the sequence of the mouse phosducin-encoding gene and its 5′-flanking region
TRANSCRIPT

Gene, 133 (1993) 179-186
0 1993 Elsevier Science Publishers B.V. All rights reserved. 0378-1119/93/$06.00 179
GENE 07386
The sequence of the mouse phosducin-encoding gene and its S-flanking region
(Recombinant DNA, introns; exons; phototransduction)
Toshiaki Abe, Takanobu Kikuchi, Thomas Chang* and Toshimichi Shinohara”
Laboratory of Retinal Cell and Molecular Biology, Bldg. 6, Rm. 327, National Eye Institute, National Institutes of Health, Bethesda, MD 20892, USA
Received by J. Piatigorsky: 17 March 1993; Revised/Accepted: 26_May/28 May 1993; Received by publishers: 1 July 1993
SUMMARY
Phosducin (Pd), a principal protein of retinal photoreceptor cells, modulates the phototransduction cascade by interacting with transducin. A recent report indicated that Pd and the G-protein-inhibitor protein (GIP) in brain are virtually identical. Here, we have sequenced the complete mouse clone (Pl-AT) carrying the Pd gene, 3026 bp of its S- flanking region, and cDNAs generated from the retinal mRNAs. Gene Pd is 15 kb in length and has four exons. The splice sites for donor and acceptor were in good agreement with the GT/AG rule. Deduced Pd amino acid sequences were highly homologous to those of human, bovine and rat. In addition, we found more than one similar Pd gene and two different mRNAs. The Pl-AT clone encodes one of the Pd mRNAs. These results open the possibility that the multiple Pd genes encode multiple Pd.
INTRODUCTION
Phototransduction is a fundamental process of all ani- mals with vision. This process takes place in the highly specialized outer segments of photoreceptor cells (Stryer, 1986; 1988; Chabre and Deterre, 1989). Phosducin (33- kDa Pd protein or MEKA) is a principal water soluble phosphoprotein in rod and cone photoreceptor cells (Lolley et al., 1977; Lee et al., 1982; 1984; 1988) and pine-
Correspondence to: T. Shinohara, Laboratory of Retinal Cell and
Molecular Biology, Bldg. 6, Rm. 327, National Eye Institute, National
Institutes of Health, Bethesda, MD 20892, USA. Tel. (l-301) 496-7799;
Fax (l-301) 402-0750.
*Permanent address: Stanford University, Stanford, CA, USA. Tel.
(1-415) 497-1642.
Abbreviations: aa, amino acid(s); A-kinase, cyclic adenosine 3’,5’-
monophosphate-dependent protein kinase; bp, base pair(s);
DTT, dithiothreitob GIP, G-protein inhibitor protein; kb, kilobase
or 1000 bp; nt, nucleotide(s); PBS, phosphate-buffered saline;
PCEl, photoreceptor conserved element (recognition site required for
photoreceptor-restricted gene expression); PCR, polymerase chain reac-
tion; Pd, phosducin; Pd, gene(s) encoding Pd, Pl-AT, phage Pl clone
of Pd; Td, transducin; tsp, transcriptional start point(s).
alocytes (Schaad et al., 1991). Pd modulates the photo- transduction cascade by binding to the Pr subunit complex of transducin (Tdby) to form a Pd-Tdpy com- plex (Lee et al., 1987; 1993). Previously, we have deter- mined the cDNA sequence of Pd and showed that the deduced aa sequence is highly conserved among mamma- lian species (Kuo et al., 1989; Abe et al., 1990). In addi- tion, Pd as well as its mRNA sequences in rat retina and the pineal gland are identical (Abe et al., 1990; Craft et al., 1991).
Phosphorylation of Pd is catalyzed by cyclic adenosine 3’S’-monophosphate-dependent protein kinase (A- kinase) (Lee et al., 1987; 1988), and phosphorylated Pd loses binding affinity to the transducin, and consequently loses inhibitory activity (Bauer et al., 1992).
Further, it is important to note that Pd in the retina or pineal gland is virtually identical with the GIP found in bovine brain (Bauer et al., 1992). The Pd (GIP) inhibits both GTPase activity of G proteins (transducin) and Gs- mediated adenyl cyclase activation in the signal transduc- tion and phototransduction cascades (Bauer et al., 1992; Lee et al., 1993). Thus, it emphasizes the need to under-

180
Fig. 1. A diagram of the Pi-AT phage Pl clone carrying Pd gene and
its restriction maps. Horizontal thin line indicates the introns and the
S- and the 3’-flanking regions. Boxes represent exons t-4. Restriction
maps for HindHI, EcoRI, BamHl and XbaI are shown below. Three
hybridization probes (probes 1,2, and 3) are represented by short hori-
zontal bars. Methods: The gene fragments were subcloned into the
BamHI site of hDASH phage (Stratagene, La Jolla, CA, USA). The nt
sequence for both strands was determined by the dideoxy chain termina-
tion method in a pBluescript IISK+ vector. (Stratagene) (Sanger et al.,
1980) and specific primers which were synthesized by an automated
DNA synthesizer (Biosearch model 8700, Burlington, MA). The resul-
tant sequences were analyzed using the Ideas program written by
Kanehisa (1986).
stand the regulatory role played by Pd in both transduc- tion cascades.
receptor cells and pineal gland, it is detected at very low levels in a wide range of tissues. Thus, Pd is of con- siderable interest with respect to its function and its gene expression in the specific tissues. In this report, we present sequences of a complete Pd gene (in Pl-AT clone). In addition, we have sequenced 2.6 kb of 5’-
Although Pd is a principal protein in the photo- flanking sequences of this gene in order to provide a
tm Fxon liltrOll Exon
76 1 77 GACCCAAA~/ptgagtctaca-----o---_catcata~TA~A~~GA
162 163 2 86 ACG CAC ACA G/~aactaatg----(2)----ttaamcp(llOA CCC MA GG
Thr His Thr Gly Pro Lys 308 309
3 152 G AGC AGA MGlptaagagaag---(3)----gcgttctc&ATG AGC ATT C Ser &a LVS Met Il.9
Fig. 2. Splicing junctions and the boundary sequences of the PI-AT
clone. The left column contains nt sequences at the 3’ end of the exons
and the 5’ end of the introns. The number in parentheses indicates each
intron (see Fig. 1). The right column contains the nt sequences at the
3’ end of introns and the 5’ end of the exons. The number above the
boundary is the nt position started from the presumptive tsp. The aa
sequence at each boundary is shown below the nt sequence and the
consensus phosphorylation sequence is underlined.
