the pattern and control of isoprenoid quinone and tocopherol
TRANSCRIPT

Biochem. J. (1968) 108, 475 475Printed in Great Britain
The Pattern and Control of Isoprenoid Quinone and TocopherolMetabolism in the Germinating Grain of Wheat (Triticum vulgare)
BY G. S. HALL* AND D. L. LAIDMANDepartment of Biochemi8try and Soil Science, Univerruity College of North Wales, Bangor
(Received 21 November 1967)
1. The syntheses of ubiquinone-9 and plastoquinone-9 were used as parametersrespectively of mitochondrial and proplastid development in the germinatingwheat grain. 2. The changes in the amounts ofthe tocopherols were also studied andthe possible biological significance of these changes is discussed. During germina-tion, the dimethyl tocopherols of the resting grain are probably not utilized for thesynthesis of o-tocopherol. 3. It was demonstrated that ubiquinone synthesis, andhence probably mitochondrial development, in the aleurone cells during germina-tion, is independent ofcontrol by gibberellic acid from the embryo. 4. The influenceof light on the syntheses of the isoprenoid quinones in the etiolated wheat shootwas investigated. In particular, illumination did not stimulate the synthesis ofeither x-tocopherol or cc-tocopherolquinone.
Despite the fact that the seeds of different plantsdiffer anatomically and chemically, their basic pro-cesses of germination appear to be similar. Theinitial stages of germination transform a quiescentseed into an actively metabolizing organism withina matter of hours (Toole, Hendricks, Borthwick &Toole, 1956). Though the major biochemical eventsconcerned with the subsequent breakdown ofstorage material in germinating seeds have beenwell documented (Wareing, 1963; Mayer &Poljakoff-Mayber, 1963), it is only in recent yearsthat interest has turned to the biochemical mechan-isms that control the development of metabolicactivity and the initiation of cell growth anddivision.Though the appearance ofmany enzymes during
germination represents synthesis (Ingle & Hageman,1965), others can be accounted for by the activationofpre-existing enzymes (Rowsell & Goad, 1964). Inthe cereals it has been demonstrated that the forma-tion of some hydrolytic enzymes in the endospermis controlled by the hormone gibberellic acid.Gibberellic acid is secreted by the embryo early ingermination (Yomo, 1960; Paleg, 1960, 1965) andresponse to the hormone is localized entirely in thealeurone cells (Briggs, 1964; Varner, 1964; Paleg,1964). The hormone also stimulates the incorpora-tion of labelled precursors into RNA (Ram Chandra& Varner, 1965) and of labelled amino acids intoprotein (Varner, 1964). These results suggest thatgibberellic acid may function as a gene de-repressor
* Present address: School of Biological Sciences, Uni-versity of East Anglia, Norwich, NOR 77H.
(Varner & Ram Chandra, 1964). Evidence for thehormonal control of the production of otherenzymes during germination is less conclusive, andthe formation of some endosperm enzymes appearsto be independent of hormone control from theembryo (Marcus & Feeley, 1964; Tanner & Beevers,1965). Paleg & Hyde (1964) have described exten-sive gibberellic acid-dependent changes in the sub-cellular morphology of aleurone cells in germinatingbarley.In the studies described in this paper, ubiquinone
and plastoquinone were used as parameters ofmitochondrial and proplastid development respec-tively. Crane (1965) has pointed out that there hasbeen no extensive study of the changes in amount ofubiquinone during growth and differentiation ofcells or tissues. Seed germination provides such asystem in the plant kingdom. During germinationof the cereal grain, the cells of the embryonic axisexhibit both growth and division, followed bydifferentiation. In contrast, the scutellum and thealeurone layer consist of actively metabolizing cellsthat enlarge but do not undergo cell division orfurther differentiation.
UTbiquinone synthesis in isolated endospermhalves was studied in an attempt to illuminatemechanisms involved in the initiation and regula-tion of mitochondrial development in aleuronetissue of the germinating wheat grain. Amounts ofthe structurally related tocopherols, present inconsiderable quantities in the wheat grain andseedling, were also investigated. [The IIUPAC-IIUBnomenclature of quinones with isoprenoid side

G. S. HALL AND D. L. LAIDMANchains [Biochem. J. (1967) 102, 15] has been usedthroughout this paper. In addition the followingtrivial names have been used: trimethyl toco-pherols, oc-tocopherol + oc-tocotrienol; dimethyltocopherols, any mixture of two or more from,B-tocopherol, f-tocotrienol, y-tocopherol, y-tocotrienol.]A brief summary of this work has already
appeared (Hall & Laidman, 1966).
METHODS
Sterilization and germination of the wheat grains. Wheatgrains (var. Atle; 1963 harvest) were sterilized with NaOClsolution under reduced pressure (B. B. Stowe, personalcommunication). Each 5g. sample of grain (150 grains) wasplaced, along with about 20ml. of dilute NaOCl solution(0-05% available chlorine), in a 100ml. Buchner flask fittedwith a Quickfit stopper. The flask contents were placedunder reduced pressure (water pump) for 15min., thevacuum being released at 3min. intervals and immediatelyreapplied. The grain was then washed free of hypochloritewith ten 30ml. volumes of sterile water.The sterilized grains were allowed to germinate in moist
sterile vermiculite at 240 in the dark. The samples wereharvested at periods up to 8 days and dissected into theircomponent parts for analysis.
