the interaction of the histone h1-related protein φ0 with chromatin
TRANSCRIPT

Biophysical Chemistry 28 (1987) 51-57
51
Elsevier
BPC 01182
The interaction of the histone H1-related protein % with chromatin
Carlos Olivares, Fernando Azorin, Juan A. Subirana and LLuis CornudellaUnidad de Quimica Macromolecular del C. S. I. C., Universidad Politecnica de Catalunya, E. T. S.I.I. de Barcelona, Diagonal 647,
08028 Barcelona, Spain
Received 15 April 1987
Revised manuscript received 19 June 1987Accepted 19 June 1987
Protein $o ; Histone HI ; Chromatin; Nucleosome; Spermatogenesis
Protein ¢e is a unique protein which is present in the sperm of the sea cucumber, Holothuria tubulosa . It associates with histories, butits physiological role is unknown . From its amino acid composition and sequence, protein ¢ e can be considered as an HI-relatedprotein. In this paper, we have studied its interaction with chicken erythrocyte chromatin particles of different complexity, from coreparticles to polynucleosomes. Addition of protein $e results in marked chromatin insolubilization . The higher the molecular weight ofthe chromatin fragment, the lower is the $o/nucleosome molar ratio at which precipitation occurs, so that complete insolubilizationof polynucleosomes is achieved at a $ e/nucleosome molar ratio which is identical to that found in mature H. tubulosa spermatozoa.We have also found that the interaction of protein 0 0 with chromatin is cooperative . These findings contribute to clarification of thepeculiar physico-chemical properties shown by H. tubulosa sperm chromatin and the role played by the .0o protein .
1. Introduction
anomalous nucleosomal particles which containexclusively 00 and histone Hl.
In order to establish whether protein 4)o isresponsible for this unusual physico-chemical be-haviour, we have studied in detail the in vitrointeraction of protein 9so with different chromatinparticles of increasing complexity . Protein ¢o is 78amino acids long and contains 17 .0% lysine, 25.9%arginine and 29.5% alanine [6] . In spite of its higharginine content, this protein is related to histoneH1, since its sequence is very similar to the C-terminal region of histone H1 [6], as long asarginine is considered equivalent to lysine . More-over, antisera induced against protein 0 0 cross-re-act with histone H1 [7] . Therefore, protein 00appears to be equivalent to an H1 histone whichlacks the central globular region . In this paper, wehave found that addition of protein 00 to chro-matin particles of increasing size, ranging fromcore particles to polynucleosomes, results in theirinsolubilization. We have observed that the higherthe molecular weight of the particle, the lower is
The chromatin of the spermatozoa from the seacucumber, Holothuria tubulosa, has a peculiarcomposition, in which the normal complement ofhistones is accompanied by a small amount (about4%) of a strongly basic low molecular weightprotein, which we have called 0 0 [1] . Previousexperiments from our laboratory [2-5] have shownthat H. tubulosa sperm chromatin is rather re-markable. On the one hand, purified nuclei swellonly slightly at low ionic strength, and when theyare subjected to micrococcal nuclease digestiononly about 50% of the total DNA can be solubi-lized. All the 0. protein remains associated withthe insoluble fraction . Furthermore, during micro-coccal nuclease digestion protein S5o appears todisplace histones from core particles, giving rise to
Correspondence address : F. Azorin, Unidad de Quimica Mac-romolecular del C.S.I.C., Universidad Politecnica de Catalunya,E.T.S.LI . de Barcelona, Diagonal 647, 08028 Barcelona, Spain .
0301-4622/87/$03 .50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)

52
the molar ratio necessary for its insolubilization.Insolubilization of polynucleosomes occurs at a4oo/nucleosome molar ratio which is very close tothat found in mature H. tubulosa spermatozoa,suggesting that this protein is actually responsiblefor the high insolubility shown by H. tubulosasperm chromatin. We have also found that theinteraction of protein 00 with chromatin is cooper-ative.
2. Materials and methods
2.1 . Preparation of nuclei and chromatin
Chicken erythrocyte nuclei were preparedaccording to ref. 8. Chromatin fragments of vari-ous lengths were obtained by micrococcal nucleasedigestion, followed by fractionation throughsucrose gradients as described previously [9,10] .To obtain short oligomers (up to octanucleo-somes), purified nuclei were digested at an en-zyme/DNA ratio of 50 Boehringer U/mg DNAfor 15 min. Polynucleosomes were obtained afterdigestion with 15 enzyme U/mg DNA for 1 .5min. The DNA and protein contents of the sam-ples used in these experiments were analyzed bygel electrophoresis (not shown) .
