the phototroph c way of l - people.ucsc.edusaltikov/migrated/courses_backup/archive... · the...
TRANSCRIPT

CHAPTER 1.3The Phototrophic Way of Life
The Phototrophic Way of Life
JÖRG OVERMANN AND FERRAN GARCIA-PICHEL
IntroductionPhotosynthesis is the utilization of radiantenergy for the synthesis of complex organic mol-ecules. The phototrophic way of life implies thecapture of electromagnetic energy (see LightAbsorption and Light Energy Transfer inProkaryotes in this Chapter), its conversion intochemical energy (see Conversion of Light intoChemical Energy in this Chapter), and its use forcellular maintenance and growth (see Efficiencyof Growth and Maintenance Energy Require-ments in this Chapter). Photosynthesis mayencompass the reduction of carbon dioxide intoorganic molecules, a mode of growth defined asphotoautotrophy. The solar electromagneticenergy reaching the Earth’s surface (160 W·m–2;see Light energy and the spectral distribution ofradiation) surpasses the energy contributed byall other sources by four to five orders of magni-tude (electric discharge, radioactivity, volcanism,or meteoritic impacts;
~0.0062 W·m–2 on primor-dial Earth; Mauzerall, 1992; present day geother-mal energy
~0.0292 W·m
-2; K. Nealson, personalcommunication).
At present the flux of electromagnetic energysupports a total primary production of 172.5
¥109 tons dry weight·year
-1 (168 g C·m
-2·year
-1;Whittaker and Likens, 1975). If this globalprimary production is converted to energy units(39.9 kJ·g C
-1, assuming that all photosyntheticproducts are carbohydrate), 0.21 W·m
-2 and thus0.13% of the available solar energy flux are con-verted into chemical energy. Even at this lowefficiency, the chemical energy stored in organiccarbon still exceeds geothermal energy by atleast one order of magnitude. As a consequence,photosynthesis directly or indirectly drives thebiogeochemical cycles in all extant ecosystems ofthe planet. Even hydrothermal vent communi-ties, which use inorganic electron donors ofgeothermal origin and assimilate CO2 by chem-olithoautotrophy (rather than photoautotro-phy), still depend on the molecular O2 generatedby oxygenic phototrophs outside of these sys-tems (Jannasch, 1989).
Several lines of evidence indicate that in theearly stages of biosphere evolution, prokaryotic
organisms were once responsible for the entireglobal photosynthetic carbon fixation. Today, ter-restrial higher plants account for the vast major-ity of photosynthetic biomass; the chlorophyllbound in light-harvesting complex LHCII ofgreen chloroplasts alone represents 50% of thetotal chlorophyll on Earth (Sidler, 1994). In con-trast, the biomass of marine primary producersis very low (0.2% of the global value). However,the biomass turnover of marine photosyntheticmicroorganisms is some 700 times faster thanthat of terrestrial higher plants. Thus, marinephotosynthetic organisms contribute signifi-cantly to total primary productivity (55·109 tonsdry weight·year
-1, or 44% of the global primaryproduction). Because the biomass of cyanobac-terial picoplankton (see Habitats of Photo-trophic Prokaryotes in this Chapter) can amountto 67% of the oceanic plankton, and their pho-tosynthesis up to 80% in the marine environ-ment (Campbell et al., 1994; Goericke andWelschmeyer, 1993; Liu et al., 1997; Waterburyet al., 1986), prokaryotic primary productionis still significant on a global scale. A singlemonophyletic group of marine unicellularcyanobacterial strains encompassing the generaProchloroccoccus and Synechococcus with a glo-bal biomass in the order of a billion of metrictons (Garcia-Pichel, 1999) may be responsiblefor the fixation of as much as 10–25% of theglobal primary productivity. Additionally,prokaryotic (cyanobacterial) photosynthesis isstill locally very important in other habitats suchas cold (Friedmann, 1976) and hot deserts(Garcia-Pichel and Belnap, 1996) a nd hyper-trophic lakes.
Today, the significance of anoxygenic photo-synthesis for global carbon fixation is limited fortwo reasons. On the one hand, phototrophicsulfur bacteria (the dominant anoxygenicphototrophs in natural ecosystems) form denseaccumulations only in certain lacustrine environ-ments and in intertidal sandflats. The fraction oflakes and intertidal saltmarshes which harboranoxygenic phototrophic bacteria is unknown,but these ecosystems altogether contribute only4% to global primary production (Whittaker andLikens, 1975). In those lakes harboring pho-
Prokaryotes (2006) 2:32–85DOI: 10.1007/0-387-30742-7_3

CHAPTER 1.3 The Phototrophic Way of Life 33
totrophic sulfur bacteria, an average of 28.7% ofthe primary production is anoxygenic (Over-mann, 1997). Consequently, the amount of CO2
fixed by anoxygenic photosynthesis must con-tribute much less than 1% to global primaryproduction. On the other hand, anoxygenic pho-tosynthesis depends on reduced inorganic sulfurcompounds which originate from the anaerobicdegradation of or ganic carbon. Since this carbonwas already fixed by oxygenic photosynthesis,the CO2-fixation of anoxygenic phototrophicbacteria does not lead to a net increase in organiccarbon available to higher trophic levels. TheCO2-assimilation by anoxygenic phototrophicbacteria has therefore been termed “secondaryprimary production” (Pfennig, 1978). Therefore,capture of light energy by anoxygenic photosyn-thesis merely compensates for the degradation oforganic carbon in the anaerobic food chain. Geo-thermal sulfur springs are the only exceptionsince their sulfide is of abiotic origin. However,because sulfur springs are rather scarce, anoxy-genic photosynthetic carbon fixation of theseecosystems also appears to be of minor signifi-cance on a global scale.
The scientific interest in anoxygenic phototro-pic bacteria stems from 1) the simple moleculararchitecture and variety of their photosystems,which makes anoxygenic phototrophic bacteriasuitable models for biochemical and biophysicalstudy of photosynthetic mechanisms, 2) theconsiderable diversity of anoxygenic pho-totrophic bacteria, which has implications forreconstructing the evolution of photosynthesis,and 3) the changes in biogeochemical cycles ofcarbon and sulfur, which are mediated by thedense populations of phototrophic bacteria innatural ecosystems.
All known microorganisms use two functionalprinciples (both mutually exclusive and repre-sent two independent evolutionary develop-ments) for the conversion of light into chemicalenergy. Chlorophyll-based systems are wide-spread among members of the domain Bacteriaand consist of a light-harvesting antenna andreaction centers. In the latter, excitation energyis converted into a redox gradient across themembrane. In contrast, the retinal-based bacte-riorhodopsin system is exclusively found inmembers of a monophyletic group within thedomain Archaea. These prokaryotes lack anantenna system and use light energy for thedirect translocation of protons across the cyto-plasmic membrane. In both systems, photosyn-thetic energy conversion ultimately results in theformation of energy-rich chemical bonds oforganic compounds.
The advent of modern genetic and biochemi-cal methods has led to a considerable gain inknowledge of the molecular biology of pho-
totrophic prokaryotes. At the same time, micro-bial ecologists have found these microorganismsof considerable interest and now frequently usemolecular methods to investigate natural popu-lations. The present chapter is limited to the dis-cussion of phototrophic bacteria and attempts tolink the physiology, ecology, and evolution ofphototrophic bacteria to a molecular basis.Emphasis is laid on those molecular structures orfunctions that have evident adaptive value. Thisintegrating view may provide a more solid foun-dation for understanding the biology of photo-synthetic prokaryotes.
Taxonomy of Phototrophic Prokaryotes
The capacity for chlorophyll-based photosyn-thetic energy conversion is found in five of the36 currently recognized bacterial lineages (Fig. 1;Hugenholtz et al., 1998): the Chloroflexussubgroup, the green sulfur bacteria, the Proteo-bacteria, the Cyanobacteria, and the Heliobacte-riaceae. With the exception of the Cyanobacteria,phototrophic bacteria perform anoxygenic pho-tosynthesis, which is not accompanied by photo-chemical cleavage of water and therefore doesnot lead to the formation of molecular oxygen.Based on their phenotypic characters, anoxy-genic phototrophic bacteria had been dividedpreviously into the five families Rhodospiril-laceae, Chromatiaceae, Ectothiorhodospiraceae,Chlorobiaceae, and Chloroflexaceae (Trüper andPfennig, 1981). However, 16S rRNA oligonucle-otide cataloguing and 16S rRNA sequence com-parisons have reveale d that the Proteobacteriaand the Chloroflexus-subgroup both containnonphototrophic representatives (Woese, 1987;Fig. 1). Therefore the use of light as an energysource for growth is not limited to phylogeneti-cally coherent groups of bacteria. However, non-phototrophic representatives of the green sulfurbacterial and the cyanobacterial lineages havenot been isolated to date.
Within the Chloroflexus-subgroup, three dif-ferent species (Chloroflexus aurantiacus, Chlo-roflexus aggregans and Heliothrix oregonensis)of filamentous multicellular phototrophs havebeen described. All three are thermophilic andgrow photoorganoheterotrophically. In additionfour mesophilic species (Oscillochloris chrysea,Oscillochloris trichoides, Chloronema gigan-teum, Chloronema spiroideum) have been affili-ated with the Chloroflexus-subgroup based ontheir multicellular filaments, gliding motility, andthe presence of chlorosomes containing bacteri-ochlorophylls c or d (Pfennig and Trüper, 1989).The phylogenetic position of these latter bacteria

34 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
has not been investigated so far. With the excep-tion of Heliothrix oregonensis all species men-tioned contain chlorosomes as distinct light-harvesting structures (Fig. 2). Yet to be cultivatedaxenically, non-thermophilic “Chloroflexus-like”organisms are known from intertidal and hyper-saline benthic environments (Pierson et al.,1994) and from cold freshwater sulfidic springs(F. Garcia-Pichel, unpublished observation). Atleast in the case of the hypersaline enrichments,the organisms are closely related to Heliothrix interms of their 16S rRNA sequence (B.K. Pierson,personal communication to FGP). This, togetherwith recent descriptions of Oscillochloris tri-choides (Keppen et al., 1994) from freshwatersediments indicates a larger diversity and morewidespread occurrence of the Chloroflexaceaeand allied organisms than was previouslyrecognized.
Green sulfur bacteria (see The Family Chloro-biaceae Volume 7) represent a coherent and iso-lated group within the domain Bacteria. They arestrict photolithotrophs and contain chlorosomes(Fig. 3A). During the oxidation of sulfide, ele-mental sulfur is deposited extracellularly.Another typical feature of this group is the verylimited physiological flexibility (see Docile Reac-
tion). In the Proteobacteria, the
a- and
b-Proteo-bacteria comprise photosynthetic representatives(often also called the purple nonsulfur bacteria),which do not form separate phylogenetic clustersbut are highly intermixed with various other phe-notypes. Characteristically, members of these twogroups exhibit a high metabolic versatility andare capable of photoorganotrophic, photo-lithoautotrophic and chemoorganotrophicgrowth. Photosynthetic pigments are bacterio-chlorophyll a or b and a variety of carotenoids.Light-harvesting complexes, reaction centers, andthe component s of the electron transport chainare located in intracellular membrane systems ofspecies-specific architecture (Fig. 2; see LightAbsorption and Light Energy Transfer inProkaryotes in this Chapter).
Several members of the
a-Proteobacteria arecapable of bacteriochlorophyll a synthesis butcannot grow by anoxygenic photosynthesis. Thisphysiological group has therefore been desig-nated “aerobic anoxygenic phototrophic bacte-ria” (Shimada, 1995; Yurkov and Beatty, 1998),“aerobic phototrophic bacteria” (Shiba, 1989),or “quasi-photosynthetic bacteria” (Gest, 1993)and comprises a considerable number of species.So far, the marine genera Erythrobacter and
Fig. 1. Phylogenetic tree based on 16S rRNA sequences. All bacterial divisions containing culturable representatives wereincluded in the analyses so that the phototrophic nature of the bacterial strains could be confirmed. Alignments were obtainedwith CLUSTAL W and pairwise distances calculated with the algorithm of Jukes and Cantor using the DNADIST programof PHYLIP 3.57c. The tree was constructed from evolutionary distances employing the least-squares algorithm of Fitch andMargoliash as implemented by the FITCH program of the package. The Archaeon Methanopyrus kandleri DSM 6324 wasused as an outgroup to root the tree. (light green) Bacteria containing chlorosomes as light-harvesting antenna. (red) Bacteriacontaining antenna complexes within the cytoplasmic membrane and quinone/pheophytin-type reaction centers. (mediumgreen) Gram-positive phototrophic bacteria with FeS-type reaction centers. (dark green) Bacteria containing the two typesof reaction centers. Width of colored wedges indicates the phylogenetic divergence.

CHAPTER 1.3 The Phototrophic Way of Life 35
Roseobacter and the six freshwater genera Acidi-philium, Erythromonas, Erythromicrobium,Porphyrobacter, Roseococcus, Sandarcinobacter(Yurkov and Beatty, 1998) have been described.This group also includes some aerobic faculta-tively methylotrophic bacteria of the genusMethylobacterium and a Rhizobium (strainBTAi1; Evans et al., 1990; Shimada, 1995;Urakami and Komagata, 1984). The oxidation oforganic carbon compounds is the principalsource of metabolic energy. Photophosphoryla-tion can be used as a supplementary source ofenergy, with a transient enhancement of aerobicgrowth following a shift from dark to illumina-tion (Harashima et al., 1978; Shiba andHarashima, 1986). Aerobic bacteriochlorophyll-containing bacteria harbor a photosyntheticapparatus very similar to photosystem IIof anoxygenic phototrophic Proteobacteria
(Yurkov and Beatty, 1998). Photochemically active reaction centers and light-harvesting com-plexes are present, as are the components ofcyclic electron transport (e.g., a cytochrome cbound to the reaction center and soluble cyto-chrome c2). In contrast to anoxygenic pho-totrophic bacteria, however, the aerobicphototrophic bacteria cannot grow autotrophi-cally. Intracellular photosynthetic membranesystems as they are typical for anoxygenic pho-totrophic Proteobacteria are absent in most aer-obic photosynthetic bacteria; Rhizobium BTAi1being a possible exception (Fleischman et al.,1995). The presence of highly polar carotenoidsulfates and C30 carotenoid glycosides is a uniqueproperty of this group. All aerobic bacteriochlo-rophyll a-containing species group with the
a-subclass of the Proteobacteria, but aremore closely related to aerobic non-
Fig. 2. Organization of the pho-totrophic apparatus in differentgroups of phototrophic bacteria.OM
= outer membrane, CW
= cellwall, CM
= cytoplasmic membrane,RC
= reaction center, LHC
= light-harvesting complex. Question marksindicate that the organization of thecell envelope and the organization ofthe photosynthetic apparatus inHeliothrix oregonensis is not exactlyknown.

36 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
bacteriochlorophyll-contain ing organisms thanto anoxygenic phototrophs (Stackebrandt et al.,1996).
The
g-subclass comprises two families ofphototrophic species, the Chromatiaceae andEctothiorhodospiraceae (also called purple sul-
fur bacteria). Chromatiaceae accumulate sulfurglobules within the cells and represent a conspic-uous microscopic feature of these bacteria. Withone notable exception (Thiocapsa pfennigii), theintracellular membrane system is of the vesiculartype (Figs. 2 and 3B). In contrast, members of
Fig. 3. Localization and organization of the photosynthetic apparatus in three major groups of phototrophic bacteria.Electron-donating enzyme systems, like flavocytochrome or sulfide quinone reductase, and ATP formation by the membrane-bound ATP synthase are not shown. A. Green sulfur bacteria (Chlorobiaceae). B. Purple nonsulfur bacteria and Chromati-aceae. C. Cyanobacteria. OM
= outer membrane; CW
= cell wall; CM
= cytoplasmic membrane; Cyt
= cytochrome; P840 andP870 reaction center special pair
= primary electron donor; B800, B850, B875
= bacteriochlorophyll molecules bound to light-harvesting complexes II and I; A0
= primary electron acceptor in green sulfur bacteria
= Chl a; A1
= secondary electronacceptor in green sulfur bacteria
= menaquinone; QA, QB
= ubiquinone; FX, FA, FB = FeS-clusters bound to the reaction center;Fd = ferredoxin; FMO = Fenna-Matthews-Olson protein; FNR = ferredoxin NADP+ reductase; PQ = plastoquinone;PC = plastocyanin; PS = photosystem.

CHAPTER 1.3 The Phototrophic Way of Life 37
the Ectothiorhodospiraceae deposit elementalsulfur outside of the cells and contain lamellarintracellular membrane systems. Like their rela-tives of the a- and b-subclass of Proteobacteria,the purple sulfur bacteria contain bacteriochlo-rophylls a and b, and all components of thephotosynthetic apparatus are located in theintracellular membrane.
No photosynthetic species have beendescribed for the d- or e-subclass of theProteobacteria.
Heliobacteriaceae differ from other anoxy-genic phototrophic bacteria by their uniquelight-harvesting and reaction center pigment,
bacteriochlorophyll g, and by their phylogeneticaffiliation (Fig. 1). The first member of this group,Heliobacterium chlorum was described in 1983by Gest and Favinger (Gest and Favinger,1983b). Based on peptidoglycan structure studies(Beer-Romero et al., 1988), their high propor-tion of branched-chain fatty acids (Beck et al.,1990) and 16S rRNA sequencing, the Heliobac-teriaceae belong to the Gram-positive low GClineage. A close relatedness can also be deducedfrom the capability of Heliobacterium modesti-caldum and Heliobacterium gestii to formendospores. However, a detailed phylogeneticanalysis also indicated a close relatedness of
Fig. 3. Continued.

38 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
Heliobact eriaceae to the Cyanobacteria (Ver-maas, 1994). Heliobacteriaceae do not containdistinct intracellular structures of the photosyn-thetic apparatus and the reaction centers arelocated in the cytoplasmic membrane. Bacterio-chlorophyll g confers to the cells a near infraredabsorption maximum at 788 nm, which is uniqueamong photosynthetic organisms. The knownspecies of Heliobacteriaceae all grow photohet-erotrophically and are strict anaerobes.
Oxygenic photosynthesis is only found inmembers of a single bacterial lineage out of the
five that contain phototrophs (Fig. 1). TheCyanobacteria by far comprise the largest num-ber of isolated strains and described species(Table 1). The Cyanobacteria (= oxyphotobacte-ria) are defined by their ability to carry outoxygenic photosynthesis (water-oxidizing,oxygen-evolving, plant-like photosynthesis)based on the coordinated work of two photosys-tems (Fig. 3C). Phylogenetically, they constitutea coherent phylum that contains the plastids ofall eukaryotic phototrophs. They all synthesizechlorophyll a as photosynthetic pigment, and
Fig. 3. Continued.

CHAPTER 1.3 The Phototrophic Way of Life 39
most types contain phycobiliproteins as light-harvesting pigments. These multimeric proteina-ceous structures are found on the cytoplasmicface of the intracellular thylakoid membranesand contain phycobilins as light-harvesting pig-ments. All Cyanobacteria are able to grow usingCO2 as the sole sou rce of carbon, which they fixusing primarily the reductive pentose phosphatepathway (see Carbon Metabolism of Pho-totrophic Prokaryotes in this Chapter). Theirchemoorganotrophic potential typically isrestricted to the mobilization of reserve poly-mers (mainly starch but also polyhydroxyal-kanoates) during dark periods, although somestrains are known to grow chemoorganotrophi-cally in the dark at the expense of external sug-ars. Owing to their ecological role, in many casesindistinguishable from that of eukaryoticmicroalgae, the cyanobacteria had been studiedoriginally by botanists. The epithets “blue-greenalgae,” “Cyanophyceae,” “Cyanophyta,” “Myxo-phyceae,” and “Schizophyceae” all apply to thecyanobacteria. Two main taxonomic treatmentsof the Cyanobacteria exist, and are widely used,which divide them into major groups (orders) on
the basis of morphological and life-history traits.The botanical system (Geitler, 1932 recognized 3orders, 145 genera and some 1300 spe cies, but ithas recently been modernized (Anagnostidis andKomárek, 1989, Komárek and Anagnostidis,1989). The bacteriological system (Stanier, 1977;Rippka et al., 1979; Castenholz, 1989), relies onthe study of cultured axenic strains. It recognizesfive larger groups or orders, separated on thebasis of morphological characters. Genetic (i.e.,mol% GC, DNA-DNA hybridization) as well asphysiological traits have been used to separategenera in problematic cases.
Previously, a separate group of organisms withequal rank to the cyanobacteria, the so-called“Prochlorophytes” (with two genera, Prochlo-ron, a unicellular symbiont of marine inverte-brates, and Prochlorothrix, a free-livingfilamentous form) had been recognized (Lewin,1981). They were differentiated from cyanobac-teria by their lack of phycobiliproteins (Fig. 2)and the presence of chlorophyll b. The recentlyrecognized genus Prochlorococcus of marinepicoplankters could be included here, eventhough the major chlorophylls in this genus are
Table 1. Groups of photosynthetic prokaryotes and their characteristics.
aThe numbers of photosynthetic species described for each taxon are given in parenthesis.BChl = bacteriochlorophyll, car = carotenoids, Chl = chlorophyll, cls = chlorosomes, icm = intracellular membranes,PBS = phycobilisomes, thy = thylacoids.
Taxon Preferred growth mode Light harvesting Photochemical reaction
Chloroflexus subdivision (3)a Anoxygenicphotoorganoheterotroph(cls);
Aerobic chemoorganoheterotroph
BChl c, car Type II reaction center
— —
Green sulfur bacteria (15) Anoxygenic photolithoautotroph cls; BChl cldle, car Type I reaction center
a-Proteobacteria (31) Anoxygenicphotoorganoheterotroph
icm; BChl alb, car Type II reaction center
Aerobic chemoorganoheterotroph — —
a-Proteobacteria(aerobicphotosynthetic)
(23) Aerobic chemoorganoheterotroph BChl a Type II reaction center
b-Proteobacteria (4) Anoxygenicphotoorganoheterotroph
icm; BChl a, car Type II reaction center
Aerobic chemoorganoheterotroph — —
ChromatiaceaeEctothiorhodospiraceae
(31)(9)
Anoxygenic photolithoautotroph icm; BChl alb, car Type II reaction center
Heliobacteriaceae (5) Anoxygenicphotoorganoheterotroph
BChl g, car Type I reaction center
Cyanobacteria (>>1000)
Oxygenic photolithoautotroph thy; Chl a + PBSor Chl b, or Chl d; car
Type I + II reactioncenter
Prochloron,Prochlorothrix
(2) thy; Chl a/b,car
Prochlorococcus (1) thy; Chl a2/b2, car (PBS)
Acaryochloris (1) thy; Chla,d, car (PBS)
Halobacteria (3) Aerobic chemoorganoheterotroph Purple membrane;bacteriorhodopsin
Bacteriorhodopsin

40 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
divinyl-Chl a and divinyl-Chl b. FourteenProchloron isolates from different localities andhosts have been found to belong to a single spe-cies by DNA-DNA hybridization studies (Stamet al., 1985; Holtin et al., 1990). Some of theoriginal distinctions leading to the separation ofthe Chl b-containing oxyphotobacteria from thecyanobacteria are questionable, since at least inone strain of Prochloroccoccus marinus, func-tional phycoerythrin (Lokstein et al., 1999), andgenes encoding for phycobiliproteins have beendetected (Lokstein et al., 1999). Additionally,phylogenetic analysis of 16S rRNA genes indi-cate that the three genera of Chl b-containingprokaryotes arose independently from eachother and from the main plastid line (see Evolu-tionary Considerations in this Chapter), a resultthat is supported by the comparative sequenceanalysis of the respective Chl a/b binding pro-teins (Laroche et al., 1996; Vanders taay et al.,1998). Thus “Prochlorophytes” are just greenishcyanobacteria, and are not treated separatelyhere. The recent discovery of Chl d-containingsymbionts in ascidians (Acaryochloris marina,Miyashita et al., 1996) once again demonstratesthe evolutionary diversification of light-harvest-ing capabilities among oxyphotobacteria (seeCompetition for Light in this Chapter). Whilethe phylogenetic affiliation of Acaryochlorismarina has not been presented as yet, ultrastruc-tural and chemotaxonomic characters predictthat A. marina belongs to the cyanobacterialradiation as well.
According to phylogenetic analysis of 16SrRNA sequences, the Cyanobacteria are adiverse phylum of organisms within the bacterialradiation, well separated from their closest rela-tives (Giovanonni, 1988; Wilmotte, 1995; Turner,1887; Garcia-Pichel, 1999; Fig. 1). These analysessupport clearly the endosymbiotic theory for theorigin of plant chloroplasts, as they place plastids(from all eukaryotic algae and higher plantsinvestigated) in a diverse, but monophyletic,deep-branching cluster (Nelissen et al., 1995).Phylogenetic reconstructions show that thepresent taxonomic treatments of the cyanobac-teria diverge considerably from a natural systemthat reflects their evolutionary relationships. Forexample, separation of the orders Chroococcalesand Oscillatoriales (Nelissen et al., 1995; Reeves,1996), and perhaps also the Pleurocapsales(Turner, 1887; Garcia-Pichel et al., 1998) is notsupported by phylogenetic analysis. The hetero-cystous cyanobacteria (comprising the twoorders Nostocales and Stigonematales) formtogether a monophyletic group, with relativelylow sequence divergence, as low as that pre-sented by the single accepted genus Spirulina(Nübel, 1999). A grouping not corresponding toany official genus, the Halothece cluster, gathers
unicellular strains of diverse morphology thatare extremely tolerant to high salt and stem fromhypersaline environments (Garcia-Pichel et al.,1998). A second grouping, bringing together verysmall unicellular ope n-ocean cyanobacteria(picoplankton) includes only marine picoplank-tonic members of the genera Synechococcus andall Prochlorococcus. Several other statisticallywell-supported groups of strains that may or maynot correspond to presently defined taxa can bedistinguished. The botanical genus “Microcystis”of unicellular colonial freshwater plankton spe-cies is very well supported by phylogeneticreconstruction, as is the genus Trichodesmium offilamentous, nonheterocystous nitrogen-fixingspecies typical from oligotrophic marine plank-ton of the tropics. The picture that emerges fromthese studies is that sufficient knowledge of eco-logical and physiological characteristics can leadto a taxonomic system that is largely congruentto the 16S rRNA phylogeny.
A different principle of conversion of lightenergy into chemical energy is found in the Halo-bacteria. These archaea are largely confined tosurface layers of hypersaline aquatic environ-ments and grow predominantly by chemoorga-noheterotrophy with amino or organic acids aselectron donors and carbon substrates, generat-ing ATP by respiration of molecular oxygen. Inthe absence of oxygen, several members arecapable of fermentation or nitrate respiration.At limiting concentrations of oxygen, at leastthree of the described species of Halobacteria(Halobacterium halobium, H. salinarium, H. sod-omense) synthesize bacteriorhodopsin (Oester-helt and Stoeckenius, 1973), a chromoproteincontaining a covalently bound retinal. Bacterior-hodopsin is incorporated in discrete patches inthe cytoplasmic membrane (“purple mem-brane”). However, these prokaryotes have onlya very limite d capability of light-dependentgrowth. Only slow growth and one to two celldoublings could be demonstrated experimentally(Hartmann et al., 1980; Oesterhelt and Krippahl,1983). The fact that rhodopsin-based photosyn-thesis has been found only in the phylogeneti-cally tight group of Halobacteria may indicatethat, because of its lower efficiency, this type oflight utilization is of selective advantage onlyunder specific (and extreme) environmental con-ditions. Further information on the biochemistry,physiology and ecology of this group may befound in the chapters, Introduction to theClassification of Archaea and The Family Halo-bacteriaceae.
During the past years, culture-independent16S rDNA-based methods have been used forthe investigation of the composition of naturalcommunities of phototrophic prokaryotes. Thesestudies have provided evidence that more than

CHAPTER 1.3 The Phototrophic Way of Life 41
one genotype of Chloroflexus occur in one hotspring microbial mat and that four previouslyunkown sequences of cyanobacteria dominate inthe same environment (Ferris et al., 1996; Ruff-Roberts et al., 1994; Weller et al., 1992). Simi-larly, nine different partial 16S rDNA sequencesof Chromatiaceae and green sulfur bacteria,which differed from all sequences previouslyknown, were retrieved from two lakes and oneintertidal marine sediment (Coolen and Over-mann, 1998; Overmann et al., 1999a).
However, 16S RNA signatures from naturalpopulations were indistinguishable from those ofcultured strains in the case of cyanobacteria withconspicuous morphologies, such as the cosmopol-itan Microcoleus chthonoplastes (Garcia-Pichelet al., 1996) from intertidal and hypersalinemicrobial mats or Microcoleus vaginatus fromdesert soils (F. Garcia-Pichel, C. López-Cortésand U. Nübel, unpublished observations). In asimilar manner, the 16S rRNA sequence of anisolated strain of Amoebobacter purpureus(Chromatiaceae) was found to be identical to theenvironmental sequence dominating in thechemocline of a meromictic salt lake (Coolen andOvermann, 1998; Overmann et al., 1999a). Obvi-ously, the limited number of isolated and charac-terized bacterial strains rather than an alleged“nonculturability,” at least in some cases,accounts for our inability to assign ecophysiolog-ical properties to certain 16S rRNA sequencetypes. This point is illustrated for extremely hal-otolerant unicellular cyanobacteria by the factthat only after a physiologically coherent groupof strains was defined on the basis of newly char-acterized isolates (Garcia-Pichel et al., 1998)could the molecular signatures retrieved fromfield samples be assigned correctly.
It has to be concluded that 1) the numbers ofspecies listed in Table 1 do not reflect the fullphylogenetic breadth at least in the four groupsof anoxygenic phototrophic prokaryotes as wellas in morphologically simple Cyanobacteria, and2) that the physiology and ecology of those spe-cies of phototrophic prokaryotes that are domi-nant in the natural environment in some casesmay differ considerably from known type strains.
Habitats of Phototrophic Prokaryotes
Bacteria of the Chloroflexus-subgroup formdense microbial mats in geothermal springs,often in close association with cyanobacteria.Chloroflexus aurantiacus is a thermophilic bacte-rium which grows optimally between 52 and60!C and thrives in neutral to alkaline hotsprings up to 70–72!C. Of all anoxygenic pho-totrophic bacteria isolated so far, only Chlorof-
lexus aurantiacus is capable of growth up to74!C. In contrast to the domain Archaea, nohyperthermophilic species are known from thedomain Bacteria. The phylogenetically relatedHeliothrix oregonensis grows optimally between50 and 55!C and is abundant as a flocculant sur-face layer in a few alkaline springs in Oregon.Hydrothermal springs of 56–66!C, which containsulfide of geothermal origin, are dominated by asurface layer or a “unispecific” mat of Chlorof-lexus (Castenholz and Pierson, 1995). Because ofthe absence of cyanobacteria in some of thesesystems, Chloroflexus presumably growsautotrophically (Pierson and Castenholz, 1995).In the presence of O2, the mats exhibit an orangecolor whereas they are green under anoxic con-ditions (Castenholz and Pierson, 1995). Theorange color is the result of the enhanced caro-tenoid biosynthesis under oxic conditions (seeChemotrophic Growth with O2 in this Chapter).In the absence of sulfide, Chloroflexus is presentas a distinct orange layer beneath a surface layerof cyanobacteria and may utilize their exudatesor the fermentation products generated duringdecomposition of cyanobacteria. Molecular oxy-gen represses bacteriochlorophyll synthesis inChloroflexus and often is present at saturationlevels in the orange layers. Since bacteriochlorophylls a and c are still present in this layer, how-ever, it must be assumed that bacteriochloro-phylls are synthesized at anoxic conditionsduring nightime (Castenholz and Pierson, 1995).
Green and purple sulfur bacteria often formconspicuous blooms in non-thermal aquatic eco-systems (Figs. 4, 5A, 5B), although moderately
Fig. 4. Bright field photomicrograph of the bacterioplanktoncommunity thriving in the chemocline of the meromicticBuchensee (near Radolfzell, Germany) during autumn. Thedominant anoxygenic phototroph at this time of the year isthe green sulfur bacterium Pelodictyon phaeoclathratiforme(brown cells, which appear in chains or netlike colonies). Inaddition, phototrophic consortia (“Pelochromatium roseum,”one consortium in the center) are found. Similar to Pld.phaeoclathratiforme, most of the colorless bacterial cellsfound in the chemocline contain gas vesicles as is evidentfrom their highly refractile appearance in the bright field.

42 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
thermophilic members of the genera Chroma-tium and Chlorobium have been described fromhot spring mats (Castenholz et al., 1990). Chlo-robium tepidum occurs in only a few NewZealand hot springs at pH values of 4.3 and 6.2and temperatures up to 56!C. Chromatium tepi-dum was found in several hot springs of westernNorth America at temperatures up to 58!C andmight represent the most thermophilic proteo-bacterium (Castenholz and Pierson, 1995). In arecent compilation (van Gemerden and Mas,1995), 63 different lakes and 7 sediment ecosys-tems harboring phototrophic sulfur bacteriawere listed. Cell densities between 104 and107·ml-1 and biomass concentrations between 10and 1000 mg bacteriochlorophyll·l-1 are commonin pelagic habitats. Of the purple sulfur bacteria,Chromatiaceae are typically found in freshwaterand marine environments (Fig. 5A, B) whereasEctothiorhodospiraceae inhabit hypersalinewaters. The phototrophic sulfur bacteria growpreferentially by photolithoautotrophic oxida-
tion of reduced sulfur compounds and are there-fore limited to those environments where lightreaches anoxic, sulfide-containing bottom layers.Because light and sulfide occur in opposing gra-dients, growth of phototrophic sulfur bacteria isconfined to a narrow zone of overlap and is onlypossible if the chemical gradient of sulfide is sta-bilized against vertical mixing. In pelagic envi-ronments like lakes or lagoons, chemicalgradients are stabilized by density differencesbetween the oxic and anoxic water layers. Suchdensity differences are either the result of ther-mals tratification and mostly transient (as inholomictic lakes) or are caused by high salt con-centrations of the bottom water layers, in whichcase stratification is permanent (meromicticlakes). Pelagic layers of phototrophic sulfur bac-teria extend over a vertical distance of 10 cm(van Gemerden and Mas, 1995; Overmann et al.,1991a) up to 30 m (Repeta et al., 1989) and reachbiomass concentrations of 28 mg bacteriochloro-phyll·l-1 (Overmann et al., 1994).
Fig. 5. Multilayered microbial mat as it is regularly found in the sandflats of Great Sippewissett Salt Marsh (Cape Cod,Massachusetts, USA). A. In most instances, the mats consist of a top green layer, an intermediate purple layer, and a grayishto blackish bottom layer. B. Fully developed microbial mats consist (from top) of an olive-green layer of diatoms andcyanobacteria, a green layer consisting mostly of cyanobacteria, a purple layer of purple sulfur bacteria, a peach-colored layerformed by BChl b-containing purple sulfur bacteria (morphologically similar to Thiocapsa pfennigii), and a greyish to blackishbottom layer.

