transcriptional complementarity in breast cancer: application to detection of circulating tumor...
TRANSCRIPT

Transcriptional Complementarity in Breast Cancer:Application to Detection of Circulating Tumor Cells
RAYMOND L. HOUGHTON, PhD,* DAVIN C. DILLON, PhD,*DAVID A. MOLESH, BA,* BARBARA K. ZEHENTNER, PhD,*
JIANGCHUN XU, MD, PhD,* JOHN JIANG, PhD,* CHERYL SCHMIDT, MS,*ANTHONY FRUDAKIS, PhD,† ELIZABETH REPASKY, PhD,‡
ARISTIDES MALTEZ FILHO, MD,§ MARCOS NOLASCO, MD,§
ROBERTO BADARO, MD,§ XINQUN ZHANG, MS,* PATRICK C. ROCHE, PhD,�
DAVID H. PERSING, MD, PhD,* STEVEN G. REED, PhD*
Sarasota, Florida; Buffalo, New York; Rochester, Minnesota; Bahia, Brazil; Seattle, Washington
Background: We used a combination of genetic subtraction, silicon DNA micro-array analysis, and quantitative PCR to identify tissue- and tumor-specific genes asdiagnostic targets for breast cancer.Methods and Results: From a large number of candidate antigens, several specificsubsets of genes were identified that showed concordant and complementary ex-pression profiles. Whereas transcriptional profiling of mammaglobin resulted in thedetection of 70% of tumors in a panel of 46 primary and metastatic breast cancers,the inclusion of three additional markers resulted in detection of all 46 specimens.Immunomagnetic epithelial cell enrichment of circulating tumor cells from theperipheral blood of patients with metastatic breast cancer, coupled with RT-PCR–based amplification of breast tumor–specific transcripts, resulted in the de-tection of anchorage-independent tumor cells in the majority of patients with breastcancer with known metastatic disease.Conclusion: Complementation of mammaglobin with three additional genes in RT-PCR increases the detection of breast cancers in tissue and circulating tumor cells.Key words: breast cancer, genes, quantitative PCR, complementation.
Breast cancer is second only to lung cancer inmortality among women in the United States, re-sulting in approximately 40,000 to 50,000 deaths
annually [1]. The recent introduction of immuno-therapeutic approaches to breast cancer treatmenttargeted toHer2/neu have provided significant mo-tivation to identify additional breast cancer–specificgenes as targets for therapeutic antibodies and T-cell vaccines [2–8]. Mammaglobin, identified asone of the most breast-specific genes discovered todate, is expressed in approximately 70% to 80% ofbreast cancers and is a candidate for T-cell vaccinedevelopment [9–11].
Early detection of breast cancer is a critical de-terminant in the outcome of therapies [12,13]. Be-cause of its highly tissue-specific distribution, de-tection of mammaglobin gene expression has been
From *Corixa Corporation, Seattle, WA; †DNAPrint Ge-nomics, Sarasota, FL; ‡Roswell Park Cancer Institute, Buffalo,NY; �Department of Pathology and Laboratory Medicine, MayoClinic, Rochester, MN; and §Hospital Aristides Maltez, Salva-dor, Bahia, Brazil.
Supported in part by grants no. CA-75794 and CA-80518from the National Cancer Institute.
Reprint requests: Raymond L. Houghton, PhD, Corixa Cor-poration, 1124 Columbia St, Ste 200, Seattle, WA 98104.Email: [email protected]
Copyright © 2001 by Churchill Livingstone�1084-8592/01/0602-0002$35.00/0doi:10.1054/modi.2001.23098
Molecular Diagnosis Vol. 6 No. 2 2001
79

used to identify micrometastatic lesions in lymphnode tissues [10,14,15] and, more recently, detectcirculating breast cancer cells in peripheral blood ofpatients with breast cancer with known primary andmetastatic lesions [16]. However, because mamma-globin gene expression is not a universal feature ofbreast cancer, additional markers likely will be nec-essary to achieve adequate sensitivity for the detec-tion of micrometastases in lymph nodes and bonemarrow or recognition of anchorage-independentcells in the peripheral circulation [14–16].
Our laboratory used a combination of geneticsubtraction, DNA microarray analysis, and real-time PCR to discover therapeutic and diagnostictarget candidates in breast cancer. In this report, wedescribe the expression profiles of several such can-didates, with a goal of identifying a combination oftargets expressed in all breast cancers. Preliminarydata are presented to suggest that detection of cir-culating cancer cells in peripheral blood by immu-nomagnetic cell capture followed by RT-PCR forthree or more of the described targets may be auseful diagnostic procedure.
