the plant journal 47 light activates the degradation of ... files/abiotic/oh et al_pj06_pil5.pdf ·...
TRANSCRIPT

Light activates the degradation of PIL5 protein to promoteseed germination through gibberellin in Arabidopsis
Eunkyoo Oh1, Shinjiro Yamaguchi2, Yuji Kamiya2, Gabyong Bae1, Won-Il Chung1 and Giltsu Choi1,*
1Department of Biological Sciences, KAIST, Daejeon 305-701, Korea, and2RIKEN Plant Science Center, Yokohama, Kanagawa 230-0045, Japan
Received 25 January 2006; revised 14 March 2006; accepted 21 March 2006.*For correspondence (fax þ82 42 869 2610; e-mail [email protected]).
Summary
Angiosperm seeds integrate various environmental signals, such as water availability and light conditions, to
make a proper decision to germinate. Once the optimal conditions are sensed, gibberellin (GA) is synthesized,
triggering germination. Among environmental signals, light conditions are perceived by phytochromes.
However, it is not well understood how phytochromes regulate GA biosynthesis. Here we investigated
whether phytochromes regulate GA biosynthesis through PIL5, a phytochrome-interacting bHLH protein, in
Arabidopsis. We found that pil5 seed germination was inhibited by paclobutrazol, the ga1 mutation was
epistatic to the pil5 mutation, and the inhibitory effect of PIL5 overexpression on seed germination could be
rescued by exogenous GA, collectively indicating that PIL5 regulates seed germination negatively through GA.
Expression analysis revealed that PIL5 repressed the expression of GA biosynthetic genes (GA3ox1 and
GA3ox2), and activated the expression of a GA catabolic gene (GA2ox) in both PHYA- and PHYB-dependent
germination assays. Consistent with these gene-expression patterns, the amount of bioactive GA was higher
in the pil5 mutant and lower in the PIL5 overexpression line. Lastly, we showed that red and far-red light
signals trigger PIL5 protein degradation through the 26S proteasome, thus releasing the inhibition of bioactive
GA biosynthesis by PIL5. Taken together, our data indicate that phytochromes promote seed germination by
degrading PIL5, which leads to increased GA biosynthesis and decreased GA degradation.
Keywords: PIL5, phytochrome, seed germination, gibberellin, protein degradation, Arabidopsis.
Introduction
As photosynthetic organisms, plants must carefully monitor
external light conditions and adjust their growth and
development accordingly. In angiosperms, at least three
photoreceptor systems – phototropins (PHOTs); crypto-
chromes (CRYs); and phytochromes (PHYs) – are respon-
sible for monitoring light conditions and responding by
making adjustments to various physiological and develop-
mental processes (Fankhauser and Staiger, 2002). Among
these, phytochromes perceive red and far-red lights and
regulate various processes, including seed germination,
seedling development, chloroplast development, shade
avoidance and flowering (Chory et al., 1996; Neff et al., 2000;
Sullivan and Deng, 2003).
In Arabidopsis, phytochromes consist of five members
that can be grouped into the type I phytochrome (PHYA),
which functions as a far-red light receptor; and the type II
phytochromes (PHYB, PHYC, PHYD, and PHYE), which
function as red light receptors (Quail, 1998). Regardless of
their spectral specificities, both types of phytochrome
regulate similar physiological processes by triggering tran-
scriptional cascades that ultimately alter the expression of
10–30% of the entire transcriptome (Ma et al., 2001; Tepper-
man et al., 2001). Various transcription factors are known to
be centrally involved in the transcriptional cascades of light
signalling, including COG1 and OBP3 (encoding DOF family
members); HY5 and HYH (encoding bZIP family members);
LAF1 (encoding a MYB family member); and HFR1, PIL1,
PIL5, PIL6, PIF3 and PIF4 (encoding bHLH family members)
(Ballesteros et al., 2001; Chattopadhyay et al., 1998; Fairchild
et al., 2000; Fujimori et al., 2004; Holm et al., 2002; Huq and
Quail, 2002; Huq et al., 2004; Kim et al., 2003; Monte et al.,
2004; Ni et al., 1998; Oh et al., 2004; Park et al., 2003; Salter
et al., 2003; Ward et al., 2005). Phytochromes initiate tran-
scriptional cascades by modulating the activities of these
124 ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd
The Plant Journal (2006) 47, 124–139 doi: 10.1111/j.1365-313X.2006.02773.x

transcription factors at the transcriptional or post-transla-
tional levels (Bauer et al., 2004; Duek et al., 2004; Fairchild
et al., 2000; Holm et al., 2002; Huq and Quail, 2002;
Jang et al., 2005; Oyama et al., 1997; Park et al., 2003,
2004; Salter et al., 2003; Seo et al., 2003; Yamashino et al.,
2003; Yang et al., 2005).
Different signalling components regulate different light
responses. The best studied of these is the inhibition of
hypocotyl elongation by phytochromes, in which a large
proportion of the light signalling components interact in
varying manners. Some signalling components regulate
both PHYA- and PHYB-mediated inhibition of hypocotyl
elongation (e.g. HY5); some regulate only PHYB-mediated
inhibition of hypocotyl elongation (e.g. PIF3); while others
regulate only PHYA-mediated inhibition of hypocotyl elon-
gation (e.g. HFR1; Fairchild et al., 2000; Fankhauser and
Chory, 2000; Kim et al., 2003; Koornneef et al., 1980; Soh
et al., 2000). The exact molecular pathway that leads to the
inhibition of hypocotyl elongation is not clear, but microar-
ray analysis suggests that this process involves expressional
changes of various genes related to the cell-wall metabolism
(Ma et al., 2001). However, while the hypocotyl elongation
process is relatively well understood, only a few signalling
components have been identified in other important light
responses, such as seed germination.
Seed germination is regulated by various factors, inclu-
ding abscisic acid (ABA), brassinosteroids (BR), ethylene
and gibberellin (GA; Chiwocha et al., 2005; Debeaujon
and Koornneef, 2000; Kepczynski and Kepczynska, 1997;
Koornneef and van der Veen, 1980; Koornneef et al., 2002;
Leubner-Metzger, 2001; Steber and McCourt, 2001). Among
these, ABA and GA play antagonistic roles. During seed
maturation, ABA levels increase and seed dormancy is
established (Karssen et al., 1983). When the dormant seed is
transferred to conditions favourable for germination, the
level of ABA decreases and de novo GA biosynthesis
commences, disrupting dormancy and triggering germina-
tion (Ogawa et al., 2003). ABA biosynthetic mutants such as
aba1 and aba2 display reduced seed dormancy, while GA
biosynthetic mutants such as ga1 are unable to germinate
even under favourable conditions (Karssen et al., 1983;
Koornneef and van der Veen, 1980; Marin et al., 1996). The
importance of GA during seed germination was further
shown in various GA-signalling mutants. A loss-of-function
mutation in RGL2, a negative regulator of GA responses,
allowed plants to germinate even in the absence of de novo
GA biosynthesis, as did a mutation in SPINDLY, a Ser/Thr
O-linked N-acetyl glucosamine (O-GlcNAc) transferase that
also functions as a negative regulator of GA signalling
(Jacobsen and Olszewski, 1993; Jacobsen et al., 1996; Lee
et al., 2002; Tyler et al., 2004). Light-independent germina-
tion of a ga1 rgl2 rga gai quadruple mutant further suggests
that GAI and RGA, homologues of RGL2, also play roles in
seed germination (Cao et al., 2005). Collectively, these
previous results indicate that ABA and GA biosynthesis
are critical for seed dormancy and seed germination
respectively.
Various external factors, such as light, also play critical
roles in regulating seed germination. The promotion of seed
germination by light was noted as early as the 19th century,
and reversible regulation of lettuce seed germination by red
and far-red light is reported by Borthwick et al. (1952). Later,
studies in Arabidopsis mutants revealed that this reversible
regulation of seed germination by red and far-red light was
determined by phytochromes (Hennig et al., 2002; Shinom-
ura et al., 1994). A direct connection between phytochrome
signalling and de novo GA biosynthesis during seed germi-
nation was demonstrated by inhibition of light-induced seed
germination in the presence of a GA biosynthesis inhibitor;
by the insensitivity of the ga1 mutant to light induction; by
the epistasis of the ga1 mutation to the phyB mutation (Peng
and Harberd, 1997); and by the direct determination of
increased GA levels after light induction (Hilhorst and
Karssen, 1988; Koornneef and van der Veen, 1980; Ogawa
et al., 2003). The increased de novo GA biosynthesis during
light-induced germination of Arabidopsis seeds is due to
increased expression of GA biosynthetic enzymes such as
GA 3b-hydroxylase (Yamaguchi et al., 1998). It seems likely
that some of the phytochrome-interacting proteins trans-
duce light signals, leading to activation of GA biosynthetic
gene transcription during seed germination.
We showed previously that PIF3-Like 5 (PIL5/PIF1/
bHLH015), a phytochrome-interacting bHLH protein, is a
key negative regulator of seed germination, and that light
promotes seed germination partly by inhibiting the function
of PIL5 (Oh et al., 2004). Here we demonstrate that PIL5
mediates seed germination by simultaneously regulating
the expression of GA biosynthetic and catabolic genes, and
also show that light inhibits PIL5 function by activating the
phytochrome-mediated degradation of the PIL5 protein.
Results
PIL5 regulates seed germination through gibberellin
Our previous work showed that PIL5 is a key negative
regulator in PHY-mediated promotion of seed germination
(Oh et al., 2004). As phytochromes are known to promote
seed germination by activating de novo GA biosynthesis, we
examined whether PIL5 negatively regulates seed germina-
tion by repressing de novo GA biosynthesis (Yamaguchi
et al., 1998). We first tested the effects of paclobutrazol, an
inhibitor of GA biosynthesis, on the ability of the pil5 mutant
to germinate irrespective of light conditions.
