tight-junctional strands first appea in regionr s where ... · differentiating olfactory epithelium...
TRANSCRIPT

Tight-junctional strands first appear in regions where three cells meet in
differentiating olfactory epithelium: a freeze-fracture study
BERT Ph. M. MENCO
Department of Neurobiology and Physiology, 0. T. I logon Hall, Northwestern University, Evanston, 1L 6020S, USA
Summary
Tight junctions of the olfactory epithelium of ratembryos were studied at the 14th day of gestationand during their subsequent development. Twodifferent epithelial morphologies could be dis-tinguished at the 14th gestational day. In onegroup of embryos the epithelial surface appearedundifferentiated, with tight-junctional strandsfound exclusively in regions where three cellsmet. The main orientation of these strands is in adirection parallel to the longitudinal orientationof the epithelial cells. These junctions resembletight junctions that interconnect three cells, i.e.tricellular tight junctions, in that respect. How-ever, unlike these the junctions mainly havesingle strands of particles, whereas tricellularjunctions usually consist of paired strands ofparticles. Tight-junctional strands were com-pletely absent in areas where two cells met. Theseareas, i.e. those of incipient bicellular tight junc-tions, had gap-junction-like aggregates of intra-membranous particles. Another group of 14-day-old embryos displayed a differentiating olfactoryepithelial surface with bicellular as well as tri-cellular tight-junctional strands. The latter oneswere paired. Here too the tight-junctional belts
displayed some gap-junction-like aggregates ofparticles, but there were considerably fewer ofthese than earlier. As one or the other tight-junctional appearance was always seen in asingle freeze-fracture replica, it is reasonable toassume that the two tight-junctional appearancesreflect a sequential pattern of differentiationpeculiar to the whole surface of the olfactoryepithelium, i.e. to surfaces of receptor cells aswell as to surfaces of supporting cells. It wouldappear that, at the onset of olfactory epithelialdifferentiation, tight junctions first interconnectcells in regions where three cells meet and thattricellular strand formation precedes the forma-tion of bicellular strands. When strands werepresent at the 14th day of embryonic develop-ment, their numbers were lower than those foundlater. However, strand packing, expressed as thedensity per micrometre of strands parallel to theepithelial surface, increased beginning at the 16thday of embryonic development.
Key words: tight junctions, freeze-fracturing, olfactory
epithelium, pre-natal development, rat.
Introduction
At E14 (El = day that the dams were sperm-positive;E23 = PI or day of birth) the developing olfactoryepithelium of the rat undergoes major transformations.Until E14 the epithelial surface of the rat's olfactoryreceptor cells has a globular topography; the olfactorysupporting cells appear flat. Neither cell type hassurface structure other than primary cilia. E14 embryosdisplaying this surface morphology were called E14.1(Menco & Farbman, 1985a,6; Menco, 1988). Anothergroup of E14 embryos has olfactory epithelial surfacesJournal of Cell Science 89, 495-505 (1988)Printed in Great Britain © The Company of Biologists Limited 1988
that are beginning to acquire an appearance typical ofthat of the olfactory epithelium in adults. The epi-thelium displays olfactory dendrites that acquire sec-ondary cilia with prolonged development. These den-drites are surrounded by microvillous supporting cells.This stage was called E14.2 (Menco & Farbman,1985a,6; Menco, 1988).
Each olfactory receptor cell is linked at first to bothreceptor cells and supporting cells, and later only tosupporting cells. As the circumference of dendriticendings of vertebrate olfactory receptor cells is small, alarge proportion of their membranes can be exposed by
495

freeze-fracturing. Consequently, one often finds sev-eral tricellular in addition to bicellular tight junctionsin freeze-fracture replicas of olfactory receptor cells.Tricellular tight junctions are located where three cellsmeet and bicellular tight junctions interlink two cells.From studies of other tissues it is known that bicellulartight junctions consist of several strands of particles orshort bars. Tricellular tight junctions consist of isolatedsets of paired strands. These have an orientationparallel to the longitudinal axes of epithelial cells andperpendicular to the main orientation of the bicellulartight-junctional belt. Strands of bicellular and tricellu-lar tight junctions interconnect and form one continu-ous complex (Staehelin, 1974). Similar observationshave been made in adult olfactory epithelia (Kerjaschki& Horandner, 1976; Menco, 1980, 1984; Miiller,1980). The present study and its preliminary report(Menco, 1987) address morphological features of tightjunctions occurring at the onset of epithelial differen-tiation. Previous studies on developing olfactory epi-thelial tight junctions (Kerjaschki & Horandner, 1976;Mendoza el al. 1980) did not investigate this aspect ofthe subject.
