the mammalian amp-activated protein kinase complex … · 2016-12-09 · the mammalian...
TRANSCRIPT

FEBS Letters 588 (2014) 2070–2077
journal homepage: www.FEBSLetters .org
The mammalian AMP-activated protein kinase complex mediatesglucose regulation of gene expression in the yeast Saccharomycescerevisiae
http://dx.doi.org/10.1016/j.febslet.2014.04.0390014-5793/� 2014 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
⇑ Corresponding author. Fax: +46 31 786 2599.E-mail address: [email protected] (S. Hohmann).
1 Present address: Department of Functional Biology, Faculty of Medicine, Univer-sity of Oviedo, Oviedo, Spain.
Tian Ye a, Loubna Bendrioua a, David Carmena b, Raúl García-Salcedo a,1, Peter Dahl a, David Carling b,Stefan Hohmann a,⇑a Department of Chemistry and Molecular Biology, University of Gothenburg, Box 462, S-40530 Göteborg, Swedenb MRC Clinical Sciences Centre, Cellular Stress Group, Imperial College London, Hammersmith Hospital Campus, Du Cane Road, London W12 0NN, UK
a r t i c l e i n f o a b s t r a c t
Article history:Received 9 March 2014Revised 21 April 2014Accepted 24 April 2014Available online 8 May 2014
Edited by Francesc Posas
Keywords:AMP-activated protein kinaseSnf1 protein kinaseInvertase activityMig1 transcriptional repressorSalt stressActive compoundsSaccharomyces cerevisiae
The AMP-activated protein kinase (AMPK) controls energy homeostasis in eukaryotic cells. Here weexpressed hetero-trimeric mammalian AMPK complexes in a Saccharomyces cerevisiae mutantlacking all five genes encoding yeast AMPK/SNF1 components. Certain mammalian complexescomplemented the growth defect of the yeast mutant on non-fermentable carbon sources.Phosphorylation of the AMPK a1-subunit was glucose-regulated, albeit not by the Glc7-Reg1/2phosphatase, which performs this function on yeast AMPK/SNF1. AMPK could take over SNF1 func-tion in glucose derepression. While indirectly acting anti-diabetic drugs had no effect on AMPK inyeast, compound 991 stimulated a1-subunit phosphorylation. Our results demonstrate a remark-able functional conservation of AMPK and that glucose regulation of AMPK may not be mediatedby regulatory features of a specific phosphatase.� 2014 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
The AMP-activated protein kinase (AMPK) is the masterregulator of energy homeostasis in eukaryotic cells [1–3]. AMPKis also involved in the regulation of whole-body energy metabo-lism and responds to hormonal and nutritional signals in thecentral nervous system and peripheral tissues that control foodintake and energy expenditure [2,4]. In yeast, the best known roleof AMPK/SNF1 is to control gene expression in response toavailability of the preferred energy source glucose: glucose repres-sion/derepression [5–7]. However, based on multi-scale modellingof different omics data it appears that the cell-physiological rolesof AMPK and SNF1 are similar [8].
AMPK and SNF1 form hetero-trimeric protein complexes con-sisting of the catalytic a-subunit (in yeast Snf1), the regulatoryand scaffolding b-subunit (Gal83, Sip1 or Sip2) and the regulatory
c-subunit (Snf4) [2,7]. While in yeast only the b-subunit exists inthree isoforms, in mammals each subunit is encoded by multiplegenes (a1, a2, b1, b2, c1, c2, c3). AMPK is activated by phosphor-ylation on T172 within the activation loop of the kinase catalyticdomain. Structural and in vitro analyses of mammalian and yeastAMPK have revealed mechanisms that control AMPK (reviewedin [2,3]). The c-subunit appears to bind adenine nucleotides onthree sites where binding of ADP and AMP stimulate enzyme activ-ity by allosteric activation, promotion of phosphorylation on T172by upstream activating kinase and protection from dephosphoryl-ation by protein phosphatases [9–15]. Yeast and mammalianAMPK appear to differ in the responsiveness to AMP and ADP:AMP may be more effective on AMPK while ADP may be the phys-iological activator of SNF1 [16–19]. Moreover, yeast SNF1 may alsobe activated by ADP-binding to the a-subunit [16]. Activation ofAMPK results in the phosphorylation of downstream targets thatcontrol the rates of ATP-consuming (e.g. fatty acid synthesis) andATP-generating (e.g. fatty acid oxidation) pathways (reviewed in[2,3]). Because of its central roles in cellular and organismal metab-olism AMPK is a potential target for treating metabolic conditionsassociated with type 2 diabetes, the metabolic syndrome, cancer

Table 1Yeast strains used in this study. All strains used have W303-1A background.
Strain ID Genotype Ref.
W303-1A MATa leu2-3,112 trp1-1 can1-100 ura3-1 ade2-1his3-11,15
[61]
YSH1701 (W303-1A) MATa snf1D::KanMX This workYSH2168 (W303-1A) MATa snf1D::KanMX snf4D::KanMX
sip1D::KanMX sip2D::KanMX gal83D::KanMXThis work
YSH2248 (W303-1A) MATa snf1D::KanMX snf4D::KanMXsip1D::KanMX sip2D::KanMX gal83D::KanMXMIG1-HA-TRP1
This work
YSH1832 (W303-1A) MATa reg1D::KanMX snf1D::KanMX This workYSH1969 (W303-1A) MATa reg2D::KanMX snf1D::KanMX This workYSH1971 (W303-1A) MATa reg1D::KanMX snf1D::KanMX This work
T. Ye et al. / FEBS Letters 588 (2014) 2070–2077 2071
and inflammation. Numerous compounds are known to act onAMPK, both indirectly (metformin, phenformin [20–22]) as wellas directly as AMP analogue (AICAR [23,24] or by association withthe carbohydrate-binding site of the b-subunit, such as compoundsA-769662 and 991 as well as salicylic acid [25–28].
In yeast, glucose starvation leads to rapid phosphorylation ofSnf1 on T210 (the analogous site to T172 in AMPK) in the activa-tion loop by upstream activating kinases (reviewed in [5–7]).While it now has been established that ADP activates yeast SNF1[16,17] a firm correlation between the cellular adenosine nucleo-tide levels and Snf1 activity has not yet been established. Glucoseregulation of the Snf1 phosphorylation state seems to be mediatedvia access of the protein phosphatase Glc7 to the SNF1 complex[29]. Glc7 is targeted to Snf1 by two proteins, Reg1 and Reg2,where Reg1 plays a major role [29–31]. In the presence of glucose,when Snf1 is inactive, the transcriptional regulator Mig1 binds totarget gene promoters, such as SUC2 encoding invertase, to represstranscription [32–35]. At low glucose levels, or its completeabsence, Snf1 is activated and phosphorylates Mig1, which resultsin dissociation of the co-repressors Ssn6/Cat8 and Tup1, export ofMig1 from the nucleus to the cytosol and derepression of glu-cose-repressed genes [36–38]. Phosphorylation of Snf1 appears tobe required but not sufficient for glucose derepression since Snf1activated by salt stress or genetic changes in the presence of glu-cose does not lead to Mig1 phosphorylation [39,40]. Hence, itappears that glucose derepression via the Snf1–Mig1 pathway isa multi-step process.
