the role of the transcriptional activator protein dbp in ... role of the transcriptional activator...
TRANSCRIPT

Journal of Cell Science, Supplem ent 16, 123-127 (1992)Printed in G reat Britain © The Company o f Biologists Limited 1992
123
The role of the transcriptional activator protein DBP in circadian liver gene
expression
JÉRÔME WUARIN, EILEEN FALVEY, DAN LAVERY, DALE TALBOT, ED SCHMIDT, VINCENT OSSIPOW,
PHILIPPE FONJALLAZ and UELI SCHIBLER*
Department o f M olecular Biology, Sciences II, 30 Quai Ernest Ansermet, CH-1211 Geneva-4, Switzerland
^Corresponding author
Summary
DBP, a liver-enriched transcriptional activator protein of the leucine zipper protein family, accumulates according to a very strong circadian rhythm (amplitude approx. 1000-fold). In rat parenchymal hepatocytes, the protein is barely detectable during the morning hours. At about 2 p.m., DBP levels begin to rise, reach maximal levels at 8 p.m. and decline sharply during the night. This rhythm is free-running: it persists with regard to both its amplitude and phase in the absence of external time cues, such as daily dark/light switches. Also, fasting of rats for several days influences neither the amplitude nor the phase of circadian DBP expression. Since the levels of DBP mRNA and nascent transcripts also oscillate with a strong amplitude, circadian DBP expression is transcriptionally controlled. While DBP mRNA fluctuates with a similar phase and amplitude in most tissues examined, DBP protein accumulates to high concentrations only in liver nuclei. Hence, at least in
nonhepatic tissues, cyclic DBP transcription is unlikely to be controlled by a positive and/or negative feedback mechanism involving DBP itself. More likely, the circadian DBP expression is governed by hormones whose peripheral concentrations also oscillate during the day. Several lines of evidence suggest a pivotal role of glucocorticoid hormones in establishing the DBP cycle.
Two genes whose mRNAs and protein products accumulate according to a strong circadian rhythm with a phase compatible with regulation by DBP encode enzymes with key functions in cholesterol metabolism: HMG-coA reductase is the rate-limiting enzyme in cholesterol synthesis; cholesterol 7-a hydroxylase performs the rate-limiting step in the conversion of cholesterol to bile acid. DBP may thus be involved in regulating cholesterol homeostasis.
Key words: liver transcription factor, DBP, circadian rhythms.
Introduction
Our laboratory has chosen the liver as a paradigm for studying cell-type-specific gene expression. By identifying cis- acting albumin promoter elements and the transcription factors interacting with them, we hoped to gain access to key regulators of hepatic gene expression. A detailed functional dissection of the albumin promoter revealed six cognate sites (A, B, C, D, E and F) for sequence-specific DNA- binding proteins (Lichtsteiner et al., 1987). Two of these, sites B and D, proved to be particularly important for efficient cell-type-specific transcription (Maire et al., 1989; Lichtsteiner and Schibler, 1989). The proteins binding to these elements have been identified, purified and their cDNA and genes isolated by molecular cloning techniques (for review, see Crabtree et al., 1992). In this paper we concentrate on our results with a protein, DBP, which binds to the albumin D promoter element (Mueller et al., 1990). Studies on DBP expression have led to the surprising discovery that in adult rats, DBP accumulates according to a
very strong circadian rhythm (Wuarin and Schibler, 1990). In the next paragraphs we will focus on the regulation of circadian DBP expression and speculate on its purpose.
DBP, a liver-enriched transcriptional activator protein
DBP cDNAs have been obtained by screening a lambda gt 11 liver cDNA expression library with a radiolabeled DNA probe encompassing the albumin promoter element D. Sequencing of close to full-length cDNAs revealed an open reading frame corresponding to 325 amino acids (Mueller et al., 1990; see corrected sequence, Cell 65, 1991, page 915). DBP dimerizes via a carboxy-terminal leucine zipper and binds DNA via an adjacent basic region (Vinson et al.,1989). The three-dimensional structure of at least one leucine zipper, that of the yeast transcription factor GCN4, has been resolved by X-ray crystallography (O’Shea et al., 1991). Moreover, the amino acid residues located at func

124 ./ . Wuarin and others
Fig. 1. Schematic representation (see O ’Shea et al., 1992) of dimerized coiled coils (leucine zippers) within a DBP homodimer. The N- to C-polarity of the dimerization helices is from left to right. The right hand panel shows a vertical projection of the coiled coils. The D and A heptad positions
containing the hydrophobic amino acids are shown in black and shaded, respectively. Hypothetical salt bridges between glutamic acids (E) and arginines (R) are indicated by arrows (four on each side of the paired amphipathic helices).
