the yeast 5s rrna binding robosomal protein yl3 is phosphorylated in vivo
TRANSCRIPT

142 Biochimica et Biophysica Acta. 1087 (1990) 142- 146 Elsevier
BBAEXP 92172
The yeast 5S rRNA binding robosomal protein YL3 is phosphorylated in vivo
Francisco Campos, Matilde Corona-Reyes and Samuel Zinker Departamento de Gen~tica y Biologla Molecular, Centro de Investigaclon y de Estudios A vanzados de[ 1. P.N.. Mexico (i(v (Mexico)
(Received 8 May 1990)
Key words: Ribosomal protein; Yeast; Phosphorylation
The 5S rRNA-ribosomal protein YL3 ribonueleoprotein particle was isolated from yeast cells labeled with [3z Plortho- phosphate. The protein moiety was purified and found to be radioactive. Labeled phosphoserine was detected after partial hydrolysis of the protein. Up to two phosphate residues are sterified per mole of YL3 ribosomal protein.
Introduction
The role of individual ribosomal components, or the structural features of the ribosome which are essential for protein synthesis, requires detail knowledge of pro- tein-protein and protein-rRNA interaction in the ribosome. The ribonucleoprotein (RNP) complex re- leased from the large ribosomal subunits by treatment with EDTA provides a simplified system to carry on such studies. The RNP complex composition is 5S rRNA associated to three [1,2] two [3] or one protein [4-6] in Escherichia coli, archaebacteria, and eukaryotes, respec- tively. This complex exhibits both GTPase and ATPase activities [7,8], it is implicated in ribosomal subunit association [9,10], and it also represents a precursor to ribosome assembly in mammalian cells [11].
In Saccharomyces cerevisiae, the 5S rRNA binding protein, known as YL3 [12] or L la [13], modulates the synthesis of the 5S rRNA moiety in vitro [141. The amino-terminal region of the protein confers the specificity for binding to 5S rRNA whereas the carbox- yl terminus provides the RNA-binding site [3]. This later region consists of 47 amino acids, 11 of which, 8 lysine and 3 arginine residues, are basic [15]. Chemical modifications of these basic residues abolish the capac- ity of protein YL3 to form stable interactions with 5S rRNA in vitro [16]. We here report that ribosomal protein YL3 can be chemically modified in vivo. The
Abbreviation: RNP, ribonucleoprotein.
Correspondence: S. Zinker, Departamento de Genrtica, y Biologia Molecular, CINVESTAV-IPN, Apartado Postal 14-740 07000 Mrxico, D.F.
modification consists in phosphorylation of serine re- sidues.
Materials and Methods
Strain and growth conditions. S. cerevisiae strain Hal (ATCC22244) was grown up to mid-log phase in YEP medium containing per liter: peptone 20 g, yeast extract 10 g and glucose 20 g. For labeling with 32po 4 cells were grown in phosphate-free medium [17]. In this medium, cells incorporate about 80% of the isotope during the first 60 rain.
Isolation o f ribosomes, and ribosomal subunits, Cells were harvested and washed once with sterile water. They were resuspended in buffer A (0.1 M NaCI, 0.03 M MgC12, 0.01 M Tris, pH 7.4), mixed with glass beads (0.45-0.5 mm diameter) and disrupted by alternate 30 s periods of vigorous vortexing and incubation in ice. Buffer A was added to the suspension up to 10% the original culture volume. Sodium deoxycholate was ad- ded to a final concentration of 0,5% and after 5 min in ice, Brij 58 (polyoxyethylene 20-cetyl ether) was added to a final concentration of 0.5%. After 5 more rain in ice, the suspension was centrifuged at 27000 × g. The supernatant was layered over a 4 ml cushion of 15% sucrose containing 5% (NH4)2SO 4 in buffer A and centrifuged at 214000 x g for 3 h at 4°C. For ribosomal subunits the pellet was resuspended in buffer B (0.1 M NaC1, 10 5 M MgC1, 0.01 M Tris, pH 7.4), layered over 10-25% sucrose gradients in buffer B and centrifuged at 76000 × g for 18 h at 4°C. At the end of the run the gradients were fractionated into 1.0 ml fractions. The absorbance at 260 nm was continuously recorded. When needed, fractions containing 60S ribosomal subunits
0167-4781/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

were pooled and pelleted by centrifugation at 214 000 x g for 3 h at 4°C.
