transcriptional analysis of alcohol and aldehyde dehydroge
TRANSCRIPT

Transcriptional analysis of alcohol and aldehyde dehydroge-nase gene families in Pholiota microspora, and estimation of their physiological roles
Surasit SUTTHIKHAMPA1), Yoshiko KAWAI2), Mirai HAYASHI2), Sophon BOONLUE3), Norihiro SHIMOMURA2), Takeshi YAMAGUCHI2) and Tadanori AIMI2)*
1)
The United Graduate School of Agricultural Sciences Tottori University, 4-101 Koyama-cho Minami, Tottori-shi, Tottori 680-8553, Japan
2)
Faculty of Agriculture, Tottori University, 4-101 Koyama-cho Minami, Tottori-shi, Tottori 680-8553, Japan
3)
Faculty of Science, Khon Kaen University,Khon Kaen 40002, Thailand
(Received 24 September 2015 / Accepted 10 January 2016)
Abstract
In this study, we analyzed two alcohol dehydrogenase (Adh), eight aldehyde dehydrogenase (Aldh) and two mannitol-1-phosphate dehydrogenase (Mpd) genes in Pholiota microspora. The transcription of Aldh1, 2, 3, Adh1, 2, and Mpd1, 2 was unaffected in liquid medium in the presence of 1 mM ethanol. However, Mpd1 expression was promoted by acetaldehyde. Therefore, Mpd1 is a candidate for an ethanol-producing alcohol dehydrogenase. On the other hand, transcription of Aldh1 and Adh2 was promoted with 3 mM veratryl alcohol; therefore, the role of Aldh1 and Adh2 might have evolved from alcohol metabolism to degradation of aromatic compounds. Transcription of Adh1, Mpd1, Mpd2 and Aldh1 was higher in primordia and fruiting bodies than mycelia. This phenomenon suggested that the response of Adh1, Mpd1, Mpd2 and Aldh1 indicates the presence of oxidative stress during fruiting body development.
Key words: Alcohol dehydrogenase, Aldehyde dehydrogenase, Ethanol, Pholiota microspora, qRT-PCR
Mushroom Science and Biotechnology, Vol. 24 (1) 16-23, 2016Copyright © 2016, Japanese Society of Mushroom Science and Biotechnology
*Corresponding author. E-mail: [email protected]
Regular Paper
Introduction
The ascomycetous yeast Saccharomyces cerevisiae
is used for efficient bioethanol fermentation. One of
the key enzymes for alcohol fermentation is alcohol
dehydrogenase (Adh). S. cerevisiae Adh1 is expressed
during anaerobic glucose fermentation and converts
acetaldehyde to ethanol1, 2)
. However, S. cerevisiae does
not have a specific enzyme that hydrolyzes the beta-1,
4-linked glucose polymers present in cellulose-rich bio-
mass3). In nature, cellulose is the most abundant biomass,
but is protected from degradation in wood by lignin.
Therefore, lignin is one of the obstacles in development of
a biochemical process for producing lignocellulosic bio-
fuel4). Wood decaying basidiomycetes such as white rot
fungi play the major role in lignin biodegradation5). Bio-
conversion of recalcitrant lignocellulose into bioethanol
using a white rot fungus is attractive and would greatly
enhance cost-effectiveness of bioethanol production in
one step6).
Production of alcohol and Adh activity in mush-
rooms belonging to the basidiomycetous fungi has been
reported. Pleurotus ostreatus, Agaricus blazei, Tricholoma
matsutake and Flammulina velutipes have been used in
wine, sake and beer production, producing 12.2%, 8.0%,
4.6% and 3.0% ethanol, respectively7-9)
. Additionally, F.
velutipes converted 1% D-glucose into ethanol at high
efficiency, up to 88% of the theoretical yield in liquid
medium10)
. On the other hand, Pholiota microspora, a
popular edible mushroom in Japan and widely used as
research material in our laboratory12)
, did not produce
ethanol. Therefore, in order to infer why P. microspora
cannot produce ethanol, we identified and analyzed two
Adh, two Mpd and eight Aldh genes, which include zinc-
containing alcohol dehydrogenases and aldehyde dehy-
drogenases expected to be related to ethanol metabolism.
Moreover, we investigated the effect of ethanol and
acetaldehyde on transcription of these genes, and discuss
the possibility of ethanol fermentation in P. microspora.
Materials and Methods
1. Fungal strains and culturing conditions
The monokaryotic strains P. microspora NGW19-6
(A4, pdx1), a pyridoxine auxotrophic mutant, and NGW12-
163 (A3, arg4), an arginine auxotrophic mutant,19, 20)
were
used. A dikaryotic strain, obtained by crossing NGW19-6
and NGW12-163 and referred to as NGW19-6/12-163, was
used for experiments.
In order to analyze the effects of different aromatic
compounds on gene expression, the NGW19-6/12-163
strain was grown on M4 agar at 25℃ for 1 week, and then
10 mycelial agar blocks (3 × 3 mm) were transferred into
20 mL of M4 medium13)
(per L: 2.20 g glucose, 0.92 g diam-
monium tartrate, 1.00 g KH2PO4, 0.26 g NaH2PO4•2H2O,
0.50 g MgSO4•7H2O, 1.00 × 10-4 g thiamine hydrochloride,

17日本きのこ学会誌
MUSHROOM SCIENCE AND BIOTECHNOLOGY
6.60 × 10-3 g CaCl2•2H2O, 5.00 × 10
-3 g FeSO4•7H2O, 3.00 ×
10-4 g MnSO4•H2O, 5.00 × 10
-4 g ZnSO4•7H2O, 6.40 × 10
-4 g
CuSO4, and 15 g agar for a solid formulation) in a 100-mL
Erlenmeyer flask supplemented with test compounds
(final concentration 0.01% lignosulfonate; 0.05 mM
2,5-xylidine; 3 mM veratryl alcohol; 0.1 mM guaiacol;
1 mM ferulic acid; 1 mM veratric acid; 0.1 mM O-anisic
acid; 1 mM ethanol; 1 mM propanol; or 1 mM acetalde-
hyde). After inoculation, the fungus was grown at 25℃
for 10 days and the mycelia were harvested by filtration
for RNA extraction.