CHAPTER 1.3 The Phototrophic Way of Life 43
Littoral sediments represent the second typeof habitat of phototrophic sulfur bacteria. Inthese systems, turbulent mixing is largely pre-vented by the sediment matrix, and diffusion isthe only means of mass transport. Gradients oflight and sulfide are much steeper, and thefluxes of sulfide much larger compared to thepelagic environment. These conditions allowlayers of phototrophic sulfur bacteria in sedi-ments to reach much higher biomass densities(up to 900 mg bacteriochlorophyll·dm-3; vanGemerden et al., 1989) than in lakes. At thesame time, the layers are very narrow (1.3–5 mm; van Gemerden and Mas, 1995; Fig. 5A).This vertical distribution of anoxygenic pho-totrophic biomass ultimately determines thesignificance of microbial sulfide oxidation forthe sulfur cycle in these ecosystems (see Signifi-cance of Anoxygenic Photosynthesis for thePelagic Carbon and Sulfur Cycles in thisChapter). The spectral compos ition of lightavailable for anoxygenic photosynthesis is con-siderably different between pelagic and benthichabitats (Fig. 6) and selects for different speciesof anoxygenic phototrophic bacteria. Whereaslight of the blue to yellow-green wavelengthbands dominates the depths of most lakes,infrared light is an important source of energyin benthic microbial mats (see Light Energy
and the Spectral Distribution of Radiation inthis Chapter).
The dominance of certain species of green sul-fur bacteria (Fig. 4) or Chromatiaceae in pelagicenvironments in many cases can be explained bytheir specific light-harvesting capabilities (seeLight Absorption and Light Energy Transfer inProkaryotes and Competition for Light in thisChapter) and other phenotypic traits. Typically,those species that have been isolated from natu-ral blooms in lakes are obligately photo-lithotrophic, lack assimilatory sulfate reduction,cannot reduce nitrate, and assimilate only feworganic carbon sources (see Carbon Metabolismof Phototrophic Prokaryotes in this Chapter).This applies not only to all green sulfur bacteriabut also to the dominant species of Chromati-aceae. Obviously, in the chemocline of lakes themetabolic versatile Chromatiaceae species haveno selective advantage. As judged from thephysiological characteristics of strains of pho-totrophic sulfur bacteria isolated from sedi-ments, the pronounced diurnal variations inoxygen concentrations and salinity, together withthe different light quality, select for different spe-cies composition in benthic microbial mats. Thepurple sulfur bacterium Chromatium (and themulticellular gliding colorless sulfur bacteriumBeggiatoa) are found in many microbial mats
Fig. 6. Effects of the habitat on the physical exposure of cyanobacteria. The spectral scalar irradiance (sun and sky radiation)incident at ground level at noon in a clear midsummer day at 41!N is plotted in Plate I. The rest of the plates depict the insitu scalar irradiance experienced by cyanobacterial cells thriving in several habitats exposed to the incident fluxes in plateI (note different scales). Plate II: a “strong shade” habitat (North-facing surface illuminated by extremely diffuse sky radiationonly), where scalar irradiance is very low but the relative importance of UV is enhanced. Plate III: a planktonic habitat(under 1 m of clear open-ocean water), where all fluxes remain fairly high and UVB and visible are more strongly attenuatedthan UVA. Plate IV: the surface of beach (quartz, feldspar) sand, where all UVB, UVA, and visible are higher than incident(by 120, 150, and 205%, respectively) due to light trapping effects. Plate V: 300-m deep in a wet topsoil, where UVB and UVA have been attenuated below 5% of incident but ca. 20% of the visible light remains. Plate VI: scalar irradiance withinthe thallus of the terrestrial cyanobacterial lichen Collema sp. Modified from Castenholz and Garcia-Pichel, 1999, after datafrom the following sources: F. Garcia-Pichel (unpublished observation); Garcia-Pichel, 1995; Büdel et al., 1997; and Smithand Baker, 1981.
1.5
0
300
400
500
600
700
I0.03
0
300
400
500
600
700
II1.3
0
300
400
500
600
700
III
2.9
0
300
400
500
600
700
IV0.6
0
300
400
500
600
700
V0.9
0
300
400
500
600
700
VI
Wavelength (nm)
Sca
lar
Irra
dian
ce (
W m
–2 n
m–1
)

44 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
and exhibit diurnal vertical migrations inresponse to the recurrent changes in environ-mental conditions (Jørgensen, 1982; Jørgensenand Des Marais, 1986). Microbial mats of inter-tidal sediments are typically colonized by theimmotile purple sulfur bacterium Thiocapsaroseopersicina and small motile thiobacilli (vanden Ende et al., 1996).
In contrast to the phototrophic members ofthe g-Proteobacteria, purple nonsulfur bacteriaof the a- and b-subclasses of Proteobacteria donot appear to form dense accumulations undernatural conditions (Biebl and Drews, 1969; Swo-ager and Lindstrom, 1971; Steenbergen andKorthals, 1982). However, purple nonsulfur bac-teria can be readily isolated from a wide varietyof marine, lacustrine and even terrestrialenvironments (Imhoff and Trüper, 1989; J.Overmann, unpublished observation). Whilecomprehensive comparative quantitation of theecological importance of purple nonsulfur bacte-ria is still lacking, as many as ca. 106 c.f.u. ofpurple nonsulfur bacteria could be cultivated percm3 of sediment in coastal eutrophic settings(Guyoneaud et al., 1996).
Generally, aerobic phototrophic bacteriathrive in eutrophic marine environments. Obli-gately aerobic bacteria containing bacteriochlo-rophyll a have been isolated from beach sandand seaweeds (thalli of Enteromorpha linza andSargassum horneri; Shiba et al., 1979), and insome cases also from freshwater ponds andmicrobial mats. At least some of the aerobic pho-totrophic bacteria apparently can survive in situtemperatures of up to 54!C (Yurkov and Beatty,1998). Aerobic phototrophic bacteria were iso-lated from hydrothermal plume water of a blacksmoker 2000 m below ocean surface (Yurkov andBeatty, 1998); acidophilic strains could be iso-lated from acidic mine drainage. Typically, Meth-ylobacterium species are isolated from foods,soils and leaf surfaces (Shimada, 1995). Photo-synthetic Rhizobium strains are widely distrib-uted in nitrogen-fixing stem nodules of thetropical legume Aeschynomene spp. where theyare present as symbiosomes. Similar strains havealso been found in root and hypocotyl nodulesof Lotononis bainesii (Fabaceae). These photo-synthetic rhizobial and regular symbiosomes dif-fer in that the former contains only one largespherical bacteroid. The photosynthesis of theseendosymbionts may provide energy for nitrogenfixation and permit a more efficient growth of thehost plant, since up to half of the photosynthateproduced by legumes is allocated to nitrogen fix-ation (Fleischman et al., 1995).
Heliobacteriaceae appear to be primarily soilbacteria and have been isolated from dry paddyfields or other soils throughout the world (Madi-gan and Ormerod, 1995). Bacteria of this family
may even represent the dominant anoxygenicphototrophic bacteria in soil (Madigan, 1992).Occasionally, strains also have been isolatedfrom lakeshore muds and hot springs (Amesz,1995; Madigan and Ormerod, 1995). Heliobacte-rium modesticaldum grows up to 56!C (Kimbleet al., 1995). Spore formation may offer aselective advantage to Heliobacterium modesti-caldum, Heliophilum fasciatum, and Heliobacte-rium gestii in their main habitat (rice field soil),which undergoes periodic drying and concomi-tantly becomes oxidized (Madigan, 1992). Dur-ing growth of the rice plants, organic compoundsexcreted by their roots could provide sufficientsubstrates for photoheterotrophic growth of theHeliobacteriaceae.
Cyanobacteria as a group exhibit the widestrange of habitats of all phototrophic prokaryotesdue to the ubiquity of water, their preferred elec-tron donor for the reduction of CO2. In principle,cyanobacteria can thrive in any environment thathas, at least temporarily, liquid water and sun-light. They are known from Antarctic endolithichabitats and from hot springs. More than 20 spe-cies of cyanobacteria (Castenholz and Pierson,1995) are thermophilic. Effectively, however, nocyanobacteria are known from acidic environ-ments (below pH 4.5) and competition witheukaryotic microalgae or higher plants mayrestrict their growth in other environments.Cyanobacteria are found in the plankton ofcoastal and open oceans and in freshwater andsaline inland lakes. They thrive in the benthos ofmarine intertidal (Fig. 5B), lacustrine and fluvialwaters and in a large variety of terrestrial habi-tats (soils, rocks, trees). Symbiotic associationsare common.
In the marine plankton, the phycoerythrin-containing Synechococcus often represents amajor fraction of all primary producers. Thesame holds true for Prochlorococcus (Campbelland Vaulot, 1993; Chisholm et al., 1988; Olson etal., 1990b). Compared with the high number ofcyanobacterial species found in freshwaterplankton, intertidal areas, and hypersaline envi-ronments, the diversity of this group is very lim-ited in the open ocean (Carr and Mann, 1994).The predominant group invariably consists ofsmall (<2 m) mostly nonmotile, non-nitrogen-fixing single cells assigned to the genus Synecho-coccus, which is found in the photic zone of alloceans except in the coldest areas. As a charac-teristic feature, the cells contain phycoerythrinas accessory photopigment which confers anorange autofluorescence on the cells. Despitetheir similar phenotype, marine Synechococcusstrains are genetically heterogenous (Waterburyet al., 1986). An important component of thephytoplankton in tropical and subtropical oceansare the filamentous Trichodesmium spp. (Carr

CHAPTER 1.3 The Phototrophic Way of Life 45
and Mann, 1994). The bundle and aggregateforming Trichodesmium typically develop intoblooms that can extend kilometers long and aredetected on the surface of oligotrophic tropicaland subtropical oceans with the naked eye orwith satellite imagery from space. The success ofTrichodesmium can be mainly traced to thehighly efficient nitrogen-fixing capacity of thesenonheterocystous cyanobacteria. Their activitiesattain global magnitude for the nitrogen cycle(Capone et al., 1977). Heterocystous, nitrogen-fixing cyanob acteria of the genera Nodularia,Anabaena, and Aphanizomenon bloom inmesotrophic and eutrophic fresh and brackishwaters. Together with the blooms of the nonhet-erocystous genus Microcystis, these cyanobacte-ria have become a real environmental concern,not only because of their effects of overall waterquality but also because of their ability to pro-duce toxins, which are known to have caused thedeaths of humans and cattle. In the chemoclineof stratified lakes, deep blooms of cyanobacteriaoccur frequently.
Edaphic cyanobacteria are also distributedworldwide, especially in soils of basic pH;sheathed oscillatorian forms (Microcoleus vagi-natus, “Schizothrix” spp.), along with heterocys-tous ones (Nostoc, Scytonema) are of majorecological relevance in arid and semiaridregions where growth of higher plants isrestricted. In such environments, cyanobacteriaadopt a life strategy of resistance to desiccation(Potts, 1994) making use of the few occasions inwhich liquid water is available from rain or dew.Very intense productivity spurts occur in a mat-ter of minutes after wetting (Garcia-Pichel andBelnap, 1996). The so-called “cyanobacterialdesert crusts” contribute significantly to the bio-geochemistry and to the physical stability ofarid soils. Other important terrestrial habitatsof cyanobacteria are the surface or subsurfa ceof rocks: extensive endolithic cyanobacterialcommunities, usually dominated by membersof the genus Chroococcidiopsis, have beendescribed from tropical, desert and polar envi-ronments (Friedmann, 1982; Wessels and Büdel,1995).
In the course of evolution, cyanobacteria haveentered into symbiotic associations with a multi-tude of organisms. These have reached a widerange in the degree of interdependence betweenpartners (see Symbiosis between PhototrophicBacteria and Eukaryotes in this Chapter).
Principles and Prerequisites of Photosynthesis
Bacterial photosynthesis can be divided into twodifferent types of reactions 1) the light reaction,
in which light energy is trapped and convertedinto ATP (via a proton-motive force DP) and areduced redox carrier Rred·H+, and 2) the so-called dark reaction of biosynthetic carbonreduction.
Light reaction: 2H2A + 2Rox + lightÆ 2A + 2Rred·H + DP
ADP + PI + DP Æ ATP + H2O
Dark reaction: CO2 + ATP + 2Rred·H + H2OÆ <CH2O> + ADP + Pi + 2Rox
Sum: CO2 + 2H2A Æ <CH2O> + H2O + 2A(van Niel equation)
Microorganisms have found different ways toaccomplish these two tasks.
Light Energy and the Spectral Distribution of Radiation
The present day solar irradiance at the averagedistance of Earth to the sun and outside theatmosphere (the so-called solar constant) is1353·W m–2 (Kirk, 1983). The spectral energy dis-tribution of this solar radiation approximatesthat of a black body at 6000!K (the surface tem-perature of the sun). According to Wien’s Law, ablack body at this temperature has a maximumemission of electromagnetic energy at about480 nm. The actual spectral energy distributionof solar radiation exhibits minima which reflectthe absorption bands of hydrogen in the outeratmosphere of the sun (Fig. 7). The total lightenergy received by the Earth is 5.46·1024 J·year-1,which would correspond to 339.4 W·m-2. Theactual solar (time and space-averaged) irradi-ance reaching the surface of the Earth amountsonly to 160 W·m-2 (Gates, 1962; Dietrich et al.,1975). This large reduction is due to Raleigh scat-tering by air molecules and dust particles, and oflight absorption by water vapor, O2, O3 and CO2
during the passage of radiation through theEarth’s atmosphere. Concomitantly, the spectraldistribution of solar irradiance is changed espe-cially because water vapor absorbs infrared light(Fig. 7). At sea level, light of the wavelengthregions 400–700 nm (PAR, photosyntheticallyavailable radiation) constitutes 50% of this irra-diance (Kirk, 1983).
Based on estimates for global primary produc-tivity, only 0.13% of the flux of solar energyreaching the surface of the Earth is convertedinto chemical energy by photosynthesis (Odum,1983; see Introduction in this Chapter). Undernatural conditions, photosynthesis of the variousgroups of phototrophic prokaryotes is limited bydifferent environmental factors including light,reduced sulfur compounds, organic carbon sub-strates, oxygen, and temperature. The physical

46 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
characteristics of the medium have, through pro-cesses of absorption and scattering, a large influ-ence on the available radiation (see Competitionfor Light in this Chapter). As a second majorlimiting factor, the availability of nutrients limitsthe growth of phototrophic bacteria and as aconsequence, photosynthetic energy conversion.
Surface environments exposed to sky radia-tion (as in strong shades) may be enriched inblue and UV radiation (Fig. 6). Water is themajor light-absorbing component only in veryclear open ocean and inland lakes. It stronglyabsorbs light of the ultraviolet, red and especiallyinfrared (wavelengths around 745 and 960 nm).As a consequence, tens of meters below the sur-face of clear waters the spectrum is enriched inblue wavelengths. Several meters below coastalor most lacustrine water surfaces, the spectrumis enriched towards the green wavelengths, anddeep(several millimeters) in the photic zones of
sediments and soils infrared wavelengths domi-nate. Yellow substance in lakes is mostly of ter-restrial origin and particularly absorbs light ofthe ultraviolet and blue portion of the spectrum(Kirk, 1983). In dystrophic lakes in which highconcentrations of humic compounds are themajor light-absorbing components, light of thered wavelength range prevails such that green-colored species of green sulfur bacteria have aselective advantage over their brown-coloredcounterparts or purple sulfur bacteria (Parkinand Brock, 1980a).
In benthic and soil ecosystems, light qualitydiffers fundamentally from that in the pelagicenvironment. In the visible wavelength range,radiation is strongly attenuated by mineral andbiogenic particles. In sandy sediments lightattenuation occurs preferentially in the wave-length range of blue light due to the reflection bysand grains (Jørgensen and Des Marais, 1986;Kühl and Jørgensen, 1992). The presence of ironminerals results in an enhanced attenuation ofUV and blue wavelengths (Garcia-Pichel andBelnap, 1996). In contrast, absorption of infraredlight by sediment particles is low and absorptionby water is negligible due to the short opticalpathlength. As a consequence of the opticalproperties of the sediment particulates, the redand infrared portion of the spectrum penetrateto the deepest levels. Multiple scattering causesthe light fields to become rapidly diffuse, so thatbacteria thriving within these environmentsreceive light from all directions. The parametermeasuring light received at a point in space fromall directions is called scalar irradiance (E0, orphoton fluence rate). A third important, butcounterintuitive, phenomenon is the presence ofmaximum irradiance values close to the surface,which are even larger than the incident scalarirradiance (Fig. 6). Below this surficial zonewhere the E0 maximum occurs, E0 attenuatesexponentially (Jørgensen and Des Marais, 1986;Jørgensen and Des Marais, 1988; Kühl and Jør-gensen, 1992; Lassen et al., 1992). For visiblelight, the measured photic depths (depths whereE0 is attenuated to 1% of the incident) variedbetween 3.1 mm for quartz sand and 0.45 mm forsilty muds (Garcia-Pichel and Bebout, 1996b). Inthe ultraviolet (UV) at 310 nm, the correspond-ing depths were only 1.25 and 0.23 mm.
Light Absorption and Light Energy Transfer in Prokaryotes
PrincipleThe chlorophyll-based photosystems of bacteriaconvert electromagnetic energy into a redoxgradient. The redox reactions are initiated by
Fig. 7. Spectral energy distribution of solar radiation outsidethe atmosphere and at sea level as compared to the absorp-tion spectra of various phototrophic bacteria. Absorptionspectra of the purple nonsulfur bacterium Rhodospirillumrubrum (containing BChl a, spirilloxanthin), Blastochlorisviridis (BChl b, 1,2-dihydroneurosporene), and Roseospiril-lum parvum (BChl a, spirilloxanthin, lycopenal), of the Chro-matiaceae species Thiospirillum jenense (BChl a,lycopene,rhodopin) and Chromatium okenii (BChl a, okenone), of theChlorobiaceae species Chlorobium limicola (BChl c, chloro-bactene) and Chlorobium phaeobacteroides (BChl e, i sore-nieratene) and of a cyanobacterium (Chl a, phycocyanin) aredepicted.
Irra
dian
ce (
W m
–2 n
m–1
) Black body 6000k
Outside atmosphereSea level
400 600 800 1000Wavelength (nm)
Rel
ativ
e ab
sorp
ilon
Rhodospirillum rubrumBlastochloris viridisRoseospirillum parvum
Thiospirillum jenenseChromatium okenii
Chlorobium limicolaChl phaeobacteroides
Cyanobacterium(phycocyanin)
200 2000 nm
2.0
1.5
1.0
0.5

CHAPTER 1.3 The Phototrophic Way of Life 47
absorption of electromagnetic energy, leading toa transition of specific molecules into an excitedelectronic state. An increase in the electronicenergy of a molecule requires more energy thanchanges in vibrational or rotational states. Sincethe energy of light quanta is inversely related totheir wavelength (Planck’s Law), moleculesabsorb electromagnetic radiation of short wave-lengths (ultraviolet and visible light) duringchanges in electronic energy, and longer wave-lengths during changes in vibrational (near infra-red radiation) and rotational energy (far infraredradiation and microwaves). Changes in the elec-tronic state of molecules, and thus photochemi-cally driven redox reactions by light absorption,can only occur by absorption of quanta of wave-lengths <1240 nm (i.e., an energy larger than1 eV per electron). This fact obviously limits thewavelength range that is usable for photochemi-cal reactions. The major fraction of solar energyis present in the wavelength range between 400and 750 nm. These wavelengths can only be har-vested by organic molecules containing delocal-ized p-electrons in conjugated double bonds(Fig. 7).
Pigments and Light-Harvesting ComplexesTo capture light for photosynthesis, phototrophicorganisms employ three classes of pigment mol-ecules: magnesium porphyrins (chlorophylls andbacteriochlorophylls, also called chlorins), open-chain tetrapyrrole bilin pigments (phycobilins),and carotenoids. However, other types of chro-mophores may be used in non-photosyntheticlight-harvesting, as is the case of the flavins andpterines of DNA-photolyase (Tanada et al., 1997)and in specific regulatory photoreceptors (Halo-bacteriaceae, bacteriorhodopsin). Until recentlyit appeared that only the magnesium-containing
chlorin molecules were employed as the majorphotosynthetic pigment. The aerobic photosyn-thetic bacterium Acidiphilium rubrum is the firstphotosynthetic organism known to employ zinc-containing bacteriochlorophyll a as the photo-chemically active pigment (Wakao et al., 1996).
Free molecules remain in the excited singletstate for as little as 10-8 to 10-9 sec and rapidlyreturn to the ground state (fluorescence).Through the multiplicity of vibrational and rota-tional states associated with each electronicenergy level, two different electronic energystates may overlap. In such molecules the lower-most electronic energy level (the lowest excitedsinglet state) is reached in a rapid series of radi-ationless transitions with a concomitant smalldecrease in free energy. The wavelengths emittedduring the subsequent return of the electron tothe ground state therefore is longer than thosewavelengths that were absorbed (Stokes shift).Chlorophylls and bacteriochlorophylls exhibittwo major absorption bands (Table 2) and, whenexcited in the dissolved state, a correspondingred (685 nm for chlorophyll a) or infrared(786 nm for bacteriochlorophyll a) fluorescence.In photosynthetically active c ells, however, onlyabout 1% of the absorbed light energy is lost byfluorescence. It is a characteristic of the photo-synthetic apparatus of living organisms, thatfluorescence (hence loss of already absorbedenergy) is minimized. Instead, most of theenergy absorbed by the antenna pigments ischanneled by vectorial and radiationless induc-tive dipole resonance toward the reaction cen-ters, where it drives the photochemical redoxreactions. The specific coordination of pigmentmolecules in photosynthetic organisms favorsinductive resonance and photochemical reac-tions over fluorescence. Within the photosyn-thetic antenna, a fine modulation of the
Table 2. Major absorption maxima of chlorins in whole cells and in the dissolved state, and fluorescence maxima of wholecells of phototrophic prokaryotes.
aBacteriochlorophyll g of the Heliobacteriaceae shows structural relationships to chlorophyll a because it contains a vinylgroup on tetrapyrrole ring I. Like in bacteriochlorophylls a and b, pyrrole ring II is reduced, however, and the esterifyingalcohol is famesol as in bacteriochlorophylls of green sulfur bacteria. As for bacteriochlorophyll a or b, the reduced state ofring II in bacteriochlorophyll g causes an additional though smaller absorption maximum, the Qx band at about 567nm.n.d., not determined.
Chlorin
Absorption maxima (nm) Fluorescence maxima (nm)
Whole cells Acetone extracts Whole cells
Chl a 670–675 435, 663 680–685Chl b n.d. 455, 645 (in acetone 652)Chl d 714–718 400, 697 (in acetone 745)BChl a 375, 590, 805, 830–911 358, 579, 771 907–915BChl b 400, 605, 835–850, 986–1035 368, 407, 582, 795 1040nmBChl c 457–460, 745–755 433, 663 775BChl d 450, 715–745 425, 654 763BChl e 460–462, 710–725 459, 648 738BChl ga 375, 419, 575, 788 365, 405, 566, 762 n.d.

48 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
absorption properties of the pigments occursbecause of differences in their binding to theantenna proteins, so that the vectorial excitationcascade is thermodynamically favored (i.e., in asequence involving pigments with progressivelylonger absorption maxima). The resulting smalldifferences in the energy level of antenna pig-ments directs the transfer of excitation energymore or less to the reaction center.
A second consequence of the interactionsbetween pigment molecules and proteins is theshift of the absorption peaks of the formertowards longer wavelengths. In the case of chlo-rophyll a, the shift is comparatively small whileit is larger in bacteriochlorophyll-proteincomplexes (up to 650 nm in bacteriochlorophyllb-containing phototrophic bacteria; Table 2). Theshift for most carotenoids in association withproteins is as small as for chlorophyll a. In intactcells, carotenoids absorb mainly in the 420–550 nm wavelength region. In contrast, bindingof one type of porphyrin pigment (bacteriochlo-rophyll a) by different apoproteins has led toa considerable diversification of the long-wavelength absorption maxima in purple sulfurand nonsulfur bacteria (Fig. 7). Obviously therole of proteins in pigment-protein-complexes isnot confined to the proper coordination of pig-ment molecules but als o can represent a meansto exploit wavelength regions not utilized byother phototrophic organisms. Especially inintertidal microbial mats, variations in the finestructure of the pigment-protein complexes is ameans of ecological niche separation (see Com-petition between Phototrophic Bacteria in thisChapter). The absorption spectra of whole cellsof phototrophic bacteria seem to have evolved insuch a way that almost the entire electromag-netic spectrum suitable for electrochemical reac-tions can be exploited (Fig. 7).
The first step of porphyrin synthesis is theformation of 5-amino levulinic acid (d-ALA).In Chloroflexus aurantiacus, b- and g-Proteobacteria, cyanobacteria, Heliobacteri-aceae, and green sulfur bacteria, d-ALA is syn-thesized from glutamate (C5-pathway), whichtherefore appears to represent the more ances-tral pathway. In contrast, a-Proteobacteria aswell as yeasts, fungi, and animals form d-ALA bythe ALA synthase-mediated condensation ofglycine with succinyl-CoA (Beale, 1995; Oh-Hama, 1989; Oh-Hama et al., 1991).
All (bacterio)chlorophylls exhibit two majorabsorption bands (Table 2), leaving a consider-ably wide gap in the absorption spectrum. Thelatter is partially complemented by the absorp-tion spectrum of carotenoids found in allphototrophic bacteria or by a range of phycobil-iproteins in most cyanobacteria. Owing to thepresence of up to 15 conjugated double bonds,
carotenoids absorb light at the short wavelengthend of the visible range.
The light-harvesting antenna complexes ofgreen sulfur bacteria and Chloroflexus areextramembranous ovoid organelles, so-calledchlorosomes, which are attached to the inner sur-face of the cytoplasmic membrane and containbacteriochlorophylls c, d, or e. Chlorosomes areexceptional in that proteins do not seem to beinvolved as ligands for most of the antenna bac-teriochlorophyll molecules. Instead, interactionsbetween the bacteriochlorophylls themselvesgovern the absorptive properties of the photo-synthetic antenna in green sulfur bacteria (Blan-kenship et al., 1995; Fig. 3A). In all otherphototrophic prokaryotes studied, chlorins andcarotenoid molecules occur in complexes withproteins.
Chlorins in pigment-protein complexes arenoncovalently bound by histidine imidazole res-idues, which ligate the central magnesium atomof the porphyrin (Drews and Golecki, 1995). Insome cases (e.g., heliobacterial reaction centerprotein; Vermaas, 1994) the histidine residuesare replaced by asparagine, glutamine or argin-ine, which may function as ligands. Noncovalentbinding of carotenoids seems to be mediatedlargely by hydrophobic interactions. In the pur-ple nonsulfur bacteria, the Chromatiaceae, andEctothiorhodospiraceae, all antenna complexes(and reaction centers) are located within intrac-ytoplasmic membranes that are differentiatedfrom, but contiguous to, the cytoplasmic mem-brane of the cell. In purple nonsulfur bacteria,Chromatiaceae, and Ectothiorhodospiraceae,intracellular membranes occur as vesicles, stacks,lamellae, or tubules (Figs. 2 and 3B). Most pho-tosynthetic species of the a-Proteobacteria(Rhodocyclus purpureus, Rhodocyclus tenuis,Rubrivivax gelatinosus) do not form extensiveintracellular membrane systems. The photo-chemical apparatus of purple nonsulfur bacteriais confined to the intracellular membrane sys-tem, whereas the enzyme complexes of the res-piratory chain and transport systems are locatedin the cytoplasmic membrane (Bowyer et al.,1985). This functional differentiation does notseem to exist in purple sulfur bacteria (Allochro-matium vinosum, Ectothiorhodospira mobilis;Drews and Golecki, 1995). With one knownexception, the photosynthetic apparatus incyanobacteria is located on specialized intracel-lular membranes (thylakoids). Thylakoids maybe either single or stacked, a nd are distributedconcentrically (parallel to the cytoplasmaticmembrane), radially, or randomly (Fig. 2). Likein chloroplasts, lateral heterogeneity (spatialseparation of photosystem I in stroma lamellaeand of photosystem II in grana stacks) has beenfound in “Prochlorophytes.”