-2907
-2787
-2667
-2547
-2427
-2307
-2187
.2067
-1947
-1827
-1707
-1587
-1467
-1347
-1227
-1107
-987
-867
-747
-627
-507
-387
-267
-147
-21
Fig. 3. The 5’-flanking sequence of the PI-AT clone. The tsp (see Fig. 8) is numbered as + 1 (enlarged A); two other tsp (see Fig. 8) are indicated by
arrows at nt 5 and 6. A presumptive TATA box and two CAT-like boxes are underlined between positions -21 and -26 (TTAAAT), -38 and -42
(TCAAT), and -74 and - 78 (CCTAT). The repetitive sequences are indicated by underlines between -653 and -922 and - 1396 and - 1490.
Capital letter indicates the sequences of the first exon and the 5’-flanking region, lower case indicates the first intron.
181
data base for understanding the tissue restricted Pd gene expression and regulation. Additionally, we have estab- lished that Pl-AT clone indeed carries one of the Pd genes.
RESULTS AND DISCUSSION
(a) The Pd gene
To characterize the Pd gene, an independent phage Pl clone (Pl-AT) was isolated from a mouse Pl phage li- brary (Genome Systems Inc., St. Louis, MO, USA) using the mouse cDNA (Abe et al., 1990). The fragment (100 kb) was cleaved with restriction enzymes; HindIII, EcoRI, BumHI and XbaI and the resulted restriction maps are summarized in Fig. 1.
To define the relative positions of the exons and in- trons, nt sequences in all exons and part of introns of Pl- AT were determined. The Pd gene in Pl-AT was com- prised of four exons interrupted by three introns spanning approximately 15 kb in length. Each of the splice junc- tions was in good agreement with the GT/AG rule (Kozak, 1987) (Fig. 2). The phosphorylation consensus
sequence (RKXS) for A-kinase (Kemp et al., 1977) was
split by the third splicing junction. In order to define the boundary of the Pd gene, se-
quences in the S- and 3’-flanking regions were also deter- mined (Fig. 3). There are no authentic TATA, GC and CAAT boxes in 2609 bp of the 5’-flanking region. The closest similarity we have found was a TATA-like and two CAAT-like sequences between - 26 and -21 (TTAAAT), - 38 and -42 (TCAAT), and - 74 and - 78 (CCTAT). In the 3’-flanking region, we found a polyade- nylation signal sequence (AATAAA) between nt 1206 and 1219 and a conserved 12-bp element between nt 1365 and 1376 (Renen, 1987) (Fig. 5).
(b) Genomic Southern blot analysis
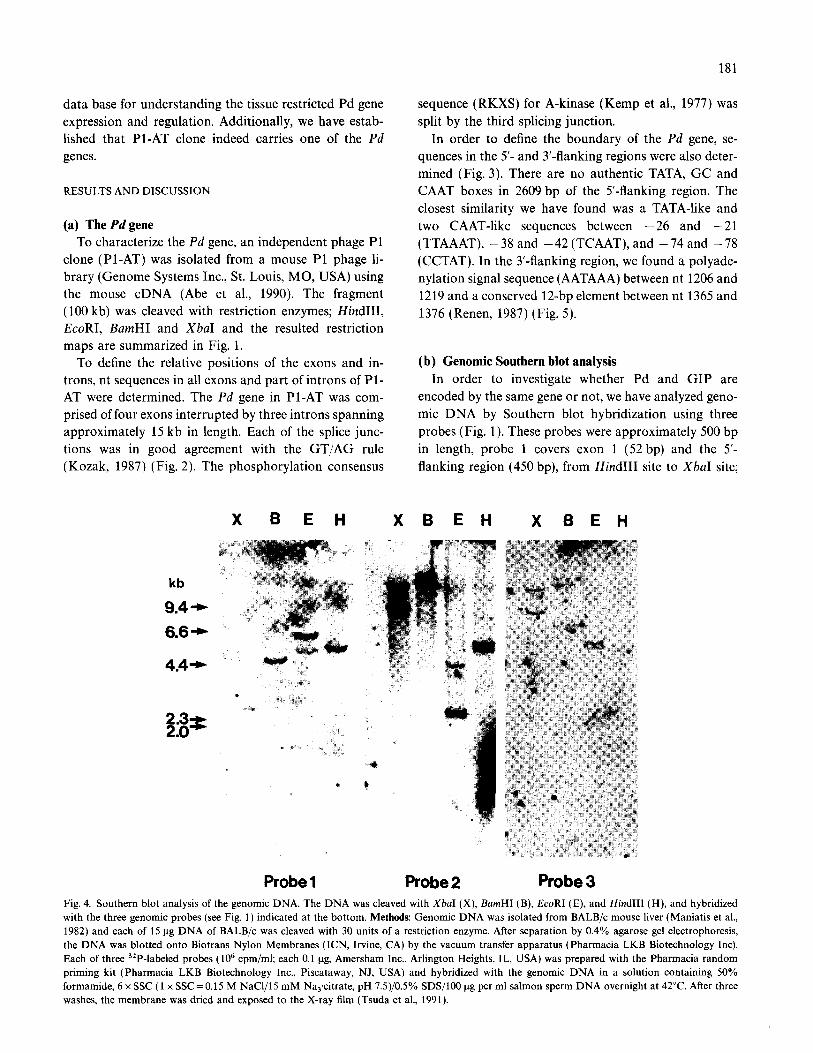
In order to investigate whether Pd and GIP are encoded by the same gene or not, we have analyzed geno- mic DNA by Southern blot hybridization using three probes (Fig. 1). These probes were approximately 500 bp in length, probe 1 covers exon 1 (52 bp) and the 5’- flanking region (450 bp), from Hind111 site to XbaI site;
X B E H XBEH X BEH
Probe 1 Probe 2 Probe 3 Fig. 4. Southern blot analysis of the genomic DNA. The DNA was cleaved with XbaI (X), BamHI (B), EcoRI (E), and Hind111 (H), and hybridized
with the three genomic probes (see Fig. 1) indicated at the bottom. Methods: Genomic DNA was isolated from BALB/c mouse liver (Maniatis et al.,
1982) and each of 15 ug DNA of BALB/c was cleaved with 30 units of a restriction enzyme. After separation by 0.4% agarose gel electrophoresis,
the DNA was blotted onto Biotrans Nylon Membranes (ICN, Irvine, CA) by the vacuum transfer apparatus (Pharmacia LKB Biotechnology Inc).
