the solution structure and dna-binding properties of the cold-shock domain of the human y-box...
TRANSCRIPT

doi:10.1006/jmbi.2001.5334 available online at http://www.idealibrary.com on J. Mol. Biol. (2002) 316, 317±326
The Solution Structure and DNA-binding Properties ofthe Cold-shock Domain of the Human Y-boxProtein YB-1
Cathelijne P. A. M. Kloks1, Christian A. E. M. Spronk1
Edwin Lasonder2, Astrid Hoffmann3, Geerten W. Vuister1
Stephan Grzesiek4 and Cornelis W. Hilbers1*
1NSR Center for MolecularStructure, Design andSynthesis, Laboratory ofBiophysical ChemistryUniversity of NijmegenToernooiveld 1, 6525 EDNijmegen, The Netherlands2Laboratory Molecular BiologyUniversity of NijmegenToernooiveld 1, 6525 EDNijmegen, The Netherlands3IBI-2, ForschungszentrumJuÈ lich, 52425 JuÈ lich, Germany4Biozentrum, UniversitaÈt BaselKlingelbergstra�e 70CH-4056 Basel, Switzerland
Present address: A. Hoffmann, AGmbH, P.O. Box 1230, 35002 Marbu
Abbreviations used: CSD, cold-shdsDNA, double-stranded DNA; ED(diethylaminopropyl)-carbodiimide;proteins from Xenopus leavis; mRNPribonucleoprotein; NHS, N-hydroxynuclear Overhauser effect; OB-fold,oligosaccharide-binding fold; PBS, psaline (2.67 mM KCl, 1.15 mM KH2
Na2HPO4, 0.1379 M NaCl, pH 7.3);ribonucleoprotein; RU, resonance uplasmon resonance; ssDNA, single-
E-mail address of the [email protected]
0022-2836/02/020317±10 $35.00/0
The human Y-box protein 1 (YB-1) is a member of the Y-box proteinfamily, a class of proteins involved in transcriptional and translationalregulation of a wide range of genes. Here, we report the solution struc-ture of the cold-shock domain (CSD) of YB-1, which is thought to beresponsible for nucleic acid binding. It is the ®rst structure solved of aeukaryotic member of the cold-shock protein family and consists of aclosed ®ve-stranded anti-parallel b-barrel capped by a long ¯exible loop.The structure of CSD is similar to the OB-fold and a comparison withbacterial cold-shock proteins shows that its structural properties are con-served from bacteria to man. Our data suggest the presence of a DNA-binding site consisting of a patch of positively charged and aromatic resi-dues on the surface of the b-barrel. Further, it is shown that CSD, whichhas a preference for binding single-stranded pyrimidine-rich sequences,binds weakly and hardly speci®cally to DNA. Binding af®nities reportedfor intact YB-1 indicate that domains other than the CSD play a role inDNA binding of YB-1.
# 2002 Elsevier Science Ltd.
Keywords: cold-shock; OB-fold; Y-box protein; single-stranded DNAbinding; solution structure
*Corresponding authorIntroduction
The human Y-box protein 1 (YB-1) is a memberof the Y-box protein family. Proteins of this familyare active as transcriptional and translational regu-lators for messenger RNA and for a wide rangeof genes that contain the Y-box sequence
ventis Behringrg, Germany.ock domain;C, N-ethyl-N0-FRG Y, Y-box, messengersuccinimide; NOE,oligonucleotide/hosphate-buffered
PO4, 8.06 mMRNP,
nit; SPR, surfacestranded DNA.ing author:
(50-CTGATTGGCCAA-30) in their promoters(reviewed by Ladomery1). The Y-box proteins were®rst identi®ed and isolated by binding to a DNAprobe containing the Y-box sequence.2 It has beendemonstrated that binding of the Y-box proteins tothe intact Y-box sequence involves speci®c inter-actions.3 Subsequent binding studies have shownthat the Y-box proteins can bind to a whole rangeof nucleic acids; namely, single-stranded DNA(ssDNA), RNA, damaged DNA, RNA and even tri-ple-stranded DNA.4,5 Although no consistent pic-ture emerged from the different binding studies, itappears that Y-box proteins have a preference forbinding ssDNA, especially when it contains the Y-box sequence.6 Further, evidence exists that Y-boxproteins have a preference for binding pyrimidine-rich DNA sequences.7
The Y-box proteins are generally composed ofthree domains: a variable N-terminal domain, thecold-shock domain (CSD) and a C-terminal domain(cf. Figure 1(a)). The function of the variableN-terminal domain is not known. The C-terminal
# 2002 Elsevier Science Ltd.

Figure 1. (a) Domain structure of Y-box proteins. The variable N-terminal (V) domain and the cold-shock domain(CSD) are indicated in purple and blue, respectively. The C-terminal domain consists of acidic (A) and basic (B)blocks of about 30 amino acid residues each, which are indicated in green and yellow. (b) Sequence alignment of theCSDs of eukaryotic Y-box proteins and prokaryotic CSD-related proteins with the human YB-1 CSD. The RNA-bind-ing RNP-1 and RNP-2-like motifs are in the green and gray boxes. Identical residues are depicted in black, homolo-gous residues are underlined and in blue. Differences in conservation of residues between prokaryotes andeukaryotes are marked by arrows. Note that the ®rst ®ve N-terminal residues (MKKVI) of the eukaryotic CSDs havebeen omitted. The following abbreviations are used: Dbp, DNA-binding protein; EF, elongation factor.
318 The Cold-shock Domain: Structure and DNA Binding
domain consists of alternating basic and aromaticregions of approximately 30 residues each.8 Itfacilitates RNA binding and is involved in protein-protein interaction.9 Thus, the C-terminal domainis part of the dimerisation site of the Y-boxproteins6,8 and it is essential for the formation ofthe messenger ribonucleoprotein (mRNP) particlesin which mRNA is stored.10
The central CSD is highly conserved within thefamily of Y-box proteins, having 93 % sequenceidentity7 (Figure 1(b)). It contains two RNA-bind-ing motifs, designated RNP-1 and RNP-211 (boxedin Figure 1(b)). The domain is involved in speci®c9
and non-speci®c RNA binding,12 and in speci®cssDNA binding.6 The non-speci®c RNA bindingoccurs during the packaging of mRNA in maturinggerm cells. Speci®c RNA binding has been pro-posed to take place in the nucleation processduring the assembly of the mRNP particles.9,13 Thespeci®c ssDNA-binding properties of the CSD arerequired for the binding of Y-box proteins tospeci®c single-stranded parts of promoters, whichthereby affect the binding of other transcriptionfactors and induce transcription.14
The sequence of CSD is similar (43 % sequenceidentity) to those of bacterial cold-shock proteins(Csps)7 (Figure 1(b)). These latter proteins areinvolved in the process in which the cell-growth ofbacteria is resumed after they have been exposedto a serious temperature drop (reviewed by Jones& Inouye15 and Wolffe16). CSD is predicted to havea fold similar to those of these Csps, denoted com-monly as the oligonucleotide/oligosaccharide fold(OB-fold).17
We have determined the solution structure ofthe CSD of the human Y-box protein YB-1 by high-resolution NMR spectroscopy. In addition, wehave investigated the ssDNA-binding properties ofthis domain and made a comparison with those ofthe intact YB-1 protein.
