the role of membrane composition in atpase activities of cirrhotic rat liver: effect of silymarin
TRANSCRIPT

JOIJRNAL OF APPLIED TOXICOLO(iY, VOL. 10(4), 281-284 (1990)
The Role of Membrane Composition in ATPase Activities of Cirrhotic Rat Liver: Effect of Sily marin
Pablo Muriel and Marisabel Mourellet Departamento de Farmacologia y Toxicologia. Centro de Investigacion y de Estudios Avanzados del Instituto Politecnico Nacional, Apartado Postal 14-740. 07000 MCxico, D.F.
Key wordc cirrhiws, ATPase?. cell plasma membrane, liver, silyinarin. carbon tetrachloride
The activities of Ca2+ - and Na + , K'-ATPases were studied in liver plasma membranes from CCI,-cirrhotic rats and from livers of rats treated with silymarin in addition to CC14. CCIJ chronic treatment produced significant decreases in Na ', K+- and Ca2 +-ATPase activities; however, the animals treated with silymarin abng with CCI4 showed no differences in ATPase activities as compared to controls. The lipid analysis performed in plasma membranes revealed increases in the cholesterollphospholipid (CHIPL) and sphingomyelinlphosphatidylcholine (SM/PC) ratios in the cirrhotic group. Again, the membranes isolated from rats receiving CCIJ + silymarin showed normal CH/PL and SM/PC values. Considering that CHlPL and SMlPC ratios are related to membrane microviscosity, this study suggests that a lower fluidity of the membrane may be responsible for the observed decreases in ArPase activities in the cirrhotic group. Additionally, the role of silymarin to improve liver function in CC'I,-cirrhosis can be attributed partially to its action at membrane level by preventing the increases in CW/PL and SM/PC ratios.
INTRODUCTION - Biological membranes are complex structures that deal with ;I multiplicity of functions, including secretion, transport, endocytosir and signal transduction. '
The functions of membrane proteins, which are embedded in, are modulated by the composition and properties of the bilayer lipid environment. A key property of the lipid., in this regard, is the fluidity or general motional freedom of the lipid molecules. A growing body of evidence indicates that the fluidity of the lipid environment influences the activity of such physiologically important membrane enzymes as the Na' , K+- and Ca2'-dependent adenosine triphos- phatasesz4 and hormone-responsive adenylate cyclase.s-6 The fluidity also modulates transmembrane transport processes' and is a determinant of the passive permeability characteristic of a given bilayer.8.y
The major lipid components of biological membranes are phosphoglycerides, sphingolipids and cholesterol. io
Increases in the molar ratios of cholesteroVphospholipid (CHIPL) and sphingomyelin/phosphatidylcholine (SM/ PC) are inversely correlated to the fluidity of a variety of natural membranes.'
A relationship bet ween membrane alteriitions and disexes has been reported.'" Furthermore, i t is known that some drugs can affect organelle functions by modifying its membrane composition.14
On the other hand, it has been shown that silymarin, which consists of the compounds silybin, silydianin and silychristin (Fig. 1) occurring in the thistle Silibum murianurn (L) Gaertn., was effective in preventing
t Author to whom correspondence should be addressed.
poisoning by several hepatotoxic substances, including CCl,.'5-17 The protective action of this flavonoid can be explained in terms of its capability of trapping free radicals and its 'stabilizing' effect on the cytoplasmic membrane.
Therefore, the aim of the present work was to study the capacity of silymarin to preserve plasma membrane ATPase activities when given along with CCI, to produce cirrhosis, as well as to study the relationship between plasma membrane lipids and ATPase activity.
EXPERIMENTAL
Animal treatments Male Wistar 30-day-old rats weighing 50-100 g initially and fed ad libitum a Purina Chow diet were used in these experiments. Liver cirrhosis was produced by intraperitoneal administration of 0.15 mi of CC1, in mineral oil, three times a week for 8 weeks. The ratio of CCI, was increased weekly as described by Ehrinpreis et al.
Four groups of ten animals each were used. The first group received CCI, treatment and the second (controls) received mineral oil. The third group con- sisted of rats that, in addition to CCI,, received a daily dose of 50 mg kg-' body wt. of silymarin for 8 weeks. Silymarin was suspended in 0.5% Carboxymethyl cellulose (CMC) and delivered into the stomach through an intragastric tube. The fourth group of animals received silymarin only, as described for animals in the third group. All the animals were sacrificed 72 h after the end of the treatments.
