the journalof biological c ... · molecularcloningandcharacterizationofnkt,ageneproduct...
TRANSCRIPT

Molecular Cloning and Characterization of NKT, a Gene ProductRelated to the Organic Cation Transporter Family That Is AlmostExclusively Expressed in the Kidney*
(Received for publication, August 8, 1996, and in revised form, January 2, 1997)
Carlos E. Lopez-Nieto‡, Guofeng You, Kevin T. Bush, Elvino J. G. Barros,Davio R. Beier§, and Sanjay K. Nigam¶
From the Renal and Genetics Divisions, Department of Medicine, Harvard Medical School and Brigham and Women’sHospital, Boston, Massachusetts 02115
We have identified a gene product (NKT) encoding anapparently novel transcript that appears to be related tothe organic ion transporter family and is expressed al-most exclusively in the kidney. Analysis of the deduced546-amino acid protein sequence indicates that NKT is aunique gene product which shares a similar transmem-brane domain hydropathy profile as well as transporter-specific amino acid motifs with a variety of bacterialand mammalian nutrient transporters. Nevertheless,the overall homology of NKT to two recently clonedorganic ion transport proteins (NLT and OCT-1) is sig-nificantly greater; together these three gene productsmay represent a new subgroup of transporters. The NKTwas characterized further with respect to its tissue dis-tribution and its expression during kidney develop-ment. A 2.5-kilobase transcript was found in kidney andat much lower levels in brain, but not in a number ofother tissues. Studies on the embryonic kidney indicatethat the NKT transcript is developmentally regulatedwith significant expression beginning at mouse gesta-tional day 18 and rising just before birth, consistentwith a role in differentiated kidney function. Moreover,in situ hybridization detected specific signals in mouserenal proximal tubules. NKT was mapped by linkagedisequilibrium to mouse chromosome 19, the same siteto which several mouse mutations localize, includingthat for osteochondrodystrophy (ocd). Although initialexperiments in a Xenopus oocyte expression systemfailed to demonstrate transport of known substrates forOCT-1, the homology to OCT-1 and other transporters,along with the proximal tubule localization, raise thepossibility that this gene may play a role in organicsolute transport or drug elimination by the kidney.
The proximal tubule of the kidney is a major site of transportof small organic molecules, including glucose, amino acids, and
uric acid. The proximal tubule also plays a key role in drugelimination by the kidney. Xenobiotics and their metabolitesare transported mainly by the organic anion and organic cationtransport systems. These two transport systems share commonsubstrates, and many functional features (2–9), and it is likelythat these transport systems may also resemble each other atthe molecular level.A complementary DNA from rat kidney (OCT-1), which has
the functional characteristics of organic cation uptake in thebasolateral membrane of renal proximal tubules has been re-cently isolated (8). At the present time, only one nucleotidesequence (NLT) with significant homology to OCT-1 has beenreported (9). NLT is a transporter protein of unknown sub-strate(s) present in the sinusoidal (basolateral) domain of hepa-tocytes. Increased expression of NLT at the time of birth cor-relates with the maturation of enterohepatic circulation. NLTis also present in the kidney although at a lower level than inliver. Organic anions, such as bilirubin and bromosulfophtha-lein, have been postulated as potential substrates for NLT,although this remains to be determined.We report here the cloning and the molecular characteriza-
tion of a transcript encoding a novel protein (NKT) apparentlyrelated to the recently identified OCT-1 and NLT. The geneproduct is almost exclusively expressed in kidney.
MATERIALS AND METHODS
Reverse Transcription and PCR1 Amplification—We have previouslyreported a method to selectively represent mammalian protein-codingregions based on statistically designed primer sets (1). This method isbased on the distribution frequency of nucleotide combinations (k-tuples) in certain genetic subsets, and the combined ability of primerpairs, based on these oligonucleotides, to detect genes. Total RNA wasprepared from various mouse tissues (brain, heart, placenta, lung, liver,spleen, kidney, and stomach) using the guanidinium thiocyanate-ce-sium chloride method (10). First strand cDNA was synthesized using acommercial kit (Life Technologies, Inc., Gaithersburg, MD). A 50-mlreaction containing 50 mM Tris-HCl (pH 8.3), 75 mM KCl, 10 mM
dithiothreitol, 3 mM MgCl2, 0.5 mM each dNTP, 10 mg/ml oligo(dT)12–18,5 mg total RNA, 200 units of Moloney murine leukemia virus (reversetranscriptase) was incubated 60 min at 37 °C, followed by PCR ampli-fication. In hot start PCR microcentrifuge tubes (Fisher Scientific,Pittsburg, PA) a 20-ml reaction mixture containing 2 ml of solution fromthe first reaction (the final concentration of buffer components was 50mM KCl, 1.425 mMMgCl2), 1 mM of each primer (Life Technologies, Inc.),12.5 mM of each dNTP, 0.5 mM [35S]dATP, and 1 unit of Taq DNApolymerase (Perkin-Elmer) was used. The reaction mixture was sub-jected to 35 PCR thermocycles at 94 °C for 30 s to denature, 50 °C for30 s for annealing, and 72 °C for 30 s for extension, followed by 5 min at72 °C. For analysis of the PCR products, the samples were electrophore-
* This work was supported in part by National Institutes of HealthGrant DK44503, a grant from Advance Genetics Inc., National Instituteof Child Health and Human Development Grant RO1 HD29028, andNational Center of Human Genomic Research Grant RO1 HG00951 (toD. R. B.). The costs of publication of this article were defrayed in part bythe payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.The nucleotide sequence(s) reported in this paper has been submitted
to the GenBankTM/EBI Data Bank with accession number(s) U52842.‡ Supported by an National Institutes of Health Individual National
Research Service Award.¶ This work was done during tenure as an Established Investigator of
the American Heart Association. To whom correspondence should beaddressed: Renal Division Brigham and Women’s Hospital, 75 FrancisSt., Boston, MA 02115. Tel.: 617-278-0436; Fax: 617-732-6392.