CHAPTER 1.3 The Phototrophic Way of Life 49
In Heliothrix, the Heliobacteriaceae, some pur-ple nonsulfur bacteria (e.g., Rhodocyclus tenuis;Wakim and Oelze, 1980) and one cyanobacte-rium (Gloeobacter violaceus), the photosyntheticapparatus is located in the cytoplasmicmembrane.
The light-harvesting antenna complexes ofpurple nonsulfur and purple sulfur bacteria arecomposed of two small, membrane-spanning a-and b-polypeptides to which bacteriochlorophylla or b, and carotenoids are noncovalentlybound. The polypeptide monomers aggregatewithin the membrane to form ring structures of16 (LHI) or 9 (LHII) subunits, respectively(McDermott et al., 1995; Fig. 3B). Accordingto the current structural model, the ring of 16LHI-subunits surrounds one reaction center.Several LHII-aggregates transfer energy to thissupercomplex.
In Cyanobacteria, light-harvesting chlorophylla is present in two different types of proteincomplexes. The CP43 and CP47 core-antennacomplexes are tightly associated with photosys-tem II (Barry et al., 1994). In photosystem I,however, antenna chlorophylls are an integralpart of the reaction center itself (Golbeck, 1994;Fig. 3C).
A third class of light-harvesting complexes arephycobilisomes. They occur in the divisionCyanobacteria (and in the plastids of red algaeand some other groups of eukaryotic algae), andin most species are the main light-harvestingantenna structures of these bacteria. Under theelectron microscope, phycobilisomes appear ashemidiscoidal to cylindrical particles attached tothe cytoplasmic side of the thylacoids. In Gloeo-bacter violaceus, the cytoplasmic membrane isunderlain by a continuous subcortical layercontaining the phycobilisomes. Light energyabsorbed by phycobilisomes is transferred pref-erentially to photosystem II, with chlorophyll aserving as antenna for photosystem I. However,short-term or partial spillover may occur, as thephycobilisomes are quite mobile (van Thor, J.J.,et al., 1998). While the blue and red wavelengthrange is absorbed mainly by chlorophyll; thephycobilisomes harvest the blue-green, yellow,and orange regions (450–655 nm) of the lightspectrum, thereby extending the spectral rangeof photosynthetic light-harvesting considerably(Fig. 7). The capacity of forming phycobilisomesis of selective advantage for the colonization oflow light aquatic habitats (see Competitionbetween Phototrophic Bacteria in this Chapter).Most (80%) of the phycobilisome mass is water-soluble phycobiliproteins, which contain open-chain tetrapyrrole chromophores (the phycobi-lins). Four types of phycobilins are known, theblue-colored phycocyanobilin (PCB), red-col-ored phycoerythrobilin (PEB), yellow-colored
phycourobilin (PUB), and purple-colored phy-cobiliviolin (PXB, also sometimes abbreviatedCV). They are found in various molar ratios, andform part of four recognized types of phycobil-iproteins: allophycocyanin (APC), phycocyanin(PC), phycoerythrocyanin (PEC), and phyco-erythr in (PE). In contrast to (bacterio)chloro-phylls, the chromophores are covalently boundby thioether linkages to cysteine residues of theapoproteins. Up to three chromophores may bebound to a single a- or b-polypeptide. The phy-cobiliproteins are heteromonomers forming(ab)3 trimeric disks. Together with chro-mophore-free linker polypeptides, these disksare assembled in aggregates, the phycobilisomes,which are attached to the cytoplasmic side ofphotosystem II (Fig. 3C). Peripheral rod ele-ments consisting of phycoerythrin (which har-bors PEB, and sometimes also PUB) orphycoerythrocyanin (with PCB and PXB), andphycocyanin (with PCB, and in some cases smallamounts of PEB) are arranged in a hemidiscoi-dal fashion around a core substructure consistinglargely of allophycocyanin (with PCB). The dif-ferent absorption properties of the phycobilinsare the result of differences in the number ofconjugated double-bonds (the conjugated p-electron system is shorter for PEB and PUB), inthe side chains of the tetrapyrrole prostheticgroups, including also chemically distinct chro-mophore-protein linkages, and in the proteinenvironments of the chromophores (Sidler,1994). Light energy is absorbed mainly by theperipheral rods, and transferred rapidly by radi-ation-less downhill energy transfer from phyco-erythrin (absorption maximum 495– 575 nm)or phycoerythrocyanin (575 nm) to phycocyanin(615–640 nm). Finally, allophycocyanin (650–655nm) transfers the energy to photosystem II.
Not all cyanobacteria possess all of these dif-ferent phycobiliproteins. Those synthesizingexclusively APC and PC appear blue-green.Many heterocystous cyanobacteria also producePEC in addition to APC and PC (Bryant et al.,1982); these strains never produce PE. Dark-colored strains of many benthic genera containlarge amounts of PC and PE. Red cyanobacteria,typical for deep lacustrine and marine watersproduce large amounts of PE, and only smallamounts of PC. Marine open ocean cyanobacte-ria (Synechoccus, Trichodesmium) contain largeamounts of a PUB-rich PE, with absorbancemaxima around 495–500 nm.
In Chl b-producing cyanobacteria (the former“Prochlorophytes”), the photosynthetic anten-nae are intrinsic to the membrane, and inProchlorothrix hollandica, they contain chloro-phyll a and b-carotene (PSI; photosystem I), orchlorophylls a and b, and zeaxanthin (PSII; pho-tosystem II). In contrast to the other two known

50 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
species, Prochlorococcus marinus contains divinyl-chlorophyll a and divinyl-chlorophyll b. The pres-ence of chlorophyll b and zeaxanthin and theirfunctional connection to the reaction center ofPSII enables these bacteria to absorb light in thewavelength range of 460–500 nm, and is of selec-tive advantage under light conditions present inthe lower euphotic zone of oligotrophic oceans(see Competition for Light in this Chapter). How-ever, chlorophyll b represents only a minorfraction of the photosynthetic pigments. InProchloron, the ratio of chlorophyll a/chlorophyllb is between 2.6 and 12.0 (Thorne et al., 1977);this ratio is even higher in Prochlorothrix (10–18), in which the ratio of PSI to PSII is > 3 : 1. InProchlorothrix hollandica, cells grown at low lightintensities exhibit the lowest chlorophyll a/chlo-rophyll b ratios (Matthijs et al., 1994).
A very interesting variation is exemplified byAcaryochloris marina, where Chl d is the majorantenna chlorin (2% of the dry weight, whereasChl a is only 0.1%) harvesting light for both pho-tosystems (Schiller et al., 1997). A. marina alsocontains traces of a Chl c-like pigment in addi-tion to more typically cyanobacterial carotenoids(a-carotene—found also in Prochlororococcus—and zeaxanthine—found in many cyanobacteria)and phycobiliproteins (APC and PC; Miyachi etal., 1997).
In purple bacteria, the size of the photosyn-thetic antenna is in the range of 20–200 bacteri-ochlorophyll a per reaction center (Zuber andCogdell, 1995). The specific bacteriochloro-phyll a content of aerobic bacteriochlorophyll-containing bacteria reaches only 5–10% of thatof anoxygenic phototrophic bacteria (Yurkov andBeatty, 1998). At least in one strain (RhizobiumBTAi1), the size of the photosynthetic unit issimilar to that of anoxygenic phototrophic bac-teria (Evans et al., 1990), indicating that the lowpigment content is due to a low number of reac-tion centers. In PSII of cyanobacteria, theantenna comprises 300–800 phycobilin chro-mophores and 47 chlorophyll a molecules (Sidler,1994; Matthijs et al., 1994 ), whereas the reactioncenter protein PsaA of PSI binds 110 chlorophylla molecules (Golbeck, 1994). The photosyntheticantenna of green sulfur bacteria is significantlylarger than that of other anoxygenic phototrophsand comprises about 1000 bacteriochlorophyllmolecules connected to one reaction center (seeThe Family Chlorobiaceae, Physiology section inVolume 7). This appears to be one major reasonfor the competitive success of green sulfur bac-teria in low-light environments (see Competitionfor Light in this Chapter). Antenna size is smallerin Chloroflexus (Olsen, 1998). About 35 mole-cules of bacteriochlorophyll g are associated withone reaction center in Heliobacteriaceae (Amesz,1995).
Efficiency of Light Harvesting
The light absorption capabilities of photosyn-thetic prokaryotes can be judged best by calcu-lating which fraction f of the light impinging ona single cell is actually absorbed. This fraction isconsiderable for purple sulfur and other bacte-ria. The highest bacteriochlorophyll-specificattenuation coefficient kB has been determinedfor a population of Amoebobacter purpureus(0.050 m2·(mg BChl a)-1; Overmann et al.,1991a). For comparison Prochlorococcus has achlorophyll-specific attenuation coefficient of0.0147–0.0232 m2·(mg Chl a-1 (Moore et al.,1998). For Amoebobacter, f is 0.36, or 36%, ascalculated from Beer’s Law and using the valueof kB, the intracellular c oncentration of light-harvesting pigments C (10.3 ¥ 106 mg BChl·m-3,calculated from a content of 85 mg BChl·(mgprotein)-1; van Gemerden and Mas, 1995;Watson et al., 1977) and the average optical path-length d of a cell (2 mm):
f = 100 ¥ exp(-kB ¥ C ¥ d)
Of the photosynthetic pigments that absorb thishigh fraction of incident light, the majority (typ-ically >97%) serves in light-harvesting and trans-fers excitation energy to the photochemicalreaction centers. The combination of antennacomplexes with one reaction center constitutesthe photosynthetic unit. The efficiency of energytransfer within the photosynthetic unit and itssize determine the fraction of the quantum fluxthat is harvested.
Large concentrations of pigments result inself-shading and thus a reduced efficiency of lightabsorption per mole of pigment. At the cell sizeand intracellular pigment concentrations typicalof most prokaryotic phototrophs, this decrease inefficiency is not very important (Garcia-Pichel,1994a), but it might be significant in someextremely low-light adapted anoxygenic pho-totrophs like the green sulfur bacterial strain iso-lated from the Black Sea chemocline (Overmannet al., 1991a).
Close proximity of photosynthetic pigmentsenables an efficient transfer of excitation energybut at the same time also causes a so-called“package effect” (Kirk, 1983) by which self-shading of the pigment molecules exceeds thatpredicted by the Lambert-Beer law. The packageeffect is seen clearly in a flattening of absorptionpeaks, commonly observed when recordingabsorption spectra of whole cells (see The FamilyChlorobiaceae, Identification section in Volume7). Because the energy requirement for biosyn-thesis of additional antenna structures is ratherconstant, the net energy gain for a photosyn-thetic cell must decrease at higher intracellularpigment concentrations, which restricts the

CHAPTER 1.3 The Phototrophic Way of Life 51
amount of light-harvesting structures a photo-synthetic cell can synthesize. Polypeptides of thephotosynthetic machinery (a significant fractionof the total cell protein) amount to 20% in pur-ple nonsulfur bacteria and >50% in phycobilip-rotein-containing cyanobacteria. Interestingly,the total protein content of cyanobacterial cellsis comparable to other phototrophic bacteria.Possibly, cyanobacteria contain reduced levels ofproteins involved in nonphotosynthetic pro-cesses to compensate for the high energy andnitrogen expenditure of the antenna proteins.
The biosynthesis of proteins requires a majorfraction of the energy expenditure of the bacte-rial cell (Gottschalk, 1986). In chlorosomes, themass ratio of protein:bacteriochlorophyll is sig-nificantly lower than in other light-harvestingcomplexes (Table 3). Probably this is one majorreason for the larger antenna size and the lowerlight energy requirements of green sulfur bacte-ria as compared to their purple and cyanobacte-rial counterparts (see Competition betweenPhototrophic Bacteria in this Chapter), andmight help explain the competitive advantagegained by Prochlorocococcus over their closerelatives Synechococcus in the open oceans.
Conversion of Light into Chemical EnergyPRINCIPLE The unifying principle of bacterialand archaeal photosynthesis is the light-drivengeneration of a proton-motive force (PMF). ThePMF is subsequently used by ATP synthase toform ATP, or for active transport and motility.
In chlorophyll-based photosynthesis, redoxreactions and charge separation precede theestablishment of the PMF. In addition, reducing
power (NAD(P)H + H+) is generated as a pri-mary product of the light reaction in Cyanobac-teria. In the photochemical reaction, only theenergy of the lowest excited singlet state (seeLight Absorption and Light Energy Transfer inProkaryotes in this Chapter) of the chlorophyllsis used. Consequently, all absorbed light quantahave the same effect irrespective of their originalenergy (wavelength). When comparing the lightenergy available in different habitats, or the lightadaptation of different phototrophic bacteria, itis therefore more meaningful to express irradi-ances in units of mol quanta·m-2·s-1 rather thanW·m-2 (see Competition for Light in thisChapter).
The standard free energy for the reduction ofCO2 depends on the redox potential of the pho-tosynthetic electron donor employed (Table 4,Fig. 8). If this energy requirement for electrontransfer is compared with the energy availableafter absorption of photons of different wave-lengths, it becomes clear that oxygenic photosyn-thesis is not feasible in photosystems containingthe known types of chlorin pigments, andrequires the absorption of two photons per elec-tron (Fig. 8).
The biological conversion of light into chemi-cal energy has been found to be remarkably effi-cient: the number of charge separation eventsper absorbed photon is 1.0 (Kok, 1973; Wraightand Clayton, 1973) and the efficiency of theentire photoconversion process of a red photonto chemical energy by oxygenic photosyntheticorganisms is 43% (Golbeck, 1994). Whereas theefficiency of energy transfer between antennabacteriochlorophyll and the reaction center inmost cases is close to 100% (Amesz, 1995), thetransfer between antenna carotenoids and thereaction center can be significantly lower, 70%in Heliobacteriaceae (Amesz, 1995) and even
Table 3. Pigment:protein ratio in different photosyntheticantenna complexes.
aChloroflexus aurantiacus.Data from Olson, 1998 or calculated from Sidler, 1994, Loachand Parkes-Loach, 1995, Zuber and Cogdell, 1995. Caro-tenoids have been neglected in these calculations because oftheir lower numbers as compared to bacteriochlorophylls(B800-850 LHII), their absence in phycobilisomes, and thecontroversy concerning their functional significance in light-harvesting (chlorosomes). Only antenna complexes whichare separate entities from reaction centers were considered.Photosystem I does not contain a distinct antenna structure;the PsaA protein of the reaction center binds 110 chlorophylla molecules.
Antenna complex type
Protein:pigment
Mass ratioPer pigment
molecule (in Da)
Chlorosomes 0.5–2.2 420–1,840B806-866 complexa 3.9–5.8 3,550–5,290B800-850 LHII 4.4 4,000B820 LHI 6.7 6,100Phycobilisomes ~22.4 ~12,300
Table 4. Standard redox potentials of different electrondonors of the photosynthetic light reaction.a
aTaken from Brune, 1989; Widdel et al., 1993; Thauer et al.,1977; Zehnder and Stumm, 1988.
Electron donor Eo-[mV]
1/2O2/H2O +820Fe(OH)3 + HCO3
-/FeCO3 +200Fumarate/Succinate +33HSO3/S0- -38SO4
2-/S0 -200SO4
2-/HS- -218Fe(OH)3/Fe2- -236S0/HS- -278HCO3
-/acetate -350S2O3
2-/HS- + HSO3- -402
H+/1/2H2 -414
Electron acceptor Eo-[mV]
CO2/<CH2O> -434
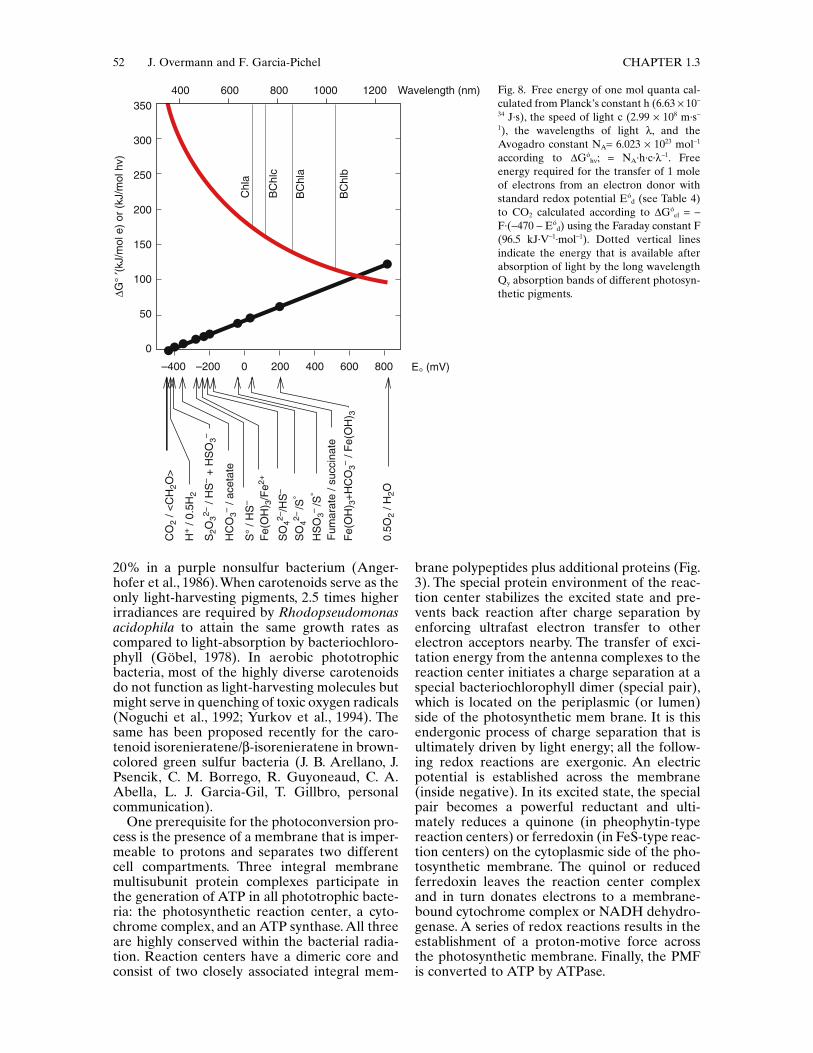
52 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
20% in a purple nonsulfur bacterium (Anger-hofer et al., 1986). When carotenoids serve as theonly light-harvesting pigments, 2.5 times higherirradiances are required by Rhodopseudomonasacidophila to attain the same growth rates ascompared to light-absorption by bacteriochloro-phyll (Göbel, 1978). In aerobic phototrophicbacteria, most of the highly diverse carotenoidsdo not function as light-harvesting molecules butmight serve in quenching of toxic oxygen radicals(Noguchi et al., 1992; Yurkov et al., 1994). Thesame has been proposed recently for the caro-tenoid isorenieratene/b-isorenieratene in brown-colored green sulfur bacteria (J. B. Arellano, J.Psencik, C. M. Borrego, R. Guyoneaud, C. A.Abella, L. J. Garcia-Gil, T. Gillbro, personalcommunication).
One prerequisite for the photoconversion pro-cess is the presence of a membrane that is imper-meable to protons and separates two differentcell compartments. Three integral membranemultisubunit protein complexes participate inthe generation of ATP in all phototrophic bacte-ria: the photosynthetic reaction center, a cyto-chrome complex, and an ATP synthase. All threeare highly conserved within the bacterial radia-tion. Reaction centers have a dimeric core andconsist of two closely associated integral mem-
brane polypeptides plus additional proteins (Fig.3). The special protein environment of the reac-tion center stabilizes the excited state and pre-vents back reaction after charge separation byenforcing ultrafast electron transfer to otherelectron acceptors nearby. The transfer of exci-tation energy from the antenna complexes to thereaction center initiates a charge separation at aspecial bacteriochlorophyll dimer (special pair),which is located on the periplasmic (or lumen)side of the photosynthetic mem brane. It is thisendergonic process of charge separation that isultimately driven by light energy; all the follow-ing redox reactions are exergonic. An electricpotential is established across the membrane(inside negative). In its excited state, the specialpair becomes a powerful reductant and ulti-mately reduces a quinone (in pheophytin-typereaction centers) or ferredoxin (in FeS-type reac-tion centers) on the cytoplasmic side of the pho-tosynthetic membrane. The quinol or reducedferredoxin leaves the reaction center complexand in turn donates electrons to a membrane-bound cytochrome complex or NADH dehydro-genase. A series of redox reactions results in theestablishment of a proton-motive force acrossthe photosynthetic membrane. Finally, the PMFis converted to ATP by ATPase.
Fig. 8. Free energy of one mol quanta cal-culated from Planck’s constant h (6.63 ¥ 10-
34 J·s), the speed of light c (2.99 ¥ 108 m·s-
1), the wavelengths of light l, and theAvogadro constant NA= 6.023 ¥ 1023 mol-1
according to DGóhn; = NA·h·c·l-1. Free
energy required for the transfer of 1 moleof electrons from an electron donor withstandard redox potential Eó
d (see Table 4)to CO2 calculated according to DGó
el = -F·(-470 - Eó
d) using the Faraday constant F(96.5 kJ·V-1·mol-1). Dotted vertical linesindicate the energy that is available afterabsorption of light by the long wavelengthQy absorption bands of different photosyn-thetic pigments.
350
300
250
200
150
100
50
0
400 600 800 1000 1200 Wavelength (nm)
!G°
"(kJ/
mol
e)
or (
kJ/m
ol h
v)
–400 –200 0 200 400 600 800 E° (mV)
Chl
a
BC
hlc
BC
hla
BC
hlb
CO
2 / <
CH
2O>
HC
O3–
/ ace
tate
S2O
32– /
HS
– +
HS
O3–
S°
/ HS
–
Fe(
OH
) 3/F
e2+
Fe(
OH
) 3+H
CO
3– / F
e(O
H) 3
0.5O
2 / H
2O
SO
42–/H
S–
SO
42–/S
°
HS
O3–
/S°
Fum
arat
e / s
ucci
nate
H+
/ 0.5
H2

CHAPTER 1.3 The Phototrophic Way of Life 53
In contrast to the (bacterio)chlorophyll-basedsystems of bacteria, light energy conversion ofHalobacteria does not involve redox reactionsand is limited to a vectorial transport of protonsby bacteriorhodopsin. Upon excitation by light,the prosthetic retinal undergoes a series ofreversible photochemical transformations (anisomerization from the all-trans to the 13-cisform) and releases a proton into the extracellularspace. The PMF thus generated is used for ATPsynthesis by ATPase. Due to its low solubility, O2
in the concentrated salt solution is present insignificantly lower amount than in freshwater.Rhodopsin-mediated formation of ATP maybecome the sole source of energy for growthunder anaerobic conditions in the light (Oester-helt and Krippahl, 1983) and has therefore beenviewed as an adaptation to the natural brine hab-itat of Halobacteria. Because of its distinctmechanism, archaeal “photosynthesis” is not dis-cussed in further detail in the present section.Additional information can be found in chapterstitled Introduction to the Classification ofArchaea and The Family Halobacteriaceae.
Molecular Architecture of the Reaction CenterAll bacteria which perform anoxygenic photo-synthesis possess—or (in the case of cyanobacte-ria which are capable of using sulfide as electrondonor) employ—only a single photosystem. Thedecrease in redox potential that a single photo-system can undergo upon excitation appears tobe limited (Blankenship, 1992, compare Fig. 8).A combination of two different photosystems isrequired for the thermodynamically unfavorableutilization of water as an electron donor for pho-tosynthesis (Fig. 3C). With the relatively simplearchitecture of their photosystems, all anoxy-genic phototrophic bacteria depend on electrondonors that exhibit standard redox potentialsmore negative than water (e.g., H2S, H2, acetate;Table 4). This molecular feature is one majorreason for the narrow ecological niche ofanoxygenic phototrophic bacteria in extantecosystems (see Habitats of PhototrophicProkaryotes in this Chapter).
Two different types of reaction centers occurin photosynthetic bacteria. Based on the chemi-cal nature of the early electron acceptors, apheophytin/quinone-type reaction center and aFeS-type reaction center are distinguished(Blankenship, 1992; Fig. 3A,B). The first type isfound in green gliding Chloroflexus species,phototrophic members of the a- andb-Proteobacteria, Chromatiaceae, Ectothiorho-dospiraceae, and in PSII of Cyanobacteria. Thereaction center of Proteobacteria consists ofthree protein subunits (L, M, H) which bind four
bacteriochlorophylls, two bacteriopheophytins,two quinones and one high-spin nonheme Fe2+
(Lancaster and Michel, 1996; Fig. 3B). Many spe-cies (e.g., Chloroflexus aurantiacus, Blastochlorisviridis and Allochromatium vinosum) contain anadditional tetraheme cytochrome c polypeptideattached to the periplasmic side of the reactioncenter.
Following the transfer of the electrons byubiquinol or plastoquinol, the redox reactions atthe cytochrome bc1 (or b6f) complex drive protontransport across the cytoplasmic membrane. Pro-tons are translocated either into the extracellularspace (anoxygenic phototrophic bacteria) or theintrathylacoidal space (cyanobacteria). The ratioof protons translocated to electrons transferred(H+/e- ratio) is 2. The reaction center and cyto-chrome bc1 in pheophytin-type reaction centersof Proteobacteria and Chloroflexus are function-ally linked by two diffusible electron carriers,ubiquinone in the hydrophobic domain of themembrane and cytochrome c2 or auracyanin(Meyer and Donohue, 1995) in the periplasmicspace. The liberated electron is transferred backto the special pair via quinone, the cytochromebc1 complex and soluble periplasmic solubleelectron carrie r (often cytochrome c2). Owing tothis cyclic electron transport, the only primaryproduct of photosynthesis is the proton-motiveforce, and the reduced pyridine nucleotiderequired for photosynthetic CO2 fixation is gen-erated by energy-dependent reverse electronflow (Fig. 3).
In oxygenic phototrophic bacteria, plasto-quinone is the electron acceptor of PSII anddonates electrons to the cytochrome b6f-complex. The special pair is reduced by themanganese-containing water-splitting systemlocated at the lumenal side of the transmem-brane PSII complex (Fig. 3C).
In the pheophytin-type reaction centers of aer-obic phototrophic bacteria, photoinduced chargeseparation occurs only in the presence of O2
(Okamura et al., 1985). It has been proposed(Yurkov and Beatty, 1998) that oxic conditionsare required for photochemical activity becausethe primary acceptor ubiquinone has a signi-ficantly higher midpoint redox potential thanin anoxygenic photosynthetic bacteria (65 to120 mV more positive). The primary acceptortherefore may stay in its oxidized, electron-accepting state only in the presence of O2.
The second type of reaction center containsiron-sulfur clusters as early electron acceptorsand occurs in green sulfur bacteria (Fig. 3A),Heliobacteriaceae, and in the photosystem I ofCyanobacteria. Functionally, the reaction centersof green sulfur bacteria, Heliobacteriaceae, andPSI of cyanobacteria are therefore similar. How-ever, the former two are homodimeric and only

54 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
one reaction center gene has been detected,whereas the reaction center of PSI of cyanobac-teria and green plants contains two nonidentical,but similar, subunits (PS I-A and PS I-B; Ver-maas, 1994). In FeS-type reaction centers, theredox potential of the special pair in its reducedstate (P*) is sufficiently low to permit a transferof electrons to ferredoxin. Until recently, it hastherefore been assumed that noncyclic electronflow can directly reduce NAD(P)+ and does notrequire further energy expenditure not only incyanobacteria but also in green sulfur bacteria.However, the sequencing of the whole genomeof Chlorobium tepidum has not provided anyindications for the presence of a ferredoxin-NADP+ oxidoreductase (D. A. Bryant, personalcommunication).
Electron DonorsAnoxygenic phototrophic bacteria of the a- andb-Proteobacteria use a wide variety of reducedorganic carbon compounds as electron-donatingsubstrates (see Carbon Metabolism in this Chap-ter; Table 4; Fig. 8). Most phototrophic sulfurbacteria are capable of using sulfide as photosyn-thetic electron donor. Other inorganic electrondonors utilized include H2, polysulfides, elemen-tal sulfur, thiosulfate, sulfite, and iron (Widdel etal., 1993). Sulfide is oxidized to zero-valent sul-fur, which in Chromatiaceae appears to be depos-ited as polysulfides or polythionates rather thanin the form of S8 rings (Steudel, 1989; Steudel etal., 1990). In addition, thiosulfate is formed as anoxidation product by some species (see The Fam-ily Chlorobiaceae in Volume 7; Steudel et al.,1990). The photosynthetic sulfide oxidation ratesof purple sulfur bacteria are higher than requiredfor growth and remains constant at all growthrates. As a result, storage of sulfur is at maximumat low growth rates (van Gemerden and Mas,1995). Zero-valent sulfur is further oxidized tosulfate. In microbial mats, polysulfides andorganic sulfur compounds may be significant asphotosynthetic electron donor. Polysulfide oxi-dation has been reported for Chlorobium limi-cola f.sp. thiosulfatophilum, Allochromatiumvinosum, Thiocapsa roseopersicina, while dime-thylsulfide is utilized and oxidized to dimethyl-sulfoxide by the two purple sulfur bacteriaThiocystis sp. and Thiocapsa roseopersicina (vanGemerden and Mas, 1995). In addition toreduced sulfur compounds, hydrogen serves aselectron donor in the majority of green sulfurbacteria, and in the metabolically more versatilespecies of purple sulfur bacteria (such as Allo-chromatium vinosum, Thiocapsa roseopersicina).In green sulfur bacteria which lack assimilatorysulfate reduction, a reduced sulfur source isrequired during growth with molecular hydro-
gen. Finally, a few species of purple nonsulfurbacteria, of Chromatiaceae, and of the green sul-fur bacteria have been found to utilize ferrousiron as photosynthetic electron donor (Widdel etal., 1993; Heising et al., 1999).
Sulfide acts as a strong poison of PSII activityin many algae and cyanobacteria. The ability ofsome Cyanobacteria to conduct anoxygenic pho-tosynthesis with sulfide as an electron donor toPSI (Cohen et al., 1975; Padan, 1979; Padan andCohen, 1982), or to continue oxygenic photosyn-thesis in the presence of sulfide (Cohen et al.,1986), may be one of the key traits that extendthe habitat of sulfide-utilizing cyanobacteria intothe temporarily anoxic, sulfide-containing, layersof hot springs (Castenholz and Utkilen, 1984),marine microbial mats (De Wit and van Gemer-den, 1987a; De Wit et al., 1988), and thechemoclines of meromictic lakes (Jørgensen etal., 1979; Camacho et al., 1996). Sulfide is aninhibitor of PSII and induces the synthesis of asulfide-oxidizing enzyme system. In contrast tophototrophic sulfur bacteria, cyanobacteria oxi-dize sulfide to elemental sulfur or thiosulfate butdo not form sulfate (De Wit and van Gemerden,1987b). However, the use of sulfide by cyanobac-teria in anoxygenic photosynthesis must beregarded as a detoxification mechanism, sincetheir low affinity for sulfide (De Wit and vanGemerden, 1987b; Garcia-Pichel and Casten-holz, 1990) renders them unable to compete withpurple or green sulfur bacteria for sulfide as anelectron donor.
In the natural habitat, growth of phototrophicsulfur bacteria is limited mainly by light and sul-fide. Sulfide often becomes the growth-limitingfactor at the top of the phototrophic sulfur bac-terial layers where light intensities are highest,while sulfide has to diffuse through the remain-der of the community. The affinity for sulfideduring photolithotrophic growth varies betweenthe different groups of anoxygenic phototrophs(including cyanobacteria growing with sulfide)and has been shown to be of selective value dur-ing competition experiments. Green sulfurbacteria and Ectothiorhodospiraceae exhibit 5 to7 times higher affinities for sulfide than Chro-matiaceae (van Gemerden and Mas, 1995). Onthe contrary, affinities for polysulfides are com-parable between green sulfur bacteria andChromatiaceae.
Efficiency of Growth and Maintenance Energy Requirements
For any photochemical reaction, the quantumyield is defined as the number of molecules con-verted per light quantum absorbed. The quantumefficiency is the ratio of energy stored in a com-

CHAPTER 1.3 The Phototrophic Way of Life 55
pound, to the radiant energy absorbed for itsformation. The quantum requirement is the recip-rocal of the quantum yield. For CO2 fixation ofpurple sulfur bacteria, a quantum requirement of8 and 10.5 mol quanta· (mol CO2)-1 is theoreti-cally expected (Brune, 1989), considering thatreverse electron transport is necessary. Experi-mentally, a quantum requirement of 12 ± 1.5 and11.7 mol quanta·(mol CO2
-1 was determined,which corresponds to a quantum yield of 0.083(Wassink et al., 1942 in Brune, 1989; Göbel, 1978).
In contrast, calculated values for the quantumrequirements of green sulfur bacteria lie between3.5 and 4.5 mol quanta·(mol CO2)-1, if noncyclicelectron transport is assumed. However, earliermeasurements had yielded much higher values(9–10; Brune, 1989). This discrepancy may beexplained by the very recent finding that a genefor ferredoxin-NADP+ oxidoreductase does notseem to be present in the genome of Chlorobiumtepidum (D. A. Bryant, personal communica-tion), which makes noncyclic electron transportrather unlikely also for green sulfur bacteria.
The quantum yield for CO2-fixation deter-mined for Prochlorococcus isolates incubated indaylight spectrum fluorescent light was between0.086 and 0.128 mol C·(mol quanta)–1 (Moore etal., 1998), thus reaching Emerson’s theoreticalmaximum for O2 evolution in oxygenic photo-synthesis. In cyanobacteria, typically thriving inoxic environments where only oxidized sourcesof nitrogen and sulfur are available, a large pro-portion of the reducing power generated in thelight reactions must be diverted to assimilatorynitrate or sulfate reduction, or to nitrogen fixa-tion, so that the quantum requirement for CO2
fixation can be substantially lower than that foroxygen evolution.
In a careful study of Rhodobacter capsulatusand Rba. acidophilus grown with lactate as elec-tron donor in a light chemostat, a value for themaintenance light energy requirement of mq =0.012 mol quanta·(g dry weight·h)–1 was deter-mined (Göbel, 1978). The maintenance energyrequirements of green sulfur bacteria are signif-icantly lower compared to their purple conter-parts (van Gemerden and Mas, 1995). This maybe explained by the fact that protein turnover ishighly energy demanding and that the proteincontent of the green sulfur bacterial antennais much lower than in purple sulfur bacteria(Table 3).
Response to Changes in Light Intensity and Quality
Phototrophic bacteria acclimate to changes inlight intensity and quality by diverse mecha-nisms. Anoxygenic phototrophic bacteria as well
as cyanobacteria respond to a step-down in irra-diance by increasing the specific pigment contentand vice versa (references compiled in Sánchezet al., 1998). These changes can be accomplishedeither by varying the number of photosyntheticunits per cell, the size of the individual photosyn-thetic unit, or both (see Long-term Adaptationsto Changes in Light Intensity in this Chapter).Besides long-term biochemical changes in thecomposition and the amount of light-harvestingcomplexes, short-term redistribution of antennacapabilities (see State Transitions in this Chap-ter) occur in oxygenic phototrophs.
Many species use vertical migration, mediatedby tactic responses (see Movement by Flagella inthis Chapter) and formation of gas vesicles toregulate their vertical position and exposure tolight. Especially in the stably stratified pelagichabitats of phototrophic sulfur bacteria, the dif-ference in buoyant density from the surroundingwater would cause a sedimentation of bacterialcells out of the photic zone and towards the lakebottom. The minimum buoyant density, whichhas been determined for phototrophic cellsdevoid of gas vesicles, was 1010 kg·m–3 (Over-mann et al., 1991b). Actively growing cells, whichcontain storage carbohydrate and—in the case ofChromatiaceae—elemental sulfur, can easilyattain much higher buoyant densities (up to 1046kg·m–3; Overmann and Pfennig, 1992). By com-parison, freshwater has a considerably lowerdensity (e.g., 996 kg·m-3; Overmann et al., 1999c).As a consequence, sedimentation losses are sig-nificant for natural populations of several speciesof phototrophic sulfur bacteria (Mas et al., 1990).Phototrophic bacteria have developed two waysto adjust their vertical position along gradientsof light intensity and spectral composition. Forpurple sulfur bacteria, motility in response tochanges in irradiance is known to be of ecologi-cal significance in both planktonic and benthicsituations. In benthic and terrestrial cyanobacte-ria, vertical locomotion by gliding is common.Planktonic cyanobacteria inhabiting stratifiedwaters perform vertical migrations by changingtheir cellular gas vesicle content and ballast mass(intracellular carbohydrates and protein) andhence their buoyant density. Planktonic anoxy-genic phototrophic bacteria do not seem to per-form vertical migrations mediated by changes ingas vesicle content but rather use these cellorganelles t o maintain their position within thechemocline (Overmann et al., 1991b; Overmannet al., 1994; Parkin and Brock, 1981).
Long-Term Adaptations to Changes in Light IntensityIn those photosynthetic bacteria in which theentire photosynthetic apparatus is confined to

56 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
the membrane, light absorption often isincreased by formation of intracellular mem-brane systems (Fig. 2). In Rhodobacter capsula-tus, the number of intracellular membranevesicles increases by a factor of 6.3 when the cellsare shifted from high to low light intensities. Asa result, the area of intracellular membranesunder these conditions is 2.7-fold larger than thearea of the whole cytoplasmic membrane. Pho-tosynthetic species of the b-Proteobacteria whichdo not form extensive intracellular membranesystems (Rhodocyclus purpureus, Rhodocyclustenuis, Rubrivivax gelatinosus) increase the den-sity of photosynthetic units in their cytoplasmicmembrane (Drews and Golecki, 1995). Intracel-lular membranes appear to be absent in Helio-bacteriaceae and Heliothrix, where pigments areconfined to the cytoplasmic membrane (Fig. 2).In Chloroflexus aurantiacus, the increase in cel-lular concentrations of bacteriochlorophylls aand c is mediated by an increase in the numberand volume of chlorosomes, and the percentageof cell membrane surface covered by chlo-rosomes (Golecki and Oelze, 1987). In a similarmanner, green sulfur bacteria adapt to low lightintensities by increasing the size and the cellularnumber of chlorosomes (see The Family Chloro-biaceae, Physiology section in Volume 7).
During induction of the photosynthesis appa-ratus in Proteobacteria, invaginations of the cyto-plasmic membrane, increases in the number andsize of the photosynthetic units, and bacterio-chlorophyll synthesis occur simultaneously.Under anoxic conditions, the amount of pigmentsynthesized by anoxygenic phototrophic bacteriais inversely related to the available light intensityand varies by a factor of up to 6.6 (Göbel, 1978).After a shift to low light intensity, the ratio oflight-harvesting complex I per reaction centerremains constant (at about 30 bacteriochloro-phylls per reaction center), whereas the relativeamount of the peripheral light-harvesting com-plex II increases. As a result, the size of the pho-tosynthetic unit changes by a factor of two tofive. Conversely, the specific NADH dehydroge-nase activity decreases as does the amount ofcytochrome and ubiquinone per reaction center.In Rba. capsulatus and Rba. spheroides thesechanges take about 2–3 generations and thegrowth rate is lowered during adaptation due toenergy limitation. In the purple sulfur bacteriumAllochromatium vinosum, low-light adaptation isalso accomplished by increasing the size of thephotosynthetic unit (Sánchez et al., 1998).Species like Rhodospirillum rubrum and Blas-tochloris viridis, which harbor only one type oflight-harvesting complex, increase the number ofphotosynthetic units (Drews and Golecki, 1995).
Similar to anoxygenic phototrophic bacteria,changes in both the number and the size of the
photosynthetic unit have also been decribed forcyanobacteria. In marine Synechococcus strains,the cellular content of the light-harvesting phy-coerythrin can be varied by a factor of 20 anddecreases with increasing light intensity. Inmarine benthic Microcoleus chthonoplastes, anincrease in the content of total phycobilines anda change in the ratio of PEC to PC occurs withdecreasing light intensity. The latter increase theratio of phycocyanin to chlorophyll a duringlow-light adaptation (Foy and Gibson, 1982; Postet al., 1985). Acclimation to very low light inten-sities usually involves an increase in the size ofthe photosynthetic unit, such as in metalimneticOscillatoria (Leptolyngbya) redekei and Oscilla-toria agha rdii. Changes in both the number andthe size of the photosynthetic units seem tooccur in Microcystis (Zevenboom and Mur,1984).
Adaptations to Low Light IntensitiesThe capability to adapt to low light intensitiesrepresents a competitive advantage for pho-totrophic organisms. An estimate of the mini-mum irradiance Imin required for survival ofphototrophic cells in the environment can be cal-culated from a few physiological parameters,namely the pigment content of the cells, P (in mgbacteriochlorophyll·g C–1); the maintenanceenergy requirement, mq (in mol quanta·g C–1·s-1);the (bacterio)chlorophyll-specific attenuationcoefficient, k (in m2·mg BChl a–1); the cellular dryweight content, D (in g C·m-3); and the meanoptical pathlength of one cell d:
Imin = mq · D · d /[1 – exp(–k · D · P · d)]
Employing the appropiate values for mq (seeEfficiency of growth and maintenance energyrequirements), k and P (see Light Energy andthe Spectral Distribution of Radiation in thisChapter), D (1.21·105 g C·m-3; Watson et al.,1977) and d (0.5 m for the smaller anoxygenicphototrophs), this yields a minimum irradiance(Imin) of 2 mmol quanta·m–2·s–1. In many naturalhabitats of anoxygenic phototrophic bacteria,irradiances of this order of magnitude or lowerhave been measured. Prochlorococcus has beenfound at deep water layers down to 300 m.However, these bacteria do not grow at lightintensities below 3.5 mol quanta·m–2·s-1 (Mooreet al., 1998) and thus appear to be less low-light adapted than the green sulfur bacterialstrain MN1 isolated from the Black Sea whichgrows a t light intensities as low as 0.25 mmolquanta·m–2·s-1 (Overmann et al., 1991a). Lowerirradiances could be used by phototrophicprokaryotes after a decrease of mq or anincrease of P or both. Both adaptations arepresent in strain MN1 (Overmann et al., 1991a).