0260-4 37>(/90/04O28 1 +$05.00 0 1 W I by John Wiley & Sons, Ltd. Received / I July I984
Accepred 1 March 1990

282 P. MURIEL AND M. MOURELLE
s i l yb in
o n 0 1 n3’
HO
CH3O
5“
o n 0
HO
no 5“
si I ydi anin on
sit ychri stin
Figure 1. Structures of the three active compounds present in silymarin: silybin (two isomers), silydianin and silychristin.
Membrane preparation Plasma membranes from rat livers were isolated accord- ing to Neville’s method]” with a modification described by Pohl et d.*” The membranes were frozen in liquid nitrogen and for further use were resuspended in 20 mM imidazole (pH 7.8). Protein determinations were performed according to the method described by Bradford.21
Na+, K+-ATPase assay The standard medium for total ATPase activity con- tained, in 0.5 ml, 150 mM NaCI, 5 mM KCI, 2.5 mM MgC12, 2.5 mM ATP, 60-80 pg of membrane protein and 20 mM imidazole buffer. The pH of the solution was adjusted to 7.8 at 37”C, which was the temperature of all enzymatic incubations. Mg2+-activated ATPase was measured in an identical medium that included 1 mM ouabain. The difference between the two reac- tions represented the Na+, K+-ATPase. Incubations were carried out at 37°C for 30 min. To stop the reaction, 100 p1 of ice-cold 35% (w/v) trichloroacetic acid were added. Aliquots of 20 pl were analysed for inorganic phosphate according to Ames.’* Spontaneous hydrolysis of ATP was monitored by eliminating protein from control assays.
CaZ+-ATPase assay A 0.5-ml aliquot of standard medium contained 1.5 mM CaCI2, 2.5 mM ATP, 60-80 pg of membrane protein and 20 mM imidazol buffer (pH 7.8). Incubations were carried out at 37°C for 30 min. The reaction was stopped as described above, and aliquots were used to determine inorganic phosphate by the method of Ames.22
Extraction of lipids The membrane lipids were extracted according to Folch et The pure lipid solutions were evaporated under a stream of nitrogen. The dried samples were dissolved in a known volume of benzene. The total lipid
phosphorus content of phospholipids and cholesterol was determined in each extract using the methods described by Ames and Abell.
Analysis of phospholipids Phospholipids species were analysed by thin-layer chromatography. Thin-layer chromatoplates (20 x 20 cm) were covered with silica gel-ammonium sul- phate. The developing solvent mixture used for phos- pholipids was cloroform-methanol-acetic acid-water 170:40:18:10 vh). Visualization of spots on all thin- f ayer chromatograms was performed by heating the
plates in an oven at 180°C for 30 min.” The silica of each spot was scraped off and the lipid
phosphorus content was determined according to the method described by Ames.” Identification of each phospholipid species was confirmed by comparison of the R F values of the pure standards chromatographed in parallel.
Chemicals CCIJ, spectroscopic grade was purchased from J.T. Baker (Mexico). Silymarin was kindly donated by Dr Madaus GmbH & Co, FRG. Silica gel G-60 was purchased from Merck & Co, FRG. Ouabain, ATP, imidazole, NaCl, KCl and CaCI? were obtained from Sigma Chemical Co (St. Louis, MO, USA). All other reagents were analytical grade, of the best quality available.
Statistics For statistical analysis, Student’s t-tests were performed and a difference was considered significant when Pc0.05.
~~
RESULTS
Previous experiments have proved that the scheme of chronic CCI4 administration consistently produces
SILYMARIN EFFECT ON LIVER MEMBRANES IN CIRKIIOSIS 283
biochcrnical and histological patterns that characterize liver cirrhosis. I x
At'tcr chronic CCI, administration, Na ' . K ' -ATPase decre;iwd by YO'% 21s compared to control values (P<O.O3) (Fig. 2, upper panel) and the Ca"-dcpen- dcnt ATPase activity showed a 70% decrease (I'<O 075) (Fig. 2, lower panel). Plasma mcmbrancs of :iniin:ils receiving silymarin in addition to CCI, showctl normal Na ' , K - and Ca?' -ATPase ;ictivities when malysed a t 8 weeks of chronic treatment (Fig. 2). In the animals recciving vehicles or silymarin only. both / \ V a s e activities wcre normal.