1 The abbreviations used are: PCR, polymerase chain reaction;RACE, rapid amplification of cDNA ends; SSCP, single strand confor-mation polymorphisms; kb, kilobase(s); TEA, triethanolamine; PAH,para-amino hipporic acid.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 272, No. 10, Issue of March 7, pp. 6471–6478, 1997© 1997 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www-jbc.stanford.edu/jbc/ 6471
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

sed in 6% sequencing-grade gels, DNA bands were visualized byautoradiography.Cloning and Sequencing—Bands from these gels that were only
present in the kidney were cut using a razor blade and DNA wasdissolved in water and subsequently precipitated in a solution of 0.3 M
sodium acetate (pH 6) and 2.5 volumes of ethanol. DNA in the pelletwas reamplified using the same primer pair and PCR conditions. Theamplified material was examined in a low-melting point 2% agarose gel,and a commercial kit (TA Cloning[Trade], Invitrogen, San Diego, CA)was employed to clone the PCR products. Positive clones (screened byblue-white changes) were grown in 1 ml of LB broth and plasmid DNAwas isolated and then sequenced on an ABI 373A DNA fluorescentautomated sequencer. Sequence homology searches were performed atthe National Center for Biotechnology Information (NCBI) using theBLAST network service (11, 12).RNA Blot Analysis—Total RNA was extracted from several mouse
tissues (see above), as well as from mouse embryonic kidney fromseveral developmental stages as has been previously described (10).Total RNA was electrophoresed on a 1% agarose/formaldehyde gel andtransferred to a nylon membrane. In addition, human multiple tissueNorthern blots I and II were purchased from Clontech (Palo Alto, CA).The probe used for hybridization was the 332-base pair fragment fromthe NKT cDNA clone originally isolated from the differential displaygels. The probe was labeled with [32P] using a random oligonucleotidelabeling kit (Pharmacia). The final washes were carried out at 65 °C.Blots were exposed to x-ray film with an intensifying screen for 3 daysat 280 °C.Rapid Amplification of 59-cDNA and 39-cDNA Ends (59- and 39-
RACE)—Adaptor-ligated mouse kidney double-stranded cDNA readyfor use as template in 59- and 39-RACE was purchased from Clontech.Gene-specific primers for 59- and 39-RACE reactions were designedbased on the sequence of the 332-base pair fragment from the NKTcDNA originally obtained from the differential display gels. RACEreactions were performed using Clontech’s Advantage[Trade] KlenTaqPolymerase Mix, 0.5 ng of template, 50 mM of each dNTP, 0.2 mM of theadapter primer (API), and 0.2 mM of either the 59 or 39 gene-specificprimer (59- and 39-RACE respectively). The PCR products obtained werecloned and sequenced as has been previously described.In Situ Hybridization—Mouse kidney was collected, rinsed in phos-