Materials and methods
E14 embryos of the Sprague Davvley rat (Holtzman Co.,Madison, Wisconsin) were used for the major part of thisstudy. Further tight-junctional development was studied inE16 through E19 embryos and in adults. Approximately eightdams for each age group were killed by asphyxiation withCO2. About 70 embryos were removed per age group. Fiveadults served as controls. For one set of observations 1 usedall embryos of four dams. Dissection of the nasal septa of allof these took 2-3 h. Immediately after dissection the septawere fixed in Karnovsky's (1965) aldehyde fixative in 0-1 M-sodium cacodylate, pH7-4, at room temperature for 3h,washed in buffer, cryo-protected with 30% glycerol for 2h,and frozen in liquid propane. The frozen specimens wereknife-fractured in a Cressington CFE 40 freeze-fracturemachine (Cressington Scientific, Watford, England) at avacuum better than 10~6mbar ( lbar=10 5 Pa) and at atemperature of — 150cC. The fracture planes were rotary-shadowed (about 300 revs min~') with platinum/carbon at anangle of 20°; the replicas were reinforced with carbonobliquely evaporated from above. They were examined on300 mesh hexagonal thin-bar uncoated grids at 100 kV in aJEOL 100 CXII transmission electron microscope equippedwith a eucentric goniometer stage. Stereo pairs were oftenused to help to identify the fracture planes and these weremade at angles of ±6°. The micrographs were printed inreverse contrast, and fracture planes were labelled accordingto Branton el al. (1975). Statistics included standard errors ofmeans, regression analyses and one-way analyses of variance.They were compiled with a Macintosh Plus microcomputerand the Statview 512+ (Brainpower, Calabas, California)statistical package.
Results
E14 embryos could clearly be divided into two groups,each of which displayed a distinctly different tight-junctional morphology (Figs 1-3 and Fig. 4, respect-ively). I never saw both tight-junctional appearances ina single freeze-fracture replica. Hence, it is reasonableto assume that the two E14 tight-junctional mor-phologies, that of E14.1 and that of E14.2, weremutually exclusive within the limited period of mydissection procedures (see Materials and methods). Adescription of the morphologies follows below.
Tight junctions in early E14 (E14.1) rat embiyos
Olfactory epithelia of E14.1 embryos never displayedtypical bicellular tight junctions, i.e. strands of par-ticles or bars with their main orientation parallel to theepithelial surface. However, P- and E-fracture planesof cells in the developing olfactory epithelium of E14.1rat embryos did have isolated strands of particlesparallel to the longitudinal axes of the epithelial cells(Figs 1, 2). Often, several such isolated strands werefound on a single cell (Fig. 1). These strands fre-quently occurred as single entities or had a morphologysomewhere in between that of single strands and pairsof strands (Fig. 1 and inset A; Fig. 2). Three cellsappeared in intimate contact in the regions where thesestrands occurred (Fig. 1), and cell membranes curvedsharply away from the area where the cells met(Figs 1-3).
Even in the absence of strands the incipient bicellu-lar tight-junctional region could be distinguished fromregions proximal and distal to it. The region displayedshallow membrane undulations and small aggregates ofparticles, which looked like gap junctions (Fig. 1 andinset B; Fig. 2). In the E-face these particles arereflected as pits (Fig. 2). The zone of developingbicellular tight junctions was often differently orien-tated and had fewer membrane-associated particles(areas between open arrows in Fig. 1) compared withmembrane zones above and below it. P-face particledensities were 310 ±50, 120 ± 20 and 470 ± 40 par-ticles/im~2 (±S.E.M.) in apical, junctional and lateralregions, respectively. These values differ significantlyat />=£ 0-05.