For the PHYB-dependent germination assay, we irradiated
seeds with a far-red light pulse and transferred them directly
to the dark with or without a second irradiation with a red
light pulse. The far-red light pulse inactivated PHYB and
PIL5 regulates seed germination via GA 125
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139
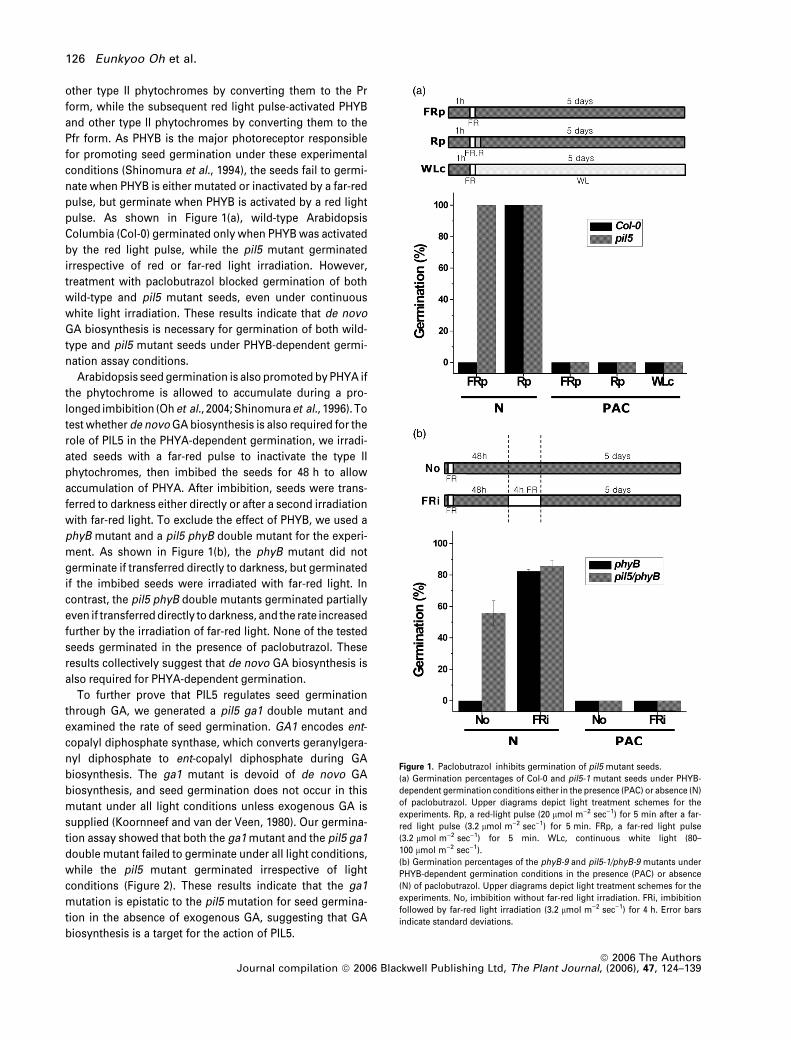
other type II phytochromes by converting them to the Pr
form, while the subsequent red light pulse-activated PHYB
and other type II phytochromes by converting them to the
Pfr form. As PHYB is the major photoreceptor responsible
for promoting seed germination under these experimental
conditions (Shinomura et al., 1994), the seeds fail to germi-
nate when PHYB is either mutated or inactivated by a far-red
pulse, but germinate when PHYB is activated by a red light
pulse. As shown in Figure 1(a), wild-type Arabidopsis
Columbia (Col-0) germinated only when PHYB was activated
by the red light pulse, while the pil5 mutant germinated
irrespective of red or far-red light irradiation. However,
treatment with paclobutrazol blocked germination of both
wild-type and pil5 mutant seeds, even under continuous
white light irradiation. These results indicate that de novo
GA biosynthesis is necessary for germination of both wild-
type and pil5 mutant seeds under PHYB-dependent germi-
nation assay conditions.
Arabidopsis seed germination is also promoted by PHYA if
the phytochrome is allowed to accumulate during a pro-
longed imbibition (Ohet al., 2004; Shinomuraet al., 1996). To
test whether de novo GA biosynthesis is also required for the
role of PIL5 in the PHYA-dependent germination, we irradi-
ated seeds with a far-red pulse to inactivate the type II
phytochromes, then imbibed the seeds for 48 h to allow
accumulation of PHYA. After imbibition, seeds were trans-
ferred to darkness either directly or after a second irradiation
with far-red light. To exclude the effect of PHYB, we used a
phyB mutant and a pil5 phyB double mutant for the experi-
ment. As shown in Figure 1(b), the phyB mutant did not
germinate if transferred directly to darkness, but germinated
if the imbibed seeds were irradiated with far-red light. In
contrast, the pil5 phyB double mutants germinated partially
even if transferred directly to darkness, and the rate increased
further by the irradiation of far-red light. None of the tested
seeds germinated in the presence of paclobutrazol. These
results collectively suggest that de novo GA biosynthesis is
also required for PHYA-dependent germination.
To further prove that PIL5 regulates seed germination
through GA, we generated a pil5 ga1 double mutant and
examined the rate of seed germination. GA1 encodes ent-
copalyl diphosphate synthase, which converts geranylgera-
nyl diphosphate to ent-copalyl diphosphate during GA
biosynthesis. The ga1 mutant is devoid of de novo GA
biosynthesis, and seed germination does not occur in this
mutant under all light conditions unless exogenous GA is
supplied (Koornneef and van der Veen, 1980). Our germina-
tion assay showed that both the ga1 mutant and the pil5 ga1
double mutant failed to germinate under all light conditions,
while the pil5 mutant germinated irrespective of light
conditions (Figure 2). These results indicate that the ga1
mutation is epistatic to the pil5 mutation for seed germina-
tion in the absence of exogenous GA, suggesting that GA
biosynthesis is a target for the action of PIL5.
Figure 1. Paclobutrazol inhibits germination of pil5 mutant seeds.
(a) Germination percentages of Col-0 and pil5-1 mutant seeds under PHYB-
dependent germination conditions either in the presence (PAC) or absence (N)
of paclobutrazol. Upper diagrams depict light treatment schemes for the
experiments. Rp, a red-light pulse (20 lmol m)2 sec)1) for 5 min after a far-
red light pulse (3.2 lmol m)2 sec)1) for 5 min. FRp, a far-red light pulse
(3.2 lmol m)2 sec)1) for 5 min. WLc, continuous white light (80–
100 lmol m)2 sec)1).
(b) Germination percentages of the phyB-9 and pil5-1/phyB-9 mutants under
PHYB-dependent germination conditions in the presence (PAC) or absence
(N) of paclobutrazol. Upper diagrams depict light treatment schemes for the
experiments. No, imbibition without far-red light irradiation. FRi, imbibition
followed by far-red light irradiation (3.2 lmol m)2 sec)1) for 4 h. Error bars
indicate standard deviations.
126 Eunkyoo Oh et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139
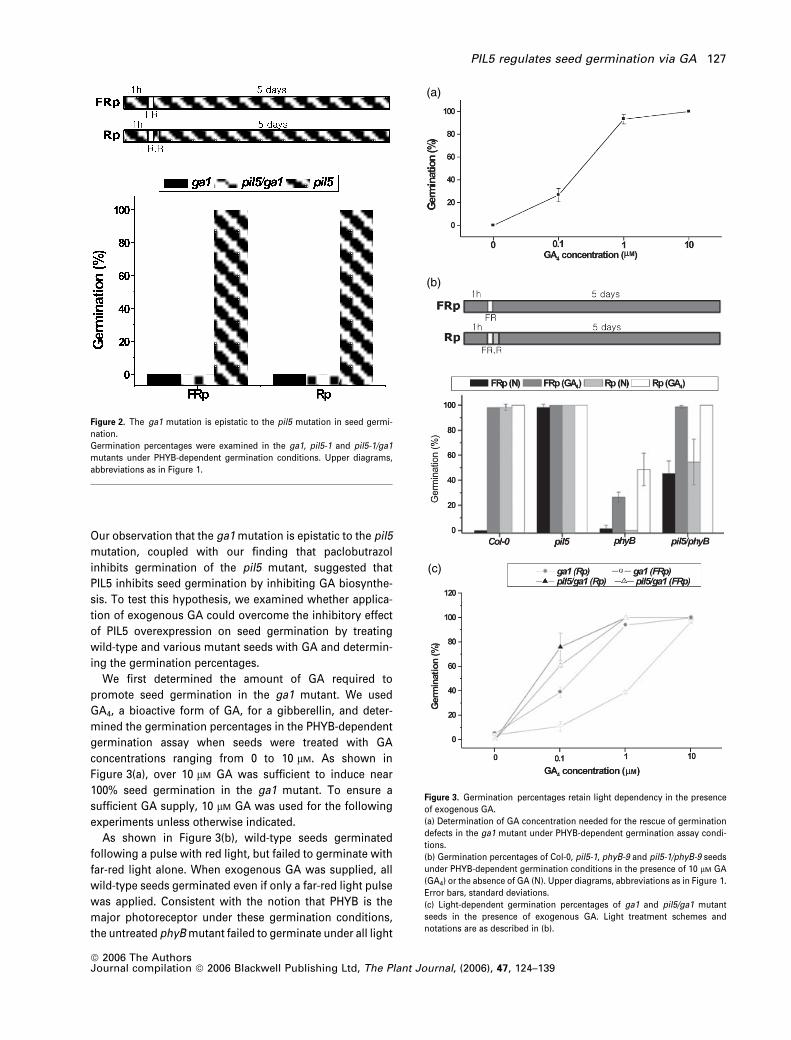
Our observation that the ga1 mutation is epistatic to the pil5
mutation, coupled with our finding that paclobutrazol
inhibits germination of the pil5 mutant, suggested that
PIL5 inhibits seed germination by inhibiting GA biosynthe-
sis. To test this hypothesis, we examined whether applica-
tion of exogenous GA could overcome the inhibitory effect
of PIL5 overexpression on seed germination by treating
wild-type and various mutant seeds with GA and determin-
ing the germination percentages.
We first determined the amount of GA required to
promote seed germination in the ga1 mutant. We used
GA4, a bioactive form of GA, for a gibberellin, and deter-
mined the germination percentages in the PHYB-dependent
germination assay when seeds were treated with GA
concentrations ranging from 0 to 10 lM. As shown in
Figure 3(a), over 10 lM GA was sufficient to induce near
100% seed germination in the ga1 mutant. To ensure a
sufficient GA supply, 10 lM GA was used for the following
experiments unless otherwise indicated.
As shown in Figure 3(b), wild-type seeds germinated
following a pulse with red light, but failed to germinate with
far-red light alone. When exogenous GA was supplied, all
wild-type seeds germinated even if only a far-red light pulse
was applied. Consistent with the notion that PHYB is the
major photoreceptor under these germination conditions,
the untreated phyB mutant failed to germinate under all light
(a)
(b)
(c)
Figure 3. Germination percentages retain light dependency in the presence
of exogenous GA.
(a) Determination of GA concentration needed for the rescue of germination
defects in the ga1 mutant under PHYB-dependent germination assay condi-
tions.
(b) Germination percentages of Col-0, pil5-1, phyB-9 and pil5-1/phyB-9 seeds
under PHYB-dependent germination conditions in the presence of 10 lM GA
(GA4) or the absence of GA (N). Upper diagrams, abbreviations as in Figure 1.
Error bars, standard deviations.
(c) Light-dependent germination percentages of ga1 and pil5/ga1 mutant
seeds in the presence of exogenous GA. Light treatment schemes and
notations are as described in (b).
Figure 2. The ga1 mutation is epistatic to the pil5 mutation in seed germi-
nation.
Germination percentages were examined in the ga1, pil5-1 and pil5-1/ga1
mutants under PHYB-dependent germination conditions. Upper diagrams,
abbreviations as in Figure 1.