phate-buffered saline, and then fixed in ice-cold freshly prepared 4%paraformaldehyde/phosphate-buffered saline for 1 h. They were thenrinsed in 0.9% NaCl and dehydrated through a graded series of ethanoland embedded in paraffin. 7-mm sections were cut, mounted on slides,dewaxed, pretreated, and prehybridized as described in Wedden et al.(13). Antisense RNA probes labeled with [a-35S]UTP (Amersham) wereproduced with T7 RNA polymerase and HindIII-linearized PCR II-NKT. Hybridization was done overnight at 50 °C. Post-hybridizationtreatments were as follows: (i) two washes in 50% formamide, 2 3 SSC,20 mMmercaptoethanol (FSM) at 60 °C for 30 min, (ii) digestion with 10mg/ml RNase A in 4 3 SSC, 20 mM Tris-HCl (pH 7.6), 1 mM EDTA at37 °C for 30 min, and (iii) two washes in FSM at 60 °C for 45 min. Slideswere dipped in Kodak NTB-2 emulsion and exposed for 10 days. Slideswere then stained in 5 mg/ml Hoechst 33258 dye in water for 2 min,followed by rinsing 2 min in water. The slides were viewed underepifluorescence optics.Chromosomal Localization—Primers were designed to amplify a re-
gion corresponding to the 39-untranslated region of NKT in order to testfor single strand conformation polymorphisms (SSCPs) between mousestrains. These were analyzed as described previously (14). Briefly,oligonucleotides were radiolabeled with [32P]ATP using polynucleotidekinase and genomic DNAs from a series of mouse strains were amplifiedusing standard protocols (anneal at 55 °C for 1 min, extend at 72 °C for2 min, and denature at 94 °C for 1 min for 40 cycles, with a finalextension at 72 °C). 2 ml of the amplified reaction was added to 8.5 mlof U. S. Biochemical Corp. stop solution, denatured at 94 °C for 5 min,and immediately placed onto ice. 2 ml of each reaction was loaded on a6% nondenaturing acrylamide sequencing gel and electrophoresed in0.5 3 TBE buffer for 2–3 h at 40 watts in a 4 °C cold room. The primerpair with the sequences CGGAGCCTGCCATTCAGAGAAAT (forward)and CTTGCAATGTCCTGGAGGTGGAA (reverse) identified polymor-phisms between C57BL/6J and Mus spretus, and was used to analyzeDNA prepared from the BSS backcross (15) (Fig. 7). The strain distri-bution pattern was analyzed using the Map Manager Program (16).Xenopus Oocyte Microinjection and Transport Measurement—Xeno-
pus oocyte expression was performed as described previously (28). Man-ually defolliculated oocytes were injected with 40–50 ng of rat kidneymedulla mRNA or NKT cRNA. Five days after injection, the uptake ofradioisotope-labeled substrates was determined. For analyzing urea
transport (positive control), 2.74 mCi of [14C]urea/ml and 1 mM ureawere added to the uptake solution containing 200 mM mannitol, 2 mM
KCl, 1 mM MgCl2, 1 mM CaCl2, 10 mM HEPES, 5 mM Tris (pH 7.4).Uptake was stopped by washing the oocytes with ice-cold uptake solu-tion containing unlabeled urea. Washed oocytes were dissolved in 10%SDS and radioactivity was counted in a scintillation counter. For or-ganic anion and cation uptake, the same procedure was followed exceptthat the uptake solution contained 100 mM NaCl, 2 mM KCl, 1 mM