Preparation and incubation of endosperm halves. Initially,some difficulty was experienced in attempting to maintainincubated endosperm halves in a sterile state. Because ofthe important results that these early experiments yielded,however, they are reported in detail.
In the first experiments, endosperm halves were dissectedfrom 5g. samples ofgrain that had been surface-sterilized bysimply soaking for 10min. in dilute NaOCl solution. Thegrains were cut in half transversely and the embryo-containing halves were discarded. This ensures the completeremoval of scutellar and embryonic tissue, which mightotherwise act as a hormone source (Paleg, 1961). Theendosperm halves dissected from different 5g. samples ofgrain were the same weight within the limits of our overallexperimental error. This obviated the need to weigh thepreparations after dissection. The halves were incubated onmoist cotton wool in a Petri dish for periods up to 5 days at240. Similar samples were incubated at 40.
In a second series ofexperiments endosperm halves, whichhad been cut from surface-sterilized grains, were soaked inan antibiotic mixture (streptomycin, 100pg./ml.; penicillin,80,ug.fml.) for 2hr. before being incubated as above oncotton wool moistened with the antibiotic mixture. Otherendosperm halves were soaked for 15hr. in 1lM-gibberellicacid solution after the 2hr. antibiotic treatment, and werethen incubated on cotton wool moistened with a solution ofboth the gibberellic acid and the antibiotics. Because theseendosperm halves also became infected with moulds andbacteria, the latter series of experiments were repeated withstrict aseptic conditions for the dissection and incubation.The incubations were carried out in Petri dishes covered byinverted filter funnels, the stems ofwhich were plugged withcotton wool. Before use, the complete incubation units weresterilized by autoclaving, and, after cooling, the cottonwool in the Petri dishes was soaked either in the antibioticsolution or in the solution of gibberellic acid and antibiotic.
In the final series of experiments 5g. samples of grainwere sterilized by vacuum infiltration with dilute NaOClsolution (see above). This procedure ensures that thesterilizing solution penetrates the intercellular spaces of theseed pericarp. Subsequent dissections and incubations wereagain carried out under aseptic conditions. In this series ofexperiments the effects of various inhibitors were alsostudied. Endosperm halves from 5g. of grain were soakedfor 24hr. under aseptic conditions in a solution of theappropriate inhibitor before they were incubated on cottonwool soaked in the same solution. In other experiments thehalves were immersed in the inhibitor solution for 24hr. at40 before incubation at 240. In addition other endospermhalves were dissected, under sterile conditions, into bran(aleurone layer and pericarp) and starchy-endosperm frac-tions, after soaking for 12 hr. in sterile water. The bran wasthen incubated with the inhibitor in the same way as theendosperm halves. All antibiotic solutions were sterilizedby ultrafiltration and samples were incubated for 3 daysbefore analysis.
Addition of exogenous 9-tocotrienol to the endosperm.Grain (4g.; 120 grains) was allowed to germinate in the darkfor 6-5 days. By this time the endosperm was virtually afluid-filled sac. In dim light, with a micro-syringe, 20-OJul.of a ,B-tototrienol emulsion (200,tg./ml.) was injected intothe endosperm of each seedling. The emulsion was preparedimmediately before use by dissolving 2mg. of 3-tocotrienolin 0-2ml. of ethanol and syringing this rapidly into 9-8ml.of water. The suspension was completely dispersed bytreating it ultrasonically for Imin. The injected seedlingswere placed in moist vermiculite and incubated for a further2 days in the dark at 240. On harvesting, the seedlings weredissected into endosperm, root, shoot and scutellum. Inthis experiment the basal portions of the root and shootwere left attached to the scutellum. The lipid extractsfrom the dissected parts were fractionated and assayed fortheir contents of plastoquinone, ubiquinone and toco-pherols.A second experiment was designed to indicate whether or
not exogenous ,B-tocotrienol was degraded by endospermfluid. Duplicate samples of grain (2-5g.) were germinatedfor 6 days and the endosperms carefully removed from theshoot and root. The endosperm fluid could then be collectedby gently crushing the endosperms with a mortar and pestleand pressing the resultant slurry through several layers ofcheesecloth. The fluid from each sample was made up to14ml. with water and placed in a 50ml. conical flask, pluggedwith cotton wool. One sample was boiled and acted ascontrol. A portion (Iml.) of /-tocotrienol emulsion(200,ug.fml.) was added to each flask and to a third flaskcontaining 14ml. of water (water control). All three flaskswere incubated at 240 in a metabolic shaker. At 0, 20 and40hr., 3ml. samples were withdrawn from each flask andplaced into 15 ml. volumes of hot propan-2-ol. The mixturewas boiled under reflux for 20min. and cooled. After addi-tion of water, the lipids were extracted with three 10ml.lots of light petroleum and evaporated to dryness under N2.The extracts were dissolved in ethanol and their tocopherolcontents determined directly by the modified Dilley &Crane (1963) method (Hall & Laidman, 1968).