2.2. Protein i0o purification
Protein 00 was obtained from H. tubulosaspermatozoa by differential acid extraction andpurified to homogeneity by chromatography asdescribed elsewhere [6] .
2.3. Protein ¢ochromatin interaction
Purified protein 4i, was dissolved in 10 mMTris-HCI, 1 mM EDTA (pH 7 .5) (buffer A) to aconcentration of 2 mg/n11. The actual concentra-tion was determined by turbidometry [11].
Chromatin in buffer A was diluted to 1 ml(A260= 0.3) with the same buffer and the saltconcentration adjusted by the addition of 4 MNaCl. Protein $o was then added at different00/nucleosome molar ratios . Following a 10 mnincubation at room temperature, complexes were
C. Olivares et al. /Protein ¢o-chromatin interaction
centrifuged in an Eppendorf microfuge for 10min . The percentage of soluble chromatin wasdetermined from the A26, of the supernatants.Proteins contained in supernatants and pellets wereanalyzed by SDS gel electrophoresis [12]. Prior toelectrophoretic analysis, proteins contained in su-pernatants were precipitated with 18% trichloro-acetic acid .
3. Results
3.1. Solubility of chromatin fragments
The solubility of the chromatin fragments usedin our experiments is shown in fig . 1 . Chromatincore particles are soluble throughout the entirerange of ionic strengths examined. The presenceof lysine-rich histones H1 and H5 confers markedinsolubility to chromatin. The maximum of in-solubility is centered around 0.15 M NaCl foroligonucleosomal particles . In particular, the frac-tion containing from penta- to octanucleosomes(5 = 5 .75) is completely insoluble at 0.15 M NaCl.In contrast, polynucleosomal particles containingon average more than 50 nucleosomes are lessinsoluble than shorter oligonucleosomes and, ad-
NaCl (M)
Fig. 1 . Solubility of different chromatin fragments as a func-tion of NaCl concentration . (•) Core particles, (0) mono-nucleosomes, (a) dinucleosomes, (n) oligonucleosomes (o =3.75), (•) oligonucteosomes (v = 5.75), (0) polynucleosomes
(ii > 50). 5, average number of nucleosomes .

ditionally, the maximum of insolubility occurs athigher salt concentrations, around 0 .20 M NaCl .This higher solubility is likely to be related withthe organization of polynucleosomes into higher-order structures [9,13] .
3.2. Addition of protein 00 to chromatin results in itsinsolubilization
The addition of increasing amounts of purifiedprotein $o to different chromatin particles resultsin their insolubilization. Fig. 2 shows the effect ofadding increasing amounts of protein % on thesolubilityy of different chromatin particles at threedifferent NaCl concentrations, namely, 0, 0 .05 and0.15 M. In all cases, marked insolubilization isdetected but the actual 4 /nucleosome molar ratioat which precipitation occurs depends on both theionic strength and complexity of the chromatinparticle. Table 1 summarizes the results shown infig. 2. In general, we have observed that for anychromatin fragment, the higher the salt concentra-tion the lower is the molar ratio required for itsinsolubilization. Likewise, large oligonucleosomesare precipitated more easily than shorter frag-ments. For instance, at 0.05 M NaCl, completeprecipitation of mononucleosomes occurs at amolar ratio of approx. 2.8 00 molecules per
TN.4
0N
10090
80
70
60
5040
30
20
10
0 1 2 3 4 5 8 7 8 9
C Oliva es et al. /Protein %-chromatin interaction
53
0,15M NaCl 0.05M NeCI OM NBCI
0 1 2 3 4 5 8 7 8 9
nucleosome while oligonucleosomal samples con-taining 5.75 nucleosomes on average precipitate ata molar ratio of about 1 .2 4 molecules pernucleosome. However, a much larger excess ofprotein $o is required to precipitate chromatincore particles which lack lysine-rich histones H1and H5 as well as internucleosomal DNA . About6 00 molecules are necessary to achieve completeprecipitation at 0 .05 M NaCl .