Isolation of the 5S rRNA-protein complex from the 60S ribosomal subunits. The pellet of 60S ribosomal subunits was resuspended in 25 mM EDTA and stirred for 20 min at 4°C. Then, it was centrifuged for 20 h at 214000 x g and 4°C [6].
Phenol-chloroform extraction of the 5S rRNA-YL3 ribosomal protein complex. To the 5S rRNA-YL3 ribo- somal protein complex MgC12 was added up to 8 mM followed by 0.7 vol. of 95% ethanol at - 2 0 ° C . The solution was stored at - 2 0 ° C overnight. The suspen- sion was centrifuged at 27000 x g and 4°C. The pellet was resuspended in 1.0 ml of 0.1 M LiC1, 10 mM EDTA, 0.1% SDS, 10 mM Tris (pH 7.4). An equal volume of phenol/chloroform (1:1, v /v) was added, stirred for 10 min at room temperature and centrifuged. The aqueous phase was removed, LiC1 was added up to 0.5 M followed by 2 volumes of 95% ethanol. The solution was stored overnight at - 2 0 ° C to precipitate the RNA. The protein from the interphase was rescued, dialyzed against 0.5% acetic acid and stored lyophilized at 4°C until its further analysis.
Extraction of total ribosomal proteins. The 241 000 x g pellet was resuspended in buffer B. Ribosomal proteins were extracted by addition of 0.1 vol. of 1 M MgC12 and 2 vol. of glacial acetic acid [18]. After stirring for 30 min at 4°C, the samples were centrifuged at 27 000 X g and 4°C for 15 min. The supernatant was dialyzed against 0.5% acetic acid at 4°C and lyophilized.
High voltage electrophoresis of amino acids. The lyophilized ribosomal protein YL3 was dissolved in 1.0 ml of 6 M HC1 and transferred into a 5 ml ampule. The ampule was sealed under vacuum and incubated at 100°C for 4 h. After drying the HC1 under vacuum, the residue was dissolved in distilled water, and applied onto Whatman (3MM) paper in the presence of nonra- dioactive phosphoserine and phosphotreonine as inter- nal markers. High-voltage electrophoresis was carried out at 2 kV for 2 h toward the cathod using 25% formic acid, 7.8% acetic acid (pH 1.85) as chambers buffer [19].
Electrophoresis of rRNA. Electrophoresis of rRNA was performed as described by Kadowaki and Halvor- son [20]. Gels (80 mm x 5 mm) were cast in 15-cm long tubes and pre-ran for 30 min at 2.5 mA per tube. Then, samples were applied and ran at 5 mA per tube for 150 min. Gels were sliced into 1 mm slices. Each gel slice was digested with Protosol (New England) and the radioactivity was determined.
Electrophoresis of ribosomal proteins. (a) One-dimen- sional polyacrylamide gel electrophoresis of ribosomal proteins in the presence of SDS (SDS-PAGE) was per- formed as described [21]. Acrylamide concentration was 4% for the stacking and 10% for the resolving gels. (b) Two-dimensional SDS-PAGE. The first dimension at pH 3.2 was as described by Panyim and Chalkley [22]
143
into 7.5% acrylamide cylindrical gels. After running, the gels were equilibrated in 1% SDS, 0.5 M Tris (pH 6.8) with gentle stirring for 60 min. The second dimension was performed as described above under one-dimen- sional SDS-PAGE.
Phosphate determination was as described by Ames and Dubin [23]. Silver staining of proteins was accord- ing to Wray et al. [24].
