the human genome and the chromosomal basis of heredity · the human genome and the chromosomal...
TRANSCRIPT

The human genome and the
chromosomal basis of heredity
• The genome
• The chromosome
Mitosis
Meiosis
• 3D
Thierry Voet([email protected])

Grown body = 100 trillion somatic cells
DNA:
*46 chromosomes• linear• ~3.1 x 109 bp : autosomes + X + Y• … genes?
*Mitochondrial DNA• circular• 16.6 x 103 bp• 37 genes

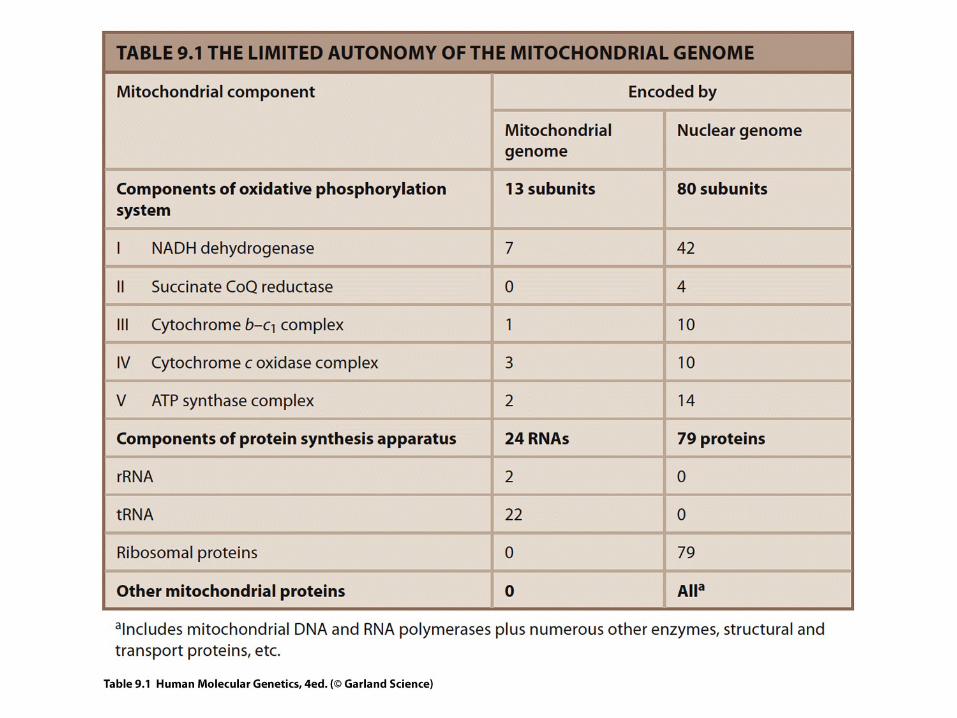

Number mtDNA-copies is cell type dependent+
Only maternal inheritance !


• Retrotransposons (copy-and-paste)
• Transposons (cut-and-paste | inactive)
4.6%4%
3%
20%
Transposons


TPRT:Target-site primed reverse transcription
Model of L1- / Alu-retrotransposition
Integration in TTTT|A (preference for AT-rich regions)
1/100 are full lengths (genome-wide average = 900bp)
80-100 full-lengths L1 (n=6000) not active
Gene mutation for L1-insertion
LINE-1 products used for retrotransposition of SINEs, mRNAs (-> processed pseudogenen) and retrogenes

Processed pseudogenes: defect copy of a gene consisting only of exons (no introns, no promoter sequences)
retrotransposition

Retrogene: integration of cDNA at a promoter + selection
Testis-specific expression of intron-less retrogenes(copies of genes from the X-chr)


Basic unit of a chromosome ?