Effect of light on tocopherol synthesis in the wheat shoot.Samples (4g.) of wheat grain were allowed to germinatein the dark for 6 days. Growth was then allowed tocontinue for periods of 24hr. or 72hr. in the light of two
1968476
TOCOPHEROL METABOLISM OF GERMINATING WHEAT40w bulbs at a distance of 50cm. The shoots were cut offjust above the scutellum and were analysed for their contentof ubiquinone, plastoquinone, phylloquinone, oc-tocopheroland f-carotene. Their chlorophyll contents were measuredby the method of Arnon (1949) on a portion of the lipidextract. Control samples of 6-day seedlings were left in thedark for 24 and 72hr. and analysed similarly.
Determination of ubiquinone, plastoquinone and toco-pherol. The ubiquinone, plastoquinone and tocopherolcontents of the dissected parts of the harvested seedlingswere determined by our routine method (Hall & Laidman,1968). The tocopherol values obtained by gas-liquidchromatography were checked (on duplicate samples) bythe modified Dilley & Crane (1963) spectrophotometricmethod. The ubiquinone contents of the endosperm halveswere determined by the same method, but, on this occasion,the tocopherols were determined as ac-tocopherol+ a-tocotrienol and ,B-tocopherol+,B-tocotrienol, by themodified Dilley & Crane (1963) procedure.
RESULTS
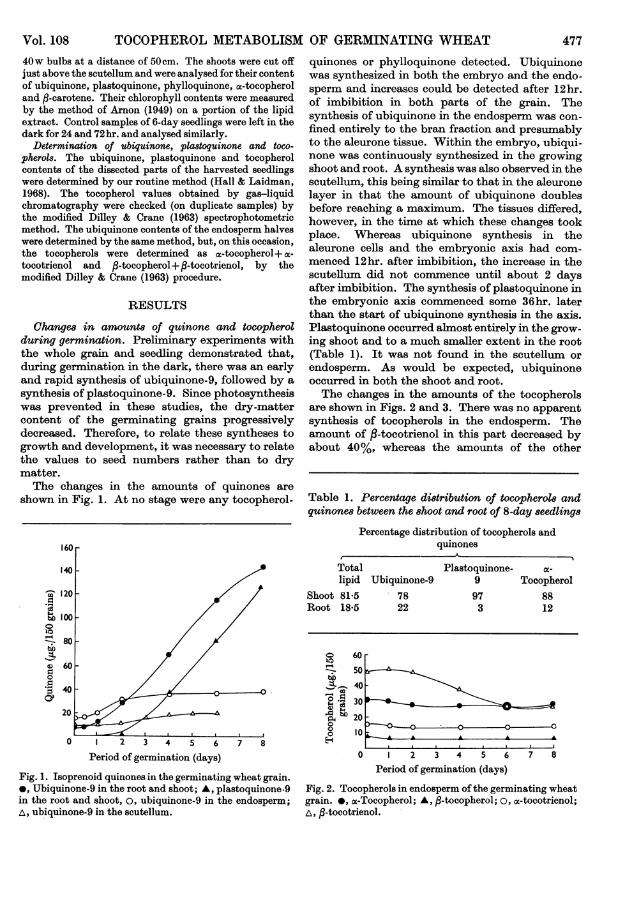
Changes in amounts of quinone and tocopherolduring germination. Preliminary experiments withthe whole grain and seedling demonstrated that,during germination in the dark, there was an earlyand rapid synthesis of ubiquinone-9, followed by asynthesis of plastoquinone-9. Since photosynthesiswas prevented in these studies, the dry-mattercontent of the germinating grains progressivelydecreased. Therefore, to relate these syntheses togrowth and development, it was necessary to relatethe values to seed numbers rather than to drymatter.The changes in the amounts of quinones are
shown in Fig. 1. At no stage were any tocopherol-
quinones or phylloquinone detected. Ubiquinonewas synthesized in both the embryo and the endo-sperm and increases could be detected after 12hr.of imbibition in both parts of the grain. Thesynthesis of ubiquinone in the endosperm was con-fined entirely to the bran fraction and presumablyto the aleurone tissue. Within the embryo, ubiqui-none was continuously synthesized in the growingshoot and root. A synthesis was also observed in thescutellum, this being similar to that in the aleuronelayer in that the amount of ubiquinone doublesbefore reaching a maximum. The tissues differed,however, in the time at which these changes tookplace. Whereas ubiquinone synthesis in thealeurone cells and the embryonic axis had com-menced 12hr. after imbibition, the increase in thescutellum did not commence until about 2 daysafter imbibition. The synthesis of plastoquinone inthe embryonic axis commenced some 36hr. laterthan the start of ubiquinone synthesis in the axis.Plastoquinone occurred ahmost entirely in the grow-ing shoot and to a much smaller extent in the root(Table 1). It was not found in the scutellum orendosperm. As would be expected, ubiquinoneoccurred in both the shoot and root.The changes in the amounts of the tocopherols
are shown in Figs. 2 and 3. There was no apparentsynthesis of tocopherols in the endosperm. Theamount of ,B-tocotrienol in this part decreased byabout 40%, whereas the amounts of the other
Table 1. Percentage distribution of tocopherols andquinones between the shoot and root of 8-day seedlings
Percentage distribution of tocopherols andquinones
120
L100/ /
80
cD 6010
n0
0
ps40O- 0
20
0 2 3 4 5 6 7 8
Period of germination (days)
Fig. 1. Isoprenoid quinones in the germinating wheat grain.*, Ubiquinone-9 in the root and shoot; A, plastoquinone-9in the root and shoot, 0, ubiquinone-9 in the endosperm;a, ubiquinone-9 in the scutellum.