Each of three 32P-labeled probes ( lo6 cpm/ml; each 0.1 ug, Amersham Inc., Arlington Heights, IL, USA) was prepared with the Pharmacia random
priming kit (Pharmacia LKB Biotechnology Inc., Piscataway, NJ, USA) and hybridized with the genomic DNA in a solution containing 50%
formamide, 6 x SSC (1 x SSC=O.15 M NaCl/lS mM Na,citrate, pH 7.5)/0.5% SDS/l00 ug per ml salmon sperm DNA overnight at 42°C. After three
washes, the membrane was dried and exposed to the X-ray film (Tsuda et al., 1991).

182
probe 2 covers exon 2 (40 bp) and the second intron with different intensity. Since there are no internal (460 bp), from Hind111 site to EcoRI site; and probe XbaI, BumHI, EcoRI and Hind111 sites in these probes, 3 covers half of exon 4 (coding sequence from nt 308 the intensity of the band represents the gene dosage. One to 612) (see Figs. 1 and 5). Southern blot analysis batch of the genomic DNA was cleaved with the above with probe 1 showed only one band in the restriction restriction enzymes and hybridized with these probes; fragments of XbaI, BumHI and HindIII, two bands in therefore, it is unlikely that Southern blot analysis gener- the EcoRI fragments, as expected, since probe 1 has ates artifactual bands with probe 2 or 3. These results an internal EcoRI site (Fig. 4). This result indicates suggest that at least two genes are present in the genome. that the Pd gene is either one copy or has different Thus, taken together more than one Pd gene with some sequences in the 5’-flanking region. In contrast, the differences in the 5’-flanking region must be present in blots with probes 2 and 3 showed at least two bands the genome.
HR -_------___------____--_----___--_-----____----_-_::~::::~:::~:::::::~_::::::::~::~:~::::~,:~:~::~::,.,:::::.:::~::::::~:~ :G:
BR -----:G::::::.T:CCA:::::TA.::::T:TC:A:.:A~':C:A. A::G:::::::::.:,::A:A-:T: ~~.:TCTAT:A:~::TA.:T:lC:::~:::A::AG:AAA:M:C:.:T: G,
RR --------~~~--------~~ :::A::::::::.C:A,':::::: A:.:':G::T::::::::':::::C::: ": :::::::G ::A. ',:::::::>::::'.:::::A:::": ::...
RP ___________::,,::::~:;: :A. -:.:::.C:A:~:~:::::A:....G::T::.: ::.::::::C:::“::::.::::G:::A.. :‘::::::z::::: ::..:A: : ::':: :
c IrPr;AeCATTm;Alrr;A~T~T~~~T~~ffi~~~ TGATT~~CCAC~C~~T~T~C
265
:G::~:.C.~::~~:~:~'~::::A::T::~~~:.:.:~:::::.::::.:~ :'::,:.:, ::::::::::G::,C::::CA:::':::,:,: '.::T.:: :::.::G::T: :::G: .: .G:' ::C :: :, : :::::T:A::T::.:.:. :'::'::::::'::' .C!: ::.:::'::::.:..G:.G:: ::::..A:::::::.G,:G .:'::: .:::::G::T:::::::: ,...:
.G:: :::,:, :, '::::::::A::.::.: :.::':::,:.::: :' :C : .:: :: ';':::::: :.:::'::::::..:F..: .:. :'::".:G::::::':::' " : :G:: ::: : ‘.‘::::::::.A:::::, :.:,::::::::.:::::::C ‘: :::‘::::: ..,.._ ., ..,,::::::::‘.::.. : ‘::::’ .,.... .::G,::::::..:. ‘:.:::.
TCCTiPGAGCAGAGiTGAC------TCk 4.
M~CPr;AIW;ATGAGCATTCAPGAATATGAACTCATTCAT~~~~~~TT~C~T~~~CA~~~T 395
,::::::::T::GA:::G:AAK;AT.::::G.::c::G:C::::::::..:::::~::::..:: .':':::A.:C::A::::G::.::G:::::A:AC.. :::.:T.:.:::::T::A: ',:T::::: ::' ~T:.:~~~:::AAAG.Ac::: ::::.:::T:C~:::::.::::::::G:'..:::::~':::::A::C~:CA:::::. :::.:::::A:A::: :::.:T.:.:: .C::A:.::T:":. :.