Linker DNA(10 bp – 60 bp)
DNA is not naked in the cell(nucleosome)

basic unit of chromosomes =nucleosome
Linker DNA(10 bp – 60 bp)
DNA is not naked in the cell


basic unit of chromosomes =nucleosome
Linker DNA(10 bp – 60 bp)
H1
DNA is not naked in the cell


basic unit of chromosomes =nucleosome
Linker DNA(10 bp – 60 bp)
H1
DNA is not naked in the cell

basic unit of chromosomes =nucleosome
Linker DNA(10 bp – 60 bp)
H1
DNA is not naked in the cell


1. Histone-modifying enzymes (N/C-tail posttranslational modification)
2. ATP-dependent chromatin-remodelling enzymes3. Histone-variants
Histone code
Chromatin (structural) modifications play key roles in DNA-related processes
A. Histone modifications:
B. DNA modification: DNA CpG methylation
Chromatin modifications are important for DNA-relatedprocesses:
-DNA-transcription (gene expression/repression)-DNA-repair-DNA-replication-DNA-compaction-chromosome segregation…
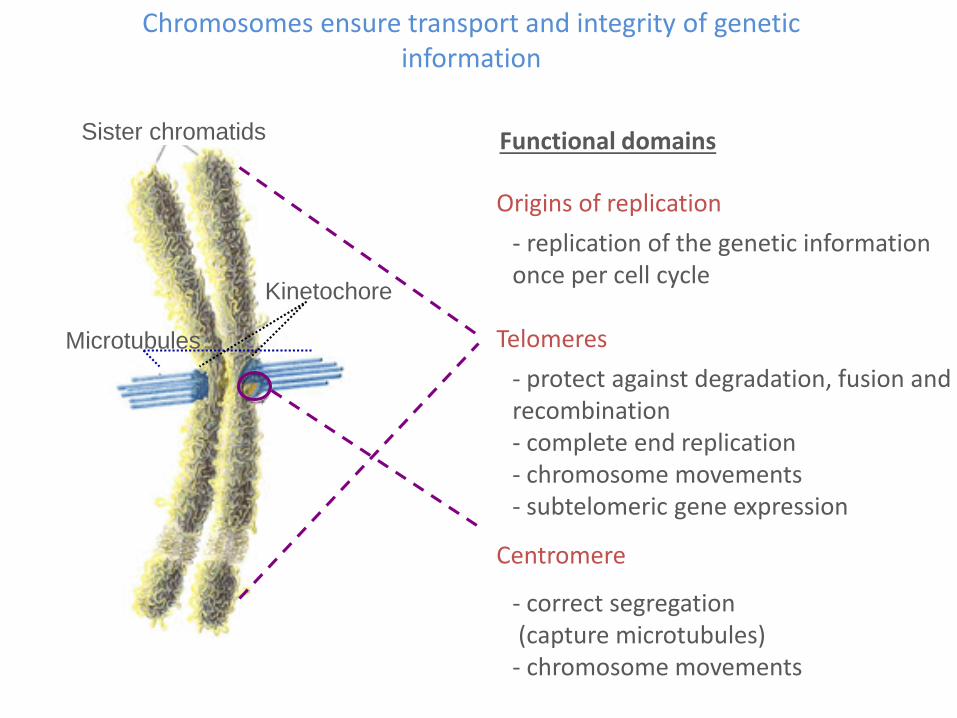
- correct segregation(capture microtubules)- chromosome movements
Centromere
- protect against degradation, fusion and recombination- complete end replication- chromosome movements- subtelomeric gene expression
Telomeres
- replication of the genetic information once per cell cycle
Origins of replication
Sister chromatids
Microtubules
Kinetochore
Functional domains
Chromosomes ensure transport and integrity of geneticinformation

Constitutive proteins are permanently associated with the centromere even during interphase, whereas facultative proteins are recruited only during mitosis to assemble the full kinetochore.
(1) Centromere
BA

Centromeric DNA:Alpha-satellite or alphoid DNA at normal human chromosomes

Centromeric DNA:Alpha-satellite or alphoid DNA at normal human chromosomes

Centromeric DNA:Alpha-satellite or alphoid DNA at normal human chromosomes
One alphoid higher order repeat can be :- specific for one chromomosome- occurring on different chromosomes
Different alphoid higher order repeats can be co-existing on the same chromosome