Totallipid Ubiquinone-9
Shoot 81-5 78Root 18-5 22
O 6010
.50t Q 40
30
4 20000 10EH
Plastoquinone-9973
ca-Tocopherol
8812
I p -if
0 1 2 3 4 5 6 7 8
Period of germination (days)
Fig. 2. Tocopherols in endosperm ofthe germinating wheatgrain. *, a-Tocopherol; A, fl-tocopherol; o, ac-tocotrienol;I,I -tocotrienol.
Vol. 108 477
G. S. HALL AND D. L. LAIDMAN
160
140-° 120
bl oo - /80
0
t 60
A 4020E- 20 -_
0 1 2 3 4 5 6 7 8
Period of germination (days)
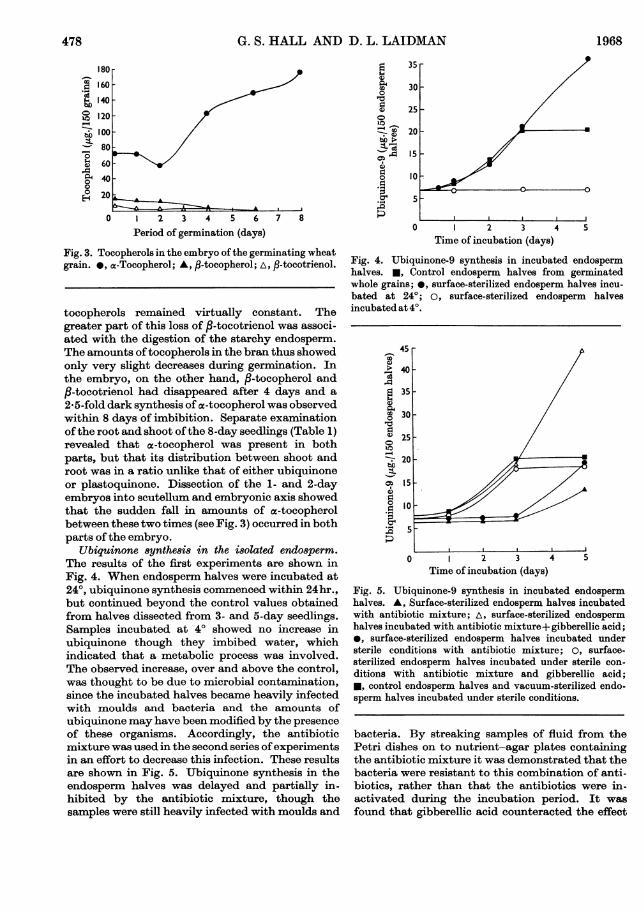
Fig. 3. Tocopherols in the embryo of the germinating wheatgrain. *, a-Tocopherol; A, ,B-tocopherol; A, /3-tocotrienol.
tocopherols remained virtually constant. Thegreater part of this loss of ,B-tocotrienol was associ-ated with the digestion of the starchy endosperm.The amounts oftocopherols in the bran thus showedonly very slight decreases during germination. Inthe embryo, on the other hand, /3-tocopherol and,-tocotrienol had disappeared after 4 days and a
2-5-fold dark synthesis of a-tocopherol was observedwithin 8 days of imbibition. Separate examinationofthe root and shoot ofthe 8-day seedlings (Table 1)revealed that ex-tocopherol was present in bothparts, but that its distribution between shoot androot was in a ratio unlike that of either ubiquinoneor plastoquinone. Dissection of the 1- and 2-dayembryos into scutellum and embryonic axis showedthat the sudden fall in amounts of ac-tocopherolbetween these two times (see Fig. 3) occurred in bothparts ofthe embryo.
Ubiqurinone synthe8i8 in the i8olated endo8perm.The results of the first experiments are shown inFig. 4. When endosperm halves were incubated at240, ubiquinone synthesis commenced within 24hr.,but continued beyond the control values obtainedfrom halves dissected from 3- and 5-day seedlings.Samples incubated at 40 showed no increase inubiquinone though they imbibed water, whichindicated that a metabolic process was involved.The observed increase, over and above the control,was thought to be due to microbial contamination,since the incubated halves became heavily infectedwith moulds and bacteria and the amounts ofubiquinone may have been modified by the presenceof these organisms. Accordingly, the antibioticmixture was used in the second series ofexperimentsin an effort to decrease this infection. These resultsare shown in Fig. 5. Ubiquinone synthesis in theendosperm halves was delayed and partially in-hibited by the antibiotic mixture, though thesamples were still heavily infected with moulds and
4)
0
4)
0
0,
4) 1S.Cbo:._-0
._.,4
35
30
25
20
15
10
0 2 3 4 5Time of incubation (days)
Fig. 4. Ubiquinone-9 synthesis in incubated endospermhalves. *, Control endosperm halves from germinatedwhole grains; 0, surface-sterilized endosperm halves incu-bated at 24°; o, surface-sterilized endosperm halvesincubated at 4°.