Late E14 (E14.2) and EJ6 through E19 lightjunctions
A second group of E14 embryos (E14.2) containeddifferentiating olfactory epithelial surfaces. These dis-played strands of particles in bi- and tricellular tight-junctional regions (Fig. 4). The tricellular tight-junc-tional regions always had paired strands (Fig. 4,inset A) and the bicellular regions single strands. Themain orientation of the bicellular tight-junctional belt is
496 B. Ph. M. Menco

Fig. 1. Olfactory epithelial surface (right to left = proximal to distal) of an E14.1 rat embryo with four isolated tight-junctional strands parallel to the longitudinal axes (small arrows). One of these strands ends in a pyramidal apex (snake-shaped arrow and rotated right inset A) near the cell's surface. Inset A suggests that the strands mainly consist of a singleline of particles (compare with inset A in Fig. 4). Another tight-junctional strand displays membrane portions of all joiningcells (marked with 1,2,3 and a large arrowhead). The open circle indicates an interstitial space in a non-strand area. Cells/ and 3 display P-faces, cell 2 an E-face portion. Small clusters of particles that resemble gap junctions (arrowheads andinset B) are present within regions of incipient bicellular tight junctions, which are further characterized by undulatingpatterns (area between open arrows) and fewer P-face intramembranous particles than regions proximal and distal to it.The curved arrow points to a primary cilium (Menco, 1988). X66 000; insets, X 237 000.
Tight-junction formation in olfactory epithelia 497

Fig. 2. Higher magnification of a tight-junctional area in the olfactory epithelial surface of an E14.1 rat embryo (right toleft = proximal to distal). Membranes curve sharply towards either side of the longitudinal strand and appose each othermore closely in regions of these strands (large arrow) and P-face (PF) particle aggregates (small arrows) than elsewhere.The latter aggregates resemble gap junctions. In the E-face (EF) these areas are reflected as areas of pits (wide arrow).X223 000.
498 B. Ph. M. Menco
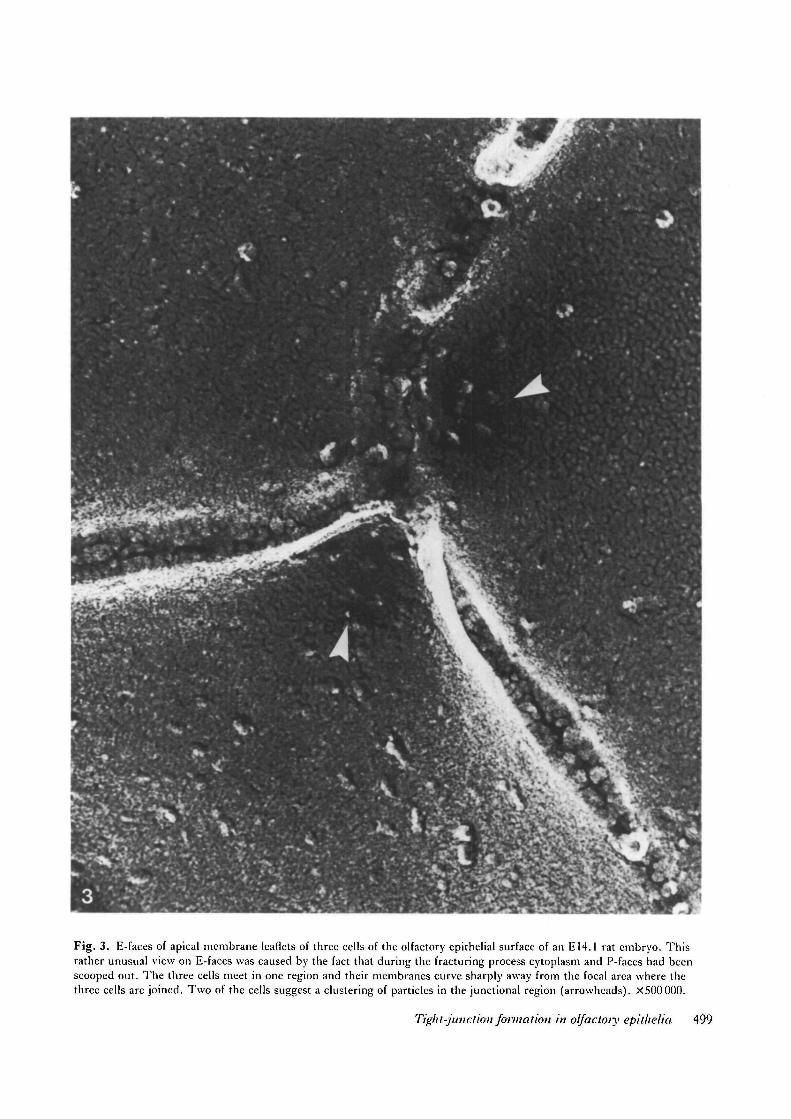
Fig. 3. E-faces of apical membrane leaflets of three cells of the olfactory epithelial surface of an E14.1 rat embryo. Thisrather unusual view on E-faces was caused by the fact that during the fracturing process cytoplasm and P-faces had beenscooped out. The three cells meet in one region and their membranes curve sharply away from the focal area where thethree cells are joined. Two of the cells suggest a clustering of particles in the junetional region (arrowheads). X500000.
