transcription of the human adenovirus e1a gene in saccharomyces cerevisiae
TRANSCRIPT

Gene, 33 (1985) 159-168
Elsevier
159
GENE 1183
Transcription of the human adenovirus Ela gene in Saccharomyces cerevisiae
(Recombinant DNA; yeast; viral gene; mRNA; E. coli shuttle vector)
Hiroshi Handa, Kiyohisa Mizumoto, Kinichiro Oda, Takashi Okamoto * and Toshio Fukasawa **
Institute for Medical Sciences, The University of Tokyo, Shirokanedai, Minato, Tokyo (Japan) Tel. 443-8111; * Department of Internal Medicine, Tel. 353-121 I, and ** Laboratory ofMolecular Genetics, Keio University School of Medicine, Shinamomachi, Shinjuku, Tokyo (Japan) Tel. 353-1211
(Received July 26th, 1984)
(Accepted September 22nd, 1984)
SUMMARY
The early region la (Ela) and its flanking sequences of human adenovirus type 5 (Ad5) have been cloned in the yeast-Escherichia coli shuttle vector YEp13 and transferred into the yeast Succharomyces cerevisiae. The Ela-specific RNAs were produced in the transformed yeast cells. The 5’ ends of these transcripts were capped but were lacking 10 to 45 nucleotides from the 5’ end of the proper Ela mRNA. These transcripts terminated approx. 1000 nucleotides downstream from the proper 3’ end. No splicing of the Ela-specific RNA could be detected in the yeast cells.
INTRODUCTION
The expression of a variety of foreign genes of eukaryotic origin in the yeast S. cerevisiae have been investigated since the establishment of the recombi- nant DNA technology. An efficient transcription was demonstrated in several cellular genes, such as the rabbit /I-globin gene (Beggs et al., 1980), the gene for glycinamide ribonucleotide transformylase from Drosophila melanogaster (Henikoff et al., 1981), the discoidin-Ia gene from Dictyostelium discoideum (Jel- linghaus et al., 1982), or the genes for cytochrome c and alcohol dehydrogenase from Schizosaccharo- myces pombe (Russell, 1983). The transcription of
Abbreviations: Ad5, adenovirus type 5; bp, base pairs; Ela, early
region la; kb, kilobases or 1000 bp; SD, see MATERIALS AND
METHODS, section b; SDS, sodium dodecyl sulfate; [I,
indicates plasmid-carrier state.
0378-I 119/85/$03.30 0 1985 Elsevier Science Publishers
those genes in S. cerevisiue generally starts near but at different sites from their proper initiation sites. On the other hand, the Helpes simplex virus thymidine kinase gene, the only viral gene that has been studied, was not transcribed at all in S. cerevisiue (Kiss et al., 1982). One might suspect, therefore, that viral genes may not contain the sequence that is recognizable as the promoter in yeast. Therefore we studied the expression of the Ela of human Ad5 in the yeast. The Ela gene has interested us since the enhancer element was found far upstream from the initiation site of Ela mRNA (Hearing and Shenk, 1983; Hen et al., 1983) and that new initiation sites were reported to appear as infection proceeds (Osborne and Berk, 1983). Thus we have constructed a hybrid DNA consisting of a yeast-E. coli shuttle vector and the Ad5 Ela gene containing its 5’- and 3’-flanking regions, with which a yeast strain was transformed. We found that the Ela gene was efficiently transcrib-

160
ed in the transformed yeast. A characterization of the Ela-specific transcript from yeast revealed that the 5’ ends of the transcripts were properly capped, despite the fact that the transcription initiated at several sites which were different from the proper start. The presumptive promoter element functional in yeast will be discussed in the light of the present result together with the information available at the present time.
MATERIALS AND METHODS
(a) Construction of YEpl3Ela
Ad5 DNA was prepared from purified vii-ions as described by Petterson and Sambrook (1973). To get rid of the terminal protein which is covalently attached to each 5’ end of the genome, Ad5 DNA was treated with T4 DNA polymerase as described by Berkner and Sharp (1982) except that dTTP was omitted from the reaction mixture. The single- stranded region generated at each 5’ end was digested with S 1 nuclease. The end was tilled in with Klenow fragment of E. coli DNA polymerase I in the presence of all four dNTPs, ligated to BamHI linker and digested with both BamHI and HindIII. The 2.8-kb HindIII-G fragment (Tooze, 1983) was in- serted into the plasmid pSV2gpt (Mulligan and Berg, 1980) at the BamHI-Hind111 sites and the resultant plasmid (pSVAdSHG, Fig. 1) was amplified in E. coli strain HB 101. About 20 bp were found to be removed from the terminus of the viral genome as judged from restriction mapping. The HindIII-G fragment was then inserted into the yeast-E. coli shuttle vector YEpl3 (Fig. 1, Broach et al., 1979) at the BamHI-Hind111 sites and amplified in strain HBlOl.
in 500 ml of leucine-free medium, called SD, containing 0.64% nitrogen base w/o amino, 2% dextrose and the necessary nutrients. At an early logarithmic phase (approx. lo7 cells/ml), cells were collected by centrifugation, washed once with distill- ed water and suspended in 25 ml of 1.2 M sorbitol. After treatment with zymolyase 60 000 (100 pg/ml, Kirin Brewery Co.) at 30’ C for 30 min, the resultant spheroplasts were mixed with an equal volume of two-fold strength SD medium containing 0.8 M MgSO, and incubated at 30°C for 30 min. During that time the synthesis of macromolecules which had stopped in the course of the zymolyase treatment would resume. The spheroplasts were then incubated for additional 2 min in the presence of cycloheximide (100 pg/ml), quickly chilled by mixing
with 25 ml of ice-cold 1 M sorbitol containing 80 pg/ml of cycloheximide, and immediately centri- fuged at 3000 rev./min for 12 min. The spheroplasts were washed twice with 1 M sorbitol containing 50 ,ug/ml of cycloheximide and disrupted thoroughly by resuspending with 25 ml of a solution containing 10mM EDTA, 10 mM NaCl and 1% SDS. The sample was mixed with an equal volume of water- saturated phenol and shaken vigorously at 60°C for 20 min. The mixture was centrifuged, and the aqueous phase separated and treated twice with phenol as above but at room temperature. The aqueous phase was further treated twice with a mixture of phenol and chloroform (3 :2) and to the final aqueous phase were added 2 ~01s. of ethanol and 0.1 vol. of 5 M LiCI; the mixture was stored at -20°C for more than 16 h. During this period of time, the bulk of RNA was precipitated. The RNA was referred to as total RNA. Poly(A) + RNA was isolated from total RNA by oligo(dT)-cellulose column chromatography.