tionally strategic positions of leucine zippers have been determined by elegant genetic (Hu et al., 1990) and biochemical work (eg. Landschulz et al., 1989, and references therein; O ’Shea et al., 1992). Leucine zipper regions form coiled-coil structures, consisting o f parallel am phipathic a - helices that interact via hydrophobic side chains o f amino acids located at positions A and D (Fig. IB). W hile no particular D position must be occupied by a leucine, a total of at least two leucines appear to be required within this heptad repeat (Hu et al., 1990). However, not all leucine zippers can form stable interactions with each other. To a large extent, the specificity o f dim erization is dictated by salt bonds between amino acids of opposite charges located in positions G and E (see O ’Shea et al., 1992 and references therein). The leucine zipper o f DBP (Fig. 1A) is somewhat unusual in two respects. Firstly, it contains only two leucines, the absolute minimum tolerated for dimerization (see above). Secondly, the G and E positions consist o f four charged amino acids which, theoretically, could establish as many as eight salt bridges. However, the intrahelical repulsion created by the runs of three consecutive arginines and glutamic acids in the E and G heptad repeat positions, respectively, may also result in a destabilization o f the dimerization helices. Indeed, our cross-linking experiments with purified recom binant proteins produced in E. coli suggest a three-fold higher dimerization dissociation constant for the DBP zipper as com pared to the zipper o f LAP, a C/EBP (Landschulz et al., 1989) related protein that also has affinity for the albumin D site (Descom bes and Schi- bler, 1991). In keeping with this observation, replacement o f the DBP zipper with the LAP zipper increases the affinity of the DBP chimeric protein to its DNA cognate site by about three-fold. Perhaps the dimerization properties of DBP have to be evaluated in the light o f its role in diurnal transcription activation (see below). The circadian accumulation (see below) and action of this protein may call for a high off-rate o f DNA binding to ensure a rapid equilibrium between specific and non-specific occupancy.
The N-terminal moiety o f DBP contains the transcription activation domain(s). In cotransfection experiments with wild-type DBP and DBP target genes, over-expression of the N-terminal two thirds o f DBP results in a strong attenuation o f DBP-m ediated transcription activation. This squelching is specific, since transcription activation through LAP is not impaired by an excess o f N-terminal DBP sequences (E. F., unpublished results). Thus, the transcription activation domains of DBP and LAP appear to inter
act with different targets o f the general transcription apparatus.
In liver, the majority of DBP is highly phosphorylated (V. O., F. Fleury and U. S., unpublished data). Dephosphorylation o f DBP with acidic potato phosphatase does not appear to influence the equilibrium DNA-binding constant o f DBP. W e cannot exclude, however, that the dynamics o f DNA binding (Kon and A"0ff) are m odulated by phosphorylation, as has recently been suggested for the serum response factor SRF (M arais et al., 1992). Alternatively, phosphate groups may be involved in regulating the transcription activation potential o f DBP.
The DBP gene
The rat gene specifying DBP has recently been isolated and characterized in our laboratory. It contains four exons and three introns and encom passes about 5.6 kb from c a p - to polyadenylation sites (J. W. and E. F., unpublished data). The prom oter is located within a CpG island and is devoid o f a canonical TATA box. As has been observed for many prom oters o f this type, transcription initiation occurs at multiple start sites spread over about 50 nucleotides. The 5 ' untranslated region of DBP m RNA is extraordinarily long (about 400 nucleotides) and contains three upstream AUGs. W e remain uncertain whether these sequences are relevant for controlling translation initiation in the animal, but deletion o f the sequences containing the three upstream AUGs increases the accum ulation of DBP both in vivo (transient transfection) and in vitro (reticulocyte lysate) (S. Rufino and U. S., unpublished data).
