the single functional blast resistance gene pi54 activates a complex
TRANSCRIPT

Journal of Experimental Botany Vol 63 No 2 pp 757ndash772 2012doi101093jxberr297 Advance Access publication 4 November 2011
RESEARCH PAPER
The single functional blast resistance gene Pi54 activatesa complex defence mechanism in rice
Santosh Kumar Gupta12 Amit Kumar Rai12 Shamsher Singh Kanwar2 Duni Chand2 Nagendera Kumar Singh1
and Tilak Raj Sharma1
1 National Research Centre on Plant Biotechnology Indian Agricultural Research Institute Pusa Campus New Delhi-110012 India2 Department of Biotechnology Himachal Pradesh University Summer Hill Shimla-171005 India
To whom correspondence should be addressed E-mail trsharmanrcpborg trsharma1965gmailcom
Received 5 July 2011 Revised 19 August 2011 Accepted 22 August 2011
Abstract
The Pi54 gene (Pi-kh) confers a high degree of resistance to diverse strains of the fungus Magnaporthe oryzae In
order to understand the genome-wide co-expression of genes in the transgenic rice plant Taipei 309 (TP) containing
the Pi54 gene microarray analysis was performed at 72 h post-inoculation of the M oryzae strain PLP-1 A total of
1154 differentially expressing genes were identified in TP-Pi54 plants Of these 587 were up-regulated whereas 567
genes were found to be down-regulated 107 genes were found that were exclusively up-regulated and 58 genes that
were down- regulated in the case of TP-Pi54 Various defence response genes such as callose laccase PAL and
peroxidase and genes related to transcription factors like NAC6 Dof zinc finger MAD box bZIP and WRKY werefound to be up-regulated in the transgenic line The enzymatic activities of six plant defence response enzymes
such as peroxidase polyphenol oxidase phenylalanine ammonia lyase b-glucosidase b-13-glucanase and
chitinase were found to be significantly high in TP-Pi54 at different stages of inoculation by M oryzae The total
phenol content also increased significantly in resistant transgenic plants after pathogen inoculation This study
suggests the activation of defence response and transcription factor-related genes and a higher expression of key
enzymes involved in the defence response pathway in the rice line TP-Pi54 thus leading to incompatible hostndash
pathogen interaction
Key words Defence enzyme global expression analysis Magnaporthe oryzae Pi-kh rice blast transcription factors transgenic
lines
Introduction
Rice (Oryza sativa L) is the most important staple food for
more than half of the worldrsquos population most of which
live in developing countries (von Braun 2007) Globally
over 150 million hectares of paddy is planted annually and
the production is close to 600 million tons Rice is mostlygrown on subsistence agriculture farms in Asia with most
grains used for local consumption (David 1991) China and
India combined account for more than half of the worldrsquos
rice areas and consume more than three-quarters of the
global rice production (Hossain 1997 MacLean et al
2002) In India rice cultivation is mostly concentrated in
the hilly areas river valleys deltas and low-lying coastal
areas which together contribute about 97 to the countryrsquos
rice production
The rice crop is affected by various abiotic and bioticstresses thus resulting in huge yield losses wherever rice is
grown Among the biotic stresses blast disease caused by
a compatible interaction between rice and Magnaporthe
oryzae results in a US$55 million loss in rice production
each year in South and South-east Asia (Padmanabhan
Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phephenylalanine CA cinnamic acid SA salicylic acid Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases PPO polyphenol oxidase Laclaccases ETI effector-triggered immunity PAMP pathogen-associated molecular patternsordf The Author [2011] Published by Oxford University Press [on behalf of the Society for Experimental Biology] All rights reservedFor Permissions please e-mail journalspermissionsoupcom
Dow
nloaded from httpsacadem
icoupcomjxbarticle632757499789 by guest on 26 D
ecember 2021
et al 1974) These losses are even higher in East Asia and
other more temperate rice-growing regions around the
world (Talbot 2003) Rice blast can be effectively managed
by using resistant rice varieties for which the identification
and characterization of appropriate blast resistance (R)
genes are vital To date more than 19 blast resistance genes
have been cloned and characterized (Sharma et al 2005b
Okuyama et al 2011) However none of them has beenshown to have resistance potential against all the strains of
M oryzae Recently it was reported in a separate study that
Pi54 (Pi-k-h cloned from the rice line Tetep) confers broad
spectrum resistance against geographically diverse strains of
M oryzae collected from various parts of India and the US
(Costanzo and Jia 2010 Rai et al 2011)
Several attempts to utilize R-genes in rice blast by de-
veloping disease-resistant cultivars have been futile owing tothe lack of information regarding the intricate mechanisms
involved in R-gene-mediated resistance pathways and the
high rate of variability in the corresponding avirulence (Avr)
genes Therefore in order to develop sustainably successful
blast-resistant rice lines a comprehensive dissection of the
reactions downstream of RndashAvr interactions is highly desir-
able The co-expression patterns of bacterial disease resistance
genes and their transcriptional regulators in transgenic ricehave indicated that the resistance genes trigger an immune
response (Sana et al 2010 Zhou et al 2010) The expression
patterns of defence response genes involved in ricendashM oryzae
interaction have been widely studied (Kim et al 2001 Xiong
et al 2001 Rao et al 2002 Jantasuriyarat et al 2005
Soderlund et al 2006 Vergne et al 2008 Kato et al 2009)
However an expression study in response to the blast
pathogen in transgenic rice containing a single gene for blastresistance still remains to be performed Further downstream
biochemical events resulting from gene-for-gene interactions
have only been studied in blast-resistant plants (Hammond-
Kosack and Jones 2000 Dangl et al 1996 Ryals et al
1996) and the impact of rice blast R-genes in the transgenic
system remains unexplored
Previously the rice blast resistance gene Pi-kh has been
isolated from the indica rice line Tetep showing resistance todifferent M oryzae strains in the North-Western Himalayan
region of India (Sharma et al 2005a) This gene is only the
third to be cloned in the series of rice blast resistance genes
after the cloning of Pib (Wang et al 1999) and Pita (Bryan
et al 2000) The gene was renamed Pi54 after it was further
re-alienated to a slightly distant location from the Pik locus
(Sharma et al 2010) The predicted Pi54 protein contains
a nucleotide-binding sitendashleucine-rich repeat (NBS-LRR)domain in addition to a small zinc finger domain (Sharma
et al 2005a b) Functional complementation of the gene
has confirmed its requirement for conferring a stable and
high level of resistance against geographically diverse strains
of M oryzae from different parts of India (Rai et al 2011)
Also this gene is widely being used in a pyramiding
programme for broad spectrum resistance to blast in
combination with Piz5 in Pusa Basmati rice varieties (Singhet al 2011) The gene has been shown to induce the
synthesis of callose (b-13-glucan) in response to pathogen
challenge The Pi54 gene is expressed constitutively at
a basal level in both resistant transgenic as well as
susceptible native lines up to 48 hpi Similar to the Pib
(Wang et al 1999) and Pi5-1 (Lee et al 2009) blast
resistance genes of rice the expression of Pi54 is induced by
M oryzae infection and a more than 2-fold higher in-
duction was observed at 72 hpi in the resistant line (Rai
et al 2011) Further studies of the expression profiles ofblast-resistant and susceptible lines at 72 hpi have yet to be
completed using microarray analysis
In this study our objectives were (i) to investigate and
compare using microarray-based transcriptome analysis the
global gene expression changes happening in response to
infection by M oryzae in transgenic rice lines containing the
rice blast resistance gene Pi54 (ii) to identify and validate the
potential genes involved in Pi54-mediated blast resistanceand (iii) to perform biochemical analysis of the enzymes and
phenolic compounds involved in the disease response path-
ways during compatible and incompatible interactions Our
results showed a Pi54-mediated reprogramming of a broad
array of genes which apparently leads to an M oryzae-
resistant phenotype via detectable changes in the activities of
the enzymes involved in the defence pathways
Materials and methods
Plant and fungal materials
The blast-resistant transgenic rice line TP-Pi54-15 used in thisstudy was originally derived from the cloned gene Pi54 (Pi-kh)transferred into Oryza sativa cv Taipei (TP) 309 which is a highlysusceptible to the M oryzae strain PLP-1 The gene was stable andexpressed in subsequent generations (T3) and was being main-tained in our laboratory for subsequent use (Rai et al 2011)Transgenic rice lines were maintained in a containment facility atthe National Phytotron Facility (NPF) Indian Agricultural Re-search Institute New Delhi India
Fifteen-day-old seedlings of TP-Pi54-15 were inoculated with anM oryzae spore suspension containing 105 spores ml1 in 025gelatin until the leaves were covered with fine droplets For themock control plants were inoculated with 025 gelatin only Theexperiment was carried out under controlled growth conditions at2562 C and 90 relative humidity for 24 h in the dark and thenshifted to a 168 h lightdark regime After 72 hpi leaves werecollected washed and pooled for RNA extraction For bio-chemical analyses challenge-inoculated leaf samples along withnon-inoculated controls of transgenic (TP-Pi54) and non-transgenic (TP) lines were harvested after 0 24 48 72 96 and120 hpi for enzyme extraction
RNA isolation and microarray analysis
Total RNA was extracted from transgenic and non-transgenic ricelines using the TRI Reagent (Invitrogen) Plant tissues werehomogenized using a mortar and pestle with liquid nitrogen and1 ml of TRI Reagent was added per 100 mg of tissue Quantitativeand qualitative assessment of the isolated RNA was done usinga UV-Vis Spectrophotometer (NanoDrop Technologies WilmingtonDE) and by gel electrophoresis respectively RNA sampleswere processed according to the standard Affymetrix GeneChipexpression analysis technical manual (Affymetrix Inc Santa ClaraCA USA) Total RNA was used for the synthesis of cDNA usinga SuperScript double-stranded cDNA synthesis kit A portion of theresulting double-stranded cDNA was used as a template to generate
758 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
biotin-tagged cRNA from an in vitro transcription reaction using anAffymetrix GeneChip IVT labelling kit Fifteen micrograms ofbiotin-tagged cRNA was fragmented (50 to 200 bp fragments) as perthe manufacturerrsquos protocols Subsequently 10 lg of this fragmentedcRNA was hybridized to rice gene chips (Affymetrix) at 45 C with16 h rotation in a GeneChip hybridization oven (Affymetrix)Duplicate biological samples from transgenic and control rice plantswere used for microarray analysis Each array contained probe setsfor 51 279 rice transcripts out of which around 48 564 were fromjaponica and 1260 from indica cultivars Seventy-two hours aftertreatment total RNA was isolated from leaf tissues inoculated withthe pathogen The samples from 20ndash30 challenge-inoculated leaveswere pooled together Total RNA was converted into biotin-labelledcRNA and hybridized to the rice gene chip Separate gene chips werehybridized for the M oryzae spore suspension prepared in 02gelatin and leaf samples mock-inoculated with 02 gelatin onlywere used as the mock control GeneChip arrays were washed andthen stained with streptavidin-phycoerythrin (Affymetrix FluidicsStation 450) followed by scanning with a GeneChip scanner(Affymetrix GeneChip Scanner 3000) FCA (fold change absolute)values were calculated for each transcript of TP-Pi54 and TP usingmock-inoculated chip data as their respective controls The FCAvalues of transgenic and native lines for each transcript werecompared
Microarray data analysis
MIAME-compliant microarray data from this study have beendeposited at NCBI Gene Expression Omnibus with the accessionnumber GSE28308 The array data set was analysed using theGeneChip Operating Software (GCOS 12) and ArrayStar(DNASTAR Inc Madison USA) software For probe levelnormalization robust multi-array analysis (RMA) was performedon Affymetrix raw files (CEL files) (Irizarry et al 2003) Theinitial data set consisted of 57 382 probes Filtration wasperformed to remove probes with lsquoAbsent Callsrsquo (poor qualityprobes) Transformation and normalization were done in order tofacilitate comparison across samples The biological significance ofdifferentially expressed genes was determined by performingfunctional classification according to Gene Ontology after filteringthe genes for gt20-fold change and lt005 P-value Gene annota-tion was performed using BLAST (httpblastncbinlmnihgovBlast) All the raw data files (CEL files two for blast-inoculatedand two for mock treatment) obtained from the GeneChipOperating Software (GCOS Affymetrix) were subjected to furtheranalysis using the ArrayStar software
Quantitative real-time PCR analysis
Part of the samples of total RNA isolated for microarray analysiswas also used to validate the functionally important genes that werefound to be differentially regulated in the microarray results Thenucleotide sequences of differentially regulated genes were down-loaded from the TIGR rice database (httpriceplantbiologymsuedu) (see Supplementary Table S5 at JXB online) The exonicsequences of selected genes were used to design primers using thePRIMER3 software (httpfrodowimiteduprimer3) For the genevalidation experiment a SYBR green-based One-Step qRT-PCR kit(Invitrogen Carlsbad CA USA) was used The thermal cyclingconditions were 50 C for 1 h followed by initial denaturation at 95C for 10 min and then 40 cycles of denaturation at 95 C for 30 sannealing at 55ndash65 C for 1 min and extension at 72 C for 1 min ina thermal cycler (Mx3000P Stratagene) Dissociation curve analysiswas performed by fluorescence reading at one-degree intervalsbetween 55ndash95 C to ensure that only one PCR product wasamplified All experiments were performed in two biological repli-cates and three technical replicates The expression data werenormalized to EF-1a using the DDCT method described by Livakand Schmittgen (2001)
Preparation of enzyme samples
To understand the role of major defence response enzymes andphenolic compounds in transgenic rice lines containing the singlerice blast resistance gene Pi54 enzymatic analysis was alsoperformed Challenge-inoculated leaf samples along with non-inoculated controls of transgenic and non-transgenic lines wereharvested after 0 24 48 72 96 and 120 hpi Total crude enzymeswere extracted from 1 g of leaf samples in a prechilled mortar andpestle and homogenized in their respective buffers (wv 11) suchas potassium phosphate buffer (10 mM pH 60) for POX(peroxidase) (18 000 g 15 min) and PPO (polyphenol oxidase)(12 000 g 20 min) sodium acetate buffer (005 M pH 46 23 000g 20 min) for laminarinase chitinase and b-glucosidase andborate buffer (25 mM pH 88) for PAL (8000 g 30 min) Thehomogenate was centrifuged at 4 C at specific speeds (given abovein parentheses) for the separate preparation of crude enzymeextract The supernatant was used as the enzyme source in all casesand for spectrophotometric estimation of enzymatic activity(Sadasivam and Manickam 2008) Spectrophotometric observa-tions were performed by Spectrophotometer GeneQuant 1300 (GEHealthcare Bio-sciences AB Uppsala Sweden) The proteincontents of the crude extract were determined by a standardmethod (Bradford 1976) using BSA (Sigma-Aldrich Chemical CoSt Louis MO USA) as the standard
Enzymatic assay
For the POX assay a reaction mixture (313 ml) was prepared byadding 3 ml of 01 M phosphate buffer (pH 70) 50 ll of 20 mMguaicol solution 30 ll of 123 mM hydrogen peroxide (014 ml of30 H2O2 to 100 ml diluted with distilled water prepared freshly)and 50 ll of enzyme extract The reaction was allowed to proceedfor 3 min A436 values were measured 30 s after adding the enzymeextract to the substrate and the change in absorbance wasrecorded
The reaction mixture for the PPO assay consisted of 15 ml of01 M sodium phosphate buffer (pH 65) and 200 ll of the enzymeextract The reaction was started with the addition of 200 ll of 10mM catechol (Mayer et al 1965) The rate of increase inabsorbance was measured at 420 nm for 1 min The activity wasexpressed as change in A436 mg1 protein min1
The PAL assay was performed using a modified procedure ofLisker et al (1983) by measuring the production of trans-cinnamicacid from L-phenylalanine The reaction mixture contained 1 ml ofenzyme extract 05 ml substrate of 50 mm L-phenylalanine and04 ml of 25 mM borate buffer (pH 88) After incubation for 2 hat 40 C the reaction was stopped by the addition of 006 ml 5 NHCl and absorbance was recorded at A290 and read against thesame volume of reaction mixture without L-phenylalanine whichserved as a blank The enzyme activity was expressed as l mol oftrans-cinnamic acid (t-CA) mg1 protein h1
For the beta-glucosidase assay enzyme activity was determinedby the modified method of Agrawal and Bahal (1969) A reactionmixture was prepared by adding 100 ll of a 25 mM solution ofp-nitrophenyl-b-D-glycopyaranoside to 350 ll of 005 M sodiumacetate buffer (pH 46) followed by initial incubation at 30 C for5 min After the addition of 50 ll of enzyme extract the mixturewas further incubated at 30 C for 15 min The reaction wasstopped by adding 700 ll of 02 M sodium carbonate The yellowcolour formed was measured at 420 nm by a spectrophotometerEnzyme activity was calculated as lmol of p-nitrophenol releasedmg1 of protein extract min1
For the b-13-glucanase assay enzyme activity was determinedby the modified method of Abeles and Forrence (1970) Thereaction mixture was prepared by adding 75 ll of enzyme extractto 175 ll of 2 (wv) Laminarin (Sigma-Aldrich USA) in waterand incubated for 60 min at 50 C in a water bath The reactionwas stopped by adding 750 ll of dinitrosalicylic acid reagent(Sigma-Aldrich USA) The tubes were then cooled to 25 C
Pi54 mediated resistance against rice blast | 759D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Whole contents were diluted 110 with distilled water Sampleswere vortexed and the A500 values recorded Glucose was used asa standard (Miller 1959) Enzyme activity was calculated as lmolof glucose released min1 mg1 protein extract
Chitinase activity was determined by quantitative estimation ofthe reducing sugars produced with colloidal chitin as the appropri-ate substrate (Sun et al 2006) Briefly the reaction mixtureconsisted of 05 ml enzyme solution and 05 ml of 1 colloidalchitin in 1 ml of 005 M sodium acetate buffer (pH 68) Themixture was incubated for 2 h at 40 C for chitinase and thereaction was stopped in boiling water for 5 min The amount ofreducing sugars released in the supernatant was measured bya method that uses dinitrosalicylic (DNS) acid reagent Theabsorbance was measured at 530 nm by a spectrophotometer Oneunit (U) of chitinase activity was defined as the amount of enzymethat liberated 1 lmol of the reducing sugar(s) min1 from N-acetylglucosamine under standard conditions
For the enzyme analysis all experiments were performed intriplicate The data obtained for each enzyme were analysedseparately Paired t test was performed in all the cases usingSigmaXL software (SigmaXL Toronto Canada) and differencesat Pfrac14005 were considered to be significant
Estimation of total phenol content
The total phenol contents in the pathogen inoculated leaf samplesof transgenic and non-transgenic rice at various intervals wereestimated following the modified procedure of Singleton and Rossi(1965) Rice leaves (1 g) were homogenized in 10 ml of 80methanol and agitated for 15 min at 70 C The homogenates werecentrifuged at 5000 g for 10 min An aliquot of 1 ml of methanolextract was made up to 5 ml with distilled water to which FolinndashCiocalteu reagent (250 ll) was added the solution was then kept at25 C After 3 min 1 ml saturated solution of Na2CO3 and 1 mldistilled water were added after which the reaction mixture wasincubated for 30 min at 25 C The A765 value of the developedblue colour was measured using a spectrophotometer (GeneQuant1300 GE Healthcare Bio-sciences AB Uppsala Sweden)A standard curve was prepared using gallic acid (Sigma-AldrichChemical Co USA) The phenol content of the extract wasexpressed as lg phenol equivalent released g1 of leaf tissue
Results
In order to observe the effect of the M oryzae-inducedexpression level of the Pi54 gene the transcriptomes of the
blast-resistant transgenic line TP-Pi54 and blast-susceptible
native TP lines were analysed at 72 hpi It has already been
shown that plants of the transgenic line TP-Pi54 have single
Pi54 gene insertions that are stably inherited up to three
generations (Rai et al 2011) The transgenic lines continu-
ally show a high degree of resistance and hypersensitive
response to the blast pathogen in the fifth generation as well(Fig 1)
Genome-wide expression analysis
For expression data analysis logarithmic (Log10) expression
values were normalized using the Robust Multichip Average(RMA) algorithm Further adopting a stringent criterion for
the expression fold change value (FCA) of gt20 and the
P value lt005 a total of 1154 differentially expressed genes
were identified in TP-Pi54 plants Of these 587 were up-
regulated whereas 567 genes were found to be down-regulated
(Table 1) However in the case of TP (susceptible) lines
a total of 1056 genes were found to be differentially regulated
of which 518 were up-regulated and 538 down-regulated A
total of 480 up- and 509 down-regulated genes were found to
be commonly expressed in both the TP-Pi54 and TP
(susceptible control) lines However 107 genes were found to