Results
NMR resonance assignments andstructure calculations
In addition to the NMR resonance assignmentsreported previously,18 85 new assignments weremade in order to determine the structure of the

The Cold-shock Domain: Structure and DNA Binding 319
YB-1 CSD. This was achieved by iterative analysisof the nuclear Overhauser effect (NOE) and rotat-ing frame Overhauser effect (ROE) experimentsand further analysis of the previously described tri-ple resonance experiments. Additional aromaticand methyl group assignments were obtained from15N-®ltered total correlated spectroscopy (TOCSY)and constant time heteronuclear single quantumcoherence (HSQC) experiments, respectively.Although the process of assigning was seriouslycomplicated due to denaturation and an equili-brium between folded (60 %) and unfolded (40 %)states of the protein, it was possible to assign 91 %of all 1H, 15N and 13C backbone nuclei and 84 % ofthe side-chain nuclei of the folded state. Foldedand unfolded states were distinguished by combin-ing the NOE and ROE data.
The total amount of restraints that could bederived is rather low, due to the lack of resonanceassignments for the region comprising residues 42to 56, which are part of a ¯exible loop (see below),overlap and the abovementioned equilibriumbetween folded and unfolded states. Finally, a totalof 470 distance restraints (of which 187 intraresi-due, 124 sequential, 26 medium-range and 133long-range) and 59 backbone torsion anglerestraints were used to calculate the structures ofthe CSD (See Table 1 and Materials and Methodsfor further details).
Description of the structure
We found that CSD folds into a ®ve-strandedanti-parallel b-barrel (Figures 2 and 3). The b-bar-rel, which forms the well-de®ned part of the pro-tein with a backbone rmsd of 0.86, is closed by a
Table 1. Structural statistics
Averaged rmsd from distance restraints (AÊ ) 0.043rmsd from experimental torsion angle
restraints (deg.) (34 f, 25 c) 0.6Number of distance restraints per core
residue 7.3Number of distance restraint
violations > 0.55 AÊ 0Number of torsion angle restraint
violations > 0.5 � 0Average rmsd deviations from idealized covalent geometry
Bonds (AÊ ) 0.0040Angles (deg.) 1.36Impropers (deg.) 2.45
Atomic rmsd differences versus average structure (AÊ )Heavy atoms of core 1.59Backbone of core 0.86Heavy atoms all residues 2.90Backbone all residues 2.04
Ramachandran region population (%) for non-glycine,non-proline residuesMost favored regions 75.6Additionally allowed regions 22.1Generously allowed regions 2.1Forbidden regions 0.2
Core residues (5-41, 57-77) were used to superimpose thestructure ensemble.
parallel line-up of strands b3 and b5. The connec-tions between the different b-strands consist ofthree tight type I turns (b1-b2, b2-b3 and b4-b5)and a loop (b3-b4). Although the local structure ofall three turns is well determined by the NMRdata, only turn b2-b3 has a ®xed orientation withrespect to the barrel (Figure 3(b)). In contrast, turnsb1-b2 and b4-b5 can adopt different orientationswith respect to the core of the protein. For turn b1-b2, the absence of 15N-HN resonances of residuesLys14, Trp15, Asn17, Val18, and Tyr22 in the NMRspectra, as well as relaxation data of the surround-ing residues (data not shown), indicate that this isdue to hinge-motions at the ends of the turn (resi-dues 15 and 23). For turn b4-b5, a similar hingemotion, suggested by the relaxation data (data notshown), is present, involving residues 66 and 69.
The long loop between strands b3 and b4 is illde®ned, except for a 310-helix turn at the C-term-inal end of b3 (Figure 3(a)). Most HN-N resonancesof residues in this loop are not visible in the NMRspectra, which is probably due to unfavorableexchange rates of the amide protons. Residues inthe loop for which the 15N-HN groups could beassigned show increased ¯exibility in relaxationexperiments (data not shown).
DNA binding
In order to probe the binding interface of CSD, itwas titrated with the core of the Y-box sequence,the ssDNA pentamer, 50-ATTGG-30. During thetitration, 15N-HSQC NMR spectra were used tomonitor chemical shift changes in CSD upon DNAbinding. As can be seen in Figure 4(a), increasingDNA concentrations cause a gradual shift of the15N-1H cross-peaks, indicating a fast chemicalexchange between the free and bound state on theNMR time-scale. The largest changes in 15N-1Hchemical shifts are observed for backbone reson-ances of residues Lys8, Thr12, Phe16, Arg19,Asn20, Gly23-Ile25, Asp33, Phe35-Thr39, Leu50,Lys58, Val64, Gly66, Lys68, Gly69 and Glu71, andfor the side-chain resonances of Trp15 and Gln38,indicating that these residues are in close proximityto the nucleic acid or that their chemical shifts areaffected indirectly by the interaction. We havedetermined the dissociation constant KD from the15N-1H proton chemical shifts as a function of theamount of added equivalents of DNA, as exempli-®ed for residue Trp15 and His37 in Figure 4(b) and(c). From the results, we estimate that KD for theprotein-DNA complex is of the order of 10ÿ4 M,assuming a 1:1 binding stoichiometry.