(c) RNA blotting and Sl mapping
(b) Transformation and RNA extraction
Transformation of E. coli strain HBlOl was per- formed according to Maniatis et al. (1982). The yeast strain SHY3 (a, ste-VC9, ura3, trpl, leu2, hi.3, adel, canl; Botstein et al. 1979) was transformed with either YEp13 or YEpl3Ela DNA by the method described by Ito et al. (1983), and leucine-indepen- dent colonies were isolated.
Cells of one of the transformant yeasts were grown
The RNA samples were glyoxalated as described by McMaster and Carmichael (1977) and fractionat- ed by electrophoresis in 1.2% neutral agarose gel. RNA was transferred to nitrocellulose membrane and hybridized with the DNA probe as indicated in the text according to Thomas (1980).
The Sl nuclease mapping was according to Berk and Sharp (1977) as modified by Weaver and Weissmann (1979).

161
Sal I (d) Determination of the cap structure at the 5’ end
of mRNA
Poly(A)+ RNA specific for the Ela gene was
purified from total poly(A)+ RNA fraction by
hybridization with denatured Ela DNA (0 to 3.8
map units on the Ad5 genome) bound to nitrocellu-
lose membrane, followed by elution from the mem-
brane according to the procedure described by
Ricciardi et al. (1979).
The Ela-specific RNA was heat-denatured at
100°C for 2 min and treated with calf intestinal
alkaline phosphatase (Boehringer Mannheim). The
cap structure was chemically removed (Fraenkel-
Conrat and Steinschneider, 1967) and RNA was
re-capped with purified vaccinia virus guanyltrans-
ferase (Saiga et al., 1982) in the presence of
[ CX-~~P] GTP. The RNA labelled at the cap structure
was treated successively with nuclease Pl (Yamasa
Shoyu Co.) and with calf intestinal alkaline phos-
phatase. To identify the capped nucleotide, the
sample was subjected to electrophoresis on What-
man DE81 paper at pH 3.5 according to the proce-
dure described by Mizumoto and Lipmann (1979).
RESULTS
(a) Blot hybridization analysis of RNA synthesized
in the yeast carrying the Ela gene on the replicating
plasmid
To determine whether the Ela-gene was tran-
scribed in yeast at all, total RNA from the trans-
formed yeast was analyzed by blot hybridization. As
a probe, the BamHI-XbaI 1.3-kb DNA fragment
from pSVAd5HG (Fig. 1) was nick-translated with
[a-32P] dCTP. The fragment covers the 5’ end of
Ela mRNA, extending from 0 to 3.8 map units of the
Ad5 genome. As clearly seen in Fig. 2, one single
species specifically hybridized to the Ela DNA was
present in the yeast with the plasmid YEpl3Ela
(lane 3) but not in the yeast with YEp13 (lane 4). The
Ela-specific transcript in yeast exhibited a size of
approx. 2000 nucleotides, which was much larger
than that of the primary transcript of the Ela gene
in HeLa cells (about 1100 nucleotides). In the same
experiment, the LEU2 transcript was also identified
+lindlll
BainHl
Fig. 1. Construction of the plasmid YEp 13E 1 a. The yeast-E. coli
shuttle vector YEpl3 (Broach et al., 1979) consists of a fragment
of pBR322 ( -), a yeast chromosomal DNA covering the
LEU2 gene (I) and a fragment of 2 pm DNA (cross-hatched
bar). The plasmid pSVAdSHG was constructed by inserting the
fragment HindHI-G (2.7 kb, open bar) which extends from 0 to
8 map units ofthe Ad5 genome into pSV2gpt (Muligan and Berg,
1980). The plasmid pSVAd5HG contains the fragment from
SV40 virus (striped bar). The arrows inside the yeast or Ad5
region indicate the approximate location and direction of the
respective gene transcripts LEU2 or Ela. Plasmid YEpl3Ela
was constructed by inserting BamHI-Hind111 segment of
pSVAd5HG into BarnHI, Hind111 sites of YEp13. SV40 DNA is
absent from YEpl3Ela.
by the 32P-labelled LEU2 DNA 1.3-kb fragment,
which was separated from the plasmid YIp32 (Bot-
stein et al., 1979) following digestion with KpnI and
Salk The fragment included about 80% of the coding
region through the 3’ flanking region (Andreadis
et al., 1983; see Fig. 1). Thus only one species of
RNA with a size of about 1300 nucleotides was
found in the yeast either with YEp13Ela or with
YEp13 (lanes 1,2). It was noted that the abundance
of the LEU2 transcript in the latter was greater than
that of the former for unknown reasons.