be exclusively up-regulated and 58 genes exclusively down-
regulated in the case of TP-Pi54 (Fig 2) Similarly in the TPline 38 genes were up-regulated and 29 down-regulated
Functional analysis of differentially regulated genes ofTP-Pi54
All the genes that showed differential expression in the
transgenic blast-resistant line TP-Pi54 were clustered and
functionally annotated The level of expression of these
genes in TP-Pi54 plants was compared with that in TP
plants Based on K-means clustering the functionally
annotated genes were classified into eight different clusters
Fig 1 Typical blast lesions as observed in the non-transgenic
susceptible rice line Taipei 309 (TP) No such disease symptoms
were observed in the resistant transgenic line (TP-Pi54) hypersen-
sitive response (HR) was observed in the TP-Pi54 line
Table 1 Gene ontology score-based categorization of differen-
tially regulated genes
Plants Up-regulated genes Down-regulated genes
TP-Pi54 (resistant) 587 [BPa 216 (1871)b 567 [BP 276 (2391)
CC 168 (1455) CC 203 (1759)
MF 274 (2374)] MF 322 (2790)]
TP (susceptible) 518 [BP 189 (1789) 538 [BP 212 (2007)
CC 150 (1420) CC 133 (1259)
MF 236 (2234)] MF 264 (25)]
a BP Biological process CC cellular component MF molecularfunction
b Numbers in parentheses represent the percentage in each set outof total differentially regulated genes of the plant
760 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
These genes were also analysed on the basis of their Gene
Ontology (GO) score into three main GO categories
including biological process (BP) molecular function (MF)
and cellular component (CC) (Table 1)
Similarly differentially regulated genes were assigned into
16 GO subcategories The percentages of up- and down-
regulated genes belonging to each category are given in Fig 3
Most (17) genes belonging to the binding-type category werefound to be up-regulated while 138 were down-regulated
Similarly 1534 of cell-part genes (GO ID004464) were up-
regulated whereas an almost equal number (1518) was
down-regulated in the transgenic line Interestingly the
number (1679) of metabolic process genes that were down-
regulated was higher than that of up-regulated (1349) ones
in this important category Likewise there were higher
numbers of down-regulated genes compared with up-regulatedones in a few other GO subcategories such as cellular
processes organelle and macromolecular complex On the
contrary 1247 462 and 341 of the genes belonging to
catalytic activity biological regulation and establishment of
localization were up-regulated respectively In some of the
subcategories such as transporter activity transcription
regulator molecular transducer and anatomical structure and
function the genes were only up-regulated (Fig 3)
Annotation information on differentially expressed genes
that were more up-regulated in TP-Pi54 was collected for
each probe set ID from GO analysis NCBI-BLAST and
NetAffx These genes were again categorized into various
functional groups A multitude of genes known for their
involvement in some way or another in stress responses (see
Supplementary Table S1 at JXB online) transcriptional
regulation (see Supplementary Table S2 at JXB online)programmed cell death (see Supplementary Table S3 at JXB
online) and signal transduction (see Supplementary Table
S4 at JXB online) were observed to be expressed differen-
tially in the transgenic line TP-Pi54 The level of expression
of these genes in the TP-Pi54 and TP lines varied from
207ndash1238-fold (see Supplementary Table S2 at JXB
online)
Functional annotation of stress response genes
Stress-response genes were specifically grouped together
according to the pathways in which they are involved Their
FCA values were represented as the average of the FCA
values of the members of each group and compared with
those of the TP (susceptible) lines (Fig 4A) Two genes for
callose biosynthesis were up-regulated 475-fold and 383-fold
in TP-Pi54 while in the TP lines these genes were up-
regulated only 2-fold following pathogen inoculation (seeSupplementary Table S1 at JXB online) One of the three
laccase enzyme-related genes was highly (31-fold) up-
regulated Similarly genes for other enzymatic categories like
phenyl ammonia lyase peroxidases lipoxygenases superoxide
dimustase amylase licheninase chitinase and glucosylcerami-
dase were up-regulated from 10-fold to 6391-fold in TP-Pi54
compared with the TP lines (see Supplementary Table S1
at JXB online) Interestingly various genes earlier reportedto be involved in other stresses such as wounding and abiotic
stresses were also more up-regulated in transgenic lines
Very high up-regulation of genes like laccase (729-fold
Fig 2 Venn diagram of genome-wide differentially expressed genes
in TP and TP-Pi54 following M oryzae inoculation Numbers in
parentheses indicate down-regulated genes U upregulated genes
Fig 3 Functional categorization of genes differentially expressed in the TP-Pi54 line The number of genes was finalized using the
filtering criteria of fold change gt20 and P-value correction ltfrac14005 by FDR (Benjamini-Hochberg) Genes were classified into 16 different
categories based on Gene Ontology The number of up- and down-regulated genes for each functional category is shown in the
histogram The functional categories are A binding B catalytic activity C transporter activity D transcription regulator activity E
molecular transducer activity F antioxidant activity G metabolic processes H cellular processes I biological regulation J
establishment of localization K response to stimulus L anatomical structure function M cell part N organelle O organelle part and
P molecular complex (A) Up-regulated genes (B) Down-regulated genes
Pi54 mediated resistance against rice blast | 761D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Os11g48060) peroxidases (1694-fold Os07g48010) ammonia
lyases (278-fold Os04g43760) and amylases (6391-fold
Os10g41550) were obtained (see Supplementary Table S1 at
JXB online) Similarly maximum up-regulation (1694ndash2545-
fold) of the genes for chemical stimulus was obtained with
a response to oxidative stress Several genes for transcription
factors (TF) were up-regulated in response to M oryzae
inoculation (see Supplementary Table S2 at JXB online)
Among the more prominently expressed are genes for NAC
domain-containing proteins (841-fold) Dof zinc finger pro-
teins (1134-fold) sigma factors (1238-fold) bZIP transcrip-
tion factors (1193-fold) and MADS box proteins (446-fold)
The up-regulation of the rest of the TF genes in transgenic
plants varied from 203-fold to 1238-fold It was interesting to
note that all these TF genes were regulated at a very low level
in the non-transgenic plants (see Supplementary Table S2 at
JXB online Fig 4B) Programmed cell death (PCD) is one of
the important responses of plants upon pathogen infection
and leads to a hypersensitive response in resistant lines In this
study four genes with predicted roles in PCD were up-
regulated up to 207ndash276-fold in transgenic lines compared
with between 067ndash159-fold in non-transgenic lines (see
Supplementary Table S3 at JXB online Fig 4B Fig 4C)
Fifteen genes associated with vital signal transduction
functions in the plants were also highly up-regulated in TP-
Pi54 (see Supplementary Table S4 at JXB online Fig 4B)
A maximum up-regulation of 1202-fold was obtained in
auxin-mediated signal pathways followed by that ingibberellic acid-mediated signalling (328-fold) Similarly
genes for two-component signal transduction pathways
were up-regulated between 244-fold and 356-fold (see
Supplementary Table S4 at JXB online Fig 4D)
Hierarchical clustering
Genome-wide analysis of the expression patterns of mock-
inoculated and pathogen-inoculated TP-Pi54 transgenicplants was performed The phylogenetic relationship be-
tween highly expressed and less expressed genes was
obtained after hierarchal clustering The expression changes
among the gene group members in six clusters (i) response
to stimulus (ii) signal transduction (iii) transcription
Fig 4 Comparison of the differential regulation of different categories of genes more up-regulated in TP-Pi54 compared to TP
(A) Stress-responsive genes 1 callose 2 laccase 3 PAL 4 Peroxidase 5 lipoxygenase 6 superoxide dismutase 7 amylase 8
licheninase 9 chitinase 10 glucosylceramidase 11 response to wounding 12 response to chemical stimulus and 13 abiotic stress
(B) 1ndash22 Transcription factor genes (C) 1ndash4 Genes for programmed cell death (D) 1ndash15 Genes for signal transduction
762 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
regulators (iv) programmed cell death (v) oxidoreductases
and (vi) lyase were represented by different colours in the
heat map The colour intensities indicate the expression
level of each gene (Fig 5)
Genes repressed in native TP lines
Out of the 107 up-regulated genes of TP-Pi54 29 were
considerably repressed in the non-transgenic TP line (Table 2)
Among these the genes Os77671S1_at (652-fold) andOs70041S1_at (2458-fold) were up-regulated at the highest
level in TP-Pi54 compared with non-transgenic lines Other
highly expressed genes in TP-Pi54 that were otherwise
repressed in TP belonged to some important categories such
as genes involved in transporter activity (2726-fold and
1692-fold) zinc ion binding (1033-fold) and lipid transport
(685-fold) Interestingly a mitochondrial located gene
(Os124301S1_at) was also highly up-regulated (15-fold) inTP-Pi54 and down-regulated in the TP lines (Table 2) Among
all the up-regulated genes in TP-54 22 were hypothetically
expressed proteins of unknown function This indicates that
these genes may be novel and may have an important role in
the Pi54-mediated defence mechanism in the blast-resistant
transgenic line TP-Pi54
Quantitative analysis of differentially regulated genes
Validation of nine up-regulated genes and one down-regulated gene in the transgenic lines was performed by
quantitative real-time PCR Genes for NAC domain-
containing proteins (NAC6) callose polysaccharide
biosynthesis proteins (CAPD) Dof zinc finger proteins
(DOF) peroxidase (POC1) MADS box proteins (MADS)
class III peroxidase (POX3) chitinase (CHIT) phenylala-
nine ammonia lyase (PAL) and endo-13-14-b-glucanase
(BG) were found to be up-regulated 124ndash58-fold whereas
genes for homeo domain-related proteins (HDR) were
down-regulated up to 14-fold (Fig 6 see Supplementary
Table S6 at JXB online Fig 4B)
Biochemical assay of enzymes involved in defenceresponse
To assess the effect of pathogen infection on plant defence
response enzymes the activities of different enzymes were
measured individually in transgenic (TP-Pi54) and non-
transgenic (TP) plants at 0 to 120 hpi (Fig 7) The data
obtained from transgenic plants were compared with those
from the non-inoculated plants used as controls M oryzae
inoculation drastically increased peroxidase activity intransgenic rice plants after 96 hpi The POX activity at 96
hpi in TP-Pi54 was higher (01623 U) than that in TP
(Fig 7A Table 3) The changes in polyphenol oxidase
activity in the plants followed more or less the same pattern
as observed for POX The PPO activity increased at 24 hpi
in transgenic plants reached its maximum at 96 hpi and
declined thereafter The PPO activity was measured as
change in A420 (DA420) mg1 protein min1 The PPOactivity at 96 h was 01736 U in TP-Pi54 compared with
01 U in TP (Fig 7B Table 3)
The time-course of PAL activity in treated plants was
quite distinct from that of the rest of the enzymes as the
profile represented a bimodal pattern in transgenic plants
indicating an early and sharp induction at 24 hpi (Fig 7C)
The decline in PAL activity continued until 72 hpi and
increased thereafter until 96 hpi before following another
dip at 120 hpi compared with that in non-inoculated
transgenic control plants which showed no induction at all
The maximum PAL activity at 24 hpi was 9305 U in TP-
Pi54 and 1768 U in TP (Fig 7C Table 3) b-Glucosidase
activity in plants gradually increased upon pathogen in-
oculation reaching its maximum at 48 hpi in the transgenic
group On the other hand a bimodal pattern of activity was
obtained in the case of non-transgenic plants showing
a slight increase in the activity at 24 and 96 hpi The activity
of b-glucosidase at 48 h was 00809 U and 00214 U in TP-
Pi54 and TP respectively (Fig 7D Table 3)
The activity of b-13-glucanase in transgenic plantswhich was already high started increasing after 24 hpi and
thereafter increased consistently up to 120 hpi Although
the b-13-glucanase activity was also higher in the trans-
genic control it did not show any significant increase after
24 hpi However in non-transgenic plants the enzyme was
slightly induced at 24 h but declined immediately until 72
hpi before reaching a normal level The highest b-1
3-glucanase activity was 8770 U in TP-Pi54 compared to727 U in TP (Fig 7E Table 3) The chitinase enzyme
concentration increased in transgenic plants at 48 hpi and
reached its maximum level after 120 hpi The maximum
chitinase activity was 1011 U in TP-Pi54 compared with
858 U in TP (Fig 7F Table 3)
Fig 5 Expression pattern as represented by the heat map profile
of mock-inoculated (control) versus TP-Pi54 (test) rice lines
inoculated with M oryzae Colour changes show the level of gene
expression in their respective clusters Yellow mildly expressed
genes blue highly suppressed genes red highly expressed
genes The clusters are genes for (A) response to stimulus (B)
signal transduction (C) transcription regulators (D) programmed
cell death (E) oxidoreductases and (F) lyase
Pi54 mediated resistance against rice blast | 763D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

et al 1974) These losses are even higher in East Asia and
other more temperate rice-growing regions around the
world (Talbot 2003) Rice blast can be effectively managed
by using resistant rice varieties for which the identification
and characterization of appropriate blast resistance (R)
genes are vital To date more than 19 blast resistance genes
have been cloned and characterized (Sharma et al 2005b
Okuyama et al 2011) However none of them has beenshown to have resistance potential against all the strains of
M oryzae Recently it was reported in a separate study that
Pi54 (Pi-k-h cloned from the rice line Tetep) confers broad
spectrum resistance against geographically diverse strains of
M oryzae collected from various parts of India and the US
(Costanzo and Jia 2010 Rai et al 2011)
Several attempts to utilize R-genes in rice blast by de-
veloping disease-resistant cultivars have been futile owing tothe lack of information regarding the intricate mechanisms
involved in R-gene-mediated resistance pathways and the
high rate of variability in the corresponding avirulence (Avr)
genes Therefore in order to develop sustainably successful
blast-resistant rice lines a comprehensive dissection of the
reactions downstream of RndashAvr interactions is highly desir-
able The co-expression patterns of bacterial disease resistance
genes and their transcriptional regulators in transgenic ricehave indicated that the resistance genes trigger an immune
response (Sana et al 2010 Zhou et al 2010) The expression
patterns of defence response genes involved in ricendashM oryzae
interaction have been widely studied (Kim et al 2001 Xiong
et al 2001 Rao et al 2002 Jantasuriyarat et al 2005
Soderlund et al 2006 Vergne et al 2008 Kato et al 2009)
However an expression study in response to the blast
pathogen in transgenic rice containing a single gene for blastresistance still remains to be performed Further downstream
biochemical events resulting from gene-for-gene interactions
have only been studied in blast-resistant plants (Hammond-
Kosack and Jones 2000 Dangl et al 1996 Ryals et al
1996) and the impact of rice blast R-genes in the transgenic
system remains unexplored
Previously the rice blast resistance gene Pi-kh has been
isolated from the indica rice line Tetep showing resistance todifferent M oryzae strains in the North-Western Himalayan
region of India (Sharma et al 2005a) This gene is only the
third to be cloned in the series of rice blast resistance genes
after the cloning of Pib (Wang et al 1999) and Pita (Bryan
et al 2000) The gene was renamed Pi54 after it was further
re-alienated to a slightly distant location from the Pik locus
(Sharma et al 2010) The predicted Pi54 protein contains
a nucleotide-binding sitendashleucine-rich repeat (NBS-LRR)domain in addition to a small zinc finger domain (Sharma
et al 2005a b) Functional complementation of the gene
has confirmed its requirement for conferring a stable and
high level of resistance against geographically diverse strains
of M oryzae from different parts of India (Rai et al 2011)
Also this gene is widely being used in a pyramiding
programme for broad spectrum resistance to blast in
combination with Piz5 in Pusa Basmati rice varieties (Singhet al 2011) The gene has been shown to induce the
synthesis of callose (b-13-glucan) in response to pathogen
challenge The Pi54 gene is expressed constitutively at
a basal level in both resistant transgenic as well as
susceptible native lines up to 48 hpi Similar to the Pib
(Wang et al 1999) and Pi5-1 (Lee et al 2009) blast
resistance genes of rice the expression of Pi54 is induced by
M oryzae infection and a more than 2-fold higher in-
duction was observed at 72 hpi in the resistant line (Rai
et al 2011) Further studies of the expression profiles ofblast-resistant and susceptible lines at 72 hpi have yet to be
completed using microarray analysis
In this study our objectives were (i) to investigate and
compare using microarray-based transcriptome analysis the
global gene expression changes happening in response to
infection by M oryzae in transgenic rice lines containing the
rice blast resistance gene Pi54 (ii) to identify and validate the
potential genes involved in Pi54-mediated blast resistanceand (iii) to perform biochemical analysis of the enzymes and
phenolic compounds involved in the disease response path-
ways during compatible and incompatible interactions Our
results showed a Pi54-mediated reprogramming of a broad
array of genes which apparently leads to an M oryzae-
resistant phenotype via detectable changes in the activities of
the enzymes involved in the defence pathways
Materials and methods
Plant and fungal materials
The blast-resistant transgenic rice line TP-Pi54-15 used in thisstudy was originally derived from the cloned gene Pi54 (Pi-kh)transferred into Oryza sativa cv Taipei (TP) 309 which is a highlysusceptible to the M oryzae strain PLP-1 The gene was stable andexpressed in subsequent generations (T3) and was being main-tained in our laboratory for subsequent use (Rai et al 2011)Transgenic rice lines were maintained in a containment facility atthe National Phytotron Facility (NPF) Indian Agricultural Re-search Institute New Delhi India
Fifteen-day-old seedlings of TP-Pi54-15 were inoculated with anM oryzae spore suspension containing 105 spores ml1 in 025gelatin until the leaves were covered with fine droplets For themock control plants were inoculated with 025 gelatin only Theexperiment was carried out under controlled growth conditions at2562 C and 90 relative humidity for 24 h in the dark and thenshifted to a 168 h lightdark regime After 72 hpi leaves werecollected washed and pooled for RNA extraction For bio-chemical analyses challenge-inoculated leaf samples along withnon-inoculated controls of transgenic (TP-Pi54) and non-transgenic (TP) lines were harvested after 0 24 48 72 96 and120 hpi for enzyme extraction
RNA isolation and microarray analysis
Total RNA was extracted from transgenic and non-transgenic ricelines using the TRI Reagent (Invitrogen) Plant tissues werehomogenized using a mortar and pestle with liquid nitrogen and1 ml of TRI Reagent was added per 100 mg of tissue Quantitativeand qualitative assessment of the isolated RNA was done usinga UV-Vis Spectrophotometer (NanoDrop Technologies WilmingtonDE) and by gel electrophoresis respectively RNA sampleswere processed according to the standard Affymetrix GeneChipexpression analysis technical manual (Affymetrix Inc Santa ClaraCA USA) Total RNA was used for the synthesis of cDNA usinga SuperScript double-stranded cDNA synthesis kit A portion of theresulting double-stranded cDNA was used as a template to generate
758 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
biotin-tagged cRNA from an in vitro transcription reaction using anAffymetrix GeneChip IVT labelling kit Fifteen micrograms ofbiotin-tagged cRNA was fragmented (50 to 200 bp fragments) as perthe manufacturerrsquos protocols Subsequently 10 lg of this fragmentedcRNA was hybridized to rice gene chips (Affymetrix) at 45 C with16 h rotation in a GeneChip hybridization oven (Affymetrix)Duplicate biological samples from transgenic and control rice plantswere used for microarray analysis Each array contained probe setsfor 51 279 rice transcripts out of which around 48 564 were fromjaponica and 1260 from indica cultivars Seventy-two hours aftertreatment total RNA was isolated from leaf tissues inoculated withthe pathogen The samples from 20ndash30 challenge-inoculated leaveswere pooled together Total RNA was converted into biotin-labelledcRNA and hybridized to the rice gene chip Separate gene chips werehybridized for the M oryzae spore suspension prepared in 02gelatin and leaf samples mock-inoculated with 02 gelatin onlywere used as the mock control GeneChip arrays were washed andthen stained with streptavidin-phycoerythrin (Affymetrix FluidicsStation 450) followed by scanning with a GeneChip scanner(Affymetrix GeneChip Scanner 3000) FCA (fold change absolute)values were calculated for each transcript of TP-Pi54 and TP usingmock-inoculated chip data as their respective controls The FCAvalues of transgenic and native lines for each transcript werecompared
Microarray data analysis