Given the overall conservation of AMPK across eukaryotes it istempting to test if mammalian AMPK could replace SNF1 in yeast.Yeast has been used successfully as a host for human/mammalianproteins to study their role, structure and function [41]. Thisapproach is complicated by the fact that AMPK is a protein com-plex consisting of three different subunits. Replacement of justthe a-subunit has apparently been tried by researchers in thefield, including ourselves, but failed (and therefore not reportedin the literature). Expression in yeast of a chimeric yeast/mamma-lian a-subunit only resulted in partial complementation of theyeast snf1D mutant [42]. We therefore decided to clear yeastfrom all endogenous SNF1 components and replace the entirecomplex by expression in different combinations mammaliansubunits.
2. Materials and methods
2.1. Yeast strains and plasmids
The Saccharomyces cerevisiae strains used in this study are iso-genic to W303-1A (Table 1). Single deletion mutants for SNF1,SNF4, SIP1, SIP2, GAL83, REG1 and REG2 were constructed in wildtype W303-1A by site-directed replacement with a KanMX dele-tion cassette derived from the global yeast knock-out collection[43]. Strains expressing an HA-tagged Mig1 carry an integratedMIG1-HA-TRP1 construct. The deletion/insertion of genes was con-firmed by PCR. Multiple deletion mutants were then obtained viaserial crosses.
Expression plasmids for mammalian AMPK subunits were gen-erated in the vectors pYX212 (URA3), pYX222 (HIS3) and pYX242(LEU2). These vectors are based on the 2 lm plasmid backbone(multi-copy) and employ the strong constitute yeast TPI1 (triose-phosphate isomerase) promoter. cDNAs were derived from rat(a1, a2, b1 and c1) and from human (b2, c2 and c3), respectively.The cDNA for subunit a1 with N-terminal myc-tag was cloned intoEcoRI/Xho1-cut pYX212. The cDNA for subunit a2 with N-terminalmyc-tag was cloned into EcoRI/-cut pYX212. The cDNA for subunitb1 was cloned into NcoI-cut pYX222. The cDNA for subunit b2 wascloned into pYX222 as EcoRI/XhoI fragment with restrictions sites
introduced by PCR. The cDNA for subunit c1 with N-terminalFLAG-tag was cloned into pYX242 as AvrII/XhoI fragment withrestriction sites introduced by PCR. The cDNA for subunits c2and c3 with N-terminal FLAG-tag were cloned into pYX242 asEcoRI/XhoI fragments.
2.2. Growth conditions and growth assays
Yeast cells were routinely grown in medium either containing2% peptone and 1% yeast extract supplemented with 2% glucoseas a carbon source (YPD) or containing YNB (yeast nitrogen base)supplemented with necessary amino acid and with 2% glucose,2% raffinose or 3% ethanol/2% glycerol as carbon sources. Selectionand growth of transformants carrying a plasmid was performed inyeast nitrogen base medium (YNB) supplemented with therequired amino acid [44]. Plate growth assays were performed bypre-growing cells in YPD medium or YNB medium supplementedwith the required amino acid. Cells were re-suspended in the samemedium to an optical density at 600 nm (OD600) of 1.0. Five micro-liters of a 10-fold serial dilution of this culture were spotted ontoagar plates supplemented with 2% glucose, 2% raffinose, 3% etha-nol/2% glycerol and 1 M NaCl (2% glucose as carbon source), asindicated. Growth was monitored after 2–3 days at 30 �C.
2.3. Western blot analysis
Cells were grown to OD600 = 0.5–0.8, harvested by centrifuga-tion, re-suspended in 500 ll of 2 M NaOH supplemented with 7%2-mercaptoethanol and incubated 2 min at room temperature.Five-hundred microliters of 50% TCA (trichloroacetic acid) wasadded, samples were vortexed and sedimented. Samples werewashed in 500 ll of 1 M Tris (pH = 7.5) and re-suspended in120 ll of 1� SDS sample buffer (62.5 mM Tris–HCl pH 6.8, 3%sodium dodecyl sulfate [SDS], 10% glycerol, 5% b-Mercaptoethanol,0.001% bromophenol blue) and incubated for 5 min at 100 �C. Totalprotein was measured after protein extraction using the BioRadcolorimetric assay (Art No. 500-0111), equal amounts were loadedin each lane, separated by SDS–PAGE using 7.5% polyacrylamidegels, transferred to nitrocellulose membranes and used forimmuno-detection as described below. The Odyssey infrared imag-ing system was used for visualisation following the instruction ofthe manufacturer (LI-COR Biosciences, Santa Clara, CA).
2.3.1. Detection of AMPK subunitsCells were grown in 10 ml cultures and 100–200 lg of protein
extracts were used for gel electrophoresis. Immuno-reagents todetect AMPK subunits were described previously. The a-subunitwas detected using a mouse monoclonal anti-c myc antibody(1:1000, Roche, Basel). The b-subunit was detected by a rabbit anti-serum raised against a GST-fusion of rat AMPK-b and detects both

2072 T. Ye et al. / FEBS Letters 588 (2014) 2070–2077
b1 and b2 [45]. The c-subunit was detected using a mouse mono-clonal anti-FLAG (Sigma–Aldrich, St. Louis, MO).
2.3.2. Detection of Mig1Cells were grown in 10 ml cultures and 40 lg of protein extract
were used for gel electrophoresis. Mig1 was analysed by immuno-blotting using anti-HA antibody (1:2,000; Santa Cruz Biotechnol-ogy, Dallas TX) and a secondary goat anti-mouse IRDye-800CW(1:15,000, Li-COR Biosciences).
2.3.3. Phosphorylation of Snf1Cells were grown in 10 ml cultures and 150 lg of protein were
applied to gel electrophoresis. Proteins were detected using rabbitanti-phospho-Snf1 (1:1000) and mouse anti-HA (1:2000) antibod-ies. Primary antibodies were detected simultaneously with goatanti-mouse IRDye-800CW (1:15,000) and anti-rabbit IRDye-680CW (1:15,000, LI-COR Biosciences).
2.3.4. Phosphorylation of AMPKCells were grown in 10 ml cultures and 100 lg of the superna-
tant were separated by SDS–PAGE. The a-subunit of AMPK wasdetected using simultaneously a monoclonal rabbit anti-Thr172Ig (1:1000; Cell Signalling Technology, Danvers, MA) and a mouseanti-c-myc Ig (1:1000; Roche). The primary antibodies were simul-taneously detected using a goat anti-rabbit (680) and a goat anti-mouse IRDye-800CW (1:15,000; LI-COR Biosciences).
2.4. Invertase activity assay
Cells were grown on 4% glucose and then half of the cells wereshifted to 0.05% glucose. Samples were harvested by centrifugationand washed three times with ice cold water. Protein extracts andmeasurements of invertase activity were performed as describedpreviously [46,47]. The protein concentration was determined byusing the DC protein assay kit (Bio-Rad, Hercules, CA).
Fig. 1. Expression of AMPK subunits in yeast. Cells were transformed with plasmids expconstitutive TPI1 promoter and grown on 4% glucose medium to OD600 = 0.8. Half of tcentrifugation, proteins were extracted, equal amounts were loaded on each lane and prsubunit, (B) an antiserum raised against a b1-GST was used to detect the b1-subunit andexpressed under high and low glucose conditions at the similar levels.
3. Results
3.1. Mammalian AMPK kinase complexes are functional in yeast
To investigate the role of the mammalian AMPK kinase complexin yeast, the genes encoding the five SNF1 subunits were deleted togenerate a quintuple snf1D gal83D sip1D sip2D snf4D mutant.cDNAs encoding human b2, c2 and c3 as well as rat a1, a2,b1and c1, AMPK subunits were cloned into a series of yeast expres-sion vectors (pYX212, pYX222, pYX242) under control of the strong,constitutively active TPI1 promoter. Each expression vector carriedcDNAs for one a-, b- or c-subunit. Three plasmids in twelve combi-nations were simultaneously transformed into yeast cells toexpress all possible combinations of AMPK subunits in the quintu-ple snf mutant. The expression of AMPK complexes was monitoredemploying immune-reagents against myc-tagged a1, against b1,and against FLAG-tagged c1, c2, c3 subunits (Fig. 1). All subunitstested except c2 were expressed at similar levels both in high andlow glucose growth medium. Expression of the myc-tagged a2 sub-unit was undetectable (data not shown) for unknown reasons.