PIL5 regulates seed germination via GA 127
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139
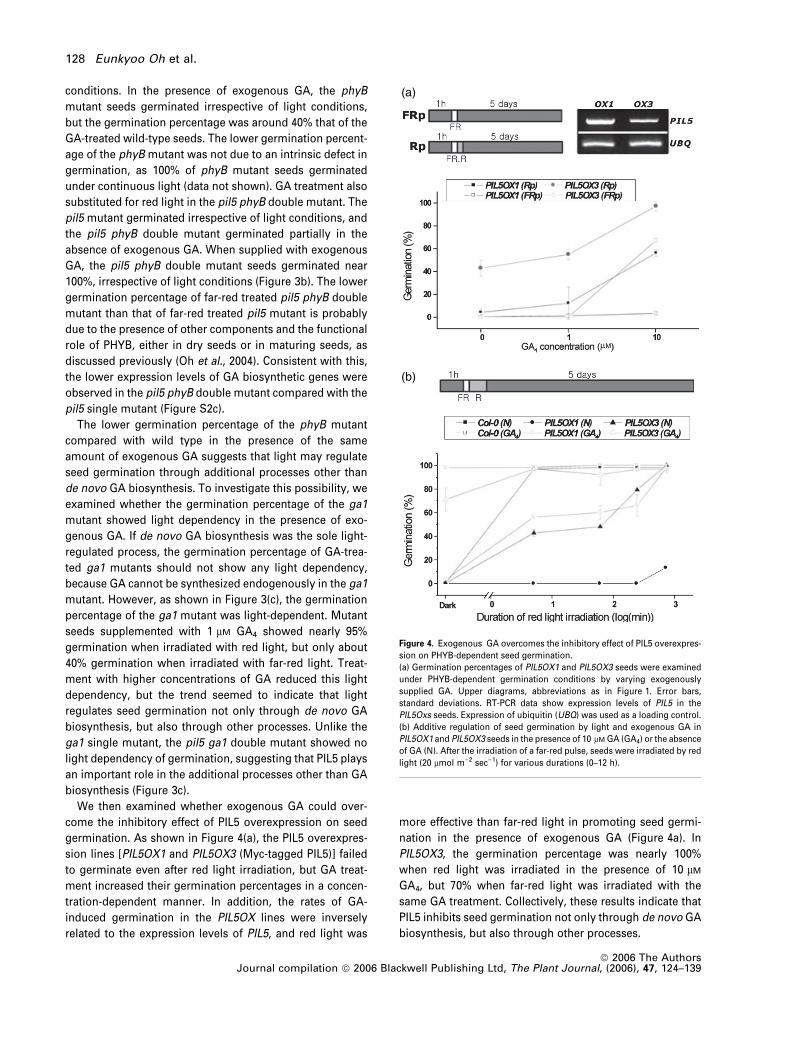
conditions. In the presence of exogenous GA, the phyB
mutant seeds germinated irrespective of light conditions,
but the germination percentage was around 40% that of the
GA-treated wild-type seeds. The lower germination percent-
age of the phyB mutant was not due to an intrinsic defect in
germination, as 100% of phyB mutant seeds germinated
under continuous light (data not shown). GA treatment also
substituted for red light in the pil5 phyB double mutant. The
pil5 mutant germinated irrespective of light conditions, and
the pil5 phyB double mutant germinated partially in the
absence of exogenous GA. When supplied with exogenous
GA, the pil5 phyB double mutant seeds germinated near
100%, irrespective of light conditions (Figure 3b). The lower
germination percentage of far-red treated pil5 phyB double
mutant than that of far-red treated pil5 mutant is probably
due to the presence of other components and the functional
role of PHYB, either in dry seeds or in maturing seeds, as
discussed previously (Oh et al., 2004). Consistent with this,
the lower expression levels of GA biosynthetic genes were
observed in the pil5 phyB double mutant compared with the
pil5 single mutant (Figure S2c).
The lower germination percentage of the phyB mutant
compared with wild type in the presence of the same
amount of exogenous GA suggests that light may regulate
seed germination through additional processes other than
de novo GA biosynthesis. To investigate this possibility, we
examined whether the germination percentage of the ga1
mutant showed light dependency in the presence of exo-
genous GA. If de novo GA biosynthesis was the sole light-
regulated process, the germination percentage of GA-trea-
ted ga1 mutants should not show any light dependency,
because GA cannot be synthesized endogenously in the ga1
mutant. However, as shown in Figure 3(c), the germination
percentage of the ga1 mutant was light-dependent. Mutant
seeds supplemented with 1 lM GA4 showed nearly 95%
germination when irradiated with red light, but only about
40% germination when irradiated with far-red light. Treat-
ment with higher concentrations of GA reduced this light
dependency, but the trend seemed to indicate that light
regulates seed germination not only through de novo GA
biosynthesis, but also through other processes. Unlike the
ga1 single mutant, the pil5 ga1 double mutant showed no
light dependency of germination, suggesting that PIL5 plays
an important role in the additional processes other than GA
biosynthesis (Figure 3c).
We then examined whether exogenous GA could over-
come the inhibitory effect of PIL5 overexpression on seed
germination. As shown in Figure 4(a), the PIL5 overexpres-
sion lines [PIL5OX1 and PIL5OX3 (Myc-tagged PIL5)] failed
to germinate even after red light irradiation, but GA treat-
ment increased their germination percentages in a concen-
tration-dependent manner. In addition, the rates of GA-
induced germination in the PIL5OX lines were inversely
related to the expression levels of PIL5, and red light was
more effective than far-red light in promoting seed germi-
nation in the presence of exogenous GA (Figure 4a). In
PIL5OX3, the germination percentage was nearly 100%
when red light was irradiated in the presence of 10 lM
GA4, but 70% when far-red light was irradiated with the
same GA treatment. Collectively, these results indicate that
PIL5 inhibits seed germination not only through de novo GA
biosynthesis, but also through other processes.
(a)
(b)
Figure 4. Exogenous GA overcomes the inhibitory effect of PIL5 overexpres-
sion on PHYB-dependent seed germination.
(a) Germination percentages of PIL5OX1 and PIL5OX3 seeds were examined
under PHYB-dependent germination conditions by varying exogenously
supplied GA. Upper diagrams, abbreviations as in Figure 1. Error bars,
standard deviations. RT-PCR data show expression levels of PIL5 in the
PIL5Oxs seeds. Expression of ubiquitin (UBQ) was used as a loading control.
(b) Additive regulation of seed germination by light and exogenous GA in
PIL5OX1 and PIL5OX3 seeds in the presence of 10 lM GA (GA4) or the absence
of GA (N). After the irradiation of a far-red pulse, seeds were irradiated by red
light (20 lmol m)2 sec)1) for various durations (0–12 h).
128 Eunkyoo Oh et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

As we previously reported that the PIL5OX lines required a
longer irradiation time for germination, we next examined
whether GA treatment altered the required irradiation time
for germination (Oh et al., 2004). We tested the germination
percentages of PIL5OX seeds in the presence or absence of
GA coupled with red light irradiations of various durations.
As shown in Figure 4(b), 5 min red light irradiation was
sufficient to fully induce germination in wild-type seeds,
while much longer irradiation times were required for the
PIL5OXs. Wild-type seeds germinated even in the absence of
red light irradiation if exogenous GA was supplied. In the
case of PIL5OX3, GA treatment reduced the required irradi-
ation time to achieve 100% germination from 720 to 5 min,
indicating that the negative role of PIL5 on seed germination
is overcome additively by light and GA.
As PIL5 is also a negative regulator of PHYA-dependent
seed germination, we investigated whether GA treatment
could overcome the inhibitory role of PIL5 on PHYA-
dependent seed germination by determining the germina-
tion percentages of wild-type and various mutant seeds in
the presence or absence of exogenous GA. In the absence of
GA, the germination percentages of wild-type seeds in-
creased in response to longer far-red light irradiation times,
while the phyA mutant failed to germinate regardless of
irradiation time (Figure 5). In the case of the PIL5OX lines,
increased far-red light irradiation times increased the ger-
mination percentages especially in the weaker line (PI-
L5OX3). In the presence of GA, both wild-type and phyA
mutant seeds germinated well, even in the absence of far-
red light irradiation (Figure 5). In the case of PIL5OX seeds,
the germination percentage increased further if GA was
supplied. Taken together with the inhibition of PHYA-
dependent seed germination by paclobutrazol, these results
indicate that PIL5 regulates seed germination negatively
through GA also under PHYA-dependent seed germination
conditions.
PIL5 regulates both GA synthesis and GA degradation
PHY-mediated light signalling induces de novo GA biosyn-
thesis by increasing the expression of two GA 3b-hydroxy-
lases that convert biologically inactive GA9 to biologically
active GA4 (Yamaguchi et al., 1998). As PIL5 is a phyto-
chrome-interacting protein that negatively regulates seed
germination through GA, we investigated the expression of
GA-biosynthesis genes in both the pil5 mutant and PIL5OX
seeds (Oh et al., 2004). For the experiment, various mutant
seeds were irradiated with a far-red pulse with or without a
subsequent red light pulse and incubated in the dark for 12 h
before sampling. This experimental condition is identical to
the PHYB-dependent germination assay condition, except
that seeds were sampled for expression analysis 12 h after
irradiation.
We first examined the expression of two representative
marker genes (EXP1 and CP1, encoding expansin 1 and
cystein proteinase 1 respectively) that are induced by GA
during seed germination (Ogawa et al., 2003; Yamauchi
et al., 2004). The wild-type seeds showed induced marker
gene expression when PHYB was activated by a red light
pulse (inductive condition), but not when PHYB was inacti-
vated by a far-red light pulse (non-inductive condition)
(Figure 6). The pil5 mutant seeds, which germinated irres-
pective of light conditions, showed increased expression of
two marker genes irrespective of light conditions. The
PIL5OX seeds, which failed to germinate under all light
conditions, showed lower levels of EXP1 and CP1 irrespect-
ive of light conditions. These results indicated that the
expression patterns of the two GA-inducible marker genes
are consistent with the germination patterns of the various
mutants.
We next determined the expression of two GA 3b-
hydroxylase genes (GA3ox1 and GA3ox2). As shown in
Figure 6, both GA3ox1 and GA3ox2 were expressed in
wild-type seeds following irradiation with red light but not
far-red light. In the pil5 mutant, GA3ox1 and GA3ox2
expression was constitutively high, irrespective of light
conditions. In contrast, no GA3ox1 and GA3ox2 expres-
sion was detected in the PIL5OX mutants, irrespective of
light conditions. Overall, the expression patterns of
GA3ox1 and GA3ox2 were very similar to those of the
two GA-inducible marker genes, collectively suggesting
that PIL5 negatively regulates seed germination, at least
partly by repressing the expression of GA biosynthetic
genes.
Figure 5. Exogenous GA overcomes the inhibitory effect of PIL5 overexpres-
sion on PHYA-dependent seed germination.
Germination percentages of Col-0, pil5-1, phyA-211, pil5-1/phyA-211, PIL5OX1
and PIL5OX3 seeds under PHYA-dependent germination conditions in the
presence of 10 lM GA (GA4) or in the absence of GA (N). After irradiation of a
far-red light pulse, seeds were imbibed for 48 h, then irradiated by far-red
light (3.2 lmol m)2 sec)1) for 0, 4 or 12 h. Error bars, standard deviations.