Results
In S. cerevisiae in vivo phosphorylation of five ribo- somal proteins was reported some time ago [21]. Among them, P1 resembles ribosomal protein YL3 in its ap- parent molecular weight, its location in the 60S ribo- somal subunit and its relative insolubility at or near neutral pH [25]. Therefore, the phosphorylation of ribosomal protein YL3 was investigated in vivo. To that end, yeast cells were labeled for 60 min with 32p and 60S ribosomal subunits isolated as described under Materials and Methods. The radioactive material re- leased from the large subunits after treatment with EDTA is shown in Fig. 1. The high radioactive back- ground toward the heavier fractions was consistently obtained and was not further analyzed. Fractions 3 to 7 which contained the main peak were pooled, and an aliquot was separated for direct analysis by electro- phoresis in SDS-PAGE (see below). The rest of the sample was extracted with phenol/chloroform. About 5000 cpm of 32p-labeled material from the aqueous phase was precipitated with ethanol, mixed with marker tritium-labeled small rRNA and coelectrophorized into polyacrylamide gels. As shown in Fig. 2, a discrete peak of 32P-labeled RNA was detected in the region of 5S rRNA. The amount of 32p recovered from this peak account for about 95% of the initial input.
The protein associated to the 5S rRNA moiety is depicted in Fig. 3. A single band was detected before (Fig. 3, lane 2) or after (Fig. 3, lane 3) phenol/chloro-
x
0
Fraction number
Fig. 1. Sedimentation profile of the radioactive material released by EDTA from the 60S ribosomal subunits. The pellet of labeled ribo- somal subunits was resuspended in 10 ml of 25 mM EDTA, pH 7.0, and centrifuged at 4°C for 20 h at 226000x g [6]. Fractions of 1.0 ml were collected and aliquots from each fraction were TCA-precip-
itated.
144
6 -
x
E t ' t 0 2 -
0 0
4s
°, , ' i i
A A s , _ _ _ ' , ) I ' I I l l I ] Q"
[jL ~ " " l A L I ' '~/ " , ¢') 1(3 2 0 50 4 0 50 6 0 70 8 0
Froction number
Fig. 2. Gel electrophoresis of rRNA. The r R N A extracted f rom the material shown in Fig. 1 was analyzed by P A G E in the presence of
tritiated r R N A markers.
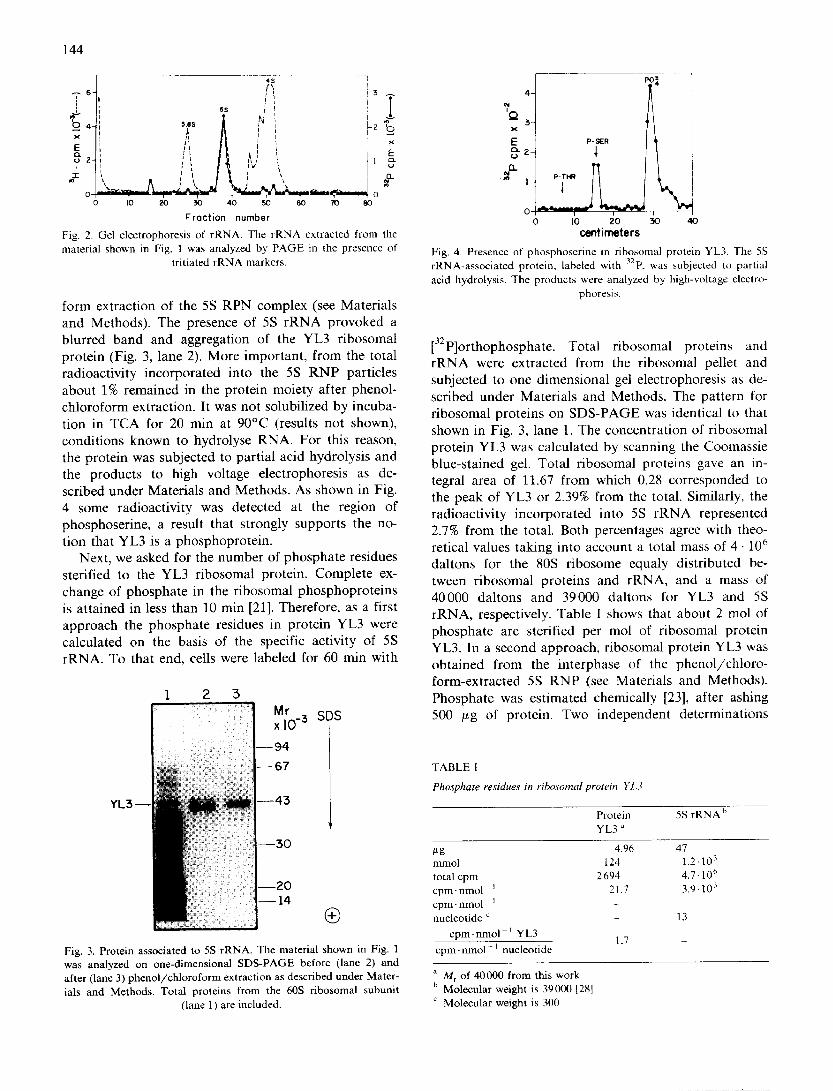
form extraction of the 5S RPN complex (see Materials and Methods). The presence of 5S rRNA provoked a blurred band and aggregation of the YL3 ribosomal protein (Fig. 3, lane 2). More important, from the total radioactivity incorporated into the 5S RNP particles about 1% remained in the protein moiety after phenol- chloroform extraction. It was not solubilized by incuba- tion in TCA for 20 min at 90°C (results not shown), conditions known to hydrolyse RNA. For this reason, the protein was subjected to partial acid hydrolysis and the products to high voltage electrophoresis as de- scribed under Materials and Methods. As shown in Fig. 4 some radioactivity was detected at the region of phosphoserine, a result that strongly supports the no- tion that YL3 is a phosphoprotein.