In order to test the function of mammalian AMPK complexes inyeast, transformants were spotted as dilution series on plasmid-selective plates containing as carbon/energy sources glucose (con-trol), raffinose (requires Snf1 and SUC2 expression) and ethanol/glycerol (requires Snf1 and expression of gluconeogenic genes;Fig. 2A). The control snf1 strain transformed with empty vectordid not or very poorly grow on raffinose or glycerol/ethanol plates.However, when transformed with certain combinations of AMPKsubunit expression constructs the strain grew almost as well asthe wild type control. All a1 subunit complexes except those withc2 subunit (which was not or poorly expressed) promoted growthon raffinose and ethanol/glycerol. None of the a2 subunit com-plexes conferred growth, consistent with the apparent lack ofexpression (data not shown). Taken together, combinations con-taining the a1-, b1- or b2- and c1- or c3- subunits mediated growth
ressing different combinations of AMPK a- b- and c-subunits under control of thehe culture was shifted to 0.05% glucose medium for 2 h. Cells were harvested byoteins were separated by SDS–PAGE. (A) An anti-myc Ig was used to detect the a1-(C) an anti-FLAG was used to detect c-subunits. All subunits tested except c2 were
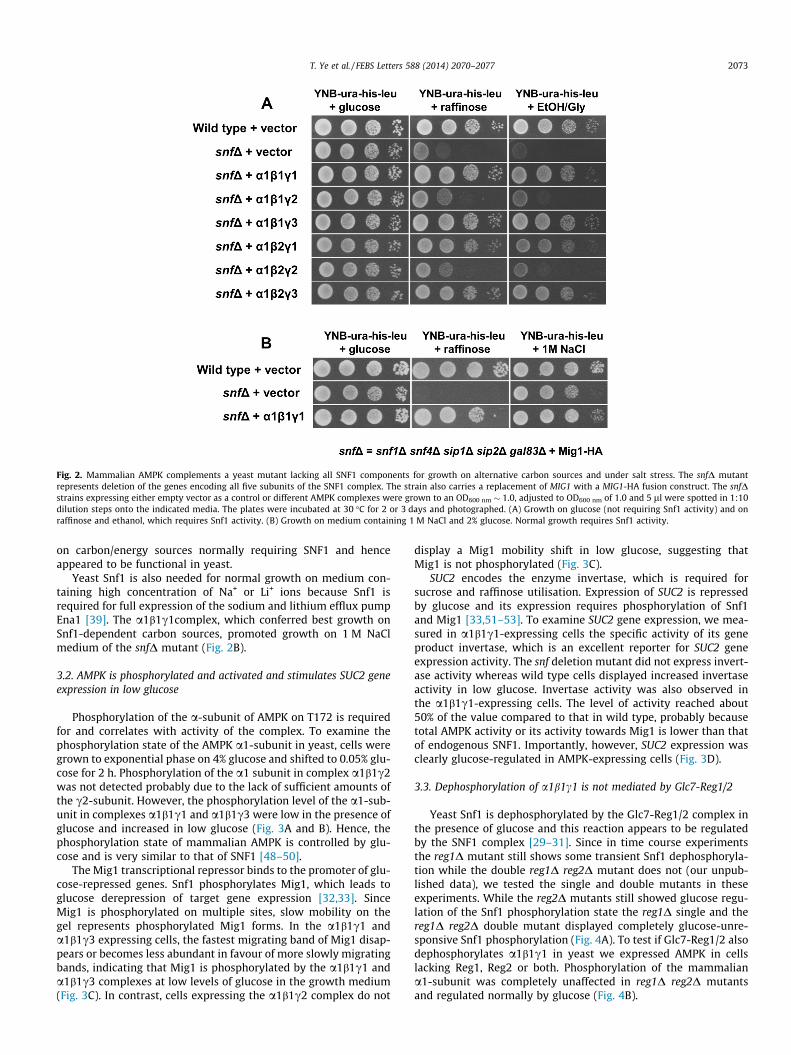
Fig. 2. Mammalian AMPK complements a yeast mutant lacking all SNF1 components for growth on alternative carbon sources and under salt stress. The snfD mutantrepresents deletion of the genes encoding all five subunits of the SNF1 complex. The strain also carries a replacement of MIG1 with a MIG1-HA fusion construct. The snfDstrains expressing either empty vector as a control or different AMPK complexes were grown to an OD600 nm � 1.0, adjusted to OD600 nm of 1.0 and 5 ll were spotted in 1:10dilution steps onto the indicated media. The plates were incubated at 30 �C for 2 or 3 days and photographed. (A) Growth on glucose (not requiring Snf1 activity) and onraffinose and ethanol, which requires Snf1 activity. (B) Growth on medium containing 1 M NaCl and 2% glucose. Normal growth requires Snf1 activity.
T. Ye et al. / FEBS Letters 588 (2014) 2070–2077 2073
on carbon/energy sources normally requiring SNF1 and henceappeared to be functional in yeast.
Yeast Snf1 is also needed for normal growth on medium con-taining high concentration of Na+ or Li+ ions because Snf1 isrequired for full expression of the sodium and lithium efflux pumpEna1 [39]. The a1b1c1complex, which conferred best growth onSnf1-dependent carbon sources, promoted growth on 1 M NaClmedium of the snfD mutant (Fig. 2B).
3.2. AMPK is phosphorylated and activated and stimulates SUC2 geneexpression in low glucose
Phosphorylation of the a-subunit of AMPK on T172 is requiredfor and correlates with activity of the complex. To examine thephosphorylation state of the AMPK a1-subunit in yeast, cells weregrown to exponential phase on 4% glucose and shifted to 0.05% glu-cose for 2 h. Phosphorylation of the a1 subunit in complex a1b1c2was not detected probably due to the lack of sufficient amounts ofthe c2-subunit. However, the phosphorylation level of the a1-sub-unit in complexes a1b1c1 and a1b1c3 were low in the presence ofglucose and increased in low glucose (Fig. 3A and B). Hence, thephosphorylation state of mammalian AMPK is controlled by glu-cose and is very similar to that of SNF1 [48–50].
The Mig1 transcriptional repressor binds to the promoter of glu-cose-repressed genes. Snf1 phosphorylates Mig1, which leads toglucose derepression of target gene expression [32,33]. SinceMig1 is phosphorylated on multiple sites, slow mobility on thegel represents phosphorylated Mig1 forms. In the a1b1c1 anda1b1c3 expressing cells, the fastest migrating band of Mig1 disap-pears or becomes less abundant in favour of more slowly migratingbands, indicating that Mig1 is phosphorylated by the a1b1c1 anda1b1c3 complexes at low levels of glucose in the growth medium(Fig. 3C). In contrast, cells expressing the a1b1c2 complex do not
display a Mig1 mobility shift in low glucose, suggesting thatMig1 is not phosphorylated (Fig. 3C).