Alpha-satellite DNA is not sufficient nor necessary for centromere function
A. Pseudo-di-centric chromosomes
Centromere function is epigenetically regulated
B. Neo-centromeres without alpha-satellite DNA

Centromere DNA elements

Localised centromeres
Diffuse centromeres
FEBS Letters 582 (2008) 1950–1959
Histone H3 variant CENP-A is the best candidate to carry the epigenetic centromere mark
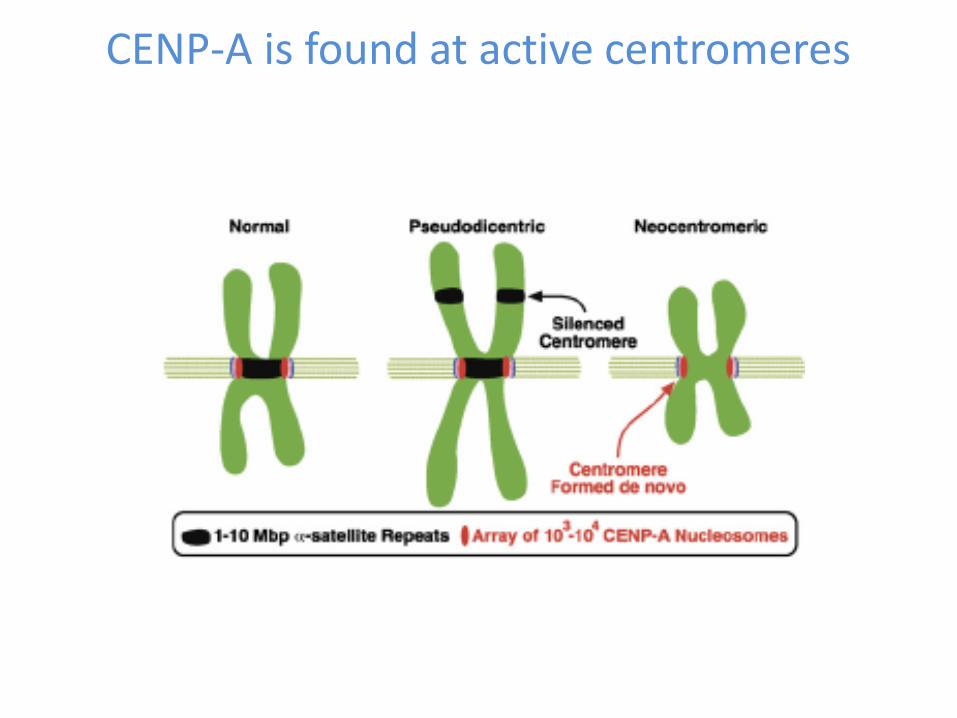
CENP-A is found at active centromeres
Current Opinion in Cell Biology 2008, 20:91–100

Achieving ordered chromatin structure at the centromere
Current Opinion in Cell Biology 2008, 20:91–100

Molecular players of functional centromeres

- correct segregation(capture microtubules)- chromosome movements
Centromere
- protect against degradation, fusion and recombination- complete end replication- chromosome movements- subtelomeric gene expression
Telomeres
- replication of the genetic information once per cell cycle
Origins of replication
Sister chromatids
Microtubules
Kinetochore
Functional domains
Chromosomes ensure transport and integrity of geneticinformation