Circadian and developmental DBP expression
DBP accum ulates in parenchym al hepatocytes according to a very robust circadian rhythm. The DBP level measured at the time of maximal accum ulation (8 p.m. in Lewis rats) exceeds the one determined at the time of minimal accumulation (8 a.m.) by about three orders o f magnitude. This rhythm is transcriptionally controlled, free-running and independent o f food or water uptake (W uarin and Schibler,1990). W hile DBP mRNA cycles with the same phase and a similarly strong am plitude in a variety o f non-hepatic tissues (lung, spleen, kidney), the DBP protein accum ulates to much lower levels in these cell types as com pared to
Circadian liver gene expression 125
hepatocytes. As DBP mRNA appears to be associated with polysom es in all tissues (E. S., unpublished results), it is conceivable that protein stability rather than translation efficiency accounts for the differential DBP accumulation in various cell types.
In rodents, circadian rhythmicity com mences shortly after weaning (for reviews on circadian rhythms, see Edmunds, 1988). As a consequence, DBP oscillation is only observed in rats older than three weeks (M ueller et al., 1990; Yano et al., 1992). In the liver of younger animals (birth to three weeks), DBP accumulates to very low levels and does not cycle during the day (D. L. and E. S., unpublished data).
How is the oscillation o f DBP transcription regulated? As circadian mRNA expression is sim ilar in tissues that do or do not accum ulate the protein, DBP itself is unlikely to control its own transcription by positive and/or negative feedback loops. M ore likely, DBP transcription is governed by hormones whose secretion is also rhythmic. Several lines o f evidence are com patible with glucocorticoid hormones participating in the “downphase” of circadian DBP expression. In rats, corticosterone shows highest peripheral levels at the dark/light switch (6 p.m. in our animal facility; for circadian glucocorticoid secretion, see Meier, 1975 and references therein), several hours after DBP transcription rates have clim axed (between 2 p.m. to 4 p.m.). Injection of dexamethasone around noon results in a strong attenuation of DBP mRNA accum ulation during the afternoon. M oreover, in homozygous fa/fa Zucker rats, which exhibit higher corticosterone levels than their w ild-type counterparts (Guillaume-Gentil et al., 1990), the DBP cycle is phase-shifted towards earlier hours and is reduced in am plitude. For the two following reasons, we consider it likely that DBP transcription is directly repressed by glucocorticoid hormones. (1) In hepatoma cells, dexamethasone addition leads to a rapid extinction of DBP mRNA accumulation (E. F., unpublished data). (2) The DBP prom oter contains a GRE, as judged from DNase 1 footprinting studies with recom binant glucocorticoid receptor.