CHAPTER 1.3 The Phototrophic Way of Life 57
Adaptations to High Light IntensitiesSessile cyanobacteria living on the surface ofbenthic microbial mats are typically adapted tovery high light conditions and contain largeamounts of sunscreen pigments. For oxygenicphototrophs, special adaptations to oxygen-dependent photoinhibition of photosynthesis areof particular relevance. The protein D1 of PSII,coded by the psbA gene, has been identified asthe central target of photoinhibition at high lightintensities. In Synechococcus PCC 7942, psbAcontains actually a multigene family coding forthree different forms of the protein D1, whichare differentially expressed according to the lightconditions. Analysis of mutants showed that theisoforms expressed under high light conditionsallow for optimal performance of PSII underphotoinhibitory conditions (Golden, 1994). Inaddition, carotenoids probably play a centralrole in avoiding oxygen-mediated pho tosensi-tized bleaching of photosynthetic pigments andphotooxidation of fatty acids under high lightconditions.They function as antioxidant quench-ers of excited molecules (such as triplet statechlorins and singlet oxygen) in many organismsand perhaps also as inhibitors of free-radicalreactions (Britton, 1995). The photoprotectivexanthophyll cycle typical of green algae andhigher plants is not present in cyanobacteria, butjudging from its increased specific content athigh light intensity, zeaxanthin seems to play animportant photoprotective role in some strains(Kana et al., 1988; Masamoto and Furukawa,1997; Millie et al., 1990). Glycosylated myxoxan-thophylls seem to attain the same role in others(Nonnengießer et al., 1996; Garcia-Pichel et al.,1998; Ehling-Schulz et al., 1997). Because thereis a considerable photooxidation of carotenoidsthemselves at high light intensities, the mainte-nance of high carotenoid contents requires anincreased expression of their biosyntheticgenes.
Chromatic AdaptationSeveral species of cyanobacteria are capable ofchanging the amount of peripheral phycoeryth-rin in response to changes in the spectral compo-sition of light. During growth in white or greenlight, red-pigmented PE hexamers are addedto the peripheral rods whereas additional blue-pigmented PC is added under red light (Sidler,1994). This complementary chromatic adapta-tion is found only in strains capable of formingPE, but not in those forming PEC. The comple-mentary change in antenna pigment compositionoptimizes the light-harvesting capabilities ofpopulations of Oscillatoria spp., which thrive indeeper layers of stratified lakes where light is
predominantly in the blue-green to green wave-length range (Utkilen et al., 1985; Fig. 6).
Genetic Regulation in Response to LightThe synthesis of the photosystem is especiallyenergy consuming because of the high amount oflight-harvesting and reaction center proteinpresent in phototrophically grown cells ofphototrophic Proteobacteria (20% in purplenonsulfur bacteria). The maintenance energyrequirements seem to be increased in low-lightadapted cells (Sánchez et al., 1998). An effectiveregulation of photosynthesis gene expressiontherefore would prevent futile synthesis of cellu-lar proteins. The synthesis of the photosystem inanoxygenic phototrophic bacteria is under thecontrol of a complex regulatory network (Bauerand Bird, 1996).
The expression of light-harvesting complex Iand reaction center genes is controlled 1) by thelinkage of genes in superoperons, 2) at the levelof transcription initiation, and 3) posttranscrip-tionally by the decay rate of mRNA (Bauer,1995).
In Rhodobacter capsulatus, the genes codingthe structural, biosynthetic and regulatory pro-teins for light-harvesting I and reaction centercomplexes are found assembled in a 46 kb-longphotosynthetic gene cluster (Alberti et al., 1995).The arrangement of the genes within the clusterseems to be conserved among different pho-totrophic species of the a-Proteobacteria, likeRhodobacter sphaeroides, Rhodocista centenariaand Rhodospirillum rubrum (Bauer et al., 1993).Only the pucBA operon which codes for struc-tural a- and b-polypeptides of light-harvestingcomplex II is found in a distant location onthe bacterial chromosome (about 18 kb of thepuhA in Rhodobacter capsulatus; Suwanto andKaplan, 1989).
In anoxygenic phototrophic bacteria, tran-scription of the photosynthesis genes occurs onlyunder anoxic conditions. Different photosynthe-sis genes exhibit varying levels of expression anddegrees of regulation (Bauer and Bird, 1996).The pufA,B,L,M genes (coding for the a- and b-polypeptide of the light-harvesting complex Iand the reaction center L and M structuralpolypeptides) as well as puhA (coding for thestructural polypeptide subunit H) are tightlycoregulated, transcribed at a high rate underanoxic conditions and strongly regulated (15- to30-fold). An inverted repeat sequence locatedbetween pufA and pufL affects the longevity ofthe respective mRNA primary transcript. Areduction of light leads to an activation of pufand puh gene expression by the hvrA gene prod-uct, which probably directly interacts with thetwo promoter regions. Light of 450 nm exhibits

58 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
the most severe repressing effect, indicating thata flavin-binding protein (possibly HvrA itself) isthe photoreceptor. Notably in aerobic photo-trophic bacteria, a blue light sensitive systemseems to regulate biosynthesis of bacteriochloro-phyll a (Shimada, 1995).
The intracellular bacteriochlorophyll concen-trations appear to affect puf and puc geneexpression not only at the transcriptional butalso the posttranscriptional level in Rhodobactercapsulatus (Rödig et al., 1999). The polycistronicorganization allows the coordinate expression ofthe structural polypeptides of light-harvestingcomplex I and the two integral membrane-proteins of the reaction center. Since, however,many light-harvesting I complexes are requiredper reaction center in Proteobacteria, additionalregulatory mechanisms must exist. Differentialdegradation of various portions of the poly-cistronic mRNA are one means to regulate thestoichiometry of different components of thephotosynthetic apparatus. The synthesis of dif-ferent amounts of gene products is achieved byposttranscriptional regulation (Rödig J. et al.,1999). Because of a h ighly stable secondary ter-minator structure at its 3´-end and the absenceof specific recognition sites for endonucleolyticcleavage, the mRNA coding the two light-harvesting polypeptides has much higher stabil-ity than that of the entire puf gene transcript. Thedegradation of the downstream pufLM sectionof the mRNA is mediated by an endonuclease.A similar regulation mechanism may exist forthe polycistronic mRNA of bacteriochloro-phyll synthesis genes (bchFNBHLM-F1696)and the puhA, and operate in regulation of light-harvesting complex II expression.
A shift to low light intensities results in anincrease especially of light-harvesting complexII. The corresponding pucBA operon is highlyexpressed but only moderately regulated (4-fold). In the purple nonsulfur bacterium Rhodo-bacter capsulatus, four-fold less puc mRNA butat the same time four times as many light-harvesting II complexes were detected after ashift from high to low-light conditions (Zucconiand Beatty, 1988). Therefore regulation by lightmost likely involves posttranscriptional regula-tion. A posttranscriptional regulation appears tooccur (Bauer, 1995).
Bacteriochlorophyll and carotenoid biosyn-thesis genes are only weakly expressed and mod-erately (2 to 4-fold) regulated. Light intensitymay control the rate of bacteriochlorophyll deg-radation (by oxidative degradation of bacterio-chlorophyll; Biel, 1986) rather than the rate ofsynthesis (Biel, 1995). This is another distinct dif-ference from the regulation by oxygen, whereinhibition of d-aminolevulinate synthase bymolecular oxygen appears to occur (see
Chemotrophic Growth with O2 in this Chapter).Bacteriochlorophyll may be stabilized by inser-tion in pigment-protein complexes, however. Thepromotor of the bacteriochlorophyll synthesisgene bchC is of the sigma-70 type and leads toone large superoperon (Yurkov and Beatty,1998). In contrast, an alternative sigma factorappears to recognize the strongly regulatedstructural puf and puh genes (Bauer, 1995).These differences explain the independent anddifferent levels of regulation observed for thetwo classes of genes.
Recently the promoter for the carotenoid bio-synthesis genes crtB and crtP were identified inSynechocystis PCC 6803, and shown to be lightregulated (Fernández-González et al., 1998).
State TransitionsIn cyanobacteria, state transitions involve redi-recting the pathways of excitation energy trans-fer from light-harvesting complexes to bothphotosystems, and can be recognized by fluores-cence analysis. Cyanobacteria can reach twoenergetically different states, in which one of thephotosystems is preferentially excited. This isachieved with fast changes in the couplingbetween the light-harvesting complexes and thereaction center (van Thor et al., 1999). Evidenceis accumulating that at least in the chlorophyll b-containing phototrophic bacteria (“Prochloro-phytes”), the short-term regulation occurs by amechanism similar to that in green chloroplasts(Matthijs et al., 1994). In the latter, polypeptidesof the PSII antenna (LHCII) are rapidly phos-phorylated during overexcitation of this photo-system, and as a consequence detach from PSIIand migrate to the stromal thylakoids. Thismechanisms ensures a bala nced energy distribu-tion between PSII and PSI. The net result of statetransitions is the balanced function of both pho-tosystems and an optimization of the quantumyield for photosynthesis during short-termchanges, such as those that planktonic cells mightexperience during vertical transport by watercurrents.
Movement by FlagellaPhototrophic Proteobacteria swim by means offlagella, whereas one species of the green sulfurbacteria (Chloroherpeton thalassium), membersof Chloroflexus subgroup and cyanobacteriamove by gliding. Of the a-Proteobacteria, mostphototrophic species are motile. Peritrichous orlateral flagella are only found in Rhodomicro-bium vannielii and the swarming phase ofRhodocista centenaria. About two thirds of theknown Chromatiaceae species are motile. Largerforms (Chromatium okenii, Chr. weissei, Chr.warmingii, Chr. buderi, Thiospirillum jenense)

CHAPTER 1.3 The Phototrophic Way of Life 59
are motile by means of bipolar multitrichoustufts of flagella. Thiospirillum jenense is bipolarlyflagellated. Forms with smaller cells are monot-richously flagel lated (small Chromatium species,Lamprocystis, Thiocystis, Thiorhodococcus,Thiorhodovibrio). All Ectothiorhodospiraceaeare flagellated. A new mode of motility has beendescribed for a unicellular cyanobacteriumwhich moves in a similar fashion to flagellatedbacteria but apparently lacks a flagellum (Water-bury et al., 1985).
True phototaxis is the ability to move towardsor away from the direction of light. Cyanobacte-ria are the only prokaryotes displaying true pho-totaxis (Garcia-Pichel and Castenholz, 1999).Phototaxis may not be of competitive value formicroorganisms adapted to live at low lightintensities in the subsurface of sediments, soilsand mats because the light fields may be closeto diffuse deep below the surface. However,directed movements can still be of much use inmicroorganisms dwelling at or close to the sedi-ment surface, where the light fields contain asignificant downward directionality. Photopho-bic responses are changes in the direction ofmovement in reaction to abrupt changes in lightintensity (Castenholz, 1982; Häder, 1987). In thestep-up photophobic response, organisms willreverse direction when sensing an increase inlight intensit y, which results in a net accumula-tion of organisms at lower light intensities. Ina step-down photophobic (or scotophobic)response, the organisms will tend to accumulatein the region of higher light intensity. Photopho-bic responses are the basis of photomovement inall flagellated bacteria (Armitage, 1997), and inmost gliding cyanobacteria (Castenholz, 1982).
In swimming cells of phototrophic Proteobac-teria, a decrease in light intensity triggers a rever-sal of flagellar rotation (Rhodospirillum rubrum,Chromatium spp.) or an increase in stopping fre-quency (Rhodobacter sphaeroides). Owing to amemory effect, cells of the latter species retain ahigher stopping frequency for up to 2 min, whichprevents the cells from being trapped in the darkbut instead permits reorientation of the cells anda return to higher light intensities (Armitageet al., 1995). As a result of this scotophobicresponse, the cells accumulate in the light and atwavelengths corresponding to the absorptionmaxima of photosynthetic pigments. A change inlight intensity of as little as 2% can be sensed(Armitage et al., 1995). Active electron transportis required for the scotophobic response.
The formation of flagella in Chromatium spe-cies is induced by low sulfide concentrations andlow light intensities. These two environmentalvariables are mutually dependent: the lower thelight intensity, the higher the sulfide concentra-tion at which a given strain can persist in its
motile stage (Pfennig and Trüper, 1989). In thenatural environment of purple sulfur bacteria,gradients of light and sulfide are opposed to eachother. The control of motility by the two inter-dependent environmental variables (instead ofonly one) enables Chromatium cells to returneither from low sulfide/high light environmentabove the chemocline or from the high sulfide/low light environment below the chemoclineback to their habitat.
In its pelagic habitat, Chromatium okenii maydisplay diurnal migrations with a vertical ampli-tude of about 2 m (Sorokin, 1970). In other lakes,vertical migrations of Chromatium minusextended over a distance of 30–35 cm (Lindholmet al., 1985; Pedrós-Alió and Sala, 1990). Verticalmigration of nonthermophilic Chromatium, andof Chromatium tepidum also has been observedin ponds and in intertidal or hot spring microbialmats (Castenholz and Pierson, 1995; Jørgensen,1982; Pfennig, 1978). In the latter environments,Chromatium cells migrate upwards to the surfaceof the mat and enter the overlaying water as aresult of positive aerotaxis during the night. Thecells contain high amounts of intracellular sulfurglobules, which are formed during incompletesulfide oxidation by anoxygenic photosynthesisduring daytime. It is assumed that migration intomicrooxic layers enables the cells to growchemoautotrophically by oxidation of sulfide orintracellular sulfur with molecular oxygen (Jør-gensen, 1982; Castenholz and Pierson, 1995).
If phototrophic sulfur bacteria would solelyfollow the light gradient, their scotophobicresponse would ultimately lead them into oxicwater layers. Both the scotophobic behavior andaerotaxis respond to the rate of intracellularelectron flow (presumably sensed as changes inthe redox state of an intermediate). Because thetwo tactic reponses interact through a commonsignal, a combination of light and molecular oxy-gen elicits a differential response. Rhodobactersphaeroides exhibits pronounced aerotaxis whenprecultivated aerobically, but negative aerotaxiswhen grown anaerobically in the light. Con-versely, cells only swim towards higher lightintensities in anoxic medium. A pulse of oxygenin the light causes a transient fall in the mem-brane potential which probably represents theprimary tactic signal. As a result, the bacteriamove towards environments where electrontransport rate is increased (Armitage et al.,1995).
Rhodocista centenaria exhibits a characteristicswarming behavior. In liquid media, cells movewith a single polar flagellum. Upon contact withsolid agar media, formation of a large number oflateral flagella is induced. Lateral flagella allowwhole colonies to swarm towards or away fromthe light (Ragatz et al., 1994). The supposedly

60 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
true phototaxis of these swarming colonies(Ragatz et al., 1995) has later been proven toactually be aerotaxis following microgradientswithin the colonies (Sackett et al., 1997). Thelight sensing system in this species appears to bemore complex, since infrared light leads to posi-tive, and visible light to negative phototaxis. Inmicrobial mats, infrared light penetrates to muchgreater depths than light of the visible wave-length range (see Competition for Light in thisChapter). It has been suggested that the ratio ofvisible to in frared light may be used to maintainan optimum position in such environments(Armitage et al., 1995; Ragatz et al., 1995).
Cyanobacteria are the only prokaryotes dis-playing true phototaxis (Garcia-Pichel and Cas-tenholz, 1999). Surface dwelling cyanobacteriasuch as Lyngbya spp. from hot springs mats andintertidal sediments and the motile phases (hor-mogonia) of terrestrial Nostoc spp. from desertsoils exhibit this type of movement. The bundle-forming Microcoleus chthonoplastes also is ableto display a “populational phototaxis” in thatbundles of trichomes of this cyanobacterium areable to steer in the direction of the incominglight, whereas single trichomes are apparentlynot able to do so (Prufert-Bebout and Garcia-Pichel, 1994). True phototaxis is a mechanism forthe orientation of cells at or close to the sedimentsurface, where the light field contains a signifi-cant downward directionality. In contrast, photo-taxis does not provide a selective advantage forbacteria thriving in the subsurface of sediments,soils and mats because of the diffuse light field.In natural microbial mats photophobic responsesto changes in light intensity are probablyinvolved in the migrations of gliding bacteria(Nelson and Castenholz, 1982; Pentecost, 1984).In microbial mats, some strains of cyanobacteriaare able to migrate vertically following theiroptimal light intensity over the diel cycle(Garcia-Pichel et al., 1996). The upward migra-tions of cyanobacteria in mats is preferentiallyprevented by short wavelengths, especially byUV radiation (Garcia-Pichel and Castenholz,1994b; Bebout and Garcia-Pichel, 1995; Kruscheland Castenholz, 1988) and not by red nor greenlight.
Phototrophic consortia are structural associa-tions between a colorless central bacterium andseveral surrounding cells of pigmented epibionts(see Interactions between Phototrophic Bacteriaand Chemotrophic Bacteria in this Chapter; TheFamily Chlorobiaceae in Volume 7; Fig. 5). Intactconsortia of the type “Chlorochromatium aggre-gatum” exhibit a scotophobic response and accu-mulate in a spot of white light. In phototrophicconsortia, only the central colorless bacteriumcarries a flagellum (J. Glaeser and J. Overmann,unpublished observation). The action spectrum
of scotophobic accumulation corresponds to theabsorption spectrum of the green sulfur bacterialepibionts, however. It has to be concluded that arapid signal transfer exists between the light-sensing but immotile epibionts and the colorlessmotile rod (Fröstl and Overmann, 1998).
Gas VesiclesBuoyancy-conferring gas vesicles are common ingreen sulfur bacteria, Chromatiaceae, and cyano-bacteria. Gas vesicles are cylindrical structureswith conical ends; their length and width arevariable and species-specific. The sheath of gasvesicles are composed of proteins (Walsby,1994). The gas mixture within the gas vesicles isthe same as in the surrounding medium and is atthe same partial pressures. Gas vesicles occur ina third of the species of Chromatiaceae (belong-ing to the genera Amoebobacter, Lamprobacter,Lamprocystis, Thiodictyon, Thiopedia, Thiolam-provum) and some green sulfur bacteria (generaAncalochloris, Pelodictyon, Chloroherpeton). Ofthe Ectothiorhodospiraceae, only Ectothiorho-dospira vacuolata forms gas vesicles during sta-tionary phase. This reflects the distribution ofboth families of purple sulfur bacteria in nature,where Chromatiaceae typically colonize low-light stratified aquatic environments, whereasEctothiorhodospiraceae typically inhabit moreshallow saline ponds and sediments. Gas vesiclesalso are present in Prochlorothrix hollandica. Inplanktonic habitats, cells of cyanobacteria andphototrophic sulfur bacteria often contain gasvesicles, which indicates a selective advantage ofthis cellular property.
Gas vesicle formation in the green sulfur bac-terium Pelodictyon phaeoclathratiforme isinduced exclusively at light intensities <5 molquanta·m–2·s-1 (Overmann et al., 1991b). Thisappears to be the reason for the rare observationof gas vesicles in pure cultures of green sulfurbacteria which routinely are incubated at muchhigher light intensities. A transfer of Amoebo-bacter purpureus strain ML1 to the dark resultsin an increase of the gas vesicle volume by a factorof nine (Overmann and Pfennig, 1992). Ambienttemperature controls gas vesicle formation inThiocapsa pendens (Eichler and Pfennig, 1986).
The buoyancy of many species of Cyanobacte-ria is regulated by the formation of gas vesicles.Highly buoyant cells may float towards the sur-face of stagnant water bodies. When the turgorpressure within the surrounding cytoplasm rises,such as by accumulation of low molecular weightphotosynthates during periods of intense photo-synthesis, the critical pressure may be exceededand the gas vesicles collapse. New vesicles areformed by de novo synthesis rather than by re-inflation of collapsed vesicles. Short-term regula-

CHAPTER 1.3 The Phototrophic Way of Life 61
tion of cell buoyant density occurs in cyanobac-terial species thriving in stratified lakes, likeAphanizomenon flos-aquae, Anabaena flos-aquae, and green-colored Oscillatoria spp.(Konopka et al., 1978; Oliver and Walsby, 1984;Utkilen et al., 1985). In these species, the pro-teinaceous gas v esicle sheaths are weak enoughto permit a collapse at high intracellular turgorpressures as they are reached during periods ofintense photosynthesis. By this mechanism, cellslose buoyancy within 30 minutes and thus cansink out of surface layers of stratified lakes. Afterde novo synthesis of gas vesicles in lower waterlayers, utilization of photosynthates, and adecrease of turgor pressure, cells rise back to thesurface during the night. Rapid, turgor-mediatedreduction of buoyancy together with gas vesicleformation thus represents an adaptation to thepronounced diurnal variations in light intensityand the limitation of growth by inorganic nutri-ents as they occur during summer stratificationin the surface layer of eutrophic lakes. In someinstances (e.g., Microcystis aeruginosa) diurnalmigrations are mediated by an increase of carbo-hydrate ballast alone and gas vesicles do notcollapse even at maximum turgor pressure(Kromkamp and Mur, 1984; Thomas and Walsby,1985).
In contrast, gas vesicles of red-colored Oscil-latoria aghardii and of phototrophic sulfur bac-teria are mechanically stronger and do notcollapse even at maximum cell turgor pressure.A decrease in the cellular gas vesicle content istherefore the result of their dilution duringgrowth and division of the cells, and thus pro-ceeds rather slowly (Overmann et al., 1991b;Overmann and Pfennig, 1992). Bacteria of thiscategory mostly colonize the low-light environ-ments shortly above or within the chemocline ofstratified lakes where photosynthetic rates typi-cally are strongly limited by light and hydrostaticpressure is high. Gas vesicles in green sulfur bac-teria are rigid enough to persist at hydrostaticpressures down to depths of 38 m (Overmann etal., 1991b). The cyanobacterium Trichodesmiumcontains extremely stable gas vesicles (mean critical collapse pressures up to 3.7 MPa, corre-sponding to a depth of 370 m; Walsby, 1978). Thedifferences in strength of gas vesicles formed bydifferent species is related to their shape(especially the diameter) and the primary struc-ture of the GvpC protein of their sheath (Walsby,1994).
In addition to the formation of gas vesicles, anew type of buoyant density regulation wasdetected in Pelodictyon phaeoclathratiforme.Cells of this species form large extracellularslime layers during the stationary phase whichleads to an increase of the cellular volume by afactor of three (Overmann et al., 1991b).
Advantages of the Vertical Movement by Flagella and by Gas Vesicles
Theoretically, motility based on flagellar move-ment and vertical migration by means of gas ves-icle formation have different advantages undernatural conditions. Movement by flagellarequires a permanent, (albeit sometimes low)fraction of metabolic energy (proton-motiveforce), whereas gas vesicle synthesis representsan initial one-time investment of a higher amountof metabolic energy. Once formed, gas vesicleskeep bacterial cells in their habitat without anyfurther demand for energy. The purple sulfur bac-terium Lamprobacter modestohalophilus is capa-ble of both flagella and gas vesicle formation.Motile cells are usually devoid of gas vacuolesand initially dominate during growth in freshmedia. Later, cells become immotile and formgas vesicles and slime capsules (Gorlenko et al.,1979). In a very similar manner, cells of Ectothior-hodospira vacuolata are flagellated at low sulfideconcentrations and light inte nsities, and becomeimmotile and form gas vesicles in stationary phase(Imhoff et al., 1981). This supports the view thatflagellar movement of purple sulfur and purplenonsulfur bacteria is favored under conditions ofcontinuous energy supply, while gas vesicle for-mation represents an adaptation to conditions ofstarvation. Within one lake ecosystem, verticalmigration of a flagellated species (Chromatiumminus) was observed while the gas-vacuolatedAmoebobacter did not change its vertical position(Pedrós-Alió and Sala, 1990).
A minimum quantum requirement of flagellarmotility can be estimated from data in the liter-ature. A vertical migration over a distance of2 m (the maximum amplitude of vertical migra-tion observed in nature) during 6 hours corre-sponds to a swimming speed of 93 mm·s-1. At asimilar speed of 100 mm·s-1 the frequency offlagellar rotation is >100 s-1 in Rhodobactersphaeroides and requires between 200 and 1000 H+
per rotation (Armitage et al., 1995). This yieldsa proton translocation rate of ~6 ¥ 104 H+·s-1 at aswimming velocity of 100 m·s-1. Based on anabsorbing cross sectional area of the cell of 1 m2,an absorption of 36% of the incident light (seeEfficiency of Light Harvesting in this Chapter),a ratio of protons translocated to electrons trans-ferred (H+/e- ratio) of 2 (see Conversion of Lightinto Chemical Energy in this Chapter), andassuming that each photon absorbed leads totransport of an electron, the proton translocationrate of 6 ¥ 104 H+·s-1 would be reached at anunderwater irradiance of 0.2 mol quanta·m-2·s-1.However, all available quanta would be requiredjust for motility at this irradiance and no verticalmigration would be possible during the night.Therefore motility by flagella will be of compet-

62 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
itive advantage only at significantly higher irra-diances. In many lakes, underwater irradiancesin layers of phototrophic sulfur bacteria are £1mol quanta·m–2·s-1 (Overmann and Tilzer, 1989a;Overmann et al., 1999a). Under these conditions,purple sulfur bacteria harboring gas vesiclesdominate over flagellated forms in thechemocline community (Fig. 4). At least in somelakes, gas vesicles appear to be of selectiveadvantage also at higher underwater irradiances(Overmann et al., 1991b; Overmann andPfennig, 1992).
Interestingly, the extremely low-light adaptedChlorobium phaeobacteroides strain MN1 iso-lated from the chemocline of the Black Sea wasnot capable of gas vesicle formation. The greensulfur bacterial layer is located at an 80-m depthand with respect to light intensity represents thelower limit for growth of a phototrophic organ-ism (see The Family Chlorobiaceae in Volume 7).The isolated strain exhibits an extremely lowmaintenance energy requirement. It thereforeappears that gas vesicle formation is too energydemanding at the very low light intensitiesavailable at an 80-m depth in the Black Sea.
Carbon Metabolism of Phototrophic Prokaryotes
In the natural environment, the principal carbonsource of phototrophic bacteria in manyinstances is CO2 (Madigan et al., 1989; SinningheDamsté et al., 1993; Takahashi et al., 1990). InCyanobacteria, Chromatiaceae, Ectothiorho-dospiraceae and purple nonsulfur bacteria, CO2
is assimilated by the reductive pentose phos-phate or Calvin cycle. Employing this cycle, theformation of one molecule of glyceraldehyde-3-phosphate requires 6 NAD(P)H+H+ and 9 ATP.By comparison, the reductive tricarboxylic acidcycle used for CO2-assimilation by green sulfurbacteria requires 4 NADH+H+, 2 reduced ferre-doxins, and only 5 ATP. As two of the reactionsof the reductive tricarboxylic acid cycle (thea-oxoglutarate synthase and pyruvate synthaserea ctions) require reduced ferredoxin as elec-tron donor, this pathway of CO2 fixation can onlyproceed under strongly reducing conditions.Furthermore, reduced ferredoxin is a primaryproduct of the light reaction only in FeS-typereaction centers. Ultimately, the lower demandfor ATP is possible because of the adapatationof green sulfur bacteria to the strongly reducingconditions of their natural environment. CO2-fixation by the hydroxypropionate cycle inChloroflexus aurantiacus requires 8 ATP perglyceraldehyde-3-phosphate and therefore isenergetically less favorable than in green sulfurbacteria.
Organic carbon as it is present in canonicalmicrobial biomass (<C4H8O2N>; Harder andvan Dijken, 1976) is considerably more reducedthan CO2. Given the high energy demand ofautotrophic growth, the capability for assimila-tion of organic carbon compounds is of selectiveadvantage especially if natural populations arelimited by light or by low concentrations ofelectron-donating substrates, as is typically thecase for phototrophic sulfur bacteria. At limitingconcentrations of sulfide or thiosulfate, the cellyield of green sulfur bacteria is increased threetimes if acetate is available as an additional car-bon source (Overmann and Pfennig, 1989b).Acetate represents one of the most importantintermediates of anaerobic degradation oforganic matter (Wu et al., 1997). That almost allanoxygenic phototrophic bacteria (with theexception of Rhodopila globiformis; Imhoff andTrüper, 1989) are capable of acetate assimilationis therefore not surprising. In most phototrophicProteobacteria, acetate is assimilated by acetyl-CoA synthetase and the enzymes of theglyoxylate cycle. In green sulfur bacteria, theferredoxin-dependent pyruvate synthetase, PEPsynthetase, and reactions of the reductive tricar-boxylic acid cycle serve this purpose. The capac-ity for organotrophic growth seems to correlatewith the presence of a-oxoglutarate dehydroge-nase. The latter is a key enzyme for the com-plete oxidation of the carbon substrates in thetricarboxylic acid cycle (Kondratieva, 1979),whereas a complete cycle is not needed for thephotoassimilation during the presence of inor-ganic electron donors. The range of carbonsubstrates utilized and the capacity forphotoorganotrophy or chemoorganotrophy var-ies considerably among the different groups ofphototrophic pr okaryotes (Pfennig and Trüper,1989).
Organic carbon compounds not only areassimilated but also can serve as photosyntheticelectron donors in purple nonsulfur bacteria,some Chromatiaceae and Ectothiorhodospi-raceae, all Heliobacteriaceae, and members of theChloroflexus subdivision.
Green sulfur bacteria are the least versatile ofall phototrophic prokaryotes. All known speciesare obligately photolithotrophic and assimilateonly very few simple organic carbon compounds(acetate, propionate, pyruvate). Few strains havebeen shown to assimilate fructose or glutamate.Whereas green sulfur bacteria have a highergrowth affinity for sulfide than purple sulfur bac-teria, acetate seems to be used by purple sulfurbacteria at an affinity 30 times higher than ingreen sulfur bacteria (Veldhuis and van Gemer-den, 1986). In addition, uptake of acetate inChlorobium phaeobacteroides is inhibited bylight (Hofman et al., 1985).