SUC2 encodes the enzyme invertase, which is required forsucrose and raffinose utilisation. Expression of SUC2 is repressedby glucose and its expression requires phosphorylation of Snf1and Mig1 [33,51–53]. To examine SUC2 gene expression, we mea-sured in a1b1c1-expressing cells the specific activity of its geneproduct invertase, which is an excellent reporter for SUC2 geneexpression activity. The snf deletion mutant did not express invert-ase activity whereas wild type cells displayed increased invertaseactivity in low glucose. Invertase activity was also observed inthe a1b1c1-expressing cells. The level of activity reached about50% of the value compared to that in wild type, probably becausetotal AMPK activity or its activity towards Mig1 is lower than thatof endogenous SNF1. Importantly, however, SUC2 expression wasclearly glucose-regulated in AMPK-expressing cells (Fig. 3D).
3.3. Dephosphorylation of a1b1c1 is not mediated by Glc7-Reg1/2
Yeast Snf1 is dephosphorylated by the Glc7-Reg1/2 complex inthe presence of glucose and this reaction appears to be regulatedby the SNF1 complex [29–31]. Since in time course experimentsthe reg1D mutant still shows some transient Snf1 dephosphoryla-tion while the double reg1D reg2D mutant does not (our unpub-lished data), we tested the single and double mutants in theseexperiments. While the reg2D mutants still showed glucose regu-lation of the Snf1 phosphorylation state the reg1D single and thereg1D reg2D double mutant displayed completely glucose-unre-sponsive Snf1 phosphorylation (Fig. 4A). To test if Glc7-Reg1/2 alsodephosphorylates a1b1c1 in yeast we expressed AMPK in cellslacking Reg1, Reg2 or both. Phosphorylation of the mammaliana1-subunit was completely unaffected in reg1D reg2D mutantsand regulated normally by glucose (Fig. 4B).
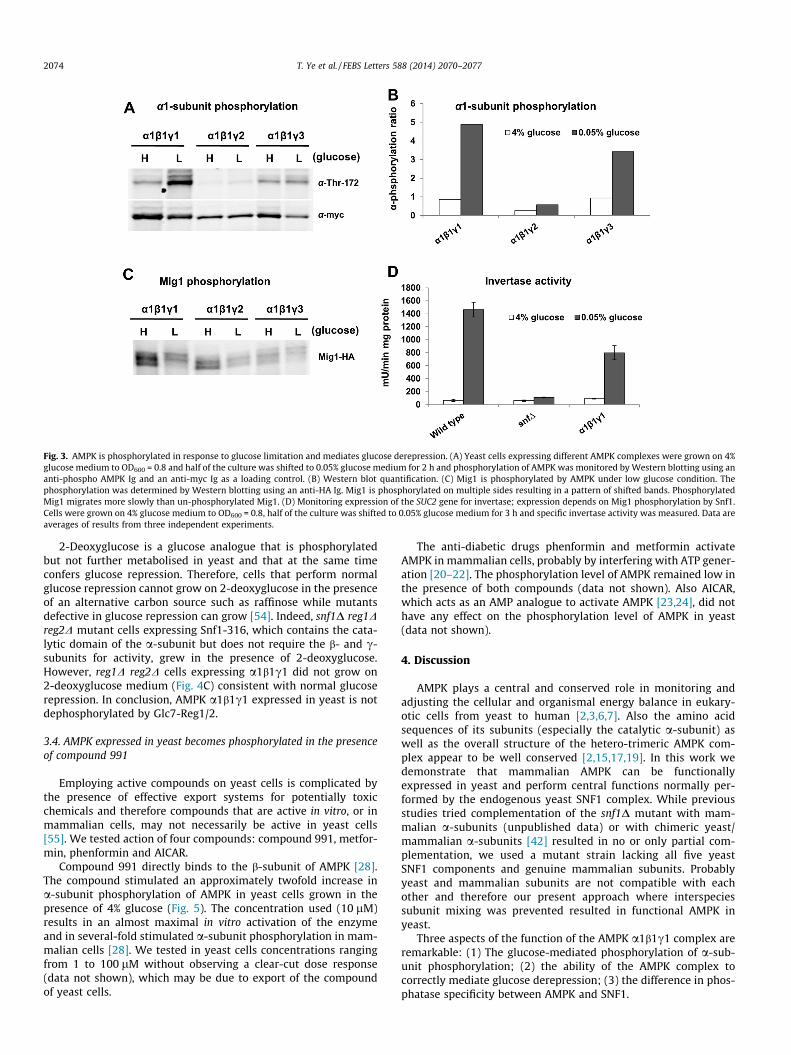
Fig. 3. AMPK is phosphorylated in response to glucose limitation and mediates glucose derepression. (A) Yeast cells expressing different AMPK complexes were grown on 4%glucose medium to OD600 = 0.8 and half of the culture was shifted to 0.05% glucose medium for 2 h and phosphorylation of AMPK was monitored by Western blotting using ananti-phospho AMPK Ig and an anti-myc Ig as a loading control. (B) Western blot quantification. (C) Mig1 is phosphorylated by AMPK under low glucose condition. Thephosphorylation was determined by Western blotting using an anti-HA Ig. Mig1 is phosphorylated on multiple sides resulting in a pattern of shifted bands. PhosphorylatedMig1 migrates more slowly than un-phosphorylated Mig1. (D) Monitoring expression of the SUC2 gene for invertase; expression depends on Mig1 phosphorylation by Snf1.Cells were grown on 4% glucose medium to OD600 = 0.8, half of the culture was shifted to 0.05% glucose medium for 3 h and specific invertase activity was measured. Data areaverages of results from three independent experiments.
2074 T. Ye et al. / FEBS Letters 588 (2014) 2070–2077
2-Deoxyglucose is a glucose analogue that is phosphorylatedbut not further metabolised in yeast and that at the same timeconfers glucose repression. Therefore, cells that perform normalglucose repression cannot grow on 2-deoxyglucose in the presenceof an alternative carbon source such as raffinose while mutantsdefective in glucose repression can grow [54]. Indeed, snf1D reg1Dreg2D mutant cells expressing Snf1-316, which contains the cata-lytic domain of the a-subunit but does not require the b- and c-subunits for activity, grew in the presence of 2-deoxyglucose.However, reg1D reg2D cells expressing a1b1c1 did not grow on2-deoxyglucose medium (Fig. 4C) consistent with normal glucoserepression. In conclusion, AMPK a1b1c1 expressed in yeast is notdephosphorylated by Glc7-Reg1/2.
3.4. AMPK expressed in yeast becomes phosphorylated in the presenceof compound 991
Employing active compounds on yeast cells is complicated bythe presence of effective export systems for potentially toxicchemicals and therefore compounds that are active in vitro, or inmammalian cells, may not necessarily be active in yeast cells[55]. We tested action of four compounds: compound 991, metfor-min, phenformin and AICAR.
Compound 991 directly binds to the b-subunit of AMPK [28].The compound stimulated an approximately twofold increase ina-subunit phosphorylation of AMPK in yeast cells grown in thepresence of 4% glucose (Fig. 5). The concentration used (10 lM)results in an almost maximal in vitro activation of the enzymeand in several-fold stimulated a-subunit phosphorylation in mam-malian cells [28]. We tested in yeast cells concentrations rangingfrom 1 to 100 lM without observing a clear-cut dose response(data not shown), which may be due to export of the compoundof yeast cells.
The anti-diabetic drugs phenformin and metformin activateAMPK in mammalian cells, probably by interfering with ATP gener-ation [20–22]. The phosphorylation level of AMPK remained low inthe presence of both compounds (data not shown). Also AICAR,which acts as an AMP analogue to activate AMPK [23,24], did nothave any effect on the phosphorylation level of AMPK in yeast(data not shown).