MIAME-compliant microarray data from this study have beendeposited at NCBI Gene Expression Omnibus with the accessionnumber GSE28308 The array data set was analysed using theGeneChip Operating Software (GCOS 12) and ArrayStar(DNASTAR Inc Madison USA) software For probe levelnormalization robust multi-array analysis (RMA) was performedon Affymetrix raw files (CEL files) (Irizarry et al 2003) Theinitial data set consisted of 57 382 probes Filtration wasperformed to remove probes with lsquoAbsent Callsrsquo (poor qualityprobes) Transformation and normalization were done in order tofacilitate comparison across samples The biological significance ofdifferentially expressed genes was determined by performingfunctional classification according to Gene Ontology after filteringthe genes for gt20-fold change and lt005 P-value Gene annota-tion was performed using BLAST (httpblastncbinlmnihgovBlast) All the raw data files (CEL files two for blast-inoculatedand two for mock treatment) obtained from the GeneChipOperating Software (GCOS Affymetrix) were subjected to furtheranalysis using the ArrayStar software
Quantitative real-time PCR analysis
Part of the samples of total RNA isolated for microarray analysiswas also used to validate the functionally important genes that werefound to be differentially regulated in the microarray results Thenucleotide sequences of differentially regulated genes were down-loaded from the TIGR rice database (httpriceplantbiologymsuedu) (see Supplementary Table S5 at JXB online) The exonicsequences of selected genes were used to design primers using thePRIMER3 software (httpfrodowimiteduprimer3) For the genevalidation experiment a SYBR green-based One-Step qRT-PCR kit(Invitrogen Carlsbad CA USA) was used The thermal cyclingconditions were 50 C for 1 h followed by initial denaturation at 95C for 10 min and then 40 cycles of denaturation at 95 C for 30 sannealing at 55ndash65 C for 1 min and extension at 72 C for 1 min ina thermal cycler (Mx3000P Stratagene) Dissociation curve analysiswas performed by fluorescence reading at one-degree intervalsbetween 55ndash95 C to ensure that only one PCR product wasamplified All experiments were performed in two biological repli-cates and three technical replicates The expression data werenormalized to EF-1a using the DDCT method described by Livakand Schmittgen (2001)
Preparation of enzyme samples
To understand the role of major defence response enzymes andphenolic compounds in transgenic rice lines containing the singlerice blast resistance gene Pi54 enzymatic analysis was alsoperformed Challenge-inoculated leaf samples along with non-inoculated controls of transgenic and non-transgenic lines wereharvested after 0 24 48 72 96 and 120 hpi Total crude enzymeswere extracted from 1 g of leaf samples in a prechilled mortar andpestle and homogenized in their respective buffers (wv 11) suchas potassium phosphate buffer (10 mM pH 60) for POX(peroxidase) (18 000 g 15 min) and PPO (polyphenol oxidase)(12 000 g 20 min) sodium acetate buffer (005 M pH 46 23 000g 20 min) for laminarinase chitinase and b-glucosidase andborate buffer (25 mM pH 88) for PAL (8000 g 30 min) Thehomogenate was centrifuged at 4 C at specific speeds (given abovein parentheses) for the separate preparation of crude enzymeextract The supernatant was used as the enzyme source in all casesand for spectrophotometric estimation of enzymatic activity(Sadasivam and Manickam 2008) Spectrophotometric observa-tions were performed by Spectrophotometer GeneQuant 1300 (GEHealthcare Bio-sciences AB Uppsala Sweden) The proteincontents of the crude extract were determined by a standardmethod (Bradford 1976) using BSA (Sigma-Aldrich Chemical CoSt Louis MO USA) as the standard
Enzymatic assay
For the POX assay a reaction mixture (313 ml) was prepared byadding 3 ml of 01 M phosphate buffer (pH 70) 50 ll of 20 mMguaicol solution 30 ll of 123 mM hydrogen peroxide (014 ml of30 H2O2 to 100 ml diluted with distilled water prepared freshly)and 50 ll of enzyme extract The reaction was allowed to proceedfor 3 min A436 values were measured 30 s after adding the enzymeextract to the substrate and the change in absorbance wasrecorded
The reaction mixture for the PPO assay consisted of 15 ml of01 M sodium phosphate buffer (pH 65) and 200 ll of the enzymeextract The reaction was started with the addition of 200 ll of 10mM catechol (Mayer et al 1965) The rate of increase inabsorbance was measured at 420 nm for 1 min The activity wasexpressed as change in A436 mg1 protein min1
The PAL assay was performed using a modified procedure ofLisker et al (1983) by measuring the production of trans-cinnamicacid from L-phenylalanine The reaction mixture contained 1 ml ofenzyme extract 05 ml substrate of 50 mm L-phenylalanine and04 ml of 25 mM borate buffer (pH 88) After incubation for 2 hat 40 C the reaction was stopped by the addition of 006 ml 5 NHCl and absorbance was recorded at A290 and read against thesame volume of reaction mixture without L-phenylalanine whichserved as a blank The enzyme activity was expressed as l mol oftrans-cinnamic acid (t-CA) mg1 protein h1
For the beta-glucosidase assay enzyme activity was determinedby the modified method of Agrawal and Bahal (1969) A reactionmixture was prepared by adding 100 ll of a 25 mM solution ofp-nitrophenyl-b-D-glycopyaranoside to 350 ll of 005 M sodiumacetate buffer (pH 46) followed by initial incubation at 30 C for5 min After the addition of 50 ll of enzyme extract the mixturewas further incubated at 30 C for 15 min The reaction wasstopped by adding 700 ll of 02 M sodium carbonate The yellowcolour formed was measured at 420 nm by a spectrophotometerEnzyme activity was calculated as lmol of p-nitrophenol releasedmg1 of protein extract min1
For the b-13-glucanase assay enzyme activity was determinedby the modified method of Abeles and Forrence (1970) Thereaction mixture was prepared by adding 75 ll of enzyme extractto 175 ll of 2 (wv) Laminarin (Sigma-Aldrich USA) in waterand incubated for 60 min at 50 C in a water bath The reactionwas stopped by adding 750 ll of dinitrosalicylic acid reagent(Sigma-Aldrich USA) The tubes were then cooled to 25 C
Pi54 mediated resistance against rice blast | 759D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Whole contents were diluted 110 with distilled water Sampleswere vortexed and the A500 values recorded Glucose was used asa standard (Miller 1959) Enzyme activity was calculated as lmolof glucose released min1 mg1 protein extract
Chitinase activity was determined by quantitative estimation ofthe reducing sugars produced with colloidal chitin as the appropri-ate substrate (Sun et al 2006) Briefly the reaction mixtureconsisted of 05 ml enzyme solution and 05 ml of 1 colloidalchitin in 1 ml of 005 M sodium acetate buffer (pH 68) Themixture was incubated for 2 h at 40 C for chitinase and thereaction was stopped in boiling water for 5 min The amount ofreducing sugars released in the supernatant was measured bya method that uses dinitrosalicylic (DNS) acid reagent Theabsorbance was measured at 530 nm by a spectrophotometer Oneunit (U) of chitinase activity was defined as the amount of enzymethat liberated 1 lmol of the reducing sugar(s) min1 from N-acetylglucosamine under standard conditions
For the enzyme analysis all experiments were performed intriplicate The data obtained for each enzyme were analysedseparately Paired t test was performed in all the cases usingSigmaXL software (SigmaXL Toronto Canada) and differencesat Pfrac14005 were considered to be significant
Estimation of total phenol content
The total phenol contents in the pathogen inoculated leaf samplesof transgenic and non-transgenic rice at various intervals wereestimated following the modified procedure of Singleton and Rossi(1965) Rice leaves (1 g) were homogenized in 10 ml of 80methanol and agitated for 15 min at 70 C The homogenates werecentrifuged at 5000 g for 10 min An aliquot of 1 ml of methanolextract was made up to 5 ml with distilled water to which FolinndashCiocalteu reagent (250 ll) was added the solution was then kept at25 C After 3 min 1 ml saturated solution of Na2CO3 and 1 mldistilled water were added after which the reaction mixture wasincubated for 30 min at 25 C The A765 value of the developedblue colour was measured using a spectrophotometer (GeneQuant1300 GE Healthcare Bio-sciences AB Uppsala Sweden)A standard curve was prepared using gallic acid (Sigma-AldrichChemical Co USA) The phenol content of the extract wasexpressed as lg phenol equivalent released g1 of leaf tissue
Results
In order to observe the effect of the M oryzae-inducedexpression level of the Pi54 gene the transcriptomes of the
blast-resistant transgenic line TP-Pi54 and blast-susceptible
native TP lines were analysed at 72 hpi It has already been
shown that plants of the transgenic line TP-Pi54 have single
Pi54 gene insertions that are stably inherited up to three
generations (Rai et al 2011) The transgenic lines continu-
ally show a high degree of resistance and hypersensitive
response to the blast pathogen in the fifth generation as well(Fig 1)
Genome-wide expression analysis
For expression data analysis logarithmic (Log10) expression
values were normalized using the Robust Multichip Average(RMA) algorithm Further adopting a stringent criterion for
the expression fold change value (FCA) of gt20 and the
P value lt005 a total of 1154 differentially expressed genes
were identified in TP-Pi54 plants Of these 587 were up-
regulated whereas 567 genes were found to be down-regulated
(Table 1) However in the case of TP (susceptible) lines
a total of 1056 genes were found to be differentially regulated
of which 518 were up-regulated and 538 down-regulated A
total of 480 up- and 509 down-regulated genes were found to
be commonly expressed in both the TP-Pi54 and TP
(susceptible control) lines However 107 genes were found to
be exclusively up-regulated and 58 genes exclusively down-
regulated in the case of TP-Pi54 (Fig 2) Similarly in the TPline 38 genes were up-regulated and 29 down-regulated
Functional analysis of differentially regulated genes ofTP-Pi54
All the genes that showed differential expression in the
transgenic blast-resistant line TP-Pi54 were clustered and
functionally annotated The level of expression of these
genes in TP-Pi54 plants was compared with that in TP
plants Based on K-means clustering the functionally
annotated genes were classified into eight different clusters
Fig 1 Typical blast lesions as observed in the non-transgenic
susceptible rice line Taipei 309 (TP) No such disease symptoms
were observed in the resistant transgenic line (TP-Pi54) hypersen-
sitive response (HR) was observed in the TP-Pi54 line
Table 1 Gene ontology score-based categorization of differen-
tially regulated genes
Plants Up-regulated genes Down-regulated genes
TP-Pi54 (resistant) 587 [BPa 216 (1871)b 567 [BP 276 (2391)
CC 168 (1455) CC 203 (1759)
MF 274 (2374)] MF 322 (2790)]
TP (susceptible) 518 [BP 189 (1789) 538 [BP 212 (2007)
CC 150 (1420) CC 133 (1259)
MF 236 (2234)] MF 264 (25)]
a BP Biological process CC cellular component MF molecularfunction
b Numbers in parentheses represent the percentage in each set outof total differentially regulated genes of the plant
760 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
These genes were also analysed on the basis of their Gene
Ontology (GO) score into three main GO categories
including biological process (BP) molecular function (MF)
and cellular component (CC) (Table 1)
Similarly differentially regulated genes were assigned into
16 GO subcategories The percentages of up- and down-
regulated genes belonging to each category are given in Fig 3
Most (17) genes belonging to the binding-type category werefound to be up-regulated while 138 were down-regulated
Similarly 1534 of cell-part genes (GO ID004464) were up-
regulated whereas an almost equal number (1518) was
down-regulated in the transgenic line Interestingly the
number (1679) of metabolic process genes that were down-
regulated was higher than that of up-regulated (1349) ones
in this important category Likewise there were higher
numbers of down-regulated genes compared with up-regulatedones in a few other GO subcategories such as cellular
processes organelle and macromolecular complex On the
contrary 1247 462 and 341 of the genes belonging to
catalytic activity biological regulation and establishment of
localization were up-regulated respectively In some of the
subcategories such as transporter activity transcription
regulator molecular transducer and anatomical structure and
function the genes were only up-regulated (Fig 3)
Annotation information on differentially expressed genes
that were more up-regulated in TP-Pi54 was collected for
each probe set ID from GO analysis NCBI-BLAST and
NetAffx These genes were again categorized into various
functional groups A multitude of genes known for their
involvement in some way or another in stress responses (see
Supplementary Table S1 at JXB online) transcriptional
regulation (see Supplementary Table S2 at JXB online)programmed cell death (see Supplementary Table S3 at JXB
online) and signal transduction (see Supplementary Table
S4 at JXB online) were observed to be expressed differen-
tially in the transgenic line TP-Pi54 The level of expression
of these genes in the TP-Pi54 and TP lines varied from
207ndash1238-fold (see Supplementary Table S2 at JXB
online)
Functional annotation of stress response genes
Stress-response genes were specifically grouped together
according to the pathways in which they are involved Their
FCA values were represented as the average of the FCA
values of the members of each group and compared with
those of the TP (susceptible) lines (Fig 4A) Two genes for
callose biosynthesis were up-regulated 475-fold and 383-fold
in TP-Pi54 while in the TP lines these genes were up-
regulated only 2-fold following pathogen inoculation (seeSupplementary Table S1 at JXB online) One of the three
laccase enzyme-related genes was highly (31-fold) up-
regulated Similarly genes for other enzymatic categories like
phenyl ammonia lyase peroxidases lipoxygenases superoxide
dimustase amylase licheninase chitinase and glucosylcerami-
dase were up-regulated from 10-fold to 6391-fold in TP-Pi54
compared with the TP lines (see Supplementary Table S1
at JXB online) Interestingly various genes earlier reportedto be involved in other stresses such as wounding and abiotic
stresses were also more up-regulated in transgenic lines
Very high up-regulation of genes like laccase (729-fold
Fig 2 Venn diagram of genome-wide differentially expressed genes
in TP and TP-Pi54 following M oryzae inoculation Numbers in
parentheses indicate down-regulated genes U upregulated genes
Fig 3 Functional categorization of genes differentially expressed in the TP-Pi54 line The number of genes was finalized using the
filtering criteria of fold change gt20 and P-value correction ltfrac14005 by FDR (Benjamini-Hochberg) Genes were classified into 16 different
categories based on Gene Ontology The number of up- and down-regulated genes for each functional category is shown in the
histogram The functional categories are A binding B catalytic activity C transporter activity D transcription regulator activity E
molecular transducer activity F antioxidant activity G metabolic processes H cellular processes I biological regulation J
establishment of localization K response to stimulus L anatomical structure function M cell part N organelle O organelle part and
P molecular complex (A) Up-regulated genes (B) Down-regulated genes
Pi54 mediated resistance against rice blast | 761D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Os11g48060) peroxidases (1694-fold Os07g48010) ammonia
lyases (278-fold Os04g43760) and amylases (6391-fold
Os10g41550) were obtained (see Supplementary Table S1 at
JXB online) Similarly maximum up-regulation (1694ndash2545-
fold) of the genes for chemical stimulus was obtained with
a response to oxidative stress Several genes for transcription
factors (TF) were up-regulated in response to M oryzae
inoculation (see Supplementary Table S2 at JXB online)
Among the more prominently expressed are genes for NAC
domain-containing proteins (841-fold) Dof zinc finger pro-
teins (1134-fold) sigma factors (1238-fold) bZIP transcrip-
tion factors (1193-fold) and MADS box proteins (446-fold)
The up-regulation of the rest of the TF genes in transgenic
plants varied from 203-fold to 1238-fold It was interesting to
note that all these TF genes were regulated at a very low level
in the non-transgenic plants (see Supplementary Table S2 at
JXB online Fig 4B) Programmed cell death (PCD) is one of
the important responses of plants upon pathogen infection
and leads to a hypersensitive response in resistant lines In this
study four genes with predicted roles in PCD were up-
regulated up to 207ndash276-fold in transgenic lines compared
with between 067ndash159-fold in non-transgenic lines (see
Supplementary Table S3 at JXB online Fig 4B Fig 4C)
Fifteen genes associated with vital signal transduction
functions in the plants were also highly up-regulated in TP-
Pi54 (see Supplementary Table S4 at JXB online Fig 4B)
A maximum up-regulation of 1202-fold was obtained in
auxin-mediated signal pathways followed by that ingibberellic acid-mediated signalling (328-fold) Similarly
genes for two-component signal transduction pathways
were up-regulated between 244-fold and 356-fold (see
Supplementary Table S4 at JXB online Fig 4D)
Hierarchical clustering
Genome-wide analysis of the expression patterns of mock-
inoculated and pathogen-inoculated TP-Pi54 transgenicplants was performed The phylogenetic relationship be-
tween highly expressed and less expressed genes was
obtained after hierarchal clustering The expression changes
among the gene group members in six clusters (i) response
to stimulus (ii) signal transduction (iii) transcription
Fig 4 Comparison of the differential regulation of different categories of genes more up-regulated in TP-Pi54 compared to TP
(A) Stress-responsive genes 1 callose 2 laccase 3 PAL 4 Peroxidase 5 lipoxygenase 6 superoxide dismutase 7 amylase 8
licheninase 9 chitinase 10 glucosylceramidase 11 response to wounding 12 response to chemical stimulus and 13 abiotic stress
(B) 1ndash22 Transcription factor genes (C) 1ndash4 Genes for programmed cell death (D) 1ndash15 Genes for signal transduction
762 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
regulators (iv) programmed cell death (v) oxidoreductases
and (vi) lyase were represented by different colours in the
heat map The colour intensities indicate the expression
level of each gene (Fig 5)
Genes repressed in native TP lines
Out of the 107 up-regulated genes of TP-Pi54 29 were
considerably repressed in the non-transgenic TP line (Table 2)
Among these the genes Os77671S1_at (652-fold) andOs70041S1_at (2458-fold) were up-regulated at the highest
level in TP-Pi54 compared with non-transgenic lines Other
highly expressed genes in TP-Pi54 that were otherwise
repressed in TP belonged to some important categories such
as genes involved in transporter activity (2726-fold and
1692-fold) zinc ion binding (1033-fold) and lipid transport
(685-fold) Interestingly a mitochondrial located gene
(Os124301S1_at) was also highly up-regulated (15-fold) inTP-Pi54 and down-regulated in the TP lines (Table 2) Among
all the up-regulated genes in TP-54 22 were hypothetically
expressed proteins of unknown function This indicates that
these genes may be novel and may have an important role in
the Pi54-mediated defence mechanism in the blast-resistant
transgenic line TP-Pi54
Quantitative analysis of differentially regulated genes
Validation of nine up-regulated genes and one down-regulated gene in the transgenic lines was performed by
quantitative real-time PCR Genes for NAC domain-
containing proteins (NAC6) callose polysaccharide
biosynthesis proteins (CAPD) Dof zinc finger proteins
(DOF) peroxidase (POC1) MADS box proteins (MADS)
class III peroxidase (POX3) chitinase (CHIT) phenylala-
nine ammonia lyase (PAL) and endo-13-14-b-glucanase
(BG) were found to be up-regulated 124ndash58-fold whereas
genes for homeo domain-related proteins (HDR) were
down-regulated up to 14-fold (Fig 6 see Supplementary
Table S6 at JXB online Fig 4B)
Biochemical assay of enzymes involved in defenceresponse
To assess the effect of pathogen infection on plant defence
response enzymes the activities of different enzymes were
measured individually in transgenic (TP-Pi54) and non-
transgenic (TP) plants at 0 to 120 hpi (Fig 7) The data
obtained from transgenic plants were compared with those
from the non-inoculated plants used as controls M oryzae
inoculation drastically increased peroxidase activity intransgenic rice plants after 96 hpi The POX activity at 96
hpi in TP-Pi54 was higher (01623 U) than that in TP
(Fig 7A Table 3) The changes in polyphenol oxidase
activity in the plants followed more or less the same pattern
as observed for POX The PPO activity increased at 24 hpi
in transgenic plants reached its maximum at 96 hpi and
declined thereafter The PPO activity was measured as
change in A420 (DA420) mg1 protein min1 The PPOactivity at 96 h was 01736 U in TP-Pi54 compared with
01 U in TP (Fig 7B Table 3)
The time-course of PAL activity in treated plants was
quite distinct from that of the rest of the enzymes as the
profile represented a bimodal pattern in transgenic plants
indicating an early and sharp induction at 24 hpi (Fig 7C)
The decline in PAL activity continued until 72 hpi and
increased thereafter until 96 hpi before following another
dip at 120 hpi compared with that in non-inoculated
transgenic control plants which showed no induction at all
The maximum PAL activity at 24 hpi was 9305 U in TP-
Pi54 and 1768 U in TP (Fig 7C Table 3) b-Glucosidase
activity in plants gradually increased upon pathogen in-
oculation reaching its maximum at 48 hpi in the transgenic
group On the other hand a bimodal pattern of activity was
obtained in the case of non-transgenic plants showing
a slight increase in the activity at 24 and 96 hpi The activity
of b-glucosidase at 48 h was 00809 U and 00214 U in TP-
Pi54 and TP respectively (Fig 7D Table 3)
The activity of b-13-glucanase in transgenic plantswhich was already high started increasing after 24 hpi and
thereafter increased consistently up to 120 hpi Although
the b-13-glucanase activity was also higher in the trans-