Fruiting bodies were cultivated on a sawdust
substrate, which was prepared as follows. Beech sawdust
was mixed with rice bran at a gravimetric ratio of 5:1
and adjusted to 65% moisture content using tap water,
and this medium was placed into a 100-mL Erlenmeyer
flask, followed by autoclaving at 121℃ for 60 min. After
cooling the medium in the air, five mycelial agar blocks
(5 × 5 mm) containing NGW19-6/12-163 were inocu-
lated and incubated at 25℃. When the mycelia had
colonized the substrate (about 40 days after inoculation),
the surface layer was scratched with a spatula, and then
50 mL of sterilized distilled water was poured into the
flask. Water was removed after flasks were incubated at
15℃ overnight, and then cultivation continued at 15℃
until fruiting bodies developed. We defined the day after
water removal as day 0. Samples for RNA extraction
were taken from triplicate cultures at different stages of
the mushroom developmental cycle: mycelia at 30 and 90
days, primordia, and fruiting bodies (1 cm).
2. Genomic DNA and total RNA preparation
Genomic DNA was extracted from mycelia grown
in liquid medium according to the method of Dellaporta
et al.14)
Harvested mycelia were frozen in liquid nitrogen
and ground to a fine powder in a mortar and pestle. RNA
was extracted using a MagExtractor™
Kit (Toyobo, Osaka,
Japan) according to the manufacturer’s instructions.
cDNA was synthesized using total RNA as a template
with ReverTra Ace® qPCR RT Master Mix with gDNA
Remover kit (Toyobo). PCR was carried out using
Takara Ex Taq® polymerase (Takara Bio, Japan). The
oligonucleotide primers used in this study are listed in
Table 1. Amplified fragments were subcloned into pMD20
T-vector (Takara Bio, Japan) and sequenced.
3. P. microspora genome and retrieved genes
Whole genomic sequences of monokaryon P.
microspora NGW19-6 were determined using Illumina
HiSeq 2000 paired-end technology with CASAVA ver.
1.8.1 software, provided by Hokkaido System Science Co.,
Ltd. (Sapporo, Hokkaido, Japan), as described by Funo
et al.15)
. This sequencing run yielded 30,935,254 high-
quality filtered reads with 101-bp paired-end sequencing.
The genome was assembled using Velvet assembler (hash
length, 85 bp)16)
. The final assembly contained 4,770
contigs with a total length of 33,400,256 bp, and an N50
length of 72,431 bp. The deduced amino acid sequences
Table 1. Primer sets for cDNA and plasmid construction.
Genes Primers 5’ → 3’
Act1 PnActin1_F1 CGAAATTTCAGCTCTCGTCGT
PnActin1_R1 CTGGAGCACGGAATCGCT
Adh1 PnADH1_F1 CGATGTTATTGTCAAGCTTGCA
PnADH1_R1 CGAGACGTTCTTGTTATAGCACT
Adh2 PnADH2_F1 CCTCCAAGGGCTCATGAAGT
PnADH2_R1 ATCCTTCGCTGATCTCTGACA
Aldh1 ALDHFF2 CTGGACGTCGCATGGTATTTTCTGG
ALDHFR3 CCAACAGGGCCAAAGATCTCC
Aldh 2 PnALDH2_F1 TCATTGCGTCTGTTGTAGCAG
PnALDH2_R1 CGAGTTCCAGCGACACCT
Aldh 3 PnALDH3F2 GTATCATGGAAGCTCGGGC
PnALDH3_R1 TGAGAGACTTGAGGACCGTG
Aldh 4 PnALDH4_F1 CTGGCTGAAGGAAAGGGAGA
PnALDH4_R1 GCCGACGAGAATCTGCGC
Aldh 5 PnALDH5_F1 AGAGTTCGTGGATATTTGCGA
PnALDH5_R1 TGTTGTTCCATTCAATAGCCTGT
Aldh 6 PnALDH6_F1 GTAGCGGCACCCTTGTATTG
PnALDH6_R1 ACGACAGCAAACACCATGAC
Aldh 7 PnALDH7_F1 TGAATATAATGCTCCAGACGCA
PnALDH7_R1 ACGCGATCCCTAAATATGCTAA
Aldh 8 PnALDH8_F1 CTCGCTGCAACTATCCTTGG
PnALDH8_R1 ACGGTCGGTCGAGAAGATTG
Mpd1 Mpd-probe-F GCACATACTGCGAATAGCCC
Mpd-probe-R TAGCAATGACACGGCGACCC
Mpd2 MPD2-F2 ATCCAGTGACTCGACCTGCT
MPD2-R2 CCAAATCCTTGAGGACGTCAT
Pdc1 PD-DiF2 CAAAAGTGGAATACAGGACATCA
PDC-pR AGTGCTCTAGCCGTGCTGTCAAT
Table 2. Primer sets for qPCR.