MgCl2, 1 mM CaCl2, 10 mM HEPES, 5 mM Tris (pH 7.4). The uptake wasstopped by washing the oocytes with ice-cold choline solution (100 mM
choline, 2 mM KCl, 1 mM MgCl2, 1 mM CaCl2, 10 mM HEPES, 5 mM Tris,pH 7.4) and the radioactivity was counted as described above.
RESULTS
Isolation of NKT cDNA—Using a new approach to selectivelyrepresent mammalian protein-coding regions (1), we identifieda novel cDNA with a kidney-specific pattern of expression (Fig.1). This clone is referred to as NKT cDNA.NKT cDNA Nucleotide and Primary Amino Acid Sequence—
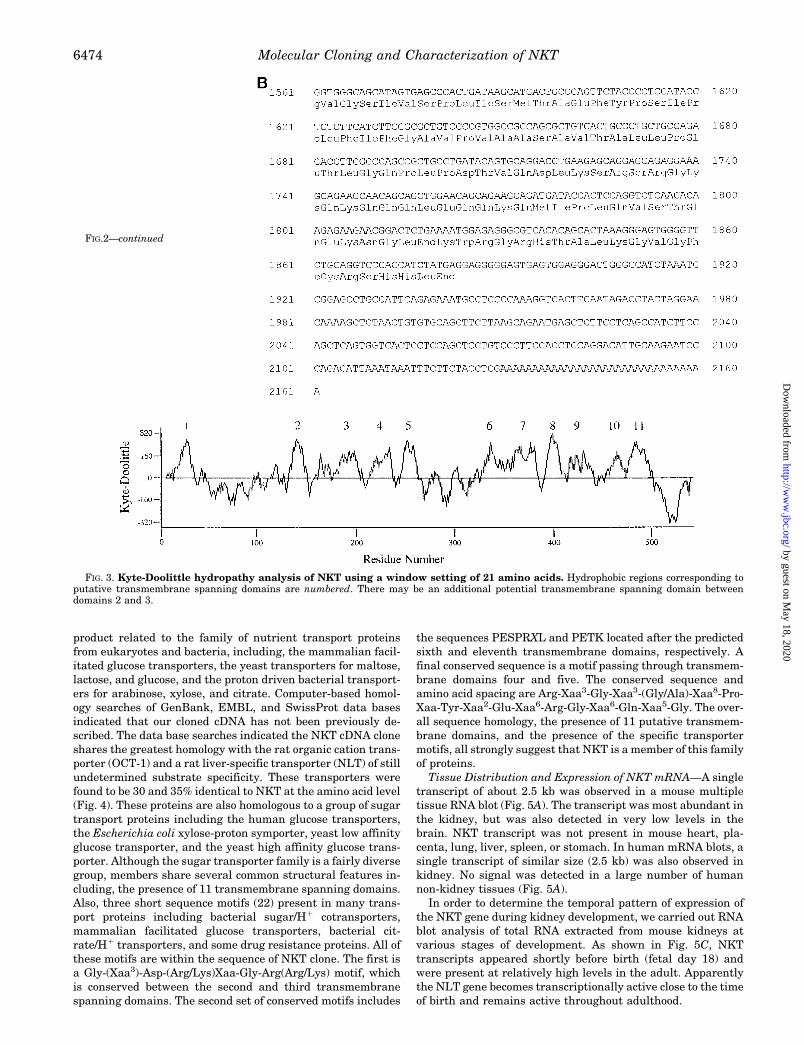
The NKT cDNA is 2161 nucleotides in length and contains botha consensus polyadenylation signal (AATAAA), and a nucleo-tide poly(A) tract defining the 39 end of the clone (Fig. 2). Theopen reading frame is 1638 nucleotides long and encodes aprotein of 546 amino acids. The deduced primary amino acidsequence of NKT is shown in Fig. 2. The deduced amino acidsequence was separately confirmed from a cDNA clone ampli-fied from mouse kidney mRNA. The AUG located in nucleotideposition 182 has the strongest translation initiation consensussequence according to Kozak’s rules and was tentatively as-signed as the first codon (17). An analysis of the primary aminoacid sequence using the Kyte and Doolittle algorithm predicts11 a-helical transmembrane spanning domains (18) (Fig. 3).These same domains were identified as likely transmembranedomains using the Eisenberg algorithm (19). The rather large100-amino acid loop between putative transmembrane regionsone and two is presumably located extracellularly. This loopcontains four N-linked glycosylation consensus sites (Asn-X-Ser/Thr) at positions Asn-56, Asn-86, Asn-91, and Asn-107, aswell as four cysteine residues Cys-49, Cys-78, Cys-99, and
FIG. 1. Portion of a sequencing grade gel showing multipledifferentially expressed bands. The lanes shown correspond to totalRNA from eight mouse tissues: heart (H), brain (B), placenta (P), lung(Lu), liver (Li), spleen (Sp), kidney (K), and stomach (St). These tissueswere reverse transcribed and PCR amplified using the following primerpair TGTGGATGGGGTTG and GTGGTGCTG(G/C)TCAT. The topband in the kidney lane (indicated by arrow) was cloned and sequenced.The preferential expression of this band in kidney was confirmed byNorthern blot.
Molecular Cloning and Characterization of NKT6472
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Cys-122 that may be involved in the formation of disulfidebridges. In addition, two hydroxyl amino acids (Ser-265 andSer-270) located in the large intracellular loop between puta-tive transmembrane domains six and seven represent potential
targets for kinase C phosphorylation (20, 21).Computer Searches and Conserved Motifs—Comparison of
the deduced peptide sequence of this protein with those foundin available data banks revealed that NKT is a novel gene
FIG. 2. Nucleotide sequence and de-duced amino acid sequence of theNKT cDNA.