CHAPTER 1.3 The Phototrophic Way of Life 63
Based on their metabolic flexibility, two groupscan be distinguished among the Chromatiaceae.Several species (Chromatium okenii, Chr. weissii,Chr. warmingii, Chr. buderi, Chr. tepidum,Thiospirillum jenense, Lamprocystis roseopersic-ina, Thiodictyon elegans, Thiodictyon bacillo-sum, Thiocapsa pfennigii, Thiopedia rosea) areobligately phototrophic, strictly anaerobic andphotoassimilate acetate and pyruvate only in thepresence of CO2 and sulfide. Assimilatory sulfatereduction is absent in these species (Pfennig andTrüper, 1989). However, particularly those spe-cies with limited metabolic flexibility form denseblooms under natural conditions (see Coexist-ence of Phototrophic Sulfur Bacteria in thisChapter). The second physiological group withinthe Chromatiaceae comprises the small Chroma-tium species (Chr. gracile, Chr. minus, Chr.minutissimum), Allochromatium vinosum,Lamprobacter modestohalophilus, as well asThiocystis spp., Thiocapsa. Most of these speciesuse thiosulfate as electron donor and a widerange of organic carbon compounds includingglucose, fructose, glycerol, fumarate, malate,succinate, formate, propionate, and butyratefor photoassimilation, and often are capableof assimilatory sulfate reduction. In somespecies (especially Allochromatium vinosum),these organic carbon substrates also serveas electron-donor for phototrophic orchemotrophic growth.
Most Ectothiorhodospiraceae species arecapable of photoorganotrophic growth, withEctothiorhodospira halophila and Ectothiorho-dospira halochloris being the exceptions. Thespectrum of electron-donating carbon substratesfor photoorganotrophic growth resembles thatfound in the versatile Chromatium species (Pfen-nig and Trüper, 1989). Assimilation of acetateand propionate proceeds by carboxylation andtherefore depends on the presence of CO2.
Chloroflexus aurantiacus grows preferably byphotoorganoheterotrophy (Pierson and Casten-holz, 1995). The carbon substrates utilizedcomprise acetate, pyruvate, lactate, butyrate,C4-dicarboxylic acids, some alcohols, sugars andamino acids (glutamate, aspartate). This versatil-ity has been seen as the major cause for theprofuse growth of Chloroflexus in microbial matswhere accompanying microorganisms, especiallycyanobacteria, may provide the required carbonsubstrates (Sirevåg, 1995). However, high ratesof formation of low-molecular-weight organiccarbon substrates by the anaerobic food chainhave also been observed in other stratified sys-tems, where the dominating anoxygenic pho-totrophs could utilize only a narrow range ofcarbon substrates (Overmann, 1997; Overmannet al., 1996). The refore, the presence of low-molecular-weight organic carbon substrates is
not necessarily the most selective factor in thenatural environment.
Slow photolithoautotrophic growth with H2Sor H2 as electron-donating substrates has beenshown in laboratory cultures of Chloroflexusaurantiacus and in hot spring populations (Pier-son and Castenholz, 1995). Carbon fixation pro-ceeds by carboxylation of acetyl-CoA and viahydroxypropionyl-CoA as an intermediate andyields glyoxylate as the net product (hydroxypro-pionate cycle; Holo, 1989; Strauß and Fuchs,1993; Eisenreich et al., 1993). So far this cycle hasnot been found in any other member of the Bac-teria. Glyoxylate is further assimilated into cellmaterial with tartronate semialdehyde and 3-phosphoglycerate as intermediates (Menendezet al., 1999).
The highest metabolic versatility is found inphototrophic a- and b-Proteobacteria (purplenonsulfur bacteria). All representatives growphotoorganoheterotrophically and (with theexception of Blastochloris viridis) photolithoau-totrophically with H2 in the light. In addition tothe substrates used by versatile purple sulfur bac-teria, the spectrum of substrates that can serveas electron donors comprise long-chain fattyacids (like pelargonate), amino acids (aspartate,arginine, glutamate), sugar alcohols (sorbitol,mannitol), or aromatic compounds (benzoate;Imhoff and Trüper, 1989). With the exception ofRubrivivax gelatinosus, none of the purple non-sulfur bacteria is capable of degradation of poly-mers and therefore depends on the anaerobicfood chain for the supply of electron-donatingsubstrates required for growth. This dependenceand the competition with chemotrophs for thecarbon substr ates might be the major reason whydense blooms of purple nonsulfur bacteria do notoccur under natural conditions (see Habitats ofPhototrophic Prokaryotes in this Chapter). Somespecies are capable of also using reduced sulfurcompounds as electron donors. However, mostspecies oxidize sulfide to elemental sulfur only(Hansen and van Gemerden, 1972).
In Heliobacteriaceae, only a limited number ofcarbon substrates can serve as photosyntheticelectron donor including pyruvate, ethanol, lac-tate, acetate, and butyrate. High levels of sulfideare inhibitory (Madigan, 1992; Madigan andOrmerod, 1995).
Cyanobacteria are obligate autotrophs parexcellence; however, small molecular weightorganic compounds such as acetate, sugars andamino acids are assimilated. In the case of aminoacids, the presence of various efficient uptakesystems has been interpreted as a means ofrecovery of leaked organic nitrogen, rather thana true chemotrophic capability (Montesinos etal., 1997). Certain strains of cyanobacteria cangrow facultatively as chemoheterotrophs in the

64 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
dark (Rippka et al., 1979), but even under theseconditions all of the photosynthetic machinery issynthesized. This lack of regulation implies thatchemotrophy has played no significant evolu-tionary role in these organisms.
Chemotrophic Growth with O2
Ecophysiology of Chemotrophic GrowthIn lakes, purple sulfur and green sulfur bacteriaare confined to environments where lightreaches sulfide-containing water layers. Thephysiological properties restrict the distributionof these bacteria in the pelagic habitat (Pfennig,1978). Dense accumulations of anoxygenic pho-totrophic bacteria, which apparently are growingchemotrophically, are only known for Chlorof-lexus (see Habitats of Phototrophic Prokaryotesin this Chapter). Although populations of otheranoxygenic phototrophic bacteria do not seem togrow permanently by chemotrophy, the ability ofmany strains to shift to an aerobic chemotrophicmode of growth is of selective advantage in envi-ronments like intertidal sediments.
Green sulfur bacteria and Heliobacteriaceaeare obligate anaerobes. Under oxic conditions,the reaction of reduced ferredoxin of the type Ireaction center with molecular oxygen wouldcreate superoxide and other activated oxygenspecies. Heliobacteriaceae are rapidly damagedby exposure to molecular oxygen. This has beenattributed not only to the formation of toxic oxy-gen radicals but also the destruction of the unsat-urated fatty acids present in the cell membraneby activated oxygen species (Madigan andOrmerod, 1995). In green sulfur bacteria, ithas been observed that the energy transfer fromlight-harvesting bacteriochlorophylls c/d/e tobacteriochlorophyll a drops by a factor of 10after an increase in redox potential due to thequenching by chlorobium quinone. This mecha-nism may protect the cells during brief anoxic/oxic transitions. (see The Family Chlorobiaceae,Physiology section in Volume 7).
All other groups of phototrophic prokaryotescomprise species that not only generate meta-bolic energy by photosynthesis but are also capa-ble of chemosynthesis with O2.
Chloroflexus aurantiacus is capable of growthas an aerobic heterotroph. During phototrophicgrowth, b-carotene, g-carotene, and hydroxy-g-carotene-glucoside are the major carotenoids,whereas echinenone and myxobactone predom-inate in aerobically grown cells (Pierson andCastenholz, 1995). Unlike in purple nonsulfur orpurple sulfur bacteria, synthesis of some caro-tenoids by C. aurantiacus is greatly enhancedunder aerobic conditions (Pierson and Casten-
holz, 1974). The expression of the chlorosomeCsmA protein is transcriptionally or posttran-scriptionally regulated by oxygen (Theroux et al.,1990).
Almost all known species of phototrophic a-and b-Proteobacteria (purple nonsulfur bacte-ria) are capable of microaerophilic or aerobicchemoorganoheterotrophic growth with oxygenas terminal electron acceptor. Of the purple sul-fur bacteria, Ectothiorhodospira species, andeight small-celled species of the Chromatiaceae(Thiocapsa rosea; Chromatium gracile; Chr.minus; Allochromatium vinosum; Thiocystis vio-lascens; Thiocapsa roseopersicina; Thiocystis vio-lacea; Thiorhodovibrio winogradskyi) can growby chemolithotrophy, oxidizing sulfide or thiosul-fate with molecular oxygen (De Wit and vanGemerden, 1987b; Kämpf and Pfennig, 1980;Overmann and Pfennig, 1992). Only few speciesgrow also chemoorganotrophically with organiccarbon substrates as electron donor of respira-tion. The group of facultatively chemotrophicChromatiaceae includes typical inhabitants ofbenthic microbial mats like Thiocapsa roseoper-sicina and Thiorhodovibrio winogradskyi. This isnot surprising considering the pronounced oxic/anoxic fluctuations in this type of habitat. Thecells of purple sulfur bacteria in benthic systemsare often immotile and form aggregates togetherwith sand grains, apparently as an adaptation tothe hydrodynamic instability of the habitat (vanden Ende et al., 1996). At the same time, how-ever, immotile cells are exposed to strong diurnalvariations in oxygen concentrations. The growthaffinities for sulfide are lower for chemotrophi-cally growing Thiocapsa roseopersicina than forcolorless sulfur bacteria, which may explain thatno natural populations of purple sulfur bacteriaare known that grow permanently by chemotro-phy (see Interactions between PhototrophicSulfur Bacteria and Chemotrophic Bacteria inthis Chapter).
When grown anaerobically in the light, facul-tatively chemotrophic species of the purple non-sulfur and purple sulfur bacteria contain apotentially active repiratory system and exhibit"50% of the respiratory activity of chemotroph-ically growing cells (De Wit and van Gemerden,1987a; Kämpf and Pfennig, 1980; Overmann andPfennig, 1992; Pfennig, 1978). In cells that stillcontain bacteriochlorophyll, respiration is inhib-ited by light. This indicates that respiration andphotosynthesis are coupled (e.g., by the mem-brane potential or common redox carriers; Rich-aud et al., 1986). An example is the solublecytochrome c2 which has a dual function inRhodobacter sphaeroides where it is needed forelectron transfer from the cytochrome bc1 com-plex to the reaction center during ph otosynthe-sis, and to the cytochrome c oxidase during

CHAPTER 1.3 The Phototrophic Way of Life 65
respiration with molecular oxygen. During pho-tosynthetic growth, expression of cytochromec2 is increased. At limiting concentrations ofelectron donating substrate, photosynthesis ispreferred over respiration as long as theintracellular bacteriochlorophyll content ismaintained at a sufficiently high level (4–7 g bac-teriochlorophyll a·mg protein–1 in Thiocapsaroseopersicina at light saturation; De Wit andvan Gemerden, 1990a).
Growth continues after a shift to microoxic oraerobic conditions. Under oxic conditions thesynthesis of pigments and of pigment-bindingproteins of the photosynthetic apparatus ceases.The number of intracellular membrane vesiclesis reduced dramatically and the composition ofmembrane lipids is altered. The pigment contentin purple sulfur bacteria is inversely related tothe ambient oxygen concentration (Kämpf andPfennig, 1986). At 25% air saturation (52 M) ofoxygen, pigment synthesis in Thiocapsa roseop-ersicina is completely repressed and cells becomecolorless (De Wit and van Gemerden, 1987b). Incontinuous cultures of purple sulfur bacteria,active degradation has not been observed andintracellular bacteriochlorophyll concentrationsfollow the washout curve. Thus bacteriochloro-phyll does not seem to be actively degraded butis diluted out by cell division (De Wit and vanGemerden, 1987b). Concomitantly, the activitiesof respiratory enzymes (NADH dehydrogenase,cytochrome c oxidases) are increased inchemotrophically grown cells. When the cells ofThiocapsa roseopersicina become colorless, theyuse only one third of the electron donor forreduction of CO2. The remaining two thirds areused for energy generation and respired. Corre-spondingly, the protein yield reaches one third ofthat of phototrophically grown cells (De Wit andvan Gemerden, 1987b; De Wit and van Gemer-den, 1990b).
In aerobic phototrophic bacteria, aerobicgrowth is stimulated by light that is absorbed bybacteriochlorophyll a. This stimulation is onlytransient, however, since bacteriochlorophyllsynthesis is repressed even by low light intensi-ties (Yurkov and van Gemerden, 1993) thusleading to a loss of the photosynthetic apparatusunder continuous illumination.
Respiration in cyanobacteria involves a fullrespiratory chain including a cytochrome aa3 ter-minal oxidase. Monomeric sugars are degradedusing the oxidative pentose phosphate cycle. Acomplete tricarboxylic acid cycle has never beenshown for any cyanobacterium. The NADPHformed in sugar catabolism is fed to themembrane-bound electron transport chain at thelevel of plastoquinone. This is in contrast togreen chloroplasts, in which plastoquinol isautoxidized (Peltier and Schmidt, 1991). The res-
piratory electron transport chain of cyanobacte-ria is located in both the plasma and thethylakoidal membrane, and it shares many func-tional components with photosynthetic electrontransport. The role of exogenous respiration oforganic substrates is probably minor under natu-ral conditions. Under anoxia, the known electronacceptor alternatives to oxygen for cyanobacte-rial chemoorganotrophy are some organic com-pounds and elemental sulfur. Fermentationseems to be a relatively widespread ability inbenthic and bloom-forming cyanobacteria, but itis not universal (Moezelaar and Stal, 1994).
Genetic Regulation by O2
A shift from anoxic to oxic growth conditionsrequires the expression of new proteins andcofactors. On the genetic level the formation ofthe photosynthetic apparatus and the intracyto-plasmic membrane system is regulated by twomain environmental variables, light intensity (seeResponse to Changes in Light Intensity and Qual-ity in this Chapter) and molecular oxygen. Thetwo factors act independently of one another andare involved in different mechanism of regulationof bacteriochlorophyll synthesis (Arnheim andOelze, 1983). Compared to light, molecular oxy-gen acts as a stronger repressor, however.Although oxygen is a major factor controlling theformation of the photosynthetic apparatus inmost of the facultatively phototrophic Proteobac-teria, Rhodovulum sulfidophilum and Rhodocistacentenaria are exceptional in that these speciesform the photosynthetic apparatus under bothaerobic and anaerobic conditions (Hansen andVeldkamp, 1973; Nickens et al., 1996). Photopig-ment synthesis is not repressed by O2 inRhodocista centenaria.
The regulation of bacteriochlorophyll synthe-sis in purple nonsulfur bacteria is complex. Thecells synthesize very little bacteriochlorophyll,probably because of the inhibition of bacterio-chlorophyll biosynthesis enzymes (the d–ami-nolevulinic acid synthesis and enzymes for theconversion of coproporphyrin; Oelze, 1992) byO2. Oxygen does not seem to exert an effectivetranscriptional control. Under oxic conditionsthe transcription of bacteriochlorophyll synthe-sis genes decreases 2-fold, while that of light-harvesting I and reaction-center genes decreasesby a factor of 30–100 (Bauer, 1995). The tetrapy-rrole synthesis pathway has four differentbranches (leading to heme, bacteriochlorophyll,siroheme and vitamin B12). While the bacterio-chlorophyll content is drastically reduced in thepresence of oxygen (Arnheim and Oelze, 1983),heme synthesis remains unaffect ed (Lascelles,1978). The intracellular activity of d-aminole-vulinic acid synthase, the key enzyme of tetrapy-

66 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
rrol synthesis in a-Proteobacteria, is reduced inthe presence of oxygen. Regulation by oxygenmay occur also during some later steps of tet-rapyrrole synthesis. It appears that oxygen inhib-its magnesium chelatase, thereby increasing theprotoporphyrin IX pool, which in turn leads toincreased formation of heme. Feedback inhibi-tion of d-aminolevulinate synthase by hemewould then slow down the synthesis of interme-diates but still guarantee the amount needed forheme biosynthesis (Beale, 1995; Biel, 1995;Rebeiz and Lascelles, 1982).
After return to anoxic conditions the synthesisof the photosynthetic apparatus and intracellu-lar membranes occurs in a light-independentmanner. Anoxygenic photosynthetic bacteriacontain a distinct light-independent protochloro-phyllide reductase, composed of probably threesubunits (BchN, BchB, and BchL). In angio-sperms, the reduction of the fourth ring of theMg-tetrapyrrole intermediate by NADPH-protochlorophyllide oxidoreductase is a light-dependent step in the chlorophyll biosyntheticpathway. This protein represents one of the onlytwo enzymatic transformations known to requirelight (Suzuki and Bauer, 1995). Cyanobacteria,green algae and gymnosperms contain both, thelight-dependent and light-independent pro-tochlorophyllide reductase. The capacity to syn-thesize (bacterio)chlorophyll in the dark is ofsignificance for the competitive success of Chro-matiaceae in intertidal microbial mats. Duringanoxic conditions in the dark, Thiocapsa roseop-ersicina can synthesize bacteriochlorophyll a atmaximum rate. Under the fluctuating conditionsas they are observed in benthic microbial mats(oxic light, anoxic dark phase), purple sulfur bac-teria therefore can maintain a photosyntheticmode of growth as long as bacteriochlorophyllsynthesis during the night compensates for thewash out of pigments during the day (De Wit andvan Gemerden, 1990b).
A multicomponent regulatory cascade con-trols the coordinate expression of the light-harvesting and reaction center puf, puh, and pucgenes and involve various transcription factors(Bauer, 1995; Bauer and Bird, 1996). In Rhodo-bacter capsulatus, a redox-sensitive repressor(CrtJ) binds under oxic conditions to a con-served palindrome sequence in promotors ofbacteriochlorophyll, carotenoid, and light-harvesting complex II genes. A second system forthe regulation of the puf, puh, and puc operonsprobably consists of three components, a mem-brane-spanning sensor kinase (RegB), a solubleresponse regulator (RegA), and a hypotheticalactivator of the nonspecific alternative sigma fac-tor sP (RegX). A decrease in oxygen tensioncauses autophos phorylation of the membrane-spanning sensor kinase RegB, which then phos-
phorylates the cytoplasmic response regulatorRegA. The latter acts as intermediate and prob-ably transfers its phosphate to a putative thirdDNA-binding component that activates geneexpression. The RegA-RegB system also isinvolved in regulation of the expression of cyto-chrome c2 and the Calvin cycle CO2 fixationgenes and therefore is of general significance forthe regulation of cellular metabolism.
The transcripts of the photosynthetic genecluster exceed 10 kb and extend from pigmentbiosynthesis genes across promoter regions andinto the genes for light-harvesting complex I andreaction center proteins. In Rhodobacter capsu-latus, transcription of the genes coding structuralpolypeptides of the reaction center and light-harvesting complex I are not the only peptidesinitiated at their respective promotors. The tran-scripts of the bacteriochlorophyll biosynthesisbchCA operon extends through the promoterand coding sequences of the downstream pufBALM operon, and the transcript of the caro-tenoid biosynthesis crtEF operon extendsthrough both (Wellington et al., 1992). Similarly,the bchFBKHLM-F1696 and puhA operons aretranscriptionally linked. The linkage of operonsof different components of the phot osyntheticapparatus in such superoperons also has beendetected in other species of purple nonsulfurbacteria and may play a significant role in theadaptation of cells to changes in environmentaloxygen tension. According to a model (Welling-ton et al., 1992), the presence of superoperonsensures a rapid physiological response to adecrease in oxygen tension. In the presence ofoxygen, a basal level of light-harvesting I andreaction center polypeptides is constantlyformed and incorporated into the membrane,but these polypeptides disappear again in theabsence of bacteriochlorophyll (Dierstein, 1984;Drews and Golecki, 1995) due to degradation.After a shift from oxic to anoxic conditions, thepresence of a basal level of structural polypep-tides considerably shortens the lag time for thechange from aerobic respiratory to anaerobicphotosynthetic growth. During this lag ph ase,the cellular amount of structural polypeptides ofthe photosynthetic apparatus is further increasedby increasing the transcription rate of the pufand puh genes.
Oxygen does not only regulate the transcrip-tion of photosynthesis genes but also later stepsin gene expression. Posttranscriptional regula-tion involves mRNA processing (mRNA degra-dation) and possibly some later steps (Rödig J.et al., 1999).
In most bacteria, the formation of multiplesigma factors is a prerequisite for the coordina-tion of the regulation of a large number of genesin response to changes in environmental condi-

CHAPTER 1.3 The Phototrophic Way of Life 67
tions. Sigma factors are dissociable subunits thatconfer promoter specificity on eubacterial coreRNA polymerase and are required for transcrip-tion initiation. In phototrophic bacteria, thediversity of sigma factors of the s70 family as theyare present in the different phylogenetic groupsappears to be correlated with their metabolic flex-ibility. In the unicellular cyanobacteria Synecho-coccus sp. and Synechocystis sp., nine differentsigma factors (one member of group 1, four mem-bers of group 2, and four members of group 3)have been found, whereas one group 1 and threegroup 2 sigma factors have been found in Chlo-roflexus spp. In contrast to most other bacteria,the green sulfur bacterium Chlorobium tepidumcontains only one group 1, but no alternativegroup 2 sigma factor (Gruber and Bryant, 1998).In Chloroflexus, one group 2 s70 factor (SigB) istranscribed at fourfold higher levels during aer-obic growth and therefore appears to be involvedin the shift in metabolism. It has been proposedthat SigB is involved in regulation of pigmentsynthesis (Gruber and Bryant, 1998).
Significance of Anoxygenic Photosynthesis for the Pelagic Carbon and Sulfur CyclesThe carbon fixation of phototrophic sulfur bac-teria has been determined in a wide range ofhabitats, mostly inland lakes (Overmann, 1997;van Gemerden and Mas, 1995). The theoreticalmaximum of primary production by photo-trophic sulfur bacteria has been estimated to be10,000 mg C·m–2·d–1. Purple and green sulfur bac-teria can contribute up to 83% of total primaryproductivity in these environments. This highnumber notwithstanding, anoxygenic primaryproduction only represents a net input of organiccarbon to the food web if 1) the anaerobic foodchain is fueled by additional allochthonous car-bon from outside and 2) aerobic grazers haveaccess to the biomass of phototrophic sulfur bac-teria. Based on recent experimental evidence,these conditions are met at least in some aquaticecosystems (Overmann, 1997).
With the exception of geothermal springs, thesulfide required by phototrophic sulfur bacteriafor CO2-assimilation originates from sulfate orsulfur reduction during the terminal degradationof organic matter. This organic matter cannotbe provided solely by anoxygenic phototrophicbacteria, since growth (hence accumulation ofreduced carbon) constantly diverts electronsfrom their cycling between anoxygenic photo-trophic bacteria and sulfate-reducing bacteria.At least part of the sulfide formation is thereforefueled by carbon that has already been fixed byoxygenic photosynthetic organisms within oroutside the ecosystem. Consequently anoxygenicphotosynthesis represents not new, but second-
ary primary production. A complete degradationof the carbon fixed by phototrophic sulfur bacte-ria in the anaerobic food chain (and thus an effi-cient recycling of electrons) in an anoxygenicprimary production has been estimated toexceed oxygenic photosynthesis by as much asten times (Overmann, 1997). In reality, anoxy-genic photosynthesis surpasses that of phyto-plankton mostly in oligotrophic lakes. In manyoligotrophic lakes, the input of allochthonouscarbon derived from terrestrial sources in thewatershed is significant (Rau, 1980; Sorokin,1970). In an oligotrophic saline meromictic lake(Mahoney Lake, B.C., Canada), purple sulfurbacteria together with the anaerobic food chainefficiently converted allochthonous organic car-bon into easily degradable bacterial biomass(Overmann, 1997). It appears likely that pho-totrophic sulfur bacteria have this ecologicalfunction also in other aquatic ecosystems.
The presence of hydrogen sulfide in layers ofphototrophic sulfur bacteria may prevent theirbiomass from entering the grazing food chain.This has been substantiated by stable carbon andsulfur isotope data, which indicated that pho-totrophic sulfur bacteria are not consumed to asignificant extent by higher organisms (Fry,1986). In addition, a quantitative analysis of lossprocesses conducted in a few lakes indicates thatpredation must be of minor significance (Mas etal., 1990; van Gemerden and Mas, 1995). In con-trast, recent investigations have revealed that atleast in one lake ecosystem, a major fraction ofpurple sulfur bacterial biomass enters the aero-bic food chain via rotifers and calanoid copepods(Overmann et al., 1999b; Overmann et al.,1999c). The key environmental factors thatcaused this efficient link between anoxic and oxicwater layers were the autumnal upwelling ofphototrophic bacteria into oxic water layers bymixing currents, and the formation of gas vesiclesand large cell aggregates by the dominant spe-cies, Amoebobacter purpureus.
Sulfide formation by sulfate- and sulfur-reducing bacteria and sulfide oxidation back tosulfur and sulfate occur at comparable rates inseveral lakes (Overmann et al., 1996; Parkin andBrock, 1981). This leads to a closed sulfur cycleand a detoxification of sulfide without concomi-tant depletion of oxygen (Pfennig, 1978).
The significance of phototrophic sulfur bacte-ria for the oxidation of sulfide in stratified envi-ronments is critically dependent on their celldensity rather than the absolute biomass per sur-face area of the ecosystem (Jørgensen, 1982).Dense populations in laminated microbial matscan account for 100% of the total sulfide oxida-tion in those systems, whereas some dilutepelagic populations oxidize only very smallamounts (e.g., 4% in the Black Sea) of the sulfide

68 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
diffusing from below into the chemocline (Over-mann et al., 1991a; Overmann et al., 1996).
No information on the ecological significanceof aerobic phototrophic bacteria is available todate.
Interactions with Other MicroorganismsCompetition for Light Blue light prevails invery clear open oceans (Fig. 6) where marineSynechococcus cells thrive under conditions oflow photon flux (~10 mol quanta·m–2·s–1; Carrand Mann, 1994). Two ecotypes of the marineSynechococcus exist which differ in the intracel-lular ratio of phycourobilin to phycoerythrobilin(Waterbury et al., 1986). Two subpopulations aredistinguished according to the predominantchromophore associated with the phycoerythrin.Phycourobilin-rich strains are characteristic ofthe open oceans whereas strains with a lowerPUB content predominate in shelf waters (Olsonet al., 1990a). Compared to PEB-containingantennae (absorption maximum, ~550 nm),incorporation of PUB (absorption maximum,~495 nm) increases the efficiency of light absorp-tion significantly in deeper water lay ers of olig-otrophic oceans.
Similarly, coexisting and phylogeneticallyclosely related but genetically distinct popula-tions of Prochlorococcus are adapted for growthat different light intensities, which results in theirbroad depth distribution (Moore et al., 1998).The low-light-adapted ecotype has a higherintracellular content of chlorophylls a and b, ahigher chlorophyll b/a ratio, and exhibits ahigher maximum quantum yield reaching thetheoretical maximum of 0.125 mol C·(molquanta)–1. Its properties enable this ecotype tocolonize very low water layers. It has been sug-gested that the distribution of different ecotypesin the same water column would result in greaterintegrated production than could be achieved bya single ecotype (Moore et al., 1998).
Based on the specific physiological propertiesof oxygenic and anoxygenic phototrophic bacte-ria, multilayered microbial communities fre-quently develop in stratified pelagic and inbenthic (Fig. 5A,B) habitats. Cyanobacteria,eukaryotic algae and even plants (Lemna) formthe topmost layers overlying populations ofChromatiaceae and green sulfur bacteria (Dubi-nina and Gorlenko, 1975; Caldwell and Tiedje,1975; Pfennig, 1978; Camacho et al., 1996; Pier-son et al., 1990; Pierson et al., 1990).
Phototrophic sulfur bacteria require the simul-taneous presence of light and sulfide, which usu-ally restricts their occurrence to layers well belowthe surface of lakes and sediments. As a conse-quence of the absorption of light in the overlyingwater, the light energy available to phototrophic
sulfur bacteria in most pelagic environments israther low (0.02–10% of surface light intensity;van Gemerden and Mas, 1995; Parkin and Brock,1980b; Camacho et al., 1996). Similar values havebeen determined for purple layers in benthicmicrobial mats (Kühl and Jørgensen, 1992; Pier-son et al., 1990; Garcia-Pichel et al., 1994c). Atight correlation between anoxygenic photosyn-thesis and the amount of light reaching pho-totrophic sulfur bacteria strongly suggests thatlight i s the main environmental variable control-ling the anoxygenic photosynthesis (van Gemer-den and Mas, 1995). Therefore, a selectivepressure for efficient light harvesting andmaximum quantum yield exists in anoxygenicphototrophs. The same holds true for a few niche-specialized, deep-dwelling cyanobacteria.
The ecological niches of green sulfur bacteriaand Chromatiaceae show considerable overlapbecause both groups grow preferably or exclu-sively by photolithotrophic metabolism, usingambient sulfide as electron-donating substrate.Different species of the same group should beeven more competitive. Besides differences inmaintenance energy demand, in adaptation tolow light intensities and metabolic flexibility,another important factor determining the speciescomposition of phototrophic sulfur bacteria intheir natural habitats is the spectral compositionof underwater light. In the overlying layers, lightis absorbed by water itself, dissolved yellow sub-stance (gilvin), phytoplankton and inanimateparticulates. The limited wavelength range avail-able at great depth selects for species of anoxy-genic phototrophic bacteria with complementaryabsorption spectra. In many lacustrine habitats,light absorption by phytoplankton exceeds thatof gilvin or water itself (Kirk, 1983), and light ofthe blue green to green wavelength range reacheslayers of phototrophic sulfur bacteria. ThoseChromatiaceae which contain the carotenoidokenone (Fig. 7) dominated in 63% of the naturalcommunities studied (van Gemerden and Mas,1995). It was proposed that energy transfer fromcarotenoid antenna pigments to the reaction cen-ter is more efficient in okenone-forming strainsthan in other purple sulfur bacteria (Guerrero etal., 1986). In addition, the capability of gas vesicleformation, and the different kinetics of sulfideoxidation (see Coexistence of Phototrophic Sul-fur Bacteria in this Chapter) appear to be ofselective value for the colonization of pelagichabitats. Below accumulations of purple sulfurbacteria, the green-colored forms of the greensulfur bacteria dominate because of their supe-rior capability to harvest the light reaching them,which has its spectrum shifted to a maximumintensity at 420–450 nm (Table 2) (Montesinoset al., 1997). In contrast, the brown-colored formsof the green sulfur bacteria dominate in lakes

CHAPTER 1.3 The Phototrophic Way of Life 69
where the chemocline is located at depths greaterthan 9 m and in eutrophic lakes with a pro-nounced light absorption in the oxic zone.
A similar niche separation occurs in the pho-totrophic consortia (see The Family Chlorobi-aceae in Volume 7), which encompass green-colored or brown-colored epibionts (Overmannet al., 1999b). The ecological niche of the brown-colored green sulfur bacteria may be attributedto their use of significantly lower light intensitiesthan purple sulfur bacteria for phototrophicgrowth and to their lower maintenance energyrequirements (see Light Absorption and LightEnergy Transfer in Prokaryotes in this Chapter;The Family Chlorobiaceae in Volume 7). Anextremely low-light adapted strain of the greensulfur bacterium Chlorobium phaeobacteroideshas been isolated from the chemocline of theBlack Sea located at an 80-m depth (Overmannet al., 1991a). This isolate (strain MN1) couldgrow at light intensities as low as 0.25 mmolquanta·m–2·s-1.
In sedimentary environments with their partic-ular optical properties (Fig. 6), the irradiancereaching anoxygenic phototrophic bacteria maybe reduced to ¨1% of the surface value for lightin the visible region, while >10% of the nearinfrared light is still available (Kühl and Jør-gensen, 1992; see Light energy and the spectraldistribution of radiation). As a consequence, thelong wavelength Qy bands of bacteriochloro-phylls are significant for light-harvesting in sedi-ments, whereas light absorption of anoxygenicphototrophic bacteria in lakes is mediated bycarotenoids and the Soret bands of bacteriochlo-rophylls. In microbial mats, the spectral qualityof the scalar irradiance is strongly modified as itpenetrates. The presence of populations of pho-totrophic microorganisms impose strong absorp-tion signatures on the spectrum of the scalarirradiance (Jørgensen and Des Marais, 1988;Pierson et al., 1987). As a result of vertical nicheseparation, benthic microbial mats can consist ofup to five distinctly colored layers that areformed (from the top) by diatoms and cyanobac-teria, cyanobacteria alone, purple sulfur bacteriawith bacteriochlorophyll a, purple sulfur bacteriawith bacteriochlorophyll b, and green sulfur bac-teria (Nicholson et al., 1987). In this verticalsequence different wavelength bands of red andinfrared light (compare Table 2, Fig. 7) are suc-cessively absorbed by the different microbiallayers (Pierson et al., 1990). Distinct bloomsof bacteriochlorophyll b-containing anoxygenicphototrophic bacteria have been observed onlyin benthic habitats. Employing this pigment, thephototrophic Proteobacteria Blastochlorisviridis, Blastochloris sulfoviridis, Thiocapsapfennigii, Halorhodospira halochloris, Halorho-dospira abdelmalekii harvest light of a wave-
length range (1020–1035 nm), which cannot beexploited by any other photosynthetic organism.
Until recently, no strain of anoxygenic photo-synthetic bacteria was known that could absorblight in the wavelength range between 900 and1020 nm. Because of the prevalence of infraredradiation in the anoxic layers of microbialmats and the strong competition for this wave-length region, bacteria containing other types ofphotosynthetic antenna complexes would havea high selective advantage. Recently, the a-Proteobacterium Rhodospira trueperi was iso-lated, which contains bacteriochlorophyll b in alight-harvesting complex with a maximumabsorption at 986 nm (Pfennig et al., 1997).Employing a selective enrichment strategy, thea-Proteobacterium Roseospirillum parvumcould be isolated which harbors another newtype of photosynthetic antenna complex. Here,bacteriochlorophyll a is the light-harvesting pig-ment and in vivo exhibits an absorption maxi-mum at 911 nm (Glaeser and Overmann, 1999,Fig. 7). Both isolates originate from benthicmicrobial mats, indicating that the diversity ofpigment-protein complexes in Proteobacteria ishigher than previously assumed. The variation inthe in vivo absorption spectra of the same pig-ment must be the result of differences in bindingto light-harvesting proteins. In contrast, changesin the absorption spectra of the light-harvestingcomplex of green sulfur bacteria are the result ofchemical alterations (e.g., methylation) of thepigment molecules (Bobe et al., 1990) becausepigment-pigment interactions dominate in thechlorosomes (see Light Absorption and LightEnergy Transfer in Prokaryotes in this Chapter).
Because methanogenesis is the predominantpathway of terminal degradation in rice fields,Heliobacteriaceae probably compete with thephotoheterotrophic purple nonsulfur bacteria intheir natural environment (Madigan and Orm-erod, 1995). Owing to the presence of bacterio-chlorophyll g, Heliobacteriaceae take advantageof a wavelength region of the electromagneticspectrum, which is not absorbed by other pho-totrophic bacteria. As a result of the small andfixed size of the photosynthetic antenna (see LightAbsorption and Light Energy Transfer inProkaryotes in this Chapter), these bacteria areadapted to higher light intensities than otheranoxygenic phototrophic bacteria (ª1,000 molquanta·m–2·s-1).
In addition to the capacity of absorbing lightin the long wavelength range, metabolic flexibil-ity is of highly selective value for the colonizationof benthic habitats with their high fluctuationsin oxygen and sulfide concentrations (seeChemotrophic growth with O2).
However, the composition of communities ofphototrophic sulfur bacteria is not solely deter-