Nucleoprotein complex: (TTAGGG)2500 + shelterin
TRF1 Telomeric Repeat-Binding Factor 1TRF2 Telomeric Repeat-Binding Factor 2Rap1 Telomeric Repeat Binding Factor 2, Interacting ProteinTIN2 TRF1-Interacting Nuclear Factor 2TPP1 TIN2 And POT1-Interacting ProteinPOT1 Protection Of Telomeres 1
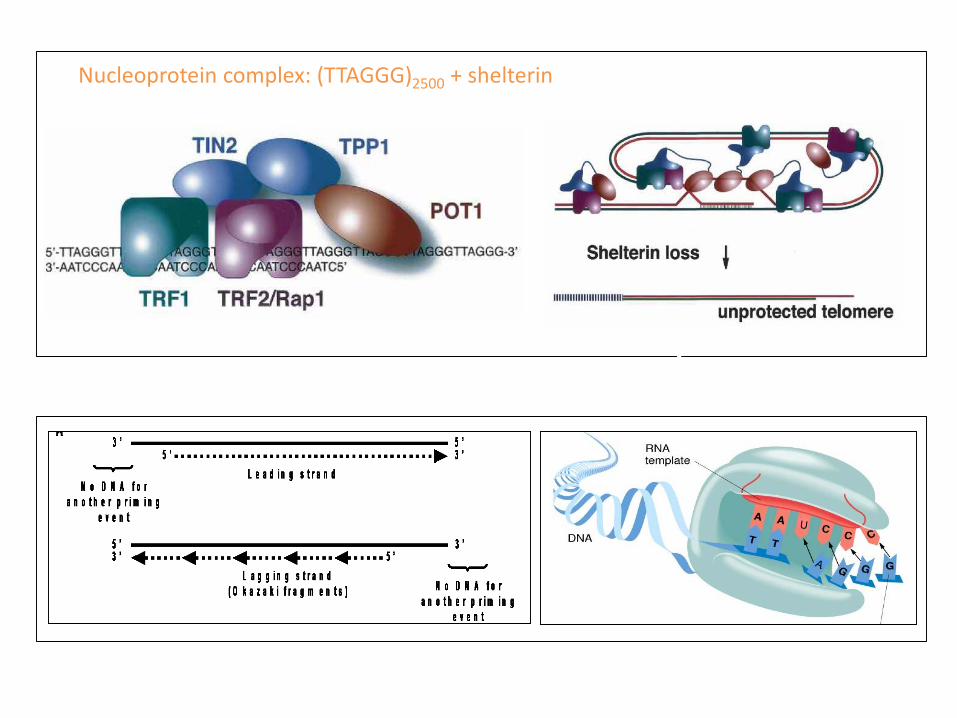
Nucleoprotein complex: (TTAGGG)2500 + shelterin


- correct segregation(capture microtubules)- chromosome movements
Centromere
- protect against degradation, fusion and recombination- complete end replication- chromosome movements- subtelomeric gene expression
Telomeres
- replication of the genetic information once per cell cycle
Origins of replication
Sister chromatids
Microtubules
Kinetochore
Functional domains
Chromosomes ensure transport and integrity of geneticinformation

ORC (multiprotein origin of replication complex)
Origins of DNA-replication

Chromosomal segregation during mitosis

Haploid cell:• n (# different chrs = 23; chromosome set)• C (DNA-content) = ~ 3.5 pg
Diploid cell:• 2n• 2C
Nulliploid cells / Polyploid cells

interfaze

Interphase
Prophase
Prometaphase
Metaphase
Anaphase
Telophase
Cytokinesis
DNAtubulin

Interphase
Prophase
Prometaphase
Metaphase
Anaphase
Telophase
Cytokinesis

Interphase
Prophase
Prometaphase
Metaphase
Anaphase
Telophase
Cytokinesis
Foutieve vasthechting van microtubuli aan kinetochoren komt tijdens de prometafase meer voor, maar moet geremedieerd worden alvorens anafase start