(b) Analysis of 5’ end of Ela-specific RNA synthe- sized in yeast
To assign the 5’ end of Ela transcript on the Ela
DNA sequence, an approx. lOOO-bp SmaI-BamHI
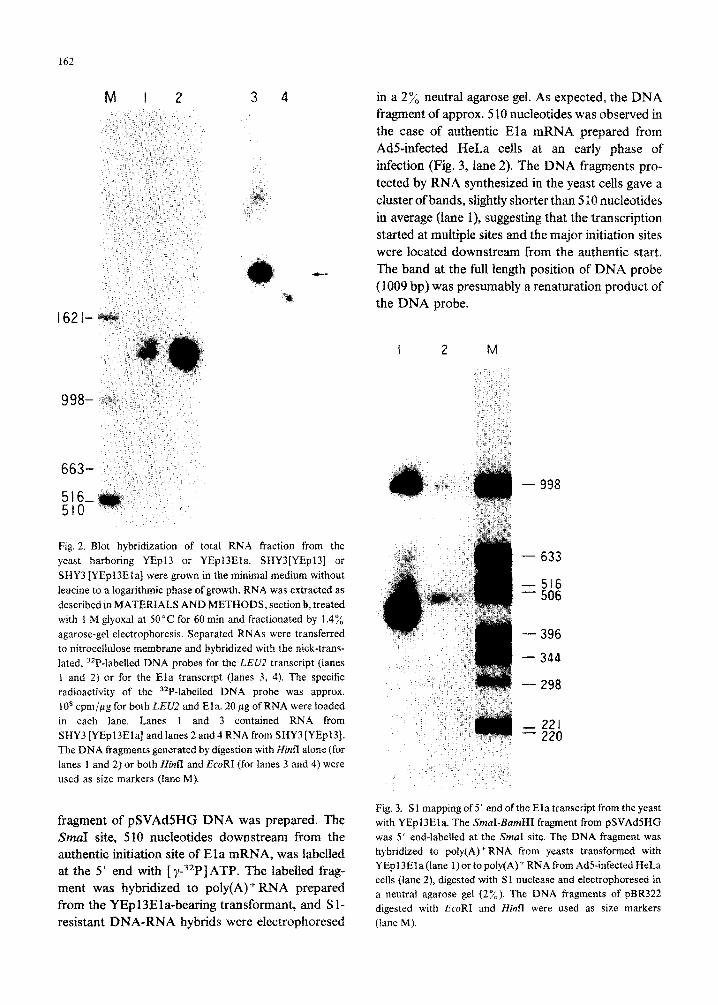
162
M I 2
1621-
998-
663-
in a 2% neutral agarose gel. As expected, the DNA fragment of approx. 5 10 nucleotides was observed in the case of authentic Ela mRNA prepared from Ad%infected HeLa cells at an early phase of infection (Fig. 3, lane 2). The DNA fragments pro- tected by RNA synthesized in the yeast cells gave a cluster of bands, slightly shorter than 5 10 nucleotides in average (lane I), su~esting that the ~~sc~ption started at multiple sites and the major initiation sites were located downstream from the au~cntic start,
- The band at the full length position of DNA probe (1009 bp) was presumably a renaturation product of the DNA probe.
I 2 M
Fig. 2. Blot hybridization of total RNA fraction from the yeast harboring YEpl3 or YEpl3Ela. SHY3[YEpl3] or SHY3 [YEpl3Ela] were grown in the minimal medium without leucine to a iogarithm~c phase of growth. RNA was extracted as described in MATERIALS AND METHODS, section b, treated with I M giyoxal at 50°C for 60 min and fractionated by 1.4% agarose-gel electrophoresis. Separated RNAs were transferred to nitrocellulose membrane and hybridized with the nick-trans- lated, 3ZP-labelled DNA probes for the LEUZ transcript (lanes 1 and 2) or for the Ela transcript (lanes 3, 4). The specific radioactivity of the 32P-labelled DNA probe was approx. lOa cpmjgg for both LEU2 and Eia. 20 pg of RNA were loaded in each lane. Lanes 1 and 3 contained RNA from SHY3[YEp13Ela]andlanes2and4RNAfromSHY3[YEpl3]. The DNA fragments generated by digestion with Hinfl alone (for lanes 1 and 2) or both Hi& and EcoRI (for lanes 3 and 4) were used as size markers (lane M).
- 633
= ;A;
- 396
- 344
- 298
fragment of pSVAd5HG DNA was prepared* The SmaI site, 510 nucleotides downstream from the authentic initiation site of Ela mRNA, was labelled at the 5’ end with [ Y-~~P]ATP. The labelled frag- ment was hyb~dized to poly(A)+ RNA prepared from the YEp13Ela-bearing transformant, and Sl-
Fig. 3. S 1 mapping of 5’ end of the Ela transcript from the yeast with YEpl3Ela. The SmaI-BarnHI fragment from pSVAd5HG was 5’ end-labelled at the SmaI site. The DNA fragment was hybridized to poIy(A)+‘RNA from yeasts transformed with YEpl3~la(lane 1)or to poly(A)+RNA from Ad%infected HeLa ceils {lane 21, digested with Sl nuclease and electrophoresed in a neutral agarose gel (2%). The DNA fragments of pBR322 digested with EcoRI and Hinff were used as size markers
resistant DNA-RNA hybrids were electrophoresed (lane M).