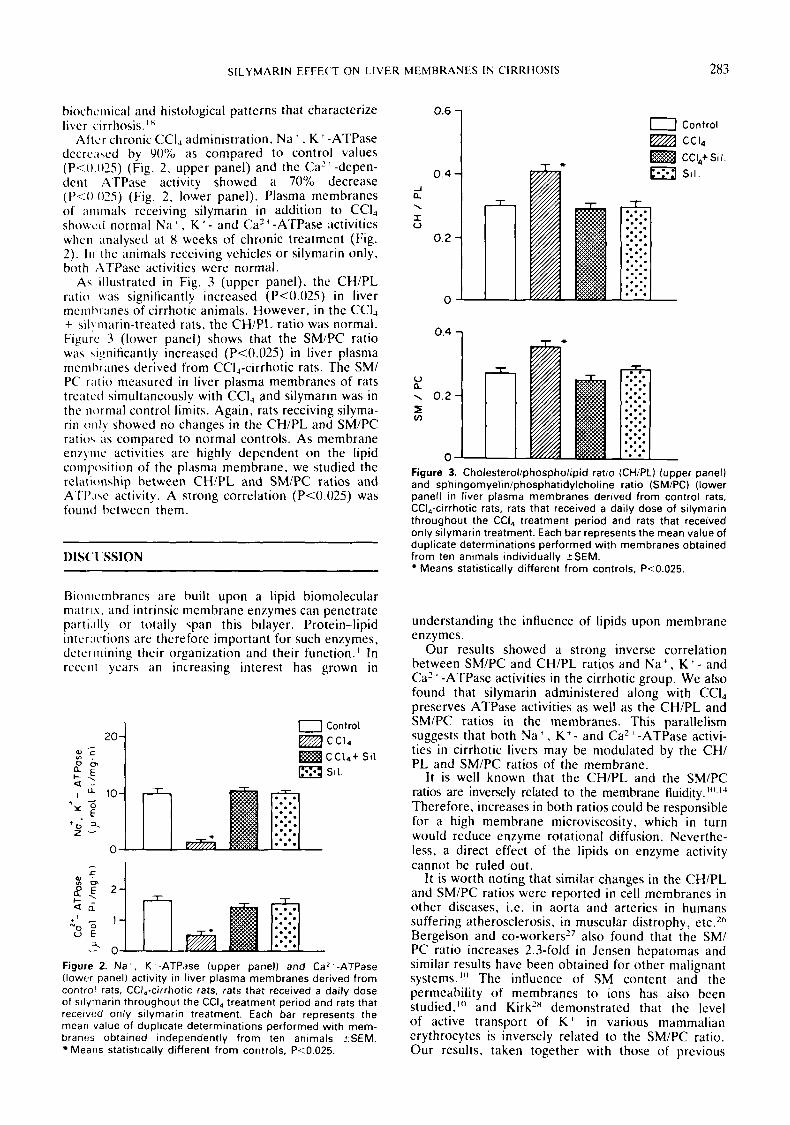
As illustrated in Fig. 3 (upper panel). the CH/PL ratio was significantly increased (P<0.025) in liver mcml~ranes of cirrhotic animals. However, in the CCI, + ~il!m:irin-treated rats, the CH/PL ratio was normal. Figurc 3 (lower panel) shows that the SM/PC ratio was b.ignificantly increased (P<0.025) in liver plasma rncnil3r:ines derived from CCI,-cirrhotic rats. The SM/ PC r i i t i o measurcd in liver plasma membrancs of rats trcatetl simultaneously with CCI, and silymarin was in the normal control limits. Again, rats receiving silyma- rin o n l y showed no changes in the CH/PL and SM/PC ratio\ ;is cornpared to normal controls. As membrane enzytiic activities arc highly dependent on the lipid comp)sition of the plasma membrane, we studied the rclnt ionship between CH/PL and SMlPC ratios and ATI'<iw activity. A strong correlation (P<0.025) was founci bctwcen them.
Bionic*inbrancs are built upon a lipid biomolecular matrix. and intrinsic mcmbrane enzymes can penctrate parti'illy or totally span this bilayer. Protein-lipid intcr:ictions are thcreforc important for such enzymes, detcimining thcir organization and their t'unction.' In receiil ycars a n increasing interest has grown in
0 Control 20 c c14 .-
c Cis+ SI1 O l Y I n .
2 : SII
Y E 0
+d 2
I- .. u - t -
I to
z 0
Figure 2. Na ' , K ' -ATPase (upper panel) and Ca2 ' -ATPase (lower panel) activity in liver plasma membranes derived from control rats, CCI,-cirrhotic rats, rats that received a daily dose of silymarin throughout the CCI, treatment period and rats that received only silymarin treatment. Each bar represents the meari value of duplicate determinations performed with mem- branes obtained independently from ten animals ! SEM.