..,, . . . . . . . . . :::AAAGAC::G::.:.:::.:::::::..:...: ::::I :...I ,::::::A:::. :::G::: ~':::::A&::: :::::~:::::~:C::A:. ::::':: ::, ,.. ,, :: ;AAAGAC:.G;;;,;,:;:.;;.:;;:;.;:;::...:;;::: :,. : ':.A:::::::,G,:::. ;~~::~:ffi.:.:~:~:~:T::~. ::C::A.. ::,:::: ::
,530
: : : ::C::::::C.: ::::::::::::A:::::::::~::~:::~:::::::::T::AA:::::~:::~A:::::A::T:~A::::~A:T::::A::..:.:::~:~:T:: C:':' :T..A:::::,AT: ~:.~:C.:::::C::::'::.::::::::A.::::.::::::::::.::.::.:T:T::::.::::::~:: :,::. ::T::A:::.:A:' ::Ak::::::T::C. T,:.C.T,:::.T:,A::.:::A:T: ..................... .C,:::C:: :::::::::.:::::::::::::::::::.::::::::.::.::..::..::::::::~~:.A::::::.:....~;;~;.~~C~~C; . :I.: -::’ :..:::.:::.:::
............. . . .. :C::::C:::..:::::~:::::::::::::::::: ::::::::::::::,:,:~:: ::::::::::::.A::::::.::::.:.::::::C::C::.::.: ::: ::.:::'.,::'::
-imm&AGcAGcT TnoCorCfiCrrGCK;T~~~~C~~T~T~~~~~~T~A~A~C~A~~ 664
AG::T.:T:.:::T::A::.::T::T:::A:A:::::::.::C:::A:.:::T:,A::T:::::T:::::::::::::::,:G:::::A::T.:T::::"":':::::TT:::T::: :T::::,AC:::CA: A&::.,T:v:::T::A:: T:::::GATA.::.:::..:CC.:A:.:.:T.::':::::::T:::::::::::G:::::~ ::.:A::T,:C::A:. '::':::: :T..::T::::::..C:~.C:..:T: . . . . . . :T:,C:::::::: :::::T::::AA ::::::.::C,.:::': .:. :::::::::::'::::::G::::::G:::::T:::,:T::: :::':: ::T::::'::::::::G: A,:..:CA: . . . . . ..T..c....::::,: .::T::::AA:::.:::,::C::.A:::::...,: ::::::::,::::::::G::: .:G.::.:T:::.:T::.: ::...:: :T:::::::::::.G: A' ::C::
GTACAkG TOGGGAACPCATA~G&TTTTATT~TGSCT&M TPTGCPG~~~mG~A~~~~~~TA~~T~~~~~~T~~~~~ 800
C::T::' ; :::~::~:::::::.::::~~.::.:T::::::::G::::~~ :::::::::::::::G:::::':.::::: :::::::::::::'::G:::':.::T:::::::::G:.::::T.:..: c:.: ::',: :::.:.:C::::::.:::C~:::::.:TA:::::::GC:G~.:::..:.:::~~:A:::G:~:: ::,::::':' :v::::::-::.::G:::T:. :T.::.A:::.::G,:::T,:' A::: ': : ,:.:G::::::::::.::::::::::::C:::::::::::'.:C,::::T.:::.C::::C, :. ::::::,': .: .:::::.:::::C:::::'.:. ': '::: ':':::: :. G, A:::.:... ":,.G :::::::::::.::"::::,:C:::::'::::::.:C:.' :T:::..C::.:C ::,'::::::':, :. ::' ::: :::C:,:::. :'.::',::: . . . . . . . . . . . . G.
CAGAccAAcirn~~ A~~~TCACCATGTOIGTATCTCATOTCTA-------T~T~~G~~A~~~T~T~TAT~ATAT~~~~~T 929
::T:'::.A:.A,::'.A::A::TG::::.,&AGA:::::T :::::A::::::::::T::------- ::::::GGT:T::------:' GA:GG ::-'.':::v:::: .:G::.:T:TG:..::AG.G .:::G::::::'.::::G---::T::G::.zAGA:::::T.:: ~:A~G:T::.:A:T:CGTATTTC.::::::Ic:.T~~------A~CACGA: ::-:::':::TC C :',T:..::GG::T::C:::C
:::::C. :.. T::A::T::C~:G~::A(Xj:::GG:.cAE~:::::G::A:::------..:.::G:::: ..M.CATC::.GG~-: :-,:,:::::::::,::.::.:::::: "'::
::::::.:::C~':::.T::A::T::C::G~::A!X GG::CIY;-::::::G::A.::-------,:::::G.: YAACATC:::G&.:.: :-::"::::.::::::::::::"::'.: ":
A~A~AIAcT(~KY~CC*~G---------G 1055
.'. ~~AC::::::GCT:::T:ACAA,UAT-------GAcGACTG:TG:A:C ::CGA::TAT:::::T::CA:::::T::G:::::::::':::T:C::.GC :G.~:A:::M~A:GC!ETC.:C--.,:':CA ::A:C:'::: :T::CF:T:T::::AA AAAAACTCKXXCTG:.::.:~~~~: A:GAT::'.:TO.C:::A.:T:~G::::Y':GC::T::::C:C ::. CT':AAC:TG.AGTC.:C--::: :TA :cA:T:~.c.__--__::.;:~~::::________#;:~::..:::~~ .., ,:A.:: ::-::,: ::..:: ::,:T:: : ::,:.: ::::: ::.,: ':.AAA. :::' .': :C:G::' :: ~cA:T~~c~____-_:~;:;~~:::;_-______#;:a;;;:.:::~:~; :, .A :: '_::; : ;,::.::.T. : ,:,:,:~:::~,:::~: :AA#::.:: ..:C:G:v::.::
AAmbMGcAGmP A~~TGTcM~TTT~cA~;~AAAMTCCCTTTCAA 1188
TTA:TffiT,:ACA.A:::AT:-:GGATAAT::TA:G:C::TT::C-:::::::':::: :: ::AT::T::GTC..,:TTA:TC::":.C::TCATCAC:G::ATTA,':::A: ,:,CA::lTAC:T::T:CA TG-~TPGT::ACA.A..:AATA-:G:AAAAT.CTG:~A:~T::TC.:.:~:::::::.:.::ATATGT.(X;CT::::TA:T:CATOC:::T:.:~ATTG:.A::~--C.A:.TT..:C-A::TTAT:T:OP:AA :::::C:G,A:: ~:":.:::ZL'GATGTC:ACG:: :G:::::,":':::.:::':,C..:::::' :.C::A:TAC~Cl'cC::::.:C:T GCA:::AT--.::T::GC.: TG:GG:: :: G::A: ::::.C:G:A::::.::~:::-G'lGATGTC:ACA::::G:: :: .:. :::::.::':C::::: ::~::::C..::TACT~CPCC:::::::C:T GCA:::AT--,::T::GC:::'E:ffi:: ': G,:A.