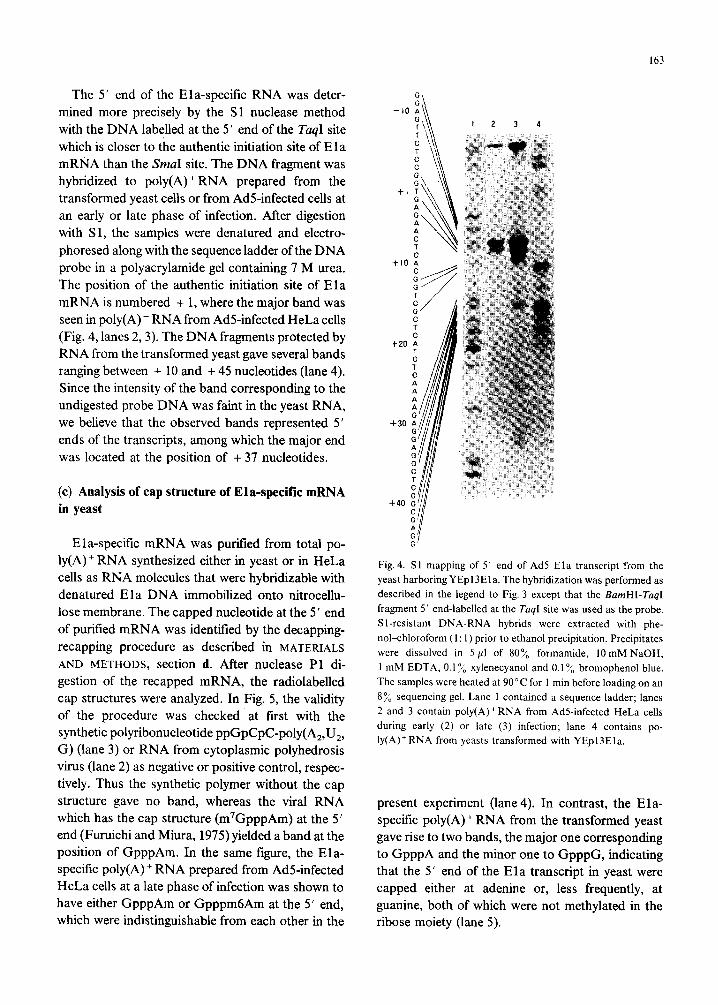
163
The 5 ’ end of the Ela-specific RNA was deter- mined more precisely by the Sl nuclease method with the DNA labelled at the 5’ end of the TuqI site which is closer to the authentic initiation site of Ela mRNA than the SmaI site. The DNA fragment was hybridized to poly(A)+ RNA prepared from the transformed yeast cells or from AdS-infected cells at an early or late phase of infection. After digestion with Sl, the samples were denatured and electro- phoresed along with the sequence ladder of the DNA probe in a polyacrylamide gel containing 7 M urea. The position of the authentic initiation site of Ela mRNA is numbered + 1, where the major band was seen in poly(A) + RNA from Ad5infected HeLa cells (Fig. 4, lanes 2,3). The DNA fragments protected by RNA from the transformed yeast gave several bands ranging between + 10 and + 45 nucleotides (lane 4). Since the intensity of the band corresponding to the undigested probe DNA was faint in the yeast RNA, we believe that the observed bands represented 5’ ends of the transcripts, among which the major end was located at the position of + 37 nucleotides.
(c) Analysis of cap structure of Ela-specific mRNA
in yeast
Ela-specific mRNA was purified from total po- ly(A) + RNA synthesized either in yeast or in HeLa cells as RNA molecules that were hybridizable with denatured Ela DNA immobilized onto nitrocellu- lose membrane. The capped nucleotide at the 5’ end of purified mRNA was identified by the decapping- recapping procedure as described in MATERIALS AND METHODS, section d. After nuclease Pl di- gestion of the recapped mRNA, the radiolabelled cap structures were analyzed. In Fig. 5, the validity of the procedure was checked at first with the synthetic polyribonucleotide ppGpCpC-poly(A,,U,, G) (lane 3) or RNA from cytoplasmic polyhedrosis virus (lane 2) as negative or positive control, respec- tively. Thus the synthetic polymer without the cap structure gave no band, whereas the viral RNA which has the cap structure (m7GpppAm) at the 5’ end (Furuichi and Miura, 1975) yielded a band at the position of GpppAm. In the same figure, the Ela- specific poly(A) + RNA prepared from AdS-infected HeLa cells at a late phase of infection was shown to have either GpppAm or Gpppm6Am at the 5’ end, which were indistinguishable from each other in the
+I
+10
+20
+30
Fig. 4. Sl mapping of 5’ end of Ad5 Ela transcript from the
yeast harboring YEpl3Ela. The hybridization was performed as
described in the legend to Fig. 3 except that the BumHI-TuqI
fragment 5’ end-labelled at the TuqI site was used as the probe.
Sl-resistant DNA-RNA hybrids were extracted with phe-
nol-chloroform (1: 1) prior to ethanol precipitation. Precipitates
were dissolved in 5 ~1 of 80% formamide, 10mM NaOH,
1 mM EDTA, 0.1% xylenecyanol and 0.1% bromophenol blue.
The samples were heated at 90°C for I min before loading on an
8% sequencing gel. Lane 1 contained a sequence ladder; lanes
2 and 3 contain poly(A)+RNA from AdS-infected HeLa cells
during early (2) or late (3) infection; lane 4 contains po-
ly(A)+RNA from yeasts transformed with YEpl3Ela.
present experiment (lane 4). In contrast, the Ela- specific poly(A)+ RNA from the transformed yeast gave rise to two bands, the major one corresponding to GpppA and the minor one to GpppG, indicating that the 5 ’ end of the Ela transcript in yeast were capped either at adenine or, less frequently, at guanine, both of which were not methylated in the ribose moiety (lane 5).