4. Discussion
AMPK plays a central and conserved role in monitoring andadjusting the cellular and organismal energy balance in eukary-otic cells from yeast to human [2,3,6,7]. Also the amino acidsequences of its subunits (especially the catalytic a-subunit) aswell as the overall structure of the hetero-trimeric AMPK com-plex appear to be well conserved [2,15,17,19]. In this work wedemonstrate that mammalian AMPK can be functionallyexpressed in yeast and perform central functions normally per-formed by the endogenous yeast SNF1 complex. While previousstudies tried complementation of the snf1D mutant with mam-malian a-subunits (unpublished data) or with chimeric yeast/mammalian a-subunits [42] resulted in no or only partial com-plementation, we used a mutant strain lacking all five yeastSNF1 components and genuine mammalian subunits. Probablyyeast and mammalian subunits are not compatible with eachother and therefore our present approach where interspeciessubunit mixing was prevented resulted in functional AMPK inyeast.
Three aspects of the function of the AMPK a1b1c1 complex areremarkable: (1) The glucose-mediated phosphorylation of a-sub-unit phosphorylation; (2) the ability of the AMPK complex tocorrectly mediate glucose derepression; (3) the difference in phos-phatase specificity between AMPK and SNF1.
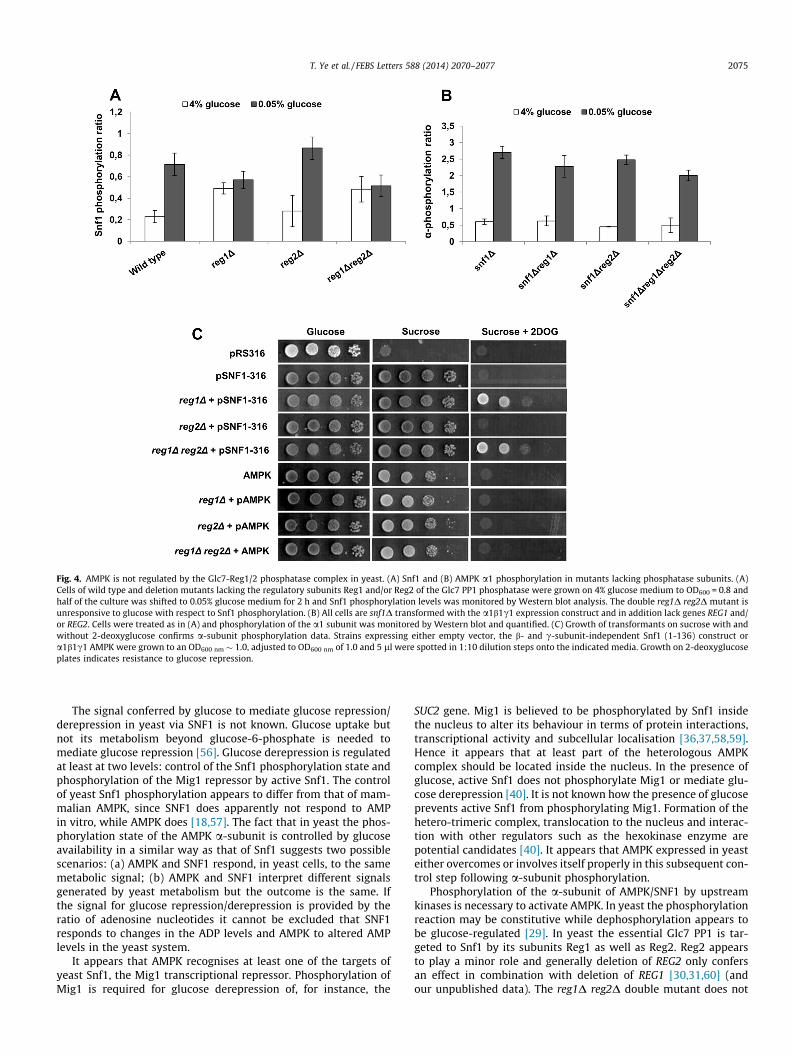
Fig. 4. AMPK is not regulated by the Glc7-Reg1/2 phosphatase complex in yeast. (A) Snf1 and (B) AMPK a1 phosphorylation in mutants lacking phosphatase subunits. (A)Cells of wild type and deletion mutants lacking the regulatory subunits Reg1 and/or Reg2 of the Glc7 PP1 phosphatase were grown on 4% glucose medium to OD600 = 0.8 andhalf of the culture was shifted to 0.05% glucose medium for 2 h and Snf1 phosphorylation levels was monitored by Western blot analysis. The double reg1D reg2D mutant isunresponsive to glucose with respect to Snf1 phosphorylation. (B) All cells are snf1D transformed with the a1b1c1 expression construct and in addition lack genes REG1 and/or REG2. Cells were treated as in (A) and phosphorylation of the a1 subunit was monitored by Western blot and quantified. (C) Growth of transformants on sucrose with andwithout 2-deoxyglucose confirms a-subunit phosphorylation data. Strains expressing either empty vector, the b- and c-subunit-independent Snf1 (1-136) construct ora1b1c1 AMPK were grown to an OD600 nm � 1.0, adjusted to OD600 nm of 1.0 and 5 ll were spotted in 1:10 dilution steps onto the indicated media. Growth on 2-deoxyglucoseplates indicates resistance to glucose repression.
T. Ye et al. / FEBS Letters 588 (2014) 2070–2077 2075
The signal conferred by glucose to mediate glucose repression/derepression in yeast via SNF1 is not known. Glucose uptake butnot its metabolism beyond glucose-6-phosphate is needed tomediate glucose repression [56]. Glucose derepression is regulatedat least at two levels: control of the Snf1 phosphorylation state andphosphorylation of the Mig1 repressor by active Snf1. The controlof yeast Snf1 phosphorylation appears to differ from that of mam-malian AMPK, since SNF1 does apparently not respond to AMPin vitro, while AMPK does [18,57]. The fact that in yeast the phos-phorylation state of the AMPK a-subunit is controlled by glucoseavailability in a similar way as that of Snf1 suggests two possiblescenarios: (a) AMPK and SNF1 respond, in yeast cells, to the samemetabolic signal; (b) AMPK and SNF1 interpret different signalsgenerated by yeast metabolism but the outcome is the same. Ifthe signal for glucose repression/derepression is provided by theratio of adenosine nucleotides it cannot be excluded that SNF1responds to changes in the ADP levels and AMPK to altered AMPlevels in the yeast system.
It appears that AMPK recognises at least one of the targets ofyeast Snf1, the Mig1 transcriptional repressor. Phosphorylation ofMig1 is required for glucose derepression of, for instance, the
SUC2 gene. Mig1 is believed to be phosphorylated by Snf1 insidethe nucleus to alter its behaviour in terms of protein interactions,transcriptional activity and subcellular localisation [36,37,58,59].Hence it appears that at least part of the heterologous AMPKcomplex should be located inside the nucleus. In the presence ofglucose, active Snf1 does not phosphorylate Mig1 or mediate glu-cose derepression [40]. It is not known how the presence of glucoseprevents active Snf1 from phosphorylating Mig1. Formation of thehetero-trimeric complex, translocation to the nucleus and interac-tion with other regulators such as the hexokinase enzyme arepotential candidates [40]. It appears that AMPK expressed in yeasteither overcomes or involves itself properly in this subsequent con-trol step following a-subunit phosphorylation.