PIL5 regulates seed germination via GA 129
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

Our germination analyses indicated that both light and
PIL5 regulate seed germination through additional proces-
ses other than GA biosynthesis. One candidate process is
the degradation of biologically active GA. During seed
germination, the level of a bioactive GA shows an initial
rapid increase and reaches a subsequent plateau concom-
itant with the increase of an inactive GA, suggesting that
both synthesis and degradation of biologically active GA
occur during seed germination (Ogawa et al., 2003). Thus it
is possible that PIL5 inhibits seed germination not only by
repressing the expression of GA biosynthetic genes, but also
by activating the expression of GA catabolic genes.
To investigate this possibility, we examined the GA2ox
genes, which encode the GA 2-oxidases responsible for
converting biologically active GA4 and/or its precursors to
biologically inactive forms. Of the eight GA2ox genes
present in the Arabidopsis genome, we focused on the
expression of GA2ox2, because it was expressed at high
levels and its expression was light-dependent in germina-
ting seeds (data not shown). As shown in Figure 6(a),
GA2ox2 was highly expressed in seeds that failed to
germinate: far-red light-irradiated wild-type seeds, and both
red and far-red light-irradiated PIL5OX seeds. In contrast,
GA2ox2 was expressed at low levels in seeds that germina-
ted well: red light-irradiated wild-type seeds, and both red
and far-red light-irradiated pil5 mutant seeds. Taken
together, these results indicate that PIL5 negatively regu-
lates the biosynthesis of bioactive GA in germinating seeds,
not only by repressing GA3ox1 and GA3ox2, but also by
activating the expression of GA2ox2.
Although GA2ox2 was expressed at high levels in PIL5OX
mutant seeds, irrespective of light, the expression level of
GA2ox2 was higher in far-red light-irradiated PIL5OX seeds
than in red light-irradiated PIL5OX seeds (Figure 6a). This
light-dependent expression of GA2ox2 in the PIL5OX seeds
might partly explain the light-dependent germination per-
centage of PIL5OX in the presence of exogenous GA
(Figure 4a). Similarly, light-dependent germination was also
observed in the ga1 mutant in the presence of exogenous
GA (Figure 3c), although the pil5 ga1 double mutant did not
show any light dependency. To investigate whether light-
dependent expression of GA2ox2 is also partly responsible
for the light-dependent germination percentages of these
mutants, we determined the expression of GA2ox2 in the
ga1 mutant and the pil5 ga1 double mutant. As shown in
Figure 6(b), GA2ox2 expression was higher in the far-red
light-irradiated ga1 mutant seeds than in the red light-
irradiated ga1 mutant seeds. In the pil5 ga1 double mutant,
GA2ox2 expression was slightly higher in the far-red light-
irradiated seeds, but the overall expression level was lower
than that of far-red light-irradiated ga1 seeds. As GA 2-
oxidase degrades biologically active GAs, these results
suggest that the light-dependent expression of GA2ox2
partly explains the light-dependent germination percent-
ages of PIL5OXs and ga1 mutant seeds in the presence of
exogenous GA (Thomas et al., 1999).
To investigate the relationship between PIL5 and PHYB in
seed germination, we determined the expression levels of
GA3ox1, GA3ox2 and GA2ox2 in the phyB mutant and in
the pil5 phyB double mutant. Under PHYB-dependent
(a)
(b)
(c)
Figure 6. PIL5 regulates GA biosynthesis by repressing the expression of GA
3b-hydroxylase genes (GA3ox1 and GA3ox2) and by activating the expression
of a GA 2-oxidase gene (GA2ox2) under PHYB-dependent germination
conditions (for quantitive RT-PCR data see Figure S2).
(a) Expression patterns of GA3ox1, GA3ox2, GA2ox2, EXP1 and CP1 in Col-0,
pil5-1 and PIL5OX1 seeds under PHYB-dependent germination conditions.
Upper diagrams, abbreviations as in Figure 1. Expression of S18 was used as
a loading control. Germination percentages indicated below figure.
(b) Light-dependent expression of GA2ox2 in ga1 and pil5/ga1 mutant seeds.
Light treatment scheme and notations are as described in (a).
(c) Expression patterns of GA3ox1, GA3ox2 and GA2ox2 in Col-0, pil5-1,
PIL5OX1, phyB-9 and pil5-1/phyB-9 seeds.
130 Eunkyoo Oh et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

germination assay conditions, the phyB and PIL5OX seeds
failed to germinate, while the pil5 phyB double mutant
germinated only partially (Figure 3b). Consistent with these
findings, seeds that germinated under the assay conditions
(wild type and pil5 mutant) showed increased GA3ox1 and
GA3ox2 expression levels, but decreased expression levels
of GA2ox2 (Figure 6c), while seeds that failed to germinate
(PIL5OX and phyB mutant) showed decreased GA3ox1 and
GA3ox2 expression and increased GA2ox2 expression. The
pil5 phyB double mutant showed intermediate expression
levels of both GA biosynthetic and GA catabolic genes,
which is consistent with its intermediate germination per-
centage. These data indicate that both PHYB and PIL5
regulate bioactive GA levels during seed germination via
mediating both synthesis and degradation of GA. As PIL5 is
a PHYB-interacting, downstream-signalling component,
these data further suggest that PHYB promotes seed germi-
nation partly by inhibiting the negative role of PIL5 in the
biosynthesis of bioactive GA.
As PIL5 is also a negative regulator of seed germination
under PHYA-dependent germination conditions, we investi-
gated whether PIL5 regulates the expression levels of
GA3ox1, GA3ox2 and GA2ox2 under these conditions. For
the experiment, far-red-irradiated seeds were imbibed for
48 h to allow the accumulation of PHYA. After imbibition,
far-red light was applied again to activate PHYA, and the
seeds were incubated for 8 h in the dark before sampling.
Under the PHYA-dependent germination assay conditions,
wild-type, pil5 mutant and pil5 phyA double mutant seeds
germinated following far-red irradiation, but PIL5OX and
phyA mutant seeds did not. Consistent with our findings
under PHYB-dependent germination conditions, the GA-
inducible marker genes (EXP1 and CP1) were highly
expressed in seeds that germinated under PHYA-dependent
germination conditions, but were expressed at low levels in
seeds that did not germinate (Figure 7a). The seeds that
germinated showed higher expression levels of GA3ox1 and
GA3ox2, but lower expression levels of GA2ox2. In contrast,
the seeds that failed to germinate showed lower expression
levels of GA3ox1 and GA3ox2, but higher expression levels
of GA2ox2 (Figure 7a,b). These data indicate that PIL5
regulates both GA synthesis and degradation under PHYA-
dependent germination conditions. Our data suggest that
PHYA promotes seed germination partly by inhibiting the
negative role of PIL5 on the biosynthesis of bioactive GA,
being similar to the relationship observed between PIL5 and
PHYB. In addition, the higher expression of GA biosynthetic
genes in the pil5 single mutant compared with the pil5 phyA
double mutant after inductive FR irradiation further suggests
the presence of other factors that mediate PHYA signalling
to GA biosynthesis.
Our expression analysis of GA biosynthetic genes
suggested that the amount of bioactive GA was likely to
be increased in the pil5 mutant and decreased in PIL5OX
plants. To confirm this, we irradiated various mutant
seeds with far-red pulses with or without subsequent red
light pulses, and quantified the GA4 contents. As shown
in Figure 8, the GA4 level was twofold higher in red light-
treated wild-type seeds versus far-red light-treated wild-
type seeds. In the pil5 mutant, GA4 levels were similar to
those found in red-light treated wild-type seeds, regard-
less of light conditions. In PIL5OX plants, the GA4 level
(a)
(b)
Figure 7. PIL5 regulates GA biosynthesis by repressing the expression of GA
3b-hydroxylase genes (GA3ox1 and GA3ox2) and activating the expression of
a GA 2-oxidase gene (GA2ox2) under PHYA-dependent germination condi-
tions (for quantitive RT–PCR data see Figure S3).
(a) Expression patterns of GA3ox1, GA3ox2, GA2ox2, EXP1 and CP1 in Col-0,
pil5-1 and PIL5OX1 seeds under PHYA-dependent germination conditions.
After irradiation of a far-red light pulse (3.2 lmol m)2 sec)1), seeds were
imbibed for 48 h, then irradiated with far-red light (3.2 lmol m)2 sec)1) for 4 h
(FR) and transferred to the dark, or left unirradiated (N) and transferred to the
dark. Expression of S18 was used as a loading control. Germination
percentages indicated below figure.
(b) Expression patterns of GA3ox1, GA3ox2 and GA2ox2 in Col-0, pil5-1,
PIL5OX1, phyA-211 and pil5-1/phyA-211 seeds.
PIL5 regulates seed germination via GA 131
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139
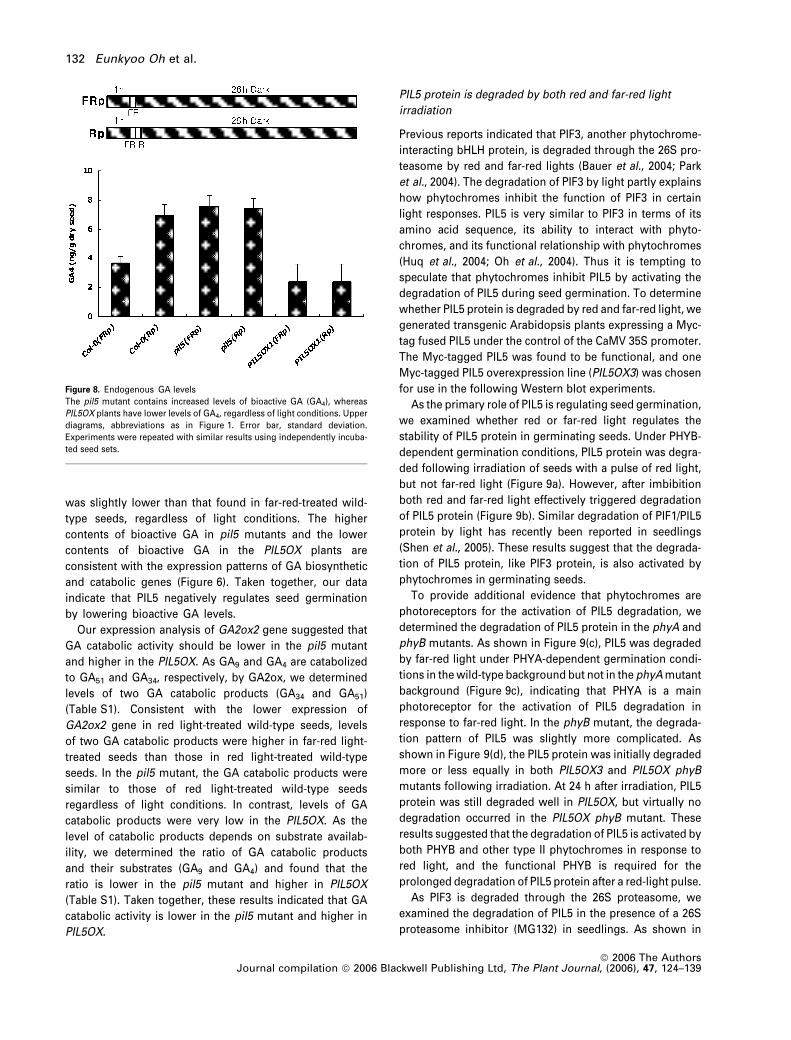
was slightly lower than that found in far-red-treated wild-
type seeds, regardless of light conditions. The higher
contents of bioactive GA in pil5 mutants and the lower
contents of bioactive GA in the PIL5OX plants are
consistent with the expression patterns of GA biosynthetic
and catabolic genes (Figure 6). Taken together, our data
indicate that PIL5 negatively regulates seed germination
by lowering bioactive GA levels.