454-m
"4)
_
4-
4)
0
0
o
:.
m
2 3 ATime of incubation (days)
Fig. 5. Ubiquinone-9 synthesis in incubated endospermhalves. A, Surface-sterilized endosperm halves incubatedwith antibiotic mixture; A, surface-sterilized endospermhalves incubated with antibiotic mixture+ gibberellic acid;*, surface-sterilized endosperm halves incubated understerile conditions with antibiotic mixture; 0, surface-sterilized endosperm halves incubated under sterile con-
ditions with antibiotic mixture and gibberellic acid;*, control endosperm halves and vacuum-sterilized endo-sperm halves incubated under sterile conditions.
bacteria. By streaking samples of fluid from thePetri dishes on to nutrient-agar plates containingthe antibiotic mixture it was demonstrated that thebacteria were resistant to this combination of anti-biotics, rather than that the antibiotics were in-activated during the incubation period. It was
found that gibberellic acid counteracted the effect
478 1968

TOCOPHEROL METABOLISM OF GERMINATING WHEATof the antibiotics since a large increase in ubiquinonewas detected when both solutions were used to-gether. When the experiment was repeated withaseptic dissection techniques the increase of ubiqui-none in the endosperm halves was still significantlydelayed in the presence of antibiotics, but in thepresence of antibiotics and gibberellic acid apattern more typical of the changes in the intactseed was observed (Fig. 5). However, the sampleswere still heavily contaminated, suggesting that themicro-organisms were not entirely responsible forthe high level of synthesis of ubiquinone in the firstexperiment.
These results suggested that the infections mustbe seed-borne and that the surface-sterilizationprocedure was unsatisfactory. The vacuum-infiltration technique was then introduced. Dilutesodium hypochlorite solution was found to have noeffect on the germination of the intact grains, ascompared with untreated controls, with respect tooxygen uptake, amounts of ubiquinone and visiblegrowth. By this procedure, endosperm halves couldbe incubated for periods ofup to 8 days in the sterilePetri dishes without any sign of microbial infection.Analyses then demonstrated that the pattern ofubiquinone synthesis in the endosperm halvesdissected from germinated grains, and the increasein isolated endosperm halves incubated undergermination conditions, either with or withoutgibberellic acid, were identical (Fig. 5). By incu-bating dissected whole endosperms as well as theendosperm halves it was found that the pattern ofdevelopment of ubiquinone was the same in bothcases. That is, there was no differential increase ofubiquinone between the proximal and distal halvesof the endosperm, owing to progressive diffusion ofthe hormone from the embryo, as has been observedin oc-amylase formation (Briggs, 1964).When endosperm halves were incubated in the
absence of gibberellic acid their amounts of toco-pherol remained constant. The partial loss duringgermination of fl-tocotrienol from the endosperm ofthe intact grain may therefore be indirectlydependent on the hormonal action of gibberellicacid on the aleurone cell. Direct proof of this wasnot possible because endosperm halves that areincubated in the presence ofgibberellic acid undergoextensive degradation of their starchy endospermas a result of hormonally induced a-amylaseactivity. The soluble products flow out from thehalves and therefore there would, in any case, be aloss of the tocopherols of the starchy endosperm.
Table 2 shows the effect of various metabolicinhibitors on the synthesis of ubiquinone in incu-bated endosperm halves and starch-free bran.Neither actinomycin D nor puromycin, at eitherrelatively high or low concentrations, preventedubiquinone synthesis in either endosperm halves or
Table 2. Ubiquinone content of grain parts afterincubation with inhibitors for 3 days
Values are given as ug./150 grain parts. Experimentaldetails are given in the text.