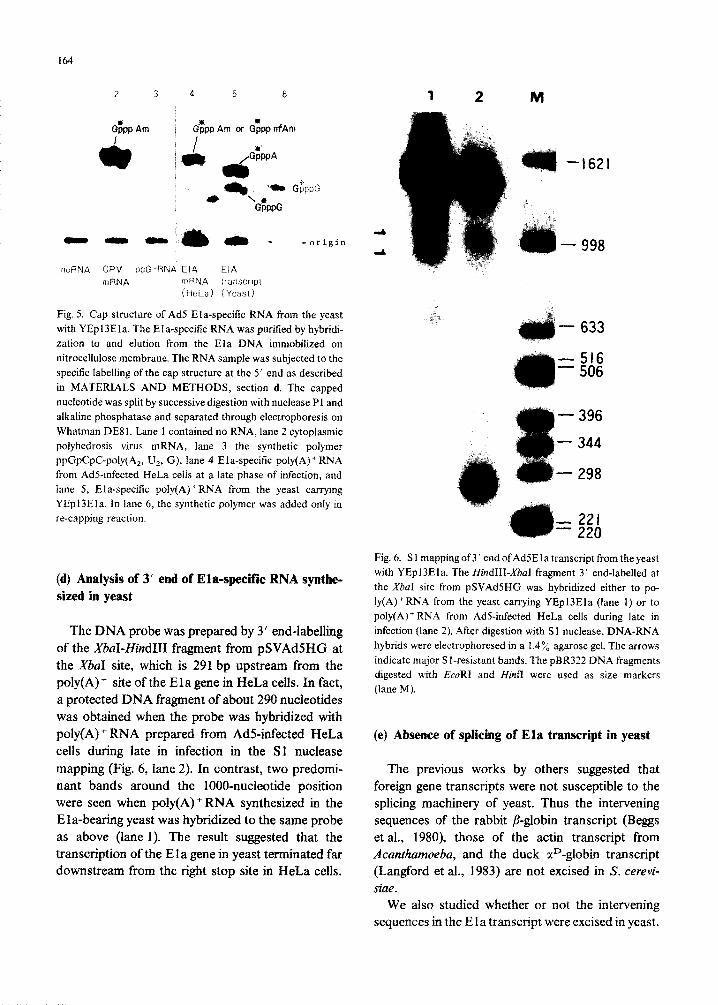
164
2 3 4 5 6
noRNA CPV [)pG-RNA EIA EIA
mRNA mRNA :ranscrlpt (HeLa) (Yeast)
Fig. 5. Cap structure of Ad5 Ela-specific RNA from the yeast with YEpI3Eta. The EIa-specific RNA was purified by hybridi- zation to and elution from the EIa DNA immobilized on nitrocellulose membrane. The RNA sample was subjected to the specific labelling of the cap structure at the 5’ end as described in MATERIALS AND METHODS, section d. The capped nucleotide was split by successive digestion with nuclease P 1 and alkaline phosphatase and separated through electrophoresis on Whatman DE8 1. Lane 1 contained no RNA, lane 2 cytoplasmic polyhedrosis virus mRNA, lane 3 the synthetic polymer ppGpCpC-poiy(A,, Uz, G), lane 4 Eia-specific poly(A) + RNA from Ad%infected HeLa cells at a late phase of infection, and lane 5, Ela-specitic poiy(A)+RNA from the yeast carrying YEpl3Ela. In lane 6, the synthetic polymer was added only in re-capping reaction.
(d) Analysis of 3’ end of Ela-specific RNA synthe- sized in yeast
The DNA probe was prepared by 3’ end-labelling of the XbaI-N&d111 fragment from pSVAd5HG at the J&z1 site, which is 291 bp upstream from the poly(A) + site of the Ela gene in HeLa cells. In fact, a protected DNA fragment of about 290 nucleotides was obtained when the probe was hybridized with poly(A) + RNA prepared from Ad5infected HeLa cells during late in infection in the Sl nuclease mapping (Fig. 6, lane 2). In contrast, two predomi- nant bands around the 1000~nucleotide position were seen when poly(A) + RNA synthesized in the Ela-bearing yeast was hybridized to the same probe as above (lane 1). The result suggested that the transcription of the Ela gene in yeast terminated far downstream from the right stop site in HeLa cells.
-
- 633
I621
- 396
- 344
- 298
- 221 - 220
Fig. 6. S 1 mapping of 3’ end ofAd5E la transcript from the yeast with YEpl3Ela. The HindHI-XbnI fragment 3’ end-labelled at the XbaI site from pSVAd5HG was hybridized either to po- ly(A)+RNA from the yeast carrying YEpl3Ela (lane 1) or to poly(A) + RNA from Ad%infected HeLa cells during late in infection (lane 2). After digestion with S 1 nuclease, DNA-RNA hybrids were electrophoresed in a 1.4% agarose gel. The arrows indicate major S l-resistant bands. The pBR322 DNA fragments digested with EcoRI and Hi&I were used as size markers (lane M).
(e) Absence of splicing of Ela transeript in yeast
The previous works by others suggested that foreign gene transcripts were not susceptible to the splicing machinery of yeast. Thus the intervening sequences of the rabbit /?-globin transcript (Bergs et al., 1980), those of the actin transcript from Acanthamoeba, and the duck orD-globin transcript (Langford et al., 1983) are not excised in S. cerevi- siae.
We also studied whether or not the inte~ening sequences in the E 1 a transcript were excised in yeast.