In an alternative attem pt to localize cis-acting DBP promoter elements, DNase 1 hypersensitive sites in rat liver nuclei were mapped (D. T., unpublished data). Such sites have been observed exclusively within the 1 kb 5 ' flanking region of the DBP gene. Some of these appear to be more efficiently cleaved in nuclei isolated during the afternoon, when the DBP gene is most active, than in nuclei isolated in the morning, when the DBP gene is not or very weakly transcribed. The importance o f these elements for circadian DBP expression is currently being tested in transgenic mice.
Purpose of circadian DBP expression
In order to decipher the physiological significance of circadian DBP expression, one needs to identify bona fide DBP target genes. As m entioned earlier, DBP has been recognized as a transcription factor w hich binds to an albumin prom oter element in vitro. Indeed, in adult rats, albumin transcription appears to fluctuate with a phase com patible with that o f DBP accum ulation (W uarin and Schibler, 1990; J. W., unpublished results). However, this circadian tran-
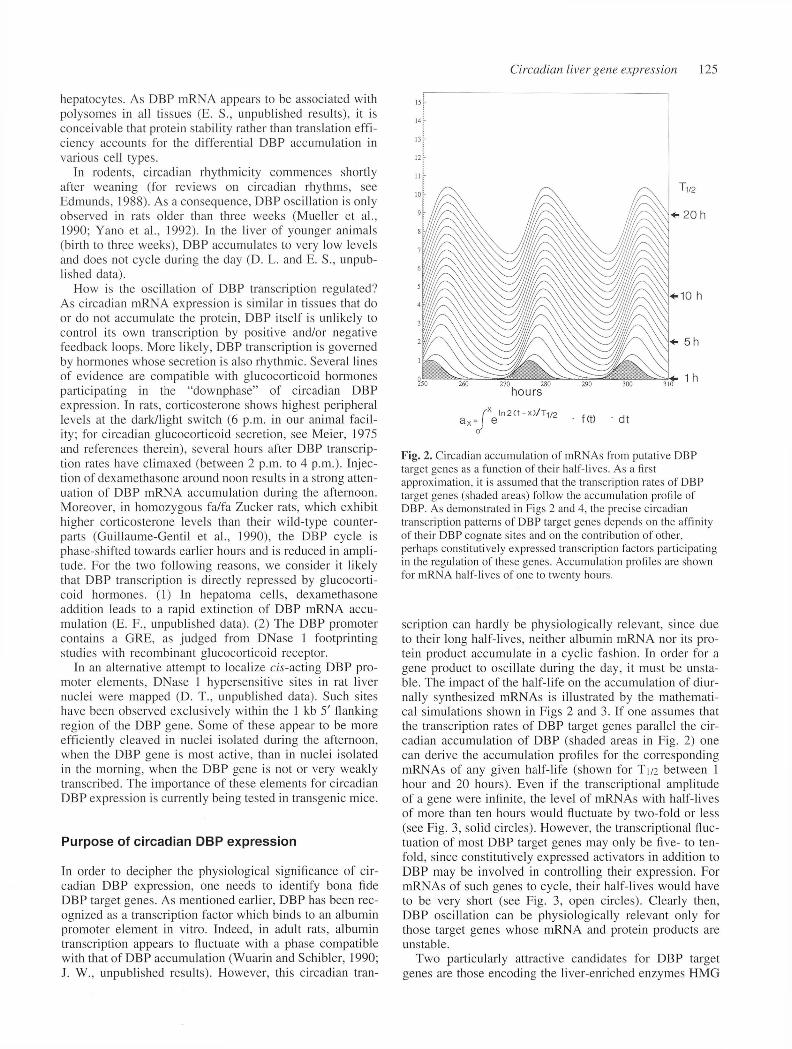
Fig. 2. Circadian accumulation of mRNAs from putative DBP target genes as a function of their half-lives. As a first approximation, it is assumed that the transcription rates of DBP target genes (shaded areas) follow the accumulation profile of DBP. As demonstrated in Figs 2 and 4, the precise circadian transcription patterns o f DBP target genes depends on the affinity o f their DBP cognate sites and on the contribution of other, perhaps constitutively expressed transcription factors participating in the regulation of these genes. Accumulation profiles are shown for mRNA half-lives of one to twenty hours.
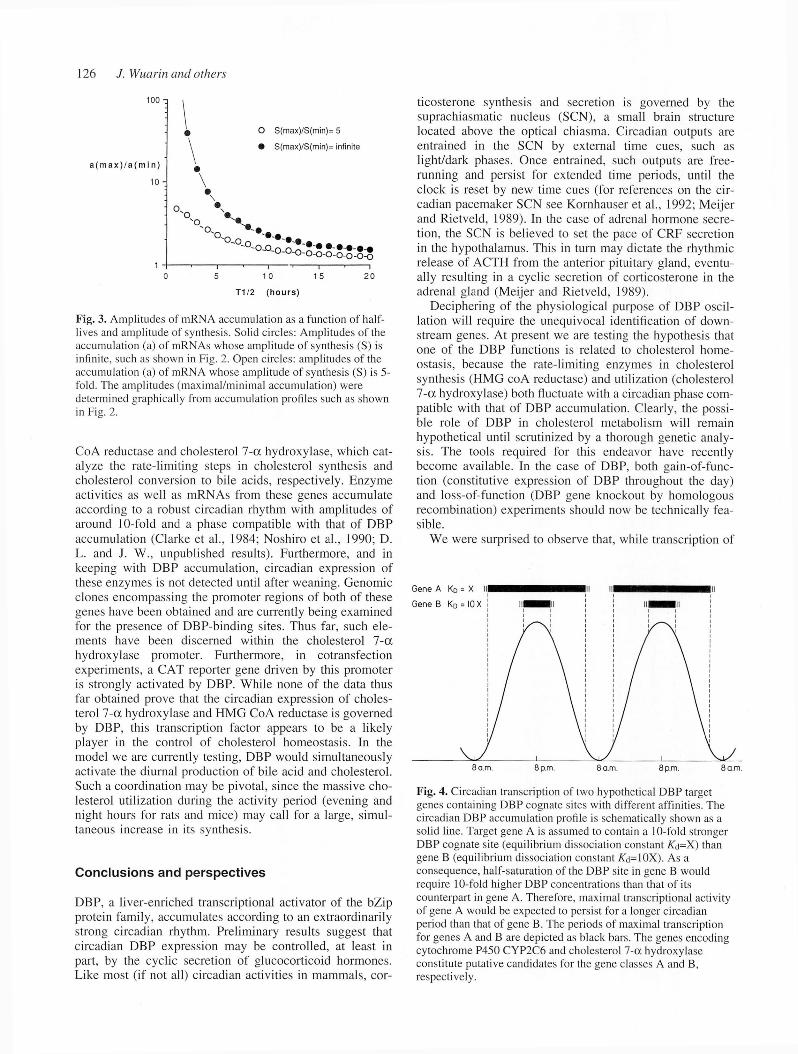
scription can hardly be physiologically relevant, since due to their long half-lives, neither albumin m RNA nor its protein product accum ulate in a cyclic fashion. In order for a gene product to oscillate during the day, it must be unstable. The impact o f the half-life on the accum ulation of diurnally synthesized mRNAs is illustrated by the m athem atical simulations shown in Figs 2 and 3. If one assumes that the transcription rates o f DBP target genes parallel the circadian accum ulation of DBP (shaded areas in Fig. 2) one can derive the accum ulation profiles for the corresponding mRNAs of any given half-life (shown for T 1/2 between 1 hour and 20 hours). Even if the transcriptional am plitude of a gene were infinite, the level o f mRNAs with half-lives of more than ten hours would fluctuate by two-fold or