To de®ne the DNA-binding site and the residuesinvolved in the binding, we ®rst considered theresidues showing shifts of their 15N-1H amide res-onances and belonging to the barrel. Most of theaffected residues belong to the RNP-1 and RNP-2motifs on b2 and b3, respectively, or are on thestrand neighboring these RNA-binding motifs.They are clearly present in different layers of the b-barrel (see Figure 4(d)). They were divided into
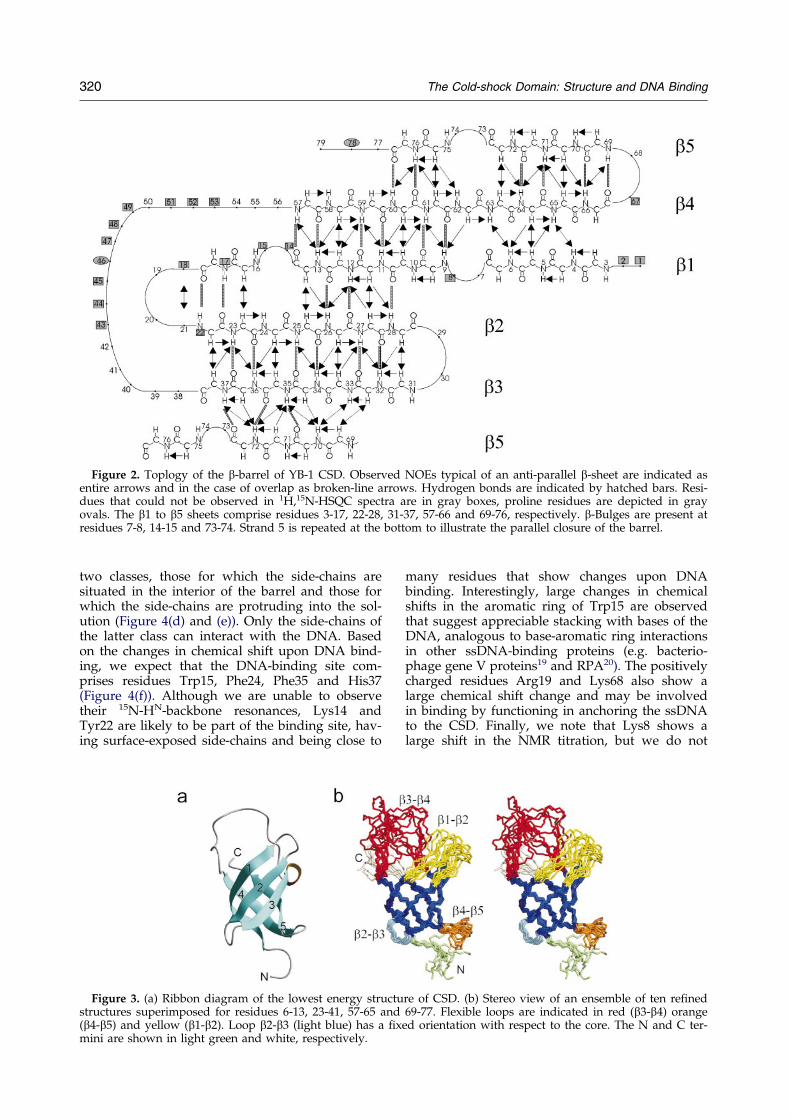
Figure 2. Toplogy of the b-barrel of YB-1 CSD. Observed NOEs typical of an anti-parallel b-sheet are indicated asentire arrows and in the case of overlap as broken-line arrows. Hydrogen bonds are indicated by hatched bars. Resi-dues that could not be observed in 1H,15N-HSQC spectra are in gray boxes, proline residues are depicted in grayovals. The b1 to b5 sheets comprise residues 3-17, 22-28, 31-37, 57-66 and 69-76, respectively. b-Bulges are present atresidues 7-8, 14-15 and 73-74. Strand 5 is repeated at the bottom to illustrate the parallel closure of the barrel.
320 The Cold-shock Domain: Structure and DNA Binding
two classes, those for which the side-chains aresituated in the interior of the barrel and those forwhich the side-chains are protruding into the sol-ution (Figure 4(d) and (e)). Only the side-chains ofthe latter class can interact with the DNA. Basedon the changes in chemical shift upon DNA bind-ing, we expect that the DNA-binding site com-prises residues Trp15, Phe24, Phe35 and His37(Figure 4(f)). Although we are unable to observetheir 15N-HN-backbone resonances, Lys14 andTyr22 are likely to be part of the binding site, hav-ing surface-exposed side-chains and being close to
Figure 3. (a) Ribbon diagram of the lowest energy structustructures superimposed for residues 6-13, 23-41, 57-65 and(b4-b5) and yellow (b1-b2). Loop b2-b3 (light blue) has a ®xmini are shown in light green and white, respectively.
many residues that show changes upon DNAbinding. Interestingly, large changes in chemicalshifts in the aromatic ring of Trp15 are observedthat suggest appreciable stacking with bases of theDNA, analogous to base-aromatic ring interactionsin other ssDNA-binding proteins (e.g. bacterio-phage gene V proteins19 and RPA20). The positivelycharged residues Arg19 and Lys68 also show alarge chemical shift change and may be involvedin binding by functioning in anchoring the ssDNAto the CSD. Finally, we note that Lys8 shows alarge shift in the NMR titration, but we do not
re of CSD. (b) Stereo view of an ensemble of ten re®ned69-77. Flexible loops are indicated in red (b3-b4) orange
ed orientation with respect to the core. The N and C ter-

The Cold-shock Domain: Structure and DNA Binding 321
understand the reason for this shift, since the resi-due is situated at the opposite side of the bindingsite on the b-barrel surface. Summarizing, we con-clude that, with the exception of Lys8, all residuesthat show large chemical shift changes upon DNAbinding are located between the b1-b2 and b4-b5turns.
We further investigated the DNA binding byCSD using surface plasmon resonance (SPR). Foursequences, namely a Y-box-containing sequence(sequence 1), a pyrimidine-rich sequence (sequence2) and two control sequences (sequence 3 and 4),were used (Figure 5(a)). A superposition of tenreal-time SPR response curves recorded at differentconcentration of CSD is shown in Figure 5(b). Thesteady-state values are reached too rapidly (withinless than ®ve seconds) to determine on and offrates. Binding constants were therefore determinedby steady-state kinetics. To a ®rst approximation,the binding constants can be derived on the basisof the assumption that each DNA molecule carriesa number of independent binding sites. In this situ-ation the relationship:
1
R� 1
Rmax� 1
KAPRmax
is applicable, where R is the steady-state value ofthe response functions at a particular CSD concen-tration, P, and Rmax is the maximum responseobtained at CSD concentrations leading to satur-ation of the available binding sites. Thus, for thissituation, a linear correlation between 1/R and 1/Pis expected. Indeed, such a linear relationship isfound for sequences 1, 2 and 4, as shown forsequence 2 in Figure 5(c). In the case of sequence 3,binding turned out to be too weak to be studied bySPR with the concentrations of CSD available. Thiscan be explained by the formation of double-stranded structures, which were found forsequence 3 by optical melting experiments (datanot shown). In Table 2, the equilibrium constantKD, the number of CSD molecules per DNA mol-ecule and Rmax are given for the differentsequences, showing that the Y-box-containingsequence 1 and sequence 4 bind more weakly toCSD, with a KD almost two orders of magnitudelarger, than the pyrimidine-rich sequence 2. The
Table 2. Binding properties of the different oligonucleotide s
Sequence Length Pu:Py A G C
1 34 1.13 7 11 72 26 0.08 2 0 203 20 1.86 4 9 54 19 1.11 6 4 45 5 1.50 1 2 0
Column 1 lists the sequence numbers as indicated in Figure 5. SeqThe length of the sequence, the ratio between the pyrimidine and pthe state of the DNA strands (i.e. single-stranded (ss) or double-stranare given in the following columns for the four sequences used in tmated uncertainties of 50 % and 10 %, respectively. The stoichiometry
a Too weak to determine.
number of proteins bound to one DNA moleculefollows from:
n � Rmax=MCSD
RU�DNAbound�=MDNA
In this formula, MCSD and MDNA stand for the mol-ecular mass of CSD and the DNA sequence,respectively, and RU(DNAbound) stands for thenumber of response units of DNA coupled to theused ¯ow-cell.