Materials and Methods
Tumor Samples
Normal and tumor-tissue samples were providedby Dr. Elizabeth Repasky (Roswell Park CancerInstitute, Buffalo, NY) and Dr. Roberto Badaro(Hospital Aristides Maltez, Salvador, Brazil). Nor-mal and tumor-tissue samples were also obtainedfrom the National Disease Research Interchangeand Cooperative Human Tissue Network and in-clude clinical information and pathology reports.
Blood Samples
Complementary DNA (cDNA) prepared fromblood samples (4 mL) from patients with metastaticbreast cancer were obtained from Dr. Patrick Roche(Mayo Medical Laboratories, Rochester, MN). Nor-mal blood samples were obtained from an in-housedonor program at Corixa Corporation (Seattle,WA). cDNA was isolated as described next.
RNA Extraction
Total RNA was extracted from liquid nitrogen–frozen tumors and normal tissues as follows. Tissuesamples were homogenized in Trizol reagent
(Gibco BRL, Bethesda, MD) using a polytron ho-mogenizer (Kinematica, Brinkmann Instruments,Inc, Westbury, NY). Poly A+ RNA was isolatedusing a Qiagen oligotex spin column messengerRNA (mRNA) purification kit.
Differential display was performed as describedin the literature [17], but with the following modi-fications: (1) PCR amplification products were vi-sualized on silver-stained gels, (2) geneticallymatched pairs of tissues were used to eliminatepolymorphic variation, and (3) two different dilu-tions of cDNA were used as templates to eliminatedilutional effects [18].
cDNA library subtraction was performed as de-scribed with some modification [19]. The breasttumor library (tracer) made from a pool of threebreast tumors was subtracted with normal breastlibrary (driver) to identify breast tumor–specificgenes. More recently, subtractions have used six toten normal tissues as driver to subtract commongenes more efficiently, with an emphasis on essen-tial tissues along with one immunologic tissue (e.g.,spleen, lymph node, or peripheral-blood mono-nuclear cells [PBMCs]), to assist in the removal ofcDNAs derived from lymphocyte infiltration in tu-mors. The breast tumor–specific subtracted cDNAlibrary was generated as follows: driver cDNA li-brary was digested withEcoR I, Not I, andSfu I (SfuI cleaves the vector), filled in with DNA polymer-ase klenow fragment. After phenol-chloroform ex-traction and ethanol precipitation, DNA was labeledwith Photoprobe biotin (Vector Labs, Burlingame,CA) and dissolved in water. Tracer cDNA librarywas digested withBamH I and Xho I and phenolchloroform was extracted, passed through Chro-maspin 400 columns (Clontech Labs, Palo Alto,CA), ethanol precipitated, and mixed with driverDNA for hybridization at 68°C for 20 hours (longhybridization). The reaction mixture was then sub-jected to the streptavidin treatment followed byphenol-chloroform extraction four times. Sub-tracted DNA was precipitated and subjected to hy-bridization at 68°C for 2 hours with driver DNAagain (short hybridization). After removal of bioti-nylated double-stranded DNA, subtracted cDNAwas ligated into pBCSK+ and transformed intoElectroMax Escherichia coli DH10B cells (LifeTechnologies, Rockville, MD) by electroporation.To analyze the subtracted cDNA library, plasmidDNA was prepared from 100 to 200 randomlypicked independent clones and characterized by
80 Molecular Diagnosis Vol. 6 No. 2 June 2001

DNA sequencing and searching the most recentGenbank and human expressed sequence tag data-bases.
PCR Subtraction
PCR subtraction was performed as described inthe literature [20,21]. Briefly, this type of subtrac-tion works by ligating two different adapters to dif-ferent aliquots of a restriction enzyme–digestedtester (breast tumor) cDNA sample, followed bymixing tester aliquots separately with excess driver(without adapters). This first hybridization resultsin normalization of single-stranded tester-specificcDNA because of the second-order kinetics of hy-bridization. These separate hybridization reactionsare then mixed without denaturation, and a secondhybridization is performed that produces the targetmolecules: double-stranded cDNA fragments con-taining both of the different adapters. Two roundsof PCR are performed, which results in the expo-nential amplification of the target population mol-ecules (normalized tester-specific cDNAs), whereasother fragments are either unamplified or only am-plified in a linear manner.
The subtractions performed included a pool ofbreast tumors subtracted with a pool of normal breastand a pool of breast tumors subtracted with a pool ofnormal tissues, including PBMCs, brain, pancreas,liver, small intestine, stomach, heart, and kidney.
RT. Before cDNA synthesis, RNA was treatedwith DNase I (Ambion, Austin, TX) in the presenceof RNasin (Promega Biotech, Madison, WI) to re-move DNA contamination. The cDNA for use inreal-time PCR tissue panels was prepared using 25�L Oligo dT (Boehringer-Mannheim) primer withsuperscript II reverse transcriptase (Gibco BRL).