less (see Fig. 3, solid circles). However, the transcriptional fluctuation of most DBP target genes may only be five- to tenfold, since constitutively expressed activators in addition to DBP may be involved in controlling their expression. For mRNAs of such genes to cycle, their half-lives would have to be very short (see Fig. 3, open circles). Clearly then, DBP oscillation can be physiologically relevant only for those target genes whose mRNA and protein products are unstable.
Two particularly attractive candidates for DBP target genes are those encoding the liver-enriched enzymes HM G
126 J. Wuarin and others
a (m a x ) /a (m in )
10
O S(max)/S(min)= 5
• S(max)/S(min)= infinite
'O - .O -o.>-0 - 0-0
5 10 15
T1/2 (h ou rs )
2 0
Fig. 3. Amplitudes of mRNA accumulation as a function of halflives and amplitude of synthesis. Solid circles: Amplitudes of the accumulation (a) of mRNAs whose amplitude of synthesis (S) is infinite, such as shown in Fig. 2. Open circles: amplitudes of the accumulation (a) of mRNA whose amplitude of synthesis (S) is 5fold. The amplitudes (maximal/minimal accumulation) were determined graphically from accumulation profiles such as shown in Fig. 2.
Co A reductase and cholesterol 7 -a hydroxylase, which catalyze the rate-lim iting steps in cholesterol synthesis and cholesterol conversion to bile acids, respectively. Enzym e activities as well as mRNAs from these genes accum ulate according to a robust circadian rhythm with am plitudes of around 10-fold and a phase com patible with that o f DBP accum ulation (Clarke et a l , 1984; Noshiro et al., 1990; D. L. and J. W., unpublished results). Furtherm ore, and in keeping with DBP accum ulation, circadian expression of these enzymes is not detected until after weaning. Genomic clones encom passing the prom oter regions o f both o f these genes have been obtained and are currently being examined for the presence of DBP-binding sites. Thus far, such elem ents have been discerned within the cholesterol 7 -a hydroxylase promoter. Furthermore, in cotransfection experiments, a CAT reporter gene driven by this prom oter is strongly activated by DBP. W hile none o f the data thus far obtained prove that the circadian expression of cholesterol 7 -a hydroxylase and HM G CoA reductase is governed by DBP, this transcription factor appears to be a likely player in the control o f cholesterol homeostasis. In the model we are currently testing, DBP would sim ultaneously activate the diurnal production of bile acid and cholesterol. Such a coordination may be pivotal, since the m assive cholesterol utilization during the activity period (evening and night hours for rats and mice) may call for a large, sim ultaneous increase in its synthesis.