Our expression analysis of GA2ox2 gene suggested that
GA catabolic activity should be lower in the pil5 mutant
and higher in the PIL5OX. As GA9 and GA4 are catabolized
to GA51 and GA34, respectively, by GA2ox, we determined
levels of two GA catabolic products (GA34 and GA51)
(Table S1). Consistent with the lower expression of
GA2ox2 gene in red light-treated wild-type seeds, levels
of two GA catabolic products were higher in far-red light-
treated seeds than those in red light-treated wild-type
seeds. In the pil5 mutant, the GA catabolic products were
similar to those of red light-treated wild-type seeds
regardless of light conditions. In contrast, levels of GA
catabolic products were very low in the PIL5OX. As the
level of catabolic products depends on substrate availab-
ility, we determined the ratio of GA catabolic products
and their substrates (GA9 and GA4) and found that the
ratio is lower in the pil5 mutant and higher in PIL5OX
(Table S1). Taken together, these results indicated that GA
catabolic activity is lower in the pil5 mutant and higher in
PIL5OX.
PIL5 protein is degraded by both red and far-red light
irradiation
Previous reports indicated that PIF3, another phytochrome-
interacting bHLH protein, is degraded through the 26S pro-
teasome by red and far-red lights (Bauer et al., 2004; Park
et al., 2004). The degradation of PIF3 by light partly explains
how phytochromes inhibit the function of PIF3 in certain
light responses. PIL5 is very similar to PIF3 in terms of its
amino acid sequence, its ability to interact with phyto-
chromes, and its functional relationship with phytochromes
(Huq et al., 2004; Oh et al., 2004). Thus it is tempting to
speculate that phytochromes inhibit PIL5 by activating the
degradation of PIL5 during seed germination. To determine
whether PIL5 protein is degraded by red and far-red light, we
generated transgenic Arabidopsis plants expressing a Myc-
tag fused PIL5 under the control of the CaMV 35S promoter.
The Myc-tagged PIL5 was found to be functional, and one
Myc-tagged PIL5 overexpression line (PIL5OX3) was chosen
for use in the following Western blot experiments.
As the primary role of PIL5 is regulating seed germination,
we examined whether red or far-red light regulates the
stability of PIL5 protein in germinating seeds. Under PHYB-
dependent germination conditions, PIL5 protein was degra-
ded following irradiation of seeds with a pulse of red light,
but not far-red light (Figure 9a). However, after imbibition
both red and far-red light effectively triggered degradation
of PIL5 protein (Figure 9b). Similar degradation of PIF1/PIL5
protein by light has recently been reported in seedlings
(Shen et al., 2005). These results suggest that the degrada-
tion of PIL5 protein, like PIF3 protein, is also activated by
phytochromes in germinating seeds.
To provide additional evidence that phytochromes are
photoreceptors for the activation of PIL5 degradation, we
determined the degradation of PIL5 protein in the phyA and
phyB mutants. As shown in Figure 9(c), PIL5 was degraded
by far-red light under PHYA-dependent germination condi-
tions in the wild-type background but not in the phyA mutant
background (Figure 9c), indicating that PHYA is a main
photoreceptor for the activation of PIL5 degradation in
response to far-red light. In the phyB mutant, the degrada-
tion pattern of PIL5 was slightly more complicated. As
shown in Figure 9(d), the PIL5 protein was initially degraded
more or less equally in both PIL5OX3 and PIL5OX phyB
mutants following irradiation. At 24 h after irradiation, PIL5
protein was still degraded well in PIL5OX, but virtually no
degradation occurred in the PIL5OX phyB mutant. These
results suggested that the degradation of PIL5 is activated by
both PHYB and other type II phytochromes in response to
red light, and the functional PHYB is required for the
prolonged degradation of PIL5 protein after a red-light pulse.
As PIF3 is degraded through the 26S proteasome, we
examined the degradation of PIL5 in the presence of a 26S
proteasome inhibitor (MG132) in seedlings. As shown in
Figure 8. Endogenous GA levels
The pil5 mutant contains increased levels of bioactive GA (GA4), whereas
PIL5OX plants have lower levels of GA4, regardless of light conditions. Upper
diagrams, abbreviations as in Figure 1. Error bar, standard deviation.
Experiments were repeated with similar results using independently incuba-
ted seed sets.
132 Eunkyoo Oh et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

Figures 9(e) and 8(f), the degradation of PIL5 by red and far-
red lights was greatly reduced in the presence of MG132,
suggesting that PIL5 is also degraded through the 26S
proteasome both under red and far-red lights. MG132 had a
much weaker effect on PIL5 degradation in seeds, when
compared with seedlings, possibly due to lower permeation
by this chemical (Figure S1). Taken together, our results
indicate that phytochromes act as photoreceptors to activate
the degradation of PIL5 protein through the 26S proteasome.
Discussion
We report here that PIL5, a phytochrome-interacting bHLH
protein, negatively regulates seed germination by repress-
ing two GA 3b-hydroxylases genes (GA3ox1 and GA3ox2)
and activating a GA 2-oxidase gene (GA2ox2) at the tran-
scriptional level. Repression of GA biosynthetic genes and
activation of GA catabolic genes by PIL5 leads to increased
levels of bioactive GA in the pil5 mutant and decreased
levels of bioactive GA in PIL5OX seeds, regardless of light
conditions. Our functional relationship studies further
suggest that phytochromes promote seed germination by
inhibiting PIL5, at least partly via activation of PIL5 protein
degradation through the 26S proteasome. Based on these
results, we propose a photogermination model in which
PIL5 plays a negative role in the biosynthesis of bioactive
GA, and PIL5 degradation by phytochromes is activated in
response to inductive light signals (Figure 10).
GA synthesis and degradation are regulated during
phytochrome-mediated promotion of seed germination
De novo GA biosynthesis plays a critical role in the phyto-
chrome-mediated promotion of seed germination, as shown
by the ability of a GA biosynthetic inhibitor (paclobutrazol)
to inhibit seed germination, and the fact that treatment with
exogenous GA could initiate germination in the absence of
light stimulation (Hilhorst and Karssen, 1988; Yang et al.,
1995). The observation that GA biosynthetic mutants such as
ga1 failed to germinate even after inductive light treatment
further supports the role of GA in the phytochrome-medi-
ated promotion of seed germination (Koornneef and van der
(a) (b)
(d)
(c)
(e) (f)
Figure 9. Phytochromes activate the degrada-
tion of PIL5 protein.
(a) Degradation of PIL5 protein by red or far-red
light under PHYB-dependent germination condi-
tions. PIL5 indicates Myc-tagged PIL5 protein
detected by an anti-Myc antibody. Tubulin (TUB)
was detected by anti-tubulin antibody as a load-
ing control.
(b) Degradation of PIL5 protein by red or far-red
light under PHYA-dependent germination condi-
tions.
(c) Degradation of PIL5 protein in PIL5OX3 and
PIL5OX3/phyA mutant seeds by far-red light
under PHYA-dependent germination conditions.
(d) Degradation of PIL5 protein in PIL5OX3 and
PIL5OX3/phyB mutant seeds by red light under
PHYB-dependent germination conditions.
(e) Inhibition of PIL5 degradation by MG132 in
far-red light-irradiated seedlings.
(f) Inhibition of PIL5 degradation by MG132 in red
light-irradiated seedlings. Five DAG dark-grown
seedlings were treated by either DMSO or
MG132 (200 lM), irradiated by red light pulse
for 5 min, and incubated in the dark for 30 min
until harvesting.
PIL5 regulates seed germination via GA 133
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

Veen, 1980). Previous studies have shown that GA biosyn-
thesis increases during seed germination due to the tran-
scriptional activation of GA 3b-hydroxylase genes, which
encode enzymes required for the rate-limiting step of GA
biosynthesis (Yamaguchi et al., 1998).
The present work reveals that light regulates the biosyn-
thesis of bioactive GA not only by activating the transcrip-
tion of GA biosynthetic genes, but also by suppressing the
transcription of at least one GA catabolic gene. Expression of
two GA 3b-hydroxylase genes (GA3ox1 and GA3ox2)
increased dramatically on the perception of inductive light
signals, whereas expression of a GA 2-oxidase gene
(GA2ox2) decreased under inductive conditions and in-
creased under non-inductive conditions. The net effect of
this reciprocal regulation of biosynthetic and catabolic
genes is probably a sharper increase or decrease of bioactive
GA levels, compared with regulation of either alone. This
reciprocal regulation was found to be under the control of
phytochromes. Under PHYB-dependent germination assay
conditions, induction of the GA 3b-hydroxylase genes and
repression of the GA 2-oxidase gene were abolished in the
phyB mutant. Similarly, under PHYA-dependent germina-
tion assay conditions, this reciprocal regulation was abol-
ished in the phyA mutant. These data indicate that both
PHYA and PHYB promote the biosynthesis of bioactive GA
during seed germination by activating expression of GA 3b-
hydroxylase genes and repressing the expression of at least
one GA 2-oxidase gene.
We further found that PIL5 plays a critical role for this
reciprocal regulation of GA synthesis and catabolism
during phytochrome-mediated promotion of seed germi-
nation. We previously showed that PIL5 is a phytochrome-
interacting bHLH protein that preferentially interacts with
the Pfr forms of both PHYA and PHYB. Our characteriza-
tions of mutant and overexpression lines revealed that
PIL5 is a key negative regulator for both PHYA- and PHYB-
dependent germination processes (Oh et al., 2004). In the
present work we found that expression of GA3ox1 and
GA3ox2 is increased in the pil5 mutant and decreased in
PIL5OX seeds, indicating that PIL5 represses expression of
these GA biosynthetic genes. Similar higher expression of
GA3ox1 and GA3ox2 in the pil5 mutant have been
reported recently by Penfield et al. (2005). In contrast,
GA2ox2 is expressed at lower levels in the pil5 mutant
and at higher levels in PIL5OX seeds, indicating that PIL5
activates expression of this GA catabolic gene. The
expression patterns of GA3ox1, GA3ox2 and GA2ox2 in
the phyB, pil5 phyB, phyA and pil5 phyA mutants further
indicated that PIL5 reciprocally regulates these genes
during phytochrome-mediated promotion of seed germi-
nation. The consequences of GA biosynthetic and cata-
bolic gene-expression patterns are reflected in the levels
of bioactive GA in germinating seeds. Consistent with the
higher expression of GA biosynthetic genes and the lower
expression of GA catabolic genes, the levels of bioactive
GA were increased and decreased in the pil5 and PIL5OX
plants, respectively, regardless of light conditions. Taken
together, these data indicate that PIL5 is a key negative
component that links phytochrome signals to the expres-
sion of GA biosynthetic and catabolic genes during seed
germination.
