics 102, 737–749.
Treisman, J.E., 1999. A conserved blueprint for the eye? Bioessays 21,843–850.
Treisman, J.E., Rubin, G.M., 1995. wingless inhibits morphogenetic fur-row movement in the Drosophila eye disc. Development 121, 3519–1527.
Wagner-Bernholz, J.T., Wilson, C., Gibson, G., Schuh, R., Gehring, W.,1991. Identification of target genes of the homeotic gene Antennapediaby enhancer detection. Genes Dev. 5, 2467–2480.
Weber, U., Siegel, V., Mlodzik, M., 1995. pipsqueak encodes a novelnuclear protein required downstream of seven-up for the developmentof photoreceptors R3 and R4. EMBO J. 14, 6247–6257.
Wirz, J., Fessler, L.I., Gehring, W.J., 1986. Localization of the Antenna-pedia protein in Drosophila embryos and imaginal discs. EMBO J. 5,3327–3334.
Wu, J., Cohen, S.M., 1999. Proximal distal axis formation in the Drosoph-ila leg: primary subdivision into proximal and distal domains by Ho-mothorax, Teashirt and Distal-less expression. Development 126, 109–117.
Yamaguchi, M., Hirose, F., Inoue, Y.H., Shiraki, M., Hayashi, Y., Nishi,Y., Matsukage, A., 1999. Ectopic expression of human p53 inhibitsentry into S phase and induces apoptosis in the Drosophila eye imag-inal disc. Oncogene 18, 6767–6775.
483M. Suzanne et al. / Developmental Biology 260 (2003) 465–483