Next, we asked for the number of phosphate residues sterified to the YL3 ribosomal protein. Complete ex- change of phosphate in the ribosomal phosphoproteins is attained in less than 10 min [21]. Therefore, as a first approach the phosphate residues in protein YL3 were calculated on the basis of the specific activity of 5S rRNA. To that end, cells were labeled for 60 min with
1 2 5
)-3 SDS
YL3
®
Fig. 3. Protein associated to 5S rRNA. The material shown in Fig. 1 was analyzed on one-d imens ional S D S - P A G E before (lane 2) and after (lane 3) p h e n o l / c h l o r o f o r m extract ion as described unde r Mater- ials and Methods . Tota l proteins f rom the 60S r ibosomal subuni t
(lane 1) are included.
PO~,
x P-SlUR
0 1(3 20 50 40 centimeters
Fig. 4. Presence of phosphoser ine in r ibosomal protein YL3. ] 'he 5S rRNA-assoc ia t ed protein, labeled with 32p. was subjected to partial
acid hydrolysis . The products were analyzed by high-voltage electro- phoresis.
[32p]orthophosphate. Total ribosomal proteins and rRNA were extracted from the ribosomal pellet and subjected to one dimensional gel electrophoresis as de- scribed under Materials and Methods. The pattern for ribosomal proteins on SDS-PAGE was identical to that shown in Fig. 3, lane 1. The concentration of ribosomal protein YL3 was calculated by scanning the Coomassie blue-stained gel. Total ribosomal proteins gave an in- tegral area of 11.67 from which 0.28 corresponded to the peak of YL3 or 2.39% from the total. Similarly, the radioactivity incorporated into 5S rRNA represented 2.7% from the total. Both percentages agree with theo- retical values taking into account a total mass of 4 • 106 daltons for the 80S ribosome equaly distributed be- tween ribosomal proteins and rRNA, and a mass of 40000 daltons and 39000 daltons for YL3 and 5S rRNA, respectively. Table I shows that about 2 tool of phosphate are sterified per mol of ribosomal protein YL3. In a second approach, ribosomal protein YL3 was obtained from the interphase of the phenol/chloro- form-extracted 5S RNP (see Materials and Methods). Phosphate was estimated chemically [23], after ashing 500 ttg of protein. Two independent determinations
T A B L E I
Phosphate residues in ribosomal protein YL3
Protein 5S r R N A b
YL3 ~
ttg 4.96 47 mmol 124 1.2- 10 :t
total cpm 2 694 4.7-10 C~ cpm- nmol i 21.7 3.9-103
cpm. nmol - 1
nucleotide " 13
c p m . n m o l 1 YL3 1.7
cpm- nmol - 1 nucleot ide
a Mr of 40000 f rom this work b Molecular weight is 39 000 [28]
Molecular weight is 300

SDS
®
, - pH 5.2 ®
STAIN
145
Fig. 7. Two-dimensional PAGE of protein YL3. Pure ribosomal protein YL3, kindly provided by Dr. Ross N. Nazar, was analyzed by one dimensional (A) or two dimensional PAGE (B). (C) Protein spots in (B) were excised and digested with trypsin. After one-dimensional
SDS-PAGE the gel was stained with silver [24].