70 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
mined by competition. The simultanous presenceof green sulfur bacteria and Chromatiaceae pos-sibly is also based on syntrophic interactions (seeCoexistence of Phototrophic Sulfur Bacteria inthis Chapter).
Coexistence of Phototrophic BacteriaWithin the Chromatiacea the small-celled genusChromatium species exhibit a considerablygreater metabolic flexibility than the large-celledspecies (see Carbon Metabolism and Che-motrophic Growth with O2 in this Chapter). Inaddition, small-celled species like Allochroma-tium vinosum have a higher growth affinity forsulfide. Based on these pure culture data, it istherefore unexpected that large-celled forms infact dominate in natural ecosystems. The large-celled Chromatium weissei oxidizes sulfide twiceas fast as the small-celled Allochromatium vino-sum. Whereas the former preferentially oxidizessulfide to zero-valent sulfur, the latter oxidizes alarger fraction directly to sulfate. Under fluctu-ating conditions as they occur in the chemoclineof lakes, Chromatium weissei is capable of rap-idly oxidizing sulfide at the onset of illumination,thereby accumulating zero-valent sulfur. Duringthe remaining light period and because of itshigher affinity for sulfide, Allochromatium vino-sum utilizes most of the sulfide. Continuouscocultures of both species have thus beenestablished by illumination in light-dark cycles(van Gemerden, 1974).
Furthermore, stable coexistence of two organ-isms is feasible in the presence of two substratesfor which the two competitors have complemen-tary affinities. Stable syntrophic interactions canbe established in laboratory cocultures of purplesulfur (Allochromatium vinosum) and green sul-fur bacteria (Chlorobium limicola f.sp. thiosulf-atophilum; van Gemerden and Mas, 1995).Because of its higher affinity, the green sulfurbacterium oxidizes sulfide to zero-valent sulfur.The extracellular sulfur is mobilized as polysul-fide, which can be used instantaneously as elec-tron donor of the purple sulfur bacterium. Thepresence of sulfide inhibits the green sulfur bac-terium from using polysulfide (see The FamilyChlorobiaceae in Volume 7). Sulfide and polysul-fide thus are the mutual substrates for the twodifferent phototrophic sulfur bacteria.
Purple and green sulfur bacteria also have com-plementary affinities for sulfide and acetate(see Carbon metabolism). Accordingly, stablecontinuous cocultures of Chlorobium phaeo-bacteroides and Thiocapsa roseopersicina can beestablished (Veldhuis and van Gemerden, 1986).
Interactions Between Phototrophic SulfurBacteria and Chemotrophic Bacteria A con-siderable number of strains of Chromatiacae is
capable of switching to a chemolithotrophicgrowth mode after prolonged incubation inthe presence of molecular oxygen (seeChemotrophic Growth with O2 in this Chapter).Under these conditions, purple sulfur bacteriacompete with colorless sulfur bacteria like Thio-bacillus spp. Compared to thiobacilli, the purplesulfur bacterium Thiocapsa roseopersicinaattains a higher growth yield under chem-olithotrophic conditions (De Wit and vanGemerden, 1987a). However, the growth affinityfor sulfide of the colorless sulfur bacteria is up to47 times higher than that of Chromatiacae (DeWit and van Gemerden, 1987b; van Gemerdenand Mas, 1995). Therefore Chromatiacae grow-ing exclusively by chemolithotrophy would berapidly outcompeted by colorless sulfur bacteria.
Culture experiments indicate that Thiocapsaroseopersicina, a typical inhabitant of laminatedmicrobial mats in temperate environments, canreplenish its photosynthetic pigments duringanoxic periods in the dark, thereby maintaininga phototrophic growth mode also during the sub-sequent oxic light period (De Wit and vanGemerden, 1990b). Based on microelectrodemeasurements, purple sulfur bacteria in marinemicrobial mats of the North Sea barrier islandsare exposed to oxygen during most of the day,whereas anoxic conditions prevail during thenight (De Wit et al., 1989). Thus, the anoxygenicphototrophs cannot grow during the night andface competition for sulfide by colorless sulfurbacteria during the day. Because of their higheraffinity for sulfide, the latter would be expectedto outcompete phototrophically growing purplesulfur bacteria. In cocultures of Thiocapsaroseopersicina and Thiobacillus thioparus, sul-fide is indeed entirely used by the colorless sulfurbacterium in the presence of oxygen. If oxygenconcentrations are limiting, however, sulfide isoxidized incompletely by the chemolithotrophand soluble zero-valent sulfur formed (either aspolysulfide or polythionates) that in turn is usedby the purple sulfur bacterium for phototrophicgrowth (van den Ende et al., 1996). Both diurnalfluctuations between oxic light and anoxic darkperiods and syntrophism based on sulfur com-pounds may permit a stable coexistence of thesegroups and explain their simultaneous presencein natural microbial mats.
Stable associations can be established betweengreen sulfur bacteria and sulfur- or sulfate-reducing bacteria (see The Family Chlorobiaceaein Volume 7; Interactions with ChemotrophicBacteria in this Chapter). These associations arebased on a cycling of sulfur compounds but notcarbon (see Significance of Anoxygenic Photo-synthesis for the Pelagic Carbon and SulfurCycles in this Chapter). The simultaneousgrowth of both types of bacteria is fueled by the

CHAPTER 1.3 The Phototrophic Way of Life 71
oxidation of organic carbon substrates and light.In a similar manner, cocultures of Chromatiaceaewith sulfate-reducing bacteria have been estab-lished in the laboratory (van Gemerden, 1967).
The most spectacular type of associationinvolving phototrophic bacteria is represented bythe phototrophic consortia. These consortia con-sist of green sulfur bacterial epibionts that arearranged in a regular fashion around a centralchemotrophic bacterium. A rapid signal transferexists between the two partners and permits pho-totrophic consortia to scotophobotactically accu-mulate at preferred light intensities andwavelengths. In this association, the immotilegreen sulfur bacteria attain motility like purplesulfur bacteria. The high numbers of pho-totrophic consortia found in many lakes indicatethat this strategy must be of high competitivevalue under certain environmental conditions.
A commensal relationship may exist betweencoccoid epibiotic bacteria and the purple sulfurbacterium Chromatium weissei (Clarke et al.,1993). This unidentified epibionts attaches tohealthy cells but does not form lytic plaques onlawns of host cells like the morphologically sim-ilar parasite Vampirococcus (see Significance ofBacteriophages and Parasitic Bacteria in thisChapter). Possibly, the epibiont grows chemo-trophically on carbon compounds excreted bythe purple sulfur bacterium.
A syntrophic interaction between cyanobacte-ria and sulfate-reducing bacteria appears to existin microbial mats where both types of micro-organisms occur in close spatial proximity, ifnot intermixed with each other. In these ecosys-tems, the excretion of organic carbon substratesby cyanobacteria may provide the electron-donating substrates for sulfate-reducing bacteria(Jørgensen and Cohen, 1977; Skyring and Bauld,1990; Fründ and Cohen, 1992). The glycolateproduced by photorespiration (Fründ andCohen, 1992), as well as the formate, acetate andethanol produced by glycogen fermentation(Moezelaar and Stal, 1994) most likely are thesubstrates excreted by cyanobacteria.
Despite a pronounced limitation of sulfatereduction by carbon substrates (Overmann et al.,1996; Overmann, 1997), no close syntrophic rela-tionship was found between purple sulfur andsulfate-reducing bacteria in a meromictic lake. Inthis specific environment degradation of biomassby the entire anaerobic food chain rather thanexcretion of small carbon molecules and theirdirect utilization by sulfate-reducing bacteriaprovides the electron-donating substrates forsulfate-reducing and sulfur-reducing bacteria.
Symbioses Between Phototrophic Bacteriaand Eukaryotes Only one example is knownfor an intracellular symbiosis of anoxygenic pho-
totrophic bacteria with an eukaryotic organism.The ciliate Strombidium purpureum inhabits thephotic zone of sulfide-containing marine sandsand harbors 200–700 purple endosymbionts.Symbionts are arranged along the periphery ofthe host cell and contain intracellular tubular orvesicular membranes, bacteriochlorophyll a andspirilloxanthin (Fenchel and Bernard, 1993a;Fenchel and Bernard, 1993b). The ciliate showsa photosensory behavior, accumulating at wave-length that corresponds to the absorption max-ima of the endosymbionts. It has been suggestedthat the intracytoplasmic purple bacteriaincrease the efficiency of the fermentative hostby using its end products for anoxygenic photo-synthesis. Furthermore, respiration of the bacte-ria may protect the host against oxygen toxicity.
In the course of evolution, Cyanobacteria haveentered into symbiotic associations with a multi-tude of organisms (Schenk, 1992). Besides alleukaryotic phototrophs, from microalgae toSequoia sempervirens, which have intracellularcyanobacterial symbioses, the most commonextracellular symbioses of nonheterocystouscyanobacteria are in the form of cyanolichensand involve the unicellular genera Chroococcid-iopsis, Gloeocapsa, “Chroococcus,” and Gloeoth-ece, as well as members of the genera Nostoc,Calothrix, Scytonema, Stigonema, and Fischer-ella as photobionts. Heterocystous cyanobacte-ria in the genus Nostoc form extracellularsymbioses with liverworts and hig her plants(Cycads, duckweed). Anabaena enters in symbi-osis with water ferns of the genus Azolla.Prochloron strains, large-celled Synechocystisand small-celled Acaryochloris marina areknown from extracellular symbioses with ascidi-ans in tropical or subtropical marine waters;Prochloron is found as ectosymbiont on themarine didemnid ascidian Lissoclinum patella(Lewin and Withers, 1975). Extracellular symbi-oses of the Pseudanabaena-like “Konvophoron”occur in Mediterranean invertebrates. Finally,intracellular symbioses of nonheterocystouscyanobacteria are known with tropical sponges(“Aphanocapsa”, Oscillatoria, Synechocystis,Proc hloron), with green algae (Phormidium)and dinoflagellates (unidentified). Heterocys-tous cyanobacteria occur intracellularly in oce-anic diatoms of the genera Hemiaulus andRhizosolenia (and the cyanobacterium Richeliaintracellularis). The cyanobacterial symbiontconsists of a short cell filament with a terminalheterocyst (Mague et al., 1977). The numbers offilaments varies with host species. Nostoc thrivesintracellularly in Trifolium (clover) and also inthe terrestrial non-lichenic fungus Geosiphonpyriforme. With the notable exception of lichenicphotobionts, many symbiotic cyanobacteria haveresisted cultivation in spite of continued efforts.

72 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
Significance of Bacteriophages and ParasiticBacteria In addition to grazing, light and nutri-ent limitation, cyanophage infection of cyano-bacteria may be a significant factor limitingprimary productivity in the marine environment.However, because of inactivation by solar radia-tion and resistence of the host cells, the role ofcyanophages has remained unclear (Bergh et al.,1989; Proctor and Fuhrman, 1990; Suttle et al.,1990; Suttle et al., 1993; Waterbury and Valois,1993).
Several bacteria have been discovered thatattack phototrophic bacteria (Guerrero et al.,1986; Nogales et al., 1997). Vampirococcusattaches to the cell surface of Chromatium spp.where it divides, forming chains of up to threecells. Concomitantly, the cytoplasm of the hostcell appears to be degraded. Daptobacter pene-trates the cell envelope and divides intracel-lularly by binary fission. In contrast toVampirococcus, Daptobacter has been cultivatedin the absence of the host and grows by fermen-tative metabolism. Bdellovibrio has a broad hostrange, and under laboratory conditions attacksalso purple sulfur bacteria. Bdellovibrio formsdaughter cells by multiple division in theperiplasmic space of the host cell. The Gram-negative chemotrophic bacterium Stenotroph-omonas maltophilia is a non-obligatory parasiteof green sulfur bacteria, which causes cell lysisand ghost formation (Nogales et al., 1997). Itshost range is not limited to green sulfur bacteria.The presence of parasitic bacteria in water sam-ples becomes evident by the formation of lyticplaques on lawns of host bacteria (Esteve et al.,1992; Nogales et al., 1997). Up to 94% of the cellsof phototrophic sulfur bacteria may be infectedby parasitic bacteria in natural samples. Sinceinfection is largely limited to nongrowing cells,the impact of parasitism on populations of pho-totrophic sulfur bacteria appears to be limited(van Gemerden and Mas, 1995).
Evolutionary ConsiderationsPorphyrins are found in all organisms fromarchaebacteria through plants to animals, andare indispensable as prosthetic groups for energyconservation. In contrast, the partially reducedderivates of porphyrins, the (bacterio)chloro-phylls, are synthesized by members of only a fewbacterial divisions (Fig. 1). This indicates that thecapability for synthesis of porphyrins is a veryancient trait, whereas only a few prokaryotesacquired the capability to form photosyntheticpigments. Photosynthesis requires the presenceof various complex protein structures and cofac-tors, and thus the expression of a large numberof different genes (see Photosynthetic GeneCluster in this Chapter). Previously, it had there-
fore appeared justified to consider all pho-totrophic prokaryotes as a monophyletic grouponly distantly related to nonphototrophic bacte-ria (Pfennig and Trüper, 1974; Trüper and Pfen-nig, 1978). Two lines of evidence have been usedto recon struct the evolution of photosynthesis.
Fossil Evidence The oldest fossils of micro-organisms have been dated back to the earlyArchaean (3.8 billion years ago) and may repre-sent remains of cyanobacteria (Awramik, 1992).They consist of chemical fossils and stromatolitesthat have been detected especially in sedimen-tary rocks of the Pilbara region, Western Austra-lia, and the Barberton Mountain Land, SouthAfrica. Stromatolites are laminated convexdomes and columns of cm to dm size and havebeen found in 3.5 to 0.8 billion year old rocks.Although scarce in biosynthetic molecular skel-etons, the insoluble, high-molecular-weightorganic matter (kerogen) contains isotopic evi-dence for autotrophic carbon fixation. The ratioof stable carbon isotopes (d13C values) are inthe range of –35.4 to –30.8!/oo, which is typicalfor CO2-carbon fixed by the ribulose-1,5-bisphosphate cycle (Hayes et al., 1983). In addi-tion, the se ancient sediments contain laminateddomes and columns of cm to dm size, which inanalogy to extant stromatolites have been inter-preted as organosedimentary structures pro-duced by the trapping, binding, and precipitationactivity of filamentous microorganisms, mostlikely cyanobacteria.
Alternatively, it has been proposed that anox-ygenic photosynthetic bacteria and not theoxygenic cyanobacteria formed the oldeststromatolites. Based on the phylogenetic analysisof the 16S rRNA gene sequence (Oyaizu et al.,1987) and the ecophysiology (Ward et al., 1989)of the filamentous green photosynthetic bacte-rium Chloroflexus aurantiacus, similar anoxy-genic phototrophic bacteria may be the morelikely candidate microorganisms that built themost ancient stromatolites. However, accordingto analyses of the nucleotide sequences of itsreaction center polypeptides and primary sigmafactor (see Molecular Evidence in this Chapter),Chloroflexus aurantiacus does not represent adeep branch of bacterial evolution. Gypsum lay-ers within the supposed stromatolites have beeninterpreted as indicators of sulfide oxidation byeither anoxygenic phototrophs or colorlesssulfur-oxidizing bacteria (Awramik, 1992). How-ever, similar structures have been discovered inlacustrine, and thus sulfur-depleted, settings withlittle input of allochthonous organic carbon(Buick, 1992). Therefore, at least some 2.7 billionyear-old stromatolites are more likely tohave harbored oxygenic cyanobacteria. Takentogether with the fossil evidence, this would indi-

CHAPTER 1.3 The Phototrophic Way of Life 73
cate that diversification of the major groups ofphototrophic microorganisms did occur duringthe early Archaean (Awramik, 1992).
Because of the indefinite character of the fossilevidence, 16S rRNA sequences and componentsof the photosynthetic apparatus of the differentphotosynthetic prokaryotes have been used togain additional insight into the evolution ofphotosynthesis.
Molecular Evidence Chlorophyll-based pho-tosystems are only found in the Bacteria andchloroplasts, suggesting that this type of energyconversion originated in the bacterial lineageafter the divergence of Archaea and Eukarya. Sofar, photosynthetic species have not been discov-ered in the very early lineages of the bacterialradiation (e.g., the thermophilic oxygen reducersand Thermotogales; Fig. 1). Because most speciesof these lineages are chemolithotrophic, it hasbeen proposed that chemolithoautotrophy pre-ceded phototrophy during the evolution of theBacteria (Pace, 1997). This conclusion is sup-ported by the fact that in phylogenetic treesbased on protein sequences of elongation factorEF-Tu and the b-subunit of ATP synthase, onlythe Aquificales and Thermotogales branchdeeper than the majority of the bacterial divi-sions, while the Chloroflexus subdivision doesnot (Stackebrandt et al., 1996), thus indicatingthat Chloroflexus does not represent the descen-dant of a more ancient ancestor than other pho-totrophic bacteria.
At present, five of the known bacterial lin-eages comprise phototrophic species (Fig. 1, seeTaxonomy of Phototrophy among Prokaryotesin this Chapter). Based on 16S rRNA sequences,extant phototrophic species of different lineagesare only very distantly related to each other. Fur-thermore, one lineage, the Chloroflexus sub-group, represents an early branch in theevolution of the Bacteria. Given the complexityof the photosynthetic apparatus, it is unlikelythat photosynthesis has evolved more than onceduring the evolution of the domain Bacteria(Woese, 1987). The phylogenetic analysis indi-cates that either an early ancestor of most knownbacteria had acquired the capacity for photosyn-thetic growth (Stackebrandt et al., 1988) or,alternatively, that the genes coding the photosyn-thetic apparatus were transferred laterallybetween phylogenetically distant bacteria. Theevidence for the various scenarios of the evolu-tion of bacterial photosynthesis is discussed inthe present section.
Originally, it had been proposed (Oparin,1938; Gest and Schopf, 1983a) that anaerobic,heterotrophic prokaryotes capable of fermentinghexose sugars were among the earliest life formsand that electron transport and photosynthesis
evolved as a response to the depletion of organicnutrients from the primordial soup. Based onone hypothesis (the Granick hypothesis; Gran-ick, 1965), the biosynthetic pathway of photosyn-thesis pigment molecules may be taken as arecapitulation of evolution such that compoundswith shorter biosynthetic pathways reflect themore ancestral state. The synthesis of bacterio-chlorophyll requires one additional enzymaticreduction than that of chlorophyll. Because chlo-rophyll precedes bacteriochlorophyll in the bio-synthetic pathway, the former should haveexisted earlier in nature. It has been proposed(Pierson and Olson, 1989) that a non-oxygenicphotosynthetic ancestor containing chlorophyll aand the two types of reaction centers evolvedprior to the major radiation event of the Bacte-ria. During the subsequent radiation, oxygenevolution appeared in one line of descentwhereas either the quinone or the FeS-type pho-tosystem was lost in other lineages, concomitantwith the emergence of the different bacteriochlo-rophylls. Besides avoiding an a priori lateral genetransfer of the complete photosynthetic genecluster, this Pierson-Olson hypothesis takes intoaccount the ecological conditions of the earlybiosphere in which the absence of oxygen andozone caused a predominance of radiation in theblue and UV wavelength range, which in turnwould render the red-shifted absorption maximaof bacteriochlorophylls of little selective advan-tage (Boxer, 1992).
As an argument against the Granick andPierson-Olson hypotheses, several types of pho-totrophic bacteria that would be expected areapparently missing in nature. As an example,anoxygenic chlorophyll-containing forms havenever been found, although it has been arguedthat the 8-hydroxychlorophyll-containing Helio-bacteriaceae represents this type inasmuch asbacteriochlorophyll g is easily converted to chlo-rophyll a by oxidation. Bacteriochlorophyllsoccur in both types of reaction centers, the pheo-phytin-type (Proteobacteria, Chloroflexus) andthe FeS-type. This could indicate that the pres-ence of bacteriochlorophyll represents a primi-tive trait. The chlorophyll-first hypothesispostulates that bacteriochlorophyll has replacedchlorophyll independently in at least three dif-ferent bacterial lineages. Chlorophyll, however,is presently only found in oxygen-evolvingorganisms of the phylum Cyanobacteria whic h,based on 16S rRNA sequence comparison,represents the most recently evolved group ofphototrophic bacteria (Woese, 1987, Fig. 1).Cyanobacteria contain two different photosys-tems and thus have the most complex photosyn-thetic apparatus. In addition, the much highercomplexity of the oxygen-evolving PSII of oxy-genic phototrophic organisms may imply that it

74 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
appeared later than the other photosystems dur-ing evolution.
As another argument against the Pierson-Olson hypothesis, chlorophyll itself should havebeen of little selective advantage in Earth’s earlybiosphere and it has been proposed thatquinone-iron complexes represented the firstphotosynthetic unit (Boxer, 1992). In contrast tothe complex porphyrin pigments, quinones canform spontaneously from acetyl thioesters (Hart-mann, 1992). Furthermore, the discrepancybetween the presence of chlorophyll exclusivelyin the most highly evolved bacteria and itsshorter biosynthetic pathway may be explainedby the finding that the chlorin reductase, whichcatalyzes the additional step of the biosyntheticpathway for bacteriochlorophyll, is phylogeneti-cally older than the enzyme (protochlorophyl-lide reductase) that catalyzes the preceedingstep. This enzyme is present in both the chloro-phyll- and bacteriochlorophyll-containing bacte-ria (Burke et al., 1993). An ancien t reductasemay have been able to perform both, the reduc-tion of protochlorophyllide and of chlorin, suchthat bacteriochlorophyll was the photochemi-cally active pigment in the last common ancestorof all extant phototrophic bacteria.
An analysis of the distribution of the differenttypes of reaction centers among the differentbacterial phyla and the amino acid sequences ofreaction center proteins (Blankenship, 1992)provides an alternative hypothesis for the evolu-tion of photosynthesis, namely the possibility oflateral transfer of photosynthesis genes. Both thepheophytin/quinone and the FeS-type reactioncenters are found in phylogenetically distantgroups (e.g., a pheophytin/quinone reaction cen-ter in Chloroflexus and phototrophic members ofthe a-Proteobacteria). Even more significantly,a phylogenetic analysis of the amino acidsequences of pheophytin-type reaction centerpolypeptides from the three different bacteriallineages Chloroflexaceae, cyanobacteria and a-Proteobacteria indicated that the reaction centerof Chloroflexus aurantiacus is more closelyrelated to that of phototrophic members of thea-Proteobacteria than to the PSII reaction cen-ter of cyanobacteria (Blankenship, 1992). Thusthe reaction center of Chloroflexus must haveevolved after (and not prior to) the divergenceof the D1/D2 branch from the L/M line ofdescent. Another essential component of thephotosynthetic apparatus of Chloroflexus andgreen sulfur bacteria are the light-harvestingchlorosomes. Based on amino acid sequencecomparison of protein constituents, chlorosomesof both groups have a common evolutionary ori-gin (Wagner-Huber et al., 1988). Similarly, acomparison of the amino acid sequences of thegroup 1 s70 primary sigma factor also has dem-
onstrated a close relationship to the green sulfurbacteria with respect to this component ofthe central housekeeping function (Gruber andBryant, 1998). Other features of Chloroflexusaurantiacus appear to be unique (like the lipidand carotenoid composition), or ancient (like thehydroxypropionate pathway of CO2-fixation).Recently, the activity of the key enzymes of thispathway have been reported for some archaea(Menendez et al., 1999) such that Chloroflexusaurantiacus seems to represent a “chimeric”organism.
Based on the most parsimonious assumptionthat homodimeric reaction centers are ancestralto homodimeric ones, the reaction centers ofgreen sulfur bacteria and Heliobacteriaceaewould resemble most the reaction center of theancestor of all extant bacteria. It has beenhypothesized (Gruber and Bryant, 1998) that thereaction center of Chloroflexus aurantiacus wasacquired by a recent lateral gene transfer eventthat may have replaced a type I reaction centerwith a type II (FeS) reaction center, whereasother features like primary sigma factor or chlo-rosomes still reflect the common descent ofChloroflexus and the green sulfur bacteria.Alternatively, it has been suggested that transferof the genetic information of the relatively sim-ple chlorosomes occurred after the evolution ofthe two classes of reaction centers and that thegreen sulfur bacteria represent a relatively mod-ern evolutionary invention (Stackebrandt et al.,1996).
The presence of two homologous polypeptidesin all known reaction centers would suggest asingle gene duplication event in an early ancestorof all phototrophic bacteria. As an additionalresult of the phylogenetic analysis of the aminoacid sequences of pheophytin-type reaction cen-ter polypeptides from the three different bacte-rial lineages (Chloroflexaceae, cyanobacteria anda-Proteobacteria; Blankenship, 1992), the mostlikely occurrence of two independent gene dupli-cations is suggested—one leading to the reactioncenter of PSII in cyanobacteria and green plants(polypeptides D1 and D2) and another to thereaction center of Chloroflexus and purple non-sulfur bacteria (polypeptides L and M). Another,third, independent gene duplication has to beassumed during the evolution of the FeS-typereaction center. The reason for the paraphyleticdevelopment of the three lineages may be a func-tional advan tage of dimeric reaction centersover monomeric ones.
Yet another evolutionary scenario for photo-synthetic reaction centers (Vermaas, 1994) hasbeen based on the finding that the sixth mem-brane-spanning region of the heliobacterial(FeS- or PSI-type) reaction center shows a greatsimilarity to the sixth membrane-spanning

CHAPTER 1.3 The Phototrophic Way of Life 75
region of the CP47 antenna polypeptide of (thequinone-type) PSII, and the preceeding N-terminal five hydrophobic regions still show sig-nificantly greater similarity to CP47 (and toanother PSII antenna protein, CP43) than to therespective portion of PSI. According to thismodel, an ancestral homodimeric antenna/reac-tion center complex comprised 11 putative trans-membrane regions and contained two quinonesand an Fx-type Fe4S4 iron-sulfur center. Rela-tively few modifications may have led to thehomodimeric complex of green sulfur bacteriaand Heliobacteriaceae, whereas a gene duplica-tion event and divergent evolution led to theheterodimeric PSI. As a para llel line of descent,splitting of the ancestral reaction center complexinto a reaction center and a separate antennaprotein may have occurred. Operon duplication,loss of the FeS, and divergent evolution areassumed to have resulted in two separate lin-eages. By association with an additional water-splitting enzyme system, PSII was formed. Incontrast, the separate antenna polypeptide waslost and replaced by a modified antenna complex(light-harvesting I) during evolution of the reac-tion center of Proteobacteria and Chloroflexus.Significantly, however, this theory does notexplain the occurrence of the quinone-type reac-tion center in these latter two groups, which arephylogenetically very distant. In addition, thecombination of a reaction center typical for Pro-teobacteria with an antenna structure character-istic for green sulfur bacteria would still need tobe explained by lateral gene transfer of either ofthe two components.
Based on the obvious discrepancy between thephylogeny of ribosomal RNA and reaction cen-ter proteins, the hypothesis of lateral transfer ofphotosynthesis genes between distantly relatedgroups of bacteria has been put forward. Lateralgene transfer as yet seems to provide the sim-plest explanation for the distribution pattern ofphotosynthesis genes within the bacterial radia-tion (Blankenship, 1992; Nagashima et al., 1993;Nagashima et al., 1997). Such a lateral genetransfer would encompass reaction center struc-tural genes, genes coding for other electrontransfer proteins, and genes needed for the bio-synthesis of pigments and cofactors. In purplenonsulfur bacteria the majority of these genesindeed form a single cluster of 46 kb (which doesnot encompass the genes for the light-harvestingII complex, however; Bauer and Bird, 1996;Wellington et al., 1992; Yildiz et al., 1992). Thegenetic organization may be taken as evidencefor lateral gene transfer as the cluster representsonly ~1.3% of the total genome size. It should bementioned, however, that clustering of mostphotosynthesis genes may also be due to struc-tural or regulatory constraints. Supporting the
latter argument (Yildiz et al., 1992), photosyn-thesis genes in a-Proteobacteria are transcrip-tionally coupled in superoperons involvingoverlapping transcripts. The particular geneticorganization is the prerequisite for adaptation ofthe cells to changing light intensity (see GeneticRegulation in Response to Light in this Chapter)and oxygen tension (see Genetic Regulation byO2 in this Chapter). Therefore a selective pres-sure may exist to retain the linkage order andwould make the genetic organization of the pho-tosynthesis genes less suitable for phylogeneticinference. Furthe rmore, the high correlationbetween the phylogenetic trees for 16S rRNAand cytochrome c in phototrophic members ofthe a-Proteobacteria has been taken as evidencethat a lateral transfer of photosynthesis genes didnot occur at least within this phylogenetic group(Woese et al., 1980). Thus, the presence of reac-tion centers in aerobic bacteriochlorophyll-containing a-Proteobacteria may represent anatavistic trait, and the genes coding the reactioncenter might have been lost frequently duringthe evolution of aerobic representatives in thisgroup (Stackebrandt et al., 1996).
Because the pigment composition of the oxy-genic photosynthetic “Prochlorophytes” is verysimilar to that of green plant chloroplasts, andlike the latter “Prochlorophytes” have appressedthylakoid membranes, it has been proposed thatthe chloroplasts of green plants evolved from anendosymbiotic “prochlorophyte” (van Valen andMaiorana, 1980; Lewin, 1981). In contrast to theother oxygenic phototrophs, Prochlorococcuscontains divinyl isomers of chlorophylls a and b,and a- instead of b-carotene (Chrisholm et al.,1992; Goericke and Repeta, 1992). However,based on sequence comparison of 16S rRNA(Urbach et al., 1992) and the rpoC1 (Palenik andHaselkorn, 1992) genes, the three knownprochlorophyte lineages (Prochloron, Prochlo-rothrix, and Prochlorococcus) are no directancestors of chloroplasts. In addition, these anal-yses revealed that “Prochlorophytes” most likelyare of polyphyletic origin and that the use ofchlorophyll b as additional light-harvesting pig-ments must have developed at least four timesduring evolution. In this case, too, a horizontaltransfer of the respective biosynthesis genescould be invoked to explain the distribution pat-tern of chlorophyll b among the different mem-bers of the cyanobacterial division (Palenik andHaselkorn, 1992). Immunological studies anddifferences in the chlorophyll a/chlorophyll bratio of the antennae isolated from different“Prochlorophytes” indicate that the capacity tobind chlorophyll b arose several times and inde-pendently from the cyanobacterial ancestors,and thus confirm the results of s equence com-parisons of the 16S rRNA and rpoC1 genes.