Tight-junction formation in olfactory epithelia 499

B
Fig. 4. A cluster of olfactory receptor cells in an E14.2 rat embryo. Bicellular tight junctions consist of single strands ofparticles here mainly seen lying in E-face (EF) grooves, with the exception of a few P-face patches from neighbouring cells.There are six tricellular tight-junctional areas (small arrows), which consist of paired strands of particles parallel to thelongitudinal axes of the cells. The one with two arrows is presented at a higher magnification in inset A (compare with insetA of Fig. 1). A pitted E-face array present in an area where three tight-junctional strands intersect (arrowhead and inset B)reflects the complementary image of a gap-junction-like structure (see also Fig. 2). PF, P-face. X73 000; insets, X242 000.
500 B. Ph. M. Memo

Number of strandsStrands;tm~' horizontal
Olfactoryreceptor cells
Olfactorysupporting cells
2 UJUJ
Nasalrespiratory
epithelial cells
Fig. 5. Histograms of numbers and densities of strands of bicellular tight junctions between olfactory receptor cells andbetween receptor and supporting cells (first set of bars), between olfactory supporting cells only (second set of bars), andbetween respiratory cells (third set of bars) in rat embryos and adults. The numbers are presented with S.E.M. values.Densities parallel to the epithelial surface (strands/Urn"1 horizontal) as a function of the developmental period from E14.2through E19 fit: v = \-lx + 12-3, /-= 0-36, P ^ 0-001, elf =202, for the first group. For the second group, i.e. that of tightjunctions between supporting cells, the fit is: v = 0-&v + 11-4, / = 0-34, P ^ 0-001, df= 161. Strands jUm"1 vertical = densityof strands perpendicular to the epithelial surface.
perpendicular to the tricellular strands, and the orien-tation of the tricellular tight junctions was parallel tothe longitudinal axes of the cells. Gap-junction-likeaggregates of intramembranous particles (Fig. 4,inset B, here reflected as E-face pits) were considerablyless often encountered within tight-junctional belts ofE14.2 and older embryos than in those of E14.1embryos.
The general appearance of tight junctions of olfac-tory receptor and supporting cells in older embryos andin adults was similar to that in E14.2 embryos (Fig. 4;see also figs 5, 6, Menco, 1988). However, tightjunctions between receptor cells, and between receptorand supporting cells, had fewer strands in the E14.2age group than in older developmental age groups(/•^O-OS); El4.2 embryos had, on average, sixstrands. Strand numbers did not show a systematictrend in age groups E16 through E19 and in adults,where these numbers varied from seven to ten. Tightjunctions between supporting cells in these age groupshad, on average, five strands at E14.2 and seven to nineat age groups E16 through E19 and in adults (Fig. 5).Numbers of tight-junctional strands between cells ofthe nasal respiratory epithelium (a ciliated epithelialtype adjacent to the olfactory epithelium, which hasbeen used for comparison) ranged from five to eightand were consistently lower than those in the olfactoryepithelium. Densities of strands with their main orien-tation parallel to the epithelial surface ('strands/.im~l
horizontal' in Fig. 5), i.e. the tight-junctional packing,increased for tight junctions between all cells of theolfactory epithelium with about one strand jUm~' day"for the developmental period E14.2 to E19. Linearregressions fit the data. Such increases were not foundfor tight junctions of nasal respiratory epithelia(Fig. 5). Adults had about 20 strandsjitm"1 in olfactoryas well as in respiratory epithelia. Densities of the shortstrands interconnecting the horizontal strands('strands ^m~' vertical' in Fig. 5) were always lowerthan strands/xm~{ horizontal and did not systemati-cally change with development.