genic control it did not show any significant increase after
24 hpi However in non-transgenic plants the enzyme was
slightly induced at 24 h but declined immediately until 72
hpi before reaching a normal level The highest b-1
3-glucanase activity was 8770 U in TP-Pi54 compared to727 U in TP (Fig 7E Table 3) The chitinase enzyme
concentration increased in transgenic plants at 48 hpi and
reached its maximum level after 120 hpi The maximum
chitinase activity was 1011 U in TP-Pi54 compared with
858 U in TP (Fig 7F Table 3)
Fig 5 Expression pattern as represented by the heat map profile
of mock-inoculated (control) versus TP-Pi54 (test) rice lines
inoculated with M oryzae Colour changes show the level of gene
expression in their respective clusters Yellow mildly expressed
genes blue highly suppressed genes red highly expressed
genes The clusters are genes for (A) response to stimulus (B)
signal transduction (C) transcription regulators (D) programmed
cell death (E) oxidoreductases and (F) lyase
Pi54 mediated resistance against rice blast | 763D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

biotin-tagged cRNA from an in vitro transcription reaction using anAffymetrix GeneChip IVT labelling kit Fifteen micrograms ofbiotin-tagged cRNA was fragmented (50 to 200 bp fragments) as perthe manufacturerrsquos protocols Subsequently 10 lg of this fragmentedcRNA was hybridized to rice gene chips (Affymetrix) at 45 C with16 h rotation in a GeneChip hybridization oven (Affymetrix)Duplicate biological samples from transgenic and control rice plantswere used for microarray analysis Each array contained probe setsfor 51 279 rice transcripts out of which around 48 564 were fromjaponica and 1260 from indica cultivars Seventy-two hours aftertreatment total RNA was isolated from leaf tissues inoculated withthe pathogen The samples from 20ndash30 challenge-inoculated leaveswere pooled together Total RNA was converted into biotin-labelledcRNA and hybridized to the rice gene chip Separate gene chips werehybridized for the M oryzae spore suspension prepared in 02gelatin and leaf samples mock-inoculated with 02 gelatin onlywere used as the mock control GeneChip arrays were washed andthen stained with streptavidin-phycoerythrin (Affymetrix FluidicsStation 450) followed by scanning with a GeneChip scanner(Affymetrix GeneChip Scanner 3000) FCA (fold change absolute)values were calculated for each transcript of TP-Pi54 and TP usingmock-inoculated chip data as their respective controls The FCAvalues of transgenic and native lines for each transcript werecompared
Microarray data analysis
MIAME-compliant microarray data from this study have beendeposited at NCBI Gene Expression Omnibus with the accessionnumber GSE28308 The array data set was analysed using theGeneChip Operating Software (GCOS 12) and ArrayStar(DNASTAR Inc Madison USA) software For probe levelnormalization robust multi-array analysis (RMA) was performedon Affymetrix raw files (CEL files) (Irizarry et al 2003) Theinitial data set consisted of 57 382 probes Filtration wasperformed to remove probes with lsquoAbsent Callsrsquo (poor qualityprobes) Transformation and normalization were done in order tofacilitate comparison across samples The biological significance ofdifferentially expressed genes was determined by performingfunctional classification according to Gene Ontology after filteringthe genes for gt20-fold change and lt005 P-value Gene annota-tion was performed using BLAST (httpblastncbinlmnihgovBlast) All the raw data files (CEL files two for blast-inoculatedand two for mock treatment) obtained from the GeneChipOperating Software (GCOS Affymetrix) were subjected to furtheranalysis using the ArrayStar software
Quantitative real-time PCR analysis
Part of the samples of total RNA isolated for microarray analysiswas also used to validate the functionally important genes that werefound to be differentially regulated in the microarray results Thenucleotide sequences of differentially regulated genes were down-loaded from the TIGR rice database (httpriceplantbiologymsuedu) (see Supplementary Table S5 at JXB online) The exonicsequences of selected genes were used to design primers using thePRIMER3 software (httpfrodowimiteduprimer3) For the genevalidation experiment a SYBR green-based One-Step qRT-PCR kit(Invitrogen Carlsbad CA USA) was used The thermal cyclingconditions were 50 C for 1 h followed by initial denaturation at 95C for 10 min and then 40 cycles of denaturation at 95 C for 30 sannealing at 55ndash65 C for 1 min and extension at 72 C for 1 min ina thermal cycler (Mx3000P Stratagene) Dissociation curve analysiswas performed by fluorescence reading at one-degree intervalsbetween 55ndash95 C to ensure that only one PCR product wasamplified All experiments were performed in two biological repli-cates and three technical replicates The expression data werenormalized to EF-1a using the DDCT method described by Livakand Schmittgen (2001)
Preparation of enzyme samples
To understand the role of major defence response enzymes andphenolic compounds in transgenic rice lines containing the singlerice blast resistance gene Pi54 enzymatic analysis was alsoperformed Challenge-inoculated leaf samples along with non-inoculated controls of transgenic and non-transgenic lines wereharvested after 0 24 48 72 96 and 120 hpi Total crude enzymeswere extracted from 1 g of leaf samples in a prechilled mortar andpestle and homogenized in their respective buffers (wv 11) suchas potassium phosphate buffer (10 mM pH 60) for POX(peroxidase) (18 000 g 15 min) and PPO (polyphenol oxidase)(12 000 g 20 min) sodium acetate buffer (005 M pH 46 23 000g 20 min) for laminarinase chitinase and b-glucosidase andborate buffer (25 mM pH 88) for PAL (8000 g 30 min) Thehomogenate was centrifuged at 4 C at specific speeds (given abovein parentheses) for the separate preparation of crude enzymeextract The supernatant was used as the enzyme source in all casesand for spectrophotometric estimation of enzymatic activity(Sadasivam and Manickam 2008) Spectrophotometric observa-tions were performed by Spectrophotometer GeneQuant 1300 (GEHealthcare Bio-sciences AB Uppsala Sweden) The proteincontents of the crude extract were determined by a standardmethod (Bradford 1976) using BSA (Sigma-Aldrich Chemical CoSt Louis MO USA) as the standard
Enzymatic assay
For the POX assay a reaction mixture (313 ml) was prepared byadding 3 ml of 01 M phosphate buffer (pH 70) 50 ll of 20 mMguaicol solution 30 ll of 123 mM hydrogen peroxide (014 ml of30 H2O2 to 100 ml diluted with distilled water prepared freshly)and 50 ll of enzyme extract The reaction was allowed to proceedfor 3 min A436 values were measured 30 s after adding the enzymeextract to the substrate and the change in absorbance wasrecorded
The reaction mixture for the PPO assay consisted of 15 ml of01 M sodium phosphate buffer (pH 65) and 200 ll of the enzymeextract The reaction was started with the addition of 200 ll of 10mM catechol (Mayer et al 1965) The rate of increase inabsorbance was measured at 420 nm for 1 min The activity wasexpressed as change in A436 mg1 protein min1
The PAL assay was performed using a modified procedure ofLisker et al (1983) by measuring the production of trans-cinnamicacid from L-phenylalanine The reaction mixture contained 1 ml ofenzyme extract 05 ml substrate of 50 mm L-phenylalanine and04 ml of 25 mM borate buffer (pH 88) After incubation for 2 hat 40 C the reaction was stopped by the addition of 006 ml 5 NHCl and absorbance was recorded at A290 and read against thesame volume of reaction mixture without L-phenylalanine whichserved as a blank The enzyme activity was expressed as l mol oftrans-cinnamic acid (t-CA) mg1 protein h1
For the beta-glucosidase assay enzyme activity was determinedby the modified method of Agrawal and Bahal (1969) A reactionmixture was prepared by adding 100 ll of a 25 mM solution ofp-nitrophenyl-b-D-glycopyaranoside to 350 ll of 005 M sodiumacetate buffer (pH 46) followed by initial incubation at 30 C for5 min After the addition of 50 ll of enzyme extract the mixturewas further incubated at 30 C for 15 min The reaction wasstopped by adding 700 ll of 02 M sodium carbonate The yellowcolour formed was measured at 420 nm by a spectrophotometerEnzyme activity was calculated as lmol of p-nitrophenol releasedmg1 of protein extract min1
For the b-13-glucanase assay enzyme activity was determinedby the modified method of Abeles and Forrence (1970) Thereaction mixture was prepared by adding 75 ll of enzyme extractto 175 ll of 2 (wv) Laminarin (Sigma-Aldrich USA) in waterand incubated for 60 min at 50 C in a water bath The reactionwas stopped by adding 750 ll of dinitrosalicylic acid reagent(Sigma-Aldrich USA) The tubes were then cooled to 25 C
Pi54 mediated resistance against rice blast | 759D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Whole contents were diluted 110 with distilled water Sampleswere vortexed and the A500 values recorded Glucose was used asa standard (Miller 1959) Enzyme activity was calculated as lmolof glucose released min1 mg1 protein extract
Chitinase activity was determined by quantitative estimation ofthe reducing sugars produced with colloidal chitin as the appropri-ate substrate (Sun et al 2006) Briefly the reaction mixtureconsisted of 05 ml enzyme solution and 05 ml of 1 colloidalchitin in 1 ml of 005 M sodium acetate buffer (pH 68) Themixture was incubated for 2 h at 40 C for chitinase and thereaction was stopped in boiling water for 5 min The amount ofreducing sugars released in the supernatant was measured bya method that uses dinitrosalicylic (DNS) acid reagent Theabsorbance was measured at 530 nm by a spectrophotometer Oneunit (U) of chitinase activity was defined as the amount of enzymethat liberated 1 lmol of the reducing sugar(s) min1 from N-acetylglucosamine under standard conditions
For the enzyme analysis all experiments were performed intriplicate The data obtained for each enzyme were analysedseparately Paired t test was performed in all the cases usingSigmaXL software (SigmaXL Toronto Canada) and differencesat Pfrac14005 were considered to be significant
Estimation of total phenol content
The total phenol contents in the pathogen inoculated leaf samplesof transgenic and non-transgenic rice at various intervals wereestimated following the modified procedure of Singleton and Rossi(1965) Rice leaves (1 g) were homogenized in 10 ml of 80methanol and agitated for 15 min at 70 C The homogenates werecentrifuged at 5000 g for 10 min An aliquot of 1 ml of methanolextract was made up to 5 ml with distilled water to which FolinndashCiocalteu reagent (250 ll) was added the solution was then kept at25 C After 3 min 1 ml saturated solution of Na2CO3 and 1 mldistilled water were added after which the reaction mixture wasincubated for 30 min at 25 C The A765 value of the developedblue colour was measured using a spectrophotometer (GeneQuant1300 GE Healthcare Bio-sciences AB Uppsala Sweden)A standard curve was prepared using gallic acid (Sigma-AldrichChemical Co USA) The phenol content of the extract wasexpressed as lg phenol equivalent released g1 of leaf tissue
Results
In order to observe the effect of the M oryzae-inducedexpression level of the Pi54 gene the transcriptomes of the
blast-resistant transgenic line TP-Pi54 and blast-susceptible
native TP lines were analysed at 72 hpi It has already been
shown that plants of the transgenic line TP-Pi54 have single
Pi54 gene insertions that are stably inherited up to three
generations (Rai et al 2011) The transgenic lines continu-
ally show a high degree of resistance and hypersensitive
response to the blast pathogen in the fifth generation as well(Fig 1)
Genome-wide expression analysis
For expression data analysis logarithmic (Log10) expression
values were normalized using the Robust Multichip Average(RMA) algorithm Further adopting a stringent criterion for
the expression fold change value (FCA) of gt20 and the
P value lt005 a total of 1154 differentially expressed genes
were identified in TP-Pi54 plants Of these 587 were up-
regulated whereas 567 genes were found to be down-regulated
(Table 1) However in the case of TP (susceptible) lines
a total of 1056 genes were found to be differentially regulated
of which 518 were up-regulated and 538 down-regulated A
total of 480 up- and 509 down-regulated genes were found to
be commonly expressed in both the TP-Pi54 and TP
(susceptible control) lines However 107 genes were found to
be exclusively up-regulated and 58 genes exclusively down-
regulated in the case of TP-Pi54 (Fig 2) Similarly in the TPline 38 genes were up-regulated and 29 down-regulated
Functional analysis of differentially regulated genes ofTP-Pi54
All the genes that showed differential expression in the
transgenic blast-resistant line TP-Pi54 were clustered and
functionally annotated The level of expression of these
genes in TP-Pi54 plants was compared with that in TP
plants Based on K-means clustering the functionally
annotated genes were classified into eight different clusters
Fig 1 Typical blast lesions as observed in the non-transgenic
susceptible rice line Taipei 309 (TP) No such disease symptoms
were observed in the resistant transgenic line (TP-Pi54) hypersen-
sitive response (HR) was observed in the TP-Pi54 line
Table 1 Gene ontology score-based categorization of differen-
tially regulated genes
Plants Up-regulated genes Down-regulated genes
TP-Pi54 (resistant) 587 [BPa 216 (1871)b 567 [BP 276 (2391)
CC 168 (1455) CC 203 (1759)
MF 274 (2374)] MF 322 (2790)]
TP (susceptible) 518 [BP 189 (1789) 538 [BP 212 (2007)
CC 150 (1420) CC 133 (1259)
MF 236 (2234)] MF 264 (25)]
a BP Biological process CC cellular component MF molecularfunction
b Numbers in parentheses represent the percentage in each set outof total differentially regulated genes of the plant
760 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
These genes were also analysed on the basis of their Gene
Ontology (GO) score into three main GO categories
including biological process (BP) molecular function (MF)
and cellular component (CC) (Table 1)
Similarly differentially regulated genes were assigned into
16 GO subcategories The percentages of up- and down-
regulated genes belonging to each category are given in Fig 3
Most (17) genes belonging to the binding-type category werefound to be up-regulated while 138 were down-regulated
Similarly 1534 of cell-part genes (GO ID004464) were up-
regulated whereas an almost equal number (1518) was
down-regulated in the transgenic line Interestingly the
number (1679) of metabolic process genes that were down-
regulated was higher than that of up-regulated (1349) ones
in this important category Likewise there were higher
numbers of down-regulated genes compared with up-regulatedones in a few other GO subcategories such as cellular
processes organelle and macromolecular complex On the
contrary 1247 462 and 341 of the genes belonging to
catalytic activity biological regulation and establishment of
localization were up-regulated respectively In some of the
subcategories such as transporter activity transcription
regulator molecular transducer and anatomical structure and
function the genes were only up-regulated (Fig 3)
Annotation information on differentially expressed genes
that were more up-regulated in TP-Pi54 was collected for
each probe set ID from GO analysis NCBI-BLAST and
NetAffx These genes were again categorized into various
functional groups A multitude of genes known for their
involvement in some way or another in stress responses (see
Supplementary Table S1 at JXB online) transcriptional
regulation (see Supplementary Table S2 at JXB online)programmed cell death (see Supplementary Table S3 at JXB
online) and signal transduction (see Supplementary Table
S4 at JXB online) were observed to be expressed differen-
tially in the transgenic line TP-Pi54 The level of expression
of these genes in the TP-Pi54 and TP lines varied from
207ndash1238-fold (see Supplementary Table S2 at JXB
online)
Functional annotation of stress response genes
Stress-response genes were specifically grouped together
according to the pathways in which they are involved Their
FCA values were represented as the average of the FCA
values of the members of each group and compared with
those of the TP (susceptible) lines (Fig 4A) Two genes for
callose biosynthesis were up-regulated 475-fold and 383-fold
in TP-Pi54 while in the TP lines these genes were up-
regulated only 2-fold following pathogen inoculation (seeSupplementary Table S1 at JXB online) One of the three
laccase enzyme-related genes was highly (31-fold) up-
regulated Similarly genes for other enzymatic categories like
phenyl ammonia lyase peroxidases lipoxygenases superoxide
dimustase amylase licheninase chitinase and glucosylcerami-
dase were up-regulated from 10-fold to 6391-fold in TP-Pi54
compared with the TP lines (see Supplementary Table S1
at JXB online) Interestingly various genes earlier reportedto be involved in other stresses such as wounding and abiotic
stresses were also more up-regulated in transgenic lines
Very high up-regulation of genes like laccase (729-fold
Fig 2 Venn diagram of genome-wide differentially expressed genes
in TP and TP-Pi54 following M oryzae inoculation Numbers in
parentheses indicate down-regulated genes U upregulated genes
Fig 3 Functional categorization of genes differentially expressed in the TP-Pi54 line The number of genes was finalized using the
filtering criteria of fold change gt20 and P-value correction ltfrac14005 by FDR (Benjamini-Hochberg) Genes were classified into 16 different
categories based on Gene Ontology The number of up- and down-regulated genes for each functional category is shown in the
histogram The functional categories are A binding B catalytic activity C transporter activity D transcription regulator activity E
molecular transducer activity F antioxidant activity G metabolic processes H cellular processes I biological regulation J
establishment of localization K response to stimulus L anatomical structure function M cell part N organelle O organelle part and
P molecular complex (A) Up-regulated genes (B) Down-regulated genes
Pi54 mediated resistance against rice blast | 761D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Os11g48060) peroxidases (1694-fold Os07g48010) ammonia
lyases (278-fold Os04g43760) and amylases (6391-fold
Os10g41550) were obtained (see Supplementary Table S1 at
JXB online) Similarly maximum up-regulation (1694ndash2545-
fold) of the genes for chemical stimulus was obtained with
a response to oxidative stress Several genes for transcription
factors (TF) were up-regulated in response to M oryzae
inoculation (see Supplementary Table S2 at JXB online)
Among the more prominently expressed are genes for NAC
domain-containing proteins (841-fold) Dof zinc finger pro-
teins (1134-fold) sigma factors (1238-fold) bZIP transcrip-
tion factors (1193-fold) and MADS box proteins (446-fold)
The up-regulation of the rest of the TF genes in transgenic
plants varied from 203-fold to 1238-fold It was interesting to
note that all these TF genes were regulated at a very low level
in the non-transgenic plants (see Supplementary Table S2 at
JXB online Fig 4B) Programmed cell death (PCD) is one of
the important responses of plants upon pathogen infection
and leads to a hypersensitive response in resistant lines In this
study four genes with predicted roles in PCD were up-
regulated up to 207ndash276-fold in transgenic lines compared
with between 067ndash159-fold in non-transgenic lines (see
Supplementary Table S3 at JXB online Fig 4B Fig 4C)
Fifteen genes associated with vital signal transduction
functions in the plants were also highly up-regulated in TP-
Pi54 (see Supplementary Table S4 at JXB online Fig 4B)
A maximum up-regulation of 1202-fold was obtained in
auxin-mediated signal pathways followed by that ingibberellic acid-mediated signalling (328-fold) Similarly
genes for two-component signal transduction pathways
were up-regulated between 244-fold and 356-fold (see
Supplementary Table S4 at JXB online Fig 4D)
Hierarchical clustering
Genome-wide analysis of the expression patterns of mock-
inoculated and pathogen-inoculated TP-Pi54 transgenicplants was performed The phylogenetic relationship be-
tween highly expressed and less expressed genes was
obtained after hierarchal clustering The expression changes
among the gene group members in six clusters (i) response
to stimulus (ii) signal transduction (iii) transcription
Fig 4 Comparison of the differential regulation of different categories of genes more up-regulated in TP-Pi54 compared to TP
(A) Stress-responsive genes 1 callose 2 laccase 3 PAL 4 Peroxidase 5 lipoxygenase 6 superoxide dismutase 7 amylase 8
licheninase 9 chitinase 10 glucosylceramidase 11 response to wounding 12 response to chemical stimulus and 13 abiotic stress
(B) 1ndash22 Transcription factor genes (C) 1ndash4 Genes for programmed cell death (D) 1ndash15 Genes for signal transduction
762 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
regulators (iv) programmed cell death (v) oxidoreductases
and (vi) lyase were represented by different colours in the
heat map The colour intensities indicate the expression
level of each gene (Fig 5)
Genes repressed in native TP lines
Out of the 107 up-regulated genes of TP-Pi54 29 were
considerably repressed in the non-transgenic TP line (Table 2)
Among these the genes Os77671S1_at (652-fold) andOs70041S1_at (2458-fold) were up-regulated at the highest
level in TP-Pi54 compared with non-transgenic lines Other
highly expressed genes in TP-Pi54 that were otherwise
repressed in TP belonged to some important categories such
as genes involved in transporter activity (2726-fold and
1692-fold) zinc ion binding (1033-fold) and lipid transport
(685-fold) Interestingly a mitochondrial located gene
(Os124301S1_at) was also highly up-regulated (15-fold) inTP-Pi54 and down-regulated in the TP lines (Table 2) Among
all the up-regulated genes in TP-54 22 were hypothetically
expressed proteins of unknown function This indicates that
these genes may be novel and may have an important role in