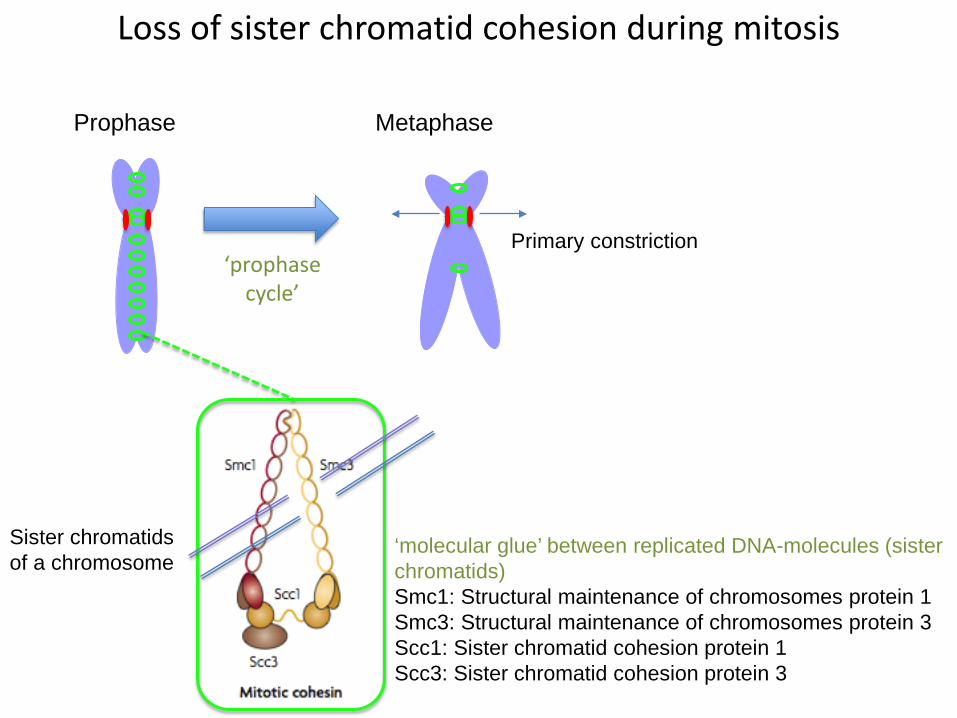
Molecular glue between replicated DNA = cohesin complex
Prophase
‘molecular glue’ between replicated DNA-molecules (sister chromatids)Smc1: Structural maintenance of chromosomes protein 1Smc3: Structural maintenance of chromosomes protein 3Scc1: Sister chromatid cohesion protein 1Scc3: Sister chromatid cohesion protein 3
Sister chromatids of a chromosome

Loss of sister chromatid cohesion during mitosis
Metaphase
‘prophasecycle’
‘molecular glue’ between replicated DNA-molecules (sister chromatids)Smc1: Structural maintenance of chromosomes protein 1Smc3: Structural maintenance of chromosomes protein 3Scc1: Sister chromatid cohesion protein 1Scc3: Sister chromatid cohesion protein 3
Sister chromatids of a chromosome
Prophase
Primary constriction

Prometaphase Metaphase
Loss of sister chromatid cohesion during mitosis
Metaphase
‘prophasecycle’
‘molecular glue’ between replicated DNA-molecules (sister chromatids)Smc1: Structural maintenance of chromosomes protein 1Smc3: Structural maintenance of chromosomes protein 3Scc1: Sister chromatid cohesion protein 1Scc3: Sister chromatid cohesion protein 3
Sister chromatids of a chromosome
Prophase
Primary constriction

Loss of sister chromatid cohesion during mitosis
‘prophasecycle’
‘separase’
‘point of no return’
MetaphaseProphase Anaphase
‘molecular glue’ between replicated DNA-molecules (sister chromatids)Smc1: Structural maintenance of chromosomes protein 1Smc3: Structural maintenance of chromosomes protein 3Scc1: Sister chromatid cohesion protein 1Scc3: Sister chromatid cohesion protein 3




Loss of sister chromatid cohesion during mitosis
Pro(meta)phase
Metaphase
Metaphase
Anaphase
Cohesin complex

Metaphase
Metaphase
Anaphase
Pro(meta)phase
Loss of sister chromatid cohesion during mitosis

Metaphase
Metaphase
Anaphase
Pro(meta)phase
Loss of sister chromatid cohesion during mitosis

Physical dislocation of Sgo-PP2A allows phosphorylation and separase-mediated
cleavage of centromeric cohesin