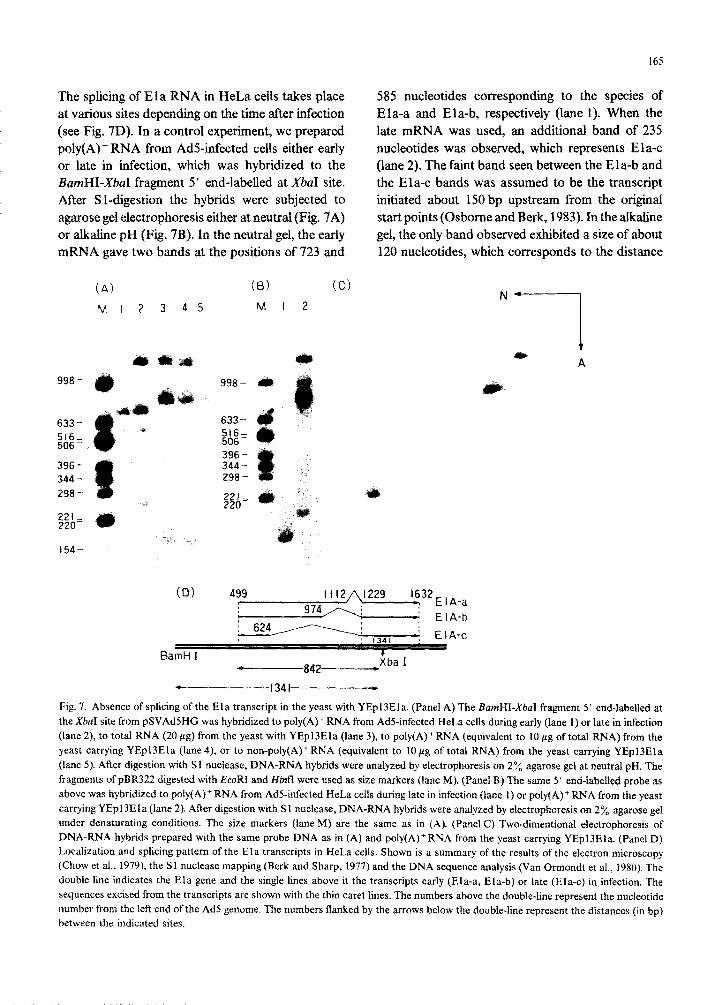
165
The splicing of Ela RNA in HeLa cells takes place 585 nucleotides corresponding to the species of
at various sites depending on the time after infection Ela-a and Ela-b, respectively (lane 1). When the
(see Fig. 7D). In a control experiment, we prepared late mRNA was used, an additional band of 235
poly(A) + RNA from Ads-infected cells either early nucleotides was observed, which represents Ela-c or late in infection, which was hybridized to the (lane 2). The faint band seen between the Ela-b and BarnHI-X&I fragment 5’ end-labelled at XbaI site. the Ela-c bands was assumed to be the transcript After Sl-digestion the hybrids were subjected to initiated about 150 bp upstream from the original
agarose gel electrophoresis either at neutral {Fig. 7A) start points (Osborne and Berk, 1983). In the alkaline
or alkaline pH (Fig. 7B). In the neutral gel, the early gel, the only band observed exhibited a size of about mRNA gave two bands at the positions of 723 and 120 nucleotides, which corresponds to the distance
(A) (B) CC)
MI 2 3 45 M I 2
998-
633 - 516_ 506-,
396-
344 -
238 -
221_ 220- jP
154-
CD)
998- dIl@
499 I Il2Al229 1632
9744 h EIA-a
j 624-i ; EIA-b
! ,341 j EIA-c
N--l
I Q*
A
BamH I * 842
i?baI
w 1341 *
Fig. 7. Absence of splicing of the Ela transcript in the yeast with YEpl3Ela. (Panel A) The RamHI-XbaI fragment 5’ end-labelled at the XbaI site from pSVAd5HG was hybridized to poly(A)+ RNA from AdS-infected HeLa cells during early (lane 1) or late in infection (lane 2), to total RNA (20 pg) from the yeast with YEpl3Ela (lane 3), to poly(A)‘RNA (equivalent to 10 pg of total RNA) from the yeast carrying YEpl3Ela (lane 4) or to non-poly(A)‘RNA (equivalent to 1Opg of total RNA) from the yeast carrying YEpl3Ela (lane 5). After digestion with S 1 nuclease, DNA-RNA hybrids were analyzed by electrophoresis on 2 % agarose gel at neutral pH. The fragments of pBR322 digested with EcoRI and H&I were used as size markers (lane M). (Panel 8) The same 5’ end-labelled probe as above was hybridized to poly(A)* RNA from AdS-infected HeLa cells during late in infection (lane 1) or poly(A)+ RNA from the yeast carrying YEpl3Ela (lane 2). After digestion with Sl nudease, DNA-RNA hybrids were analyzed by electrophoresis on 2% agarose gel under denaturating conditions. The size markers (lane M) are the same as in (A). (Panel C) Two-dimentional electrophoresis of DNA-RNA hybrids prepared with the same probe DNA as in (A) and poly(A)‘RNA from the yeast carrying YEpl3Ela. (Panel D) Localization and splicing pattern of the Ela transcripts in HeLa cells. Shown is a summary of the results of the electron microscopy (Chow et al., 1979), the Sl nuclease mapping (Berk and Sharp, 1977) and the DNA sequence analysis (Van Ormondt et al., 1980). The double line indicates the Ela gene and the single lines above it the transcripts early (Eta-a, Ela-b) or late (Ela-c) in infection. The sequences excised from the transcripts are shown with the thin caret lines. The numbers above the double-line represent the nucleotide uumber from the left end of the Ad5 genome. The numbers flanked by the arrows below the double-line represent the distances (in bp) between the indicated sites.

166
between the XbaI site and the 3’-end of the inter- vening sequence (lane 1). These results indicated that splicing occurred normally in HeLa cells in the present experiment. When poly(A) + RNA synthe- sized in yeast was similarly studied, the size of the protected fragment of about 800 nucleotides did not vary with pH at which the gel electrophoresis was performed (Fig. 7, A, B). The result was confirmed by two-dimensional electrophoresis as shown in Fig. 7C. We believe that the minor band of about 160 nucleotides was an artifact in the Sl mapping. The band may not be a transcript initiated about 640 bp downstream from the major start points, because no corresponding band was observed in the blot hybrid- ization analysis as shown in Fig. 2. From these results, we concluded that the Ela transcript synthe- sized in yeast contained the intervening sequence which was, however, not amenable to splicing in yeast.
DISCUSSION
We studied transcription of the Ela gene of human Ad5 in the yeast S. cerevisiae. We inserted the Ela gene with its flanking regions into the multi-copy plasmid YEpl3 (Broach et al., 1979), which carries the yeast gene LEUZ. The recombinant plasmid was then transferred into leu2 mutant yeast and the ieucine-independent transformants have been inves- tigated. The blot hybridization experiment suggested that the viral gene in question was transcribed more efficiently than the yeast gene in the transformant yeast: the Ela-specific RNA was found more abun- dantly than the LEU2-specific RNA in the yeast harboring YEpl3Ela. It is tempting to assume that the enhancer element contained in the inserted Ela DNA (Hearing and Shenk, 1983; Hen et al., 1983) is responsible for its efficient transcription beyond the normal host range. This idea is now under our experimental verification.