TTAT~TTGTA~A~~~~~T~CATCA~~T~~~TC~T~C~ AAAT&AAATWl+CATTTAT&TCTC 1334
. . . . ..TT.TC...::GTu. .G ::::TC::::::.m .:::T:T:C::::GT..':G:~G::TfX:Ab&::AAAAAAAAAAAAAAAA .:: .T:::.:::::TG. ,.G=G:::: '-:~.:::~.A.::.::.:C:::::T::GC:G.::.:Ck.G. ,,.... :Y. .TTT
':::: T.: :::":TG:: .G-G:::::':.:.-
CAATAkATAAATAiGCAT 1388
&TA~CCLACUTATA
Fig. 5. Sequence comparison of Pd in mammalian species. The mouse Pd gene (clone Pl-AT) sequence was compared with that of cDNAs of human
retina (HR), bovine retina (RR), rat retina (RR) and rat pineal gland (RP). The nt are numbered from the presumptive tsp of the mouse Pl-AT clone
(A at + 1). These nt identical to the mouse Pd are indicated by colons. The underlined ATG and TGA (or TAA) indicate the start and the stop
codons, respectively. The Pd gene had two repeats of AATAAA polyadenylation sites between nt 1212 and 1223 (underlined). A consensus 13-bp
element (TTRNNNNTTTTTT) is present between nt 1360 and 1371 and is underlined (R, purine; N any nt). Arrows indicate the splicing junctions.

183
(c) Sequence conservation of the Pd in mammalian species
To study sequence conservation of Pd in mammalian species, we have compared the sequences between clone Pl-AT and the previously published Pd cDNAs of human, bovine and rat. Mouse Pd nt sequence is highly conserved when compared with that of human (78%), rat (87%) and bovine (74%) (Fig. 5). Furthermore, marked protein sequence conservation over the 244-aa residues was revealed with the mouse sequence having high ho- mology (more than 98%) and an identity with rat (95%) and with human or bovine (88%) aa sequence (Fig. 6). Thus, the Pd and its mRNA sequences are highly con- served in mammalian species.
(d) Sequences in the S-non-coding regions of Pd mRNAs isolated from the retinas
The Southern blot analysis suggests that the Pd genes have heterogeneous sequences in the 5’ region, and differences can be detected in the sequence at the 5’-non- coding regions of the corresponding mRNAs. For this reason, retinal mRNAs were converted into cDNAs using a specific primer (GSP I) and partial sequence of the nine cDNAs was determined. The presence of heterogeneous sequences in the 5’-non-coding region (the first exon) of cDNAs was obvious (Fig. 7). The cDNAs can be classi- fied into two groups by their sequence similarity, one group (five cDNAs; ARl, AR2, AR3, AR4, and AR5) has an identical sequence with the corresponding region of the Pl-AT clone, and the other (four cDNAs; BC6, BC7, BC8 and BC9) has more than eight nt differences at the same positions in their sequences. The clones AR4 and AR5 had a different nt such as G, 53 and T, 72, respec- tively. Also ARl, AR2, and AR3 had one nt mismatch at the 5’ end. BC8 had C, not A, at position 25. These small
1 t -. . 60 PI-AT MSEAASQSLEEDFEGQATHTGPKGVINDWRKFKLESEMjD HR ::::K:::::::::::::::::::::::::::::::Q:S::::::::::::::::::::N
BR ::K:K::::::::::::S::::::::::::::::::::s::v*::::............. . . . . . . . . . . . . .
:::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: ~_sK~~~::::::::':::::::::i:::::::::l:::::::::~:::::::::~::::~2~
1QEYELIHQDKeDESCLRKYRRQCm)DMHoKLSFOPRYGFELETGEQFLE Gia::::V:;;;;::::::::KE::.. ..N::::::::::::::::::::::::::::::::K:::: :KD::::F:;1;;":::::::K:::::N::::::::::::::::::::::::::::::S:::::: :KD:::.....................G::::::::::::::::::::::::::::::::::::: ._._..._.......... :KD::::::;;;;::::::::::::::G:::::::::::::::::::::::::::::::::::::
TIEKEQ~TI.VVNIYEDGGCD~SSLACLAVE;PMSNTGAGDRFSTDVLPTLL 188
:::::L:I:::::H:::::IK:::::::::T:::A:::~::::::::::::::::::L::::::: . . . . . . . . . . . . . . ~:::::H:::::IK:::::::::~:::A::::::::::::::::::::::S::::::: :::::::::::::::::::::::::::::: E:::A::::::::::R:::::::::::S::::::: ::::::::::::::::::::::::::::::E:::A::::::::::R:::::::::::S:::::::
";KGGEL~SNF;SVAEQFAEEFFAVDVESFLNEYGLLPERE (2441 1::::::::::::::::::::::: G::::::::::::::::":v::H:KI:E::V: (2461 :::::::L::: :::T::L:::::TG::::::::::::::K:M:":::S:::-::M: (245) ::::::i::::::::::::: D:::A::::::::::::::::::::G:::T:::::: (2461 I:::::::::::::::::::D:::A::::::::::::::::::::G:::T:::::: (24.5)
Fig. 6. Comparison of aa sequences (for symbols see Fig. 5) in the mam-
malian species. The aa are numbered from the N-terminal, and residues
identical to those of mouse are indicated by colons. The numbers in
parentheses indicate the total residues. The phosphorylation site (at aa
position 71 in the mouse and 73 in the other species) is underlined and
splicing junctions are indicated by arrows.
differences might be PCR artifacts. Thus, our results indi- cate that two types of cDNA are present in the retinal preparation and one cDNA has an identical sequence with that of Pl-AT clone.
