the stable evolutionary fixation of a bifunctional tyrosine-pathway protein in enteric bacteria
TRANSCRIPT

FEMS Microbiology Letters 52 (1988) 109-116 109 Published by Elsevier
FEM 03237
The stable evolutionary fixation of a bifunctional tyrosine-pathway protein in enteric bacteria
Suhail Ahmad and Roy A. Jensen
Department of Microbiology and Cell Science, University of Florida, Gainesoille, FL, U.S.A.
Received 8 April 1988 Accepted 11 April 1988
Key words: Enterobacteriaceae; Tyrosine biosynthesis; Bifunctional protein; (Comparative enzymology)
1. SUMMARY 2. INTRODUCTION
The bifunctional T-protein (chorismate mutase- T:cyclohexadienyl dehydrogenase) of L-tyrosine biosynthesis was found to be present in all genera making up the enteric bacteria. The dehydro- genase component of the T-protein was active with both prephenate and L-arogenate, showing it to be a cyclohexadienyl dehydrogenase. The dehy- drogenase component, but not the mutase compo- nent, of the T-protein was feedback-inhibited by L-tyrosine. Unlike some other bifunctional pro- teins, the T-protein has evolved recently and is not ubiquitous. However, once the biochemical spe- cialization of bifunctionality becomes established, the results indicate that such character states are strongly conserved through evolutionary time. Thus, bifunctional proteins can provide particu- larly reliable markers for small (recent origin), intermediate, and large (ancient origin) phylo- genetic clusters.
Correspondence to: Roy A. Jensen, Department of Microbi- ology and Cell Science, 1059 McCarty Hall, IFAS, University of Florida, Gainesville, FL 32611, U.S.A. Florida Agricultural Experiment Station Journal Series No. 8972.
Bifunctional proteins have proven to be stable character states, and thus far, none are known to have originated more than once. Escherichia coli possesses as many as four bifunctional proteins participating in the biosynthesis of aromatic amino acids, these bifunctional proteins enjoying differ- ent hierarchical levels of phylogenetic distribution [1-3]. The bifunctional P-protein (chorismate mutase-P : prephenate dehydratase, E.C. 5.4.99.5- 4.2.1.51) of phenylalanine biosynthesis is of an- cient origin [4]. This bifunctional P-protein (CM- P: PDT) is present in all members of the Super- family-A and Superfamily-B subdivisions of Gram-negative bacteria, and not a single example of its evolutionary loss has yet been documented [4-6].
The two bifunctional proteins that participate in tryptophan biosynthesis exhibit different levels of hierarchical distribution. Phosphoribosylanth- ranilate isomerase : indoleglycerolphosphate syn- thase (E.C. 4.1.1.48) [trp(C)F] is present at least throughout the cluster comprising the enteric lin- eage, while anthranilate synthase:anthranilate 5- phosphoribosylpyrophosphate phosphoribosyl- transferase (E.C. 4.1.3.27-2.4.2.18) [trp(G)D] has been found in the Escherichia/Citrobacter/
0378-1097/88/$03.50 © 1988 Federation of European Microbiological Societies

110
Salmonella/Klebsiella/Enterobacter genera only, and is absent from other enteric bacteria [2,3].
The bifunctional T-protein (chorismate mutase- T : cyclohexadienyl dehydrogenase, E.C. 5.4.99.5- 1.3.1.12) of tyrosine biosynthesis is of relatively recent origin and is present in only a small cluster of Superfamily-B organisms designated as the en- teric lineage (includes traditional enteric bacteria, Aeromonas and Alteromonas) [5]. This bifunc- tional protein (CM-T:CDH) has been shown to be present in E. coli, Klebsiella pneumoniae, Erwinia sp., Serratia marcescens and Proteus mirabilis as representatives of genera within the enteric bacteria [7-9]. Since the enteric bacteria represent a grouping of genera of special microbiological interest, all the genera that make up the enteric bacteria were screened for presence of the bifunc- tional T-protein. The aim of this study was to assess the reliability of this bifunctional protein as a stable character state for marking the identity of genera within the enteric lineage.