Genes Primers 5’ → 3’
Act1 PnActin1_CF1 GCTATGCTATGTCGCGCTTGAT
PnActin1_R1 CTGGAGCACGGAATCGCT
Adh1 PnADH1_CF1 GTACAGCCGTTCTCGAGGTCGT
PnADH1_CR1 GTGGTGCTCCGTGGACTCCA
Adh2 PnADH2_CF1 GTCGGTTGTTGCCGTCAAC
PnADH2_CR1 GCCACAGTCGAGCCCTTGAT
Aldh1 ALDHFF3 GATTTCCCAGATCCAATACGATCGC
ALDHFR3 CCAACAGGGCCAAAGATCTCC
Aldh 2 PnALDH2_CF1 GCGTCTGTTGTAGCAGGTTCT
PnALDH2_CR1 GAGCAACCTAGAGCGTTCCCA
Aldh 3 PnALDH3_CF1 CAACATCGTCAACGGATATGGA
PnALDH3_CR1 CCGCTGACGCTTTCATGACT
Aldh 4 PnALDH4_CF1 CCATCTGCTATGATCACCCGA
PnALDH4_CR1 CCAGCACGTTGTGAGAGTTCA
Aldh 5 PnALDH5_CF1 CTTTACTGGCAGCGAGCACGT
PnALDH5_CR1 CAGGCATGATGATCGAAGCGT
Aldh 6 PnALDH6_CF1 GGTGTTCTGAACTTTCTGCCCA
PnALDH6_CR1 CGCGATCACTTCCAGTAAAGTT
Aldh 7 PnALDH7_CF1 GGACACTGGCAAGACGTTGA
PnALDH7_CR1 CTTGATAGCGTCTTCGCCGTA
Aldh 8 PnALDH8_CF1 GCCGACCTGCATATGACAA
PnALDH8_CR1 CCTTTCGTGTCATCACCTGA
Mpd1 MPDFF1 GCCGCAAGGGATACGAAC
MPDFR1 TACACAGTAAGACCAGCG
Mpd2 PnMPD2-F2 ATCCAGTGACTCGACCTGCT
PnMPD2-CR1 GCCGTGGCTACTTGAGAAGTCA
Pdc1 PD-FF3 CGTTACGGTGGTGTATGTATTATCT
PD-FR3 GTGACGGCGAACGGGGATG

18 Vol.24 No. 1
of known proteins from public databases were searched
against the P. microspora genome using the BLASTp
algorithm. Two alcohol dehydrogenase genes were
assigned as Adh1 and Adh2, two mannitol-1-phosphate
dehydrogenase genes were assigned as Mpd1 and Mpd2,
eight aldehyde dehydrogenase genes were assigned as
Aldh1 to Aldh8, and a pyruvate decarboxylase1 gene was
assigned as Pdc1. The coding sequences of intron-exon
junctions based on GT-AG rules17, 18)
and open reading
frames based on generic rules (start codon, ATG and stop
codon TAA, TAG, or TGA)19)
were then predicted. The
nucleotide sequences of genomic DNA fragments of P.
microspora Adh, Aldh, Mpd and Pdc genes were deposited
in the DDBJ under the following accession numbers:
Adh1 (LC102238); Adh2 (LC102239); Aldh1 (LC102241);
Aldh2 (LC102242); Aldh3 (LC102243); Aldh4 (LC102244);
Aldh5 (LC102245); Aldh6 (LC102246); Aldh7 (LC102247);
Aldh8 (LC102248); Mpd1 (AB686426); Mpd2 (LC102237);
and Pdc1 (LC102240). Protein motifs in deduced amino
acid sequences were identified using the MOTIF search
program (http://motif.genome.jp) and NCBI’s conserved
domain database20)
. Subcellular localization was predicted
using the PSORTII21)
(http://psort.hgc.jp/form2.html),
IntroPro (http://www.ebi.ac.uk/interpro)22)
and SOSUI23)
(http://harrier.nagahama-i-bio.ac.jp/sosui/sosui_submit.
html) online tools.
4. Quantitative RT-PCR (qRT-PCR) assays
The actin gene (Act1) was used as a reference gene.
Primer pairs for amplification of Adh1-2, Aldh1-8, Mpd1-2,
Pdc1 and Act1 cDNA were designed based on their cDNA
sequences using Genetyx ver. 10.0.3 software (Genetyx,
Tokyo, Japan). Amplification of genomic DNA was
prevented by designing primers for exon-exon junctions.
All primers were tested to ensure that they amplified a
single band with no primer-dimers, as shown in Table 2.
Plasmids with target gene (Adh1-2, Aldh1-8, Mpd1-2,
and Pdc1) and housekeeping gene (Act1) inserts were
extracted as described by Birnboim24)
. Standard curves
were constructed using four ten-fold dilutions of plasmid.
Real-time PCR was performed using a KOD SYBR® qPCR
Mix kit (Toyobo). Thermocycling was carried out using a
PikoReal™ 96 system (Thermo Fisher Scientific) with an
initial incubation for 1 min at 95℃, followed by 40 cycles
of 95℃ for 10 s, 60℃ for 1 min. Each run was completed
with a melting curve analysis to confirm the specificity of
amplification and absence of primer-dimers. Data analysis
was performed in accordance with the manufacturer’s
instructions.
5. Analysis of sequences and phylogenetic tree
Nucleotide and protein sequence data were analyzed
using Genetyx software. Protein sequence similarity was
analyzed using the BLASTP algorithm25)
. Peroxidase
genes were retrieved from public domain databases (NCBI
and UniProt). A phylogenetic tree was constructed by
MEGA 6.06 software26)
using the neighbor joining meth-
od with a bootstrap value of 1,000 replicates. Multiple
alignment was performed using ClustalW software27)
.