Discussion
Here, we have shown that the cold-shockdomain (SCD) of YB-1 adopts a ®ve-stranded anti-parallel b-barrel structure. Its structural character-istics are very similar to proteins of the OB-foldfamily, with ®ve b-strands and a shear number of8.17 A notable difference with respect to most OB-fold proteins is that CSD contains an extra b-bulgeat Thr7 and Lys8 (in addition to the bulge at Lys14and Trp15 generally found in OB-folds) that allowsthe relatively long b1 to follow the curvature of thebarrel.
The N-terminal part of CSD and the loopbetween b3 and b4 are appreciably longer than thecorresponding parts in the bacterial cold-shockproteins, CspA of Escherichia coli,21,22 and CspB ofBacillus subtilis.23,24 In CSD and these bacterial pro-teins, the b3-b4 loop is largely unstructured, exceptfor a single 310-helix turn at the start of the loop.Moreover, unlike in many other OB-fold proteins,it does not contain an a-helix. At this point, thefunctional role of this long loop is not clear,although it might be a scaffold to orient the DNA-binding wing b1-b2 as in the ssDNA-binding geneV proteins of bacteriophages M13 and Pf3, whereit probably serves to guide the orientation of theDNA-binding wing, b1-b2.25 In this respect, it isinteresting to note that Leu50, which is located inthe center of the loop and surrounded by threepositively charged and one aromatic residue,shows signi®cant changes in chemical shifts uponDNA binding, indicating that the loop is in¯u-enced by DNA binding, although it is not part ofthe binding site situated between loops b1-b2 and
equences studied
T State KD (mM) Rmax (RU) n
9 ss 93 477 4.04 ss 1.5 396 2.32 ds (a) - -5 ss 52 326 1.42 - 100 - 1
uence 5 (ATTGG) was used in the NMR titration (see the text).urine bases, the number of the individual bases A, G, C, T andded (ds)) as determined by optical melting point measurementshe SPR experiments. KD and Rmax are average values with esti-
of the interaction is given in the last column.
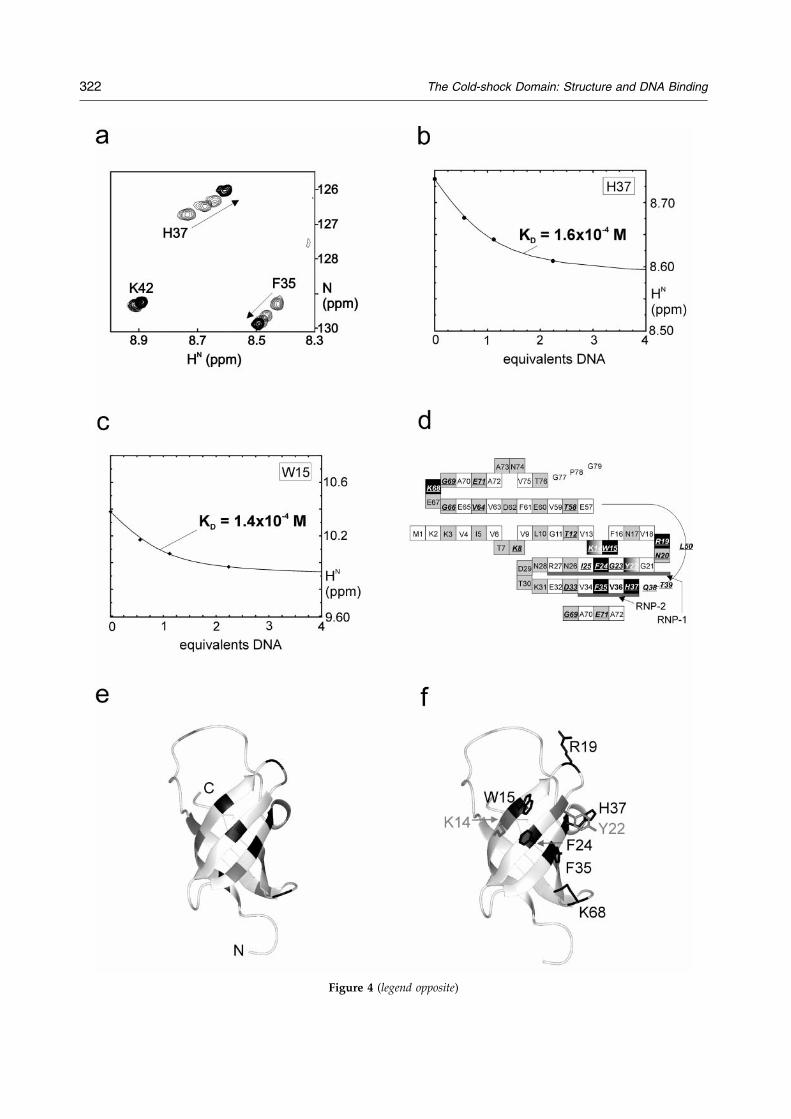
Figure 4 (legend opposite)
322 The Cold-shock Domain: Structure and DNA Binding
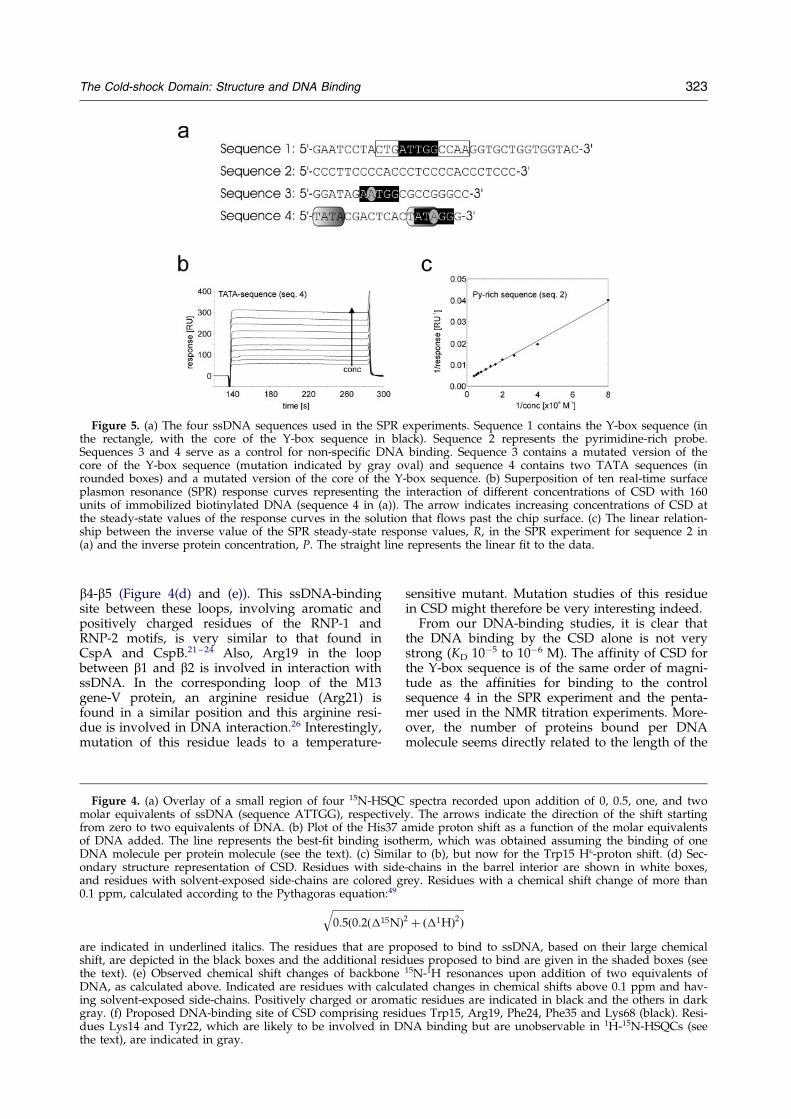
Figure 5. (a) The four ssDNA sequences used in the SPR experiments. Sequence 1 contains the Y-box sequence (inthe rectangle, with the core of the Y-box sequence in black). Sequence 2 represents the pyrimidine-rich probe.Sequences 3 and 4 serve as a control for non-speci®c DNA binding. Sequence 3 contains a mutated version of thecore of the Y-box sequence (mutation indicated by gray oval) and sequence 4 contains two TATA sequences (inrounded boxes) and a mutated version of the core of the Y-box sequence. (b) Superposition of ten real-time surfaceplasmon resonance (SPR) response curves representing the interaction of different concentrations of CSD with 160units of immobilized biotinylated DNA (sequence 4 in (a)). The arrow indicates increasing concentrations of CSD atthe steady-state values of the response curves in the solution that ¯ows past the chip surface. (c) The linear relation-ship between the inverse value of the SPR steady-state response values, R, in the SPR experiment for sequence 2 in(a) and the inverse protein concentration, P. The straight line represents the linear ®t to the data.