On the other hand, polynucleosomal particlesshowed quite different behaviour . They are ingeneral more soluble than shorter oligonucleo-somal particles (fig . 1). Similarly, their complexeswith protein .0o are also more soluble. At 0.05 MNaCl, complete insolubilization is achieved at amolar ratio of 3 .5 0,0 molecules per nucleosome. Itis worth noting that the molar ratio required toinsolubilite polynucleosomes at 0 .15 M NaCl cor-relates with the amount of protein $o present inmature spermatozoa. Since the amount required toinsolubilize polynucleosomes . at 0 M NaCl is simi-lar to that required to precipitate shorteroligonucleosomes, it is likely that the higher solu-bility of the 0o-polynucleosome complexes de-tected at 0.05 and 0.15 M NaCl might be relatedto the organization of polynucleosomes into thick30-nm chromatin fibers which are known to occurat these NaCl concentrations [9,13] .
0 1 2 3 4 5 8 7 8 9
'Q0 I Nuc leosome molar ratio
Fig. 2. Insolubilization of different chromatin fragments as a function of added protein 0 0 at three NaCl concentrations . (•) Coreparticles, (s) mononucleosomes, (a) dinucleosomes, (a) oligonucleosomes (o = 3.75), (•) oligonucleosomes (ii = 5.75), (0) poly-
nucleosomes (e > 50) . n, average number of nucleosomes .

54
C. Olioares et al. /Protein 00 -chromatin interaction
Table 1
*0 /nucleosome molar ratio at which 50% precipitation occursfor different chromatin particles at three NaCl concentrations
a 6, average number of nucleosomes .
3.3. Interaction of protein tPo with chromatin iscooperative
We have found that a given amount of .po inthe presence of more than its molecular equivalentof chromatin particles will bind extensively tosome of these particles and only partially or not atall to the rest, indicating that the interaction ofprotein $o with chromatin is cooperative . At NaClconcentrations of 0.1.5 or 0.05 M any 40-contain-ing particle precipitates . Fig. 3 shows the SDS gelelectrophoretic patterns of the supernatants andpellets obtained after addition of increasingamounts of protein 00 to different chromatin par-ticles at 0.15 M NaCl. Protein 00 is always foundin the precipitates (fig . 3, right-hand panels) sothat it is never detected in the supernatants (fig . 3,left-hand panels). Similar results were obtainedwhen the interaction was carried out at 0 .05 MNaCl. From fig. 3 it appears that soluble mono-nucleosomal particles are deficient in lysine-richhistories Hl and H5. However, since the corre-sponding mononucleosomal pellets are not en-riched in these proteins, we conclude that theapparent depletion of Hl and H5 in solublemononucleosomes is mostly due to losses occur-ring during preparation of the samples prior toelectrophoresis, as has been observed by others[15] . As shown in fig. 4, the c0/nuclcosome molarratio of the pellets is constant and independent ofthe input molar ratio until complete precipitation
IICLII$IIIS
ILIIIIIGL[ISI6tl(t.L76)IUPIIIATAITS
0 0.6
PILTIISIE1liu!t
lot
ANTI
0
0 06 126 4.6 6 10
1250 6
Fig. 3. SDS gel electrophoresis of proteins contained in thesupernatants and pellets obtained after addition of increasingamounts of protein %6o to different chromatin partcles whenthe interaction was carried out at 0 .15 M NaCl. Values indicatethe $o /nucleosome molar ratio at which the interaction took
place.
of chromatin occurs, indicating that protein .0odoes not distribute itself evenly among all accessi-ble particles but rather interacts in a cooperativefashion with chromatin. At an input 0o/ nucleo-some molar ratio of unity and assuming a randomnon-cooperative interaction, each mononucleo-some should have received one 400 molecule. Incontrast, at this input molar ratio we observed
NaCl concentration
OM 0.05M 0.15M
Cores 6.7 3.2 2.3Mononucleosomes 4.0 1.4 1 .1Dinucleosomes - - 0.6Oligonucleosomes
(V = 3.75) a 3.7 1.0 0.4Oligonucleosomes
(5 = 5 .75) 3.2 0.6 0Polynucleosomes
(5 > 50) 3.7 1.7 0.25

5
• 4Etn0vu•
3C
,moo2
u0Mauv
IL
2
COZIES
III
0 1
2 3 4 5Input Mr (0o / nucleosorne )
Fig. 4. 1su /nucleosome molar ratios (M,) of the precipitates asa function of the input molar ratio for different chromatin
fragments . u, average number of nucleosomes .
that half of the mononucleosomal particles con-tained two $a molecules while the remaining 50%contained .none.