Conclusions and perspectives
DBP, a liver-enriched transcriptional activator of the bZip protein family, accum ulates according to an extraordinarily strong circadian rhythm. Preliminary results suggest that circadian DBP expression may be controlled, at least in part, by the cyclic secretion of glucocorticoid hormones. Like most (if not all) circadian activities in mammals, cor
ticosterone synthesis and secretion is governed by the suprachiasmatic nucleus (SCN), a small brain structure located above the optical chiasma. Circadian outputs are entrained in the SCN by external tim e cues, such as light/dark phases. Once entrained, such outputs are free- running and persist for extended time periods, until the clock is reset by new time cues (for references on the circadian pacem aker SCN see Kornhauser et al., 1992; M eijer and Rietveld, 1989). In the case o f adrenal hormone secretion, the SCN is believed to set the pace o f CRF secretion in the hypothalam us. This in turn may dictate the rhythmic release o f ACTH from the anterior pituitary gland, eventually resulting in a cyclic secretion o f corticosterone in the adrenal gland (M eijer and Rietveld, 1989).
Deciphering o f the physiological purpose o f DBP oscillation will require the unequivocal identification of downstream genes. A t present we are testing the hypothesis that one of the D BP functions is related to cholesterol hom eostasis, because the rate-limiting enzymes in cholesterol synthesis (HM G coA reductase) and utilization (cholesterol 7 -a hydroxylase) both fluctuate with a circadian phase com patible with that o f DBP accumulation. Clearly, the possible role of DBP in cholesterol metabolism will remain hypothetical until scrutinized by a thorough genetic analysis. The tools required for this endeavor have recently becom e available. In the case of DBP, both gain-of-func- tion (constitutive expression of DBP throughout the day) and loss-of-function (DBP gene knockout by homologous recombination) experiments should now be technically feasible.
W e were surprised to observe that, while transcription of
Fig. 4. Circadian transcription of two hypothetical DBP target genes containing DBP cognate sites with different affinities. The circadian DBP accumulation profile is schematically shown as a solid line. Target gene A is assumed to contain a 10-fold stronger DBP cognate site (equilibrium dissociation constant A'd=X) than gene B (equilibrium dissociation constant ATd=10X). As a consequence, half-saturation of the DBP site in gene B would require 10-fold higher DBP concentrations than that of its counterpart in gene A. Therefore, maximal transcriptional activity of gene A would be expected to persist for a longer circadian period than that of gene B. The periods of maximal transcription for genes A and B are depicted as black bars. The genes encoding cytochrome P450 CYP2C6 and cholesterol 7 -a hydroxylase constitute putative candidates for the gene classes A and B, respectively.

Circadian liver gene expression 127
certain genes, such as the one encoding serum albumin, cycles during the day, the accumulation of their products remains relatively constant. Apparently, diurnally expressed transcription factors like DBP may be used for genes whose products oscillate, such as HMG coA reductase and cholesterol 7 -a hydroxylase, and genes whose products accumulate constitutively, depending on their stability.
The relative affinity of the DBP sites for their cognate factor may be another important parameter in determining the temporal pattern of putative DBP target gene expression. As illustrated by the cartoon shown in Fig. 4, DBP target genes with low affinity sites may be maximally expressed during a short circadian time window, since the critical DBP concentrations required for efficient occupancy may only be reached during a few hours during the day. In contrast, genes with high affinity sites are expected to be expressed during much longer time periods, as even low DBP levels would suffice to fill the respective recognition sequences. For example, the cytochrome P450 gene CYP2C6 contains a DBP site within its promoter that binds E. coli-derived recombinant DBP with a 17-fold lower Kc\ than the albumin D promoter element (Yano et al., 1992). Run-on transcription experiments with nuclei isolated at four hour intervals around the clock yielded more or less constant transcription rates for the CYP2C6 gene (D. L., unpublished data). While further experiments are required to identify this gene as a bona fide DBP target gene, these considerations exemplify the versatile potentials of oscillating transcription factors: circadian DBP accumulation may result in strong, weak or no oscillation of mRNAs and proteins issued from downstream genes, depending on the relative affinities of their DBP-binding sites and the stabilities of their products.
We would like to thank Prof. G. Wanner, Department of Applied Mathematics, University of Geneva, for preparing the computer program used in Fig. 2. This research was supported by the Swiss National Science Foundation and the State of Geneva.