The Cold-shock Domain: Structure and DNA Binding 323
b4-b5 (Figure 4(d) and (e)). This ssDNA-bindingsite between these loops, involving aromatic andpositively charged residues of the RNP-1 andRNP-2 motifs, is very similar to that found inCspA and CspB.21 ± 24 Also, Arg19 in the loopbetween b1 and b2 is involved in interaction withssDNA. In the corresponding loop of the M13gene-V protein, an arginine residue (Arg21) isfound in a similar position and this arginine resi-due is involved in DNA interaction.26 Interestingly,mutation of this residue leads to a temperature-
Figure 4. (a) Overlay of a small region of four 15N-HSQCmolar equivalents of ssDNA (sequence ATTGG), respectivelfrom zero to two equivalents of DNA. (b) Plot of the His37 aof DNA added. The line represents the best-®t binding isotDNA molecule per protein molecule (see the text). (c) Similaondary structure representation of CSD. Residues with sideand residues with solvent-exposed side-chains are colored g0.1 ppm, calculated according to the Pythagoras equation:49����������������������������
0:5�0:2��15N�q
are indicated in underlined italics. The residues that are proshift, are depicted in the black boxes and the additional residthe text). (e) Observed chemical shift changes of backboneDNA, as calculated above. Indicated are residues with calcuing solvent-exposed side-chains. Positively charged or aromagray. (f) Proposed DNA-binding site of CSD comprising residdues Lys14 and Tyr22, which are likely to be involved in Dthe text), are indicated in gray.
sensitive mutant. Mutation studies of this residuein CSD might therefore be very interesting indeed.
From our DNA-binding studies, it is clear thatthe DNA binding by the CSD alone is not verystrong (KD 10ÿ5 to 10ÿ6 M). The af®nity of CSD forthe Y-box sequence is of the same order of magni-tude as the af®nities for binding to the controlsequence 4 in the SPR experiment and the penta-mer used in the NMR titration experiments. More-over, the number of proteins bound per DNAmolecule seems directly related to the length of the
spectra recorded upon addition of 0, 0.5, one, and twoy. The arrows indicate the direction of the shift startingmide proton shift as a function of the molar equivalents
herm, which was obtained assuming the binding of oner to (b), but now for the Trp15 He-proton shift. (d) Sec--chains in the barrel interior are shown in white boxes,rey. Residues with a chemical shift change of more than
�������������������������2 � ��1H�2�posed to bind to ssDNA, based on their large chemicalues proposed to bind are given in the shaded boxes (see15N-1H resonances upon addition of two equivalents oflated changes in chemical shifts above 0.1 ppm and hav-tic residues are indicated in black and the others in darkues Trp15, Arg19, Phe24, Phe35 and Lys68 (black). Resi-
NA binding but are unobservable in 1H-15N-HSQCs (see

324 The Cold-shock Domain: Structure and DNA Binding
probes. Although these data indicate that bindingof CSD to the Y-box sequence occurs in a non-speci®c manner, contrary to what has been foundfor intact Y-box proteins, we do observe a prefer-ence of CSD for interacting with ssDNA andincreased Py:Pu ratios.