Means statistically different from controls, P.cO.025.
0.6
0.4 -1 (L
\
I V
0.2
0
V (L
\ 0.2
5
0 Figure 3. Cholesterol/phospholipid ratio (CHIPL) (upper panel) and sphingomyelinlphosphatidylcholine ratio (SMIPC) (lower panel) in liver plasma membranes derived from control rats, CCI,-cirrhotic rats, rats that received a daily dose of silymarin throughout the CCI, treatment period and rats that received only silymarin treatment. Each bar represents the mean value of duplicate determinations performed with membranes obtained from ten animals individually ZSEM.
Means statistically different from controls, P<0.025.
understanding the influence of lipids upon membrane enzymes.
Our results showed a strong inverse correlation between SM/PC and CH/PL ratios and Na + , K ' - and Ca' . -ATPase activities in the cirrhotic group. We also found that silymarin administered along with CCI, preserves ATPase activities as well as the CH/PL and SM/I'C ratios in the membranes. This parallelism suggests that both Na ' , K'- and Ca' ' -ATPase activi- ties in cirrhotic livers may he modulated by the CHI PL and SM/PC ratios of the membrane.
I t is well known that the CH/PL and the SM/PC ratios are inversely related to the membrane fluidity. I".'-'
Therefore, increases in both ratios could be responsible for a high membrane microviscosity, which in turn would reduce enzyme rotational diffusion. Neverthe- less, a direct effect of the lipids o n enzyme activity cannot be ruled out.
I t is worth noting that similar changes in the CH/PL and SM/PC ratios were reported in cell membranes in other discases, i.c. in aorta and arteries in humans suffering atherosclerosis, in muscular distrophy, etc.2" Bergelson and co-workers" also found that the SM/ PC ratio increases 2.3-fold in Jensen hepatomas and similar results have been obtained for other malignant
The influence of SM contcnt and the permeability of membranes to ions has also bcen studied,"' and Kirk'x demonstrated that the level of active transport of K ' in various mammalian crythrocytes is inversely related to the SM/PC ratio. Our results, taken together with those of previous

284 P. MURIEL AND M. MOURELLE
observations, lead us to suggest that several pathologi- cal processes are related to membrane alterations. Experimental alterations of lipid fluidity are reported to have consistent effects upon the activity of the hepatocyte plasma membrane adenylate cyclase and Na', K'-ATPase, and modification of the membrane environment can be achieved by pharmacological agents. '
The Ca'+-ATPase activity that we measure has been qualified as the plasma membrane calcium pump responsible for the final regulation of the total calcium content of the hepa t~cyte . '~ Penniston19 pointed out that mitochondria and endoplasmic reticulum may be responsible for determining the level of intracellular calcium in the short term, but these organelles cannot continue to accumulate calcium within themselves
indefinitely; then, in the long term, the plasma mem- brane Ca'+-ATPase must be responsible for maintain- ing the ultimate resting level of calcium within the cell.
Beneficial effects of silymarin have been reported before in several models of liver injury;"-" our results now suggest that some of its pharmacological actions on the liver could be due to its capacity to preserve the SM/PC and CH/PL ratios and consequently ATPase activities in the normal limits.