Chromosomal segregation during meiosis

Meiosis: ‘to reduce’
Diploid somatic cell (2n)
Haploid gametes4 x (n)
meiosis I: reductional division
- separation of homologous autosomes- separation of sex chromosomes
meiosis II: equational division
separation of sister chromatids(~mitosis)
Chromosomes replicated once

Meiosis: ‘to reduce’
Diploid somatic cell (2n)
Haploid gametes4 x (n)
meiosis I: reductional division
- separation of homologous autosomes- separation of sex chromosomes
meiosis II: equational division
separation of sister chromatids(~mitosis)
Chromosomes replicated onceCrossovers -> genetic diversity
-> correct homologue segregation

Meiosis: ‘to reduce’
Diploid somatic cell (2n)
Haploid gametes4 x (n)
meiosis I: reductional division
- separation of homologous autosomes- separation of sex chromosomes
meiosis II: equational division
separation of sister chromatids(~mitosis)
Chromosomes replicated once
Random assortment: 223 = 8388608
Crossovers -> genetic diversity-> correct homologue segregation


2.Zygonema 4.Diplonema
Bouquet formation
3.Pachynema
Tijdens profase I van meiose I worden chiasmata (genetische crossovers) gevormd. Profase I duurtbijgevolg lang en kan in verschillende stadia worden ingedeeld afhankelijk van de ‘nucleusarchitectuur’: homologe chromosomen aligneren, paren, gaan in synapsis en recombineren. HetDNA wordt hiertoe ondermeer opzettelijk beschadigd tijdens leptonema. De chromosomen zulleneveneens condenseren.
1.Leptonema
DSB
SC
Genetic crossover
DSB
5.Diakinesis

Sister chromatids of the maternal homologue
Meiotic cohesin complex(SMC1β / SMC3 / REC8 / STAG3SMC1α / SMC3 / REC8 / STAG3SMC1β / SMC3 / RAD21 / SA1/2)
DSB
SPO11
Tijdens leptonema van profase I zullen meiotische cellen opzettelijk hun DNA beschadigen (creatie van dubbelstrengige DNA-breuken (DSB)) door middel van het SPO11 proteïne. De
breuken zullen bij voorkeur hersteld worden door een intacte DNA-matrijs te gebruiken van het homoloog chromosoom.

Sister chromatids of the maternal homologue
Meiotic cohesin complex(SMC1β / SMC3 / REC8 / STAG3SMC1α / SMC3 / REC8 / STAG3SMC1β / SMC3 / RAD21 / SA1/2)
DSB
SPO11
Sister chromatids of the paternal homologue
meiosis
Tijdens leptonema van profase I zullen meiotische cellen opzettelijk hun DNA beschadigen (creatie van dubbelstrengige DNA-breuken (DSB)) door middel van het SPO11 proteïne. De
breuken zullen bij voorkeur hersteld worden door een intacte DNA-matrijs te gebruiken van het homoloog chromosoom.

Mlh1Mlh3
Spo11
RPARad51Dmc1
Homologous Recombination(generation of genetic crossovers)
Maternal chromatid
Paternal chromatid
Szostak model
Crossover ± Gene conversion
Recombination nodules
>200 DSBs are generated <-> ~1 crossover per chromosome arm (~ sex)

Synthesis-dependent strand annealing
Majority of >200 DSBs is repaired as a non-crossover
Spo11
Homologous Recombination(non-crossovers)
Paternal chromatid
Maternal chromatid
Gene conversion

Sister chromatids of the maternal homologue
Meiotic cohesin complex(SMC1β / SMC3 / REC8 / STAG3SMC1α / SMC3 / REC8 / STAG3SMC1β / SMC3 / RAD21 / SA1/2)
DSB
SPO11
Sister chromatids of the paternal homologue
meiosis
Synaptonemal complex
Transverse filaments (SCP1)
Axial/lateral element (SCP2 / SCP3)
Tussen leptonema en pachynema zullen deze dubbelstrengige breuken hersteld worden als crossovers en non-crossovers. De homologe chromosomen zullen hiertoe aligneren, paren en
in synapsis gaan (vorming van synaptonemal complex tussen homologe chromosomen).