the Pi54-mediated defence mechanism in the blast-resistant
transgenic line TP-Pi54
Quantitative analysis of differentially regulated genes
Validation of nine up-regulated genes and one down-regulated gene in the transgenic lines was performed by
quantitative real-time PCR Genes for NAC domain-
containing proteins (NAC6) callose polysaccharide
biosynthesis proteins (CAPD) Dof zinc finger proteins
(DOF) peroxidase (POC1) MADS box proteins (MADS)
class III peroxidase (POX3) chitinase (CHIT) phenylala-
nine ammonia lyase (PAL) and endo-13-14-b-glucanase
(BG) were found to be up-regulated 124ndash58-fold whereas
genes for homeo domain-related proteins (HDR) were
down-regulated up to 14-fold (Fig 6 see Supplementary
Table S6 at JXB online Fig 4B)
Biochemical assay of enzymes involved in defenceresponse
To assess the effect of pathogen infection on plant defence
response enzymes the activities of different enzymes were
measured individually in transgenic (TP-Pi54) and non-
transgenic (TP) plants at 0 to 120 hpi (Fig 7) The data
obtained from transgenic plants were compared with those
from the non-inoculated plants used as controls M oryzae
inoculation drastically increased peroxidase activity intransgenic rice plants after 96 hpi The POX activity at 96
hpi in TP-Pi54 was higher (01623 U) than that in TP
(Fig 7A Table 3) The changes in polyphenol oxidase
activity in the plants followed more or less the same pattern
as observed for POX The PPO activity increased at 24 hpi
in transgenic plants reached its maximum at 96 hpi and
declined thereafter The PPO activity was measured as
change in A420 (DA420) mg1 protein min1 The PPOactivity at 96 h was 01736 U in TP-Pi54 compared with
01 U in TP (Fig 7B Table 3)
The time-course of PAL activity in treated plants was
quite distinct from that of the rest of the enzymes as the
profile represented a bimodal pattern in transgenic plants
indicating an early and sharp induction at 24 hpi (Fig 7C)
The decline in PAL activity continued until 72 hpi and
increased thereafter until 96 hpi before following another
dip at 120 hpi compared with that in non-inoculated
transgenic control plants which showed no induction at all
The maximum PAL activity at 24 hpi was 9305 U in TP-
Pi54 and 1768 U in TP (Fig 7C Table 3) b-Glucosidase
activity in plants gradually increased upon pathogen in-
oculation reaching its maximum at 48 hpi in the transgenic
group On the other hand a bimodal pattern of activity was
obtained in the case of non-transgenic plants showing
a slight increase in the activity at 24 and 96 hpi The activity
of b-glucosidase at 48 h was 00809 U and 00214 U in TP-
Pi54 and TP respectively (Fig 7D Table 3)
The activity of b-13-glucanase in transgenic plantswhich was already high started increasing after 24 hpi and
thereafter increased consistently up to 120 hpi Although
the b-13-glucanase activity was also higher in the trans-
genic control it did not show any significant increase after
24 hpi However in non-transgenic plants the enzyme was
slightly induced at 24 h but declined immediately until 72
hpi before reaching a normal level The highest b-1
3-glucanase activity was 8770 U in TP-Pi54 compared to727 U in TP (Fig 7E Table 3) The chitinase enzyme
concentration increased in transgenic plants at 48 hpi and
reached its maximum level after 120 hpi The maximum
chitinase activity was 1011 U in TP-Pi54 compared with
858 U in TP (Fig 7F Table 3)
Fig 5 Expression pattern as represented by the heat map profile
of mock-inoculated (control) versus TP-Pi54 (test) rice lines
inoculated with M oryzae Colour changes show the level of gene
expression in their respective clusters Yellow mildly expressed
genes blue highly suppressed genes red highly expressed
genes The clusters are genes for (A) response to stimulus (B)
signal transduction (C) transcription regulators (D) programmed
cell death (E) oxidoreductases and (F) lyase
Pi54 mediated resistance against rice blast | 763D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

Whole contents were diluted 110 with distilled water Sampleswere vortexed and the A500 values recorded Glucose was used asa standard (Miller 1959) Enzyme activity was calculated as lmolof glucose released min1 mg1 protein extract
Chitinase activity was determined by quantitative estimation ofthe reducing sugars produced with colloidal chitin as the appropri-ate substrate (Sun et al 2006) Briefly the reaction mixtureconsisted of 05 ml enzyme solution and 05 ml of 1 colloidalchitin in 1 ml of 005 M sodium acetate buffer (pH 68) Themixture was incubated for 2 h at 40 C for chitinase and thereaction was stopped in boiling water for 5 min The amount ofreducing sugars released in the supernatant was measured bya method that uses dinitrosalicylic (DNS) acid reagent Theabsorbance was measured at 530 nm by a spectrophotometer Oneunit (U) of chitinase activity was defined as the amount of enzymethat liberated 1 lmol of the reducing sugar(s) min1 from N-acetylglucosamine under standard conditions
For the enzyme analysis all experiments were performed intriplicate The data obtained for each enzyme were analysedseparately Paired t test was performed in all the cases usingSigmaXL software (SigmaXL Toronto Canada) and differencesat Pfrac14005 were considered to be significant
Estimation of total phenol content
The total phenol contents in the pathogen inoculated leaf samplesof transgenic and non-transgenic rice at various intervals wereestimated following the modified procedure of Singleton and Rossi(1965) Rice leaves (1 g) were homogenized in 10 ml of 80methanol and agitated for 15 min at 70 C The homogenates werecentrifuged at 5000 g for 10 min An aliquot of 1 ml of methanolextract was made up to 5 ml with distilled water to which FolinndashCiocalteu reagent (250 ll) was added the solution was then kept at25 C After 3 min 1 ml saturated solution of Na2CO3 and 1 mldistilled water were added after which the reaction mixture wasincubated for 30 min at 25 C The A765 value of the developedblue colour was measured using a spectrophotometer (GeneQuant1300 GE Healthcare Bio-sciences AB Uppsala Sweden)A standard curve was prepared using gallic acid (Sigma-AldrichChemical Co USA) The phenol content of the extract wasexpressed as lg phenol equivalent released g1 of leaf tissue
Results
In order to observe the effect of the M oryzae-inducedexpression level of the Pi54 gene the transcriptomes of the
blast-resistant transgenic line TP-Pi54 and blast-susceptible
native TP lines were analysed at 72 hpi It has already been
shown that plants of the transgenic line TP-Pi54 have single
Pi54 gene insertions that are stably inherited up to three
generations (Rai et al 2011) The transgenic lines continu-
ally show a high degree of resistance and hypersensitive
response to the blast pathogen in the fifth generation as well(Fig 1)
Genome-wide expression analysis
For expression data analysis logarithmic (Log10) expression
values were normalized using the Robust Multichip Average(RMA) algorithm Further adopting a stringent criterion for
the expression fold change value (FCA) of gt20 and the
P value lt005 a total of 1154 differentially expressed genes
were identified in TP-Pi54 plants Of these 587 were up-
regulated whereas 567 genes were found to be down-regulated
(Table 1) However in the case of TP (susceptible) lines
a total of 1056 genes were found to be differentially regulated
of which 518 were up-regulated and 538 down-regulated A
total of 480 up- and 509 down-regulated genes were found to
be commonly expressed in both the TP-Pi54 and TP
(susceptible control) lines However 107 genes were found to
be exclusively up-regulated and 58 genes exclusively down-
regulated in the case of TP-Pi54 (Fig 2) Similarly in the TPline 38 genes were up-regulated and 29 down-regulated
Functional analysis of differentially regulated genes ofTP-Pi54
All the genes that showed differential expression in the
transgenic blast-resistant line TP-Pi54 were clustered and
functionally annotated The level of expression of these
genes in TP-Pi54 plants was compared with that in TP
plants Based on K-means clustering the functionally
annotated genes were classified into eight different clusters
Fig 1 Typical blast lesions as observed in the non-transgenic
susceptible rice line Taipei 309 (TP) No such disease symptoms
were observed in the resistant transgenic line (TP-Pi54) hypersen-
sitive response (HR) was observed in the TP-Pi54 line
Table 1 Gene ontology score-based categorization of differen-
tially regulated genes
Plants Up-regulated genes Down-regulated genes
TP-Pi54 (resistant) 587 [BPa 216 (1871)b 567 [BP 276 (2391)
CC 168 (1455) CC 203 (1759)
MF 274 (2374)] MF 322 (2790)]
TP (susceptible) 518 [BP 189 (1789) 538 [BP 212 (2007)
CC 150 (1420) CC 133 (1259)
MF 236 (2234)] MF 264 (25)]
a BP Biological process CC cellular component MF molecularfunction
b Numbers in parentheses represent the percentage in each set outof total differentially regulated genes of the plant
760 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
These genes were also analysed on the basis of their Gene
Ontology (GO) score into three main GO categories
including biological process (BP) molecular function (MF)
and cellular component (CC) (Table 1)
Similarly differentially regulated genes were assigned into
16 GO subcategories The percentages of up- and down-
regulated genes belonging to each category are given in Fig 3
Most (17) genes belonging to the binding-type category werefound to be up-regulated while 138 were down-regulated
Similarly 1534 of cell-part genes (GO ID004464) were up-
regulated whereas an almost equal number (1518) was
down-regulated in the transgenic line Interestingly the
number (1679) of metabolic process genes that were down-
regulated was higher than that of up-regulated (1349) ones
in this important category Likewise there were higher
numbers of down-regulated genes compared with up-regulatedones in a few other GO subcategories such as cellular
processes organelle and macromolecular complex On the
contrary 1247 462 and 341 of the genes belonging to
catalytic activity biological regulation and establishment of
localization were up-regulated respectively In some of the
subcategories such as transporter activity transcription
regulator molecular transducer and anatomical structure and
function the genes were only up-regulated (Fig 3)
Annotation information on differentially expressed genes
that were more up-regulated in TP-Pi54 was collected for
each probe set ID from GO analysis NCBI-BLAST and
NetAffx These genes were again categorized into various
functional groups A multitude of genes known for their
involvement in some way or another in stress responses (see
Supplementary Table S1 at JXB online) transcriptional
regulation (see Supplementary Table S2 at JXB online)programmed cell death (see Supplementary Table S3 at JXB
online) and signal transduction (see Supplementary Table
S4 at JXB online) were observed to be expressed differen-
tially in the transgenic line TP-Pi54 The level of expression
of these genes in the TP-Pi54 and TP lines varied from
207ndash1238-fold (see Supplementary Table S2 at JXB
online)
Functional annotation of stress response genes
Stress-response genes were specifically grouped together
according to the pathways in which they are involved Their
FCA values were represented as the average of the FCA
values of the members of each group and compared with
those of the TP (susceptible) lines (Fig 4A) Two genes for
callose biosynthesis were up-regulated 475-fold and 383-fold
in TP-Pi54 while in the TP lines these genes were up-
regulated only 2-fold following pathogen inoculation (seeSupplementary Table S1 at JXB online) One of the three
laccase enzyme-related genes was highly (31-fold) up-
regulated Similarly genes for other enzymatic categories like
phenyl ammonia lyase peroxidases lipoxygenases superoxide
dimustase amylase licheninase chitinase and glucosylcerami-
dase were up-regulated from 10-fold to 6391-fold in TP-Pi54
compared with the TP lines (see Supplementary Table S1
at JXB online) Interestingly various genes earlier reportedto be involved in other stresses such as wounding and abiotic
stresses were also more up-regulated in transgenic lines
Very high up-regulation of genes like laccase (729-fold
Fig 2 Venn diagram of genome-wide differentially expressed genes
in TP and TP-Pi54 following M oryzae inoculation Numbers in
parentheses indicate down-regulated genes U upregulated genes
Fig 3 Functional categorization of genes differentially expressed in the TP-Pi54 line The number of genes was finalized using the
filtering criteria of fold change gt20 and P-value correction ltfrac14005 by FDR (Benjamini-Hochberg) Genes were classified into 16 different
categories based on Gene Ontology The number of up- and down-regulated genes for each functional category is shown in the
histogram The functional categories are A binding B catalytic activity C transporter activity D transcription regulator activity E
molecular transducer activity F antioxidant activity G metabolic processes H cellular processes I biological regulation J
establishment of localization K response to stimulus L anatomical structure function M cell part N organelle O organelle part and
P molecular complex (A) Up-regulated genes (B) Down-regulated genes
Pi54 mediated resistance against rice blast | 761D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Os11g48060) peroxidases (1694-fold Os07g48010) ammonia
lyases (278-fold Os04g43760) and amylases (6391-fold
Os10g41550) were obtained (see Supplementary Table S1 at
JXB online) Similarly maximum up-regulation (1694ndash2545-
fold) of the genes for chemical stimulus was obtained with
a response to oxidative stress Several genes for transcription
factors (TF) were up-regulated in response to M oryzae
inoculation (see Supplementary Table S2 at JXB online)
Among the more prominently expressed are genes for NAC
domain-containing proteins (841-fold) Dof zinc finger pro-
teins (1134-fold) sigma factors (1238-fold) bZIP transcrip-
tion factors (1193-fold) and MADS box proteins (446-fold)
The up-regulation of the rest of the TF genes in transgenic
plants varied from 203-fold to 1238-fold It was interesting to
note that all these TF genes were regulated at a very low level
in the non-transgenic plants (see Supplementary Table S2 at
JXB online Fig 4B) Programmed cell death (PCD) is one of
the important responses of plants upon pathogen infection
and leads to a hypersensitive response in resistant lines In this
study four genes with predicted roles in PCD were up-
regulated up to 207ndash276-fold in transgenic lines compared
with between 067ndash159-fold in non-transgenic lines (see
Supplementary Table S3 at JXB online Fig 4B Fig 4C)
Fifteen genes associated with vital signal transduction
functions in the plants were also highly up-regulated in TP-
Pi54 (see Supplementary Table S4 at JXB online Fig 4B)
A maximum up-regulation of 1202-fold was obtained in
auxin-mediated signal pathways followed by that ingibberellic acid-mediated signalling (328-fold) Similarly
genes for two-component signal transduction pathways
were up-regulated between 244-fold and 356-fold (see
Supplementary Table S4 at JXB online Fig 4D)
Hierarchical clustering
Genome-wide analysis of the expression patterns of mock-
inoculated and pathogen-inoculated TP-Pi54 transgenicplants was performed The phylogenetic relationship be-
tween highly expressed and less expressed genes was
obtained after hierarchal clustering The expression changes
among the gene group members in six clusters (i) response
to stimulus (ii) signal transduction (iii) transcription
Fig 4 Comparison of the differential regulation of different categories of genes more up-regulated in TP-Pi54 compared to TP
(A) Stress-responsive genes 1 callose 2 laccase 3 PAL 4 Peroxidase 5 lipoxygenase 6 superoxide dismutase 7 amylase 8
licheninase 9 chitinase 10 glucosylceramidase 11 response to wounding 12 response to chemical stimulus and 13 abiotic stress
(B) 1ndash22 Transcription factor genes (C) 1ndash4 Genes for programmed cell death (D) 1ndash15 Genes for signal transduction
762 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
regulators (iv) programmed cell death (v) oxidoreductases
and (vi) lyase were represented by different colours in the
heat map The colour intensities indicate the expression
level of each gene (Fig 5)
Genes repressed in native TP lines
Out of the 107 up-regulated genes of TP-Pi54 29 were
considerably repressed in the non-transgenic TP line (Table 2)
Among these the genes Os77671S1_at (652-fold) andOs70041S1_at (2458-fold) were up-regulated at the highest
level in TP-Pi54 compared with non-transgenic lines Other
highly expressed genes in TP-Pi54 that were otherwise
repressed in TP belonged to some important categories such
as genes involved in transporter activity (2726-fold and
1692-fold) zinc ion binding (1033-fold) and lipid transport
(685-fold) Interestingly a mitochondrial located gene
(Os124301S1_at) was also highly up-regulated (15-fold) inTP-Pi54 and down-regulated in the TP lines (Table 2) Among
all the up-regulated genes in TP-54 22 were hypothetically
expressed proteins of unknown function This indicates that
these genes may be novel and may have an important role in
the Pi54-mediated defence mechanism in the blast-resistant
transgenic line TP-Pi54
Quantitative analysis of differentially regulated genes
Validation of nine up-regulated genes and one down-regulated gene in the transgenic lines was performed by
quantitative real-time PCR Genes for NAC domain-
containing proteins (NAC6) callose polysaccharide
biosynthesis proteins (CAPD) Dof zinc finger proteins
(DOF) peroxidase (POC1) MADS box proteins (MADS)
class III peroxidase (POX3) chitinase (CHIT) phenylala-
nine ammonia lyase (PAL) and endo-13-14-b-glucanase
(BG) were found to be up-regulated 124ndash58-fold whereas
genes for homeo domain-related proteins (HDR) were
down-regulated up to 14-fold (Fig 6 see Supplementary
Table S6 at JXB online Fig 4B)
Biochemical assay of enzymes involved in defenceresponse
To assess the effect of pathogen infection on plant defence
response enzymes the activities of different enzymes were
measured individually in transgenic (TP-Pi54) and non-
transgenic (TP) plants at 0 to 120 hpi (Fig 7) The data
obtained from transgenic plants were compared with those
from the non-inoculated plants used as controls M oryzae
inoculation drastically increased peroxidase activity intransgenic rice plants after 96 hpi The POX activity at 96
hpi in TP-Pi54 was higher (01623 U) than that in TP
(Fig 7A Table 3) The changes in polyphenol oxidase
activity in the plants followed more or less the same pattern
as observed for POX The PPO activity increased at 24 hpi
in transgenic plants reached its maximum at 96 hpi and
declined thereafter The PPO activity was measured as
change in A420 (DA420) mg1 protein min1 The PPOactivity at 96 h was 01736 U in TP-Pi54 compared with
01 U in TP (Fig 7B Table 3)
The time-course of PAL activity in treated plants was
quite distinct from that of the rest of the enzymes as the
profile represented a bimodal pattern in transgenic plants
indicating an early and sharp induction at 24 hpi (Fig 7C)
The decline in PAL activity continued until 72 hpi and
increased thereafter until 96 hpi before following another
dip at 120 hpi compared with that in non-inoculated
transgenic control plants which showed no induction at all
The maximum PAL activity at 24 hpi was 9305 U in TP-
Pi54 and 1768 U in TP (Fig 7C Table 3) b-Glucosidase
activity in plants gradually increased upon pathogen in-
oculation reaching its maximum at 48 hpi in the transgenic
group On the other hand a bimodal pattern of activity was
obtained in the case of non-transgenic plants showing
a slight increase in the activity at 24 and 96 hpi The activity
of b-glucosidase at 48 h was 00809 U and 00214 U in TP-
Pi54 and TP respectively (Fig 7D Table 3)
The activity of b-13-glucanase in transgenic plantswhich was already high started increasing after 24 hpi and
thereafter increased consistently up to 120 hpi Although
the b-13-glucanase activity was also higher in the trans-
genic control it did not show any significant increase after
24 hpi However in non-transgenic plants the enzyme was
slightly induced at 24 h but declined immediately until 72
hpi before reaching a normal level The highest b-1
3-glucanase activity was 8770 U in TP-Pi54 compared to727 U in TP (Fig 7E Table 3) The chitinase enzyme
concentration increased in transgenic plants at 48 hpi and
reached its maximum level after 120 hpi The maximum
chitinase activity was 1011 U in TP-Pi54 compared with
858 U in TP (Fig 7F Table 3)
Fig 5 Expression pattern as represented by the heat map profile
of mock-inoculated (control) versus TP-Pi54 (test) rice lines
inoculated with M oryzae Colour changes show the level of gene
expression in their respective clusters Yellow mildly expressed
genes blue highly suppressed genes red highly expressed
genes The clusters are genes for (A) response to stimulus (B)
signal transduction (C) transcription regulators (D) programmed
cell death (E) oxidoreductases and (F) lyase
Pi54 mediated resistance against rice blast | 763D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
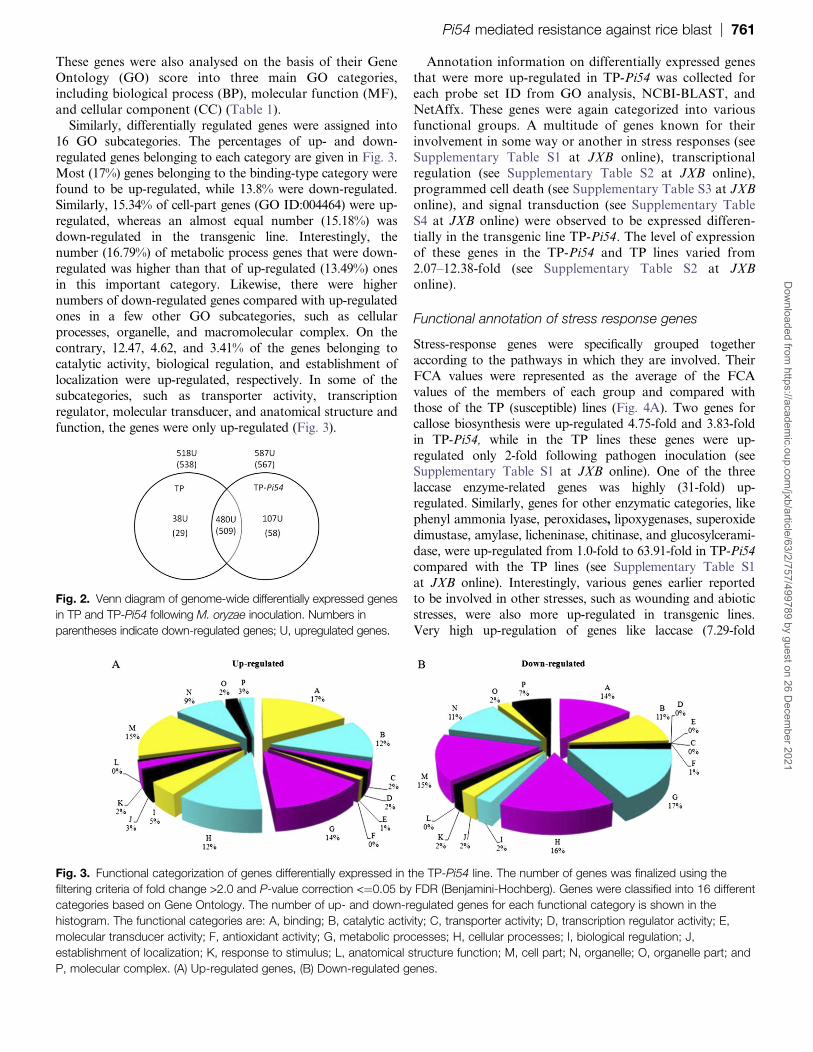
These genes were also analysed on the basis of their Gene
Ontology (GO) score into three main GO categories
including biological process (BP) molecular function (MF)
and cellular component (CC) (Table 1)
Similarly differentially regulated genes were assigned into
16 GO subcategories The percentages of up- and down-
regulated genes belonging to each category are given in Fig 3
Most (17) genes belonging to the binding-type category werefound to be up-regulated while 138 were down-regulated
Similarly 1534 of cell-part genes (GO ID004464) were up-
regulated whereas an almost equal number (1518) was
down-regulated in the transgenic line Interestingly the
number (1679) of metabolic process genes that were down-
regulated was higher than that of up-regulated (1349) ones
in this important category Likewise there were higher