AUTORADIOGRAPH Fig. 5. Two-dimensional PAGE of total ribosomal proteins. Cells were labeled with 32p. Total ribosomal proteins were isolated and analyzed by pH 3.2 X SDS-PAGE. The gel was stained with Coomassie blue
(A), dried and subjected to autoradiography (B).
showed 1.44 and 1.6 mol of phosphate per mol of YL3 protein. These numbers closely agree with that of Table I.
In our hands, YL3 remains near or at the origin of conventional pH 8.6 x pH 4.5 or pH 5.0 x SDS two-di- mensional PAGE. When pure, it enters into solution at acid pH [25]. This property was considered to explore the behavior of YL3 on pH 3.2 X SDS two-dimensional PAGE. The results of such an analysis, using total 32p labeled ribosomal proteins, are shown in Fig. 5. A1-
SDS - - ~ 3.2 • ®
03
Fig. 6. Ribosomal proteins from ribosomes treated with EDTA. Ribosomes were washed with 25 mM EDTA (pH 7.4) and sedimented by centrifugation. Ribosomal proteins were extracted from the pellet and analyzed by pH 3.2 X SDS two-dimensional PAGE. The arrow
points to the migration site of YL3.
though a substantial amount of YL3 migrated into the first dimension of the gel, some of it remained at the origin interconnected with the former by a bridge-like strike. From the two protein spots and their bridge-like connection, unexpectedly, only the one at the origin was radioactive (Fig. 5B). As shown in Fig. 6, neither of them were detected after EDTA extraction of the ribo- somal pellet, suggesting that no other protein com- igrated in the region of ribosomal protein YL3. Further- more, pure ribosomal protein YL3 (kindly provided by Dr. Ross N. Nazar) gave the same split pat tern when analyzed by pH 3.2 X SDS two dimensional PAGE (Fig. 7B), indicating again that this protein is chemically heterogeneous.
Discussion
In S. cereoisiae, we reported some time ago [21] on the in vivo phosphorylation of a ribosomal protein from the 60S ribosomal subunit with an M r of 40000 by one-dimensional SDS-PAGE analysis. The eukaryotic 5S rRNA binding protein named YL3 [12], L la [13] or L5 [26] shares all of the physicochemical properties with our former P1, i.e., location on the 60S ribosomal sub- unit, similar molecular weight and poor migration into two-dimensional gels. In this study we show that when yeast cells are labeled with [32p]orthophosphate, the 5S rRNA-associated protein becomes radioactive. From the fact that the ribosomal protein associated to 5S rRNA is phosphorylated (Fig. 4), and since no other protein was detected in the same region after EDTA extraction of ribosomes (Fig. 6), it can be concluded that almost certainly P1 and YL3 are the same protein. The number of phosphate residues sterified to YL3 was calculated by two independent methods. Firstly, as a function of the specific activity of 5S rRNA taking advantage from

146
the rapid turnover of phosphate in the protein [21]. Secondly, by chemical analysis after ashing the protein [23]. The results of both methods were in close agree- ment, and led us to conclude that up to two phosphate residues are sterified per mol of protein (Table I). One of them, at least, must be bound to a serine residue (Fig. 4).
The complete primary structure of ribosomal protein YL3 from S. cerevisiae has not been solved yet, but that for rat liver ribosomal protein L5, equivalent to YL3, was published recently [26]. The protein has 298 amino acid residues. The primary structure flanking serine residue 287 is: Lys-Lys-Ala-Ser-Phe which resembles that of Arg-Arg-Ala-Ser-Phe, a synthetic peptide effi- ciently phosphorylated by the catalytic subunit of a cAMP-stimulated protein kinase from rat liver [27]. Therefore, it is tempting to predict that serine residue 287 in ribosomal protein L5 will be found phospho- rylated in vivo, and also a similar sequence to be found in ribosomal protein YL3 from S. cerevisiae.