Microarray. mRNA expression of the genes ofinterest was performed as follows. cDNA for thedifferent genes was prepared as described previ-ously and arrayed on a glass slide (Incyte, PaloAlto, CA). The arrayed cDNA was then hybridizedwith a 1:1 mixture of Cy3 or Cy5 fluorescent-labeled first-strand cDNAs obtained from polyA+RNA from breast tumors, normal breast and normaltissues, and other tumors [22]. Typically, Cy3(probe 1) is attached to cDNAs from breast tumors,and Cy5 (probe 2), to normal breast tissue or othernormal tissues. Both probes are allowed to competewith the immobilized gene-specific cDNAs on thechip, washed, then scanned for fluorescence inten-
sity of the individual Cy3 and Cy5 fluorescence todetermine the extent of hybridization. Data wereanalyzed using GEMTOOLS software (Incyte),which enables the relative overexpression patternsof breast tumors to be compared with normal tissuesby the ratios of Cy3-Cy5. Mammaglobin,B511S,B305D, andB311D were analyzed with one set ofchips and probes, whereasB726P and�-aminobu-tyrate type A receptor� (GABA�) subunits wereanalyzed on a second set of chips.
Quantitative Real-time PCR. Specificity andsensitivity of the different genes were determinedby using quantitative PCR analysis. Breast metas-tases, breast tumors, benign breast disorders, andnormal breast tissue, along with other normal tis-sues and tumors, were tested in quantitative (real-time) PCR. This was performed either on the ABI7700 Prism using specific primers and Taqmanprobes [23] or on a GeneAmp 5700 sequence de-tection system (Applied Biosystems, Foster City,CA) using SYBR green, a fluorescent dye that onlybinds to double-stranded DNA and similar primersto the 7700 with no probe [24]. Matching primersand fluorescent probes were designed for each ofthe genes according to the Primer Express program(PE Biosystems). Forward primer, reverse primer,and probe sequences are listed in Table 1. Concen-trations used in the quantitative PCR for the for-ward primers for mammaglobin,GABA�, B305DC-form, B311D, B511S, B533S, and B726P were900, 900, 300, 900, 900, 300, and 300 nM, respec-tively. Concentrations for the reverse primers were300, 900, 900, 900, 300, 900, and 900 nM, respec-tively. Primers and probes were used in the univer-sal thermal cycling program in real-time PCR. Thereaction was performed in 25-�L volumes. The fi-nal probe concentration in all cases was 160 nM.dATP, dCTP, and dGTP were 0.2 mM, and dUTPwas 0.4 mM. AmpliTaq gold and Amperase UNG(PE Biosystems) were used at 0.625 and 0.25 unitsper reaction, respectively. MgCl2 was at a final con-centration of 5 mM. Trace amounts of glycerol,gelatin, and Tween 20 (Sigma Chemical Co, StLouis, MO) were added to stabilize the reaction.Each reaction contained 2�L diluted template. ThecDNA from RT reactions prepared as described isdiluted 1:10 for the gene of interest and 1:100 for�-actin (PE Biosystems). Primers and probes for�-actin were used in a similar manner to quantitatethe presence of�-actin in the samples. In the case ofthe SYBR green assay, the reaction mix (25�L)
Gene Complementation in Breast Cancer • Houghton et al. 81

included 2.5�L SYBR green buffer, 2�L cDNAtemplate, and 2.5�L each forward and reverseprimers for the gene of interest. This mix also con-tained 3 mM MgCl2, 0.25 units AmpErase UNG,0.625 units AmpliTaq gold, 0.08% glycerol, 0.05%gelatin, 0.0001% Tween 20, and 1 mM dNTP mix.In both formats, 40 cycles of amplification wereperformed.
To quantitate the amount of specific cDNA (andhence initial mRNA) in the sample, a standardcurve was generated for each run using a plasmidcontaining the gene of interest. Standard dilutionsranging from 20 to 2 × 106 copies of the gene ofinterest were used for this purpose. In addition, astandard curve is generated for the housekeepinggene�-actin ranging from 200 fg to 2,000 pg toenable normalization to a constant amount of�-ac-tin. This allowed the evaluation of expression levelsseen for each gene.
Quantitative PCR in Peripheral Bloodof Patients With Breast Cancer
Epithelial cells were enriched from bloodsamples with an immunomagnetic bead separationmethod (Dynal AS, Oslo, Norway) using magneticbeads coated with monoclonal antibodies specificfor glycopolypeptide antigens on the surface of hu-man epithelial cells [25–31]. Cells isolated this waywere lysed, and the magnetic beads were removed.The lysate was then processed for poly A+ mRNAisolation using magnetic beads (Dynabeads) coatedwith Oligo (dT)25. After washing the beads in thekit buffer, and eluting polyA+ RNA, samples werefinally suspended in 10 mM Tris-HCl, pH 8, and
subjected to RT followed by real-time PCR usinggene-specific primers and probes with the reactionconditions outlined in Materials and Methods.�-Actin content was also determined and used fornormalization.