Molecular Cloning and Characterization of NKT 6473
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from
product related to the family of nutrient transport proteinsfrom eukaryotes and bacteria, including, the mammalian facil-itated glucose transporters, the yeast transporters for maltose,lactose, and glucose, and the proton driven bacterial transport-ers for arabinose, xylose, and citrate. Computer-based homol-ogy searches of GenBank, EMBL, and SwissProt data basesindicated that our cloned cDNA has not been previously de-scribed. The data base searches indicated the NKT cDNA cloneshares the greatest homology with the rat organic cation trans-porter (OCT-1) and a rat liver-specific transporter (NLT) of stillundetermined substrate specificity. These transporters werefound to be 30 and 35% identical to NKT at the amino acid level(Fig. 4). These proteins are also homologous to a group of sugartransport proteins including the human glucose transporters,the Escherichia coli xylose-proton symporter, yeast low affinityglucose transporter, and the yeast high affinity glucose trans-porter. Although the sugar transporter family is a fairly diversegroup, members share several common structural features in-cluding, the presence of 11 transmembrane spanning domains.Also, three short sequence motifs (22) present in many trans-port proteins including bacterial sugar/H1 cotransporters,mammalian facilitated glucose transporters, bacterial cit-rate/H1 transporters, and some drug resistance proteins. All ofthese motifs are within the sequence of NKT clone. The first isa Gly-(Xaa3)-Asp-(Arg/Lys)Xaa-Gly-Arg(Arg/Lys) motif, whichis conserved between the second and third transmembranespanning domains. The second set of conserved motifs includes
the sequences PESPRXL and PETK located after the predictedsixth and eleventh transmembrane domains, respectively. Afinal conserved sequence is a motif passing through transmem-brane domains four and five. The conserved sequence andamino acid spacing are Arg-Xaa3-Gly-Xaa3-(Gly/Ala)-Xaa8-Pro-Xaa-Tyr-Xaa2-Glu-Xaa6-Arg-Gly-Xaa6-Gln-Xaa5-Gly. The over-all sequence homology, the presence of 11 putative transmem-brane domains, and the presence of the specific transportermotifs, all strongly suggest that NKT is a member of this familyof proteins.Tissue Distribution and Expression of NKT mRNA—A single
transcript of about 2.5 kb was observed in a mouse multipletissue RNA blot (Fig. 5A). The transcript was most abundant inthe kidney, but was also detected in very low levels in thebrain. NKT transcript was not present in mouse heart, pla-centa, lung, liver, spleen, or stomach. In human mRNA blots, asingle transcript of similar size (2.5 kb) was also observed inkidney. No signal was detected in a large number of humannon-kidney tissues (Fig. 5A).In order to determine the temporal pattern of expression of
the NKT gene during kidney development, we carried out RNAblot analysis of total RNA extracted from mouse kidneys atvarious stages of development. As shown in Fig. 5C, NKTtranscripts appeared shortly before birth (fetal day 18) andwere present at relatively high levels in the adult. Apparentlythe NLT gene becomes transcriptionally active close to the timeof birth and remains active throughout adulthood.
FIG. 3. Kyte-Doolittle hydropathy analysis of NKT using a window setting of 21 amino acids. Hydrophobic regions corresponding toputative transmembrane spanning domains are numbered. There may be an additional potential transmembrane spanning domain betweendomains 2 and 3.
FIG.2—continued
Molecular Cloning and Characterization of NKT6474
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

In situ hybridization using sense and antisense cRNA onmouse kidney paraffin sections showed that the most intensesignal was present in kidney cortex, following a pattern char-acteristic of proximal tubular localization (Fig. 6). There was nodetectable signal in the glomeruli, distal tubules, or collectingducts. RNA blot analysis done in mouse microdissected kidneyalso showed an intense signal in the cortex, a moderate signalin outer stripe of the outer medulla, a faint signal in innerstripe, and no signal in inner medulla (Fig. 5B), once moreconsistent with a proximal tubular distribution.Chromosomal Localization of the NKT Gene—SSCP analysis
was used to map the chromosomal localization of NKT (14, 23).Two primer pairs corresponding to non-overlapping regions of
the 39-untranslated region of NKT were analyzed and found toidentify SSCPs between mouse species (see “Materials andMethods” and Fig. 7). The BSS interspecific backcross wasgenotyped and the strain distribution pattern, which wereidentical for the two primer pairs, was analyzed using the MapManager program. NKT was found to map to chromosome 19with a LOD likelihood score of 27.1. No recombinants werefound between NKT and D19Mit32 in 94 progeny; NKT is there-fore the most proximal gene mapped on chromosome 19 on theBSS cross. This is the very site to which a number of unknownmurine mutations have been mapped (see “Discussion”).Evaluation of Microinjected NKT cRNA Capacity to Trans-
port in Xenopus Oocytes—Since TEA and PAH are the proto-type substrates for organic cation and organic anion transport-ers, respectively (8, 24), and the organic anion transporter wasreported to transport PAH by exchanging with intracellulara-ketoglutarate (30), we examined these possibilities with NKTcRNA-injected oocytes under different conditions. The uptakeof [14C]urea into rat medulla mRNA-injected oocytes was usedas a positive control. Uptake of [14C]urea (1 mM) into rat me-dulla mRNA-injected oocytes resulted in a '4-fold increaseabove that of water control level. This is consistent with aprevious study reported by You and co-workers (29). Neverthe-less, NKT cRNA injected oocytes did not demonstrate a signif-icant amount of transport of either PAH and TEA under 100 mM
concentration (Fig. 8a). When the concentration of these sub-strates (and, in addition, cimetidine) were increased to 1 mM
(Fig. 8b), still no transport was activity observed. Next, wepreincubated 100 mM a-ketoglutarate for 30 min before theuptake of 100 mM PAH was measured (Fig. 8c); however, wewere not able to show any transport activity. Our results sug-gest that NKT cRNA-injected oocytes do not demonstrate sig-
FIG. 4. Comparison of the NKT amino acid sequence withOCT-1 and NLT transport proteins. At the amino acid level, OCT-1and NLT are 30 and 35% identical to NKT, respectively. Conservedresidues are bold and underlined. All three proteins also share commonstructural features, including: 11 putative a-helical transmembranespanning domains; a large extracellular loop between putative trans-membrane regions 1 and 2; several N-linked glycosylation consensussites and four conserved cysteine residues located in this large extra-cellular loop; a large intracellular loop located between putative trans-membrane domains 6 and 7, with several potential target sites forprotein kinase C; intracellular location of both the amino and carboxyltermini.