6. Statistical analysis
Mean values and standard deviation of the relative
results for each treatment were calculated. Comparisons
between control and treatment groups were made using
Student’s t-test. Differences were regarded as statistically
significant for P values under 5% (P<0.05).
Results
1. Protein sequences of Adhs and Mpds and phylogenetic
analysis
A phylogenetic tree constructed using the deduced
amino acid sequences of alcohol dehydrogenases (Adhs)
and mannitol-1-phosphate dehydrogenases (Mpds)
is shown in Fig. 1. Adhs and Mpds were clustered in
different clades. Clustered in the same clade with Mpds
were alcohol dehydrogenases from S. cerevisiae and
Aspergillus nidulans. Adh2 clustered within a group of
class III Adhs including Sfa1 [S-(hydroxymethyl) glutath-
ione dehydrogenase] from S. cerevisiae, which is related to
resistance to formaldehyde. Adh1 was clustered with A.
nidulans Adh (AN8406).
Zinc-binding dehydrogenase (PF00107) and alcohol
dehydrogenase (PF08240) motifs, and the zinc-containing
Adh signature [GHEX2GX5(G, A)X2(I, V, A, C, S), where X
is any other amino acid]28)
, were present in all Adhs and
Mpds. The NAD+-binding domains of several dehydro-
genases were analyzed. A conserved domain [GXGXXG,
where X is any other amino acid)29)
in the medium-chain
alcohol dehydrogenase/reductase) superfamily appeared
in P. microspora Mpd1 and Mpd2, a sequence belonging to
the Class I alcohol dehydrogenase Adh1 from S. cerevisiae,
which converts acetaldehyde to ethanol30)
. However,
P. microspora Mpds were clustered with A. nidulans
alcB (AN3741), which has a lower substrate specificity
to ethanol than to other alcohols such as 2-propanol,
2-pentanol and 2,3-butanediol31)
. This suggests that the
Mpd proteins of P. microspora might have lost activity
toward ethanol, and therefore, transcription of their genes
in liquid medium containing ethanol should be assessed.
2. Protein sequences of Aldhs and phylogenetic analysis
A phylogenetic tree based on protein sequences of
aldehyde dehydrogenases (Aldhs) is shown in Fig. 2. All
eight P. microspora Aldhs were clustered in different
clades. Aldh1 clustered with Ustilago maydis Iad1
(indole-3-acetaldehyde dehydrogenase), involved in
indole-3-acetic acid metabolism32)
, as well as the ethanol
metabolism related Aldh of S. cerevisiae and A. nidulans,
a predicted Aldh in Arabidopsis thaliana, and retinalde-
hyde dehydrogenase of Homo sapiens. Each cluster of
Aldh2 and Aldh3 grouped with other predicted proteins
of basidiomycetes. However, Aldh2 and Aldh3 clusters
were close to Aldh1, which suggests that Aldh1, 2 and
3 might have similar roles in P. microspora. In addition,
Aldh4 to Aldh8 had predicted functions based on
similarities to succinate semialdehyde dehydrogenase of

19日本きのこ学会誌
MUSHROOM SCIENCE AND BIOTECHNOLOGY
A. bisporus33)
, Δ1-piperideine-6-carboxylate dehydroge-
nases related to human Aldh7A134)
, a putative salicylalde-
hyde dehydrogenase, a protein involved in meiotic sister-
chromatid recombination of eukaryotic organisms35)
and
Δ-1-pyrroline-5-carboxylate dehydrogenase of A. bisporus
pruA36)
. All Aldh proteins belonged to the aldehyde
dehydrogenase family (PF0017) and were predicted to be
soluble cytoplasmic proteins.
3. Ethanol was ineffective in inducing transcription of
Adh, Aldh or Mpd
To investigate the relationship between ethanol
metabolism and Adh and Aldh, transcription of the Adh
and Aldh genes in mycelia grown in liquid medium
supplemented with 1 mM ethanol, 1 mM propanol or
1 mM acetaldehyde was analyzed by qRT-PCR. Tran-
scription levels of Aldh1, 2 and 3 are shown in Fig. 3A
and levels of Adh1 and 2 and Pdc1 are shown in Fig. 3B.
Ethanol, propanol and acetaldehyde did not promote the
transcription of Adh, Aldh or Pdc1. Mpd2 also showed
a relatively low level of transcription in liquid medium
supplemented with ethanol, propanol or acetaldehyde.
Moreover, ethanol did not promote transcription of
either Mpd gene (Fig. 3C). However, the presence of 1
mM acetaldehyde significantly promoted transcription
of only Mpd1, which has a protein sequence most similar
to that of Adhs of A. nidulans (AN3741) and S. cerevisiae
Fig. 1. Phylogenetic tree of alcohol dehydrogenases. The tree was
constructed by the neighbor joining method with 1000 bootstrap
replications of P. microspora Adh1, Adh2, Mpd1 and Mpd2 with
corresponding sequences from Agaricus bisporus, Arabidopsis
thaliana, Aspergillus nidulans, Coprinopsis cinerea, Homo sapiens,
Laccaria bicolor, Pleurotus ostreatus and Saccharomyces
cerevisiae. The branches are identified by organism and the gene
name, then protein ID.
Fig. 2. Phylogenetic tree of aldehyde dehydrogenases. The tree was
constructed by the neighbor joining method with 1000 bootstrap
replications of P. microspora aldehyde dehydrogenases (Aldh1-
8) and Aldhs of eukaryotic organisms grouped according to gene
family: white-rot fungi Agaricus bisporus, Coprinopsis cinerea,
Hypholoma sublateritium, Laccaria bicolor and Phanerochaete
carnosa; a single-celled fungus, Saccharomyces cerevisiae;
filamentous fungus Aspergillus nidulans; dimorphic fungus Ustilago
maydis; Animalia Homo sapiens; and Planta Arabidopsis thaliana.