We further showed by the Sl-nuclease mapping that the transcription of Ela gene in the transformed yeast started at multiple sites, mostly scattering downstream from the authentic initiation site ( + 1). The majority of the transcripts started at a position of + 37. Some of the minor 5’ ends observed far upstream from the + 1 position seemed to coincide with the additional initiation sites of the Ela gene
which appear at the late phase of infection in HeLa cells (Osborne and Berk, 1983). We have compared the available data on the 5’ ends of the RNAs transcribed in yeast from genes of multicellular eukaryotes. It is interesting to note that the major ends generally are located between 50 and 70 bp downstream from the first T in the TATAAA box (Fig. 8). Homology in the sequence around the start site is not obvious, however. In this context, it would be worthwhile to recall a remarkable difference between yeast and multicellular eukaryotes in the transcription initiation by RNA polymerase II, viz. the position of TATAAA relative to the transcription start. Although the majority of the sequenced yeast protein-encoding genes have TATAAA elements somewhere within 200 bp of sequence preceding the ATG codon, the distance between TATAAA and the 5 ’ end of the corresponding mRNA varies with the gene, ranging from 37 to 180 bp (see Sentenac and Hall, 1983), in contrast to the distance of 30 bp in multicellular eukaryotes (Breathnach and Cham- bon, 198 1; see also Fig. 8). The evolutionary diver- gence of the transcription initiation machinery would therefore involve a factor that determines the dis- tance between these elements. It looks as if such a factor is absent in yeast. Many more data must be accumulated on the transcription of heterologous as well as homologous genes in the yeast before we could draw any conclusion as to the elements involved in the eflicient transcription in this microor- ganism.
We also demonstrated that the 5’ ends of the Ela transcripts synthesized in yeast, like transcripts of yeast genes, were capped at purine nucleotides, in which the ribose moiety was not methylated. This result supports the notion that the 5’ termini of
nascent pre-mRNAs in yeast are susceptible to guanylyltransferase (Sripati et al., 1976; DeKloet and Andrean, 1976).
The 3’ ends of the Ela transcripts in yeast were also heterogeneous, and were far downstream from the normal polyadenylation site generated in Ad5- infected HeLa cells. The DNA sequences required for efficient transcription termination in yeast have been proposed (Zaret and Sherman, 1982; Henikof et al., 1983). Our experiment did not reach the level of precision which could be sufficient for determining whether or not such sequences were present in the inserted Ela DNA.

167
2) F\TA~"~~AA~TTTCACA~T~T~GTTAAAA~TGGTGAGTG~TTTTTTTTTTCAAATAA~~ATGGTTGGTTTA~AAAAAAAAA
(. . ,tiAIAAAnAAA"GAAGTTTTT~TTTTTTTTA~T~ATTAAATTGAAAATCAAAATTT~ATTA~TTCAAATT, :.I '
*‘r== ! , t
ATTTAT C$GGTGAGTTCCTCAAGAGGCCACTCTTG GTG&GAGTAG?i?iTTTTCTCCTCCk&
Fig. 8. Transcriptional initiation site of foreign genes of multi-cellular eukaryotic origin in S. cerevisiae. Shown are the sequences of the 5’-flanking regions down-stream of TATAAA boxes in the non-coding strands. (1) Rabbit /?-globin gene (Beggs et al., 1980). (2) Discoidin- gene of Dicfyostelium discoideum (Jellingshaus et al., 1982). The sequence in the parentheses locates 171 bp downstream from the first TATAAA, which contains the second TATAAA and the start site functional in the slime mold. (3) The major late protein gene of the human Ad5 (H.H. and T.F., unpublished). (4) The Ela gene of Ad5 (this report). The major transcriptional starts in yeast are represented by tilled boxes above the sequences, while the minor starts by open boxes. The upward arrows indicate the initiation sites in the original host.
In agreement with the results of others (Beggs et al., 1980; Langford et al., 1983), the ~te~en~g sequences contained in the Ela RNA were not excised in yeast. As a result, the aberrant EIa transcripts accumulated to a substantial amount in yeast, which could be useful for the in vitro study as the substrate of the splicing enzyme.
ACKNOWLEDGEMENTS
This work was supported by a Grant-in-Aid for Special Project from the Ministry of Education, Science and Culture of Japan, No. 58113006 to H. Handa and No. 57106009 to T. Fukasawa. Cyto- plasmic polyhydrosis virus was a generous gift from Dr. K. Miura (The University of Tokyo), to whom our thanks are due.
REFERENCES
Andreadis, A., Hsu, Y-P., Kohlhaw, G.B. and Sahimmai, P.: Nucleotide sequence of yeast LEU.2 shows 5’-noncoding region has sequences cognate to leucine. CelI 31 (1982) 319-325.
Beggs, J.D., Van den Berg, J., Van Ooyen, A. and Weissmann, C.: Abnormal expression of chromosomal rabbit /?-globin gene in Saccharomyces cerevisiae Nature 283 (1980) 835-840.
Berk, A.J. and Sharp, P.A.: Sizing and mapping of early adenovirus mRNAs by gel electrophoresis of Sl endonu- clease-digested hybrids. Cell 12 (1977) 721-732.
Berkner, K.L. and Sharp, P.A.: Preparation of adenovirus recombinants using plasmids of viral DNA, in Gluzman, Y. (Ed.), Eukaryotic Viral Vectors. Cold Spring Harbor Labora- tory, Cold Spring Harbor, NY, 1982, pp. 193-198.
Botstein, D., Falco, S.C., Stewart. SE., Brennan, M., Scherer, S., Stinchcomb, D.T., Struhi, K. and Davis, R.W.: Sterile host yeasts (SHY): a eukaryotic system of biological contain- ment for recombinant DNA experiments. Gene 8 (1979) 17-24.
Breathnach, R. and Chambon, P.: Organization and expression of eukaryotic split genes coding for proteins. Annu. Rev, Biochem. 50 (1981) 349-383.