(e) The fsp in the Pl-AT clone Further studies were focused on the determination of
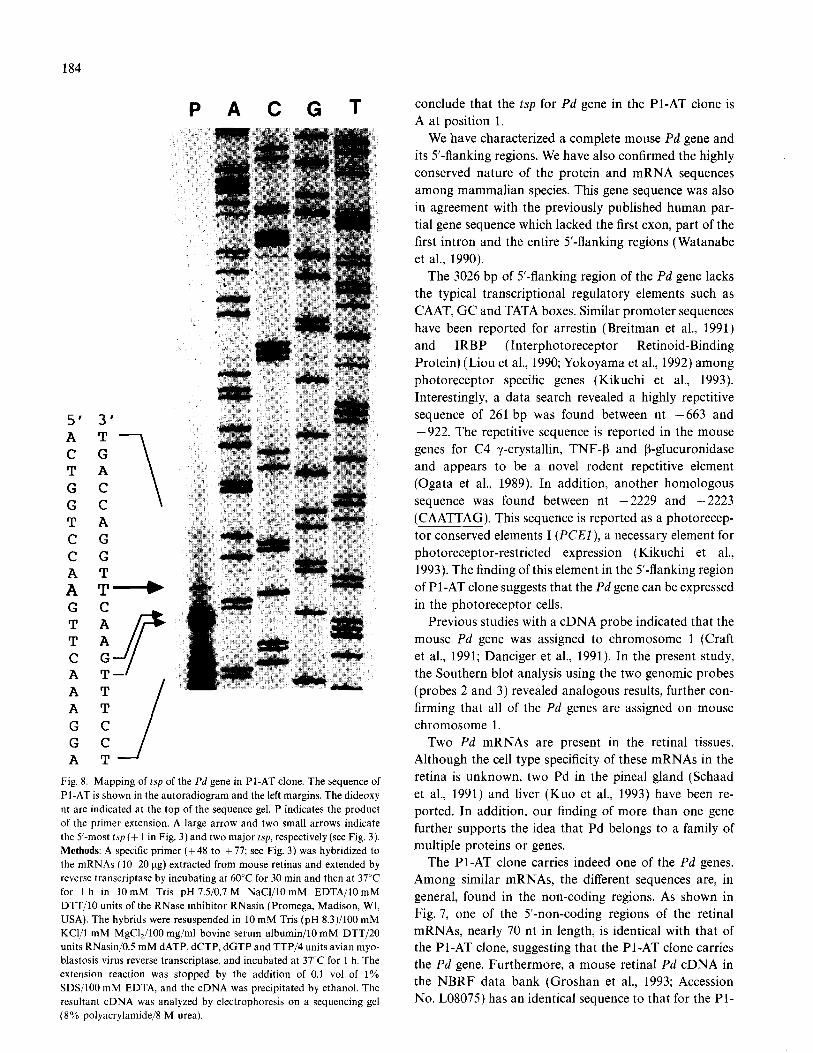
the tsp for Pd in the Pl-AT clone. For products from the primer extension with a specific primer (+ 48 to + 77), the largest product was 102 bp in length, as adenine as the 5’ nt (Fig. 8, also see Fig. 3; numbered as 1). In addi- tion, two high intensity bands corresponding to C and A at positions + 5 and + 6, respectively, were also found (Fig. 8 indicated by arrows). No extended product was found when yeast RNA was substituted in the primer extension experiment using the same probe. Thus, we
1 10 20 30 40 50 60 70 80 86
PI-AT - .
AGTTCAAAGGAGTTCAGTGGACAGCGATTCTCACTCCCTGGACATATTCTAG~CACCAGG~CAGAGACCC-AAACTgtgagtct
AR1 T::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::-::::: AR2 A::::::::::::::::::::................................................::::: . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . AR3 T::::::..._.....__........................_...._..................._~~~~: . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . AR4 ::::::::::::::::.... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ..G..:::........ . . . . .._..r.: . . . . . . . . . . . . . . AR5 :::::::: :::::::::::::::::::::::::::::::::::::::::::::::: T-:::::
BC6 GTT:CCT:CGCAAAT::C:A:::::::::C:A:::::::::A:::::G::T::::::::::::::::C::::: BC7 GTT:CCT:CGCAAAT::C:A:::::::::C:A:::::::::A:::,::G::T::::::::::::::::C::::: BC8 GTT:CCT:CGCAAAT::C:::::....-.C:A:::::::::A:::::G::T::::::::::::::::C::::: . . . . . . BC9 GTT:CCT:CGCAAAT::C:A:::::::::C:A:::::::::A:::::G::T::::::::::::::::C:::::
Fig. 7. Sequences in the first exon and part of the first intron of the Pd gene (clone Pl-AT) and part of the non-coding region of the Pd cDNAs (AR1
through AR5 and BC6 through BC9) generated from mouse retinal mRNAs. The nt positions are numbered from the tsp of the Pl-AT clone. Capital
and lower-case letters in Pl-AT clone indicate the sequences of the first exon and the first intron, respectively. Retinal mRNAs were isolated and
10 pmole of primer GSP I (nt + 595 to +615 of Pl-AT clone; see Fig. 5) were hybridized to 0.8 ug of RNAs and extended by reverse transcriptase
(Boehringer Mannheim, Indianapolis, IN, USA). A poly(C) tail was attached to the 5’ end of the cDNA by terminal transferase. The specific GSP
II (nt + 354 to + 374 of Pl-AT; see Fig. 5) and poly(G) (Anchor Primer) primers were used for PCR amplification as described elsewhere (Frohman
et al., 1988). The resultant double-stranded cDNAs were digested with SalI+HindIII, fragments larger than 200 bp were separated by the Spin
Cartridge System (BRL) and subcloned into a pBluescript vector (Abe et al., 1990).
184
P A C G T
5’ 3’ A T C G T A G C G C \
T A C G C G
: ;- G C T A T A C G
r A T- A T A T G C G C A T /
Fig. 8. Mapping of tsp of the Pd gene in Pl-AT clone. The sequence of
Pl-AT is shown in the autoradiogram and the left margins. The dideoxy
nt are indicated at the top of the sequence gel. P indicates the product
of the primer extension. A large arrow and two small arrows indicate
the 5’-most tsp (+ 1 in Fig. 3) and two major tsp, respectively (see Fig. 3).