When the interaction is carried out at 0 MNaCl the behaviour is quite different, since underthese salt conditions protein 0 0 is found either inthe supernatants, at low molar ratios, or in thepellets, at higher molar ratios (fig. 5). That, at lowinput molar ratios, protein 0e actually interactswith chromatin under these salt conditions is dem-onstrated by the fact that, following incubationwith protein 0. at an input molar ratio of 0 .6, thisprotein is found associated with mononucleo-somes purified by sucrose gradient centrifugation.In order to establish whether the interaction isalso cooperative at this ionic strength the follow-ing experiment was performed . Either core par-ticles or mononucleosomes, were incubated at 0 MNaCl with, protein ,4,,, at, different molar ratios .Following incubation for -10 min, the salt con-centration was raised to 0.15 M NaCl and thepercentage of soluble particles determined. Weobtained a similar value fbr' the percentage ofsoluble particles as if the interaction had beenperformed directly at 0 .15 M NaCl, suggestingthat the interaction had also been cooperative at 0
C. Olivares et al. /Protein ¢b -chromatin interaction 55
cellsSIPEIIATAATS
0 Q6 125 2$ 5
IIILEISIIESII PEIIATAITS
0 00 1,25 2p
ILIIIIISLEIAI#ES(ds 4 .78)SIPEIIATAITS
PILTIISLEISII/ES
SFPEIIATAITI
PELLETS
10
PELLETS
5 10
+0a
PELLETS
PELLETS
Fig. 5 . SDS gel electrophoresis of proteins contained in thesupernatants and pellets obtained after addition of increasingamounts of protein $v to different chromatin particles whenthe interaction was carried out at 0 M NaCI . Values indicatethe 4o/nucleosome molar ratio at which interaction took
place .
M NaCl. These results might also be interpretedas being the consequence of redistribution of pro-tein % at 0.15 M NaCl. However, when purifiedH. tubulosa sperm nuclei were incubated at 0 .15 M
56
zo
is
M 5
0
/
0.05M•
OASM
O 1 s
s