Although our finding that light signals trigger induction
of GA 3b-hydroxylase genes is consistent with the previ-
ous report, the relationship between phytochromes and
these genes observed in the present study differs slightly
from the earlier findings (Yamaguchi et al., 1998). The
previous study showed that PHYB controlled GA3ox2
expression, while GA3ox1 expression was controlled by
other phytochromes. However, our data clearly indicate
that these genes were not expressed in the phyB mutant
under the PHYB-dependent germination assay conditions,
showing that the expression of both GA3ox1 and GA3ox2
are both controlled by PHYB (Figure S2c). We further
showed that PHYA also regulates the expression of both
GA3ox1 and GA3ox2 under PHYA-dependent germination
assay conditions. The basis of these discrepancies is not
immediately clear, but may be related to the use of
different Arabidopsis ecotypes. The previous reports were
Figure 10. Proposed model for the phytochrome-mediated promotion of
seed germination in Arabidopsis.
PIL5, a phytochrome-interacting bHLH protein, inhibits seed germination by
repressing the transcription of GA 3b-hydroxylase genes and activating the
transcription of a GA 2-oxidase gene. Under inductive light conditions,
phytochromes activate the degradation of PIL5 through the 26S proteasome,
thus increasing the biosynthesis of bioactive GAs. Bioactive GAs promote
seed germination by activating the degradation of GAI, RGA and RGL2
through SCFSLY1,SNE. For simplicity, other factors implicated in mediating
light signalling to GA biosynthesis and the degradation of GA9 to GA51 are not
indicated in the model.
134 Eunkyoo Oh et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

based on analyses of the Landsberg erecta (Ler) ecotype,
while we utilized the Col-0 ecotype in the present work.
Differences have been reported in light signalling and
seed germination between the Ler and Col ecotypes,
suggesting the possibility that light-signalling processes
such as expression of GA3ox1 might differ between these
ecotypes (Borevitz et al., 2002; van der Schaar et al., 1997;
Wolyn et al., 2004; Yanovsky et al., 1997). It will be
interesting to determine if any of the quantitative trait
loci between Ler and Col are related to the expression of
GA biosynthetic genes, such as GA3ox1. Alternatively, the
use of slightly different protocols in the PHYB-dependent
germination assays might account for the differences
between our results and those of other groups.
It is not yet known how PIL5 reciprocally regulates the
transcription of GA biosynthetic and catabolic genes. Many
transcription factors have been shown to activate one set of
genes while repressing another, although the molecular
mechanisms for these reciprocal regulations vary (Holm
et al., 2002; Monte et al., 2004). For example, PIL5 may
activate transcription of two different downstream transcrip-
tion factors, one that activates GA2ox2 transcription and one
that represses expression of GA3ox1 and GA3ox2. Alternat-
ively, PIL5 may regulate these genes directly, with its activity
as an activator or a repressor depending on the context
created by promoters or binding cofactors. Further analysis
will be required to elucidate the reciprocal regulation of
these genes by PIL5, leading to greater understanding of
light signalling in general.
Phytochromes inhibit PIL5 by promoting its protein
degradation
Protein degradation plays a key role in PHY-mediated light
responses. Mutations in the COP/DET/FUS genes cause
constitutive photomorphogenic phenotypes, even in the
dark, and analysis of these genes indicated that the
majority of them encode proteins involved in protein
degradation (Wei and Deng, 1996). Among these, COP1
encodes a RING-finger type ubiquitin E3 ligase; COP10
encodes a ubiquitin E2 conjugation enzyme; and most of
the others encode subunits of a regulatory complex sim-
ilar to the 19S lid complex of the 26S proteasome (von
Arnim and Deng, 1993; Suzuki et al., 2002; Wei and Deng,
1996). In the light-signalling process, HFR1, HY5 and LAF1,
three positive light-signalling components, are ubiquiti-
nated by COP1 in the dark, and the ubiquitinated compo-
nents are subsequently degraded by 26S proteasome
(Duek et al., 2004; Jang et al., 2005; Osterlund et al., 2000;
Seo et al., 2003; Yang et al., 2005). Photoreceptors such as
phytochromes or cryptochromes inhibit the E3 ligase
activity of COP1 partly by excluding COP1 from the nuc-
leus (von Arnim and Deng, 1994). Thus photoreceptors
potentiate light responses that are mediated by HFR1, HY5
and LAF1 by inhibiting the degradation of these factors by
COP1.
Phytochromes also modulate the stability of directly
interacting components. PIF3, a bHLH protein, was the first
identified phytochrome-interacting protein (Ni et al., 1998).
The physiological function of PIF3 is complex, depending on
the light responses examined. For hypocotyl elongation, it
acts as a negative component in PHYB- but not PHYA-
mediated inhibition of hypocotyl elongation. For anthocy-
anin synthesis, PIF3 acts as a positive component in PHY-
mediated accumulation of anthocyanin; in terms of chloro-
plast development, PIF3 plays a positive role during the
transition from dark to light (Kim et al., 2003; Monte et al.,
2004). Expression analysis indicated that the level of PIF3
transcript is not significantly affected by light, whereas PIF3
protein is rapidly degraded by red or far-red light (Baueret al.,
2004; Park et al., 2004). This degradation is activated by PHYA
in response to far-red light, and by PHYB and PHYD in
response to red light. In both cases, PIF3 is degraded by
ubiquitination followed by degradation via the 26S protea-
some.
We show here that phytochromes also activate the
degradation of PIL5, another phytochrome-interacting
bHLH protein, through the 26S proteasome. Similarly to
PIF3, PIL5 regulates various light responses, including
seed germination, hypocotyl elongation, shoot gravitro-
pism and protochlorophyllide biosynthesis (Huq et al.,
2004; Oh et al., 2004). Genetically, phytochromes inhibit
the function of PIL5 in those PHY-mediated light
responses. Thus our data, showing the degradation of
PIL5 by red or far-red light, demonstrate a mechanism by
which phytochromes inhibit the function of PIL5 to initiate
those light responses.
Our work, and the previous analyses, show that phyt-
ochromes are able to regulate the degradation of various
light-signalling components through at least two different
pathways. One pathway acts through COP1, which leads
to degradation of largely positive components such as
HFR1, HY5 and LAF1 in the dark (Duek et al., 2004; Jang
et al., 2005; Osterlund et al., 2000; Seo et al., 2003; Yang
et al., 2005). The regulation of COP1 activity by phyto-
chromes is partly due to the exclusion of COP1 from the
nucleus under light conditions (von Arnim and Deng,
1994). However, HFR1 accumulates more quickly under
far-red light irradiation than COP1 exclusion proceeds,
indicating that this exclusion may not be the sole
regulatory mechanism (Yang et al., 2005). The other
pathway, acting through a yet-unknown mechanism,
degrades directly interacting, largely negative components
such as PIF3 and PIL5 through the 26S proteasome under
light conditions (Bauer et al., 2004; Park et al., 2004).
Further investigations will be required to determine how
phytochromes activate the degradation of their directly
interacting proteins in the light.
PIL5 regulates seed germination via GA 135
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

Experimental procedures
Plant materials and growth conditions
Arabidopsis thaliana plants were grown in a growth room with a 16-h light/8-h dark cycle at 22–24�C for general growth and seed har-vesting. The ga1 mutant was obtained from the Arabidopsis StockCenter (Salk_109115; Alonso et al., 2003). For generation of the PI-L5OX3 transgenic line, the full-length PIL5 cDNA was amplified withspecific primers (5¢-AGA GTG ATC AAA AAT GCA TCA TTT TGT CCCTGA C-3¢ and 5¢-AGA GTG ATC ACC ACC TGT TGT GTG GTT TCC-3¢),cloned into a pBI-HTM vector for expression of a fusion proteinbearing both His and Myc tags, and transformed into wild-typeplants (Col-0) by Agrobacterium-mediated transformation. Threeindependent homozygous lines (PIL5OX3, PIL5OX4 and PIL5OX5)were established. As they all showed phenotypes consistent withthe previously established PIL5OX1 and PIL5OX2 lines expressingthe native PIL5 gene, we used PIL5OX3 seeds for our experimentswhere indicated. All plants used in the experiments (pil5-1, phyA-211, phyB-9, ga1, PIL5OX1 and PIL5OX3) were Col-0 ecotype back-ground. Different mutants used in each figure were grown at thesame time, in the same growth room, in the same tray. Seeds werestored at 22�C in white paper bags.
Germination assay
For the PHYB-dependent germination assay, triplicate sets of 60seeds for each mutant were surface sterilized and plated on aqueousagar medium (0.6% phytoagar, pH 5.7). At 1 h after the start of seedsterilization, the plated seeds were irradiated with red(20 lmol m)2 sec)1) or far-red (3.2 lmol m)2 sec)1) light for 5 min.After 5 days’ incubation in the dark, germinated seeds were deter-mined by the emergence of radicles. For the PHYA-dependent ger-mination assay, the plated seeds were first irradiated with far-redlight (3.2 lmol m)2 sec)1) for 5 min, incubated in the dark for 48 h,then further irradiated with far-red light (3.2 lmol m)2 sec)1) forvarious times. Four days later, the germinated seeds were counted.To determine the effect of paclobutrazol on seed germination, trip-licate sets of 60 seeds for each mutant were spotted on agar platescontaining 100 lM paclobutrazol, incubated under PHYA- and PHYB-dependent germination assay conditions, and counted for germi-nated seeds. For the germination assay in the presence of exogen-ous GA, triplicate sets of 60 seeds for each mutant were spotted onagar plates containing various concentrations of GA4 (0–10 lM).
Gene-expression analysis
A total of 50 ll seeds were plated on wet filter paper and incubatedunder PHYA- or PHYB-dependent germination assay conditions,with the exception that the plates were incubated for 12 h beforeseeds were harvested for expression analysis. Total RNA wasextracted from the seeds using an Ambion RNA extraction kitaccording to the manufacturer’s guidelines (Ambion, Austin, TX,USA).