Discussion
General
The major findings of this study are summarized inFig. 6. Just before differentiation olfactory epithelialcells were seen to be joined in regions where three cellsmet by isolated strands of mainly unpaired particlesresembling tricellular tight-junctional strands and inregions where two cells met by gap-junction-like arraysof particles (Fig. 6A). Once differentiation sets in, twocells were firmly joined by a network of single bicellulartight-junctional strands and three cells were joined bytricellular tight junctions consisting of individualstrands of paired particles. There are still some gap-junction-like aggregates of particles, but considerably
Tight-junction formation in olfactory epithelia 501

Fig. 6. Diagram of the tight-junctional intercellular linkages just before (A) and after the onset of differentiation of theolfactory epithelial surface (B). Developmental changes in cellular outlines do occur but are not considered in the diagrams.A. Regions where three cells meet are marked by large dots from which thin lines parallel to the longitudinal axes of thecells emerge. These lines suggest the presence of linear arrays of tight-junctional strands. Regions where two cells meet aremarked by spotted areas, suggesting the presence of gap-junction-like aggregates of intramembranous particles. B. Tight-junctional strands are present in regions where three cells meet as well as in regions where two cells meet. Tricellular tight-junctional strands are paired. The small arrows point to spotted areas, which represent gap-junction-like arrays of particles.There are considerably fewer of these here than in the E14.1 situation shown in A.
fewer than just before the onset of epithelial differen-tiation (Fig. 6B).
I never saw both tight-junctional appearances in asingle freeze-fracture replica, or a mixed population ofcells with both E14.1 and E14.2 epithelial appearances(Menco & Farbman, 1985a; Menco, 1988). Therefore,it is reasonable to assume that the two tight-junctionalappearances reflect a distinct and sequential pattern ofdifferentiation peculiar to the whole surface of theolfactory epithelium, i.e. to receptor cells as well as tosupporting cells. The above observations suggest thatthe transition from the E14.1 to the E14.2 situation israpid, and that intermediate stages are hard to catch forpurposes of electron microscopy. Because tight-junc-tional strands can form within minutes (Kachar &Pinto da Silva, 1981), such a rapid transition is feasibleas far as the process of tight junction genesis isconcerned.
None of the previous studies on the development oftight junctions in olfactory epithelia dealt with featuresoccurring on the day that epithelial differentiation setsin. Mendoza et al. (1980) considered features in thechick before and after that stage. Kerjaschki & Hor-andner (1976) studied, in the mouse, only tight-junctional features of differentiating olfactory epi-thelia, i.e. features similar to those of developmentalage groups older than E14.1, which have been studiedhere in the rat. All of these had a tight-junctionalmorphology resembling that of E14.2 embryos (Figs 4,6B; figs 5 and 6, Menco, 1988).
Tri- and bicelhdar tight junctionsThe following arguments serve to substantiate thehypothesis that the isolated strands of particles presentjust before epithelial differentiation are focal tightjunctions peculiar to regions where three cells meet.
(1) The strands were found parallel to the longitudinalaxes of the olfactory epithelial cells (Fig. 1) as is thecase for tricellular tight-junctional strands (Fig. 4;Kerjaschki & Horandner, 1976; Menco, 1980, 1986).(2) Occasionally fracture planes were obtained in whichmembrane leaflets of all three cells were still seen to bejoined in the region of such strands (Figs 1, 3).Otherwise, membranes of different cells are closelyapposed in the region of these strands (Fig. 1). (3) As isthe case for tricellular strands in mature tight junctions(e.g. see Kerjaschki & Horandner, 1976; Menco, 1980,1986; Muller, 1980; Staehelin, 1974; Suzuki &Nagano, 1978), the isolated strands extend well belowthe regions of incipient bicellular tight-junctionalareas, i.e. to the right of the right-hand sets of openarrows in Fig. 1.
One important difference exists between tricellulartight junctions present in cells that are about todifferentiate and in those present later in development",tricellular tight junctions of cells that are about todifferentiate often have only single strands or strandssomewhere between single and parallel sets of strands(Fig. 1, inset A; Fig. 2). On the other hand, those ofcells present later in development and in adults (Ker-jaschki & Horandner, 1976; Menco, 1980, 1986) alwayshave pairs of strands (Fig. 4, inset A). Hence, itappears that in E14.1 embryos one strand might
502 B. Ph. M. Menco

temporarily interconnect all three joining cells,whereas in E14.2 embryos, and later, the epithelial cellsare interconnected by paired tricellular strands(Menco, 1980; Staehelin, 1974).