The radioactive fraction remaining at the origin of the two-dimensional PAGE, as well as the bridge-like strike binding the phosphorylated and dephospho- rylated forms of ribosomal protein YL3 (Fig. 5A), indi- cate that phosphate esters in this protein may be labile. Dephosphorylation could take place either during ex- traction from the total ribosomal pellet or throughout electrophoresis in the first dimension at pH 3.2. Alter- natively, it is possible that both the phosphorylated and dephosphorylated forms of ribosomal protein YL3 coexist in the ribosome. This remains to be clarified.
Acknowledgements
We thank Rosa Maria Cruces for typing this manuscript. This work was supported in part by grant ICEXCNA-050570, from the Consejo Nacional de Ciencia y Tecnologia (M6xico).
References
1 Erdmann, V.A. (1976). Progr. Nucleic Acid Res. Mol. Biol. 18, 45-90.
2 Chen-Schmeisser, U. and Garret, R.A. (1977) FEBS Lett. 74, 287 291.
3 Smith, N., Matheson, A.T., Yaguchi, M., Willick, G.E. and Nazar, R.N. (1978). Eur. J. Biochem. 89, 501-509.
4 Blobel, G. (1971). Proc. Natl. Acad. Sci. USA 68, 1881-1885. 5 Terao, K., Uchimi, T. and Ogata, K. (1982) Biochim. Biophys.
Acta 697, 20-24. 6 Nazar, R.N. (1979) J. Biol. Chem. 254, 7724-7729. 7 Grummt, F., Grummt, I. and Erdmann. V.A. (1974) Eur. J.
Biochem. 43, 343-348. 8 0 g a t a , K., Terao, K. and Uchiumi, T. (1980) J. Biochem. 87,
517-524. 9 Azad, A.A. and Lane, B.G. (1973) Canad. J. Biochem. 51, 1669-
1672. 10 Metspalu, E., Ustav, M. and Villeins, R. (1983) FEBS Lett. 153,
125-127. 11 Steitz, J.A., Berg, C., Hendrick, J.P., La Branche-Chabot, H.,
Metspalu, A., Rinke, J. and Yorio, T. (1988) J. Cell Biol. 106, 545-556.
12 Nazar, R.N., Yaguchi, M., Willick, G.E., Rollin, C.F. and Roy. C. (1979) Eur. J. Biochem. 102, 573-582.
13 Michael, S., Traut, R.R. and Lee, J.C. (1983) Mol. Gen. Genet. 191, 251-256.
14 Brow, D.A. and Geiduschek, E.P. (1987). J Biol. Chem. 262, 13953 13958.
15 Yaguchi, M., Rollin, C.F., Roy, C. and Nazar, R.N. (1984) Eur. ,l. Biochem. 139, 451-457.
16 Viogue. A., Hernandez, G. and Palacian, E. (19871 Mol. Cell Biochem. 76. 141-146.
17 Rubin, G.M., (1973) J. Biol. Chem. 248, 3860-3875. 18 Hardy, J.S., Kurland, C.G., Voynon, P. and Mora, G. (1969)
Biochemistry 8, 2897-2905. 19 Labrie, F., Lamaine, S., Poitier, G., Pelletier, G. and Baucher, R.
(1971) J. Biol. Chem. 246, 731t-7317. 20 Kadowaki, K. and Halvorson, H.O. (1971) J. Bact. 105, 826-830. 21 Zinker, S. and Warner, J.R. (1976) J. Biol. Chem. 251, 1799-1807. 22 Panyim, S. and Chalkley, R. (1969) Arch. Biochem. Biophys. 130,
337 346. 23 Ames, B.N. and Dubin, D.T. (1960) J. Biol. Chem. 235, 769-775. 24 Wray, W., Boulikas, T., Wray, V. and Hancock, R. (198l) Anal.
Biochem. 118, 197-203. 25 Willick, G.E., Nazar, R.N. and Van, N.T. (1980) Biochemistry 19.
2738-2742. 26 Yueng-Lin, Ch., Lin, A., McNally, J. and Wool, I.G. (1987) J.
Biol. Chem. 262, 12879 12886. 27 Engstr~Sm, k. (1980). Recently-discovered systems of enzyme regu-
lation by reversible phosphorylation (Cohen, P., ed.),Vol. 1, 11-31, Elsevier/North Holland Biomedical Press, Amsterdam.
28 Helser, T.L. and McLaughlin, C.S. (1975) J. Biol. Chem. 250, 2003-2007.