The following primers were used for RT–PCR analysis of geneexpression: GA3ox1 (5¢-CCG AAG GTT TCA CCA TCA CT-3¢ and 5¢-CCC CAA AGG AAT GCT ACA GA-3¢); GA3ox2 (5¢-TAG ATC GCA TCCCAT TCA CA-3¢ and 5¢- TGG ATA ACT GCT TGG GTT CC-3¢); GA2ox2(5¢-TGA CTC GGT TAG AGC AGG AG-3¢ and 5¢-CTT GAA CCT CCCGTT AGT CA-3¢); EXP1 (5¢-TCA CAT GTC AAT GGT TAC GC-3¢ and 5¢-TGT CCC CAG TTT CTT GAC AT-3¢); CP1 (5¢-TGA TGA GTC CAT CATCAA CG-3¢ and 5¢-TGT AGG ATT TGT CGC AGT CA-3¢); S18 (5¢-CCAGCG ATC GTT TAT TGC TT-3¢ and 5¢-AGT CTT TCC TCT GCG ACC
AG-3¢); PIL5 (5¢-GGG GAT TTT AAT AAC GGT-3¢ and 5¢-GAG ATTATG AAC TTC AGC AGC ACG-3¢); UBQ (5¢-GAT CTT TGC CGG AAAACA ATT GGA GGA TGG T-3¢ and 5¢-CGA CTT GTC ATT AGA AAGAAA GAG ATA ACA GG-3¢).
Analysis of endogenous GAs
Various mutant seeds were surface sterilized and plated onaqueous agar medium (0.6% phytoagar, pH 5.7). At 1 h after thestart of seed sterilization, the plated seeds were irradiated withred (20 lmol m)2 sec)1) or far-red (3.2 lmol m)2 sec)1) light for5 min. After 26 h incubation in the dark, seeds were collected forGA quantification. Quantitative analysis of GAs was carried out byGC-selected ion monitoring (SIM) using 2H-labelled GAs as inter-nal standards, as described previously (Gawronska et al., 1995).Briefly, a pre-purified ethyl acetate-soluble fraction containingGAs was subjected to HPLC purification using a reverse-phasecolumn (Capcell Pak C18 SG120; Shiseido Fine Chemicals, Tokyo,Japan). When necessary, GA-containing fractions after reverse-phase HPLC were further purified through another round of HPLCusing an ion-exchange column (Senshu Pak N[CH3]2, 1151-N;Senshu Scientific, Tokyo, Japan) to ensure the removal ofimpurities for reliable quantification. The purified fractions weresubjected to GC–SIM analysis using a mass spectrometer (Auto-mass Sun; JEOL, Tokyo, Japan) equipped with a GC (6890N;Agilent Technologies, Palo Alto, CA, USA) and a capillary column(DB-1; Agilent Technologies) after derivatization. Authentic GAsamples and 2H-labelled internal standards were purchasedfrom Professor Lewis Mander (Australian National University,Canberra).
Protein extraction and protein gel blotting
A total of 50 ll seeds were plated on wet filter paper, incubatedunder PHYA- or PHYB-dependent germination assay conditions,and harvested at the times indicated. The harvested seeds wereground in liquid nitrogen and solubilized in buffer (100 mM NaH2-
PO4, 10 mM Tris–Cl, 8 M urea pH 8.0) by vigorous vortexing. Thedebris was cleared by centrifugation at 20 000 g for 10 min at 4�C.Western blot analyses were performed as described by Park et al.(2004) to determine the PIL5 protein levels. The Myc tag was used toassess PIL5 protein levels with an anti-Myc antibody. To determinethe effects of MG132, five DAG dark-grown seedlings were pre-treated by either DMSO or MG132 (200 lM) and incubated undervarious light conditions until harvesting.
Acknowledgements
We thank Ms Masayo Sekimoto and Mr Atushi Hanada (RIKEN PlantScience Center) for GA analysis. This work was supported in part bygrants from the KRF (C00044), KOSEF (R21-2003-000-10002-0), thePlant Diversity Research Center of the 21st Frontier Research Pro-gram (PF0330508-00), and the Plant Metabolism Research Centerfunded by KOSEF.
Supplementary Material
The following supplementary material is available for this articleonline:Figure S1. The inhibition of PIL5 degradation by MG132 requires aprolonged pretreatment in seeds. The degradation of PIL5 without(A) or with (B) 24 hours pretreatment.
136 Eunkyoo Oh et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

Figure S2. Quantitive RT-PCR data corresponding to Figure 6.Figure S3. Quantitive RT-PCR data corresponding to Figure 7.Table S1 (A) Endogenous GA levels (ng/g dry seed) in germinatingseeds of Col-0, pil5, and PIL5OX1. Two sets of data were obtainedfrom independently incubated seed samples. (B) Ratio of GAcatabolic products and their substrates ([GA34 þ GA51]/[GA4 þ GA9])This material is available as part of the online article from http://www.blackwell-synergy.com
References
Alonso, J.M., Stepanova, A.N., Leisse, T.J. et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science,301, 653–657.
von Arnim, A.G. and Deng, X.W. (1993) Ring finger motif of Ara-bidopsis thaliana COP1 defines a new class of zinc-binding do-main. J. Biol. Chem. 268, 19626–19631.
von Arnim, A.G. and Deng, X.W. (1994) Light inactivation of Ara-bidopsis photomorphogenic repressor COP1 involves a cell-spe-cific regulation of its nucleocytoplasmic partitioning. Cell, 79,1035–1045.
Ballesteros, M.L., Bolle, C., Lois, L.M., Moore, J.M., Vielle-Calzada,
J.P., Grossniklaus, U. and Chua, N.H. (2001) LAF1, a MYB tran-scription activator for phytochrome A signaling. Genes Dev. 15,2613–2625.
Bauer, D., Viczian, A., Kircher, S. et al. (2004) Constitutive photo-morphogenesis 1 and multiple photoreceptors control degrada-tion of phytochrome interacting factor 3, a transcription factorrequired for light signaling in Arabidopsis. Plant Cell, 16, 1433–1445.
Borevitz, J.O., Maloof, J.N., Lutes, J. et al. (2002) Quantitative traitloci controlling light and hormone response in two accessions ofArabidopsis thaliana. Genetics, 160, 683–696.
Borthwick, H.A., Hendricks, S.B., Parker, M.W., Toole, E.H. and To-
ole, V.K. (1952) A reversible photoreaction controlling seed ger-mination. Proc. Natl Acad. Sci. USA 38, 662–666.
Cao, D., Hussain, A., Cheng, H. and Peng, J. (2005) Loss of functionof four DELLA genes leads to light- and gibberellin-independentseed germination in Arabidopsis. Planta, 223, 105–113.
Chattopadhyay, S., Ang, L.H., Puente, P., Deng, X.W. and Wei, N.
(1998) Arabidopsis bZIP protein HY5 directly interacts with light-responsive promoters in mediating light control of gene expres-sion. Plant Cell, 10, 673–683.
Chiwocha, S.D., Cutler, A.J., Abrams, S.R., Ambrose, S.J., Yang, J.,
Ross, A.R. and Kermode, A.R. (2005) The etr1–2 mutation inArabidopsis thaliana affects the abscisic acid, auxin, cytokininand gibberellin metabolic pathways during maintenance of seeddormancy, moist-chilling and germination. Plant J. 42, 35–48.
Chory, J., Chatterjee, M., Cook, R.K. et al. (1996) From seed ger-mination to flowering, light controls plant development via thepigment phytochrome. Proc. Natl Acad. Sci. USA 93, 12066–12071.
Debeaujon, I. and Koornneef, M. (2000) Gibberellin requirement forArabidopsis seed germination is determined both by testa char-acteristics and embryonic abscisic acid. Plant Physiol. 122, 415–424.
Duek, P.D., Elmer, M.V., van Oosten, V.R. and Fankhauser, C. (2004)The degradation of HFR1, a putative bHLH class transcriptionfactor involved in light signaling, is regulated by phosphorylationand requires COP1. Curr. Biol. 14, 2296–2301.
Fairchild, C.D., Schumaker, M.A. and Quail, P.H. (2000) HFR1 en-codes an atypical bHLH protein that acts in phytochrome A signaltransduction. Genes Dev. 14, 2377–2391.
Fankhauser, C. and Chory, J. (2000) RSF1, an Arabidopsislocus implicated in phytochrome A signaling. Plant Physiol. 124,39–45.
Fankhauser, C. and Staiger, D. (2002) Photoreceptors in Arabidopsisthaliana: light perception, signal transduction and entrainment ofthe endogenous clock. Planta, 216, 1–16.
Fujimori, T., Yamashino, T., Kato, T. and Mizuno, T. (2004) Circa-dian-controlled basic/helix-loop-helix factor, PIL6, implicated inlight-signal transduction in Arabidopsis thaliana. Plant CellPhysiol. 45, 1078–1086.
Gawronska, H., Yang, Y.Y., Furukawa, K., Kendrick, R.E., Takahashi,
N. and Kamiya, Y. (1995) Effects of low irradiance stress ongibberellin levels in pea seedlings. Plant Cell Physiol. 36, 1361–1367.
Hennig, L., Stoddart, W.M., Dieterle, M., Whitelam, G.C. and Scha-
fer, E. (2002) Phytochrome E controls light-induced germinationof Arabidopsis. Plant Physiol. 128, 194–200.
Hilhorst, H.W.M. and Karssen, C.M. (1988) Dual effect of light on thegibberellin- and nitrate-stimulated seed germination of Sisym-brium officinale and Arabidopsis thaliana. Plant Physiol. 86, 591–597.
Holm, M., Ma, L.G., Qu, L.J. and Deng, X.W. (2002) Two interactingbZIP proteins are direct targets of COP1-mediated control of light-dependent gene expression in Arabidopsis. Genes Dev. 16, 1247–1259.
Huq, E. and Quail, P.H. (2002) PIF4, a phytochrome-interacting bHLHfactor, functions as a negative regulator of phytochrome B sign-aling in Arabidopsis. EMBO J. 21, 2441–2450.
Huq, E., Al-Sady, B., Hudson, M., Kim, C., Apel, K. and Quail, P.H.
(2004) Phytochrome-interacting factor 1 is a critical bHLH regu-lator of chlorophyll biosynthesis. Science, 305, 1937–1941.
Jacobsen, S.E. andOlszewski, N.E. (1993) Mutations at the SPINDLYlocus of Arabidopsis alter gibberellin signal transduction. PlantCell, 5, 887–896.
Jacobsen, S.E., Binkowski, K.A. and Olszewski, N.E. (1996) SPIND-LY, a tetratricopeptide repeat protein involved in gibberellin sig-nal transduction in Arabidopsis. Proc. Natl Acad. Sci. USA 93,9292–9296.
Jang, I.C., Yang, J.Y., Seo, H.S. and Chua, N.H. (2005) HFR1 is tar-geted by COP1 E3 ligase for post-translational proteolysis duringphytochrome A signaling. Genes Dev. 19, 593–602.
Karssen, C., Brinkhorst-van der Swan, D.L.C., Breekland, A.E. and
Koornneef, M. (1983) Induction of dormancy during seed devel-opment by endogenous abscisic acid: studies on abscisic aciddeficient genotypes of Arabidopsis thaliana (L.) Heynh. Planta,157, 158–165.