In our treatment we assumed that the bindingsites on the DNA are independent, which is sup-ported by the linear relationship found between1/R and 1/P. This implies that there is no coopera-tivity in binding. This corresponds nicely with thebinding behavior of the gene V protein of bacterio-phage M13, which shows high cooperativity forlong ssDNA-chains, while this factor is two ordersof magnitude smaller, if not equal to one, for oligo-nucleotides with a length comparable to that of theDNA used in our study.27 We estimate the numberof nucleotides per CSD to be roughly between 7and 12. For CspB of B. subtilis, it was found thatfor binding to a 23-mer ssDNA, each protein mol-ecule covered six to seven nucleotides.28,29
The preference of CSD for binding pyrimidine-rich sequences was found for other ssDNA-bindingproteins,30 and might be related to the in¯uence ofstacking interactions. Protein binding to pyrimi-dine-rich ssDNA would not require unstacking ofthe bases, in contrast to purine-rich DNA, andwould be energetically more favorable.30 The pre-sence of stacking interactions in the CSD-DNAcomplexes is indicated by large changes in chemi-cal shift of the aromatic ring protons of Trp15 inthe NMR titration experiment. In all, we surmisethat the interaction between CSD and ssDNA isgoverned mainly by non-speci®c, for a large partelectrostatic, interactions. In addition, stackinginteractions are present.
The weak interactions and the apparent absenceof highly speci®c interactions between CSD andthe Y-box sequence con®rm previous ®ndings thatthe ¯anking domains of CSD in intact YB-1 play animportant role in nucleic acid binding. DNA-bind-ing studies on Y-box proteins indicated that,although CSD on its own is suf®cient to bind tossDNA, the C-terminal domain facilitates ssDNAbinding and is needed for strong interaction.6,31,32
Intact Y-box proteins, such as FRG Y-proteins,6
chkYB-1 and 2,31,32 mRNP3 � 433 and the humandbpA/B protein,5 which are highly homologous tothe human YB-1 protein, bind the Y-box sequencewith dissociation constants of 10ÿ7 to 10ÿ9 M,whereas for CSD alone, KD is 10ÿ4 to 10ÿ6 M. Poss-ibly, these ¯anking domains are involved inadditional interactions with ssDNA or provideextra stability of the CSD domain, resulting in atighter binding.
Materials and Methods
NMR spectroscopy and structure calculations
Uniformly 13C,15N-labeled and 15N-labeled NMR-samples were prepared as described.18 The samples con-sisted of 0.5 to 1.5 mM protein (8740 Da) in 250 ml of
95 % H2O/5 % 2H2O or 100 % 2H2O at pH 6.7 in Shige-mi-tubes. All NMR experiments were recorded at 298 Kon Bruker DRX 600 MHz and Varian UnityInova spec-trometers operating at 500 and 750 MHz. Spectra wereprocessed using NMRPipe34 and assigned using PIPP.35
The 15N-®ltered TOCSY36 and constant time HSQCexperiments37 were used for additional assignment ofaromatic and methyl groups, respectively. Titration ofthe YB-1 CSD, monitored with 15N-HSQC-spectra, wasperformed by adding 0, 0.5, 1.0 and 2.0 equivalents ofsingle-stranded 50-ATTGG-30 DNA (OliGold, Eurogentec)to an NMR sample containing 2.7 mg of CSD.
Distance restraints for structure calculations werederived from 15N-NOESY-HSQC, 13C-NOESY-HSQC and15N-ROESY-HSQC experiments with mixing times of 90,80, and 34 ms, respectively. NOEs resulting from spindiffusion were identi®ed by careful inspection of the15N-NOESY-HSQC and 15N-ROESY-HSQC experimentsand not converted to distance restraints. After calibrationagainst known distances in proteins, NOEs were classi-®ed as corresponding to 2.8, 3.5, 4.5 and 5.0 AÊ andassigned upper bounds of 3.3, 4.0, 5.0 and 5.5 AÊ , respect-ively. For overlapping NOEs, an upper bound of 7.3 AÊ
was introduced. All lower bounds were set to 1.8 AÊ .Dihedral f and c torsion angle restraints were derivedfrom a 3D-HNHA experiment38 combined with infor-mation from Ca and Cb chemical shifts according to themethod described by Spera & Bax.39
Initially, 80 structures were calculated using the Tor-sion Angle Dynamics protocol40,41 in X-PLOR42 using theCHARMM19 force-®eld including dihedral energy termsand starting from extended conformations. Subsequently,the ten lowest-energy structures were re®ned byrestrained molecular dynamics for 35 ps in a water-boxwith three chloride counterions using force-®eldCHARMM22 in X-PLOR similar to the method describedby Spronk et al.43 Analysis of structures was done usingAQUA/PROCHECK-NMR44 and WHAT IF.45
Figures were generated using the program MOLMOL2.6.46
Surface plasmon resonance experiments
All SPR binding experiments were performed using aBiacoreTM 2000, using research grade CM5 chips. Chipsand required chemicals N-ethyl-N0-(diethylaminopropyl)-carbodiimide (EDC), N-hydroxysuccinimide (NHS) andethanolamine were purchased from Biacore AB (Uppsa-la, Sweden), and streptavidin was purchased fromSigma. The 30-biotinylated ssDNA sequences were pur-chased from Biolegio (Wijchen, The Netherlands), orwere synthesized for us at the EMBL (Heidelberg,Germany).
In all experiments, PBS (pH 7.3), 0.1 M NaCl, 0.05 %(v/v) Tween20 was used as running buffer (withinexperimental accuracy the binding was the same withoutdetergent). A ¯ow-rate of 5 ml/minute was used unlessstated otherwise. Approximately 10,000 units of strepta-vidin was immobilized on each ¯ow-cell of a CM5 sen-sor chip47 by activating the chip surface with 75 ml ofEDC/NHS (1:1, v/v) and subsequent coupling with50 mg/ml of streptavidin in 10 mM sodium acetate buf-fer (pH 5.0) for 24 minutes. Then 1 M ethanolamine(pH 8.5) was added and left for 15 minutes to deactivatethe surface, followed by a regeneration step of 15 ml of0.1 mM NaOH. At least one ¯ow-cell was coupled withonly streptavidin to serve as a reference channel. Biotiny-lated DNA was added to obtain the required immobiliz-

The Cold-shock Domain: Structure and DNA Binding 325
ation level, followed by a regeneration step with 15 ml of0.1 M NaOH. Regeneration of the chip between exper-iments was performed in the same way with 0.1 MNaOH.
Possible non-speci®c binding of CSD to the chip waschecked using two types of sensor surfaces. First, theprotein was allowed to bind to a new untreated CM5chip, which showed that CSD binds non-speci®cally tothe chip. Secondly, the protein was tested for binding toa CM5 chip occupied by streptavidin. This showed thatCSD did not bind to the chip that was coupled to a verylarge amount of streptavidin (approximately 10,000units).