numbers of down-regulated genes compared with up-regulatedones in a few other GO subcategories such as cellular
processes organelle and macromolecular complex On the
contrary 1247 462 and 341 of the genes belonging to
catalytic activity biological regulation and establishment of
localization were up-regulated respectively In some of the
subcategories such as transporter activity transcription
regulator molecular transducer and anatomical structure and
function the genes were only up-regulated (Fig 3)
Annotation information on differentially expressed genes
that were more up-regulated in TP-Pi54 was collected for
each probe set ID from GO analysis NCBI-BLAST and
NetAffx These genes were again categorized into various
functional groups A multitude of genes known for their
involvement in some way or another in stress responses (see
Supplementary Table S1 at JXB online) transcriptional
regulation (see Supplementary Table S2 at JXB online)programmed cell death (see Supplementary Table S3 at JXB
online) and signal transduction (see Supplementary Table
S4 at JXB online) were observed to be expressed differen-
tially in the transgenic line TP-Pi54 The level of expression
of these genes in the TP-Pi54 and TP lines varied from
207ndash1238-fold (see Supplementary Table S2 at JXB
online)
Functional annotation of stress response genes
Stress-response genes were specifically grouped together
according to the pathways in which they are involved Their
FCA values were represented as the average of the FCA
values of the members of each group and compared with
those of the TP (susceptible) lines (Fig 4A) Two genes for
callose biosynthesis were up-regulated 475-fold and 383-fold
in TP-Pi54 while in the TP lines these genes were up-
regulated only 2-fold following pathogen inoculation (seeSupplementary Table S1 at JXB online) One of the three
laccase enzyme-related genes was highly (31-fold) up-
regulated Similarly genes for other enzymatic categories like
phenyl ammonia lyase peroxidases lipoxygenases superoxide
dimustase amylase licheninase chitinase and glucosylcerami-
dase were up-regulated from 10-fold to 6391-fold in TP-Pi54
compared with the TP lines (see Supplementary Table S1
at JXB online) Interestingly various genes earlier reportedto be involved in other stresses such as wounding and abiotic
stresses were also more up-regulated in transgenic lines
Very high up-regulation of genes like laccase (729-fold
Fig 2 Venn diagram of genome-wide differentially expressed genes
in TP and TP-Pi54 following M oryzae inoculation Numbers in
parentheses indicate down-regulated genes U upregulated genes
Fig 3 Functional categorization of genes differentially expressed in the TP-Pi54 line The number of genes was finalized using the
filtering criteria of fold change gt20 and P-value correction ltfrac14005 by FDR (Benjamini-Hochberg) Genes were classified into 16 different
categories based on Gene Ontology The number of up- and down-regulated genes for each functional category is shown in the
histogram The functional categories are A binding B catalytic activity C transporter activity D transcription regulator activity E
molecular transducer activity F antioxidant activity G metabolic processes H cellular processes I biological regulation J
establishment of localization K response to stimulus L anatomical structure function M cell part N organelle O organelle part and
P molecular complex (A) Up-regulated genes (B) Down-regulated genes
Pi54 mediated resistance against rice blast | 761D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Os11g48060) peroxidases (1694-fold Os07g48010) ammonia
lyases (278-fold Os04g43760) and amylases (6391-fold
Os10g41550) were obtained (see Supplementary Table S1 at
JXB online) Similarly maximum up-regulation (1694ndash2545-
fold) of the genes for chemical stimulus was obtained with
a response to oxidative stress Several genes for transcription
factors (TF) were up-regulated in response to M oryzae
inoculation (see Supplementary Table S2 at JXB online)
Among the more prominently expressed are genes for NAC
domain-containing proteins (841-fold) Dof zinc finger pro-
teins (1134-fold) sigma factors (1238-fold) bZIP transcrip-
tion factors (1193-fold) and MADS box proteins (446-fold)
The up-regulation of the rest of the TF genes in transgenic
plants varied from 203-fold to 1238-fold It was interesting to
note that all these TF genes were regulated at a very low level
in the non-transgenic plants (see Supplementary Table S2 at
JXB online Fig 4B) Programmed cell death (PCD) is one of
the important responses of plants upon pathogen infection
and leads to a hypersensitive response in resistant lines In this
study four genes with predicted roles in PCD were up-
regulated up to 207ndash276-fold in transgenic lines compared
with between 067ndash159-fold in non-transgenic lines (see
Supplementary Table S3 at JXB online Fig 4B Fig 4C)
Fifteen genes associated with vital signal transduction
functions in the plants were also highly up-regulated in TP-
Pi54 (see Supplementary Table S4 at JXB online Fig 4B)
A maximum up-regulation of 1202-fold was obtained in
auxin-mediated signal pathways followed by that ingibberellic acid-mediated signalling (328-fold) Similarly
genes for two-component signal transduction pathways
were up-regulated between 244-fold and 356-fold (see
Supplementary Table S4 at JXB online Fig 4D)
Hierarchical clustering
Genome-wide analysis of the expression patterns of mock-
inoculated and pathogen-inoculated TP-Pi54 transgenicplants was performed The phylogenetic relationship be-
tween highly expressed and less expressed genes was
obtained after hierarchal clustering The expression changes
among the gene group members in six clusters (i) response
to stimulus (ii) signal transduction (iii) transcription
Fig 4 Comparison of the differential regulation of different categories of genes more up-regulated in TP-Pi54 compared to TP
(A) Stress-responsive genes 1 callose 2 laccase 3 PAL 4 Peroxidase 5 lipoxygenase 6 superoxide dismutase 7 amylase 8
licheninase 9 chitinase 10 glucosylceramidase 11 response to wounding 12 response to chemical stimulus and 13 abiotic stress
(B) 1ndash22 Transcription factor genes (C) 1ndash4 Genes for programmed cell death (D) 1ndash15 Genes for signal transduction
762 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
regulators (iv) programmed cell death (v) oxidoreductases
and (vi) lyase were represented by different colours in the
heat map The colour intensities indicate the expression
level of each gene (Fig 5)
Genes repressed in native TP lines
Out of the 107 up-regulated genes of TP-Pi54 29 were
considerably repressed in the non-transgenic TP line (Table 2)
Among these the genes Os77671S1_at (652-fold) andOs70041S1_at (2458-fold) were up-regulated at the highest
level in TP-Pi54 compared with non-transgenic lines Other
highly expressed genes in TP-Pi54 that were otherwise
repressed in TP belonged to some important categories such
as genes involved in transporter activity (2726-fold and
1692-fold) zinc ion binding (1033-fold) and lipid transport
(685-fold) Interestingly a mitochondrial located gene
(Os124301S1_at) was also highly up-regulated (15-fold) inTP-Pi54 and down-regulated in the TP lines (Table 2) Among
all the up-regulated genes in TP-54 22 were hypothetically
expressed proteins of unknown function This indicates that
these genes may be novel and may have an important role in
the Pi54-mediated defence mechanism in the blast-resistant
transgenic line TP-Pi54
Quantitative analysis of differentially regulated genes
Validation of nine up-regulated genes and one down-regulated gene in the transgenic lines was performed by
quantitative real-time PCR Genes for NAC domain-
containing proteins (NAC6) callose polysaccharide
biosynthesis proteins (CAPD) Dof zinc finger proteins
(DOF) peroxidase (POC1) MADS box proteins (MADS)
class III peroxidase (POX3) chitinase (CHIT) phenylala-
nine ammonia lyase (PAL) and endo-13-14-b-glucanase
(BG) were found to be up-regulated 124ndash58-fold whereas
genes for homeo domain-related proteins (HDR) were
down-regulated up to 14-fold (Fig 6 see Supplementary
Table S6 at JXB online Fig 4B)
Biochemical assay of enzymes involved in defenceresponse
To assess the effect of pathogen infection on plant defence
response enzymes the activities of different enzymes were
measured individually in transgenic (TP-Pi54) and non-
transgenic (TP) plants at 0 to 120 hpi (Fig 7) The data
obtained from transgenic plants were compared with those
from the non-inoculated plants used as controls M oryzae
inoculation drastically increased peroxidase activity intransgenic rice plants after 96 hpi The POX activity at 96
hpi in TP-Pi54 was higher (01623 U) than that in TP
(Fig 7A Table 3) The changes in polyphenol oxidase
activity in the plants followed more or less the same pattern
as observed for POX The PPO activity increased at 24 hpi
in transgenic plants reached its maximum at 96 hpi and
declined thereafter The PPO activity was measured as
change in A420 (DA420) mg1 protein min1 The PPOactivity at 96 h was 01736 U in TP-Pi54 compared with
01 U in TP (Fig 7B Table 3)
The time-course of PAL activity in treated plants was
quite distinct from that of the rest of the enzymes as the
profile represented a bimodal pattern in transgenic plants
indicating an early and sharp induction at 24 hpi (Fig 7C)
The decline in PAL activity continued until 72 hpi and
increased thereafter until 96 hpi before following another
dip at 120 hpi compared with that in non-inoculated
transgenic control plants which showed no induction at all
The maximum PAL activity at 24 hpi was 9305 U in TP-
Pi54 and 1768 U in TP (Fig 7C Table 3) b-Glucosidase
activity in plants gradually increased upon pathogen in-
oculation reaching its maximum at 48 hpi in the transgenic
group On the other hand a bimodal pattern of activity was
obtained in the case of non-transgenic plants showing
a slight increase in the activity at 24 and 96 hpi The activity
of b-glucosidase at 48 h was 00809 U and 00214 U in TP-
Pi54 and TP respectively (Fig 7D Table 3)
The activity of b-13-glucanase in transgenic plantswhich was already high started increasing after 24 hpi and
thereafter increased consistently up to 120 hpi Although
the b-13-glucanase activity was also higher in the trans-
genic control it did not show any significant increase after
24 hpi However in non-transgenic plants the enzyme was
slightly induced at 24 h but declined immediately until 72
hpi before reaching a normal level The highest b-1
3-glucanase activity was 8770 U in TP-Pi54 compared to727 U in TP (Fig 7E Table 3) The chitinase enzyme
concentration increased in transgenic plants at 48 hpi and
reached its maximum level after 120 hpi The maximum
chitinase activity was 1011 U in TP-Pi54 compared with
858 U in TP (Fig 7F Table 3)
Fig 5 Expression pattern as represented by the heat map profile
of mock-inoculated (control) versus TP-Pi54 (test) rice lines
inoculated with M oryzae Colour changes show the level of gene
expression in their respective clusters Yellow mildly expressed
genes blue highly suppressed genes red highly expressed
genes The clusters are genes for (A) response to stimulus (B)
signal transduction (C) transcription regulators (D) programmed
cell death (E) oxidoreductases and (F) lyase
Pi54 mediated resistance against rice blast | 763D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

Os11g48060) peroxidases (1694-fold Os07g48010) ammonia
lyases (278-fold Os04g43760) and amylases (6391-fold
Os10g41550) were obtained (see Supplementary Table S1 at
JXB online) Similarly maximum up-regulation (1694ndash2545-
fold) of the genes for chemical stimulus was obtained with
a response to oxidative stress Several genes for transcription
factors (TF) were up-regulated in response to M oryzae
inoculation (see Supplementary Table S2 at JXB online)
Among the more prominently expressed are genes for NAC
domain-containing proteins (841-fold) Dof zinc finger pro-
teins (1134-fold) sigma factors (1238-fold) bZIP transcrip-
tion factors (1193-fold) and MADS box proteins (446-fold)
The up-regulation of the rest of the TF genes in transgenic
plants varied from 203-fold to 1238-fold It was interesting to
note that all these TF genes were regulated at a very low level
in the non-transgenic plants (see Supplementary Table S2 at
JXB online Fig 4B) Programmed cell death (PCD) is one of
the important responses of plants upon pathogen infection
and leads to a hypersensitive response in resistant lines In this
study four genes with predicted roles in PCD were up-
regulated up to 207ndash276-fold in transgenic lines compared
with between 067ndash159-fold in non-transgenic lines (see
Supplementary Table S3 at JXB online Fig 4B Fig 4C)
Fifteen genes associated with vital signal transduction
functions in the plants were also highly up-regulated in TP-
Pi54 (see Supplementary Table S4 at JXB online Fig 4B)
A maximum up-regulation of 1202-fold was obtained in
auxin-mediated signal pathways followed by that ingibberellic acid-mediated signalling (328-fold) Similarly
genes for two-component signal transduction pathways
were up-regulated between 244-fold and 356-fold (see
Supplementary Table S4 at JXB online Fig 4D)
Hierarchical clustering
Genome-wide analysis of the expression patterns of mock-
inoculated and pathogen-inoculated TP-Pi54 transgenicplants was performed The phylogenetic relationship be-
tween highly expressed and less expressed genes was
obtained after hierarchal clustering The expression changes
among the gene group members in six clusters (i) response
to stimulus (ii) signal transduction (iii) transcription
Fig 4 Comparison of the differential regulation of different categories of genes more up-regulated in TP-Pi54 compared to TP
(A) Stress-responsive genes 1 callose 2 laccase 3 PAL 4 Peroxidase 5 lipoxygenase 6 superoxide dismutase 7 amylase 8
licheninase 9 chitinase 10 glucosylceramidase 11 response to wounding 12 response to chemical stimulus and 13 abiotic stress
(B) 1ndash22 Transcription factor genes (C) 1ndash4 Genes for programmed cell death (D) 1ndash15 Genes for signal transduction
762 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
regulators (iv) programmed cell death (v) oxidoreductases
and (vi) lyase were represented by different colours in the
heat map The colour intensities indicate the expression
level of each gene (Fig 5)
Genes repressed in native TP lines
Out of the 107 up-regulated genes of TP-Pi54 29 were
considerably repressed in the non-transgenic TP line (Table 2)
Among these the genes Os77671S1_at (652-fold) andOs70041S1_at (2458-fold) were up-regulated at the highest
level in TP-Pi54 compared with non-transgenic lines Other
highly expressed genes in TP-Pi54 that were otherwise
repressed in TP belonged to some important categories such
as genes involved in transporter activity (2726-fold and
1692-fold) zinc ion binding (1033-fold) and lipid transport
(685-fold) Interestingly a mitochondrial located gene
(Os124301S1_at) was also highly up-regulated (15-fold) inTP-Pi54 and down-regulated in the TP lines (Table 2) Among
all the up-regulated genes in TP-54 22 were hypothetically
expressed proteins of unknown function This indicates that
these genes may be novel and may have an important role in
the Pi54-mediated defence mechanism in the blast-resistant
transgenic line TP-Pi54
Quantitative analysis of differentially regulated genes
Validation of nine up-regulated genes and one down-regulated gene in the transgenic lines was performed by
quantitative real-time PCR Genes for NAC domain-
containing proteins (NAC6) callose polysaccharide
biosynthesis proteins (CAPD) Dof zinc finger proteins
(DOF) peroxidase (POC1) MADS box proteins (MADS)
class III peroxidase (POX3) chitinase (CHIT) phenylala-
nine ammonia lyase (PAL) and endo-13-14-b-glucanase
(BG) were found to be up-regulated 124ndash58-fold whereas
genes for homeo domain-related proteins (HDR) were
down-regulated up to 14-fold (Fig 6 see Supplementary
Table S6 at JXB online Fig 4B)
Biochemical assay of enzymes involved in defenceresponse
To assess the effect of pathogen infection on plant defence
response enzymes the activities of different enzymes were
measured individually in transgenic (TP-Pi54) and non-
transgenic (TP) plants at 0 to 120 hpi (Fig 7) The data
obtained from transgenic plants were compared with those
from the non-inoculated plants used as controls M oryzae
inoculation drastically increased peroxidase activity intransgenic rice plants after 96 hpi The POX activity at 96
hpi in TP-Pi54 was higher (01623 U) than that in TP
(Fig 7A Table 3) The changes in polyphenol oxidase
activity in the plants followed more or less the same pattern
as observed for POX The PPO activity increased at 24 hpi
in transgenic plants reached its maximum at 96 hpi and
declined thereafter The PPO activity was measured as
change in A420 (DA420) mg1 protein min1 The PPOactivity at 96 h was 01736 U in TP-Pi54 compared with
01 U in TP (Fig 7B Table 3)
The time-course of PAL activity in treated plants was
quite distinct from that of the rest of the enzymes as the
profile represented a bimodal pattern in transgenic plants
indicating an early and sharp induction at 24 hpi (Fig 7C)
The decline in PAL activity continued until 72 hpi and
increased thereafter until 96 hpi before following another
dip at 120 hpi compared with that in non-inoculated
transgenic control plants which showed no induction at all
The maximum PAL activity at 24 hpi was 9305 U in TP-
Pi54 and 1768 U in TP (Fig 7C Table 3) b-Glucosidase
activity in plants gradually increased upon pathogen in-
oculation reaching its maximum at 48 hpi in the transgenic
group On the other hand a bimodal pattern of activity was
obtained in the case of non-transgenic plants showing
a slight increase in the activity at 24 and 96 hpi The activity
of b-glucosidase at 48 h was 00809 U and 00214 U in TP-
Pi54 and TP respectively (Fig 7D Table 3)
The activity of b-13-glucanase in transgenic plantswhich was already high started increasing after 24 hpi and
thereafter increased consistently up to 120 hpi Although
the b-13-glucanase activity was also higher in the trans-
genic control it did not show any significant increase after
24 hpi However in non-transgenic plants the enzyme was
slightly induced at 24 h but declined immediately until 72
hpi before reaching a normal level The highest b-1
3-glucanase activity was 8770 U in TP-Pi54 compared to727 U in TP (Fig 7E Table 3) The chitinase enzyme
concentration increased in transgenic plants at 48 hpi and
reached its maximum level after 120 hpi The maximum
chitinase activity was 1011 U in TP-Pi54 compared with
858 U in TP (Fig 7F Table 3)
Fig 5 Expression pattern as represented by the heat map profile
of mock-inoculated (control) versus TP-Pi54 (test) rice lines
inoculated with M oryzae Colour changes show the level of gene
expression in their respective clusters Yellow mildly expressed
genes blue highly suppressed genes red highly expressed
genes The clusters are genes for (A) response to stimulus (B)
signal transduction (C) transcription regulators (D) programmed
cell death (E) oxidoreductases and (F) lyase
Pi54 mediated resistance against rice blast | 763D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

regulators (iv) programmed cell death (v) oxidoreductases
and (vi) lyase were represented by different colours in the
heat map The colour intensities indicate the expression
level of each gene (Fig 5)
Genes repressed in native TP lines
Out of the 107 up-regulated genes of TP-Pi54 29 were
considerably repressed in the non-transgenic TP line (Table 2)
Among these the genes Os77671S1_at (652-fold) andOs70041S1_at (2458-fold) were up-regulated at the highest
level in TP-Pi54 compared with non-transgenic lines Other
highly expressed genes in TP-Pi54 that were otherwise
repressed in TP belonged to some important categories such
as genes involved in transporter activity (2726-fold and
1692-fold) zinc ion binding (1033-fold) and lipid transport
(685-fold) Interestingly a mitochondrial located gene
(Os124301S1_at) was also highly up-regulated (15-fold) inTP-Pi54 and down-regulated in the TP lines (Table 2) Among
all the up-regulated genes in TP-54 22 were hypothetically
expressed proteins of unknown function This indicates that
these genes may be novel and may have an important role in
the Pi54-mediated defence mechanism in the blast-resistant
transgenic line TP-Pi54
Quantitative analysis of differentially regulated genes
Validation of nine up-regulated genes and one down-regulated gene in the transgenic lines was performed by
quantitative real-time PCR Genes for NAC domain-
containing proteins (NAC6) callose polysaccharide
biosynthesis proteins (CAPD) Dof zinc finger proteins
(DOF) peroxidase (POC1) MADS box proteins (MADS)
class III peroxidase (POX3) chitinase (CHIT) phenylala-
nine ammonia lyase (PAL) and endo-13-14-b-glucanase
(BG) were found to be up-regulated 124ndash58-fold whereas
genes for homeo domain-related proteins (HDR) were
down-regulated up to 14-fold (Fig 6 see Supplementary
Table S6 at JXB online Fig 4B)
Biochemical assay of enzymes involved in defenceresponse
To assess the effect of pathogen infection on plant defence
response enzymes the activities of different enzymes were
measured individually in transgenic (TP-Pi54) and non-
transgenic (TP) plants at 0 to 120 hpi (Fig 7) The data
obtained from transgenic plants were compared with those
from the non-inoculated plants used as controls M oryzae
inoculation drastically increased peroxidase activity intransgenic rice plants after 96 hpi The POX activity at 96
hpi in TP-Pi54 was higher (01623 U) than that in TP
(Fig 7A Table 3) The changes in polyphenol oxidase
activity in the plants followed more or less the same pattern
as observed for POX The PPO activity increased at 24 hpi
in transgenic plants reached its maximum at 96 hpi and
declined thereafter The PPO activity was measured as
change in A420 (DA420) mg1 protein min1 The PPOactivity at 96 h was 01736 U in TP-Pi54 compared with
01 U in TP (Fig 7B Table 3)
The time-course of PAL activity in treated plants was
quite distinct from that of the rest of the enzymes as the