Kepczynski, J.a.K. and Kepczynska, E. (1997) Ethylene in seed dor-mancy and germination. Physiol. Plant. 101, 720–726.
Kim, J., Yi, H., Choi, G., Shin, B., Song, P.S. and Choi, G. (2003)Functional characterization of phytochrome interacting factor 3 inphytochrome-mediated light signal transduction. Plant Cell, 15,2399–2407.
Koornneef, M. and van der Veen, J.H. (1980) Induction and analysisof gibberellin sensitive mutants in Arabidopsis thaliana (L.) Hey-nh. Theor. Appl. Genet. 58, 257–263.
Koornneef, M., Rolff, E. and Spruit, C. (1980) Genetic control of light-inhibited hypocotyl elongation in Arabidopsis thaliana (L.) Heynh.Z. Pflanzenphysiol. 100, 147–160.
Koornneef, M., Bentsink, L. and Hilhorst, H. (2002) Seed dormancyand germination. Curr. Opin. Plant Biol. 5, 33–36.
Lee, S., Cheng, H., King, K.E., Wang, W., He, Y., Hussain, A., Lo, J.,
Harberd,N.P.andPeng,J. (2002)Gibberellin regulatesArabidopsisseedgerminationviaRGL2,aGAI/RGA-likegenewhoseexpressionis up-regulated following imbibition. Genes Dev. 16, 646–658.
PIL5 regulates seed germination via GA 137
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

Leubner-Metzger, G. (2001) Brassinosteroids and gibberellins pro-mote tobacco seed germination by distinct pathways. Planta, 213,758–763.
Ma, L., Li, J., Qu, L., Hager, J., Chen, Z., Zhao, H. and Deng, X.W.
(2001) Light control of Arabidopsis development entails coordi-nated regulation of genome expression and cellular pathways.Plant Cell, 13, 2589–2607.
Marin, E., Nussaume, L., Quesada, A., Gonneau, M., Sotta, B.,
Hugueney, P., Frey, A. and Marion-Poll, A. (1996) Molecularidentification of zeaxanthin epoxidase of Nicotiana plumbagini-folia, a gene involved in abscisic acid biosynthesis and corres-ponding to the ABA locus of Arabidopsis thaliana. EMBO J. 15,2331–2342.
Monte, E., Tepperman, J.M., Al-Sady, B., Kaczorowski, K.A., Alon-
so, J.M., Ecker, J.R., Li, X., Zhang, Y. and Quail, P.H. (2004) Thephytochrome-interacting transcription factor, PIF3, acts early,selectively, and positively in light-induced chloroplast develop-ment. Proc. Natl Acad. Sci. USA 101, 16091–16098.
Neff, M.M., Fankhauser, C. and Chory, J. (2000) Light: an indicator oftime and place. Genes Dev. 14, 257–271.
Ni, M., Tepperman, J.M. and Quail, P.H. (1998) PIF3, a phytochrome-interacting factor necessary for normal photoinduced signaltransduction, is a novel basic helix–loop–helix protein. Cell, 95,657–667.
Ogawa, M., Hanada, A., Yamauchi, Y., Kuwahara, A., Kamiya, Y. and
Yamaguchi, S. (2003) Gibberellin biosynthesis and responseduring Arabidopsis seed germination. Plant Cell, 15, 1591–1604.
Oh, E., Kim, J., Park, E., Kim, J.I., Kang, C. and Choi, G. (2004) PIL5, aphytochrome-interacting basic helix–loop–helix protein, is a keynegative regulator of seed germination in Arabidopsis thaliana.Plant Cell, 16, 3045–3058.
Osterlund, M.T., Wei, N. and Deng, X.W. (2000) The roles of pho-toreceptor systems and the COP1-targeted destabilization of HY5in light control of Arabidopsis seedling development. PlantPhysiol. 124, 1520–1524.
Oyama, T., Shimura, Y. and Okada, K. (1997) The Arabidopsis HY5gene encodes a bZIP protein that regulates stimulus-induceddevelopment of root and hypocotyl. Genes Dev. 11, 2983–2995.
Park, D.H., Lim, P.O., Kim, J.S., Cho, D.S., Hong, S.H. and Nam, H.G.
(2003) The Arabidopsis COG1 gene encodes a Dof domain tran-scription factor and negatively regulates phytochrome signaling.Plant J. 34, 161–171.
Park, E., Kim, J., Lee, Y., Shin, J., Oh, E., Chung, W.I., Liu, J.R. and
Choi, G. (2004) Degradation of phytochrome interacting factor 3in phytochrome-mediated light signaling. Plant Cell Physiol. 45,968–975.
Penfield, S., Josse, E.M., Kannangara, R., Gilday, A.D., Halliday, K.J.
and Graham, I.A. (2005) Cold and light control seed germinationthrough the bHLH transcription factor SPATULA. Curr. Biol. 15,1998–2006.
Peng, J. and Harberd, N.P. (1997) Gibberellin deficiency and re-sponse mutations suppress the stem elongation phenotype ofphytochrome-deficient mutants of Arabidopsis. Plant Physiol.113, 1051–1058.
Quail, P.H. (1998) The phytochrome family: dissection of functionalroles and signalling pathways among family members. Phil.Trans. R. Soc. Lond. B, Biol. Sci. 353, 1399–1403.
Salter, M.G., Franklin, K.A. and Whitelam, G.C. (2003) Gating of therapid shade-avoidance response by the circadian clock in plants.Nature, 426, 680–683.
van der Schaar, W., Alonso-Blanco, C., Leon-Kloosterziel, K.M.,
Jansen, R.C., van Ooijen, J.W. and Koornneef, M. (1997) QTLanalysis of seed dormancy in Arabidopsis using recombinantinbred lines and MQM mapping. Heredity, 79, 190–200.
Seo, H.S., Yang, J.Y., Ishikawa, M., Bolle, C., Ballesteros, M.L. and
Chua, N.H. (2003) LAF1 ubiquitination by COP1 controls photo-morphogenesis and is stimulated by SPA1. Nature, 423, 995–999.
Shen, H., Moon, J. and Huq, E. (2005) PIF1 is regulated by light-mediated degradation through the ubiquitin-26S proteasomepathway to optimize photomorphogenesis of seedlings in Ara-bidopsis. Plant J. 44, 1023–1035.
Shinomura, T., Nagatani, A., Chory, J. and Furuya, M. (1994) Theinduction of seed germination in Arabidopsis thaliana is regula-ted principally by phytochrome B and secondarily by phyto-chrome A. Plant Physiol. 104, 363–371.
Shinomura, T., Nagatani, A., Hanzawa, H., Kubota, M., Watanabe,
M. and Furuya, M. (1996) Action spectra for phytochrome A- andB-specific photoinduction of seed germination in Arabidopsisthaliana. Proc. Natl Acad. Sci. USA 93, 8129–8133.
Soh, M.S., Kim, Y.M., Han, S.J. and Song, P.S. (2000) REP1, a basichelix–loop–helix protein, is required for a branch pathway ofphytochrome A signaling in Arabidopsis.PlantCell,12, 2061–2074.
Steber, C.M. and McCourt, P. (2001) A role for brassinosteroids ingermination in Arabidopsis. Plant Physiol. 125, 763–769.
Sullivan, J.A. and Deng, X.W. (2003) From seed to seed: the role ofphotoreceptors in Arabidopsis development. Dev. Biol. 260, 289–297.
Suzuki, G., Yanagawa, Y., Kwok, S.F., Matsui, M. and Deng, X.W.
(2002) Arabidopsis COP10 is a ubiquitin-conjugating enzymevariant that acts together with COP1 and the COP9 signalosome inrepressing photomorphogenesis. Genes Dev. 16, 554–559.
Tepperman, J.M., Zhu, T., Chang, H.S., Wang, X. and Quail, P.H.
(2001) Multiple transcription-factor genes are early targets ofphytochrome A signaling. Proc. Natl Acad. Sci. USA 98, 9437–9442.
Thomas, S.G., Phillips, A.L. and Hedden, P. (1999) Molecular cloningand functional expression of gibberellin 2 – oxidases, multifunc-tional enzymes involved in gibberellin deactivation. Proc. NatlAcad. Sci. USA 96, 4698–4703.
Tyler, L., Thomas, S.G., Hu, J., Dill, A., Alonso, J.M., Ecker, J.R. and
Sun, T.P. (2004) Della proteins and gibberellin-regulated seedgermination and floral development in Arabidopsis. Plant Phys-iol. 135, 1008–1019.
Ward, J.M., Cufr, C.A., Denzel, M.A. and Neff, M.M. (2005) The Doftranscription factor OBP3 modulates phytochrome and crypto-chrome signaling in Arabidopsis. Plant Cell, 17, 475–485.
Wei, N. and Deng, X.W. (1996) The role of the COP/DET/FUS genesin light control of Arabidopsis seedling development. PlantPhysiol. 112, 871–878.
Wolyn, D.J., Borevitz, J.O., Loudet, O., Schwartz, C., Maloof, J.,
Ecker, J.R., Berry, C.C. and Chory, J. (2004) Light-responsequantitative trait loci identified with composite interval and eX-treme array mapping in Arabidopsis thaliana. Genetics, 167, 907–917.
Yamaguchi, S., Smith, M.W., Brown, R.G., Kamiya, Y. and Sun, T.
(1998) Phytochrome regulation and differential expression ofgibberellin 3beta-hydroxylase genes in germinating Arabidopsisseeds. Plant Cell, 10, 2115–2126.
Yamashino, T., Matsushika, A., Fujimori, T., Sato, S., Kato, T., Ta-
bata, S. andMizuno, T. (2003) A link between circadian-controlledbHLH factors and the APRR1/TOC1 quintet in Arabidopsis thali-ana. Plant Cell Physiol. 44, 619–629.
Yamauchi, Y., Ogawa, M., Kuwahara, A., Hanada, A., Kamiya, Y. and
Yamaguchi, S. (2004) Activation of gibberellin biosynthesis andresponse pathways by low temperature during imbibition ofArabidopsis thaliana seeds. Plant Cell, 16, 367–378.
Yang, Y.Y., Nagatani, A., Zhao, Y.J., Kang, B.J., Kendrick, R.E. and
Kamiya, Y. (1995) Effects of gibberellins on seed germination of
138 Eunkyoo Oh et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139

phytochrome-deficient mutants of Arabidopsis thaliana. PlantCell Physiol. 36, 1205–1211.
Yang, J., Lin, R., Sullivan, J., Hoecker, U., Liu, B., Xu, L., Deng, X.W.
and Wang, H. (2005) Light regulates COP1-mediated degradationof HFR1, a transcription factor essential for light signaling inArabidopsis. Plant Cell, 17, 804–821.
Yanovsky, M.J., Casal, J.J. and Luppi, J.P. (1997) The VLF loci,polymorphic between ecotypes Landsberg erecta and Columbia,dissect two branches of phytochrome A signal transduction thatcorrespond to very-low-fluence and high-irradiance responses.Plant J. 12, 659–667.
PIL5 regulates seed germination via GA 139
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 47, 124–139