3. MATERIALS A N D METHODS
3.1. Bacterial strains and culture conditions Citrobacter freundii ATCC29935, Shigella dys-
enteriae ATCCl1456a, Salmonella enteritidis ATCC13076, Enterobacter aerogenes ATCC13048, Enterobacter cloacae ATCC13047, Enterobacter agglomerans ATCC29915, Enterobacter ag- glomerans ATCC27155 , Serratia rubidaea ATCC27614, Cedecae devisae ATCC33431, Kluyvera ascorbata ATCC33433, Hafnia alvei ATCC13337, Morganella morganii ATCC25830, Edwardsiella tarda ATCC15947, Yersinia enteroc- olitica ATCC9610, Proteus vulgaris ATCC29905 and Providencia alcalifaciens ATCC9886 were ob- tained from American Type Culture Collection, Rockville, MD, U.S.A. Citrobacterfreundii (37 o C), S. enteritidis (37 o C), E. aerogenes (30 o C), E. cloacae (30 o C), E. agglomerans (26 ° C) and S. rubidaea (26 ° C) were grown on M9 medium as described [10]. C. divisae (26°C), K. ascorbata (37 o C), H. alvei (30 o C), Y. enterocolitica (30 o C), and P. alcalifaciens were grown on M9 medium supplemented with 1 mg/1 each of nicotinamide, p-aminobenzoate, D-biotin, calcium pantothenate
and thiamine. S. dysenteriae (37 o C), M. morganii (37 o C) and E. tarda (37 o C) were grown on M9 medium supplemented with the above-mentioned vitamins and 0.1% (w/v) acid-hydrolyzed casein (Difco).
3.2. Preparation of cell extracts and enzyme assays Crude cell-free extracts (free of small mole-
cules) were prepared as described elsewhere [6]. Chorismate mutase was assayed by the method
of Cotton and Gibson [11]. Prephenate dehydro- genase and arogenate dehydrogenase activities were assayed by following the continuous spectro- photofluorometric formation of reduced nico- tinamide adenine dinucleotide (NADH) as de- scribed by Patel et al. [12].
Protein in the crude extract was estimated by the method of Bradford [13] with bovine serum albumin as the standard.
3.3. DE52 column chromatography Approx. 100 mg of crude-extract protein were
applied to a DEAE cellulose (DE52) column (1.5 × 20.0 cm) equilibrated in 10 mM K-phosphate buffer (pH 7.0) containing 1 mM dithiothreitol (DTT). The column was washed with two bed volumes of equilibrating buffer, and then the bound proteins were eluted with 300 ml of a linear gradient of KC1 (0.0-0.35 M) contained in the equilibration buffer. Fractions of 2.2 ml were col- lected.
3.4. Biochemicals and chemicals Amino acids, NAD, NADP, DTT, bovine serum
albumin and Sephadex G-25 were purchased from Sigma (St. Louis, MO). DE52 was purchased from Whatman (Clifton, N J). Prephenate was prepared as the barium salt from culture supernatants of a tyrosine auxotroph of Salmonella typhimurium [14] and was converted to the potassium salt before use. Chorismate was isolated from the accumula- tion medium of a triple auxotroph of K. pneumo- niae 62-1 and purified as the free acid [15]. L- Arogenate was prepared from a triple auxotroph of Neurospora crassa [16]. All other chemicals were of standard reagent grade.