C. ONvares et al. /Protein #0-chromatin interaction
4 5 6
I/
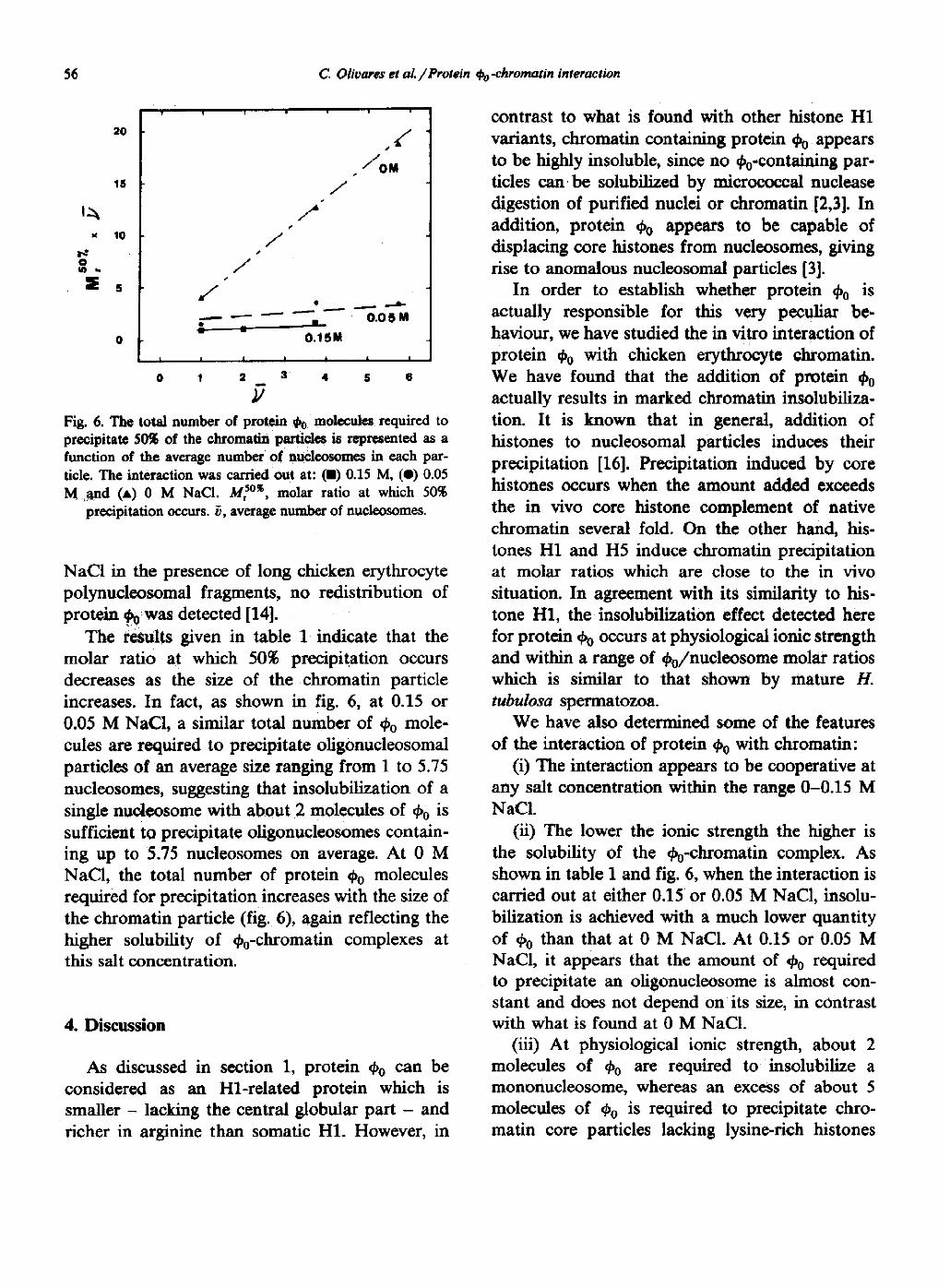
Fig. 6. The total number of protein
molecules required toprecipitate 50% of the chromatin particles is represented as afunction of the average number of nucleosomes in each par-ticle. The interaction was carried out at : (0) 0.15 M, (41) 0.05M and (A) 0 M NaCl. M,50*, molar ratio at which 50%
precipitation occurs. u, average number of nucleosomes .
NaCl in the presence of long chicken erythrocytepolynucleosomal fragments, no redistribution ofprotein was detected [14].
The results given in table 1 indicate that themolar ratio at which 50% precipitation occursdecreases as the size of the chromatin particleincreases. In fact, as shown in fig . 6, at 0.15 or0.05 M NaCl, a similar total number of 00 mole-cules are required to precipitate oligonucleosomalparticles of an average size ranging from 1 to 5 .75nucleosomes, suggesting that insolubilization of asingle nucleosome with about 2 molecules of 0 0 issufficient to precipitate oligonucleosomes contain-ing up to 5 .75 nucleosomes on average. At 0 MNaCl, the total number of protein 00 moleculesrequired for precipitation increases with the size ofthe chromatin particle (fig. 6), again reflecting thehigher solubility of 40ochromatin complexes atthis salt concentration .
4. Discussion
As discussed in section 1, protein 0 0 can beconsidered as an H1-related protein which issmaller - lacking the central globular part - andricher in arginine than somatic Hl . However, in
contrast to what is found with other histone Hivariants, chromatin containing protein 0 0 appearsto be highly insoluble, since no 0ocontaining par-ticles can be solubilized by micrococcal nucleasedigestion of purified nuclei or chromatin [2,3] . Inaddition, protein 00 appears to be capable ofdisplacing core histones from nucleosomes, givingrise to anomalous nucleosomal particles [3] .