Actinomycin D10,g./ml. (8PM)
100lg./ml.lOO,uLg./ml. (initialperiod4°)100l,g./ml.+KBrO3 (1mM)
Puromycin hydrochloride50,Mg./ml. (O mmM)
500,jg./ml.Chloramphenicol
lmg./ml.2,4-Dinitrophenol1Omm (pH 5.0)0-05mM0-1mMOlmM+gibberellic acid
(0 05mM)
Halves steeped for 3 daysControlsOday (ungerminated)3 days
Pericarp+ Endospermaleuronelayer halves
18 2021-7
- 1921*5
20 2020
- 14*4
15
11*8
619-6
1616-5
12*9
820
isolated bran layers. Chloramphenicol partiallyinhibited the synthesis. When the halves weresoaked in the inhibitor solution for 24hr. at 40before incubation at 240 the grain still absorbedwater, but ubiquinone synthesis during incubationwas not inhibited. It was hoped that, by using thismethod, the antibiotic would have time to penetratethe aleurone cells before active metabolism com-menced. At 40 RNA and protein synthesis isappreciably decreased (Marcus & Feeley, 1965).The addition of potassium bromate to inhibitpossible breakdown of actinomycin D by proteo-lytic enzymes (Varner & Ram Chandra, 1964) hadno effect on the results.When endosperm halves were steeped in water
for 3 days there was only a partial synthesis ofubiquinone, indicating an oxygen requirement.Similarly 2,4-dinitrophenol, either alone or incombination with gibberellic acid, caused a partialbut smaller inhibition of ubiquinone synthesis inboth endosperm halves and isolated aleurone layers.The importance of the oxidative metabolism as asource of energy for the aleurone cell and forubiquinone synthesis is thus established.The observation that the amounts of ubiquinone
in the control samples of the endosperm halves andisolated aleurone layers were identical (Table 2)confirmed that ubiquinone synthesis outside the
Vol. 108 479

480
embryo is confirgests that the s
hydrate or othe:endosperm. It vall traces of st,layer, and it is cthe aleurone cel]
Addition of e'sperm. Seedlingtinued to grow rcontrols. Thou
Table 3. Analysf-tocotri
Values are givedetails are given ir
ShootExperimentalControl
RootExperimentalControl
ScutellumExperimentalControl
TotalExperimentalControl
Table 4. Incuba
Values are giventhe text.
G. S. HALL AND D. L. LAIDMAN 1968ied to the aleurone layer, and sug- injected into 120 seedlings, after a further 2 days of3ynthesis is independent of carbo- growth no trace of this compound could be detectedr energy reserves from the starchy in the root, shoot or scutellum, though a smallwas, however, impossible to remove amount (36utg.) was found in the endosperm. This,arch from the dissected aleurone however, could be accounted for by the endogenousonceivable that this was utilized by fl-tocotrienol present in the bran. The amounts ofIs. a-tocopherol, plastoquinone and ubiquinone in thexogenou8s -tocotrienol to the endo- injected seedlings were not different from those ofrs injected with ,-tocotrienol con- the controls (Table 3).iormally, compared with untreated In the experiments in which ,B-tocotrienol was.gh 400,ug. of ,B-tocotrienol was incubated with samples of endosperm fluid it was
found that there was no rapid breakdown ofP-tocotrienol by enzymic or other agents (Table 4).All the samples, including the controls, showed onlyis of wheat 8eedlings after injection of a relatively slow breakdown over the 40hr. incuba-
wenol into the endosperm sac tion period, though there was an initial sharp de-
n as ,ug./120 seedlings. Experimental crease in the flasks that contained endosperm fluid.a the text. In view of the natural instability of the tocopherol
emulsion and the similarity of the values obtainedPlasto- fromboth the control and experimental incubations,
there was no appreciable destruction of,-tocotrienol88 59 97 in vitro due to enzymic activity of the endosperm83 64 94 fluid. No ,B-tocotrienolquinone could be detected in
any of the incubations or in the seedlings that had2 17 13 been injected with ,-tocotrienol.3 24 16 Effect of light on the synthesi8 of isoprenoid
quinones in the shoot. Though the syntheses of5 24 23 plastoquinone, phylloquinone, ,8-carotene and3 25 20 chlorophyll were stimulated after the illumination
of etiolated shoots, the syntheses of ubiquinone and95 100 133 ac-tocopherol were not (Table 5). In contrast with89 113 130 the results of Threlfall & Griffiths (1967) in barley
and maize only insignificant amounts of oc-toco-pherolquinone could be detected in the wheatshoots that had been illuminated for 24 or 72hr.,
otion of P-tocotrienol with endosperm and we were unable to detect phylloquinone in thefluid etiolated shoot. The light-independent nature of
in ,ig. Experimental details are given in o-tocopherol synthesis in the wheat seedling is inagreement with that demonstrated in the pea by
Time Gaunt & Stowe (1967) and in maize by Threlfall &_____________ l Griffiths (1967).
Olhr. 20hr. 40hr.Endosperm fluid+ ,-tocotrienolBoiled endosperm fluid+
fi-tocotrienolWater control+ fi-tocotrienol
150 134 109146 122 117
202 135 132
DISCUSSION
The increases in amounts of ubiquinone duringthe early stages of germination support the con-
Table 5. Change8 in isoprenoid compounds in etiolated 6-day wheat 8hoots after additional growth periodsin the light and in the dark
Values are given as ug./120 shoots. Experimental details are given in the text.Total
Additional growth period Phylloquinone Ubiquinone-9 Plastoquinone-9 oc-Tocopherol fl-Carotene chlorophyllDark (24hr.) - 57 81 60 16Light (24hr.) Trace 53 176 67 104 3440Dark (72hr.) - 68 87 96 20Light (72hr.) 25 74 293 92 336 5210

TOCOPHEROL METABOLISM OF GERMINATING WHEATclusions of Lund, Vatter & Hanson (1958) that a
significant growth of mitochondria occurs duringcell growth. Available histological evidence in-dicates that mitochondria in scutellar tissues do notincrease in number, but that they do increase insize and degree of development of their cristae afterabout 3 days of germination (Hanson, Vatter,Fisher & Bils, 1959; Nieuwdorp & Buys, 1964).The precise timing of ubiquinone synthesis andmitochondrial development in the various tissues ispresumably synchronized with demands for energy.