Aldhs fell into six clusters: Aldh1-3 clustered with H. sapiens, A.
thaliana, A. nidulans, U. maydis and S. cerevisiae. Aldh4 was
identified as a succinate semialdehyde dehydrogenase and Aldh5
as related to Δ1-piperideine-6-carboxylate dehydrogenases of A.
thaliana and H. sapiens. Aldh6 was closely related to a putative
salicylaldehyde dehydrogenase of A. nidulans (AN4050). Aldh7
was closely related to a protein involved in meiotic sister-chromatid
recombination in S. cerevisiae. Aldh8 was a Δ1-pyrroline-5-
carboxylate dehydrogenase. The branches are identified by
organism and the gene name, then protein ID.

20 Vol.24 No. 1
(YOL086C). Adhs and Mpd2 from P. microspora do not
seem related to ethanol metabolism. However, based on
these results, if Mpd1 has high enough activity to convert
acetaldehyde to ethanol, it is possible that P. microspora
can become an alcohol producer.
4. Gene expression of Aldh and Adh in the presence of
aromatic compounds
In order to predict the roles of Adh1 and 2 and Aldh1,
2 and 3, their transcription in mycelia grown in liquid
medium supplemented with several aromatic compounds
was examined by qRT-PCR. The transcription levels
of Aldh1, 2 and 3 are shown in Fig. 4A. Transcription
of Aldh1 was significantly promoted by 3 mM veratryl
alcohol and 0.1 mM guaiacol. Transcription of Aldh2 was
unaffected by any aromatic compounds tested. Tran-
scription of Aldh3 was promoted significantly by 0.1 mM
guaiacol. On the other hand, the transcriptional levels of
Adh1 and 2 are shown in Fig. 4B. Transcription of Adh1
was unaffected by any tested aromatic compounds, but
transcription of Adh2 was significantly promoted by 3
mM veratryl alcohol. These results indicated that Aldh
and Adh transcription are responsive to the presence of
aromatic compounds; therefore, the role of the Aldh and
Adh genes might have evolved from ethanol fermentation
to degradation of aromatic compounds related to the ones
we tested, such as lignin and lignin-like compounds. More-
over, because veratryl alcohol promotes fruiting body
formation in P. ostreatus, we investigated gene expression
during cultivation of fruiting bodies on sawdust medium.
5. Adh2 was highly transcribed during mycelial growth
in sawdust medium, but Adh1, Aldh1 and Mpd1 were
highly transcribed in fruiting bodies
Transcription of Adh1 and 2, Aldh1, 2 and 3, and
Mpd1 and 2 genes during mycelial growth and in
primordia and fruiting bodies was examined by qRT-PCR
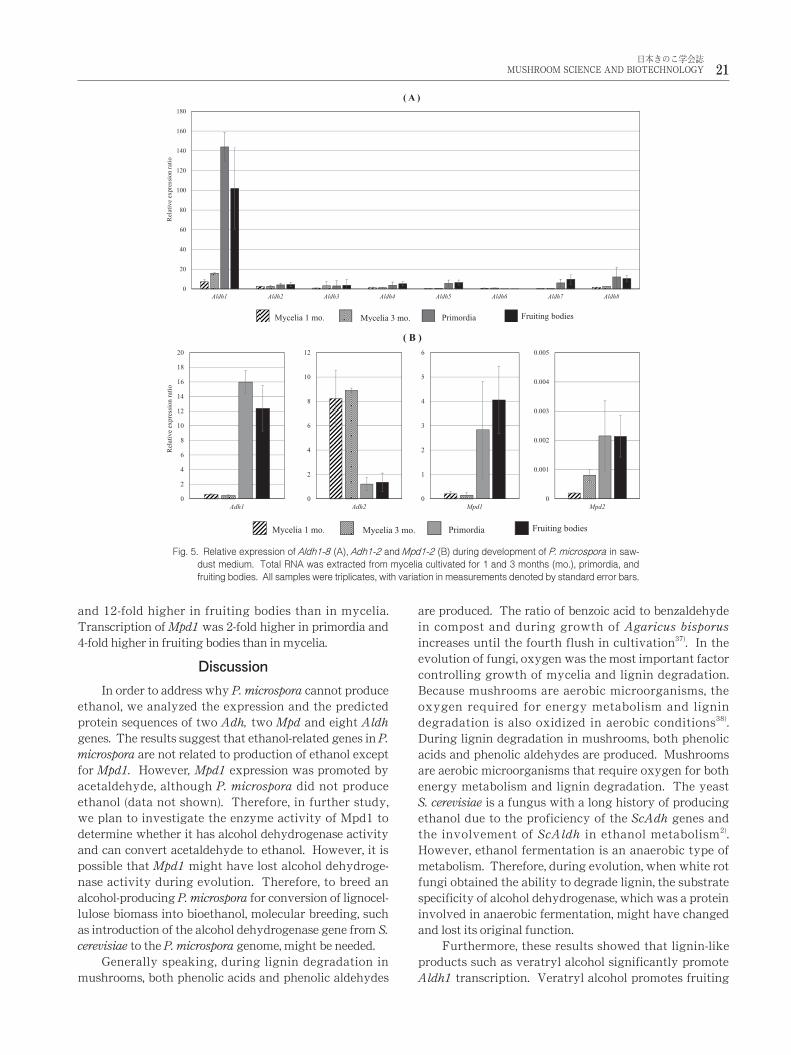
(Fig. 5). The expression of the eight aldehyde dehydroge-
nase genes is displayed in Fig. 5A. Transcription of Aldh1
in primordia and fruiting bodies was respectively 130- and
100-fold higher than in mycelia. However, transcription of
the other Aldh genes remained the same in all developmen-
tal stages including mycelia. Transcription of Adhs and
Mpd genes during mycelial growth and in primordia and
fruiting bodies was also examined by qRT-PCR (Fig. 5B).