111
4. RESULTS AND DISCUSSION
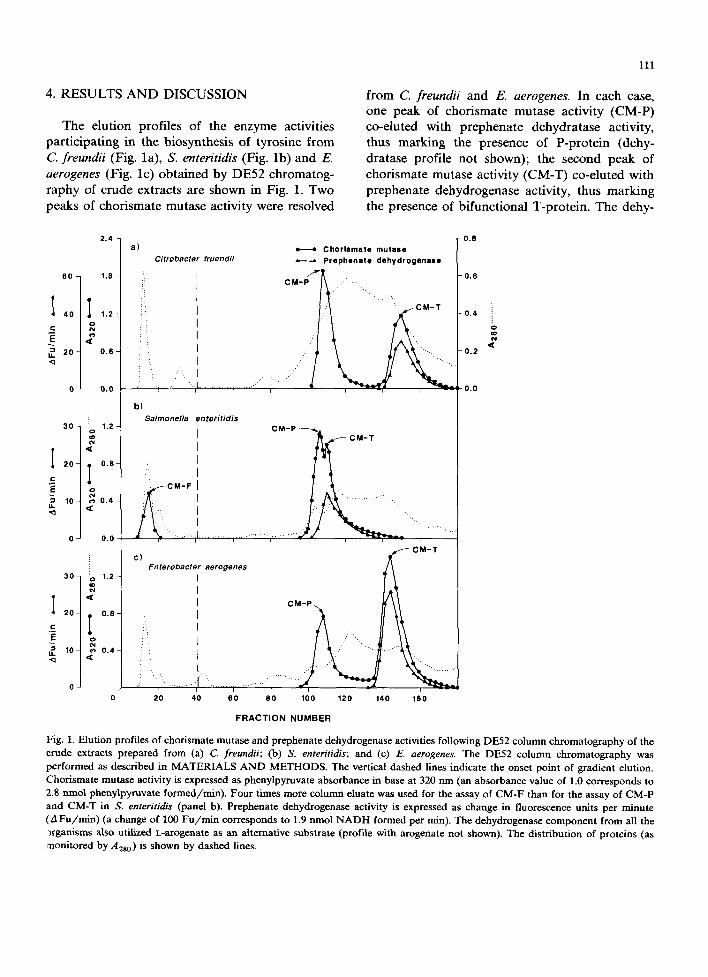
The elution profiles of the enzyme activities participating in the biosynthesis of tyrosine from C. freundii (Fig. la), S. enteritidis (Fig. lb) and E. aerogenes (Fig. lc) obtained by DE52 chromatog- raphy of crude extracts are shown in Fig. 1. Two peaks of chorismate mutase activity were resolved
from C. freundii and E. aerogenes. In each case, one peak of chorismate mutase activity (CM-P) co-eluted with prephenate dehydratase activity, thus marking the presence of P-protein (dehy- dratase profile not shown); the second peak of chorismate mutase activity (CM-T) co-eluted with prephenate dehydrogenase activity, thus marking the presence of bifunctional T-protein. The dehy-
2 .4 - 0 . 8 a) : ~ Chor lsmate mutase
Citrobacter fruendii • * Prephenate dehydrogenase
60 - 1.8 - i!ii: CM-P / t . "~"~ . . , . ' .... ' " " " " " . . - 0.6
40 1.2 - :: 0.4 ._: o
N ~.~ 20 0 . 6 - :. ::". ....i.....y.. 0.2
0 0 .0 " :"i '''' ''~ '""',~'"'""""'i ........ " 0.0 b)
Salmonella enteritidis 3 0 - :o 1 . 2 - C M _ p ~ a
~ - - C M - T <
,
O - o,0 ' " ' - ' " i I . . . . . . . . . . . . . . . . . . i ' " " " ........... i i i i
c ) R ~ C M - T
3 0 - "o 1 , 2 - Enterobacter aerogenes ~ 1
g. ~o- ,2 0 " 4 -
o .- i" ................ " ......... ................ i ............ , , _
20 40 60 80 100 120 140 160
F R A C T I O N N U M B E R
Fig. 1. Elution profiles of chorismate mutase and prephenate dehydrogenase activities following DE52 column chromatography of the crude extracts prepared from (a) C. freundii; (b) S. enteritidis; and (c) E. aerogenes. The DE52 column chromatography was performed as described in MATERIALS AND METHODS. The vertical dashed lines indicate the onset point of gradient elution. Chorismate mutase activity is expressed as phenylpyruvate absorbance in base at 320 nm (an absorbance value of 1.0 corresponds to 2.8 nmol phenylpyruvate formed/rain). Four times more column eluate was used for the assay of CM-F than for the assay of CM-P and CM-T in S. enteritidis (panel b). Prephenate dehydrogenase activity is expressed as change in fluorescence units per minute (A Fu/min) (a change of 100 Fu/min corresponds to 1.9 nmol NADH formed per min). The dehydrogenase component from all the ~rganisms also utilized L-arogenate as an alternative substrate (profile with arogenate not shown). The distribution of proteins (as monitored by A2s0) is shown by dashed lines.
1 1 2
drogenase was also active with L-arogenate (data not shown) as an alternative substrate, thus identi- fying the dehydrogenase component as a broad- specificity cyclohexadienyl dehydrogenase [9]. Similar results were obtained from S. dysenteriae and E. agglomerans 29915 (data not shown).
Data obtained from S. enteritidis (Fig. lb) dif- fered from the above in that an additional third
peak of chorismate mutase designated as CM-F eluted in the wash fractions. It was not associated with any other activity of aromatic biosynthesis.