References
Clarke, C. F., Fogelman, A. M. and Edwards, P. A. (1984). Diurnal rhythm of rat liver mRNAs encoding 3-hydroxy-3-methylglutaryl coenzyme A reductase. J. Biol. Chem. 259, 10439-10447.
Crabtree, G., R., Schibler, U. and Scott, M. P. (1992). Transcriptional regulatory mechanisms in liver and midgut morphogenesis of vertebrates and invertebrates. In Transcriptional Regulation (ed. K. R. Yamamoto and S. L. McKnight). The Cold Spring Harbor Monograph (in press)
Descombes, P. and Schibler, U. (1991). A liver-enriched transcriptional activator, LAP, and a transcriptional inhibitory protein, LIP, are translated from the same mRNA. Cell 67,569-580
Edmunds, L. N. (1988). Cellular and M olecular Bases o f Biological Clocks. 497pp. New York: Springer Verlag.
Guillaume-Gentil, C., Rohner-Jeanrenaud, F., Bestetti, G. E., Abramo, F., Rossi, G. L. and Jeanrenaud, B. (1990). Abnormal regulation of the hypothalamo-pituitary-adrenal axis in the genetically obese fa/fa rat. Endocrinology 126, 1873-1879.
Hu, J. C., O ’Shea, E. K., Kim, P. S. and Sauer, R. T. (1990). Sequence requirements for coiled coils: Analysis with lambda repressor-GCN4 leucine zipper fusions. Science 250, 1400-1403.
Kornhauser, J. M., Nelson, D. E., M ayo, K. E. and Takahashi, J. S. (1992). Regulation o f jun-B mRNA and AP-1 activity by light and a circadian clock. Science 255, 1581 -1584.
Landschulz, W . H., Johnson, P. F. and M cKnight, S. L. (1989). The DNA-binding domain of the rat liver nuclear protein C/EBP is bipartite. Science 243, 1681-1688.
Lichtsteiner, S. and Schibler, U. (1989). A glycosylated liver-specific transcription factor stimulates transcription o f the albumin gene. Cell 57, 1179-1187.
Lichtsteiner, S., W uarin, J. and Schibler, U. (1987). The interplay of DNA-binding proteins on the promoter o f the mouse albumin gene. Cell 51,963-973.
M aire, P., W uarin, J. and Schibler, U. (1989). The role o f cis-acting elements in tissue-specific albumin gene transcription. Science 244, 343346.
M arais, R. M., Hsuan, J. J., McGuigan, C., W ynne, J. and Treisman, R.(1992). Casein kinase II phosphorylation increases the rate of serum response factor-binding site exchange. EM BO J. 11,97-105.
Meier, A. H. (1975). Daily variations o f concentration o f plasma corticosteroid in hypophysectomized rats. Endocrinology 98, 14751479.
M eijer, J. H. and Rietveld, W. J. (1989). Neurophysiology of the suprachiasmatic circadian pacemaker in rodents. Physiol. Rev. 69, 671 - 707.
Mueller, C. R., Maire, P. and Schibler, U. (1990). DBP, a liver-enriched transcriptional activator, is expressed late in ontogeny and its tissue- specificity is determined post-transcriptionally. Cell 61, 279-291.
Noshiro, M ., Nishimoto, M. an d O kuda, K. (1990). Rat liver cholesterol 7-alpha hydroxylase: Pretranslational regulation for circadian rhythm. J. Biol. Chem. 265, 10036-10041.
O ’Shea, E. K., Klemm, J. D ., Kim, P. S. and Alber, T (1991). X-ray structure of the GCN4 leucine zipper, a two-stranded, parallel, coiled coil. Science 254, 539-544.
O ’Shea, E. K., Rutkowski, R. and Kim, P. S. (1992). Mechanism of specificity in the Fos-Jun oncoprotein heterodimer. Cell 68, 699708.
Vinson, C. R., Sigler, P. B. and M cKnight, S. L. (1989). Scissor-grip model for DNA recognition by a family o f leuzine zipper proteins. Science 246, 911-916.
W uarin J. and Schibler, U. (1990). Expression of the liver-enriched activator protein DBP follows a stringent circadian rhythm. Cell 63, 1257-1266
Yano, M ., Falvey, E. and Gonzalez, F. J. (1992). Role o f the liver-enriched transcription factor DBP in expression of the cytochrome P450 CYP2C6 gene. Mol. Cell. Biol. 12, 2847-2854.