Only transcription of Adh2 in mycelia was higher than
that of Adh2 (by 6-fold) in primordia and fruiting bodies.
The transcription of Adh1 was 16-fold higher in primordia
Fig. 3. Relative expression of Aldh1-3 (A), Adh1-2 and Pdc1 (B) and
Mdh1-2 (C) in P. microspora mycelia cultured in M4 medium with
ethanol, propanol and acetaldehyde. Total RNA was measured on
day 10 by qRT-PCR in triplicate, with variation denoted by standard
error bars. Asterisks indicate a significant difference in expression
between control basal medium and medium supplemented with
ethanol, propanol or acetaldehyde (t-test, p < 0.05).
Fig. 4. Relative expression of Aldh1-3 (A) and Adh1-2 (B) in P.
microspora mycelia cultured in M4 medium with aromatic
compounds. Total RNA was measured on day 10 by qRT-
PCR in triplicate, with variation denoted by standard error bars.
Asterisks indicate a significant difference in expression between
control basal medium and medium supplemented with aromatic
compounds (t-test, p < 0.05).
21日本きのこ学会誌
MUSHROOM SCIENCE AND BIOTECHNOLOGY
and 12-fold higher in fruiting bodies than in mycelia.
Transcription of Mpd1 was 2-fold higher in primordia and
4-fold higher in fruiting bodies than in mycelia.
Discussion
In order to address why P. microspora cannot produce
ethanol, we analyzed the expression and the predicted
protein sequences of two Adh, two Mpd and eight Aldh
genes. The results suggest that ethanol-related genes in P.
microspora are not related to production of ethanol except
for Mpd1. However, Mpd1 expression was promoted by
acetaldehyde, although P. microspora did not produce
ethanol (data not shown). Therefore, in further study,
we plan to investigate the enzyme activity of Mpd1 to
determine whether it has alcohol dehydrogenase activity
and can convert acetaldehyde to ethanol. However, it is
possible that Mpd1 might have lost alcohol dehydroge-
nase activity during evolution. Therefore, to breed an
alcohol-producing P. microspora for conversion of lignocel-
lulose biomass into bioethanol, molecular breeding, such
as introduction of the alcohol dehydrogenase gene from S.
cerevisiae to the P. microspora genome, might be needed.
Generally speaking, during lignin degradation in
mushrooms, both phenolic acids and phenolic aldehydes
are produced. The ratio of benzoic acid to benzaldehyde
in compost and during growth of Agaricus bisporus
increases until the fourth flush in cultivation37)
. In the
evolution of fungi, oxygen was the most important factor
controlling growth of mycelia and lignin degradation.
Because mushrooms are aerobic microorganisms, the
oxygen required for energy metabolism and lignin
degradation is also oxidized in aerobic conditions38)
.
During lignin degradation in mushrooms, both phenolic
acids and phenolic aldehydes are produced. Mushrooms
are aerobic microorganisms that require oxygen for both
energy metabolism and lignin degradation. The yeast
S. cerevisiae is a fungus with a long history of producing
ethanol due to the proficiency of the ScAdh genes and
the involvement of ScAldh in ethanol metabolism2)
.
However, ethanol fermentation is an anaerobic type of
metabolism. Therefore, during evolution, when white rot
fungi obtained the ability to degrade lignin, the substrate
specificity of alcohol dehydrogenase, which was a protein
involved in anaerobic fermentation, might have changed
and lost its original function.
Furthermore, these results showed that lignin-like
products such as veratryl alcohol significantly promote
Aldh1 transcription. Veratryl alcohol promotes fruiting
Fig. 5. Relative expression of Aldh1-8 (A), Adh1-2 and Mpd1-2 (B) during development of P. microspora in saw-
dust medium. Total RNA was extracted from mycelia cultivated for 1 and 3 months (mo.), primordia, and
fruiting bodies. All samples were triplicates, with variation in measurements denoted by standard error bars.

22 Vol.24 No. 1
body formation and shortens the culture period in P.
ostreatus39)
. Therefore, Aldh1 might have an important
role involving various aromatic compounds in fruiting
bodies. In this study, we investigated Ald, Aldh and
Mpd expression in P. microspora, finding that all were
incapable of producing ethanol. Therefore, we will
further study introduction and expression of S. cerevisiae
Adh in P. microspora to investigate the feasibility of
ethanol production by P. microspora.
Acknowledgment This work was partially supported by
Grant-in-Aid for Scientific Research (C) 15K07514 by the
Japan Society for the Promotion of Science (JSPS).