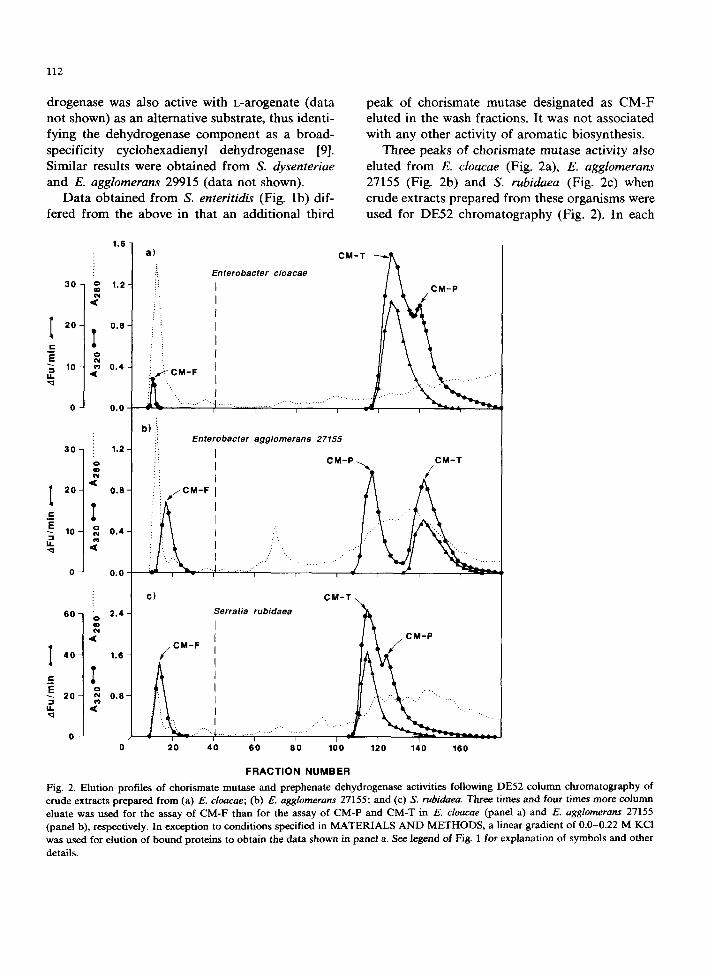
Three peaks of chorismate mutase activity also eluted from E. cloacae (Fig. 2a), E. agglomerans 27155 (Fig. 2b) and S. rubidaea (Fig. 2c) when crude extracts prepared from these organisms were used for DE52 chromatography (Fig. 2). In each
r-
E :3 u. '<3
t- in E :3 u. '*:3
e.
:3 i i
1.6
30 . ~ 1.2
20 0.8
10 ~ 0.4
0 0.0
30 %2
20 0.8
10 - 0.4
0 0.0
60 - 2.4 -
40 - 1.6 -
20 - 0.8 -
0 0
a) C M - T - - " * ' 7 ~ ~ E n t e r o b a c t e r c loacae
ii ~ eM-p
i ~ C M - F
i ......... '"""1 ........... "
b) ~ ii E n t e r o b a c t e r a g g l o m e r a n s 27155
I C M - P -~.~
I f C a - F I
C M - T
C) C M - T
Ser ra t ia r u b i d a e a - -
C M - F
2 0 4 0 6 0 8 0 1 0 0 1 2 0 1 4 0 160
F R A C T I O N N U M B E R
Fig. 2. Elution profiles of chorismate mutase and prephenate dehydrogenase activities following DE52 column chromatography of crude extracts prepared from (a) E. cloacae; (b) E. agglomerans 27155; and (c) S. rubidae~ Three times and four times more column eluate was used for the assay of CM-F than for the assay of CM-P and CM-T in E. cloacae (panel a) and E. agglomerans 27155 (panel b), respectively. In exception to conditions specified in MATERIALS A N D METHODS, a linear gradient of 0.0-0.22 M KC1 was used for elution of bound proteins to obtain the data shown in panel a. See legend of Fig. 1 for explanation of symbols and other
details.
case, the peak of activity eluting in the wash fractions (CM-F) was not associated with any other activity of aromatic biosynthesis, while the remaining two activities proved to be CM-P and CM-T as indicated by the coincidence of their elution profiles with prephenate dehydratase and prephenate dehydrogenase activities, respectively (dehydratase profile not shown). The dehydro- genase enzyme was able to utilize arogenate as an
113
alternative substrate in all 3 organisms (data not shown).