profile represented a bimodal pattern in transgenic plants
indicating an early and sharp induction at 24 hpi (Fig 7C)
The decline in PAL activity continued until 72 hpi and
increased thereafter until 96 hpi before following another
dip at 120 hpi compared with that in non-inoculated
transgenic control plants which showed no induction at all
The maximum PAL activity at 24 hpi was 9305 U in TP-
Pi54 and 1768 U in TP (Fig 7C Table 3) b-Glucosidase
activity in plants gradually increased upon pathogen in-
oculation reaching its maximum at 48 hpi in the transgenic
group On the other hand a bimodal pattern of activity was
obtained in the case of non-transgenic plants showing
a slight increase in the activity at 24 and 96 hpi The activity
of b-glucosidase at 48 h was 00809 U and 00214 U in TP-
Pi54 and TP respectively (Fig 7D Table 3)
The activity of b-13-glucanase in transgenic plantswhich was already high started increasing after 24 hpi and
thereafter increased consistently up to 120 hpi Although
the b-13-glucanase activity was also higher in the trans-
genic control it did not show any significant increase after
24 hpi However in non-transgenic plants the enzyme was
slightly induced at 24 h but declined immediately until 72
hpi before reaching a normal level The highest b-1
3-glucanase activity was 8770 U in TP-Pi54 compared to727 U in TP (Fig 7E Table 3) The chitinase enzyme
concentration increased in transgenic plants at 48 hpi and
reached its maximum level after 120 hpi The maximum
chitinase activity was 1011 U in TP-Pi54 compared with
858 U in TP (Fig 7F Table 3)
Fig 5 Expression pattern as represented by the heat map profile
of mock-inoculated (control) versus TP-Pi54 (test) rice lines
inoculated with M oryzae Colour changes show the level of gene
expression in their respective clusters Yellow mildly expressed
genes blue highly suppressed genes red highly expressed
genes The clusters are genes for (A) response to stimulus (B)
signal transduction (C) transcription regulators (D) programmed
cell death (E) oxidoreductases and (F) lyase
Pi54 mediated resistance against rice blast | 763D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
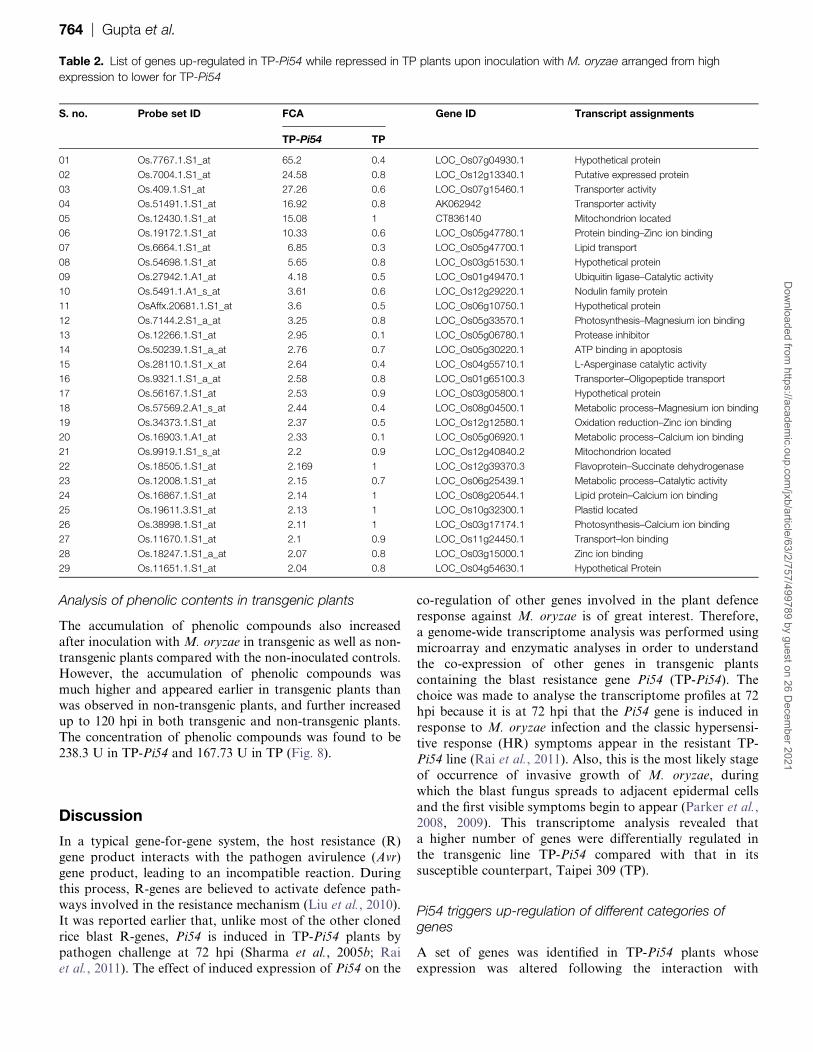
Analysis of phenolic contents in transgenic plants
The accumulation of phenolic compounds also increased
after inoculation with M oryzae in transgenic as well as non-
transgenic plants compared with the non-inoculated controls
However the accumulation of phenolic compounds was
much higher and appeared earlier in transgenic plants than
was observed in non-transgenic plants and further increasedup to 120 hpi in both transgenic and non-transgenic plants
The concentration of phenolic compounds was found to be
2383 U in TP-Pi54 and 16773 U in TP (Fig 8)
Discussion
In a typical gene-for-gene system the host resistance (R)
gene product interacts with the pathogen avirulence (Avr)
gene product leading to an incompatible reaction During
this process R-genes are believed to activate defence path-ways involved in the resistance mechanism (Liu et al 2010)
It was reported earlier that unlike most of the other cloned
rice blast R-genes Pi54 is induced in TP-Pi54 plants by
pathogen challenge at 72 hpi (Sharma et al 2005b Rai
et al 2011) The effect of induced expression of Pi54 on the
co-regulation of other genes involved in the plant defence
response against M oryzae is of great interest Therefore
a genome-wide transcriptome analysis was performed using
microarray and enzymatic analyses in order to understand
the co-expression of other genes in transgenic plants
containing the blast resistance gene Pi54 (TP-Pi54) The
choice was made to analyse the transcriptome profiles at 72hpi because it is at 72 hpi that the Pi54 gene is induced in
response to M oryzae infection and the classic hypersensi-
tive response (HR) symptoms appear in the resistant TP-
Pi54 line (Rai et al 2011) Also this is the most likely stage
of occurrence of invasive growth of M oryzae during
which the blast fungus spreads to adjacent epidermal cells
and the first visible symptoms begin to appear (Parker et al
2008 2009) This transcriptome analysis revealed thata higher number of genes were differentially regulated in
the transgenic line TP-Pi54 compared with that in its
susceptible counterpart Taipei 309 (TP)
Pi54 triggers up-regulation of different categories ofgenes
A set of genes was identified in TP-Pi54 plants whose
expression was altered following the interaction with
Table 2 List of genes up-regulated in TP-Pi54 while repressed in TP plants upon inoculation with M oryzae arranged from high
expression to lower for TP-Pi54
S no Probe set ID FCA Gene ID Transcript assignments
TP-Pi54 TP
01 Os77671S1_at 652 04 LOC_Os07g049301 Hypothetical protein
02 Os70041S1_at 2458 08 LOC_Os12g133401 Putative expressed protein
03 Os4091S1_at 2726 06 LOC_Os07g154601 Transporter activity
04 Os514911S1_at 1692 08 AK062942 Transporter activity
05 Os124301S1_at 1508 1 CT836140 Mitochondrion located
06 Os191721S1_at 1033 06 LOC_Os05g477801 Protein bindingndashZinc ion binding
07 Os66641S1_at 685 03 LOC_Os05g477001 Lipid transport
08 Os546981S1_at 565 08 LOC_Os03g515301 Hypothetical protein
09 Os279421A1_at 418 05 LOC_Os01g494701 Ubiquitin ligasendashCatalytic activity
10 Os54911A1_s_at 361 06 LOC_Os12g292201 Nodulin family protein
11 OsAffx206811S1_at 36 05 LOC_Os06g107501 Hypothetical protein
12 Os71442S1_a_at 325 08 LOC_Os05g335701 PhotosynthesisndashMagnesium ion binding
13 Os122661S1_at 295 01 LOC_Os05g067801 Protease inhibitor
14 Os502391S1_a_at 276 07 LOC_Os05g302201 ATP binding in apoptosis
15 Os281101S1_x_at 264 04 LOC_Os04g557101 L-Asperginase catalytic activity
16 Os93211S1_a_at 258 08 LOC_Os01g651003 TransporterndashOligopeptide transport
17 Os561671S1_at 253 09 LOC_Os03g058001 Hypothetical protein
18 Os575692A1_s_at 244 04 LOC_Os08g045001 Metabolic processndashMagnesium ion binding
19 Os343731S1_at 237 05 LOC_Os12g125801 Oxidation reductionndashZinc ion binding
20 Os169031A1_at 233 01 LOC_Os05g069201 Metabolic processndashCalcium ion binding
21 Os99191S1_s_at 22 09 LOC_Os12g408402 Mitochondrion located
22 Os185051S1_at 2169 1 LOC_Os12g393703 FlavoproteinndashSuccinate dehydrogenase
23 Os120081S1_at 215 07 LOC_Os06g254391 Metabolic processndashCatalytic activity
24 Os168671S1_at 214 1 LOC_Os08g205441 Lipid proteinndashCalcium ion binding
25 Os196113S1_at 213 1 LOC_Os10g323001 Plastid located
26 Os389981S1_at 211 1 LOC_Os03g171741 PhotosynthesisndashCalcium ion binding
27 Os116701S1_at 21 09 LOC_Os11g244501 TransportndashIon binding
28 Os182471S1_a_at 207 08 LOC_Os03g150001 Zinc ion binding
29 Os116511S1_at 204 08 LOC_Os04g546301 Hypothetical Protein
764 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
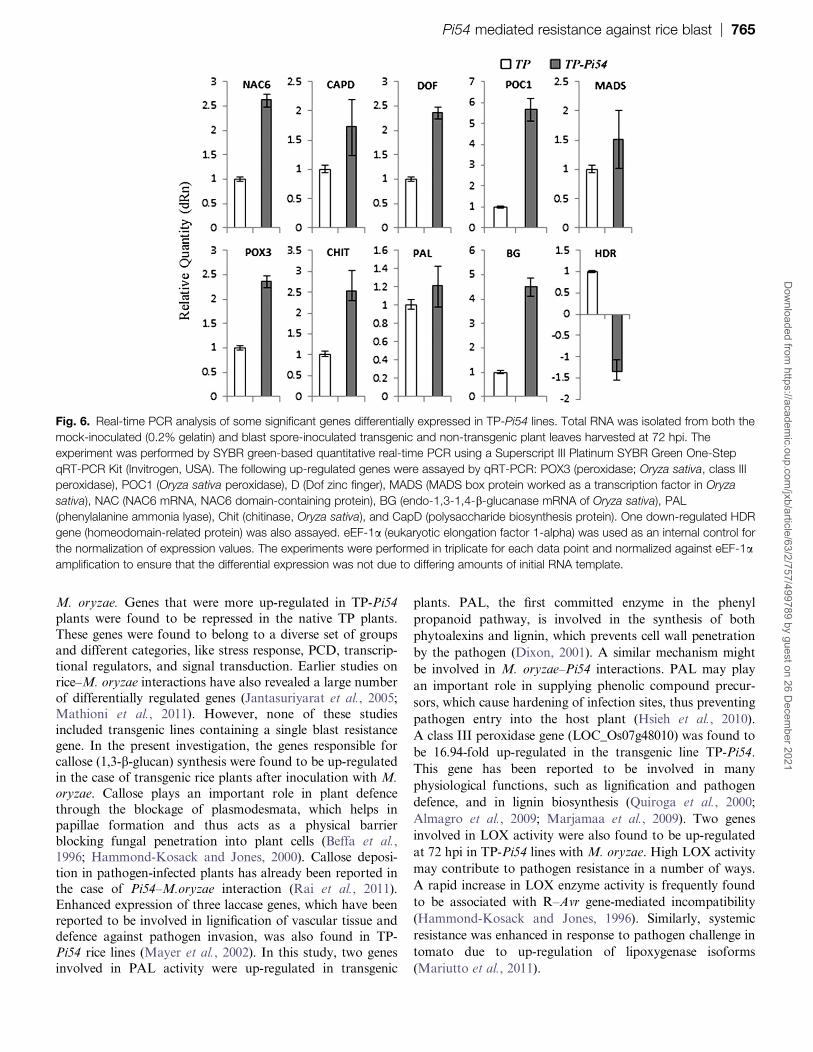
M oryzae Genes that were more up-regulated in TP-Pi54
plants were found to be repressed in the native TP plants
These genes were found to belong to a diverse set of groups
and different categories like stress response PCD transcrip-
tional regulators and signal transduction Earlier studies on
ricendashM oryzae interactions have also revealed a large number
of differentially regulated genes (Jantasuriyarat et al 2005
Mathioni et al 2011) However none of these studies
included transgenic lines containing a single blast resistancegene In the present investigation the genes responsible for
callose (13-b-glucan) synthesis were found to be up-regulated
in the case of transgenic rice plants after inoculation with M
oryzae Callose plays an important role in plant defence
through the blockage of plasmodesmata which helps in
papillae formation and thus acts as a physical barrier
blocking fungal penetration into plant cells (Beffa et al
1996 Hammond-Kosack and Jones 2000) Callose deposi-
tion in pathogen-infected plants has already been reported inthe case of Pi54ndashMoryzae interaction (Rai et al 2011)
Enhanced expression of three laccase genes which have been
reported to be involved in lignification of vascular tissue and
defence against pathogen invasion was also found in TP-
Pi54 rice lines (Mayer et al 2002) In this study two genes
involved in PAL activity were up-regulated in transgenic
plants PAL the first committed enzyme in the phenyl
propanoid pathway is involved in the synthesis of both
phytoalexins and lignin which prevents cell wall penetration
by the pathogen (Dixon 2001) A similar mechanism might
be involved in M oryzaendashPi54 interactions PAL may play
an important role in supplying phenolic compound precur-
sors which cause hardening of infection sites thus preventing
pathogen entry into the host plant (Hsieh et al 2010)
A class III peroxidase gene (LOC_Os07g48010) was found to
be 1694-fold up-regulated in the transgenic line TP-Pi54
This gene has been reported to be involved in many
physiological functions such as lignification and pathogen
defence and in lignin biosynthesis (Quiroga et al 2000
Almagro et al 2009 Marjamaa et al 2009) Two genes
involved in LOX activity were also found to be up-regulated
at 72 hpi in TP-Pi54 lines with M oryzae High LOX activity
may contribute to pathogen resistance in a number of ways
A rapid increase in LOX enzyme activity is frequently found
to be associated with RndashAvr gene-mediated incompatibility
(Hammond-Kosack and Jones 1996) Similarly systemic
resistance was enhanced in response to pathogen challenge in
tomato due to up-regulation of lipoxygenase isoforms
(Mariutto et al 2011)
Fig 6 Real-time PCR analysis of some significant genes differentially expressed in TP-Pi54 lines Total RNA was isolated from both the
mock-inoculated (02 gelatin) and blast spore-inoculated transgenic and non-transgenic plant leaves harvested at 72 hpi The
experiment was performed by SYBR green-based quantitative real-time PCR using a Superscript III Platinum SYBR Green One-Step
qRT-PCR Kit (Invitrogen USA) The following up-regulated genes were assayed by qRT-PCR POX3 (peroxidase Oryza sativa class III
peroxidase) POC1 (Oryza sativa peroxidase) D (Dof zinc finger) MADS (MADS box protein worked as a transcription factor in Oryza
sativa) NAC (NAC6 mRNA NAC6 domain-containing protein) BG (endo-13-14-b-glucanase mRNA of Oryza sativa) PAL
(phenylalanine ammonia lyase) Chit (chitinase Oryza sativa) and CapD (polysaccharide biosynthesis protein) One down-regulated HDR
gene (homeodomain-related protein) was also assayed eEF-1a (eukaryotic elongation factor 1-alpha) was used as an internal control for
the normalization of expression values The experiments were performed in triplicate for each data point and normalized against eEF-1aamplification to ensure that the differential expression was not due to differing amounts of initial RNA template
Pi54 mediated resistance against rice blast | 765D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

Plant pathogens tend to diminish the sugar levels of thehost resulting in the induction of sugar-cleaving enzymes
like sucrose synthase invertase and amylases (Machenaud
et al 2007) As evidenced from earlier studies b-amylases
also contributed in redox-regulated starch degradation and
play a direct role in signal transduction by regulating sugar
transport in pathogen-challenged or wound-induced stress-
ful conditions (Gupta et al 2010) Five genes involved in
b-amylase activity and two genes in a-amylase activity werefound to be up-regulated upon M oryzae infection in TP-
Pi54 plants thus they might play an important role insignal transduction pathways
The up-regulation was found of 14 genes for pathogene-
sis-related (PR) proteins which are important constituents
of the defence response mechanism It has already been
reported that PR genes encoding chitinases are up-regulated
in transgenic plants and show significant improvement in
disease resistance (Gonzalez-Candelas et al 2010 Chen
et al 2010) Chitinase genes are synergistically inducedduring infection by fungal pathogens and fungal elicitors
The role of chitinases in the plant defence against fungal
pathogens is very well studied (Kasprzewska 2003) and also
supports our results Two genes for glucosylceramidase
which plays an important role in the fungal defence
response by degrading the cell wall component (b-1
6-glucan) were also found to be up-regulated in TP-Pi54
plants A host of genes in response to wounding andchemical stimulus were up-regulated in the TP-Pi54 line
supporting the notion of crosstalk between abiotic and
biotic stress responses in disease-resistant plants (Fujita
et al 2006)
Transcriptional activation of genes is a vital component
of the plant defence system against pathogens Changes in
the expression patterns of pathogen-responsive genes are
mostly influenced by native promoters Transcription
Fig 7 The activities of different enzymes as measured in the crude extract prepared from leaves harvested at various stages after
M oryzae inoculation in TP-Pi54 rice lines were compared with those in non-transgenic (TP) lines and their respective mock-inoculated
controls (A) Peroxidase (B) polyphenol oxidase (C) phenylalanine ammonia lyase (D) b-glucosidase (E) b-13-glucanase and (F)
chitinase Bars indicate the standard error at P-valuefrac14005
Table 3 Summary of maximum enzymatic activity observed upon
inoculation with M oryzae U Enzyme activity unit of respective
enzymes hpi hours post-inoculation TP-Pi54 transgenic TP
non-transgenic
Enzyme Activity inTP-Pi54 (U)
Activity inTP (U)
hpi D Foldchange
POX 184 126 96 058
PPO 263 147 96 116
PAL 34 19 24 15
b-glucosidase 266 108 48 108
b13glucanase 139 127 120 012
Chitinase 172 16 120 01
766 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
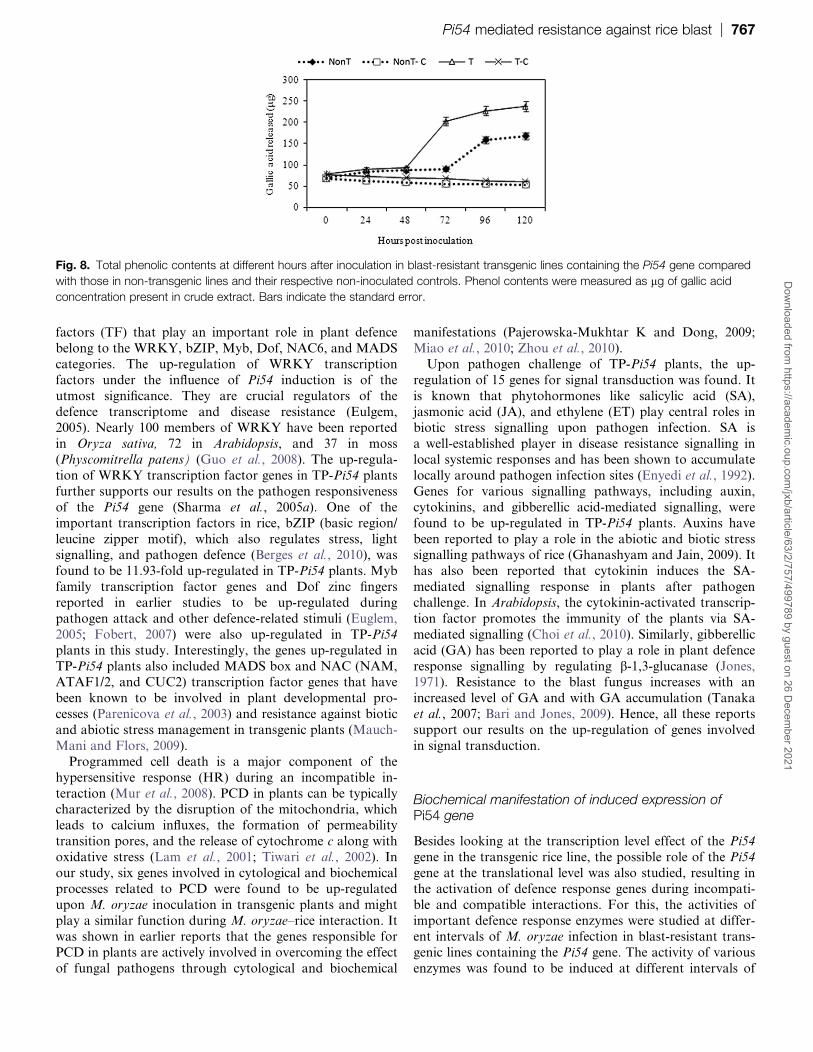
factors (TF) that play an important role in plant defence
belong to the WRKY bZIP Myb Dof NAC6 and MADS
categories The up-regulation of WRKY transcription
factors under the influence of Pi54 induction is of the
utmost significance They are crucial regulators of the
defence transcriptome and disease resistance (Eulgem2005) Nearly 100 members of WRKY have been reported
in Oryza sativa 72 in Arabidopsis and 37 in moss
(Physcomitrella patens) (Guo et al 2008) The up-regula-
tion of WRKY transcription factor genes in TP-Pi54 plants
further supports our results on the pathogen responsiveness
of the Pi54 gene (Sharma et al 2005a) One of the
important transcription factors in rice bZIP (basic region
leucine zipper motif) which also regulates stress lightsignalling and pathogen defence (Berges et al 2010) was
found to be 1193-fold up-regulated in TP-Pi54 plants Myb
family transcription factor genes and Dof zinc fingers
reported in earlier studies to be up-regulated during
pathogen attack and other defence-related stimuli (Euglem
2005 Fobert 2007) were also up-regulated in TP-Pi54
plants in this study Interestingly the genes up-regulated in
TP-Pi54 plants also included MADS box and NAC (NAMATAF12 and CUC2) transcription factor genes that have
been known to be involved in plant developmental pro-
cesses (Parenicova et al 2003) and resistance against biotic
and abiotic stress management in transgenic plants (Mauch-
Mani and Flors 2009)
Programmed cell death is a major component of the
hypersensitive response (HR) during an incompatible in-
teraction (Mur et al 2008) PCD in plants can be typicallycharacterized by the disruption of the mitochondria which
leads to calcium influxes the formation of permeability
transition pores and the release of cytochrome c along with
oxidative stress (Lam et al 2001 Tiwari et al 2002) In
our study six genes involved in cytological and biochemical
processes related to PCD were found to be up-regulated
upon M oryzae inoculation in transgenic plants and might
play a similar function during M oryzaendashrice interaction Itwas shown in earlier reports that the genes responsible for
PCD in plants are actively involved in overcoming the effect
of fungal pathogens through cytological and biochemical
manifestations (Pajerowska-Mukhtar K and Dong 2009
Miao et al 2010 Zhou et al 2010)
Upon pathogen challenge of TP-Pi54 plants the up-
regulation of 15 genes for signal transduction was found It
is known that phytohormones like salicylic acid (SA)
jasmonic acid (JA) and ethylene (ET) play central roles inbiotic stress signalling upon pathogen infection SA is
a well-established player in disease resistance signalling in
local systemic responses and has been shown to accumulate
locally around pathogen infection sites (Enyedi et al 1992)
Genes for various signalling pathways including auxin
cytokinins and gibberellic acid-mediated signalling were
found to be up-regulated in TP-Pi54 plants Auxins have
been reported to play a role in the abiotic and biotic stresssignalling pathways of rice (Ghanashyam and Jain 2009) It
has also been reported that cytokinin induces the SA-
mediated signalling response in plants after pathogen
challenge In Arabidopsis the cytokinin-activated transcrip-
tion factor promotes the immunity of the plants via SA-
mediated signalling (Choi et al 2010) Similarly gibberellic
acid (GA) has been reported to play a role in plant defence
response signalling by regulating b-13-glucanase (Jones1971) Resistance to the blast fungus increases with an
increased level of GA and with GA accumulation (Tanaka
et al 2007 Bari and Jones 2009) Hence all these reports
support our results on the up-regulation of genes involved
in signal transduction
Biochemical manifestation of induced expression ofPi54 gene
Besides looking at the transcription level effect of the Pi54
gene in the transgenic rice line the possible role of the Pi54
gene at the translational level was also studied resulting in
the activation of defence response genes during incompati-
ble and compatible interactions For this the activities of
important defence response enzymes were studied at differ-ent intervals of M oryzae infection in blast-resistant trans-
genic lines containing the Pi54 gene The activity of various
enzymes was found to be induced at different intervals of
Fig 8 Total phenolic contents at different hours after inoculation in blast-resistant transgenic lines containing the Pi54 gene compared
with those in non-transgenic lines and their respective non-inoculated controls Phenol contents were measured as lg of gallic acid
concentration present in crude extract Bars indicate the standard error
Pi54 mediated resistance against rice blast | 767D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

pathogen infection in resistant transgenic and susceptible
non-transgenic rice lines
The pattern of POX and PPO activities in transgenic
plants showed significant induction after pathogen inocula-
tion Both enzymes showed maximum activity at 96 hpi in
TP-Pi54 plants compared witho susceptible TP plants Plant
peroxidases catalyse the oxidation of various reductants and
play a key role in several metabolic responses Theseenzymes are basically involved in auxin metabolism lignin
and suberin formation cross-linking of cell wall compo-
nents phytoalexin synthesis and the metabolism of ROX
(reactive oxygen species) and RNS (reactive nitrogen
species) (Almagro et al 2009) The activities of these
enzymes were found to be altered in tomato and groundnut
upon pathogen challenge (Vanitha et al 2009 Usha Rani
and Jyothsna 2009) In earlier reports the activity of POX
was significantly induced in rice leaves after inoculation
with M oryzae (Sekizawa et al 1990) These findings
support the results obtained in our present investigationSimilar to its high-level transcriptional activation in
microarray studies the PAL enzyme was produced at
a significantly higher concentration in Pi54 transgenic
Fig 9 Schematic representation of the activation of a defence response by the Pi54 gene induced by M oryzae inoculation Genome-
wide expression analysis and enzymatic analysis of defence response genes indicate that Pi54 regulates the regulators of the defence
response against rice blast pathogens This gene plays a major role in the whole defence mechanism by its activation of the metabolic
pathway related to the phenyl propanoid mechanism The enzymes that showed high activities in transgenic lines are the core enzymes
of the phenyl propanoid pathway the first pathway for flavonoid lignin and soluble phenolic biosynthesis The genes activated in the co-
expression analysis of Pi54 are shown with their transcript assignment These are PAL (LOC_Os04g437601) glucanase
(LOC_Os02g585601) chitinase (LOC_Os11g379601) lipoxygenase (LOC_Os12g372601) peroxidases (LOC_Os03g132101) laccase
(LOC_Os03g166101) DOF (LOC_Os02g15350) and WRKY (LOC_Os01g087101) The pathway components that are affected in
a major way are shaded in pink while those affected partially are shaded in light blue The pink triangles are effector proteins secreted by
germinating M oryzae spores ( ) penetrating cell wall Abbreviations ROS reactive oxygen species NO nitric oxide RLKs receptor-
like kinases PCD programmed cell death PAL phenylalanine ammonia lyase Phe phenylalanine CA cinnamic acid SA salicylic acid
Glc glucanases Chit chitinases LPO lipoxygenases POX peroxidases Lac laccases ETI effector-triggered immunity PAMP
pathogen-associated molecular patterns
768 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

plants after 24 hpi The fold change in PAL activity between
transgenic and non-transgenic lines was 15 which indicates
a significant induction of this enzyme upon pathogen
inoculation Earlier reports related to the induction of PAL
during the early hours of pathogen infection also support
the results of this study (Vanitha et al 2009 Campos-
Vargas et al 2004) The activity profile of the b-glucosidase
enzyme in TP-Pi54 plants was found to be higher than inTP plants at 48 hpi Like PAL b-glucosidase is also a key
enzyme in phytoalexin synthesis and might play a crucial
role in Pi54-mediated rice blast resistance in TP-Pi54 plants
This enzyme also plays important roles in many biotrans-
formation processes Plant b-glucosidases may be involved
in the processing and release of fungal glucan elicitors
triggering a chain of reactions in the host including
phytoalexin formation and the biosynthesis of phenylpropanoids and lignin-like phenol aglucones by hydrolysing