76 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
Literature CitedAlberti, M., Burke, D. H., Hearst, J. E. 1995. Structure and
sequence of the photosynthesis gene cluster. In: Blan-kenship RE, Madigan MT, Bauer CE (Eds.) Anoxygenicphotosynthetic bacteria. Kluwer Academic Publishers.Dordrecht, Boston, London, 1083–1106.
Amesz, J. 1995. The antenna-reaction center complex ofHeliobacteria. In: Blankenship RE, Madigan MT, BauerCE (Eds.) Anoxygenic Photosynthetic Bacteria. KluwerAcademic Publishers. Dordrecht, The Netherlands. 687–697.
Anagnostidis, K., Komárek, J. 1988. Modern approach to theclassification system of the cyanophytes. 3. Oscillatori-ales. Arch Hydrobiol Suppl 80 Algol. Studies 53:327–472.
Angerhofer, A., Cogdell, R. J., Hipkins, M. F. 1986. A spectralcharacterization of the light-harvesting pigment-proteincomplexes from Rhodopseudomonas acidophila. Bio-chim Biohys Acta 848:333–341.
Armitage, J. P., Kelly, D. J., Sockett, R. E. 1995. Flagellatemotility, behavioral responses and active transport inpurple non-sulfur bacteria. In: Blankenship RE, Madi-gan MT, Bauer CE (Eds.) Anoxygenic PhotosyntheticBacteria. Kluwer Academic Publishers. Dordrecht, TheNetherlands. 1005–1028.
Armitage, J. P. 1997. Behavioral resposnes of bacteria to lightand oxygen. Arch Microbiol 168:249–261.
Arnheim, K., Oelze, J. 1983. Differences in the control ofbacteriochlorophyll formation by light and oxygen.Arch Microbiol 135:299–304.
Awramik, S. M. 1992. The oldest records of photosynthesis.Photosynth Res 33:75–89.
Barry, B. A., Boerner, R. J., de Paula, J. C. 1994. The use ofcyanobacteria in the study of the structure and functionof photosystem II. In: Bryant DA (Eds.) The molecularbiology of cyanobacteria. Kluwer Academic Publishers.Dordrecht, Boston, London, 217–257.
Bauer, C. E., Bollivar, D. W., Suzuki, J. Y. 1993. Geneticanalyses of photopigment biosynthesis in eubacteria: aguiding light for algae and plants. J Bacteriol 175:3919–3925.
Bauer, C. E. 1995. Regulation of photosynthesis gene expres-sion. In: Blankenship RE, Madigan MT, Bauer CE(Eds.) Anoxygenic Photosynthetic Bacteria. KluwerAcademic Publishers. Dordrecht, The Netherlands.1221–1234.
Bauer, C. E., Bird, T. H. 1996. Regulatory circuits controllingphotosynthesis gene expression. Cell 85:5–8.
Beale, S. I. 1995. Biosynthesis and structures of porphyrinsand hemes. In: Blankenship RE, Madigan MT, BauerCE (Eds.) Anoxygenic Photosynthetic Bacteria. KluwerAcademic Publishers. Dordrecht, The Netherlands. 153–177.
Bebout, B. M., Garcia-Pichel, F. 1995. UVB-induced verticalmigrations of cyanobacteria in a microbial mat. ApplEnvironm Microbiol 61:4215–4222.
Beck, H., Hegeman, G. D., White, D. 1990. Fatty acid andlipopolysaccharide analyses of three Heliobacteriumspp. FEMS Microbiol Lett 69:229–232.
Beer-Romero, P., Favinger, J. L., Gest, H. 1988. Distinctiveproperties of bacilliform photosynthetic heliobacteria.FEMS Microbiol Lett 49:451–454.
Bergh, Ø., Børsheim, K. Y., Bratbak, G., Heldal, M. 1989.High abundance of viruses found in aquatic environ-ments. Nature 340:467–468.
Biebl, H., Drews, G. 1969. Das in vivo Spektrum als taxono-misches Merkmal bei Untersuchungen zur Verbreitungder Athiorhodaceae. Zbl Bakt Abt II 123:425–452.
Biedermann, M., Drews, G., Marx, R., Schröder, J. 1967. DerEinfluß des Sauerstoffpartialdruckes und der Antibiot-ica Actinomycin und Puromycin auf das Wachstum, dieSynthese von Bacteriochlorophyll und die Thylakoid-morphogenese in Dunkelkulturen von Rhodospirillumrubrum. Arch Mikrobiol 56:133–147.
Biel, A. J. 1986. Control of bacteriochlorophyll accumulationby light in Rhodobacter capsulatus. J Bacteriol 168:655–659.
Biel, A. J. 1995. Genetic analysis and regulation of bacterio-chlorophyll biosynthesis. In: Blankenship RE, MadiganMT, Bauer CE (Eds.) Anoxygenic Photosynthetic Bac-teria. Kluwer Academic Publishers. Dordrecht, TheNetherlands. 1125–1134.
Blankenship, R. E. 1992. Origin and early evolution of pho-tosynthesis. Photosynth Res 33:91–111.
Blankenship, R. E., Olson, J. M., Miller, M. 1995. Antennacomplexes from green photosynthetic bacteria. In: Blan-kenship RE, Madigan MT, Bauer CE (Eds.) Anoxygenicphotosynthetic bacteria. Kluwer Academic Publishers.Dordrecht, The Netherlands. 399–435.
Bobe, F. W., Pfennig, N., Swanson, K. L., Smith, K. M. 1990.Red shift of absorption maxima in Chlorobiaceaethrough enzymic methylation of their antenna bacterio-chlorophylls. Biochemistry 29:4340–4348.
Bowyer, J. R., Hunter, C. N., Ohnishi, T., Niederman, R. A.1985. Photosynthetic membrane development inRhodopseudomonas sphaeroides. J Biol Chem260:3295–3304.
Boxer, S. G. 1992. Some speculations concerning the evolu-tion of photosynthetic function. Photosynth Res 33:113–119.
Brockmann, H., Lipinski, A. 1983. Bacteriochlorophyll g. Anew bacteriochlorophyll from Heliobacterium chlorum.Arch Microbiol 136:17–19.
Brune, D. C. 1989. Sulfur oxidation by phototrophic bacteria.Biochim Biophys Acta 975:189–221.
Bryant, D. A. 1982. Phycoerythrocyanin and phycoerythrin:properties and occurrence in cyanobacteria. J GenMicrobiol 128:835–844.
Büdel, B., Karsten, U., Garcia-Pichel, F. 1997. Ultraviolet-absorbing scytonemin and mycosporine-like amino acidsin exposed. rock-inhabiting cyanobacterial lichens.Oecologia 112:165–172.
Buick, R. 1992. The antiquity of oxygenic photosynthesis:evidence from stromatolites in sulphate-deficientArchaean lakes. Science 255:74–77.
Burke, D. H., Hearst, J. E., Sidow, A. 1993. Early evolutionof photosynthesis:Clues from nitrogenase and chloro-phyll iron proteins. Proc Natl Acad Sci USA 90:7134–7138.
Caldwell, D. E., Tiedje, J. M. 1975. The structure of anaerobicbacterial communities in the hypolimnia of severalMichigan lakes. Can J Microbiol 21:377–385.
Camacho, A., Garcia-Pichel, F., Vicente, E., Castenholz,R. W. 1996. Adaptation to sulfide and to the underwaterlight field in three cyanobacterial isolates from lakeArcas (Spain). FEMS Microbiol Ecol 21:293–301.
Cameron, R. E. 1966. Desert Algae: soil crusts and diapha-nous substrata as algal habitats. Jet Propulsion Lab.Tech. Rep. 32–971:1–41.
Campbell, L., Vaulot, D. 1993. Photosynthetic picoplanktoncommunity structure in the subtropical North Pacific

CHAPTER 1.3 The Phototrophic Way of Life 77
Ocean near Hawaii (station ALOHA). Deep Sea Res40:2043–2060.
Campbell, L., Nolla, H. A., Vaulot, D. 1994. The importanceof Prochlorococcus to community structure in the cen-tral North Pacific Ocean. Limnol Oceanogr 39:954–961.
Capone, D. G., Zehr, J. P., Paerl, H. W., Bergman, B., Car-penter, E. J. 1977. Trichodesmium, a globally significantcyanobacterium. Science 276:1221–1229.
Carr, N. G., Mann, N. H. 1994. The oceanic cyanobacterialpicoplankton. In: Bryant DA (Eds.) The Molecular Biol-ogy of Cyanobacteria. Kluwer Academic Publishers.Dordrecht, Boston, London, 27–48.
Castenholz, R. W. 1982. Motility and Taxes. In: Carr NG,Whitton BA (Eds.) The biology of cyanobacteria. Black-well Scientific Publications. Oxford, 413–440.
Castenholz, R. W., Utkilen, H. C. 1984. Physiology of sulfidetolerance in a thermophilic Oscillatoria. Arch Microbiol138:299–305.
Castenholz, R. W. 1989. Oxygenic Photosynthetic Bacteria.In: Staley JT, Bryant MP, Pfennig N, Holt JG (Eds.)Bergey’s Manual of Systematic Bacteriology. Williams& Wilkins. Baltimore, 1710–1806.
Castenholz, R. W., Bauld, J., Jørgensen, B. B. 1990. Anoxy-genic microbial mats of hot springs: thermophilic Chlo-robium sp. FEMS Microbio Ecol 74:325–336.
Castenholz, R. W. 1992. Species usage, concept and evolutionin the cyanobacteria (Blue-green algae). J Phycol28:735–745.
Castenholz, R. W., Pierson, B. K. 1995. Ecology of thermo-philic anoxygenic phototrophs. In: Blankenship RE,Madigan MT, Bauer CE (Eds.) Anoxygenic Photosyn-thetic Bacteria. Kluwer Academic Publishers. Dor-drecht, The Netherlands. 87–103.
Castenholz, R. W., Garcia-Pichel, F. 1999. Cyanobacterialresponses to UV-radiation. In: Whitton BA, Potts M(Eds.) Ecology of cyanobacteria: their diversity in timeand space. Kluwer Academic Publications. Dordrecht,704.
Chisholm, S. W., Olson, R. J., Zettler, E. R., Goericke, R.,Waterbury, J. B., Welschmeyer, N. A. 1988. A novel free-living prochlorophyte abundant in the oceanic euphoticzone. Nature 334:340–343.
Chisholm, S. W., Frankel, S. L., Goericke, R., Olson, R. J.,Palenik, B., Waterbury, J. B., West-Johnsrud, L., Zet-tler, E. R. 1992. Prochlorococcus marinus nov. gen.nov. sp.: an oxyphototrophic marine prokaryote con-taining divinyl chlorophyll a and b. Arch Microbiol157:297–300.
Clarke, K. J., Finlay, B. J., Vicente, E., Lloréns, H., Miracle,M. R. 1993. The complex life-cycle of a polymorphicprokaryote epibiont of the photosynthetic bacteriumChromatium weissei. Arch Microbiol 159:498–505.
Cohen, Y., Jørgensen, B. B., Padan, E., Shilo, M. 1975.Sulphide-dependent anoxygenic photosynthesis in thecyanobacterium Oscillatoria limnetica. Nature 257:489–492.
Cohen, Y., Jørgensen, B. B., Revsbech, N. P., Poplawski, R.1986. Adaptation to hydrogen sulfide of oxygenic andanoxygenic photosynthesis among cyanobacteria. Appl.Environ. Microbiol 51:398–407.
Coolen, M. J. L., Overmann, J. 1998. Analysis of subfossilmolecular remains of purple sulfur bacteria in a lakesediment. Appl Environ Microbiol 64:4513–4521.
De Wit, R., van Gemerden, H. 1987a. Chemolithotrophicgrowth of the phototrophic sulfur bacterium Thiocapsaroseopersicina. FEMS Microbiol Ecol 45:117–126.
De Wit, R., van Gemerden, H. 1987b. Oxidation of sulfideto thiosulfate by Microcholeus chtonoplastes. FEMSMicrobiol Ecol 45:7–13.
De Wit, R., van Boekel, W. H. M., van Gemerden, H. 1988.Growth of the cyanobacterium Microcoleus chtonoplas-tes on sulfide. FEMS Microbiol Ecol 53:203–209.
De Wit, R., Jonkers, H. M., van den Ende, F. P., van Gemer-den, H. 1989. In situ fluctuations of oxygen and sulphidein marine microbial sediment ecosystems. Neth J SeaRes 23:271–281.
De Wit, R., van Gemerden, H. 1990a. Growth and metabo-lism of the purple sulfur bacterium Thiocapsa roseoper-sicina under combined light/dark and oxic/anoxicregimens. Arch Microbiol 154:459–464.
De Wit, R., van Gemerden, H. 1990b. Growth of the pho-totrophic purple sulfur bacterium Thiocapsa roseoper-sicina under oxic/anoxic regimens in the light. FEMSMicrobiol Ecol 73:69–76.
Dierstein, R. 1984. Synthesis of pigment-binding protein intoluene-treated Rhodopseudomonas capsulata and incell-free systems. Eur J Biochem 138:509–518.
Dietrich, G., Kalle, K., Krauss, W., Siedler, G. 1975. Allge-meine Meereskunde. Gebrüder Bornträger. Berlin,Stuttgart, 593.
Drews, G., Golecki, J. R. 1995. Structure, molecular organi-zation, and biosynthesis of membranes of purple bacte-ria. In: Blankenship RE, Madigan MT, Bauer CE (Eds.)Anoxygenic Photosynthetic Bacteria. Kluwer AcademicPublishers. Dordrecht, The Netherlands. 231–257.
Dubinina, G. A., Gorlenko, V. M. 1975. New filamentousphotosynthetic green bacteria containing gas vacuoles.Mikrobiologiya 44:511–517.
Eaton, J. W., Moss, B. 1966. The estimation of numbers andpigment content in epipelic algal populations. LimnolOcean 11:584–594.
Ehling-Schulz, M., Bilger, W., Scherer, S. 1997. UVB inducedsynthesis of photoprotective pigments and extracellularpolysaccharides in the terrestrial cyanobacterium Nos-toc commune. J Bacteriol 179:1940–1945.
Eichler, B., Pfennig, N. 1986. Characterization of a new plate-let-forming purple sulfur bacterium Amoebobacterpedioformis sp. nov. Arch Microbiol 146:295–300.
Eisenreich, W., Strauß, G., Werz, U., Fuchs, G., Bacher, A.1993. Retrobiosynthetic analysis of carbon fixation inthe phototrophic eubacterium Chloroflexus aurantiacus.Eur J Biochem 215:619–632.
Esteve, I., Gaju, N., Mir, J., Guerrero, R. 1992. Comparisonof techniques to determine the abundance of predatorybacteria attacking Chromatiaceae. FEMS MicrobiolEcol 86:205–211.
Evans, W. R., Fleischman, D. E., Calvert, H. E., Pyati, P. V.,Alter, G. M., Rao, N. S. S. 1990. Bacteriochlorophyll andphotosynthetic reaction centers in Rhizobium strainBTA:1. Appl Environ Microbiol 56:3445–3449.
Fenchel, T., Bernard, C. 1993a. A purple protist. Nature362:300.
Fenchel, T., Bernard, C. 1993b. Endosymbiotic purple non-sulphur bacteria in an anaerobic ciliated protozoon.FEMS Microbiol Lett 110:21–25.
Ferguson, S. J., Jackson, J. B., McEwan, A. G. 1987. Anaero-bic respiration in the Rhodospirillaceae: characteriza-tion of pathways and evaluation of roles in redoxbalancing during photosynthesis. FEMS Microbiol Rev46:117–143.
Fernández-González, A., Martínez-Pérez, I. M., Vioque, A.1998. Characterization of two carotenoid gene promot-

78 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
ers in the cyanobacterium Synechocystis sp PCC6803.Biochim Biophys Acta 1443:343–351.
Ferris, M. J., Ruff-Roberts, A. L., Kopczynski, E. D., Bateson,M. M., Ward, D. M. 1996. Enrichment culture andmicroscopy conceal diverse thermophilic Synechococcuspopulations in a single hot spring microbial mat habitat.Appl Environ Microbiol 62:1045–1050.
Fleischman, D. E., Evans, W. R., Miller, I. M. 1995.Bacteriochlorophyll-containing Rhizobium species. In:Blankenship RE, Madigan MT, Bauer CE (Eds.) Anox-ygenic Photosynthetic Bacteria. Kluwer Academic Pub-lishers. Dordrecht, The Netherlands. 123–136.
Foy, R. H., Gibson, C. E. 1982. Photosynthetic characteristicsof planktonic blue-green algae: changes in photosyn-thetic capacity and pigmentation of Oscillatoria redekeiVan Goor under high and low light. Br Phycol 17:183–193.
Fröstl, J. M., Overmann, J. 1998. Physiology and tacticresponse of the phototrophic consortium “Chlorochro-matium aggregatum”. Arch Microbiol 169:129–135.
Fründ, C., Cohen, Y. 1992. Diurnal cycles of sulfate reductionunder oxic conditions in cyanobacterial mats. ApplEnviron Microbiol 58:70–77.
Friedmann, E. I., Ocampo, R. 1976. Endolithic blue-greenalgae in the Dry valleys: primary producers in the Anct-arctic Desert Ecosystem. Science 193:1247–1249.
Friedmann, E. I. 1982. Endolithic microorganims in theAntactc Cold Desert. Science 215:1045–1053.
Fritsch, F. E. 1922. The terrestrial algae. J Ecol 10:220–236.Fry, B. 1986. Sources of carbon and sulfur nutrition for con-
sumers in three meromictic lakes of New York state.Limnol Oceanogr 31:79–88.
Göbel, F. 1978. Quantum efficiencies of growth. In: ClaytonRK, Sistrom WR (Eds.) The photosynthetic bacteria.Plenum Press. New York, 907–925.
Galinski, E. A. 1995. Osmoadaptation in bacteria. AdvMicrob Physiol 37:274–328.
Gantt, E. 1994. Supramolecular membrane organization. In:Bryant DA (Eds.) The Molecular Biology of Cyanobac-teria. Kluwer Academic Publishers. Dordrecht, TheNetherlands. 119–138.
Garcia-Pichel, F., Castenholz, R. W. 1990. Comparative anox-ygenic photosynthetic capacity in 7 strains of a thermo-philic cyanobacterium. Arch Microbiol 153:344–355.
Garcia-Pichel, F., Mechling, M., Castenholz, R. W. 1994a.Diel migrations of microorganisms within a benthic,hypersaline mat community. Appl Environ Microbiol60:1500–1511.
Garcia-Pichel, F. 1994b. A model for internal self-shadingin planktonic microorganisms and its implications forthe usefulness of sunscreens. Limnol Ocean 39:1704–1717.
Garcia-Pichel, F., Castenholz, R. W. 1994c. On the signifi-cance of solar ultraviolet radiation for the ecology ofmicrobial mats. In: Stal LJ, Caumette P (Eds.) Microbialmats. Structure, development and environmental signif-icance. Springer. Heidelberg, 77–84.
Garcia-Pichel, F. 1995. A scalar irradiance fiber-optic micro-probe for the measurement of ultraviolet radiation athigh spatial resolution. Photochem Photobiol 61:248–254.
Garcia-Pichel, F., Belnap, J. 1996. Microenvironments andmicroscale productivity of cyanobacterial desert crusts.J Phycol 32:774–782.
Garcia-Pichel, F., Prufert-Bebout, L., Muyzer, G. 1996. Phe-notypic and phylogenetic analyses show Microcoleus
chthonoplastes to be a cosmopolitan cyanobacterium.Appl Environ Microbiol 62:3284–3291.
Garcia-Pichel, F., Bebout, B. 1996. The penetration of ultra-violet radiation into shallow water sediments: high expo-sure for photosynthetic communities. Mar. Ecol. Prog.Ser. 131:257–262.
Garcia-Pichel, F., Nübel, U., Muyzer, G. 1998. The phylogenyof unicellular, extremely halotolerant cyanobacteria.Arch Microbiol 169:469–482.
Garcia-Pichel, F. 1999. Cyanobacteria. In: Lederberg J (Eds.)Encyclopedia of Microbiology. Academic Press. SanDiego.
Garcia-Pichel, F., Castenholz, R. W. 1999. Photomovementsof microroganisms in sediments and soils. In: HäderD-P (Eds.) Photomovements. Elsevier. Amsterdam,
Gates, D. M. 1962. Energy exchange in the biosphere. Harper& Row. New York, 151.
Geitler, L. 1932. Cyanophyceae. Akademische Verlagsgesell-schaft. Leipzig, 1035.
Gest, H., Schopf, J. W. 1983a. Biochemical evolution ofanaerobic energy conversion: the transition from fer-mentation to anoxygenic photosynthesis. In: Schopf JW(Eds.) Earth’s Earliest Biosphere. Princeton UniversityPress. Princeton, NJ. 135–148.
Gest, H., Favinger, J. L. 1983b. Heliobacterium chlorum: ananoxygenic brownish-green photosynthetic bacteriumcontaining a new form of bacteriochlorophyll. ArchMicrobiol 136:11–16.
Gest, H. 1993. Photosynthetic and quasi-photosynthetic bac-teria. FEMS Microbiol Lett 112:1–6.
Giovanonni, S. J., Turner, S., Olsen, G. J., Barns, S., Lane,D. J., Pace, N. R. 1988. Evolutionary relatiosnhips amongcyanobacteria and green chloroplasts. J Bacteriol170:3548–3592.
Glaeser, J., Overmann, J. 1999. Selective enrichment andcharacterization of Roseospirillum parvum, gen. nov.and sp. nov., a new purple nonsulfur bacterium withunusual light absorption properties. Arch Microbiol171:405–416.
Goericke, R., Repeta, D. J. 1992. The pigments of Prochlo-rococcus marinus: The presence of divinyl chlorophyll aand b in a marine procaryote. Limnol Oceanogr 57:425–433.
Goericke, R., Welschmeyer, N. A. 1993. The marine prochlo-rophyte Prochlorococcus contributes significantly tophytoplankton biomass and primary production in theSargasso Sea. Deep-Sea Res 40:2283–2294.
Golbeck, J. H. 1994. Photosystem I in cyanobacteria. In: Bry-ant DA (Eds.) The molecular biology of cyanobacteria.Kluwer Academic Publishers. Dordrecht, Boston, Lon-don, 319–360.
Golden, S. S. 1994. Light responsive gene expression and thebiochemistry of photosystem II reaction center. In:Bryant DA (Eds.) The molecular biology of cyanobac-teria. Kluwer Academic Press. Dordrecht, 693–714.
Golecki, J. R., Oelze, J. 1987. Quantitative relationshipbetween bacteriochlorophyll content, cytoplasmic mem-brane structure and chlorosome size in Chloroflexusaurantiacus. Arch Microbiol 148:236–241.
Gorlenko, V. M., Krasilánikova, E. N., Kikina, O. G., Tatar-inova, N. Y. 1979. The new motile purple sulfur bacte-rium Lamprobacter modestohalophilus nov. gen., nov.sp. with gas vacuoles. Izv. Akad. Nauk. S. S. S. R. Ser.Biol. 5:755–767.
Gottschalk, G. 1986. Bacterial metabolism. Springer. NewYork, 359.

CHAPTER 1.3 The Phototrophic Way of Life 79
Granick, S. 1965. Evolution of heme and chlorophyll. In:Bryson V, Vogel HJ (Eds.) Evolving genes and proteins.Academic Press. New York, 67–88.
Guerrero, R., Pedrós-Alió, C., Esteve, I., Mas, J., Chase, D.,Margulis, L. 1986. Predatory prokaryotes: Predation andprimary consumption evolved in bacteria. Proc NatlAcad Sci USA 83:2138–2142.
Guyoneaud, R., Matheron, R., Baulaigue, R., Podeur, K.,Hirschler, A., Caumette, P. 1996. Anoxygenic pho-totrophic bacteria in eutrophic coastal lagoons of theFrench Mediterranean and Atlantic Coasts. Hydrobio-logia 329:33–43.
Häder, D-P. 1987. Photosensory behavior in prokaryotes.Microbiol Rev 51:1–21.
Hansen, T. A., van Gemerden, H. 1972. Sulfide utilizationby purple nonsulfur bacteria. Arch Mikrobiol 86:49–56.
Hansen, T. A., Veldkamp, H. 1973. Rhodopseudomonas sul-fidophila nov. sp., a new species of the purple nonsulfurbacteria. Arch Microbiol 92:45–58.
Harashima, K., Shiba, T., Totsuka, T., Simidu, U., Taga, N.1978. Occurrence of bacteriochlorophyll a in a strain ofan aerobic heterotrophic bacterium. Agricult Biol ChemTokyo 42:1627–1628.
Harder, W., van Dijken, J. P. 1976. Theoretical considerationson the relation between energy production and growthof methane-utilizing bacteria. In: Schlegel HG,Gottschalk G, Pfennig N (Eds.) Symposium on microbialproduction and utilization of gases (H2, CH4, CO). EGoltze KG. Göttingen, 403–418.
Hartman, H. 1992. Conjectures and reveries. Photosynth Res33:171–176.
Hartmann, R., Sickinger, H-D., Oesterhelt, D. 1980. Anaer-obic growth of halobacteria. Proc Natl Acad Sci USA77:3821–3825.
Hayes, J. M., Kaplan, I. R., Wedeking, K. W. 1983. Precam-brian organic geochemistry, preservation of the record.In: Schopf JW (Eds.) Earth’s Earliest Biosphere. Princ-eton University Press. Princeton, NJ. 93–134.
Heising, S., Richter, L., Ludwig, W., Schink, B. 1999. Chlo-robium ferrooxidans sp. nov., a phototrophic green sul-fur bacterium that oxidizes ferrous iron in coculturewith a “Geospirillum” sp. strain. Arch Microbiol172:116–124.
Hofman, P. A. S., Veldhuis, M. J. W., van Gemerden, H. 1985.Ecological significance of actetate assimilation by Chlo-robium phaeobacteroides. FEMS Microbiol Ecol31:271–278.
Holo, H. 1989. Chloroflexus aurantiacus secretes 3-hydrox-ypropionate, a possible intermediate in the assimilationof CO2 and acetate. Arch Microbiol 151:252–256.
Holtin, R. W., Stam, W. T., Boele-Bos, S. A. 1990. DNA-DNAreassociation studies with DNA from Prochloron(prochlorophyta) samples of Indo-West Pacific origin. JPhycol 26:358–361.
Hugenholtz, P., Pitulle, C., Hershberger, K. L., Pace, N. R.1998. Novel division level bacterial diversity in a Yellow-stone hot spring. J Bacteriol 180:366–376.
Imhoff, J. F., Tindall, B. J., Grant, W. D., Trüper, H. G. 1981.Ectothiorhodospira vacuolata sp. nov., a new pho-totrophic bacterium from soda lakes. Arch Microbiol130:238–242.
Imhoff, J. F., Trüper, H. G. 1989. Purple nonsulfur bacteria.In: Staley JT, Bryant MP, Pfennig N, Holt JG (Eds.)Bergey’s Manual of Systematic Bacteriology. Williamsand Wilkins. Baltimore, 1658–1682.
Jørgensen, B. B., Kuenen, J. G., Cohen, Y. 1979. Microbialtransformations of sulfur compounds in a stratified lake(Solar Lake, Sinai). Limnol Oceanogr 24:799–822.
Jørgensen, B. B. 1982. Ecology of the bacteria of the sulphurcycle with special reference to anoxic-oxic interfaceenvironments. Phil Trans Royal Soc London B 298:543–561.
Jørgensen, B. B., Des Marais, D. J. 1986. A simple fiber-opticmicroprobe for high resolution light measurements:applications in marine sediments. Limnol Oceanogr31:1374–1383.
Jørgensen, B. B., Des Marais, D. J. 1988. Optical propertiesof benthic photosynthetic communities: fiber-opticstudies of cyanobacterial mats. Limnol Ocean 33:99–113.
Jannasch, H. W. 1989. Chemosynthetically sustained ecosys-tems in the deep sea. In: Schlegel HG, Bowien B (Eds.)Autotrophic Bacteria. Springer. New York, 147–166.
Jorgensen, B. B., Cohen, Y. 1977. Solar Lake (Sinai). 5. Thesulfur cycle of the benthic microbial mats. LimnolOceanogr 22:657–666.
Kämpf, C., Pfennig, N. 1980. Capacity of Chromatiaceae forchemotrophic growth. Specific respiration rates of Thio-cystis violacea and Chromatium vinosum. Arch Micro-biol 127:125–135.
Kämpf, C., Pfennig, N. 1986. Chemoautotrophic growth ofThiocystis violacea, Chromatium gracile and C. vinosumin the dark at various O2-concentrations. J Basic Micro-biol 26:517–531.
Kühl, M., Jørgensen, B. B. 1992. Spectral light measurementsin microbenthic phototrophic communities with a fiber-optic microprobe coupled to a sensitive diode arraydetector. Limnol Oceanogr 37:1813–1823.
Kana, T., Gilbert, P. M., Goericke, R., Welschmeyer, N. A.1988. Zeaxanthin and beta-carotene in SynechococcusWH7803 respond differently to irradiation. LimnolOceanogr 33:1623–1627.
Keppen, O. I., Baulina, O. I., Kondratieva, E. N. 1994. Oscil-lochloris trichoides neotype strain DG-6. PhotosynthesisRes 41:29–33.
Kimble, L. K., Mandelco, L., Woese, C. R., Madigan, M. T.1995. Heliobacterium modesticaldum, sp. nov., a ther-mophilic heliobacterium of hot springs and volcanicsoils. Arch Microbiol 163:259–267.
Kirk, J. T. O. 1983. Light and photosynthesis in aquaticecosystems. Cambridge University Press. Cambridge,401.
Kok, B. 1973. Photosynthesis. In: Gibbs M, Hollaender A,Kok B, Krampitz LO, San Pietro A (Eds.) Proceedingsof the Workshop on Bio-solar Conversion. National Sci-ence Foundation. Bethesda, MD.
Komárek, J., Anagnostidis, K. 1989. Modern approach to theclassification system of cyanophytes. 4. Nostocales. ArchHydrobiol Suppl 823, Algol. Studies 56:247–345.
Kondratieva, E. N. 1979. Interrelation between modes ofcarbon assimilation and energy production in pho-totrophic purple and green bacteria. In: Quale JR (Eds.)Microbial Biochemistry. University Park Press. Balti-more, 117–175.
Konopka, A., Brock, T. D., Walsby, A. E. 1978. Buoyancyregulation by planktonic blue-green algae in Lake Men-dota, Wisconsin. Arch Hydrobiol 83:524–537.
Kromkamp, J. C., Mur, L. R. 1984. Buoyant density changesin the cyanobacterium Microcystis aeruginosa due tochanges in the cellular carbohydrate content. FEMSMicrobiol Lett 25:105–109.

80 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
Krutschel, C., Castenholz, R. W. 1998. The effect of solar UVand visible irradiance on the vertical movements ofcyanobacteria in microbial mats of hypesaline waters.FEMS Microbiol Ecol 27:53–72.
Lancaster, C. R. D., Michel, H. 1996. Three-dimensionalstructures of photosynthetic reaction centers. Photo-synth Res 48:65–74.
Laroche, J., Vanderstaay, G. M. W., Partensky, F., Ducret, A.,Aeberssold, R., Li, R., Golden, S. S., Hiller, R. G.,Wrench, P. M., Larkum, A. W. D., Green, B. R. 1996.Independent evolution of the Prochlorophyte and greenplant Chlorophyll a/b light-harvesting proteins. ProcNatl Acad Sci (USA) 93:15244–15248.
Lascelles, J. 1978. Regulation of pyrrole synthesis. In: Clay-ton RK, Sistrom WR (Eds.) The Photosynthetic Bacte-ria. Plenum Press. New York, 795–808.
Lassen, C., Plough, H., Jørgensen, B. B. 1992. A fiber-opticscalar irradinace microsensor: application for spectrallight measurements in sediments. FEMS Microbiol Ecol86:247–254.
Lewin, R. A., Withers, N. W. 1975. Extraordinary pigmentcomposition of a prokaryotic alga. Nature 256:735–737.
Lewin, R. A. 1981. Prochloron and the theory of symbiogen-esis. Ann N Y Acad Sci 361:325–329.
Liebl, U., Mockensturm-Wilson, M., Trost, J. T., Brune, D. C.,Blankenship, R. E., Vermaas, W. 1993. Single corepolypeptide in the reaction center of the photosyntheticbacterium Heliobacillus mobilis: structural implicationsand relations to other photosystems. Proc Natl Acad SciUSA 90:7124–7128.
Lindholm, T., Weppling, K., Jensen, H. S. 1985. Stratificationand primary production in a small brackish lake studiedby close-interval siphon sampling. Verh Internat VereinLimnol 22:2190–2194.
Liu, H., Nolla, H. A., Campbell, L. 1997. Prochlorococcusgrowth rate and contribution to primary production inthe equatorial and subtropical North Pacific Ocean.Aquat Microb Ecol 12:39–47.
Loach, P. A., Parkes-Loach, P. S. 1995. Structure-functionrelationships in core light-harvesting complexes(LHI) as determined by characterization of the struc-tural subunit and by reconstitution experiments. In:Blankenship RE, Madigan MT, Bauer CE (Eds.)Anoxygenic Photosynthetic Bacteria. Kluwer Aca-demic Publishers. Dordrecht, The Netherlands. 437–471.
Lokstein, H., Steglich, C., Hess, W. R. 1999. Light-harvestingantenna function of phycorerythrin in Prochlorococcusmarinus. Biochim Biophys Acta- Bioenergetics 1410:97–98.
Madigan, M. T., Takigiku, R., Lee, R. G., Gest, H., Hayes,J. M. 1989. Carbon isotope fractionation by thermophilicphototrophic sulfur bacteria: Evidence for autotrophicgrowth in natural populations. Appl Environ Microbiol55:639–644.
Madigan, M. T. 1992. The Family Heliobacteriaceae. In:Balows A, Trüper HG, Dworkin M, Harder W, SchleiferKH (Eds.) The Prokaryotes. Springer. New York, 1982–1992.
Madigan, M. T., Ormerod, J. G. 1995. Taxonomy, physiologyand ecology of Heliobacteria. In: Blankenship RE,Madigan MT, Bauer CE (Eds.) Anoxygenic Photosyn-thetic Bacteria. Kluwer Academic Publisher. Dordrecht,The Netherlands. 17–30.
Madigan, M. T., Martinko, J. M., Parker, J. 1997. Biology ofMicroorganisms. Prentice Hall International. London,986.
Mague, T. H. 1977. Ecological aspects of dinitrogen fixationby blue-green algae. In: Hardy RWF, Gibson AH (Eds.)A treatise on dinitrogen fixation. Wiley. New York, 85–140.
Malkin, R. 1992. Cytochrome bc1 and b6f complexes ofphotosynthetic membranes. Photosynth Res 33:121–136.
Martin, J. H., Gordon, R. M., Fitzwater, S. E. 1990. Iron inAntarctic waters. Nature 345:156–158.
Mas, J., Pedrós-Alió, C., Guerrero, R. 1990. In situ specificloss and growth rates of purple sulfur bacteria in LakeCisó. FEMS Microbiol Ecol 73:271–281.
Masamoto, K., Furukawa, K. I. 1997. Accumulation of zeax-anthin in the cell of the cyanobacterium Synechococcussp strain PCC7942 grown under high irradiance. J PlantPhysiol 151:257–261.
Matthijs, H. C. P., van der Staay, G. W. M., Mur, L. R. 1994.Prochlorophytes: The “other” cyanobacteria?. In: Bry-ant DA (Eds.) The Molecular Biology of Cyanobacteria.Kluwer Academic Publishers. Dordrecht, The Nether-lands. 49–64.
Mauzerall, D. 1992. Light, iron, Sam Granick and the originof life. Photosynth Res 33:163–170.
McDermott, G., Prince, S. M., Freer, A. A., Hawthornth-waite-Lawless, A. M., Papiz, M. Z., Cogdell, R. J., Isaacs,N. W. 1995. Crystal structure of an integral membranelight-harvesting complex from photosynthetic bacteria.Nature 374:517–521.
Menendez, C., Bauer, Z., Huber, H., Gad’on, N., Stetter,K-O., Fuchs, G. 1999. Presence of acetyl coenzyme A(CoA) carboxylase and propionyl-CoA carboxylase inautotrophic Crenarchaeota and indication for operationof a 3-hydroxypropionate cycle in autototrophic carbonfixation. J Bacteriol 181:1088–1098.
Millie, D. F., Ingram, D. A., Dionigi, C. P. 1990. Pigment andphotosynthetic responses of Oscillatoria aghardii(Cyanophyta) to photon flux and spectral quality. J Phy-col 26:660–666.
Miyachi, S., Strassdat, K., Miyashita, H., Senger, H. 1997.Quantum requirement of photosynthesis in the prima-rily Chlorophyll d containing prokaryote Acaryochlorismarina. Z Naturforsch C-A 52:636–638.
Miyashita, H., Ikemoto, H., Kurano, N., Adachi, K., Chihara,M., Miyachi, S. 1996. Chlorophyll d as a major pigment.Nature 383:402–402.
Moezelaar, R., Stal, L. J. 1994. Anaerobic dark energy gen-eration in the mat-building cyanobacterium Microcoleuschthonoplastes. In: Stal LJ, Caumette P (Eds.) MicrobialMats. Springer. New York, 273–278.
Montesinos, M. L., Herrero, A., Flores, E. 1997. Amino acidtransport in taxonomically diverse cyanobacteria andidentification of two genes encoding elements of a neu-tral aminoacid permease putatively involved in recap-ture of leaked hydrophobic amino acids. J Bacteriol179:853–862.
Moore, L. R., Rocap, G., Chisholm, S. W. 1998. Physiologyand molecular phylogeny of coexisting Prochlorococcusecotypes. Nature 393:464–467.
Nagashima, K. V. P., Shimada, K., Matsuura, K. 1993. Phylo-genetic analysis of photosynthesis genes of Rhodocyclusgelatinosus: possibility of horizontal gene transfer inpurple bacteria. Photosynth Res 36:185–191.