Broach, J.R., Strathern, J.N. and Hicks, J.B.: Transformation in yeast: development of a hybrid cloning vector and isolation of the CANi gene. Gene 8 (1979) 121-133.

168
Chow, L.T., Broker, T.R. and Lewis, J.B.: Complex splicing
patterns of RNAs from the early regions of adenovirus-2. J.
Mol. Biol. 134 (1979) 265-303.
De Kloet, S.R. and Andrean, A.G.: Methylated nucleosides in
polyadenylate-containing yeast messenger ribonucleic acid.
Biochem. Biophys. Acta 425 (1976) 401-408.
Fraenkel-Conrat, H. and Steinschneider, A.: Stepwise degra-
dation of RNA: periodate followed by aniline cleavage.
Methods Enzymol. (1967) 243-246.
Furuichi, Y. and Miura, K.: A blocked structure at the 5’
terminus of mRNA from cytoplasmic polyhedrosis virus.
Nature 253 (1975) 374-375.
Hearing, P. and Shenk, T.: The adenovirus type 5 Ela tran-
scriptional control region contains a duplicated enhancer
element. Cell 33 (1983) 695-703.
Hen, R., Borrelli, C., Sassone-Corsi, P. and Chambon, P.: An
enhancer element is located 340 base pairs upstream from the
adenovirus-2 Ela gene. Nucl. Acids Res. 11 (1983)
8747-8760.
Henikoff, S., Tatchell, K., Hall, B.D. and Nasmyth, K.A.:
Isolation of a gene from Drosophila by complementation in
yeast. Nature 289 (1981) 33-37.
Ito, H., Fukuda, Y., Murata, K. and Kimura, A.: Transformation
of intact yeast cells treated with alkali cations. J. Bacterial.
153 (1983) 163-168.
Jellinghaus, V., Schatzle, U., Schmid, W. and Roewekamp, W.:
Transcription of a Dicfyostelium discoidin- gene in yeast:
alternative promoter sites used in two different eukaryotic
cells. J.Mol. Biol. 159 (1982) 623-636.
Kiss, G.B., Pearlman, R.E., Cornish, K.V., Friesen, J.D. and
Chan, V.L.: The herpes simplex virus thymidine kinase gene
is not transcribed in Succharomyces cerevisiue. J. Bacterial.
149 (1982) 542-547.
Langford, C., Nellen, W., Niessing, J. and Gallwitz, D.: Yeast is
unable to excise foreign intervening sequences from hybrid
gene transcripts. Proc. Natl. Acad. Sci. USA 80 (1983)
1496-1500.
Maniatis, T., Fritsch, E.F. and Sambrook, J.: Molecular Cloning.
A Laboratory Manual. Cold Spring Harbor Laboratory, Cold
Spring Harbor, NY, 1982.
McMaoter, G.K. and Carmichael, G.G.: Analysis of single- and
double-stranded nucleic acids on polyacrylamide and agarose
gels by using glyoxal and acridine orange. Proc. Natl. Acad.
Sci. USA 74 (1977) 4835-4838.
Mizumoto, K. and Lipmann, F.: Transmethylation and transgua-
nylylation in 5’-RNA capping system isolated from rat liver
nuclei. Proc. Natl. Acad. Sci. USA 76 (1979) 4961-4965.
Mulligan, R.C. and Berg, P.: Expression of a bacterial gene
in mammalian cells. Science 209 (1980) 1422-1427.
Osborne, T.F. and Berk, A.J.: Far upstream initiation sites for
adenovirus early region la transcription are utilized after the
onset of viral DNA replication. J. Virol. 45 (1983) 594-599.
Petterson, U.Z. and Sambrook, J.: Amount of viral DNA in the
genome of cells transformed by adenovirus type 2. J. Mol.
Biol. 73 (1973) 125-130.
Ricciardi, R.P., Miller, J.S. and Roberts, B.: Purification and
mapping of specific mRNAs by hybridization-selection and
cell-free translation. Proc. Natl. Acad. Sci. USA 76 (1979)
4927-493 1.
Russell, P.R.: Evolutionary divergence of the mRNA tran-
scription initiation mechanism in yeast. Nature 301 (1983)
167-169.
Saiga, H., Mizumoto, K., Matsui, T. and Higashinakagawa, T.:
Determination of the transcription initiation site of Tetra-
hymena pyrijbrmis rDNA using in vitro capping of 35s
pre-rRNA. Nucl. Acids Res. 10 (1982) 4223-4236.
Sentenac, A. and Hall, B.: Yeast nuclear RNA polymerases and
their role in transcription, in Strathern, J.N., Jones, E. and
Broach, J. (Eds.), The Molecular Biology of the Yeast
Saccharomyces, Vol. 2. Cold Spring Harbor Laboratory,
Cold Spring Harbor, NY, 1983, pp. 561-606.
Sripati, C.E., Groner, Y. and Warner, J.R.: Methylated, blocked
5’ termini of yeast mRNA. .I. Biol. Chem. 251 (1976)
2898-2904.
Thomas, P.S.: Hybridization ofdenatured RNA and small DNA
fragments transferred to nitrocellulose. Proc. Natl. Acad. Sci.
USA 77 (1980) 5201-5205.
Tooze, J.: DNA Tumor Viruses. Cold Spring Harbor Laboratory,
Cold Spring Harbor, NY, 1980.
Van Ormondt, H., Maat, J. and Van Beveren, C.P.: The
nucleotide sequence of the transforming early region El of
adenovirus type 5 DNA. Gene 11 (1980) 299-309.
Weaver, R.F. and Weissmann, C.: Mapping of RNA by a
modification of the Berk-Sharp procedure: the 5’ termini of
15s /j’-globin mRNA precursor and mature 10s /I-globin
mRNA has identical map coordinates. Nucl. Acids Res. 7
(1979) 1175-l 193.
Communicated by K. Matsubara.