Phosphorylation of the a-subunit of AMPK/SNF1 by upstreamkinases is necessary to activate AMPK. In yeast the phosphorylationreaction may be constitutive while dephosphorylation appears tobe glucose-regulated [29]. In yeast the essential Glc7 PP1 is tar-geted to Snf1 by its subunits Reg1 as well as Reg2. Reg2 appearsto play a minor role and generally deletion of REG2 only confersan effect in combination with deletion of REG1 [30,31,60] (andour unpublished data). The reg1D reg2D double mutant does not
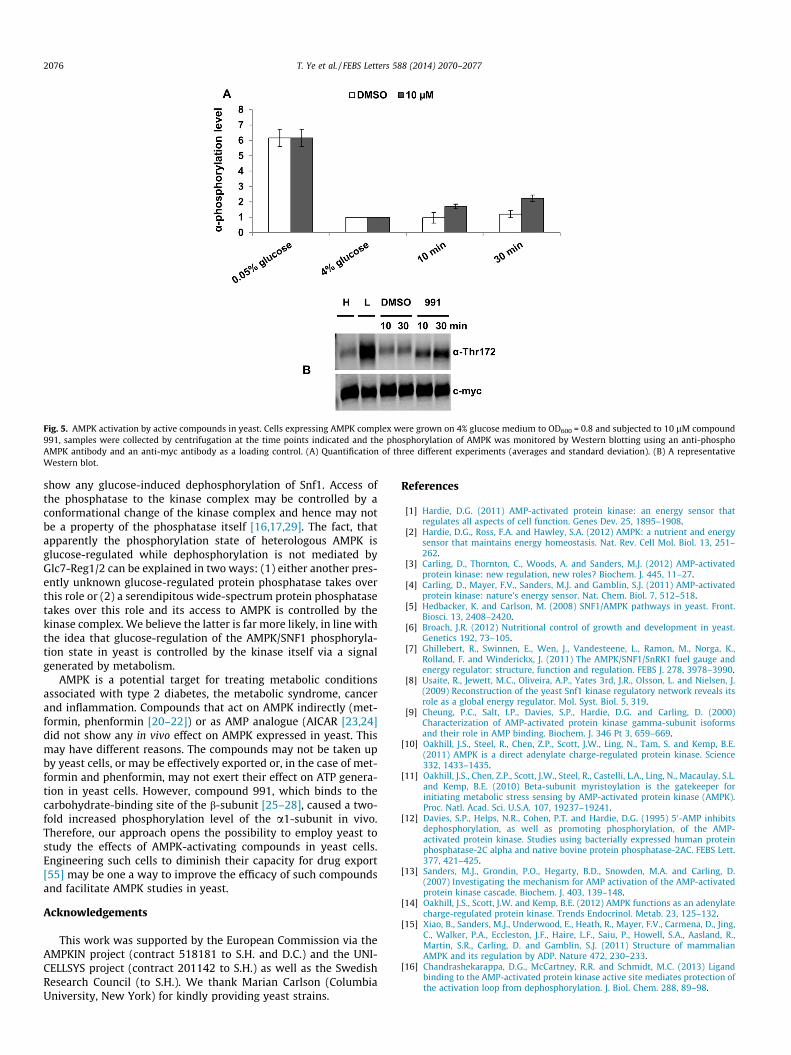
Fig. 5. AMPK activation by active compounds in yeast. Cells expressing AMPK complex were grown on 4% glucose medium to OD600 = 0.8 and subjected to 10 lM compound991, samples were collected by centrifugation at the time points indicated and the phosphorylation of AMPK was monitored by Western blotting using an anti-phosphoAMPK antibody and an anti-myc antibody as a loading control. (A) Quantification of three different experiments (averages and standard deviation). (B) A representativeWestern blot.
2076 T. Ye et al. / FEBS Letters 588 (2014) 2070–2077
show any glucose-induced dephosphorylation of Snf1. Access ofthe phosphatase to the kinase complex may be controlled by aconformational change of the kinase complex and hence may notbe a property of the phosphatase itself [16,17,29]. The fact, thatapparently the phosphorylation state of heterologous AMPK isglucose-regulated while dephosphorylation is not mediated byGlc7-Reg1/2 can be explained in two ways: (1) either another pres-ently unknown glucose-regulated protein phosphatase takes overthis role or (2) a serendipitous wide-spectrum protein phosphatasetakes over this role and its access to AMPK is controlled by thekinase complex. We believe the latter is far more likely, in line withthe idea that glucose-regulation of the AMPK/SNF1 phosphoryla-tion state in yeast is controlled by the kinase itself via a signalgenerated by metabolism.
AMPK is a potential target for treating metabolic conditionsassociated with type 2 diabetes, the metabolic syndrome, cancerand inflammation. Compounds that act on AMPK indirectly (met-formin, phenformin [20–22]) or as AMP analogue (AICAR [23,24]did not show any in vivo effect on AMPK expressed in yeast. Thismay have different reasons. The compounds may not be taken upby yeast cells, or may be effectively exported or, in the case of met-formin and phenformin, may not exert their effect on ATP genera-tion in yeast cells. However, compound 991, which binds to thecarbohydrate-binding site of the b-subunit [25–28], caused a two-fold increased phosphorylation level of the a1-subunit in vivo.Therefore, our approach opens the possibility to employ yeast tostudy the effects of AMPK-activating compounds in yeast cells.Engineering such cells to diminish their capacity for drug export[55] may be one a way to improve the efficacy of such compoundsand facilitate AMPK studies in yeast.
Acknowledgements
This work was supported by the European Commission via theAMPKIN project (contract 518181 to S.H. and D.C.) and the UNI-CELLSYS project (contract 201142 to S.H.) as well as the SwedishResearch Council (to S.H.). We thank Marian Carlson (ColumbiaUniversity, New York) for kindly providing yeast strains.
References
[1] Hardie, D.G. (2011) AMP-activated protein kinase: an energy sensor thatregulates all aspects of cell function. Genes Dev. 25, 1895–1908.
[2] Hardie, D.G., Ross, F.A. and Hawley, S.A. (2012) AMPK: a nutrient and energysensor that maintains energy homeostasis. Nat. Rev. Cell Mol. Biol. 13, 251–262.
[3] Carling, D., Thornton, C., Woods, A. and Sanders, M.J. (2012) AMP-activatedprotein kinase: new regulation, new roles? Biochem. J. 445, 11–27.
[4] Carling, D., Mayer, F.V., Sanders, M.J. and Gamblin, S.J. (2011) AMP-activatedprotein kinase: nature’s energy sensor. Nat. Chem. Biol. 7, 512–518.
[5] Hedbacker, K. and Carlson, M. (2008) SNF1/AMPK pathways in yeast. Front.Biosci. 13, 2408–2420.
[6] Broach, J.R. (2012) Nutritional control of growth and development in yeast.Genetics 192, 73–105.
[7] Ghillebert, R., Swinnen, E., Wen, J., Vandesteene, L., Ramon, M., Norga, K.,Rolland, F. and Winderickx, J. (2011) The AMPK/SNF1/SnRK1 fuel gauge andenergy regulator: structure, function and regulation. FEBS J. 278, 3978–3990.
[8] Usaite, R., Jewett, M.C., Oliveira, A.P., Yates 3rd, J.R., Olsson, L. and Nielsen, J.(2009) Reconstruction of the yeast Snf1 kinase regulatory network reveals itsrole as a global energy regulator. Mol. Syst. Biol. 5, 319.
[9] Cheung, P.C., Salt, I.P., Davies, S.P., Hardie, D.G. and Carling, D. (2000)Characterization of AMP-activated protein kinase gamma-subunit isoformsand their role in AMP binding. Biochem. J. 346 Pt 3, 659–669.