Isolated tight-junctional strands parallel to the longi-tudinal axes of the cells were also found in otherspecialized epithelia, namely developing retinal andciliary epithelia (Townes-Anderson & Raviola, 1981;Whiteley & Young, 1986). Therefore, it seems that adevelopmental feature of at least some specializedepithelia is that focal tight-junctional strands first formin regions where three cells meet. The present resultssuggest that the aforementioned tight-junctional ap-pearance occurs exclusively at the onset of epithelialdifferentiation. The results of studies on the pre-nataldevelopment of cellular (Menco & Farbman, I985a,b)and membrane structures (Menco, 1988) in the olfac-tory epithelial surface reinforce this view. The samemight be true for undifferentiated developing retinae,the epithelial surfaces of which (Whiteley & Young,1985) resemble those of developing olfactory epithelia(Menco & Farbman, 1985a).
Three distinct membrane domains (apical, junc-tional and lateral; Fig. 1) in and near zones of incipienttight-junctional belts could be distinguished at theonset of epithelial differentiation. This feature is notpeculiar to olfactory epithelia and the presence of thesedomains is probably caused by cytoskeletal elementsunderneath the membranes (Suzuki & Nagano, 1979;Townes-Anderson & Raviola, 1981). Such cytoskeletalelements are most likely responsible for the shallowmembrane undulations seen at the onset of epithelialdifferentiation (Fig. 1; and Suzuki & Nagano, 1979).According to Yamamoto & Kataoka (1986), on the basisof a study on the development of intercellular junctionsin pancreatic endocrine tissue, the shallow membraneundulations represent remnants of tight junctionsrather than initial features. Both processes might yieldthe same kind of images. During developmental disas-sembly the order of changes could be the reverse ofthose during assembly.
A tight-junctional morphology displaying rows ofparticles, as commonly observed in differentiatingolfactory epithelia (Fig. 4), is typical for newly formedtight junctions in chemically fixed, cryo-protected cells(Luciano et al. 1979; Suzuki & Nagano, 1979).Whether strand numbers increase, remain constant ordecrease during development depends apparently onthe epithelial type. My results on strand numbers ofbicellular tight junctions during the developmentalperiod following E14 (Fig. 5) are in agreement withthose of Kerjaschki & Horandner (1976), who statedthat once olfactory receptor cells begin to sprout cilia,tight-junctional strand numbers remain constant dur-ing further development. In contrast, tight-junctionalstrands in mouse epididymis increase (Suzuki &
Nagano, 1978) and in the rat's ciliary epithelium theydecrease with development (Arguillere et al. 1986).
According to their strand numbers tight junctions ofolfactory and nasal respiratory epithelia are typical ofthose having a rather high transepithelial resistance(Kerjaschki & Horandner, 1976; Menco, 1980). Thisstudy suggests that this is already the case as soon asstrands begin to form in either epithelial type, fromE14 on, even though olfactory epithelia have a fewmore tight-junctional strands and, hence, most prob-ably a somewhat higher transepithelial resistance thanrespiratory epithelia during development (Fig. 5). Inadults this distinction no longer exists (Menco, 1980).
It seems useful to relate the present results on tight-junctional appearance and strand numbers as a func-tion of development to electrophysiological exper-iments in rat embryos. Electro-olfactograms, which arethought to reflect at least in part summated generatorpotentials towards odorants (Van hset al. 1985), couldfirst be measured at E14 (Gesteland et al. 1982), thesame day that the tight-junctional transformation de-scribed here occurs. In addition, Gesteland et al.(1982) provided, with extracellular single-unit record-ings, evidence that olfactory receptor cell specificitytowards odorants only sets in after E15 and continues atleast until E20, thus after numbers of tight-junctionalstrands are established. This suggests that the develop-ment of olfactory receptor cell specificity towardsodorants is not related to tight-junctional strand num-bers and, hence probably also not to transepithelialresistance. In this context it is interesting to note thatAdamek et al. (1984) found, in the frog, that regener-ation of the electro-olfactogram, following experimen-tal epithelial destruction, is independent of tissue shortcircuits.
Strand density has not often been considered as aparameter important for transepithelial resistance. Inthe few studies that I could find this parameter did notseem to be important (Gonzalez-Mariscal et al. 1984;Palant et al. 1983). However, results of the presentstudy suggest that this feature should be consideredmore seriously in the future when relations betweenepithelial physiology and tight-junctional morphologyare investigated. In contrast to the situation duringdevelopment (Fig. 5), the packing of tight-junctionalstrands of cells of olfactory and nasal respiratoryepithelia is the same in adult vertebrates (Menco,1980).