FIG. 5. Tissue distribution and developmental expression ofNKT in mouse and human. In the mouse multiple tissue RNA blot(Panel A), a 2.5-kb transcript is only present in kidney (Ki), and, atmuch lower level, in brain (Br). No signal was detected in mouse heart(He), placenta (Pl), lung (Lu), liver (Li), spleen (Sp), or stomach (St). Asingle transcript of similar size (2.5 kb) was also observed in humankidney (Panel A), but no signal was detected in human brain, heart,placenta, lung, liver, skeletal muscle (Sk), pancreas (Pa), spleen, thy-mus (Th), prostate (Pr), testis (Te), ovary (Ov), small intestine (Sm),colon (Co), and peripheral blood leukocyte (Pb). Within the kidney(Panel B), the NKT was mainly localized to the cortex (Co) and the outerstripe (OS) of the outer medulla. RNA blot analysis of mouse kidneys atvarious stages of development (Panel C) shows that NKT becomestranscriptionally active close to the time of birth (fetal day 18).
Molecular Cloning and Characterization of NKT 6475
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

nificant transport for the substrates tested, at least under theconditions employed here (see “Discussion”).
DISCUSSIONRenal tubular cells are responsible for the reabsorption and
secretion of numerous substrates. These cells are highly polar-
ized with unique species of transporters localized to either thebasolateral or the apical domains of their plasma membranes.Using a new approach to selectively represent mammalianprotein-coding regions (1), we have identified a novel transportprotein which, by Northern analysis, is almost exclusively ex-
FIG. 6. In situ hybridization ofmouse kidney sections with anti-sense and sense probes of NKT cRNA.a, a parasagittal section of mouse kidneyshowing a strong signal for NKT probe incortex but not in the medulla (m). Scalebar, 100 mm. b, a close-up of the cortexregion shown in a. No specific signalswere detected in glomeruli (arrows). Scalebar, 25 mm. c, mouse kidney hybridizedwith sense probe of NKT. No specific sig-nals were detected. Scale bar, 100 mm.
Molecular Cloning and Characterization of NKT6476
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

pressed in the kidney.The sequence analyses of NKT suggest that it belongs to a
recently identified subgroup of transport proteins. One memberof this subgroup (OCT-1) has been shown to translocate hydro-phobic and hydrophilic organic cations of different structuresover the basolateral membrane of renal proximal tubules andhepatocytes (8). OCT-1 is currently considered a new prototypeof polyspecific transporters likely to be important in drug elim-ination, although presently little is known of its specific role invarious tissues. Xenobiotics and their metabolites are trans-ported mainly by the organic anion (PAH) and organic cationtransport systems, and there exist substrates that interactwith both the transporter for organic anions and that for or-ganic cations (2, 3). Neither transporter appears to detect thedegree of ionization in bulk solution, and they also acceptnonionizable substrates (4). Since these two transport systems(cationic and anionic) share so many common functional fea-tures, it is possible that they may also resemble each other atthe molecular level. Functional expression of renal organicanion transport in Xenopus laevis oocytes injected with ratkidney poly(A)1 RNA has shown that the active species withrespect to PAH transport was in the range of 1.8 to 3.5 kb (24).The size of NKT (2.5 kb) is within this range. Deduced aminoacid sequence analysis showed that four cysteine residues areconserved among NKT, NLT, and OCT-1. Previous studies ofthe effect of N-ethylmaleimide (NEM), an irreversible sulfhy-dryl modifying reagent, on the transport of organic cations inthe renal basolateral membrane imply that inactivation in-volves the binding of at least four molecules of N-ethylmaleim-
ide per active transport unit. This is most consistent with thepresence of four sulfhydryl groups at this site. The capability oforganic cations to alter the susceptibility to sulfhydryl modifi-cation suggests that these groups may have a dynamic role inthe transport process (25). For these and other reasons alreadydiscussed, NKT was considered to be a strong candidate for theimportant task of drug elimination by the kidney, a majorfunction of the organ.PAH and TEA are the prototype substrates of organic cation
and organic anion transporters. Therefore, we tested thesepossibilities by measuring the uptake of the radiolabeled PAH,TEA as well as cimetidine into Xenopus oocytes. However,under the conditions we employed (including measurements inthe presence of a-ketoglutarate (30)), we were unable to showany transport activity. At present it is unclear whether thisnegative result was due to suboptimal conditions for NKT
FIG. 7. A, SSCP analysis of C57BL/6J (B), M. spretus (S), and 21progeny of the ((C57BL/6J 3 M. spretus)F1 3 M. spretus). BSS inter-specific backcross is shown. Since this is a backcross, all progeny carryanM. spretus allele, and the mice are scored for the presence or absenceof a C57BL/6J allele inherited from the F1 parent. B, chromosome 19.The figure on the left is concensus map obtained from the MGD Database at the Jackson Laboratory. Mutants (nmd, mdf, ocd, oc, Dc, anddn), genes (Gstp1 and Adrbk1), and MIT microsatellite markers areshown. The figure on right is the data obtained for the analysis of Ntkand adjacent MIT microsatellite markers in the BSS cross.