[10] Oakhill, J.S., Steel, R., Chen, Z.P., Scott, J.W., Ling, N., Tam, S. and Kemp, B.E.(2011) AMPK is a direct adenylate charge-regulated protein kinase. Science332, 1433–1435.
[11] Oakhill, J.S., Chen, Z.P., Scott, J.W., Steel, R., Castelli, L.A., Ling, N., Macaulay, S.L.and Kemp, B.E. (2010) Beta-subunit myristoylation is the gatekeeper forinitiating metabolic stress sensing by AMP-activated protein kinase (AMPK).Proc. Natl. Acad. Sci. U.S.A. 107, 19237–19241.
[12] Davies, S.P., Helps, N.R., Cohen, P.T. and Hardie, D.G. (1995) 50-AMP inhibitsdephosphorylation, as well as promoting phosphorylation, of the AMP-activated protein kinase. Studies using bacterially expressed human proteinphosphatase-2C alpha and native bovine protein phosphatase-2AC. FEBS Lett.377, 421–425.
[13] Sanders, M.J., Grondin, P.O., Hegarty, B.D., Snowden, M.A. and Carling, D.(2007) Investigating the mechanism for AMP activation of the AMP-activatedprotein kinase cascade. Biochem. J. 403, 139–148.
[14] Oakhill, J.S., Scott, J.W. and Kemp, B.E. (2012) AMPK functions as an adenylatecharge-regulated protein kinase. Trends Endocrinol. Metab. 23, 125–132.
[15] Xiao, B., Sanders, M.J., Underwood, E., Heath, R., Mayer, F.V., Carmena, D., Jing,C., Walker, P.A., Eccleston, J.F., Haire, L.F., Saiu, P., Howell, S.A., Aasland, R.,Martin, S.R., Carling, D. and Gamblin, S.J. (2011) Structure of mammalianAMPK and its regulation by ADP. Nature 472, 230–233.
[16] Chandrashekarappa, D.G., McCartney, R.R. and Schmidt, M.C. (2013) Ligandbinding to the AMP-activated protein kinase active site mediates protection ofthe activation loop from dephosphorylation. J. Biol. Chem. 288, 89–98.

T. Ye et al. / FEBS Letters 588 (2014) 2070–2077 2077
[17] Mayer, F.V., Heath, R., Underwood, E., Sanders, M.J., Carmena, D., McCartney,R.R., Leiper, F.C., Xiao, B., Jing, C., Walker, P.A., Haire, L.F., Ogrodowicz, R.,Martin, S.R., Schmidt, M.C., Gamblin, S.J. and Carling, D. (2011) ADP regulatesSNF1, the Saccharomyces cerevisiae homolog of AMP-activated protein kinase.Cell Metab. 14, 707–714.
[18] Hardie, D.G., Carling, D. and Gamblin, S.J. (2011) AMP-activated proteinkinase: also regulated by ADP? Trends Biochem. Sci. 36, 470–477.
[19] Xiao, B., Heath, R., Saiu, P., Leiper, F.C., Leone, P., Jing, C., Walker, P.A., Haire, L.,Eccleston, J.F., Davis, C.T., Martin, S.R., Carling, D. and Gamblin, S.J. (2007)Structural basis for AMP binding to mammalian AMP-activated protein kinase.Nature 449, 496–500.
[20] Fryer, L.G., Parbu-Patel, A. and Carling, D. (2002) The anti-diabetic drugsrosiglitazone and metformin stimulate AMP-activated protein kinase throughdistinct signaling pathways. J. Biol. Chem. 277, 25226–25232.
[21] Sakamoto, K., Goransson, O., Hardie, D.G. and Alessi, D.R. (2004) Activity ofLKB1 and AMPK-related kinases in skeletal muscle: effects of contraction,phenformin, and AICAR. Am. J. Physiol. Endocrinol. Metabol. 287, E310–E317.
[22] Lizcano, J.M., Goransson, O., Toth, R., Deak, M., Morrice, N.A., Boudeau, J.,Hawley, S.A., Udd, L., Makela, T.P., Hardie, D.G. and Alessi, D.R. (2004) LKB1 is amaster kinase that activates 13 kinases of the AMPK subfamily, includingMARK/PAR-1. EMBO J. 23, 833–843.
[23] Corton, J.M., Gillespie, J.G., Hawley, S.A. and Hardie, D.G. (1995) 5-Aminoimidazole-4-carboxamide ribonucleoside. A specific method foractivating AMP-activated protein kinase in intact cells? Eur. J. Biochem. 229,558–565.
[24] Sullivan, J.E., Brocklehurst, K.J., Marley, A.E., Carey, F., Carling, D. and Beri, R.K.(1994) Inhibition of lipolysis and lipogenesis in isolated rat adipocytes withAICAR, a cell-permeable activator of AMP-activated protein kinase. FEBS Lett.353, 33–36.
[25] Hawley, S.A., Fullerton, M.D., Ross, F.A., Schertzer, J.D., Chevtzoff, C., Walker,K.J., Peggie, M.W., Zibrova, D., Green, K.A., Mustard, K.J., Kemp, B.E., Sakamoto,K., Steinberg, G.R. and Hardie, D.G. (2012) The ancient drug salicylate directlyactivates AMP-activated protein kinase. Science 336, 918–922.
[26] Sanders, M.J., Ali, Z.S., Hegarty, B.D., Heath, R., Snowden, M.A. and Carling, D.(2007) Defining the mechanism of activation of AMP-activated protein kinaseby the small molecule A-769662, a member of the thienopyridone family. J.Biol. Chem. 282, 32539–32548.
[27] Goransson, O., McBride, A., Hawley, S.A., Ross, F.A., Shpiro, N., Foretz, M.,Viollet, B., Hardie, D.G. and Sakamoto, K. (2007) Mechanism of action of A-769662, a valuable tool for activation of AMP-activated protein kinase. J. Biol.Chem. 282, 32549–32560.
[28] Xiao, B., Sanders, M.J., Carmena, D., Bright, N.J., Haire, L.F., Underwood, E.,Patel, B.R., Heath, R.B., Walker, P.A., Hallen, S., Giordanetto, F., Martin, S.R.,Carling, D. and Gamblin, S.J. (2013) Structural basis of AMPK regulation bysmall molecule activators. Nat. Commun. 4, 3017.
[29] Rubenstein, E.M., McCartney, R.R., Zhang, C., Shokat, K.M., Shirra, M.K., Arndt,K.M. and Schmidt, M.C. (2008) Access denied: Snf1 activation loopphosphorylation is controlled by availability of the phosphorylatedthreonine 210 to the PP1 phosphatase. J. Biol. Chem. 283, 222–230.
[30] Tu, J. and Carlson, M. (1995) REG1 binds to protein phosphatase type 1 andregulates glucose repression in Saccharomyces cerevisiae. EMBO J. 14, 5939–5946.
[31] Frederick, D.L. and Tatchell, K. (1996) The REG2 gene of Saccharomycescerevisiae encodes a type 1 protein phosphatase-binding protein thatfunctions with Reg1p and the Snf1 protein kinase to regulate growth. Mol.Cell. Biol. 16, 2922–2931.
[32] Treitel, M.A., Kuchin, S. and Carlson, M. (1998) Snf1 protein kinase regulatesphosphorylation of the Mig1 repressor in Saccharomyces cerevisiae. Mol. Cell.Biol. 18, 6273–6280.
[33] Ostling, J. and Ronne, H. (1998) Negative control of the Mig1p repressor bySnf1p-dependent phosphorylation in the absence of glucose. Eur. J. Biochem.252, 162–168.