Treharne, Mercer & Goodwin (1966) have shownthat, during the 'greening-up' of etiolated shoots,isoprenoid synthesis in the chloroplast is regulatedby a combination of enzyme segregation andspecific membrane permeability. We wish to takethe discussion one stage further. The ratio of therate of synthesis in the light to the rate of synthesisin the dark in the shoots of 6-day dark-grown seed-lings is different for each isoprenoid compound. Inthe wheat seedlings used in our 'greening-up'experiments the ratios range from unity, for a-tocopherol, to infinity, for phylloquinone. In otherwords, the syntheses of different isoprenoids in thedeveloping chloroplast are stimulated to differentdegrees by light. It is clear therefore that some otherfactor, besides that of simple compartition, isinvolved in the control of isoprenoid synthesis inthis organelle.Though the a-tocopherol content of the wheat
embryo axis had increased by 103,ug./150 embryosafter 8 days of germination in the dark, only 47 ,ug.of other tocopherols had disappeared in that timefrom the whole grain. Thus the methylation of thedimethyl tocopherols of the resting grain couldaccount for only a part of the increases in a-tocopherol. The fact that injected ,B-tocotrienol didnot influence the rate of oc-tocopherol synthesistends to uphold the view that the dimethyl toco-pherols are not utilized in this way during germina-tion. Tocopherols have been associated with theflowering process in plants (Michniewicz &Kamienska, 1965) and the presence of the dimethyltocopherols in the resting grain may simply reflecttheir distribution at the end of the grain maturationperiod; they may have no function during germina-tion. Further, the breakdown of exogenousfl-tocotrienol by the starchy endosperm may benothing more than one aspect of the general hydro-lysis and breakdown of this tissue during germina-tion. However, enzymic breakdown of exogenous
tocopherol by tissues that do not appear to becatabolizing their own endogenous tocopherol hasbeen noted (Gaunt & Stowe, 1967; Laidman, 1964).With respect to the tocopherols in the aleurone
cells, however, a more significant proposition maybe advanced. This is that the oc-tocopherol in thesecells is associated with the lipid-rich spherosomes
16
(Paleg & Hyde, 1964), and that this a-tocopheroland some of the other lipids of the spherosome areinvolved in the gibberellic acid-triggered develop-ment of the endoplasmic reticulum during germina-tion. Such a system would be similar to thatdescribed by Lichtenthaler & Sprey (1966) andSprey & Lichtenthaler (1966) as being associatedwith lamellar formation during chloroplast develop-ment.The experiments on endosperm halves have
demonstrated that the synthesis of ubiquinone, andtherefore the complete development of the mito-chondria, in the aleurone tissue are independent ofcontrol by gibberellic acid. This is in contrast withthe gibberellic acid-initiated development of theendoplasmic reticulum in these cells during theearly stages of germination (Paleg & Hyde, 1964).Some other factor must be responsible for initiatingubiquinone synthesis in this tissue during germina-tion. There is clearly yet another controllingmechanism that terminates ubiquinone synthesis inthe aleurone cell after 3 days ofgermination. Similarfactors probably initiate and terminate ubiquinonesynthesis in the scutellum during germination.The inhibitory effect of the penicillin-strepto-
mycin mixture on ubiquinone synthesis in thealeurone tissue is important. It is assumed thatthis effect is due to the streptomycin rather than thepenicillin since the former is more toxic to plants(Brian, 1957). The use of streptomycin to inhibitbacterial growth in isolated endosperm halves can-not therefore be recommended. The finding thatgibberellic acid reverses the inhibitory effect ofstreptomycin but not the inhibitory effect of 2,4-dinitrophenol suggests that the gibberellic acid-streptomycin interaction is specifically associatedwith RNA and protein synthesis. The apparentanomalies between the results of the inhibitorstudies in our aleurone system and those in someother plant systems (e.g. Marcus & Feeley, 1965;Morton & Raison, 1964; Parisi & Ciferri, 1966) canprobably be explained to some degree by thedemonstration in yeast that two distinct protein-synthesizing systems exist (Clark-Walker &Linnane, 1966). One, the cytoplasmic ribosomalsystem, is sensitive to cycloheximide but largelyinsensitive to the inhibitors of bacterial proteinsynthesis; the other, the mitochondrial system, isinhibited by chloramphenicol and other inhibitorsof bacterial protein synthesis. Thus it might beconstrued that ubiquinone synthesis is controlled bya mitochondrial DNA-RNA system. A furtheraspect of antibiotic action that must be consideredis that streptomycin and chloramphenicol alsoinhibit energy-linked processes in plant mito-chondria (Stoner, Hodges & Hanson, 1964). Such amode of action in aleurone mitochondria mightappear to be supported by the inhibitory effect of
Bioch. 1968, 108
Vol. 108 481

482 G. S. HALL AND D. L. LAIDMAN 1968
2,4-dinitrophenol on ubiquinone synthesis, butwould offer no explanation for the reversal of theinhibitory effect of streptomycin by gibberellic acid.Obviously a clearer understanding of the modes ofaction of antibiotics on plant systems is requiredbefore the present results can be fully interpreted.