Acknowledgements
The authors are indebted to Ms Marisela Vidal and Ms Concepci6n Avalos for secretarial assistance, and Mr Alfredo Padilla for preparing the figures.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
J. Stroch and A. M. Kleinfeld, The lipid structure of biological membranes. Trends Biochem. Sci. 11, 418-421 (1985). C. M. Grisham and R. E. Barnett, The interrelationship of membrane and protein structure in the functioning of the (Na' + K+)-activated ATPase. Biochim. Biophys. Acta 266,
H. K. Kimelberg and D. Papahadjopoulos, Effects of phos- pholipid acyl chain fluidity, phase transition and cholesterol on (Na' and K+)-stimulated ATPase. J. Biol. Chern. 249,
E. B. Keeffe, B. F. Scharschmidt, N. M. Blankenship, eta/., Studies of relationship among bile flow, liver plasma membrane Na', K+-ATPase, and membrane microviscosity in the rat. J. CIin. Invest. 64, 1590-1598 (1979). M. D. Houslay, T. R. Hesketh, G. A. Smith, et a/., The lipid environment of the glucagon receptor regulates adenylate cyclase activity. Biochirn. Biophys. Acta 436,495-504 ( 1 976). I. Dipple and M. D. Houslay, The activity of glucagon- stimulated adenylate cyclase from rat liver plasma mem- branes is modulated by the fluidity of its lipid environment. Biochem. J. 174, 179-190 (1978). 6. Deuticke, Properties and structural basis of simple diffusion pathways in the erythrocyte membrane. Rev. Physiol. Biochem. Pharmacol. 78, 1-97 (1977). J. De Gier, J. G. Mandersloot and L. L. M. Van Deenen, Lipid composition and permeability of liposomes. Biochim. Biophys. Acta 150, 666-675 (1968). D. Papahadjopoulos, M. Cowden, and H. Kimelberg, Role of cholesterol in membrane permeability and enzymatic activity. Biochirn. Biophys. Acta 330, 8-26 (1973). Y. Garenholz and T. E. Thompson, Sphingomyelins in bilayers and biological membranes. Biochirn. Biophys. Acta 604, 129-158 (1980). W. J. Van Blitterswijk, R. P. Van Hoeven and B. W. Van der Meer, Lipid structural order parameters (reciprocal of fluidity) in biomembranes derived from steady-state fluor- escence polarization measurements. Biochim. Biophys. Acta 644, 323-332 (1981). M. Shinitzky and M. Inbar, Differences in microviscosity induced by different cholesterol levels in surface membrane lipid layer of normal lymphocytes and malignant lymphoma cells. J. Mof. Biol. 85, 603-615 (1974). M. Shinitzky and Y. Barenholz, Fluidity parameters of lipid regions determined by fluorescence polarization Biochim. Biophys. Acta 515, 367-394 (1978). 0. Schachter, Fluidity and function of hepatocyte plasma membranes. Hepatology 4, 140-151 (1984). G. A. Desplaces, J. Choppin, G. Vogel and W. Trost, The
613-624 (1972).
1071-1 080 ( 1 974).
effects of Silymarin on experimental phalloidine-poisoning. Arzneim-Forsch. 25, 89-96 (1975).
16. G. Vogel, W. Trost, R. Braatz and M. Ulrich, Untersuchungen zu pharmakodynamik, angriffspunkt und wirkungsmechan- ismus von silymarin dem antihepatotoxischen prinzip aus Silybum mar. (L) GAERTN. Arzneim.-Forsch. 25, 82-89 (1975).
17. A. Valenzuela, C. Lagos, K. Schmidt and L. A. Videla, Silymarin protection against hepatic lipid peroxidation induced by acute ethanol intoxication in the rat. Biochim. Pharmacol. 34, 2209-2212 ( 1 985).
18. M. N. Ehrinpreis, M. A. Giambrone and M. Rojkind, Liver proline oxidase activity and collagen synthesis in rats with cirrhosis induced by carbon tetrachloride. Biochim. Biophys. Acta 629, 184-193 (1980).
19. M. D. Neville, The isolation of a cell membrane fraction from rat liver. J. Biophys. Biochem. CytoI. 8,415-422 (1960).
20. S. L. Pohl, L. Birnbaumer and M. Rodbell, The glucagon sensitive adenyl cyclase system in plasma membranes of rat liver. J. Biol. Chem. 245, 1849-1856 (1971).
21. M. M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochern. 72, 24&254 (1976).
22. B. N. Ames, Assay of inorganic phosphate, total phosphate and phosphatases. Methods Enzymol. 8, 115-118 (1966).
23. J. Folch, M. Lees and G. H. Sloane-Stanley, A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226, 497-509 (1957).
24. L. L. Abell, 6. 6. Levy, B. 6. Brodie, et a/., A simplified method for the estimation of total cholesterol in serum and demonstration of its specificity. J. Biol. Chem. 195, 357-366 (1952).
25. P. V. Skipski and M. Barclay, Thin-layer chromatography of lipids. Methods Enzymol. 14, 530-598 (1969).
26. K. Owens and 6. P. Hughes, Lipids of dystrophic and normal mouse muscle: whole tissue and particulate fractions. J. Lipid Res. 11, 486-495 (1970).
27. L. D. Bergelson, E. V. Dyateovitskaya, 1. B. Sorokina and N. P. Gorkova, Phospholipid composition of mitochondria and microsomes regenerating rat liver and hepatomas of different growth rats. Biochim. Biophys. Acta 360, 361-365 (19741.
28. G. Kirk, Potassium transport and lipid composition in mammalian red blood cell membranes. Biochim. Biophys. Acta 464, 157-164 (1977).
29. J. T. Penniston, Plasma membrane Caz+-pumping ATPases. Ann. N. Y. Acad. Sci. (U.S.A.) 296-303 (1982).