Analysis of prophase I :DSB formation and repair process / Homologue pairing and synapsis(γH2AX (marker for DSB)– SCP3 - DNA staining)
Leptonema

Analysis of prophase I :DSB formation and repair process / Homologue pairing and synapsis(Rad51 – SCP3 - DNA staining)
Early leptonema

Late leptonema - Early zygonema
Analysis of prophase I :DSB formation and repair process / Homologue pairing and synapsis(Rad51 – SCP3 - DNA staining)
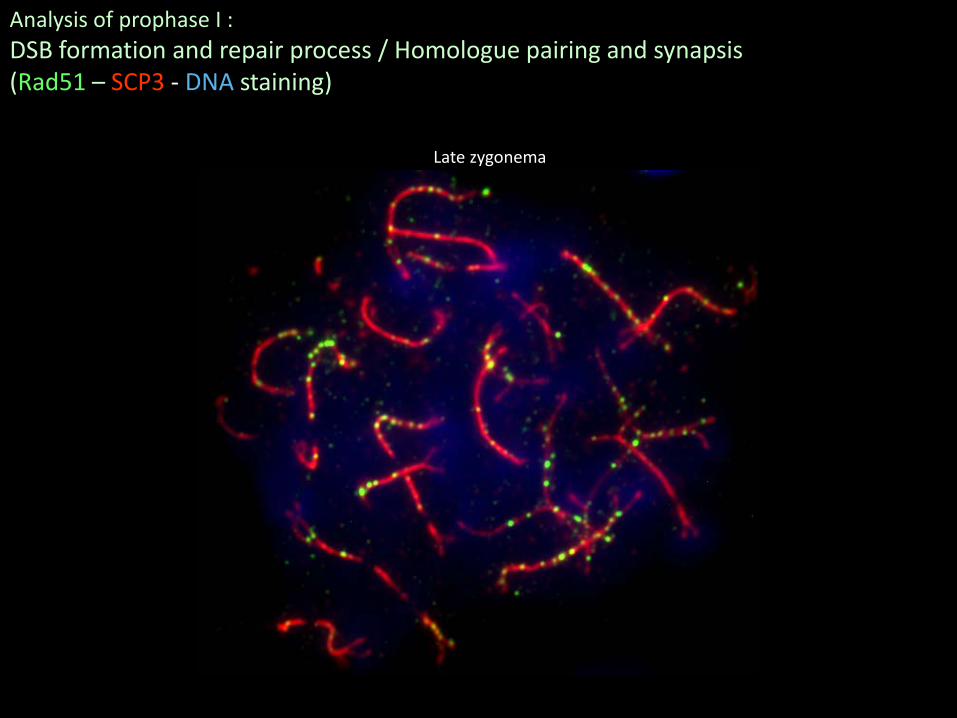
Late zygonema
Analysis of prophase I :DSB formation and repair process / Homologue pairing and synapsis(Rad51 – SCP3 - DNA staining)

Early-Pachynema
Analysis of prophase I :DSB formation and repair process / Homologue pairing and synapsis(Rad51 – SCP3 - DNA staining)

Mid-Pachynema
Analysis of prophase I :DSB formation and repair process / Homologue pairing and synapsis(Mlh1 (merker voor genetische crossover) – SCP3 - DNA staining)
Cross-over interference
h.PAR:2.6Mb Xp/Yp0.32Mb Xq/Yq

Diplonema
Analysis of prophase I :DSB formation and repair process / Homologue pairing and synapsis(Rad51 – SCP3 - DNA staining)

Analysis of metaphase I :20 bivalents in normal mouse meiosis(FISH Y – X – DNA staining)
Chiasmata