Gap-junction-like aggregates of intramembranousparticles
Gap-junction-like aggregates of intramembranous par-ticles (Figs 1, 2) were more often found just beforedifferentiation of the olfactory epithelial surface set in(Menco & Farbman, 1985a,6) than at later stages ofdevelopment (Fig. 4, inset B; Fig. 6; Kerjaschki &
Tight-junction formation in olfactory epithelia 503

Horandner, 1976; Mendoza et al. 1979). Occasionallythey were still found in olfactory epithelia of adultanimals (Menco, 1980).
The aggregates are not unique to tight-junctiorialgenesis in olfactory epithelia (Dermietzel et al. 1977;Suzuki & Nagano, 1978, 1979; Townes-Anderson &Raviola, 1981; Whiteley & Young, 1986). They havebeen assumed to be true gap junctions (e.g. seeTownes-Anderson & Raviola, 1981), although this isinconclusive (Revel, 1982). If the aggregates are,indeed, true gap junctions they could conceivably playa role in spreading the signals triggering the describedtransition from the E14.1 to the E14.2 epithelialappearance. This might be the reason why suchaggregates in the developing olfactory epithelial surface(rather than strands of particles) function as initialintercellular fusion sites in regions of incipient bicellu-lar tight junctions. In one instance (not shown) I sawsuch a gap-junction-like aggregate of particles to be inline with the linear strands of particles of tight junc-tions of olfactory epithelia of E14.1 embryos, sugges-ting that such aggregates may also serve as precursorsites for tricellular tight junctions.
I am very grateful to Drs A. I. Farbman, F. Gonzales(Northwestern University) and J.-P. Revel (California Insti-tute of Technology), and to Audrey Niffenegger and LynRosen for critical reading of this manuscript. I appreciatedthe continuous technical support of Dr P. A. Walley (Cress-ington Scientific, Watford, UK) and Mr E. W. Minner. Thiswork was funded by NIH grant NS 21555.
References
ADAMEK, G. D., GESTELAND, R. C , MAIR, R. G. &
OAKLEY, B. (1984). Transduction physiology of olfactoryreceptor cell cilia. Brain Res. 310, 87-97.
ARGUILLERE, P., PATEY, A. & HIRSCH, M. (1986).
Quantitative analysis of tight junctions during ciliaryepithelium development. Expl Eye Res. 42, 239-248.
BRANTON, D., BULLIVANT, S., GILULA, N. A.,
KARNOVSKY, M. J., MOOR, H., MOHLETHALER, K.,
NORTHCOTE, D. H., PACKER, L., SATIR, B., SATIR, P.,
SPETH, V., STAEHELIN, L. A., STEERE, R. L. &
WEINSTEIN, S. (1975). Freeze-etching nomenclature.Science 190, 54-56.
DERMIETZEL, R., MELLER, K., TETZLAFF, W. & WAELSCH,
M. (1977). In vivo and in vitro formation of thejunctional complex in choroid epithelium. A freeze-etching study. Cell Tiss. Res. 181, 427-441.
GESTELAND, R. C , YANCEY, R. A. & FARBMAN, A. I.
(1982). Development of olfactory receptor neuronselectivity in the rat fetus. Neumscience 7, 3127-3136.
GONZALEZ-MARISCAL, B., CHAVEZ DE RAMfREZ, B. &
CEREUIDO, M. (1983). Effect of temperature on theoccluding junctions of monolayers of epithelioid cells(MDCK). J. Membrane Biol. 79, 175-184.
KACHAR, B. & PINTO DA SILVA, P. (1981). Rapid massive
assembly of tight junction strands. Science 213,541-544.
KARNOVSKY, M. J. (1965). A formaldehyde-glutaraldehydefixative of high osmolarity for use in electronmicroscopy. J . Cell Biol. 27, 137-138.
KERJASCHKI, D. & HORANDNER, H. (1976). The
development of mouse olfactory vesicles and theircontacts: A freeze-etching study. J. Ullrastmct. Res. 54,420-444.
LUCIANO, L., THIELE, J. & REALE, E. (1979).
Development of follicles and of occluding junctionsbetween the follicular cells of the thyroid gland. A thin-section and freeze-fracture study in the fetal rat. J.Ultrastmct. Res. 66, 164-181.