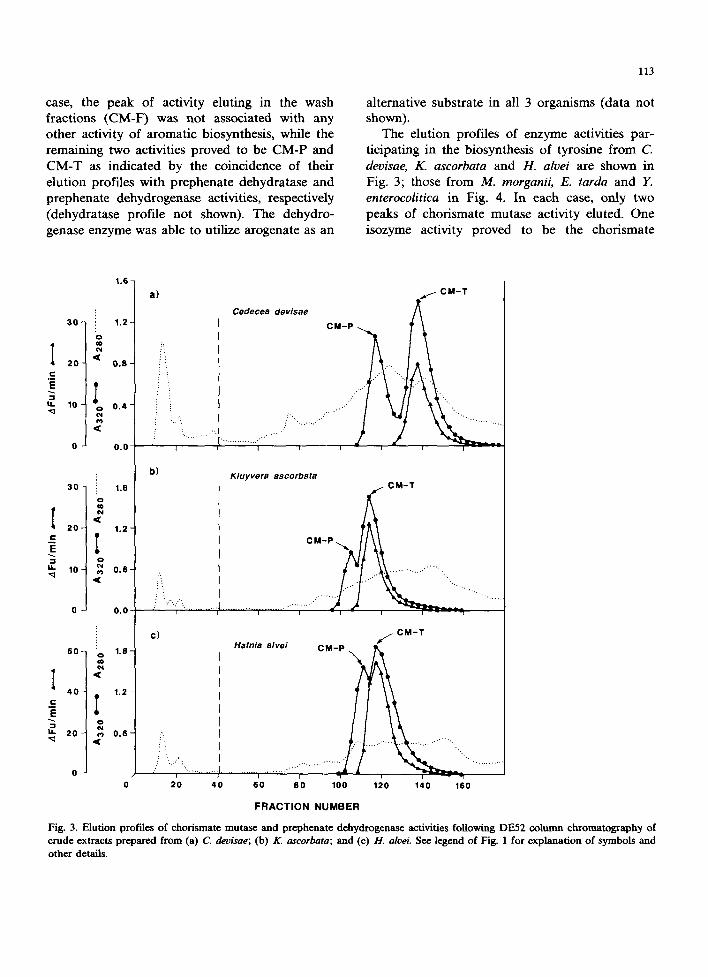
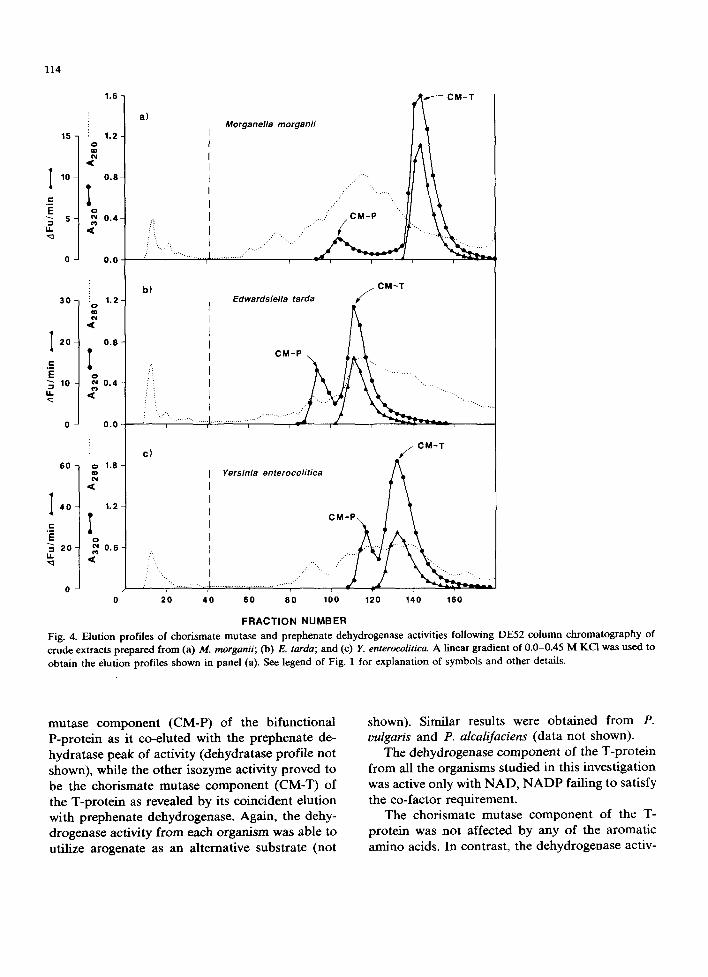
The elution profiles of enzyme activities par- ticipating in the biosynthesis of tyrosine from C. deoisae, K. ascorbata and H. alvei are shown in Fig. 3; those from M. morganii, E. tarda and Y. enterocolitica in Fig. 4. In each case, only two peaks of chorismate mutase activity eluted. One isozyme activity proved to be the chorismate
I ¢ .