Methods: A specific primer (+48 to + 77; see Fig. 3) was hybridized to
the mRNAs (lo-20 ug) extracted from mouse retinas and extended by
reverse transcriptase by incubating at 60°C for 30 min and then at 37°C
for 1 h in 10 mM Tris pH 7.5/0.7 M NaCI/lOmM EDTAjlO mM
DTTjlO units of the RNase inhibitor RNasin (Promega, Madison, WI,
USA). The hybrids were resuspended in 10 mM Tris (pH 8.3)/100 mM
KCljl mM MgClJlOO mg/ml bovine serum albumin/l0 mM DTT/20
units RNasimO.5 mM dATP, dCTP, dGTP and TTP/4 units avian myo-
blastosis virus reverse transcriptase, and incubated at 37°C for 1 h. The
extension reaction was stopped by the addition of 0.1 vol. of 1%
SDS/l00 mM EDTA, and the cDNA was precipitated by ethanol. The
resultant cDNA was analyzed by electrophoresis on a sequencing gel
(8% polyacrylamide,B M urea).
conclude that the tsp for Pd gene in the Pl-AT clone is A at position 1.
We have characterized a complete mouse Pd gene and its 5’-flanking regions. We have also confirmed the highly conserved nature of the protein and mRNA sequences among mammalian species. This gene sequence was also in agreement with the previously published human par- tial gene sequence which lacked the first exon, part of the first intron and the entire 5’-flanking regions (Watanabe et al., 1990).
The 3026 bp of 5’-flanking region of the Pd gene lacks the typical transcriptional regulatory elements such as CAAT, GC and TATA boxes. Similar promoter sequences have been reported for arrestin (Breitman et al., 1991) and IRBP (Interphotoreceptor Retinoid-Binding Protein) (Liou et al., 1990; Yokoyama et al., 1992) among photoreceptor specific genes (Kikuchi et al., 1993). Interestingly, a data search revealed a highly repetitive sequence of 261 bp was found between nt -663 and -922. The repetitive sequence is reported in the mouse genes for C4 y-crystallin, TNF-h and b-glucuronidase and appears to be a novel rodent repetitive element (Ogata et al., 1989). In addition, another homologous sequence was found between nt -2229 and -2223 (CAATTAG). This sequence is reported as a photorecep- tor conserved elements I (PCEI ), a necessary element for photoreceptor-restricted expression (Kikuchi et al., 1993). The finding of this element in the 5’-flanking region of PI-AT clone suggests that the Pd gene can be expressed in the photoreceptor cells.
Previous studies with a cDNA probe indicated that the mouse Pd gene was assigned to chromosome 1 (Craft et al., 1991; Danciger et al., 1991). In the present study, the Southern blot analysis using the two genomic probes (probes 2 and 3) revealed analogous results, further con- firming that all of the Pd genes are assigned on mouse chromosome 1.
Two Pd mRNAs are present in the retinal tissues. Although the cell type specificity of these mRNAs in the retina is unknown, two Pd in the pineal gland (Schaad et al., 1991) and liver (Kuo et al., 1993) have been re- ported. In addition, our finding of more than one gene further supports the idea that Pd belongs to a family of multiple proteins or genes.
The Pl-AT clone carries indeed one of the Pd genes. Among similar mRNAs, the different sequences are, in general, found in the non-coding regions. As shown in Fig. 7, one of the 5’-non-coding regions of the retinal mRNAs, nearly 70 nt in length, is identical with that of the Pl-AT clone, suggesting that the Pl-AT clone carries the Pd gene. Furthermore, a mouse retinal Pd cDNA in the NBRF data bank (Groshan et al., 1993; Accession No. LO8075) has an identical sequence to that for the Pl-

185
AT clone. Only two nt differences are found between the two sequences (absence of C, 914 and one extra T inser- tion between nt 1135 and 1136 in the cDNA) and a stretch of 19 nt at the 5’ end of the cDNA. These 19 nt may be rearranged, since they are located on cDNA at the same position as the Pd tsp in the Pl-AT clone. Thus, the sequence of the mouse retinal cDNA strongly sup- ports that the Pl-AT contains the Pd gene.
In order to study the cell type specificity of the Pl-AT Pd gene expression, we have constructed a fusion gene which contains 3.3 kb of S-flanking region of the Pl-AT clone and the bacterial reporter gene lacZ. Analysis of lacZ expression in various tissues of transgenic mice car- rying the fusion gene will provide exact profiles of Pd gene expression. This study is currently in progress in our laboratory.
(f ) Conclusions
A mouse Pd gene and 3026 bp of its 5’-flanking se- quences were determined. The Pd gene is 15 kb in length and has four exons. The 5’-flanking sequence has no au- thentic CAAT, CG and TATA boxes, but had a PCEl site, a necessary element for photoreceptor restricted gene expression. The splice sites for donor and acceptor were in good agreement with the GT/AG rule. The Pd se- quences in mammalian species are highly conserved. More than one Pd gene and two mRNAs were found. These results open a possibility that the Pd gene belongs to a family of multiple genes.
ACKNOWLEDGEMENTS
We thank Drs. Michael Redmond and Peggy Zelenka for critical reading and discussions of this manuscript.
REFERENCES
Abe, T., Nakabayashi, H., Tamada, H., Takagi, T., Sakuragi, S.,
Yamaki, K. and Shinohara, T.: Analysis of the human, bovine and
rat 33-kDa proteins and cDNA in retina and pineal gland. Gene 91
(1990) 209-215.
Bauer, P.H., Muller, S., Puzicha, M., Pippig, S., Obermaier, B.,
Helmreich, E.J.M. and Lohse, M.J.: Phosducin is a protein kinase
A-regulated G-protein regulator. Nature 358 (1992) 73-76.
Breitman, M.L., Tsuda, M., Usukura, J., Kikuchi, T., Zucconi, A.,
Khoo, W. and Shinohara, T.: Expression of S-antigen in retina,
pineal gland, lens, and brain is directed by 5’-flanking sequences.
J. Biol. Chem. 266 (1991) 15505515510.
Chabre, M. and Deterre, P.: Molecular mechanism of visual transduc-
tion. Eur. J. Biochem. 179 (1989) 255-266.
Craft, M.C., Lolley, R.N., Seldin, M.F. and Lee, R.H.: Rat pineal gland
phosducin: cDNA isolation, nucleotide sequence, and chromosomal
assignment in the mouse. Genomics 10 (1991) 400-409.