CHAPTER 1.3 The Phototrophic Way of Life 81
Nagashima, K. V. P., Hiraishi, A., Shimada, K., Matsuura, K.1997. Horizontal transfer of genes coding for the photo-synthetic reaction centers of purple bacteria. J Mol Evol45:131–136.
Nelissen, B., van de Peer, Y., Wilmotte, A., de Wachter, R.1995. An early origin of plastids within the cyanobacte-rial divergence is sugggested by evolutionary trees basedon complete 16S rDNA sequences. Mol Biol Evol12:1166–1173.
Nelson, D. C., Castenholz, R. W. 1982. Light responses inBeggiatoa. Arch Microbiol 131:146–155.
Nicholson, J. A. M., Stolz, J. F., Pierson, B. K. 1987. Structureof a microbial mat at Great Sippewissett Marsh,Cape Cod, Massachusetts. FEMS Microbiol Ecol 45:343–364.
Nickens, D., Fry, C. J., Ragatz, L., Bauer, C. E., Gest, H. 1996.Biotype of the purple nonsulfur photosynthetic bacte-rium, Rhodospirillum centenum. Arch Microbiol165:91–96.
Nogales, B., Guerrero, R., Esteve, I. 1997. A heterotrophicbacterium inhibits growth of several species of the genusChlorobium. Arch Microbiol 167:396–399.
Noguchi, T., Hayashi, H., Shimada, K., Takaichi, S., Tasumi,M. 1992. In vivo states and function of carotenoids in anaerobic photosynthetic bacterium, Erythrobacter lon-gus. Photosynth Res 31:21–30.
Nonnengießer, A., Schuster, A., Koenig, F. 1996. Carotenoidsand reaction center II- D1 protein in light regulation ofthe photosynthetic apparatus in Aphanocapsa. Bot Acta109:115–124.
Nübel, U. 1999. Diversität und Salinitätsabhängiges Verh-alten benthischer, cyanobakterieller Lebensgemein-schaften. Ph D. Thesis. University of Bremen. Bremen,
Oelze, J. 1992. Light and oxygen regulation of the synthesisof bacteriochlorophylls a and c in Chloroflexus auranti-acus. J Bacteriol 174:5021–5026.
Oesterhelt, D., Stoeckenius, W. 1973. Function of a new pho-toreceptor membrane. Proc Natl Acad Sci USA70:2853–2857.
Oesterhelt, D., Krippahl, G. 1983. Phototropic growth ofhalobacteria and its use for isolation of photosyntheti-cally-deficient mutants. Ann Microbiol (Inst Pasteur)134 B:137–150.
Oh-Hama, T. 1989. Evolutionary consideration of the twopathways of 5-aminolevulinic acid biosynthesis inorganelles and prokaryotes. Endocytology IV:589–592.
Oh-Hama, T., Santander, P. J., Stolowich, N. J., Scott, A. I.1991. Bacteriochlorophyll c formation via the C5 path-way of 5-aminolevulinic acid synthesis in Chloroflexusaurantiacus. FEBS Lett 281:173–176.
Okamura, K., Takamiya, K., Nishimura, M. 1985. Photosyn-thetic electron transfer system is inoperative in anaero-bic cells of Erythrobacter species strain OCh114. ArchMicrobiol 142:12–17.
Oliver, R. L., Walsby, A. E. 1984. Direct evidence for the roleof light-mediated gas vesicle collapse in the buoyancyregulation of Anabaena flos-aquae. Limnol Oceanogr29:879–886.
Olson, R. J., Chisholm, S. W., Zettler, E. R., Armbrust, E. V.1990a. Pigment, size and distribution of Synechococcusin the North Atlantic and Pacific oceans. Limnol Ocean-ogr 35:45–58.
Olson, R. J., Chisholm, S. W., Zettler, E. R., Altabet, M. A.,Dusenberry, J. A. 1990b. Spatial and temporal distribu-
tions of prochlorophyte picoplankton in the NorthAtlantic Ocean. Deep Sea Res 37:1033–1051.
Olson, J. M. 1998. Chlorophyll organization and function ingreen photosynthetic bacteria. Photochem Photobiol67:61–75.
Oparin, A. I. 1938. The origin of life. Macmillan. New York.Overmann, J., Tilzer, M. M. 1989a. Control of primary pro-
ductivity and the significance of photosynthetic bacteriain a meromictic kettle lake. Mittlerer Buchensee, West-Germany. Aquatic Sci 51:261–278.
Overmann, J., Pfennig, N. 1989b. Pelodictyon phaeoclathrat-iforme sp. nov., a new brown-colored member of theChlorobiaceae forming net-like colonies. Arch Micro-biol 152:401–406.
Overmann, J., Beatty, J. T., Hall, K. J., Pfennig, N., Northcote,T. G. 1991a. Characterization of a dense, purple sulfurbacterial layer in a meromictic salt lake. Limnol Ocean-ogr 36:846–859.
Overmann, J., Lehmann, S., Pfennig, N. 1991b. Gas vesicleformation and buoyancy regulation in Pelodictyon phae-oclathratiforme (green sulfur bacteria). Arch Microbiol157:29–37.
Overmann, J., Pfennig, N. 1992. Buoyancy regulation andaggregate formation in Amoebobacter purpureus fromMahoney Lake. FEMS Microbiol Ecol 101:67–79.
Overmann, J., Beatty, J. T., Hall, K. J. 1994. Photosyntheticactivity and population dynamics of Amoebobacter pur-pureus in a meromictic saline lake. FEMS MicrobiolEcol 15:309–320.
Overmann, J., Beatty, J. T., Krouse, H. R., Hall, K. J. 1996.The sulfur cycle in the chemocline of a meromictic saltlake. Limnol Oceanogr 41:147–156.
Overmann, J. 1997. Mahoney Lake: A case study of the eco-logical significance of phototrophic sulfur bacteria. In:Jones JG (Eds.) Advances in Microbial Ecology Vol. 15.Plenum Press. New York, 251–288.
Overmann, J., Tuschak, C., Fröstl, J. M., Sass, H. 1998. Theecological niche of the consortium “Pelochromatiumroseum”. Arch Microbiol 169:120–128.
Overmann, J., Hall, K. J., Northcote, T. G., Beatty, J. T. 1999a.Grazing of the copepod Diaptomus connexus on purplesulfur bacteria in a meromictic salt lake. Environ Micro-biol 1:213–222.
Overmann, J., Coolen, M. J. L., Tuschak, C. 1999b. Specificdetection of different phylogenetic groups ofchemocline bacteria based on PCR and denaturing gra-dient gel electrophoresis of 16S rRNA gene fragments.Arch Microbiol 172:83–94.
Overmann, J., Hall, K. J., Ebenhöh, W., Chapman, M. A.,Beatty, J. T. 1999c. Structure of the aerobic food chainin a meromictic lake dominated by purple sulfur bacte-ria. Arch Hydrobiol 144:127–156.
Oyaizu, H., Debrunner-Vossbrinck, B., Mandelco, L., Stud-ier, J. A., Woese, C. R. 1987. The green non-sulfur bac-teria: a deep branching in the eubacterial line of descent.System Appl Microbiol 9:47–53.
Pace, N. R. 1997. A molecular view of microbial diversity andthe biosphere. Science 276:734–740.
Padan, E. 1979. Impact of facultative anaerobic phototrophicmetabolism and ecology of cyanobacteria. Adv MicrobEcol 3:1–48.
Padan, E., Cohen, Y. 1982. Anoxygenic photosynthesis. In:Botanical Monographs, vol. 19 (Eds.) The biology ofcyanobacteria. Blackwell Scientific Publications.Oxford, 215–235.

82 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
Palenik, B., Haselkorn, R. 1992. Multiple evolutionary ori-gins of prochlorophytes, the chlorophyll b -containingprokaryotes. Nature 355:265–267.
Parkin, T. B., Brock, T. D. 1980a. The effects of light qualityon the growth of phototrophic bacteria in lakes. ArchMicrobiol 125:19–27.
Parkin, T. B., Brock, T. D. 1980b. Photosynthetic bacterialproduction in lakes: The effects of light intensity. LimnolOceanogr 25:711–718.
Parkin, T. B., Brock, T. D. 1981. The role of phototrophicbacteria in the sulfur cycle of a meromictic lake. LimnolOceanogr 26:880–890.
Pedrós-Alió, C., Sala, M. M. 1990. Microdistribution and dielvertical migration of flagellated vs gas-vacuolate purplesulfur bacteria in a stratified water body. Limnol Ocean-ogr 35:1637–1644.
Peltier, G., Schmidt, G. W. 1991. Chlororespiration: Anadaptation to nitrogen deficiency in Chlamydomo-nas reinhardtii. Proc Natl Acad Sci USA 88:4791–4795.
Pentecost, A. 1984. Effects of sedimentation and light inten-sity on mat-forming Oscillatoriaceae with particular ref-erence to Microcoleus lyngyaceus Gomont. J GenMicrobiol 130:983–990.
Pfennig, N., Trüper, H. G. 1974. The phototrophic bacteria.In: Buchanan RE, Gibbons NE (Eds.) Bergey’s manualof determinative bacteriology, 8th edition. Williams andWilkins. Baltimore, 24–64.
Pfennig, N. 1978. General physiology and ecology of photo-synthetic bacteria. In: Clayton RK, Sistrom WR (Eds.)The Photosynthetic Bacteria. Plenum Press. New York,3–18.
Pfennig, N., Trüper, H. G. 1989. Anoxygenic phototrophicbacteria. In: Staley JT, Bryant MP, Pfennig N, Holt JG(Eds.) Bergey’s Manual of Systematic Bacteriology. Wil-liams and Wilkins. Baltimore, 1635–1709.
Pfennig, N., Lünsdorf, H., Süling, J., Imhoff, J. F. 1997. Rho-dospira trueperi gen. nov., spec. nov., a new pho-totrophic Proteobacterium of the alpha group. ArchMicrobiol 168:39–45.
Pierson, B. K., Castenholz, R. W. 1974. Studies of pigmentsand growth in Chloroflexus aurantiacus, a phototrophicfilamentous bacterium. Arch Microbiol 100:283–305.
Pierson, B. K., Oesterle, A., Murphy, G. L. 1987. Pigments,light penetration and photosynthetic activity in themulti-layered microbial mats of Great Sippewisset SaltMarsh, Massachussets. FEMS Microbiol Ecol 45:365–376.
Pierson, B. K., Olson, J. M. 1989. Evolution of photosynthesisin anoxygenic photosynthetic procaryotes. In: Cogen Y,Rosenberg E (Eds.) Microbial Mats: Physiological Ecol-ogy of Benthic Microbial Communities. Am Soc Micro-biol. Washington, DC, 402–427.
Pierson, B. K., Sands, V. M., Frederick, J. L. 1990. Spectralirradinace and distribution of pigments in a highly lay-ered microbial mat. Appl Environ Microbiol 56:2327–2340.
Pierson, B. K., Valdez, D., Larsen, M., Morgan, E., Mack,E. E. 1994. Chloroflexus-like organims from marine andhypersaline environments: distribution and diversity.Photosynthesis Res 41:35–52.
Pierson, B. K., Castenholz, R. W. 1995. Taxonomy and phys-iology of filamentous anoxygenic phototrophs. In: Blan-kenship RE, Madigan MT, Bauer CE (Eds.) AnoxygenicPhotosynthetic Bacteria. Kluwer Academic Publishers.Dordrecht, The Netherlands. 31–47.
Post, A. F., de Wit, R., Mur, L. R. 1985. Interactions betweentemperature and light intensity on growth and photosyn-thesis of the cyanobacterium Oscillatoria agardhii. JPlankton Res 7:487–495.
Potts, M. 1994. Desiccation tolerance in prokaryotes. Micro-biol Rev 58:755–805.
Proctor, L. M., Fuhrman, J. A. 1990. Viral mortality of marinebacteria and cyanobacteria. Nature 343:60–62.
Prufert-Bebout, L., Garcia-Pichel, F. 1994. Field and culti-vated Microcoleus chthonoplastes: the search for cluesto its prevalence in marine microbial mats. In: Stal LJ,Caumette P (Eds.) Microbial Mats: Structure, develop-ment and environmental significance. Springer. NewYork, Heidelberg. 265–271.
Ragatz, L., Jiang, Z-Y., Bauer, C., Gest, H. 1994. Phototacticpurple bacteria. Nature 370:104.
Ragatz, L., Jiang, Z-Y., Bauer, C. E., Gest, H. 1995. Macro-scopic phototactic behaviour of the purple photosyn-thetic bacterium Rhodospirillum centenum. ArchMicrobiol 163:1–6.
Rau, G. H. 1980. Carbon-13/Carbon-12 variation in subalpinelake aquatic insects: Food source implications. Can JFish Aquat Sci 37:742–746.
Rebeiz, C. A., Lascelles, J. 1982. Biosynthesis of pigments inplants and bacteria. In: Godvindjee (Eds.) Energy Con-version by Plants and Bacteria, Vol I. Academic Press.New York, 699–780.
Reeves, R. H. 1996. 16S ribosomal RNA and the molecularphylogeny of the Cyanobacteria. Nova Hedwiga 112:55–67.
Repeta, D. J., Simpson, D. J., Jørgensen, B. B., Jannasch,H. W. 1989. Evidence for the existence of anoxygenicphotosynthesis from the distribution of bacteriochloro-phylls in the Black Sea. Nature 342:69–72.
Richaud, P., Marrs, B. L., Vermiglio, A. 1986. Two modes ofinteraction between photosynthetic and respiratoryelectron chains in whole cells of Rhodopseudomonascapsulata. Biochim Biophys Acta 850:265–263.
Rippka, R., Deruelles, J., Waterbury, J. B., Herdman, M.,Stanier, R. Y. 1979. Generic assignments, strain historiesand properties of pure cultures of cyanobacteria. J GenMicrobiol 111:1–61.
Rödig, J., Jock, S., Klug, G. 1999. Coregulation of the synthe-sis of bacteriochlorophyll and pigment-binding proteinsin Rhodobacter capsulatus. Arch Microbiol 171:198–204.
Rudi, K., Skulberg, D. M., Jakobsen, K. S. 1998. Evolution ofcyanobacteria by exchange of genetic material amongphylogenetically related strains. J Bacteriol 180:3453–3461.
Ruff-Roberts, A. L., Kuenen, J. G., Ward, D. M. 1994. Distri-bution of cultuvated and uncultivated cyanobacteria andChloroflexus-like bacteria in hot spring microbial mats.Appl Environ Microbiol 60:697–704.
Sackett, M. J., Armitage, J. P., Sherwood, E. E., Pitta, T. P.1997. Photoresponses of the purple nonsulfur bacteriaRhodospirillum centenum and Rhodobacter sphaeroi-des. J Bacteriol 179:6764–6768.
Sánchez, O., van Gemerden, H., Mas, J. 1998. Acclimation ofthe photosynthetic response of Chromatium vinosum tolight-limiting conditions. Arch Microbiol 170:405–410.
Schenk, H. E. A. 1992. Cyanobacterial symbioses. In: BalowsA, Trüper HG, Dworkin M, Harder W, Schleifer K-H(Eds.) The Prokaryotes. Springer. New York, 3819–3854.
Schiller, H., Senger, H., Miyashita, H., Miyashi, S., Dau, H.1997. Light-harvesting in Acaryochloris marina—

CHAPTER 1.3 The Phototrophic Way of Life 83
Spectroscopic characterization of a Chlorophyll d-dominated photosynthetic antenna system. FEBSLetters 410:433–436.
Shiba, T., Simidu, U., Taga, N. 1979. Distribution of aerobicbacteria which contain bacteriochlorophyll a. ApplEnviron Microbiol 38:43–45.
Shiba, T., Harashima, K. 1986. Aerobic photosynthetic bac-teria. Microbiological Sciences 3:377–378.
Shiba, T. 1989. Overview of the aerobic photosynthetic bac-teria. In: Harashima K, Shiba T, Murata N (Eds.)Aerobic Photosynthetic Bacteria. Springer. Berlin, 1–8.
Shimada, K. 1995. Aerobic anoxygenic phototrophs. In:Blankenship RE, Madigan MT, Bauer CE (Eds.) Anox-ygenic photosynthetic bacteria. Kluwer Academic Pub-lishers. Dordrecht, The Netherlands. 105–122.
Sidler, W. A. 1994. Phycobilisome and phycobiliprotein struc-tures. In: Bryant DA (Eds.) The Molecular Biology ofCyanobacteria. Kluwer Academic Publishers. Dor-drecht, Boston, London, 139–216.
Sinninghe Damsté, J. S., Wakeham, S. G., Kohnen, M. E. L.,Hayes, J. M., de Leeuw, J. W. 1993. A 6, 000-year sedi-mentary molecular record of chemocline excursions inthe Black Sea. Nature 362:827–829.
Sirevåg, R. 1995. Carbon metabolism in green bacteria. In:Blankenship RE, Madigan MT, Bauer CE (Eds.) Anox-ygenic Photosynthetic Bacteria. Kluwer Academic Pub-lishers. Dordrecht, The Netherlands. 871–883.
Sistrom, W. R., Griffiths, M., Stanier, R. Y. 1956. The biologyof a photosynthetic bacterium which lacks colored car-otenoids. J Cell Comp Physiol 48:473–515.
Skyring, G. W., Bauld, J. 1990. Microbial mats in coastalenvironments. Adv Microb Ecol 11:461–498.
Smith, R. C., Baker, K. S. 1981. Optical properties of theclearest seawaters. Appl. Optics 20:177–184.
Sorokin, Y. I. 1970. Interrelations between sulphur and car-bon turnover in meromictic lakes. Arch Hydrobiol66:391–446.
Stackebrandt, E., Embley, M., Weckesser, J. 1988. Phyloge-netic, evolutionary, and taxonomic aspects of pho-totrophic bacteria. In: Olson JM, Stackebrandt E,Trüper HG (Eds.) Green Photosynthetic Bacteria. Ple-num Publishing Cooperation. New York, 201–215.
Stackebrandt, E., Rainey, F. A., Ward-Rainey, N. 1996. Anox-ygenic phototrophy across the phylogenetic spectrum:current understanding and future perspectives. ArchMicrobiol 166:211–223.
Stam, W. T., Boele-Bos, S. A., Stulp, B. K. 1985. Genotypicrelationships between Prochloron samples from differ-ent localities and hosts as determined by DNA-DNAreassociations. Arch Microbiol 142:340–341.
Stanier, R. Y. 1977. The position of cyanobacteria in theworld of phototrophs. Carlsberg Res. Comm. 42:77–98.
Steenbergen, C. L. M., Korthals, H. J. 1982. Distribution ofphototrophic microorganisms in the anaerobic andmicroaerophilic strata of Lake Vechten (The Nether-lands). Limnol Oceanogr 27:883–895.
Steudel, R. 1989. On the nature of the “elmental sulfur” (So)produced by sulfur-oxidizing bacteria-a model for So
globules. In: Schlegel HG, Bowien B (Eds.) AutotrophicBacteria. Springer. New York, 289–303.
Steudel, R., Holdt, G., Visscher, P. T., van Gemerden, H.1990. Search for polythionates in cultures of Chroma-tium vinosum after sulfide incubation. Arch Microbiol153:432–437.
Strauß, G., Fuchs, G. 1993. Enzymes of a novel autotrophicCO2 fixation pathway in the phototrophic bacteriumChloroflexus aurantiacus, the hydroxypropionate cycle.Eur J Biochem 215:633–643.
Suttle, C. A., Chan, A. M., Cottrell, M. T. 1990. Infection ofphytoplankton by viruses and reduction of primary pro-ductivity. Nature 347:467–469.
Suttle, C. A., Chan, A. M., Feng, C., Garza, D. R. 1993.Cyanophages and sunlight: A paradox. In: GuerreroR, Pedrós-Alió C (Eds.) Trends in Microbial Ecol-ogy. Spanish Society for Microbiology. Barcelona,303–307.
Suwanto, A., Kaplan, S. 1989. Physical and genetic mappingof the Rhodobacter sphaeroides 2.4.1 genome: genomesize, fragment identification, and gene localization. JBacteriol 171:5840–5849.
Suzuki, J. Y., Bauer, C. E. 1995. A prokaryotic origin for light-dependent chlorophyll biosynthesis of plants. Proc NatlAcad Sci USA 92:3749–3753.
Swoager, W. C., Lindstrom, E. S. 1971. Isolation and countingof Athiorhodaceae with membrane filters. Appl Micro-biol 22:683–687.
Takahashi, K., Wada, E., Sakamoto, M. 1990. Carbon isotopediscrimination by phytoplankton and photosyntheticbacteria in monomictic Lake Fukami-ike. Arch Hydro-biol 120:197–210.
Tanada, T., Kitadokoro, K., Higuchi, Y., INaka, K., Yasui, A.,Deruiter, P. E., Eker, A. P. M., Miki, K. 1997. Crystalstructure of DNA photolyase from Anacystis nidulans.Nature Struct. Biol 4:887–891.
Thauer, R. K., Jungermann, K., Decker, K. 1977. Energyconservation in chemotrophic anaerobic bacteria. Bac-teriolog Rev 41:100–180.
Theroux, S. J., Redlinger, T. E., Fuller, R. C., Robinson, S. J.1990. Gene encoding the 5.7-kilodalton chlorosome pro-tein of Chloroflexus aurantiacus: regulated message lev-els and a predicted carboxy-terminal protein extension.J Bacteriol 172:4497–4504.
Thomas, R. H., Walsby, A. E. 1985. Buoyancy regulationin a strain of Microcystis. J Gen Microbiol 131:799–809.
Thorne, S. W., Newcomb, E. H., Osmond, C. B. 1977. Identi-fication of chlorophyll b in extracts of prokaryotic algaeby fluorescence spectroscopy. Proc Natl Acad Sci74:575–578.
Trüper, H. G., Pfennig, N. 1978. Taxonomy of the Rhodospi-rillales. In: Clayton RK, Sistrom WR (Eds.) The photo-synthetic bacteria. Plenum Publishing Corp.. New York,19–27.
Trüper, H. G., Pfennig, N. 1981. Characterization and identi-fication of the anoxygenic phototrophic bacteria. In:Starr MP, Stolp H, Trüper HG, Balows A, Schlegel HG(Eds.) The prokaryotes: A handbook on habitats, isola-tion and identification of bacteria. Springer. New York,299–312.
Turner, S. 1887. Molecular systematics of oxygenic photosyn-thetic bacteria. Plant Syst Evol 11:13–52.
Urakami, T., Komagata, K. 1984. Protomonas, a new genusof facultatively methylotrophic bacteria. Int J Syst Bac-teriol 34:188–201.
Urbach, E., Robertson, D. L., Chrisholm, S. W. 1992. Multipleevolutionary origins of prochlorophytes within thecyanobacterial radiation. Nature 355:267–270.
Utkilen, H. C., Skulberg, O. M., Walsby, A. E. 1985. Buoy-ancy regulation and chromatic adaptation in planktonicOscillatoria species: alternative strategies for optimizing

84 J. Overmann and F. Garcia-Pichel CHAPTER 1.3
light absorption in stratified lakes. Arch Hydrobiol104:407–417.
van den Ende, F. P., Laverman, A. M., van Gemerden, H.1996. Coexistence of aerobic chemotrophic and anaero-bic phototrophic sulfur bacteria under oxygen limita-tion. FEMS Microbiol Ecol 19:141–151.
van den Ende, H., van de Peer, Y., Perry, J., de Wachter, R.1990. 5S rRNA sequences of representatives of the gen-era Chlorobium, Prosthecochloris, Thermomicrobium,Cytophaga, Flavobacterium, Flexibacter and Saprospiraand a discussion of the evolution of eubacteria in gen-eral. J Gen Microbiol 136:11–18.
van Gemerden, H. 1967. In the bacterial sulfur cycle of inlandwaters. University of Leiden.
vanGemerden, H. 1974. Coexistence of organisms competingfor the same substrate: An example among the purplesulfur bacteria. Microb Ecol 1:104–119.
van Gemerden, H., Tughan, C. S., de Wit, R., Herbert, R. A.1989. Laminated microbial ecosystems on shelteredbeaches in Scapa Flow, Orkney Islands. FEMS Micro-bial Ecol 62:87–102.
van Gemerden, H., Mas, J. 1995. Ecology of phototrophicsulfur bacteria. In: Blankenship RE, Madigan MT,Bauer CE (Eds.) Anoxygenic photosynthetic bacteria.Kluwer Academic Publishers. Dordrecht, The Nether-lands. 49–85.
van Thor, J. J., Mullineaux, C. W., Matthijs, H. P. C., Helling-wert, K. J. 1998. Light harvesting and state transitions incyanobacteria. Bot Acta 111:430–443.
van Valen, L. M., Maiorana, V. C. 1980. The archaebacteriaand eukaryotic origins. Nature 287:248–250.
Vanderstaay, G. W. M., Yurkova, N., Green, B. R. 1998. The38KDa Chlorophyll a/b protein of the prokaryoteProchlorothrix hollandica is encoded by a divergent Pcbgene. Plant Mol Biol 36:706–716.
Veldhuis, M. J. W., van Gemerden, H. 1986. Competitionbetween purple and brown phototrophic bacteria instratified lakes: sulfide, acetate, and light as limiting fac-tors. FEMS Microbiol Ecol 38:31–38.
Vermaas, W. F. J. 1994. Evolution of heliobacteria: Implica-tions for photosynthetic reaction cneter complexes.Photosynth Res 41:285–294.
Wagner-Huber, R., Brunisholz, R., Frank, G., Zuber, H.1988. The BChl c/e- binding polypeptides from chlo-rosomes of green photosynthetic bacteria. FEBS Lett239:8–12.
Wakao, N., Yokoi, N., Isoyama, N., Hiraishi, A., Shimada, K.,Kobayachi, M., Kise, H., Iwaki, M., Itoh, S., Takaishi, S.,Sakurai, Y. 1996. Discovery of natural photosynthesisusing Zn-containing bacteriochlorophyll in an aerobicbacterium Acidiphilium rubrum. Plant Cell Physiol37:889–896.
Wakim, B., Oelze, J. 1980. The unique mode of adjusting thecomposition of the photosynthetic apparatus to differentenvironmental conditions by Rhodospirillum tenue.FEMS Microbiol Lett 7:221–223.
Walsby, A. E. 1978. The properties and buoyancy-providingrole of gas vacuoles in Trichodesmium Ehrenberg. BrPhycol J 13:103–116.
Walsby, A. E. 1994. Gas vesicles. Microbiol Rev 58:94–144.Ward, D. M., Weller, R., Shiea, J., Castenholz, R. W., Cohen,
Y. 1989. Hot springs microbial mats:Anoxygenic andoxygenic mats of possible evolutionary significance. In:Cohen Y, Rosenberg E (Eds.) Microbial Mats: Physio-logical Ecology of Benthic Microbial Communities. AmSoc Microbiol. Washington, DC, 3–15.
Waterbury, J. B., Wiley, J. M., Franks, D. G., Valois, F. W.,Watson, S. W. 1985. A cyanobacterium capable of swim-ming motility. Science 230:74–76.
Waterbury, J. B., Watson, S. W., Valois, F. W., Franks, D. G.1986. Biological and ecological characterization of themarine unicellular cyanobacterium Synechococcus. In:Platt T, Li WKW (Eds.) Photosynthetic Picoplankton.Can Bull Fish Aquat Sci 214. 71–120.
Waterbury, J. B., Valois, F. W. 1993. Resistance to co-occurring phages enables marine Synechococcus com-munities to coexist with cyanophages abundant in sea-water. Appl Environ Microbiol 59:3393–3399.
Watson, S. W., Novitsky, T. J., Quinby, H. L., Valois, F. W.1977. Determination of bacterial number and biomassin the marine environment. Appl Environ Microbiol33:940–946.
Weller, R., Bateson, M. M., Heimbuch, B. K., Kopczynski,E. D., Ward, D. M. 1992. Uncultivated cyanobacteria,Chloroflexus-like inhabitants, and spirochete-like inhab-itants of a hot spring microbial mat. Appl EnvironMicrobiol 58:3964–3969.
Wellington, C. L., Bauer, C. E., Beatty, J. T. 1992. Photo-synthesis gene superoperons in purple non-sulfurbacteria: The tip of the iceberg?. Can J Microbiol 38:20–27.
Wessels, D. C. J., Büdel, B. 1995. Epilithic and cryptoendo-lithic cyanobacteria of Clarens Sandstone Cliffs in theGolden Gate Highlands National Park, South Africa.Bot Acta 108:220–226.
Whatley, J. M. 1977. The fine structure of Prochloron. NewPhytol 79:309–313.
Whittaker, R. H., Likens, G. E. 1975. The biosphere andman. In: Lieth H, Whittaker RH (Eds.) Primary pro-ductivity of the biosphere. Springer. New York, 305–328.
Widdel, F., Schnell, S., Heising, S., Ehrenreich, A., Assmus,B., Schink, B. 1993. Ferrous iron oxidation by anoxy-genic phototrophic bacteria. Nature 362:834–836.
Woese, C. R., Gibson, J., Fox, G. E. 1980. Do genealogicalpatterns in photosynthetic bacteria reflect interspecificgene transfer?. Nature 283:212–214.
Woese, C. E. 1987. Bacterial evolution. Microbiol Rev51:221–271.
Wraight, C. A., Clayton, R. K. 1973. The absolute quantumefficiency of bacteriochlorophyll photooxidation in reac-tion centres of Rhodopseudomonas spheroides. Bio-chim Biophys Acta 333:246.
Wu, H., Green, M., Scranton, M. I. 1997. Acetate cycling inthe water column and surface sediment of Long IslandSound following a bloom. Limnol Oceanogr 42:705–713.
Yildiz, F. H., Gest, H., Bauer, C. E. 1992. Conservation of thephotosynthesis gene cluster in Rhodospirillum cente-num. Mol Microbiol 6:2683–2691.
Yurkov, V., van Gemerden, H. 1993. Impact of light/darkregime on growth rate, biomass formation and bacteri-ochlorophyll synthesis in Erythromicrobium hydrolyti-cum. Arch Microbiol 159:84–89.
Yurkov, V., Gad’on, N., Angerhofer, A., Drews, G. 1994.Light-harvesting complexes of aerobic bacteriochloro-phyll-containing bacteria Roseococcus thiosulfatophi-lus, RB3 and Erythromicrobium ramosum, E5 and thetransfer of excitation energy from carotenoids to bacte-riochlorophyll. Z Naturforsch 49(c):579–586.
Yurkov, W., Beatty, J. T. 1998. Isolation of aerobic anoxygenicphotosynthetic bacteria from black smoker plume

CHAPTER 1.3 The Phototrophic Way of Life 85
waters of the Juan de Fuca Ridge in the Pacific Ocean.Appl Environ Microbiol 64:337–341.
Zehnder, A. J. B., Stumm, W. 1988. Geochemistry and bio-geochemistry of anaerobic habitats. In: Zehnder AJB(Eds.) Biology of Anaerobic Microorganisms. Wiley-Liss. New York, 1–38.
Zevenboom, W., Mur, L. R. 1984. Growth and photosyntheticresponse of the cyanobacterium Microcystis aeruginosain relation to photoperiodicity and irradiance. ArchMicrobiol 139:232–239.
Zuber, H., Cogdell, R. J. 1995. Structure and organization ofpurple bacterial antenna complexes. In: BlankenshipRE, Madigan MT, Bauer CE (Eds.) Anoxygenic Photo-synthetic Bacteria. Kluwer Academic Publishers. Dor-drecht, The Netherlands. 315–348.
Zucconi, A. P., Beatty, J. T. 1988. Posttranscriptional regula-tion by light of the steady state levels of mature B800-850 light-harvesting complex in Rhodobacter capsula-tus. J Bacteriol 170:877–882.