E - s
U . . , q
I
I t .
I c
E - 1
I I . , q
1.6-
30- :: 1.2- o ¢o
20- io 0.8-
10 - 0.4 -
0 0.0
3 0 - :: 1 . e -
N d f
20- T 1.2-
10 - ~ 0.6-
0 0.0
60 - o 1.8-
<
40- i ) 1.2 -
20 - 0.6 -
o o
a)
: !
b)
CM-T
Cedecea devisae A CM-P
I I
/:
i
Kluyvera ascorbata CM-T
I i - i r I ~ - - 1
C)
i ~ ...
2~0 40
Hafnia alvei CM-P \
............. .....""'"'""'"; 60 80 100
f CM-T
i l - - "1
120 140 160
FRACTION NUMBER
Fig. 3. Elution profiles of chorismate mutas¢ and prephenate dehydrogenas¢ activities following DE52 column chromatography of crude extracts prepared from (a) C. devisae; (b) K. ascorbata; and (c) 1t. alvei. See legend of Fig. 1 for explanation of symbols and other details.
1.6
30
15 : 1.2
I 10 0.8
E B s - o.4 U.
O - 0 . 0
b ) Edwardsiella tarda ~ C M- T
I 2O
c E = 10 g.
60
I 40
=-
E = 2O U.
"o 1.2
.2 0.8
0.4
0.0
1.8
1.2
~0.6
a) Morganella morganii
...."/'"'",.,... .....
... ..
.... , .,..' C M - P '"',
/! .......... ;
114
C) ~ C M - T
Yersinia enterocolitica
20 40 60 80 100 120 140 160
FRACTION NUMBER Fig. 4. Elution profiles of chorismate mutase and prephenate dehydrogenase activities following DE52 column chromatography of crude extracts prepared from (a) M. morganii; (b) E. tarda; and (c) Y. enterocolitica. A linear gradient of 0.0-0.45 M KCI was used to obtain the elution profiles shown in panel (a). See legend of Fig. 1 for explanation of symbols and other details.
mutase component (CM-P) of the bifunctional P-protein as it co-eluted with the prephenate de- hydratase peak of activity (dehydratase profile not shown), while the other isozyme activity proved to be the chorismate mutase component (CM-T) of the T-protein as revealed by its coincident elution with prephenate dehydrogenase. Again, the dehy- drogenase activity from each organism was able to utilize arogenate as an alternative substrate (not
shown). Similar results were obtained from P. oulgaris and P. alcalifaciens (data not shown).
The dehydrogenase component of the T-protein from all the organisms studied in this investigation was active only with NAD, N A D P failing to satisfy the co-factor requirement.
The chorismate mutase component of the T- protein was not affected by any of the aromatic amino acids. In contrast, the dehydrogenase activ-

115
Table 1
Inhibition of cyclohexadienyl dehydrogenase by L-tyrosine. a
Organism % Inhibition by b
0.1 mM 0.5 mM L-Tyrosine L-Tyrosine
Citrobacter freundii 71 97 Salmonella enteritidis 85 95 Enterobacter aerogenes 81 100 Enterobacter cloacae 68 95 Enterobacter agglomerans 27155 37 92 Serratia rubidaea 42 94 Cedecae deoisae 86 98 Kluyoera ascorbata 80 97 Hafnia alvei 54 96 Morganella morganii 93 100 Edwardsiella tarda 72 96 Yersinia enterocolitica 77 99
Cyclohexadienyl dehydrogenase activities were recovered from DE52 column fractions (refer to column profiles shown in Figs. 1-4). The inhibition was studied with prephenate as substrate at a final concentration of 0.1 raM. Comparable inhibition was also obtained when L-arogenate was used as an alternative substrate (data not shown).