The ¯ow-cells contained approximately 160 units ofbiotinylated DNA of each sequence. Note that one unitcorresponds to 10ÿ12 g/mm2 of protein or DNA on thesurface.48 For sequences 1, 3 and 4, 12 concentrations ofCSD in the range 2.4 mg/ml to 0.024 mg/ml in runningbuffer were used to measure the binding kinetics for ®veminutes at a rate of 50 ml/minute at 25 �C. CSD bindingto sequence 2 was measured with concentrations rangingfrom 0.024 mg/ml to 0.001 mg/ml.
Data Bank accession numbers
The complete assignment of the protein, including sca-lar couplings and relaxation times, have been depositedat the BioMagResBank under accession number 4147(updated since Kloks et al.18). All data have been sub-mitted to the Brookhaven PDB under entry number1h95.
Acknowledgements
This research was supported by DFG grant Gr1683/1-1 and the Netherlands Foundation for Chemical Scienceswith ®nancial assistance from the Netherlands Organiz-ation for Scienti®c Research (NWO). We acknowledge J.Aelen and M. Rogowski for preparing protein samples.
References
1. Ladomery, M. (1997). Multifunctional proteinssuggest connections between transcriptional andpost-translational processes. BioEssays, 19, 903-909.
2. Dorn, A., Bollekens, J., Staub, A., Benoist, C. &Mathis, D. (1987). A multiplicity of CCAAT box-binding proteins. Cell, 50, 863-872.
3. Didier, D. K., Schiffenbauer, J., Woulfe, S. L. &Zacheis, M. (1988). Characterization of the cDNAencoding a protein binding to the major histocom-patibility complex class II Y box. Proc. Natl Acad.Sci. USA, 85, 7322-7326.
4. Koike, K., Uchiumi, T., Ohga, T., Toh, S., Wada, M.,Kohno, K. & Kuwano, M. (1997). Nuclear transloca-tion of the Y-box binding protein by ultravioletirradiation. FEBS Letters, 417, 390-394.
5. Horwitz, E. M., Maloney, K. A. & Ley, T. J. (1994).A human protein containing a ``cold-shock'' domainbinds speci®cally to H-DNA upstream from thehuman g-globin genes. J. Biol. Chem. 269, 14130-14139.
6. Tafuri, S. R. & Wolffe, A. P. (1992). DNA binding,multimerization, and transcription stimulation bythe Xenopus Y box proteins in vitro. New Biol. 4,349-359.
7. Wolffe, A. P. (1994). Structural and functional prop-erties of the evolutionarily ancient Y-box family ofnucleic acid binding proteins. BioEssays, 16, 245-251.
8. Murray, M. T., Schiller, D. L. & Franke, W. W.(1992). Sequence analysis of cytoplasmic mRNA-binding proteins of Xenopus oocytes identi®es afamily of RNA-binding proteins. Proc. Natl Acad. Sci.USA, 89, 11-15.
9. Bouvet, P., Mastumoto, K. & Wolffe, A. P. (1995).Sequence-speci®c RNA recognition by the XenopusY-box proteins. An essential role for the cold-shockdomain. J. Biol. Chem. 270, 28297-28303.
10. Matsumoto, K., Meric, F. & Wolffe, A. P. (1996).Translational repression dependent on the inter-action of the Xenopus Y-box protein FRGY2 withmRNA. Role of the cold-shock domain, tail domainand selective RNA sequence recognition. J. Biol.Chem. 271, 22706-22712.
11. Landsman, D. (1992). RNP-1, an RNA binding motifis conserved in the DNA-binding cold-shockdomain. Nucl. Acids Res. 20, 2861-2864.
12. Tafuri, S. R. & Wolffe, A. P. (1993). Selective recruit-ment of masked maternal mRNA from messengerribonucleoprotein particles containing FRGY2(mRNP4). J. Biol. Chem. 268, 24255-24261.
13. Wolffe, A. P. (1994). Nucleosome positioning andmodi®cation: chromatin structures that potentiatetranscription. Trends Biochem. Sci. 19, 240-244.
14. MacDonald, G. H., Itoh-Lindstrom, Y. & Ting, J. P.-Y. (1995). The transcriptional regulatary protein, YB-1, promotes single-stranded regions in the DRA-pro-moter. J. Biol. Chem. 270, 3527-3533.
15. Jones, P. G. & Inouye, M. (1994). The cold-shockresponse - a hot topic. Mol. Microbiol. 11, 811-818.
16. Wolffe, A. P. (1995). The cold-shock response in bac-teria. Sci. Prog. 78, 301-310.
17. Murzin, A. G. (1993). OB (oligonucleotide/oligosac-charide binding)-fold: common structural and func-tional solution for non-homologous sequences.EMBO J. 12, 861-867.
18. Kloks, C. P. A. M., Hoffmann, A., Omichinski, J. G.,Vuister, G. W., Hilbers, C. W. & Grzesiek, S. (1998).Resonance assignment and secondary structure ofthe cold shock domain of the human YB-1 protein.J. Biomol. NMR, 12, 463-464.
19. Alma, N. C. M., Harmsen, B. J. M., van Boom, J. H.,van der Marel, G. & Hilbers, C. W. (1982). 1H NMRstudies of the binding of bacteriophage M13encoded gene-5 protein to oligo(deoxyadenylicacid)s of variable lenghts. Eur. J. Biochem. 122, 319-326.
20. Bochkarev, A., Pfuetzner, R. A. P., Edwards, A. M.& Frappier, L. (1997). Structure of the single-stranded DNA binding domain of replicationprotein A bound to DNA. Nature, 385, 176-181.
21. Schindelin, H., Jiang, W., Inouye, M. & Heinemann,U. (1994). Crystal structure of Csp A, the majorcold-shock protein of Escherichia coli. Proc. Natl Acad.Sci. USA, 91, 5119-5123.
22. Newkirk, K., Feng, W., Jiang, W., Tejero, R.,Emerson, S. D., Inouye, M. & Montelione, G. T.(1994). Solution NMR structure of the major cold-shock protein (CspA) from Escherichia coli: identi®-cation of a binding epitope for DNA. Proc. NatlAcad. Sci. USA, 91, 5114-5118.
23. Schindelin, H., Marahiel, M. A. & Heinemann, U.(1993). Universal nucleic acid-binding domainrevealed by crystal structure of the B. subtilis majorcold-shock protein. Nature, 364, 164-168.

326 The Cold-shock Domain: Structure and DNA Binding
24. Schnuchel, A., Wiltscheck, R., Czisch, M., Herrier,M., Willimsky, G., Graumann, P. A. et al. (1993).Structure in solution of the major cold-shock proteinfrom Bacillus subtilis. Nature, 364, 169-171.