MENCO, B. PH. M. (1980). Qualitative and quantitativefreeze-fracture studies on olfactory and nasal respiratoryepithelial surfaces of frog, ox, rat, and dog. III. Tight-junctions. Cell Tiss. Res. 211, 361-373.
MENCO, B. PH. M. (1984). Ciliated and microvillousstructures of rat olfactory and nasal respiratory epithelia.A study using ultra-rapid cryo-fixation followed byfreeze-substitution or freeze-etching. Cell Tiss. Res. 235,225-241.
MENCO, B. PH. M. (1986). A survey of ultra-rapidcryofixation methods with particular emphasis onapplications to freeze-fracturing, freeze-etching, andfreeze-substitution. J. Election Microsc. Techn. 4,177-240.
MENCO, B. PH. M. (1987). Tight-junctions in developingrat olfactory epithelia. Client. Sens. 12, 683.
MENCO, B. PH. M. (1988). Pre-natal development of ratnasal epithelia. IV. Freeze-fracturing on apices,microvilli and primary and secondary cilia of olfactoryand respiratory epithelial cells, and on olfactory axons.Anat. Embryol. (in press).
MENCO, B. PH. M. & FARBMAN, A. I. (1985«). Genesis of
cilia and microvilli of rat nasal epithelia during pre-nataldevelopment. I. Olfactory epithelium, qualitativestudies. J . Cell Sci. 78, 283-310.
MENCO, B. PH. M. & FARBMAN, A. 1. (19856). Genesis of
cilia and microvilli of rat nasal epithelia during pre-nataldevelopment. II. Olfactory epithelium, a morphometricanalysis. J . Cell Sci. 78, 311-336.
MENDOZA, A. S., MIRAGALL, F. & BREIPOHL, W. (1980).
Intercellular junctions during the development of theolfactory epithelium in the chick. A freeze-etching study.J. siibmicmsc. Cytol. 12, 29-41.
MULLER, W. (1980). Cell junctions with funnels in theolfactory mucosa of frogs (Rana temporaria L). CellTiss. Res. 207, 165-169.
PALANT, G. E., DUFFEY, M. E., MOOKERJEE, B. K., HO,
S. & BENTZEL, C. J. (1983). Ca2+ regulation of tight-junction permeability and structure in Necturus gallbladder. Am. J. Physiol. 245, C203-C212.
REVEL, J.-P. (1982). Intercellular communication. In TheParacellular Pathway (Report of a conference) (ed. S.E. Bradley & E. F. Purcell), pp. 57-83. New York:Josiah Macy, Jr Foundation.
504 B. Ph. M. Menco

STAEHELIN, L. A. (1974). Structure and function ofintercellular junctions. Int. Rev. Cytol. 39, 191-283.
SUZUKI, F. & NAGANO, T. (1978). Development of tightjunctions in the caput epididymal epithelium of themouse. Devi Biol. 63, 321-334.
SUZUKI, F. & NAGANO, T. (1979). Morphogenesis of tightjunctions in the peritoneal mesothelium of the mouseembryo. Cell Tiss. Res. 198, 247-260.
TOWNES-ANDERSON, E. & RAVIOLA, G. (1981). The
formation and distribution of intercellular junctions inthe Rhesus monkey optic cup: The early development ofthe cilio-iridic and sensory retinas. Devi Biol. 85,209-232.
VAN AS, W., KAUER, J. S., MENCO, B. PH. M. & KOSTER,
E. P. (1985). Quantitative aspects of the electro-
olfactogram in the tiger salamander. Chein. Sens. 10,1-21.
VVHITELEY, H. E. & YOUNG, S. (1985). Cilia in the fetaland neonatal canine retina. Tissue & Cell 17, 335-340.
VVHITELEY, H. E. & YOUNG, S. (1986). The external
limiting membrane in developing normal and dysplasticcanine retina. Tissue & Cell 18, 231-239.
YAMAMOTO, M. & KATAOKA, K. (1986). Intercellular
junctions in pancreatic exocrine and endocrine cells indeveloping rats and mice: Freeze-fracture and thinsection studies. .7. Electron Micivsc. 35, Suppl.,2899-2900.
(Received 28 September J9S7 -Accepted, in revised fonn,11 December 1987)
Tight-junction formation in olfactoiy epithelia 505