FIG. 8. Expression of NKT in Xenopus oocytes. Filled bars rep-resent oocytes injected with rat kidney medulla mRNA or NKT cRNA(50 ng). Open bars represent water-injected oocytes. Columns representthe mean 1 S.E. (n 5 5–8 oocytes). a, rat kidney medullamRNA-injected oocytes induced [14C]urea uptake (1 mM) '4-fold abovewater control level. NKT cRNA-injected oocytes did not induce signifi-cant amount of uptake of 14C-labeled PAH and TEA (100 mM). b, NKTcRNA-injected oocytes did not induce significant amount of uptake of14C-labeled PAH, TEA, and cimetidine (1 mM, Amersham). c, NKTcRNA-injected oocytes did not induce a significant amount of uptake of14C-labeled PAH (100 mM) with the preincubation of 100 mM a-keto-glutarate (a-KG) (Sigma) for 30 min.
Molecular Cloning and Characterization of NKT 6477
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

transport (despite robust transport in the positive control),poor expression or insertion of an inactive (incompletely proc-essed) transporter protein, or because NKT might transportother substrates than those we have examined so far. Expres-sion of NKT protein in other mammalian cells such as COS-7cells may be required to answer these questions.Assuming NKT is a transporter, the forementioned data
raises the possibility that it may have significantly differentsubstrates from OCT-1. The low overall homology of the NKTsequence to the hexose transporters argues against its partic-ipation in sugar transport. When considering possible sub-strates for NKT, it is of importance to keep in mind that theexpression of NKT appears to be kidney-specific, or at leastrelatively so. Both NLT and OCT-1 are located within thebasolateral membrane. Other less closely related members ofthe family of nutrient transport proteins (e.g. SV2) are ex-pressed in intracellular organelles or the membrane of synap-tosomes rather than the plasma membrane (26). The subcellu-lar location of NKT awaits the development of antibodies tocarry out indirect immunofluorescent detection of the proteinin kidney sections. In situ hybridization in mouse kidneyshowed that NKT is expressed in proximal tubules, but not indistal tubules, collecting ducts, or glomeruli (a pattern similarto that observed for OCT1). Confirmatory evidence for proximaltubular distribution was also obtained by Northern blot anal-ysis with positive signals obtained in cortex and outer stripe,but not inner stripe and inner medulla. But unlike OCT-1 andNLT, transcripts were also detected in brain (mouse but nothuman), while no signal was found in liver (NLT and OCT-1) orsmall intestine (OCT-1). Of special interest is that NKT isexpressed preferentially in the kidney and that its expressionis developmentally regulated. The NKT transcripts appearshortly before birth. Studies of gene expression during kidneydevelopment have shown that genes appearing late in kidneydevelopment or at birth represent markers for highly differen-tiated kidney tubular cells, and these markers are often lostduring neoplastic transformation. Therefore, NKT cDNA inaddition to being related to organic ion transporters representsa new molecular marker for the terminally differentiatednephron.SSCP analysis was used to localize NKT to mouse chromo-
some 19, tightly linked to D19Mit32. The human homologs ofgenes in this region such as Gstp1 and Adrbk1 map to 11q13(27). Since subchromosomal linkage relationships are con-served in many cases between mouse and man, this resultsuggests that the human homolog of NKT will be found in thisregion. A number of interesting mouse mutations have beenmapped to the proximal portion of chromosome 19, including
several that affect neurological function or development (neu-romuscular degeneration (nmd), muscle deficient (mdf),Dancer (Dc), deafness (dn)) or bone development (osteochon-drodystrophy (ocd), osteosclerosis (oc)). Whether NKT plays arole in these murine mutations awaits further analyses.
Acknowledgments—We thank Christine Miller and Duane Hinds forexpert technical assistance, and Lucy Rowe for help with data analysis.