b-phenyl glucosides These aglucones are basically fungi-
toxic and fungistatic in action (Whetten et al 1998) and
may limit the spread of M oryzae in these resistant plants
Chitinase and b-13-glucanase enzyme activities were
significantly higher after pathogen challenge in TP-Pi54
plants at 120 hpi but were significantly lower in TP plants
These enzymes play an important role in the degradation offungal cell walls by hydrolysing polymers where fragments
of the cell wall are shown to induce a defence reaction in
plants (Shunja et al 2007) Earlier reports support the
finding of the current study that the induction of b-13-
glucanase upon pathogen challenge was highest at 120 hpi
(De Bolle et al 1996) Similarly increased induction in the
activity profile of chitinases in transgenic potato plants was
able to overcome the damage caused by plant pathogens(Hou et al 1998)
Many plant phenolic compounds are antimicrobial in
nature function as precursors of various structural poly-
mers such as lignin or serve as signal molecules Our study
revealed that TP-Pi54ndashM oryzae interaction results in the
higher accumulation of total phenolics compared with the
susceptible check Our analysis showed that a battery of
defence enzymes might be involved in the resistance re-action of TP-Pi54 plants against M oryzae
Pi54-mediated resistance development
The blast resistance gene Pi54 is a major resistance gene
that has already been cloned and characterized (Sharma
et al 2005b Rai et al 2011) The main purpose of this
study was to understand the functions of the co-expressedgenes involved in Pi54-mediated blast resistance in rice
Hence it is shown that M oryzaendashTP-Pi54 interaction
alters the expression level of many genes involved in the
defence response pathway due to induced expression of the
NBS-LRR gene Pi54 at 72 hpi Like other NBS-LRR genes
this gene follows the effector-triggered immunity-based
defence response and induces higher expression of some
key transcription factors and signalling and enzymaticgenes that may be involved in RndashAvr interactions (Jones
and Dangl 2006) However this was not evident in the case
of M oryzaendashTP (susceptible) interactions as TP plants
were devoid of the functional Pi54 gene The up-regulation
of genes for signal transduction and transcription factors in
tandem resulted in elevated expression levels of various key
defence response enzymes that are known to be involved in
the synthesis of vital secondary metabolites like phyto-
alexin lignin and phenolic compounds identified in various
compatible hostndashpathogen interactions Stress-responsivegenes like glucanase chitinase lipoxygenase peroxidases
and laccase which were found to be up-regulated in our
study play crucial roles in the metabolic pathways related
to the phenyl propanoid mechanism Expectedly the bio-
chemical activity of various defence response enzymes
considered to be the core of the phenyl propanoid pathway
was found to be highly expressed in the blast-resistant TP-
Pi54 plants and might be involved in the biosynthesis offlavonoid lignin and soluble phenols Thus in this way
Pi54 plays a central role in receiving signals from M oryzae
avirulence genes and activating downstream defence re-
sponse genes in the transgenic line In the process Pi54
protein is assumed to regulate the regulators of the defence
response leading to incompatible interactions and finally
into resistant phenotypes
The Pi54 gene is widely used in rice-breeding programmesacross India for the development of locally suited blast-
resistant commercial rice varieties The results of the tran-
scriptome and biochemical analyses in the present study
indicate an active role for the Pi54 gene in transgenic rice
lines following interactions with the avirulent strain of
M oryzae This interaction confirms that a single functional
NBS-LRR gene Pi54 activates a cascade of defence
response genes in transgenic rice lines leading to incompat-ible interactions (Fig 9) This study presents a detailed
analysis of blast-resistant transgenic lines performed using
microarray-based transcriptomes The involvement of de-
fence response enzymes has also been studied to support
our findings The study also provides novel insights that
would be helpful in deciphering the molecular mechanisms
involved in M oryzaendashPi54 interactions
Supplementary data
Supplementary date can be found at JXB online
Supplementary Table S1 Functional annotation andcategorization of stress-associated up-regulated genes in
TP-Pi54 following incompatible interaction with M oryzae
Supplementary Table S2 Differentially regulated tran-
scription factor genes in the TP-Pi54 and TP plants infected
with M oryzae
Supplementary Table S3 Differentially regulated pro-
grammed cell death (PCD)-associated genes in the TP-Pi54
and TP plants inoculated with M oryzaeSupplementary Table S4 Signal transduction-associated
genes differentially regulated in the TP-Pi54 and TP plants
inoculated with M oryzae
Supplementary Table S5 Primers used in qRT-PCR
validation of gene expression analysis
Pi54 mediated resistance against rice blast | 769D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

Supplementary Table S6 Up-regulated novel genes
identified in TP-Pi54 plants and their position in TP plants
Acknowledgements
The financial assistance received (FNoBTABFG-2 (PH-
II)2009-3A) from the Department of Biotechnology Gov-ernment of India by TR Sharma is gratefully acknowl-
edged The authors are thankful to Dr KV Prabhu Officer
in Charge National Phytotron Facility Indian Agricultural
Research Institute New Delhi for providing basic facilities
for growing transgenic plants to HC Upreti for helping to
maintain the transgenic lines and to Neha Jain Bagish
Mishra and G Dandapani for their valuable help and
suggestions in carrying out the microarray experimentsSKG and AKR are thankful to Mr Niraj Kumar of
Xplorigen Technologies New Delhi for his help in making
graphics
References
Abeles FB Forrence LE 1970 Temporal and hormonal control of
b-13-glucanase in Phaseolus vulgaris L Plant Physiology 45
395ndash400
Agrawal KML Bahl OP 1969 Glycosidases of Phaseolus vulgaris
Journal of Biological Chemistry 243 103ndash111
Almagro L Ros LVG Navarro SB Bru R Barcelo AR
Pedreno MA 2009 Class III peroxidases in plant defense reactions
Journal of Experimental Botany 60 377ndash390
Bari R Jones JDG 2009 Role of plant hormones in plant defence
responses Plant Molecular Biology 69 473ndash488
Beffa RS Hofer RM Thomas M Meins Jr F 1996 Decreased
susceptibility to virus disease of b-13-glucanase-deficient plants
generated by antisense transformation The Plant Cell 8 1001ndash1011
Berges ML Rispail N Rosales RCP Pietro AD 2010 A nitrogen
response pathway regulates virulence functions in Fusarium
oxysporum via the protein kinase TOR and the bZIP protein MeaB The
Plant Cell 22 2459ndash2475
Bradford MM 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
proteinndashdye binding Analytical Biochemistry 72 248ndash254
Bryan GT Wu KS Farrall L Jia Y Hershey HP McAdams SA
Faulk KN Donaldson GK Tarchini R Valent B 2000 A single
amino acid difference distinguishes resistant and susceptible alleles of
the rice blast resistance gene Pi-ta The Plant Cell 12 2033ndash2045
Campos-Vargas R Nonogaki H Suslow T Saltveit ME 2004
Isolation and characterization of a wound inducible phenylalanine
ammonia-lyase gene (LsPAL1) from Romaine lettuce leaves Plant
Physiology 121 429ndash438
Chen X Niks RE Hedley PE Morris J Druka A Marcel TC
Vels A Waugh R 2010 Differential gene expression in nearly
isogenic lines with QTL for partial resistance to Puccinia hordei in
barley BMC Genomics 1011861471-2164-11-629
Choi J Huh SU Kojima M Sakakibara H Paek KH Hawang I
2010 The cytokinin activated transcription factor ARR2 promotes
plant immunity via TGA3NPR1 dependent salicylic acid signaling in
Arabidopsis Developmental Cell 19 284ndash295
Costanzo S Jia Y 2010 Sequence variation at the rice blast
resistance gene Pi-km locus implications for the development of allele
specific markers Plant Science 178 523ndash530
Dangl JL Dietrich RA Richberg MH 1996 Death donrsquot have no
mercy cell death programs in plantndashmicrobe interactions The Plant
Cell 8 1793ndash1807
David CC 1991 The world rice economy challenges ahead In Khus
GS Toenniessen GH eds Rice biotechnology Oxford UK The Alden
Press Ltd 1ndash18
De Bolle MF Osborn RW Goderis IJ Noe L Acland D Hart CA
Torrekens S Van Leuven F Broekaert WF 1996 Antimicrobial
peptides from Mirabilis jalapa and Amaranthus caudatus expression
processing localization and biological activity in transgenic tobacco
Plant Molecular Biology 31 993ndash1008
Dixon RA 2001 Natural products and plant disease resistance
Nature 411 843ndash847
Enyedi AJ Yalpani N Silverman P Raskin I 1992 Localization
conjugation and function of salicylic acid in tobacco during the
hypersensitive reaction to tobacco mosaic virus Proceedings of the
National Academy of Sciences USA 89 2480ndash2484
Euglem T 2005 Regulation of the Arabidopsis defense
transcriptome Trends in Plant Sciences 10 1960ndash1385
Fobert PR 2007 Transcription factors regulating plant defense
responses In Varshney RK Koebner RMD eds Model plants and
crop improvement Boca Raton CRC Press 159ndash205
Fujita M Fujita Y Noutoshi Y Takahashi F Narusaka Y
Sinozaki KY Sinozaki K 2006 Crosstalk between abiotic and biotic
stress responses a current view from the points of convergence in the
stress signaling networks Current Opinion in Plant Biology 9
436ndash442
Ghanashyam C Jain M 2009 Role of auxin-responsive genes in
biotic stress responses Plant Signaling and Behavior 4 846ndash848
Gonzalez-Candelas L Alamar S Torres PS Zacarias L
Markos JF 2010 A transcriptomic approach highlights induction of
secondary metabolism in citrus fruit in response to Penicillium
digitatum infection BMC Plant Biology 1011861471-2229-10-194
Guo S Boyd J Sammynaiken R Loewen MC 2008 Identification
and characterization of a unique cysteine residue proximal to the
catalytic site of Arabidopsis thaliana carotenoid cleavage enzyme
Biochemistry and Cell Biology 86 262ndash270
Gupta S Chakraborti D Sengupta A Basu D Das S 2010
Primary metabolism of chickpea is the initial target of wound inducing
early sensed Fusarium oxysporum f sp ciceri Race I PLoS One
101371journalpone0009030
Hammond-Kosack KE Jones JDG 2000 Responses to plant
pathogens In Biochemistry and molecular biology of plants Rockville
Maryland USA American Society of Plant Physiology 1102ndash1156
Hammond-Kosack KE Jones JDG 1996 Resistance gene
dependent plant defense responses The Plant Cell 8 1773ndash1791
770 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

Hossain M 1997 Rice supply and demand in Asia a socioeconomic
and biophysical analysis In Teng PS et al Applications of systems
approaches at the farm and regional levels Vol I IRRI Dordrecht
Kluwer Academic Publishers 263ndash279
Hou WC Chen YC Lin YH 1998 Chitinase activity of sweet potato
(Ipomoea batatas [L] Lam var Tainong 57) Botanical Bulletin of
Academia Sinica 39 93ndash97
Hsieh LS Ma GT Yang CC Lee PD 2010 Cloning expression
site-directed mutagenesis and immunolocalization of phenyl ammonia-
lyase in Bambusa oldhamii Phytochemistry 71 1999ndash2009
Irizarry RA Hobbs B Collin F Beazer-Barclay YD
Antonellis KJ Scherf U Speed TP 2003 Exploration
normalization and summaries of high density oligonucleotide array
probe level data Biostatistics 4 249ndash264
Jantasuriyarat C Gowda M Haller K et al 2005 Large-scale
identification of expressed sequence tags involved in rice and rice
blast fungus interaction Plant Physiology 138 105ndash115
Jones JDG Dangl JL 2006 The plant immune system Nature 444
323ndash329
Jones RL 1971 Gibberellic acid-enhanced release of b-13-
glucanase from barley aleurone cells Plant Physiology 47 412ndash416
Kasprzewska A 2003 Plant chitinases regulation and function
Cellular and Molecular Biology Letters 8 809ndash24
Kato T Tanabe S Nishimura M et al 2009 Differential responses
of rice to inoculation with wild type and non-pathogenic mutants of
Magnaporthe oryzae Plant Molecular Biology 70 617ndash625
Kim S Ahn IP Park CH Park SG Park SY Jwa NS Lee YH
2001 Molecular characterization of the cDNA encoding an acidic
isoform of PR-1 protein in rice Molecular Cells 11 115ndash121
Lam M Oleinick NL Nieminen AL 2001 Photodynamic therapy-
induced apoptosis in epidermoid carcinoma cells Reactive oxygen
species and mitochondrial inner membrane permeabilization Journal
of Biological Chemistry 276 47379ndash47386
Lee S-K Song M-Y Seo Y-S et al 2009 Rice Pi5-mediated
resistance to Magnaporthe oryzae requires the presence of two coiled-
coil-nucleotide-binding-leucine-rich repeat genes Genetics 181
1627ndash1638
Lisker N Cohen I Chalutz E Fucus Y 1983 Fungal infections
suppress ethylene-induced phenylalanine ammonia lyase activity in
grape fruits Physiological Plant Pathology 22 331ndash338
Liu XQ Li YY Wang LY Liu XQ Wang CT Wang L Pan QH
2010 The effect of rice blast resistant gene Pi36 on expression of
disease resistance-related genes Chinese Science Bulletin 55
1881ndash1888
Livak KJ Schmittgen TD 2001 Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T))
method Methods 25 402ndash408
Maclean JL Dawe DC Hardy B Hettel GP 2002 Rice almanac
Source book for the most important activity on earth Los Banos
Philippines International Rice Research Institute West Africa Rice
Development Association International Center for Tropical Agriculture
and Food and Agriculture Organization
Machenaud J Henri R Dieuaide-Noubhani M Pracros P
Renaudin J Eveillard S 2007 Gene expression and enzymatic
activity of invertases and sucrose synthase in Spiroplasma citri or
Stolbur phytoplasma infected plants Bulletin of Insectology 60
219ndash220
Mariutto M Duby F Adam A Bureau C Fauconnier ML
Ongena M Thonart P Dommes J 2011 The elicitation of
a systemic resistance by Pseudomonas putida BPT1 in tomato
involves the stimulation of two Lipoxygenase isoforms BMC Plant
Biology 1011861471-2229-11-29
Marjamaa K Kukkola EM Fagerstedt KV 2009 The role of xylem
class III peroxidases in lignification Journal of Experimental Botany 60
367ndash376
Mathioni SM Belo A Rizzo CJ Dean RA Donofrio NM 2011
Transcriptome profiling of the rice blast fungus during invasive plant
infection and in vitro stresses BMC Genomics 1011861471-2164-
12-49
Mauch-Mani B Flors V 2009 The ATAF1 transcription factor at the
convergence point of ABA-dependent plant defense against biotic and
abiotic stresses Cell Research 19 1322ndash1323
Mayer AM Harel E Shaul RB 1965 Assay of catechol oxidase
a critical comparison of methods Phytochemistry 5 783ndash789
Mayer RT Inbar M McKenzie CL Shatters R Borowicz V
Albrecht U Powell CA Doostdar H 2002 Multitrophic interactions
of the silverleaf whitefly host plants competing herbivores and
phytopathogens Archives of Insect Biochemistry and Physiology 51
151ndash169
Miao WG Wang XB Li M Song CF Wang Y Hu DW Wang JS
2010 Genetic transformation of cotton with a harpin-encoding gene
hpaXoo confers an enhanced defense response against different
pathogens through a priming mechanism BMC Plant Biology
1011861471-2229-10-67
Miller GL 1959 Use of dinitrosalicylic acid reagent for determination
of reducing sugar Analytical Chemistry 31 426ndash428
Mur LA Kenton P Lloyd AJ Ougham H Prats E 2008 The
hypersensitive response the centenary is upon us but how much do
we know Journal of Experimental Botany 59 501ndash520
Okuyama Y Kanzaki H Abe A et al 2011 A multifaceted
genomics approach allows the isolation of the rice Pia blast resistance
gene consisting of two adjacent NBS-LRR protein genes The Plant
Journal 66 467ndash479
Padmanabhan SY Mathur SC Misra RK 1974 Breeding for blast
resistance in India genetics of blast resistance Indian Journal of
Genetics and Plant Breeding 34 424ndash429
Parker D Beckmann M Enot DP Overy DP Rios ZC Gilbert M
Talbot N Draper P 2008 Rice blast infection of Brachypodium
distachyon as a model system to study dynamic hostpathogen
interactions Nature Protocols 3 435ndash445
Parker D Beckmann M Zubair H Enot DP Caracuel-Rios Z
Overy DP Snowdon S Talbot NJ Draper J 2009 Metabolomic
analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea
The Plant Journal 59 723ndash737
Pajerowska-Mukhtar K Dong X 2009 A kiss of death
proteasome-mediated membrane fusion and programmed cell death
Pi54 mediated resistance against rice blast | 771D
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021
in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021

in plant defense against bacterial infection Genes and Development
23 2449ndash2454
Parenicova L Folter SD Kieffer M et al 2003 Molecular and
phylogenetic analysis of the complete MADS-BOX transcription factor
family in Arabidopsis new openings to the MADS world The Plant Cell
15 1538ndash1551
Quiroga C Botella MA Barcelo A Amaya I Medina MI
Alonso F De Forchetti SM Tigiert H Valpuesta V 2000
A tomato peroxidase involved in the synthesis of lignin and suberin
Plant Physiology 122 1119ndash1127
Rai AK Kumar SP Gupta SK Gautam N Singh NK Sharma TR
2011 Functional complementation of rice blast resistance gene Pi-kh
(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae
Journal of Plant Biochemistry and Biotechnology 20 55ndash65
Rao ZM Dong HT Zhuang JY Chai RY Fan YY Li DB
Zheng KL 2002 Analysis of gene expression profiles during hostndash
Magnaporthe grisea interactions in a pair of near isogenic lines of rice
Yi Chuan Xue Bao 29 887ndash893
Ryals J Neuenschwander UH Willits MG Molina A Steiner H-Y
Hunt MD 1996 Systemic acquired resistance The Plant Cell 8
1809ndash1819
Sadasivam S Manickam A 2008 Enzymes and phenolics In
Biochemical methods New Delhi India New Age International (P)
Limited Publishers 91ndash211
Sana TR Fischer S Wholgemuth G Kalrekar A Jung K
Ronald PC Fiehn O 2010 Metabolomic and transcriptomic analysis
of the rice response to the bacterial blight pathogen Xanthomonas
oryzae pv oryzae Metabolomics 6 451ndash465
Sekizawa Y Haruyama T Kano H Urushizaki S Saka H
Matsumoto K Haga M 1990 Dependence on ethylene of the
induction of peroxidase and lipoxygenase activity in rice leaf infected
with blast fungus Agricultural and Biological Chemistry 54 471ndash478
Sharma TR Shanker P Singh BK Jana TK Madhav MS
Gaikwad K Singh NK Plaha P Rathour R 2005a Molecular
mapping of rice blast resistance gene Pi-kh in rice variety Tetep
Journal of Plant Biochemistry and Biotechnology 14 127ndash133
Sharma TR Madhav MS Singh BK et al 2005b High resolution
mapping and molecular characterization of the Pi-kh gene of rice
which confers resistance to Magnaporthe grisea Molecular Genetics
and Genomics 274 569ndash578
Sharma TR Rai AK Gupta SK Singh NK 2010 Broad spectrum
blast resistance gene Pi-kh designated as Pi-54 Journal of Plant
Biochemistry and Biotechnology 19 987ndash989
Shunja TK Hanai I Galis K Suzuki K Matsuoka H Soite M
2007 Characterization of Nt chit IV a class IV chitinase induced by
b-1316-glucan elicitor from Alternaria alternate 102 antagonistic
effect of salicylic acid methyl jasmonate on the induction of Nt chit IV
Biochemical and Biophysical Research Communication 353 311ndash317
Singh AK Gopalakrishnan S Singh VP et al 2011 Marker
assisted selection a paradigm shift in Basmati breeding Indian
Journal of Genetics and Plant Breeding 71 1ndash9
Singleton VL Rossi JA 1965 Colorimetry of total phenolics with
phosphomolybdic phosphotungstic acid reagents Americal Journal of
Enology and Viticulture 16 144ndash158
Soderlund C Haller K Pampanwar V et al 2006 MGOS
A resource for studying Magnaporthe grisea and Oryza sativa
interactions Molecular PlantndashMicrobe Interactions 19 1055ndash1061
Sun Y Liu W Han B Zhang J Liu B 2006 Purification and
characterization of two types of chitosanase from a Microbacterium
sp Biotechnology Letters 28 1393ndash1399
Talbot NJ 2003 On the trail of a cereal killer exploring the biology of
Magnaporthe grisea Annual Review of Microbiology 57 177ndash202
Tanaka MU Nakajima M Katoh E et al 2007 Molecular
interactions of a soluble gibberellin receptor GID1 with a rice DELLA
protein SLR1 and gibberellin The Plant Cell 19 2140ndash2155
Tiwari BS Belenghi B Levine A 2002 Oxidative stress increased
respiration and generation of reactive oxygen species resulting in ATP
depletion opening of mitochondrial permeability transition and
programmed cell death Plant Physiology 128 1271ndash1281
Usha Rani P Jyothsna Y 2009 Physiological changes in the
groundnut Arachis hypogaea L plant due to the infection of a fungal
pathogen Cercosporidium personatum Deighton Journal of
Allelopathy 23 369ndash378
Vanitha SC Niranjana SR Umesha S 2009 Role of phenylalanine
ammonia lyase and polyphenol oxidase in host resistance to bacterial
wilt of tomato Journal of Phytopathology 157 552ndash557
Vergne E Ballini E Droc G Tharreau D Notte ghem J-L
Morel J-B 2008 Archipelago a dedicated resource for exploiting
past present and future genomic data on disease resistance
regulation in rice Molecular PlantndashMicrobe Interactions 21 869ndash878
von Braun J 2007 The world food situation new driving forces and
required actions Washington DC International Food Policy Research
Institute (IFPRI) Food policy reports 1024990896295303
Wang ZX Yano M Yamanouchi U Iwamoto M Monna L
Hayasaka H Katayose Y Sasaki T 1999 The Pib gene for rice
blast resistance belongs to the nucleotide binding and leucine-rich
repeat class of plant disease resistance genes The Plant Journal 19
55ndash64
Whetten RW Mackay JJ Sederoff RR 1998 Recent advances in
understanding lignin biosynthesis Annual Review of Plant Physiology
and Plant Molecular Biology 29 585ndash609
Xiong L Lee WM Qi M Yang Y 2001 Identification of defense
related rice genes by suppression subtractive hybridization and
differential screening Molecular PlantndashMicrobe Interactions 14
685ndash692
Zhou YL Xu MR Zhao MF Xie XW Zhu LH Fu BY Li ZK 2010
Genome-wide gene responses in a transgenic rice line carrying maize
resistance gene Rxo1 to the rice bacterial streak pathogen
Xanthomonas oryzae pv oryzicola BMC Genomics 1011861471-
2164-11-78
772 | Gupta et alD
ownloaded from
httpsacademicoupcom
jxbarticle632757499789 by guest on 26 Decem
ber 2021