和 文 摘 要
Pholiota microspora におけるアルコールと
アルデヒド脱水素酵素遺伝子ファミリー
の転写分析,その生理的役割の推定
Surasit SUTTHIKHAMPA1)・河井 祥2)・林 未来2)
Sophon BOONLUE3)・霜村典宏2)・山口武視2)
• 會見忠則2)
1) 鳥取大学大学院連合農学研究科
〒680-8553 鳥取県鳥取市湖山町南 4-101
2) 鳥取大学農学部
〒680-8553 鳥取県鳥取市湖山町南 4-101
3) Faculty of Science, Khon Kaen University,
Khon Kaen 40002, Thailand
本研究では,Pholiota microspora での 2 つのアルコール
脱水素酵素(Adh),8 つのアルデヒド脱水素酵素(Aldh)
と 2 つのマンニトール -1- リン酸脱水素酵素(Mpd)遺伝子
の発現を分析した結果,Aldh1, 2, 3, Adh1, 2,及び Mpd1, 2
の転写は,1 mM のエタノールの存在下の液体培養におい
て全く影響を受けなかった.しかし,Mpd1 の発現はアセト
アルデヒドで促進された.したがって,Mpd1 は,エタノー
ル生産のためのアルコール脱水素酵素の候補である.一方,
Aldh1 と Adh2 の転写は,3 mM のベラトリルアルコール
の存在で促進された.したがって,Aldh1とAdh2の役割は,
アルコール代謝から,芳香族化合物の分解に進化した可能
性があると考えられた.Adh1,Mpd1,Mpd2 と Aldh1 の
転写は菌糸より原基および子実体で高かった.この現象は
Adh1,Mpd1,Mpd2 と Aldh1 の応答は子実体形成中に酸
化ストレスが存在することを示唆していた.
References
1) Young, T, Williamson, V, Taguchi, A, Smith, M, Sledziewski,
A, Russell, D, Osterman, J, Denis, C, Cox, D and Beier, D: The
alcohol dehydrogenase genes of the yeast, Saccharomyces
Cerevisiae: isolation, structure, and regulation, In: Hollaender
A, DeMoss, R, Kaplan, S, Konisky, J, Savage, D and Wolfe, R,
editors, Genetic engineering of microorganisms for chemicals,
Basic Life Sciences: Springer US, pp 335-361 (1982)
2) Wills, C: Regulation of sugar and ethanol metabolism in
Saccharomyces cerevisiae, Crit Rev Biochem Mol Biol, 25, 245-
80 (1990)
3) Van Rensburg, P, Van Zyl, W H, and Pretorius, I S: Engineer-
ing yeast for efficient cellulose degradation, Yeast, 14, 67-76
(1998)
4) Kirk, T K and Farrell, R L: Enzymatic ‘combustion’: the
microbial degradation of lignin, Ann Rev Microbiol, 41, 465-
505 (1987)
5) Zeng, Y, Zhao, S, Yang, S and Ding, S Y: Lignin plays a nega-
tive role in the biochemical process for producing lignocellu-
losic biofuels, Curr Opin Biotechnol, 27, 38-45 (2014)
6) Lynd, L R, Van Zyl, W H, McBride, J E and Laser, M:
Consolidated bioprocessing of cellulosic biomass: an update,
Curr Opin Biotechnol, 16, 577-583 (2005)
7) Okamura, T, Ogata, T, Toyoda, M, Tanaka, M, Minamimoto,
N, Takeno, T, Noda, H, Fukuda, S and Ohsugi, M: Production of
sake by mushroom fermentation, Mushroom Sci Biotechnol, 8,
109-114 (2000)
8) Okamura, T, Ogata, T, Minamoto, N, Takeno, T, Noda, H,
Fukuda, S and Ohsugi, M: Characteristics of wine produced by
mushroom fermentation, Biosci Biotechnol Biochem, 65, 1596-
1600 (2001)
9) Okamura-Matsui, T, Tomoda, T, Fukuda, S and Ohsugi, M:
Discovery of alcohol dehydrogenase from mushrooms and
application to alcoholic beverages, J Mol Catalys B: Enzymatic,
23, 133-144 (2003)
10) Mizuno, R, Ichinose, H, Maehar, T, Takabatake, K and
Kaneko, S: Properties of ethanol fermentation by Flammulina
velutipes, Biosci Biotechnol Biochem, 73, 2240-2245 (2009)
11) Masuda, P, Yamanaka, K, Sato, Y and Kitamoto, Y: Nuclear
selection in monokaryotization of dikaryotic mycelia of
Pholiota nameko as described by leading and following nuclei,
Mycoscience, 36, 413-420 (1995)
12) Yi, R, Tachikawa, T, Mukaiyama, H, Mochida, Y, Ishikawa,
M and Aimi, T: DNA-mediated transformation system in a
bipolar basidiomycete, Pholiota microspora (P. nameko), Myco-
science, 50, 123-129 (2009)
13) Johansson, T, Nyman, P O and Cullen, D: Differential regu-
lation of mnp2, a new manganese peroxidase-encoding gene
from the ligninolytic fungus Trametes versicolor PRL 572,
Appl Environ Microbiol, 68, 2077-2080 (2002)
14) Dellaporta, S, Wood, J and Hicks, J: A plant DNA miniprepa-
ration: Version II, Plant Mol Biol Rep, 1, 19-21 (1983)
15) Funo, K, Kitagawa, W, Tanaka, M, Sone, T, Asano, K and
Kamagata, Y: Draft genome sequence of Tomitella biformata
AHU 1821T, isolated from a Permafrost Ice Wedge in Alaska,
Genome Announcements, 2, e00066-14 (2014)
16) Zerbino, D R and Birney, E: Velvet algorithms for de novo
short read assembly using de Bruijn graphs, Genome Res, 18,
821-829 (2008)
17) Breathnach, R, Benoist, C, O’Hare, K, Gannon, F and
Chambon, P: Ovalbumin gene: evidence for a leader sequence
in mRNA and DNA sequences at the exon-intron boundaries,
Proc Natl Acad Sci, 75, 4853-4857 (1978)
18) Wu, Q and Krainer, A R: AT-AC Pre-mRNA Splicing mecha-
nisms and conservation of minor introns in voltage-gated ion