REFERENCES
1. Lopez-Nieto, C. E., and Nigam, S. K. (1996) Nature Biotech. 14, 857–8612. Ullrich, K. J., Rumrich, G., David, C., and Fritzsch, G. (1993) Pflugers Arch.
425, 280–2993. Ullrich, K. J., Rumrich, G., David, C., and Fritzsch, G. (1993) Pflugers Arch.
425, 300–3124. Ullrich, K. J., and Rumrich, G. (1992) Pflugers Arch. 421, 286–2885. Ullrich, K. J., and Rumrich, G. (1993) Clin. Investig. 71, 843–8486. Hohage, H., Morth, D. M., Querl, I. U., and Greven, J. (1994) J. Pharmacol.
Exp. Ther. 268, 897–9017. Hohage, H., Lohr, M., Querl, I. U., and Greven, J. (1994) J. Pharmacol. Exp.
Ther. 269, 659–6648. Grundemann, D., Gorboulev, V., Gambaryan, S., Veyhl, M., and Koepsell, H.
(1994) Nature 372, 549–5529. Simonson, G. D., Vincent, A. C., Roberg, K. J., Huang, Y., and Iwanij, V. (1994)
J. Cell Sci. 107, 1065–107210. Chirgwin, J., Przybyla, A., MacDonald, R., and Rutter, W. J. (1979) Biochem-
istry 18, 5294–529911. Gish, W., and States, D. J. (1993) Nat. Genet. 3, 266–27212. Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990) J.
Mol. Biol. 215, 03–1013. Wedden, S., Pang, K., and Eichele, G. (1989) Development 105, 639–65014. Beier, D. R. (1993) Mamm. Genome 4, 627–63115. Rowe, L. B., Nadeau, J. H., Turner, R., Frankel, W. N., Letts, V. A., Eppig, J.
T., Ko, M. S. H., Thurston, S. J., and Birkenmeier, E. H. (1994) Mamm.Genome 5, 253–274
16. Manley, K. F. (1991) Mamm. Genome 4, 303–31317. Kozak, M. (1987) Nucleic Acids Res. 15, 8125–813218. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157, 105–13219. Eisenberg, D., Schwarz, E., Komaromy, M., and Wall, R. (1984) J. Mol. Biol.
179, 125–14220. Kishimoto, A., Nishiyama, K., Nakanishi, H., Uratsuji, Y., Nomura, H.,
Takeyama, Y., and Nishizuka, Y. (1985) J. Biol. Chem. 260, 12492–1249921. Kennelly, P. J., and Krebs, E. G. (1991) J. Biol. Chem. 266, 15555–1555822. Gingrich, J. A., Andersen, P. H., Tiberi, M., Mestikawy, S. E., Jorgensen, P. N.,
Fremeau, R. T., and Caron, M. G. (1992) FEBS Lett. 312, 115–12223. Beier, D. R., Dushkin, H., and Sussman, D. J. (1992) Proc. Natl. Acad. Sci.
U. S. A. 89, 9102–910624. Wolff, N. A., Philpot, R. M., Miller, A. S., and Pritchard, J. B. (1992)Mol. Cell.
Biochem. 114, 35–4125. Zimmerman, W. B., Byun, E., McKinney, T. D., and Sokol, P. P. (1991) J. Biol.
Chem. 266, 5459–546326. Bajjalieh, S. M., Peterson, K., Shinghal, R., and Sheller, R. H. (1992) Science
257, 1271–127327. Rochelle, J. M., Watson, M. L., Oakey, R. J., and Seldin, M. F. (1992) Genomics
14, 26–3128. You, G., Lee, W-S., Barros, E. J. G., Kanai, Y., Huo, T-L., Khawaja, S., Wells,
R. G., Nigam, S. K., and Hediger, M. A. (1995) J. Biol. Chem. 270,29365–29371
29. You, G., Smith, G. P., Kanai, Y., Lee, W., Stelzner, M., and Hediger, M. (1993)Nature 365, 844–847
30. Danzler, W. H., Evans, K. K., and Wright, S. H. (1995) J. Pharmacol. Exp.Ther. 272, 663–672
Molecular Cloning and Characterization of NKT6478
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

and Sanjay K. NigamCarlos E. Lopez-Nieto, Guofeng You, Kevin T. Bush, Elvino J. G. Barros, Davio R. Beier
KidneyOrganic Cation Transporter Family That Is Almost Exclusively Expressed in the Molecular Cloning and Characterization of NKT, a Gene Product Related to the
doi: 10.1074/jbc.272.10.64711997, 272:6471-6478.J. Biol. Chem.
http://www.jbc.org/content/272/10/6471Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/272/10/6471.full.html#ref-list-1
This article cites 30 references, 11 of which can be accessed free at
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from