We are indebted to the Agricultural Research Council forfinancial support, to the Science Research Council for theaward of a research studentship to G.S.H., to Dr A. L.Morrison (Roche Products Ltd., Welwyn Garden City,Herts.) for the gift of authentic tocopherol samples, to DrF. W. Hemming (University of Liverpool) for the gift ofauthentic quinone samples and to Dr J. S. Tzoumertic(Merck, Sharp and Dohme Research Laboratories, Rahway,N.J., U.S.A.) for the gift of actinomycin D.
REFERENCES
Arnon, D. I. (1949). Plant Phy8iol. 24,1.Brian, P. W. (1957). Annu. Rev. Plant Phy8iol. 8,413.Briggs, D. E. (1964). J. Inst. Brew. 70,14.Clark-Walker, G. D. & Linnane, A. W. (1966). Biochem.
biophy8. Re8. Commun. 25, 8.Crane, F. L. (1965). In Biochemi8try of Quinone8, p. 183.
Ed. by Morton, R. A. London and New York: AcademicPress Inc.
Dilley, R. A. & Crane, F. L. (1963). Analyt. Biochem. 5,531.Gaunt, J. K. & Stowe, B. B. (1967). Plant Phy8iol. 42,859.Hall, G. S. & Laidman, D. L. (1966). Biochem. J. 101, 5P.Hall, G. S. & Laidman, D. L. (1968). Biochem. J. 108,465.Hanson, J. B., Vatter, A. E., Fisher, M. E. & Bils, R. F.
(1959). Agron. J. 51, 295.Ingle, J. & Hageman, R. H. (1965). Plant Phy8iol. 40, 672.Laidman, D. L. (1964). Biochem. J. 91, 31 c.Lichtenthaler, H. K. & Sprey, B. (1966). Z. Naturf. 21b, 690.Lund, H. A., Vatter, A. E. & Hanson, J. B. (1958). J.
biophy8. biochem. Cytol. 4,87.
Marcus, A. & Feeley, J. (1964). Biochim. biophy8. Acta, 89,170.
Marcus, A. & Feeley, J. (1965). J. biol. Chem. 240, 1675.Mayer, A. M. & Poljakoff-Mayber, A. (1963). The Germina-
tion ofSeed8, pp. 101-143. Oxford: Pergamon Press Ltd.Michniewicz, M. & Kamienska, A. (1965). Naturwi8en-
8chaften, 52, 623.Morton, R. K. & Raison, J. K. (1964). Biochem. J. 91, 528.Nieuwdorp, P. J. & Buys, M. C. (1964). Acta bot. neerl. 13,
559.Paleg, L. (1960). Plant Physiol. 35, 293.Paleg, L. (1961). Plant Phy8iol. 36, 829.Paleg, L. (1964). In Regulateurs Naturels de la Croi8sance
Vegetale, p. 303. Ed. by Nitsch, J. P. Paris: CentreNationale du Recherche Scientifique.
Paleg, L. (1965). Annu. Rev. Plant Phy8iol. 16,291.Paleg, L. & Hyde, B. (1964). Plant Phy8iol. 89, 673.Parisi, B. & Ciferri, 0. (1966). Biochemi8try, 5,1638.Ram Chandra, G. & Varner, J. E. (1965). Biochim. biophy8.
Acta, 108,583.Rowsell, E. V. & Goad, L. J. (1964). Biochem. J. 90, 12P.Sprey, B. & Lichtenthaler, H. K. (1966). Z. Naturf. 21b, 697.Stoner, C. D., Hodges, T. K. & Hanson, J. B. (1964).
Nature, Lond. 203, 258.Tanner, W. & Beevers, H. (1965). Z. Pflanzenphysiol. 58,72.Threlfall, D. R. & Griffiths, W. T. (1967). In Biochemistry of
Chloropla8t8, vol. 2, p. 255. Ed. by Goodwin, T. W.London andNew York: Academic Press Inc.
Toole, E. H., Hendricks, S. B., Borthwick, H. A. & Toole,V. K. (1956). Annu. Rev. Plant Phy8iol. 7, 299.
Treharne, K. J., Mercer, E. I. & Goodwin, T. W. (1966).Biochem. J. 99, 239.
Varner, J. E. (1964). Plant Phy8iol. 89, 413.Varner, J. E. & Ram Chandra, G. (1964). Proc. nat. Acad.
Sci., Wa8h. 52, 100.Wareing, P. F. (1963). In Vistas in Botany, vol. 3, p. 195.
Ed. by Turrill, W. B. Oxford: Pergamon Press Ltd.Yomo, H. (1960). J. Ferment. A88. Japan, 18, 600; cited in
Chem. Ab8tr. (1961), 55,26145.