Mlh1Mlh3
Spo11RPARad51Dmc1
Homologous Recombination(generation of genetic crossovers)
Maternal chromatid
Paternal chromatid
Szostak model
Crossover ± Gene conversion
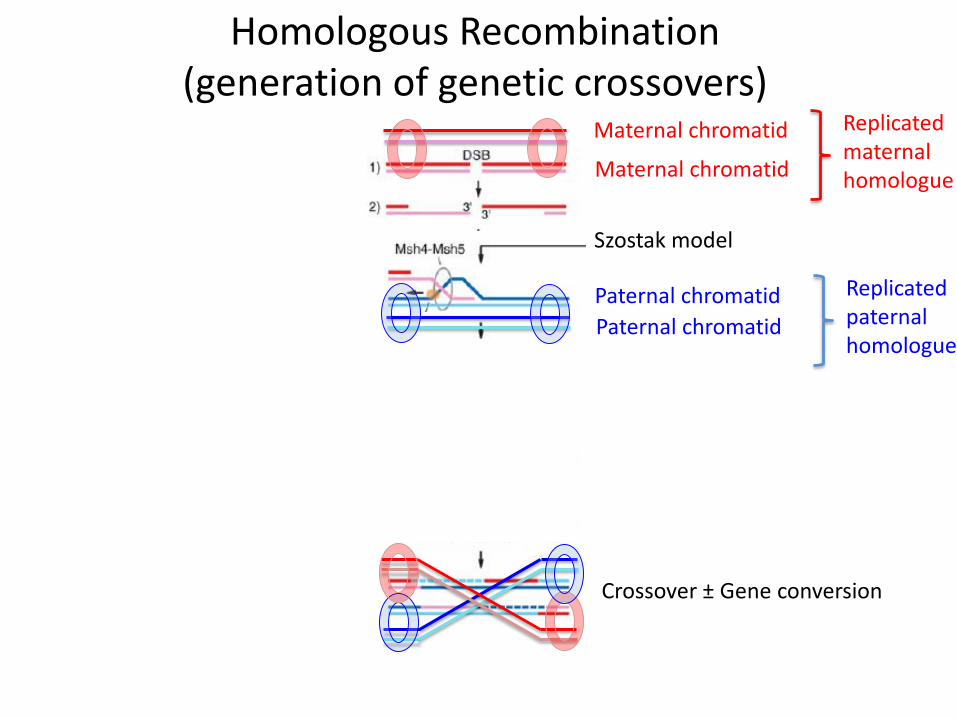
Homologous Recombination(generation of genetic crossovers)
Maternal chromatid
Paternal chromatid
Szostak model
Crossover ± Gene conversion
Maternal chromatid Replicated maternal homologue
Paternal chromatid
Replicated paternal homologue

Premeiotische S-faseProfase


separase
Cohesin rings at the chromosomal arms are opened, but remain intact at the centromeres
First meiotic division

separase
Cohesin rings at centromeres are opened
Second meiotic division

Physical dislocation of Sgo-PP2A allows phosphorylation and separase-mediated
cleavage of centromeric cohesin

Differences between spermatogenesis and
oogenesis

Continuous from puberty till death.4 spermatids per primary spermatocyte.
Discontinuous from fetal life till menopause.1 definitive oocyte per primary oocyte possible.
More genetic crossovers.
Less “strict”.
Mitosis
DNA-synthesis
Meiosis I
Meiosis II

Recombination is not random across chromosomes and differs according to sex
Sex-averaged recombination rate: 1.22cM per Mb(0.88cM / Mb in male // 1.55 cM / Mb in female) <> PAR
Physical mapGenetic map

The human genome and the
chromosomal basis of heredity
• The genome
• The chromosome
Mitosis
Meiosis
• 3D
Thierry Voet([email protected])

Individual chromosomes occupy distinct chromosome territories in the interphase nucleus
DAPI1819
Positioning of chromosomes in the nucleus is not exact, but also not random(centromeres in the G1-phase nucleus at the periphery;Non-overlapping territories; gene-poor <-> gene-rich)