25. Rietman, B. H., Folkers, P. J. M., Folmer, R. H. A.,Tesser, G. I. & Hilbers, C. W. (1996). The solutionstructure of the synthetic circular peptideCGVSRQGKPYC: NMR studies of the folding of asynthetic model for the DNA-binding loop of thessDNA-binding protein encoded by gene V of phageM13. Eur. J. Biochem. 238, 706-713.
26. van Duijnhoven, J. P. M., Folkers, P. J. M., Stassen,A. P. M., Harmsen, B. J. M., Konings, R. N. H. &Hilbers, C. W. (1990). Structure of the DNA bindingwing of the gene-V encoded single-stranded DNAbinding protein of the ®lamentous bacteriophageM13. FEBS Letters, 261, 1-4.
27. Bulsink, H., Van Resandt, R. W. W., Harmsen, B. M.& Hilbers, C. W. (1986). Different DNA-bindingmodes and cooperativities for bacteriophage M-13gene-5 protein revealed by means of ¯uorescencedepolarization studies. Eur. J. Biochem. 157, 329-334.
28. Lopez, M. M., Yutani, K. & Makhatadze, G. I.(1999). Interactions of the major cold-shock proteinof Bacillus subtilis CspB with single-stranded DNAtemplates of different base composition. J. Biol.Chem. 274, 33601-33608.
29. Kremer, W., Schuler, B., Harrieder, S., Geyer, M.,Gronwald, W., Welker, C. et al. (2001). SolutionNMR structure of the cold-shock protein from thehyperthermophilic bacterium Thermotoga maritima.Eur. J. Biochem. 268, 2527-2539.
30. Bulsink, H., Harmsen, B. J. M. & Hilbers, C. W.(1985). Speci®city of the binding of bacteriophageM-13 encoded gene-5 protein to DNA and RNAstudied by means of ¯uorescence titrations. J. Biomol.Struct. Dynam. 3, 227-248.
31. Nambiar, A., Swamynathan, S. K., Kandala, J. C.& Guntaka, R. V. (1998). Characterization of theDNA-binding domain of the avian Y-box protein,chkYB-2, and mutational analysis of its single-strandbinding motif in the Rous sarcoma virus enhancer.J. Virol. 72, 900-909.
32. Nambiar, A., Kandala, J. C., Svoboda, J. & Guntaka,R. V. (1998). Cloning of a novel Y-box homologyprotein (chkYB-1HP) cDNA lacking the cold-shockdomain. Biochim. Biophys. Acta, 1395, 1-6.
33. Murray, M. T. (1994). Nucleic acid-binding proper-ties of the Xenopus oocyte Y box protein mRNP3 � 4.Biochemistry, 33, 13910-13917.
34. Delaglio, F., Grzesiek, S., Vuister, G. W., Zhu, G.,Pfeifer, J. & Bax, A. (1995). NMRPipe: a multidimen-sional spectral processing system based on UNIXpipes. J. Biomol. NMR, 6, 277-293.
35. Garrett, D. S., Powers, R., Gronenborn, A. M. &Clore, G. M. (1991). A common sense approach topeak-picking in two-, three-, and four-dimensionalspectra using automatic computer analysis of con-tour diagrams. J. Magn. Reson. 95, 214-220.
36. Whitehead, B., Tessari, M., DuÈ x, P., Boelens, R.,Kaptein, R. & Vuister, G. W. (1997). A 15N-®ltered2D 1H TOCSY experiment for assignment of aro-matic ring resonances and selective identi®cation oftyrosine ring resonances in proteins. J. Biomol. NMR,9, 313-316.
37. Vuister, G. W. & Bax, A. (1992). Resolution enhance-ment and spectral editing of uniformly 13C enrichedproteins by homonuclear broadband 13C-13C decou-pling. J. Magn. Reson. 98, 428-435.
38. Vuister, G. W. & Bax, A. (1993). Quantitative J corre-lation: a new approach for measuring homonuclearthree-bond J(H-NH-alpha) coupling constants innitrogen-15 enriched proteins. J. Am. Chem. Soc. 115,7772-7777.
39. Spera, S. & Bax, A. (1991). Emperical correlationbetween protein backbone conformation and Ca andCb 13C nuclear magnetic resonance chemical shifts.J. Am. Chem. Soc. 113, 5490-5492.
40. Rice, L. M. & BruÈ nger, A. T. (1994). Torsion angledynamics: reduced variable conformationalsampling enhances crystallographic structure re®ne-ment. Protein: Struct. Funct. Genet. 19, 277-290.
41. Stein, E. G., Rice, L. M. & BruÈ nger, A. T. (1997). Tor-sion-angle dynamics as a new ef®cient tool for NMRstructure calculation. J. Magn. Reson. 124, 154-164.
42. BruÈ nger, A. T. (1993). X-PLOR Version 3.1: A Systemfor X-ray Crystallography and NMR, Yale UniversityPress, New Haven, CT.
43. Spronk, C. A. E. M., Bonvin, A. M. J. J., Radha, P. K.,Melacini, G., Boelens, R. & Kaptein, R. (1999). Thesolution structure of lac repressor headpiece 62 com-plexed to a symmetric lac operator sequence deter-mined by NMR and restrained molecular dynamics.Structure, 7, 1483-1492.
44. Laskowski, R. A., Rullmann, J. A. C., MacArthur,M. W., Kaptein, R. & Thorton, J. M. (1996). AQUAand PROCHECK-NMR. Programs for checking thequality of protein structures by NMR. J. Biomol.NMR, 8, 477-486.
45. Vriend, G. (1990). WHAT IF: a molecular modelingand drug design program. J. Mol. Graph. 8, 52-56.
46. Koradi, R., Billeter, M. & WuÈ thrich, K. (1996).MOLMOL: a program for display and analysis ofmacromolecular structures. J. Mol. Graph. 14, 51-55.
47. Johnsson, B., Lofas, S. & Lindquist, G. (1991). Immo-bilization of proteins to a carboxymethyldextran-modi®ed gold surface for biospeci®c interactionanalysis in surface plasmon resonance sensors. Anal.Biochem. 198, 268-277.
48. Persson, B., Stenhag, K., Nilsson, P., Larsson, A.,UhleÂn, M. & Nygren, P.-A. (1997). Analysis of oligo-nucleotide probe af®nities using surface plasmonresonance: a means for mutational scanning. Anal.Biochem. 245, 34-44.
49. Pellencchia, M., Sebbel, P., Hermanns, U., WuÈ thrich,K. & Glockshuber, R. (1999). Pilus chaperone FimC-edhesin FimH interactions mapped by TROSY-NMR. Nature Struct. Biol. 6, 336-339.
Edited by P. E. Wright
(Received 1 October 2001; received in revised form 10 December 2001; accepted 11 December 2001)