channel genes, Mol Cell Biol, 19, 3225-3236 (1999)

23日本きのこ学会誌
MUSHROOM SCIENCE AND BIOTECHNOLOGY
19) Brown, T A: Gene cloning and DNA analysis: an introduc-
tion, 6th
edition: Wiley-Blackwell, p 208 (2010)
20) Nakai, K and Kanehisa, M: A knowledge base for predicting
protein localization sites in eukaryotic cells, Genomics, 14, 897-
911 (1992)
21) Marchler-Bauer, A, Derbyshire, M K, Gonzales, N R, Lu, S,
Chitsaz, F, Geer, L Y, Geer, R C, He, J, Gwadz, M, Hurwitz, D
I, Lanczycki, C J, Lu, F, Marchler, G H, Song, J S, Thanki, N,
Wang, Z, Yamashita, R A, Zhang, D, Zheng, C and Bryant, S H:
CDD: NCBI’s conserved domain database, Nucleic Acids Res,
43, D222-D6 (2015)
22) Jones, P, Binns, D, Chang, H Y, Fraser, M, Li, W, McAnulla,
C, McWilliam, H, Maslen, J, Mitchell, A, Nuka, G, Pesseat, S,
Quinn, A F, Sangrador-Vegas, A, Scheremetjew, M, Yong, S Y,
Lopez, R and Hunter, S: InterProScan 5: genome-scale protein
function classification, Bioinformatics, 30, 1236-1240 (2014)
23) Hirokawa, T, Boon-Chieng, S and Mitaku, S : classification
and secondary structure prediction system for membrane
proteins, Bioinformatics, 14, 378-379 (1998)
24) Birnboim, H C: A rapid alkaline extraction method for the
isolation of plasmid DNA, Methods Enzymol, 100, 243-255
(1983)
25) Altschul, S F, Madden, T L, Schäffer, A A, Zhang, J, Zhang, Z,
Miller, W and Lipman, D J: Gapped BLAST and PSI-BLAST: a
new generation of protein database search programs, Nucleic
Acids Res, 5, 3389-3402 (1997)
26) Tamura, K, Stecher, G, Peterson, D, Filipski, A and Kumar, S:
MEGA6: Molecular evolutionary genetics analysis version 6.0,
Mol Biol Evol, 30, 2725-2729 (2013)
27) Larkin, M A, Blackshields, G, Brown, N P, Chenna, R,
McGettigan, P A, McWilliam, H, Valentin, F, Wallace, I M,
Wilm, A, Lopez, R, Thompson, J D, Gibson, T J and Higgins, D
G: ClustalW and ClustalX version 2, Bioinformatics, 23, 2947-
2948 (2007)
28) Persson, B, Hallborn, J, Walfridsson, M, Hahn-Hägerdal, B,
Keränen, S, Penttilä, M and Jörnvall, H: Dual relationships of
xylitol and alcohol dehydrogenases in families of two protein
types, FEBS Lett, 324, 9-14 (1993)
29) Lesk, A M: NAD-binding domains of dehydrogenases, Curr
Opin Struct Biol, 5, 775-783 (1995)
30) Bennetzen, J L and Hall, B D: The primary structure of the
Saccharomyces cerevisiae gene for alcohol dehydrogenase, J
Biol Chem, 257, 3018-3025 (1982)
31) Sealy-Lewis, H M and Fairhurst, V: Substrate specificity of
nine NAD+-dependent alcohol dehydrogenases in Aspergillus
nidulans, Microbiology, 141, 2295-300 (1995)
32) Basse, C W, Lottspeich, F, Steglich, W and Kahmann, R:
Two Potential indole-3-acetaldehyde dehydrogenases in the
phytopathogenic fungus Ustilago maydis, Eur J Biochem, 242,
648-656 (1996)
33) Baldy, P: Metabolisme du γ-aminobutyrate chez Agari-
cus bisporus III, La succinate-semialdehyde:NAD(P)+
oxydoreductase, Physiol Plant, 40, 91-97 (1977)
34) Brocker, C, Lassen, N, Estey, T, Pappa, A, Cantore, M, Orlova,
V V, Chavakis, T, Kavanagh, K L, Oppermann, U and Vasiliou,
V: Aldehyde dehydrogenase 7A1 (ALDH7A1) is a novel
enzyme involved in cellular defense against hyperosmotic
stress, J Biol Chem, 285, 18452-18463 (2010)
35) van Heemst, D and Heyting, C: Sister chromatid cohesion
and recombination in meiosis, Chromosoma, 109, 10-26 (2000)
36) Schaap, P J, Müller, Y, Sonnenberg, A S, van Griensven,
L J and Visser, J: The Agaricus bisporus pruA gene encodes
a cytosolic delta 1-pyrroline-5-carboxylate dehydrogenase
which is expressed in fruit bodies but not in gill tissue, Appl
Environ Microbiol, 63, 57-62 (1997)
37) Iiyama, K, Stone, B A and Macauley, B J: Compositional
changes in compost during composting and growth of Agaricus
bisporus, Appl Environ Microbiol, 60, 1538-1546 (1994)
38) Gessler, N N, Aver’yanov, A A and Belozerskaya, T A:
Reactive oxygen species in regulation of fungal development,
Biochemistry (Moscow), 72, 1091-1109 (2007)
39) Suguimoto, H H, Barbosa, A M, Dekker, R F H and Castro-
Gomez, RJH: Veratryl alcohol stimulates fruiting body
formation in the oyster mushroom, Pleurotus ostreatus, FEMS
Microbiol Lett, 194, 235-238 (2001)