[34] Nehlin, J.O. and Ronne, H. (1990) Yeast MIG1 repressor is related to themammalian early growth response and Wilms’ tumour finger proteins. EMBOJ. 9, 2891–2898.
[35] Westholm, J.O., Nordberg, N., Muren, E., Ameur, A., Komorowski, J. and Ronne,H. (2008) Combinatorial control of gene expression by the three yeastrepressors Mig1, Mig2 and Mig3. BMC Genomics 9, 601.
[36] DeVit, M.J. and Johnston, M. (1999) The nuclear exportin Msn5 is required fornuclear export of the Mig1 glucose repressor of Saccharomyces cerevisiae. Curr.Biol. 9, 1231–1241.
[37] De Vit, M.J., Waddle, J.A. and Johnston, M. (1997) Regulated nucleartranslocation of the Mig1 glucose repressor. Mol. Biol. Cell 8, 1603–1618.
[38] Papamichos-Chronakis, M., Gligoris, T. and Tzamarias, D. (2004) The Snf1kinase controls glucose repression in yeast by modulating interactionsbetween the Mig1 repressor and the Cyc8-Tup1 co-repressor. EMBO Rep. 5,368–372.
[39] Ye, T., Elbing, K. and Hohmann, S. (2008) The pathway by which the yeastprotein kinase Snf1p controls acquisition of sodium tolerance is different fromthat mediating glucose regulation. Microbiology 154, 2814–2826.
[40] Garcia-Salcedo, R., Lubitz, T., Beltran, G., Elbing, K., Tian, Y., Frey, S.,Wolkenhauer, O., Krantz, M., Klipp, E. and Hohmann, S. (2014) Glucose de-repression by yeast AMP-activated protein kinase SNF1 is controlled via atleast two independent steps. FEBS J. 281, 1901–1917.
[41] Botstein, D. and Fink, G.R. (2011) Yeast: an experimental organism for 21stCentury biology. Genetics 189, 695–704.
[42] Daniel, T. and Carling, D. (2002) Expression and regulation of the AMP-activated protein kinase-SNF1 (sucrose non-fermenting 1) kinase complexesin yeast and mammalian cells: studies using chimaeric catalytic subunits.Biochem. J. 365, 629–638.
[43] Winzeler, E.A., Shoemaker, D.D., Astromoff, A., Liang, H., Anderson, K., Andre,B., Bangham, R., Benito, R., Boeke, J.D., Bussey, H., Chu, A.M., Connelly, C., Davis,K., Dietrich, F., Dow, S.W., El Bakkoury, M., Foury, F., Friend, S.H., Gentalen, E.,Giaever, G., Hegemann, J.H., Jones, T., Laub, M., Liao, H., Liebundguth, N.,Lockhart, D.J., Lucau-Danila, A., Lussier, M., M’Rabet, N., Menard, P., Mittmann,M., Pai, C., Rebischung, C., Revuelta, J.L., Riles, L., Roberts, C.J., Ross-MacDonald,P., Scherens, B., Snyder, M., Sookhai-Mahadeo, S., Storms, R.K., Veronneau, S.,Voet, M., Volckaert, G., Ward, T.R., Wysocki, R., Yen, G.S., Yu, K., Zimmermann,K., Philippsen, P., Johnston, M. and Davis, R.W. (1999) Functionalcharacterization of the S. cerevisiae genome by gene deletion and parallelanalysis. Science 285, 901–906.
[44] Kotval, J., Zaret, K.S., Consaul, S. and Sherman, F. (1983) Revertants of atranscription termination mutant of yeast contain diverse genetic alterations.Genetics 103, 367–388.
[45] Woods, A., Cheung, P.C., Smith, F.C., Davison, M.D., Scott, J., Beri, R.K. andCarling, D. (1996) Characterization of AMP-activated protein kinase beta andgamma subunits. Assembly of the heterotrimeric complex in vitro. J. Biol.Chem. 271, 10282–10290.
[46] Goldstein, A. and Lampen, J.O. (1975) Beta-D-fructofuranoside fructohydrolasefrom yeast. Methods Enzymol. 42, 504–511.
[47] Hohmann, S. and Zimmermann, F.K. (1986) Cloning and expression on amulticopy vector of five invertase genes of Saccharomyces cerevisiae. Curr.Genet. 11, 217–225.
[48] Wilson, W.A., Hawley, S.A. and Hardie, D.G. (1996) Glucose repression/derepression in budding yeast: SNF1 protein kinase is activated byphosphorylation under derepressing conditions, and this correlates with ahigh AMP:ATP ratio. Curr. Biol. 6, 1426–1434.
[49] McCartney, R.R. and Schmidt, M.C. (2001) Regulation of Snf1 kinase. Activationrequires phosphorylation of threonine 210 by an upstream kinase as well as adistinct step mediated by the Snf4 subunit. J. Biol. Chem. 276, 36460–36466.
[50] Hong, S.P., Leiper, F.C., Woods, A., Carling, D. and Carlson, M. (2003) Activationof yeast Snf1 and mammalian AMP-activated protein kinase by upstreamkinases. Proc. Natl. Acad. Sci. U.S.A. 100, 8839–8843.
[51] Bu, Y. and Schmidt, M.C. (1998) Identification of cis-acting elements in theSUC2 promoter of Saccharomyces cerevisiae required for activation oftranscription. Nucleic Acids Res. 26, 1002–1009.
[52] Ozcan, S., Vallier, L.G., Flick, J.S., Carlson, M. and Johnston, M. (1997)Expression of the SUC2 gene of Saccharomyces cerevisiae is induced by lowlevels of glucose. Yeast 13, 127–137.
[53] Lutfiyya, L.L. and Johnston, M. (1996) Two zinc-finger-containing repressorsare responsible for glucose repression of SUC2 expression. Mol. Cell. Biol. 16,4790–4797.
[54] Zimmermann, F.K. and Scheel, I. (1977) Mutants of Saccharomyces cerevisiaeresistant to carbon catabolite repression. Mol. Gen. Genet. 154, 75–82.
[55] Sa-Correia, I., dos Santos, S.C., Teixeira, M.C., Cabrito, T.R. and Mira, N.P. (2009)Drug:H+ antiporters in chemical stress response in yeast. Trends Microbiol.17, 22–31.
[56] Gancedo, J.M. (2008) The early steps of glucose signalling in yeast. FEMSMicrobiol. Rev. 32, 673–704.
[57] Hardie, D.G., Carling, D. and Carlson, M. (1998) The AMP-activated/SNF1protein kinase subfamily: metabolic sensors of the eukaryotic cell? Annu. Rev.Biochem. 67, 821–855.
[58] Schmidt, M.C. and McCartney, R.R. (2000) Beta-subunits of Snf1 kinase arerequired for kinase function and substrate definition. EMBO J. 19, 4936–4943.
[59] Vincent, O., Townley, R., Kuchin, S. and Carlson, M. (2001) Subcellularlocalization of the Snf1 kinase is regulated by specific beta subunits and anovel glucose signaling mechanism. Genes Dev. 15, 1104–1114.
[60] Zhang, Y., McCartney, R.R., Chandrashekarappa, D.G., Mangat, S. and Schmidt,M.C. (2011) Reg1 protein regulates phosphorylation of all three Snf1 isoformsbut preferentially associates with the Gal83 isoform. Eukaryot. Cell 10, 1628–1636.
[61] Thomas, B.J. and Rothstein, R.J. (1989) Elevated recombination rates intranscriptionally active DNA. Cell 56, 619–630.