Danciger, M., Kozak, C.A., Abe, T., Shinohara, T. and Farber, D.B.:
The gene for retinal rod 33-kDa protein on mouse chromosome 1
near Lamb 2. Cell Genet. 56 (1991) 202-205.
Frohman, M.A., Dush, M.K. and Martin, G.R.: Rapid production of
full-length cDNAs from rare transcripts: amplification using a single
gene-specific oligonucleotide primer. Proc. Natl. Acad. Sci. USA 8.5
(1988) 8998-9002.
Groshan, K.R., Norton, J.C., Craft, C.M. and Travis, G.H.: In NBRF
data bank; Accession No. LO8075 (1993).
Kanehisa, M.: Integrated Database and Extended Analysis System for
Nucleic Acids and Proteins. National Cancer Institute, Frederick
Cancer Research Facility, Frederick, MD, USA, 1986.
Kemp, B.E., Graves, D.J., Benjamini, E. and Krebs, E.G.: Role of
multiple basic residues in determining the substrate specificity of
cyclic AMP-dependent protein kinase. J. Biol. Chem. 252 (1977)
4888-4894.
Kikuchi, T., Raju, K., Breitman, M.L. and Shinohara, T.: The proximal
promoter of the mouse arrestin gene directs gene expression in pho-
toreceptor cells and contains an evolutionarily conserved retinal
factor binding site. Mol. Cell. Biol 13 (1993) 4400-4408.
Kozak, M.: An analysis of S-noncoding sequences from 699 vertebrate
messenger RNAs. Nucleic Acids Res. 15 (1987) 812558133.
Kuo, C.-H., Akiyama, M. and Miki, N.: Isolation of a novel retina-
specific clone (MEKA cDNA) encoding a photoreceptor soluble
protein. Mol. Brain Res. 6 (1989) l-10.
Kuo, C-H., Taira, E., Takaha, N., Soma, H., Akino, T., Fukada, Y.,
Sanada, K. and Miki, N.: Purification and characterization of three
MEKA-like proteins in liver: association of a 94 kDa protein with
By subunits of G-proteins. Biochem. Biophys. Res. Commun. 191
(1993) 109771104.
Lee, R.H., Brown, B.M. and Lolley, R.N.: Autophosphorylation of rho-
dopsin kinase from retinal rod outer segments. Biochemistry 21
(1982) 3303-3307.
Lee, R.H., Brown, B.M. and Lolley, R.N.: Light-induced dephosphory-
lation of a 33K protein in rod outer segments of rat retina.
Biochemistry 23 (1984) 1972-1977.
Lee, R.H., Lieberman, B.S. and Lolley, R.N.: A novel complex from
bovine visual cells of a 33,000-dalton phosphoprotein with B- and
y-transducin: purification and subunit structure. Biochemistry 26
(1987) 3983-3990.
Lee, R.H., Whelan, J.P., Lolley, R.N. and McGinnis, J.F.: The photore-
ceptor-specific 33 kDa phosphoprotein of mammalian retina: gener-
ation of monospecific antibodies and localization by
immunocytochemistry. Exp. Eye Res. 46 (1988) 829-840.
Lee, R.H., Ting, D., Lieberman, B.S., Tobias, D.E., Lolly, D.N. and Ho,
Y.-K.: Regulation of retinal cGMP cascade by phosducin in bovine
rod photoreceptor cells: interaction of phosducin and transducin.
J. Biol. Chem. (1993) in press.
Liou. G., Geng, L.I., Al-Abaidi, M.R., Matragoon, S., Hanten, G.,
Baehr, W. and Overbeek, P.A.: Tissue-specific expression in
transgenic mice directed by the 5’-flanking sequences of the human
gene encoding interphotoreceptor retinoid binding protein. J. Biol.
Chem. 265 (1990) 837338376.
Lolley, R.N., Brown, B.M. and Farber, D.B.: Protein phosphorylation
in rod outer segments from bovine retina: cyclic nucleotide-activated
protein kinase and its endogenous substrate. Biochem. Biophys. Res.
Commun. 78 (1977) 572-578.
Maniatis, T., Fritsch, E.F. and Sambrook, J.: Molecular Cloning. A
Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring
Harbor, NY, 1982.
Ogata, R.T., Rosa, P.A. and Zepf, N.E.: Sequence of the gene for murine
complement component C4. J. Biol. Chem. 264 (1989) 16565-16572.
Renan, M.J.: Conserved 12-bp element downstream from mRNA poly-
adenylation sites. Gene 60 (1987) 245-254.
Sanger, F., Coulson, A.R., Barrel, G.B.G., Smith, A.J.H. and Roe, B.A.:
Cloning in single-stranded bacteriophage as an aid to rapid DNA
sequencing. J. Mol. Biol. 143 (1980) 161-178.
Schaad, N.C., Shinohara, T., Abe, T. and Klein, D.C.: Photoneural

186
control of the synthesis and phosphorylation of pineal MEKA (phosducin). Endocrinology 129 (1991) 3289-3298.
Stryer, L.: Cyclic GMP cascade of vision. Annu. Rev. Neurosci. 9 (1986) 87-119.
Stryer, L.: Molecular basis of visual excitation. Cold Spring Harbor Symp. Quant. Biol. 53 (1988) 283-294.
Tsuda, M.. Kikuchi, T., Yamaki, K. and Shinohara, T.: The mouse S-antigen gene: comparison with human and Drosophila. Eur. J. Biochem. 200 (1991) 95-101.
Yamaki, K., Tsuda, M., Kikuchi, T., Chan, K.-H., Huang, K.-P. and
Shinohara, T.: Structural organization of the human S-antigen gene. J. Biol. Chem. 265 (1990) 20757-20762.
Yokoyama, T., Liou, G.L., Caldwell, R.B. and Overbeek, P.A.: Photoreceptor-specific activity of the human interphotoreceptor re- tinoid-binding protein (IRBP) promoter in transgenic mice. Exp. Eye Res. 55 (1992) 225-233.
Watanabe, Y., Kawasaki, K., Miki, N. and Kuo, C-H.: Isolation and analysis of the human gene encoding a retina-specific protein. Biochem. Biophys. Res. Commun. 170 (1990) 951-956.