In order to establish whether protein ¢o isactually responsible for this very peculiar be-haviour, we have studied the in vitro interaction ofprotein 0. with chicken erythrocyte chromatin .We have found that the addition of protein 0 0actually results in marked chromatin insolubiliza-tion. It is known that in general, addition ofhistones to nucleosomal particles induces theirprecipitation [16] . Precipitation induced by corehistones occurs when the amount added exceedsthe in vivo core histone complement of nativechromatin several fold . On the other hand, his-tones H1 and H5 induce chromatin precipitationat molar ratios which are close to the in vivosituation. In agreement with its similarity to his-tone Hl, the insolubiliaation effect detected herefor protein 400 occurs at physiological ionic strengthand within a range of 0 0/nucleosome molar ratioswhich is similar to that shown by mature H.tubulosa spermatozoa.
We have also determined some of the featuresof the interaction of protein oo with chromatin :
(i) The interaction appears to be cooperative atany salt concentration within the range 0-0 .15 MNaCl.
(ii) The lower the ionic strength the higher isthe solubility of the ¢e-chromatin complex . Asshown in table 1 and fig. 6, when the interaction iscarried out at either 0 .15 or 0.05 M NaCI, insolu-bilization is achieved with a much lower quantityof 00 than that at 0 M NaCl. At 0.15 or 0.05 MNaCl, it appears that the amount of 00 requiredto precipitate an oligonucleosome is almost con-stant and does not depend on its size, in contrastwith what is found at 0 M NaCl .
(iii) At physiological ionic strength, about 2molecules of 00 are required to insolubilize amononucleosome, whereas an excess of about 5molecules of 0. is required to precipitate chro-matin core particles lacking lysine-rich histones

and internucleosomal DNA .(iv) Complexes formed with large polynucleo-
somal fragments are insoluble at a molar ratio of0.5 molecules of $c per nucleosome at 0.15 MNaCl, which is very similar to the in vivo 4 molarratio of mature H. tubulosa spermatozoa.
Acknowledgments
We are grateful to Dr. Maria Teresa Casas forproviding pure protein 00 and for helpful discus-sions. The technical assistance of the Taller de laCatedra de Tecnologia Mecanica of the E .T.S.LI .B.is also acknowledged. This work was supported bygrants from the Comision Asesora de Investiga-tion Cientifica y Tecnica (84-214), Consejo Su-perior de Investigaciones Cientificas (85-169), theFondo de Investigaciones Sanitarias de la Seguri-dad Social and the Comissio Interdepartamentalde Recerca i Innovacio Tecnologica . C' . was arecipient of a fellowship from the Instituto deCooperacion Iberoamericana and the Universityof Talca, Chile .
C. Olvares et al. /Protein $o-chromatin interaction
References
I J.A. Subirana, Exp . Cell Res. 63 (1970) 253 .2 F. Azorin, C. Olivares, A. Jordan, L. Perez-Grau, L .
Cornudella and J.A. Subirana, Exp. Cell Res. 148 (1983)331 .
3 F. Azorin, E. Rocha, L . Comudella and J.A. Subirana, Eur.J. Biochem. 148 (1985) 529 .
4 J.A. Subirana, S . Muiioz-Guerra, A .B. Martinet, L . Perez-Grau, X. Marcet and I. Fita, Chromosoma 83 (1981) 455 .
5 L. Cornudella and E. Rocha, Biochemistry 18 (1979) 3724 .6 A. Jordan, Ph .D. Thesis, Faculty of Chemistry, University
of Barcelona (1982) .7 M.T. Casas, Ph.D. Thesis, Faculty of Biology, University of
Barcelona (1986) .8 A.L. Olins, R.D. Carlson, W.B. Wright and D.E. Ohns,
Nucleic Acids Res. 3 (1976) 3271 .9 F. Azorin, L . Perez-Grau and J.A. Subirana, Chromosoma
85 (1982) 251 .10 L. Perez-Grau, F. Azorin and J.A. Subirana, Chromosoma
87 (1982) 437 .11 P.S. Rasmussen, K. Murray and J.M. Luck, Biochemistry 1
(1962) 79.12 U.K. Laemli, Nature 227 (1970) 680 .13 F. Thoma and T . Koller, J. Mol. Biol. 149 (1981) 709 .14 C. Olivares, Ph .D. Thesis, Facultad de Biologia, Universi-
dad de Barcelona (1987) .15 D.L. Bates, P.J.G. Butler, E.C. Pearson and J.O. Thomas,
Eur. J. Biochem. 119 (1981) 469 .16 0. Voordouw and H. Eisenberg, Nature 273 (1978) 446 .
57