b Other aromatic amino acids had no effect on the dehydro- genase activity when either prephenate or L-arogenate was utilized as substrate (data not shown).
single instance of their loss has been documented in the phylogenetic cluster where they have been found. Perhaps the efficient channeling of inter- mediates in the microenvironment of the two ac- tive sites catalyzing two consecutive reactions ex- erts overwhelming selective pressure for their maintenance. These findings also suggest that the fortuitous appearance of new genes specifying bi- functional proteins unequivocally establishes points of divergence at differing hierarchical levels in the evolution of microorganisms [4]. Earlier observations that the dehydrogenase activity of tyrosine biosynthesis present in Superfamily-B organisms, whether present as a monofunctional activity or as a component of the bifunctional T-protein, is a cyclohexadienyl dehydrogenase since it can utilize both prephenate and L- arogenate [9,17] are further supported.
ACKNOWLEDGEMENT
These studies were supported by grant DMB- 8615314 from the National Science Foundation.
ity was tightly regulated by t-tyrosine when as- sayed with prephenate as substrate (Table 1). The dehydrogenase activity from each organism was also completely inhibited by 0.5 mM L-tyrosine when assayed with L-arogenate as substrate (data not shown). Other aromatic amino acids had no effect upon activity.
These results and some earlier observations [8] clearly demonstrate that the bifunctional T-pro- tein of tyrosine biosynthesis is uniformly distrib- uted throughout the enteric bacteria. Unlike the P-protein which is widely distributed among Gram-negative bacteria (ancient origin), the T-protein is restricted in distribution to the enteric lineage of bacteria (recent origin). Thus, a spec- trum of bifunctional proteins of differing ubiquity is becoming evident. The data presented in this paper together with other comprehensive studies carried out on the bifunctional P-protein [1,5] support the conclusion that bifunctional proteins are highly conserved character states, since not a
REFERENCES
[1] Jensen, R.A. (1985) Mol. Biol. Evol. 2, 92-108. [2] Zalkin, H. (1980) in Mul t i func t iona l Prote ins
(Bisswanger, H. and Schmincke-Ott, E., Eds.), pp. 123-149. Wiley, New York.
[3] Crawford, I.P. (1980) in Multifunctional proteins (Bi- sswanger, H. and Schmincke-Ott, E., Eds.), pp. 151-173. Wiley, New York.
[4] Ahmad, S. and Jensen, R.A. (1986) Trends Biochem. Sci. 11,108-112.
[5] Ahmad, S. and Jensen, R.A. (1988) Origins Life 18, 41-57. [61 Abroad, S. and Jensen, R.A. (1988) Curt. Microbiol. 16,
295-302. [7] Koch, G.L.E., Shaw, D.C. and Gibson, F. (1970) Biochim.
Biophys. Acta 212, 375-386. [8] Koch, G.L.E., Shaw, D.C. and Gibson, F. (1971) Biochim.
Biophys. Acta 229, 795-804. [9] Ahmad, S. and Jensen, R.A. (1988) Mol. Biol. Evol. 5,
282-297. [10] Winlder, U.K. and Stuckman, M. (1979) J. Bacteriol. 138,
663-670. [11] Cotton, R.G.H. and Gibson, F. (1965) Biochim. Biophys.
Acta 100, 76-88.

116
[12] Patel, N., Pierson, D.L. and Jensen, R.A. (1977) J. Biol. Chem. 252, 5839-5846.
[13] Bradford, M.M. (1976) Anal. Biochem. 72, 248-254. [14] Dayan, J. and Sprinson, D.B. (1970) Methods Enzymol.
17A, 559-561. [15] Gibson, F. (1970) Methods Enzymol. 17A, 362-364.
[16] Zamir, L.O., Jensen, R.A., Arison, B.H., Douglas, A.W., Albers-Schonberg, G. and Bowen, J.R. (1980) J. Am. Chem. Soc. 102, 4499-4504.
[17] Ahmad, S. and Jensen, R.A. (1986) Arch. Microbiol. 147, 8-12.