Genes of Interest. Genbank accession numbersfor the genomic sequences are listed for each of thegenes. The known genes evaluated in this studywere mammaglobin [9–11] (AF015224) andGABA� [32] (U95367). To identify novel genesthat are overexpressed in breast cancer, we used animproved version of the differential display RT-PCR technique [17,18], cDNA library extractionmethods [19], and PCR subtraction [20,21]. Differ-ential display resulted in the recovery of two cDNAfragments designated B305D (AC018804) locatedon chromosome 2 and B311D (AL157387 match atposition 66000) located on chromosome 10 [33].B511S (AC019223, chromosome 12) and B533S(AL157387 match at position 133949, chromosome10) are two cDNA fragments isolated using thecDNA library subtraction approach, whereas theB726P cDNA fragment was derived from PCR sub-traction (AL357148) and is also located on chro-mosome 10 [34–36] (S.L. Harlocker, R. Jones, J.L.Mitcham, et al., manuscript in preparation).
Results
DNA Microarray AnalysisDNA microarray analysis was used primarily as a
screening tool to determine relative tissue and/ortumor specificity of cDNAs recovered from differ-ential display and cDNA library and PCR subtrac-tions before more rigorous analysis by quantitative
Table 1. Primer and Probe Sequences for the Genes of Interest
Gene Forward Primer Reverse Primer Probe
Mammaglobin TGCCATAGATGAATTGAAGGAATG
TGTCATATATTAATTGCATAAACACCTCA
TCTTAACCAAACGGATGAAACTCTGAGCAATG
B305D C form AAAGCAGATGGTGGTTGAGGTT
CCTGAGACCAAATGGCTTCTTC
ATTCCATGCCGGCTGCTTCTTCTG
B311D CCGCTTCTGACAACACTAGAGATC
CCTATAAAGATGTTATGTACCAAAAATGAAGT
CCCCTCCCTCAGGGTATGGCCC
B726P TCTGGTTTTCTCATTCTTTATTCATTTATT
TGCCAAGGAGCGGATTATCT
CAACCACGTGACAAACACTGGAATTACAGG
B533S CCCTTTCTCACCCACACACTGT
TGCATTCTCTCATATGTGGAAGCT
CCGGGCCTCAGGCATATACTATTCTACTGTCTG
GABA� AAGCCTCAGAGTCCTTCCAGTATG
AAATATAAGTGAAGAAAAAAATTAGTAGAT
AATCCATTGTATCTTAGAACCGAGGGATTTGTTTAGA
B511S GACATTCCAGTTTTACCCAAATGG
TGCAGAAGACTCAAGCTGATTCC
TCTCAGGGACACACTCTACCATTCGGGA
82 Molecular Diagnosis Vol. 6 No. 2 June 2001

RT-PCR, northern blotting, and immunohistochem-istry. The various discovery methods were used be-cause each introduced a different bias in the cDNAfragments that were recovered. The differential dis-play method used (see Materials and Methods) bi-ased toward the recovery of fragments with verylow or no expression in the normal sample, but didnot bias on the mRNA abundance in the tumorsample, allowing the recovery of low-abundancecDNAs. The conventional subtraction method used(see Materials and Methods) biased toward greaterabundance mRNAs and the recovery of largercDNA fragments. To clone less abundant breast tu-mor–specific genes, cDNA library subtraction wasrepeated by subtracting the tracer cDNA librarywith the driver cDNA library plus abundant cDNAsfrom primary subtractions. This resulted in thedepletion of these abundant sequences and the gen-eration of subtraction libraries that contain lessabundant sequences. The PCR-based subtractionmethod (see Materials and Methods) biased towardthe recovery of both abundant and moderatelyabundant cDNAs.
Microarray analysis was performed on two dif-ferent microchips. These two different chip sets in-cluded 3,603 subtracted cDNAs and 197 differentialdisplay templates, which were evaluated to identify40 candidates for further analysis by quantitativePCR. From these candidates, several were chosenon the basis of favorable tissue specificity profiles,including five novel candidates and two knowngenes. Figure 1 shows the microarray profiles for allnovel genes of interest,B305D, B311D, B726P,B511S, andB533S, indicating their overexpressionprofiles in breast tumors and/or normal breast ver-sus other normal tissues. Probe 1 was labeled withCy3, and probe 2, with Cy5. It is evident from thesestudies these genes show relatively increased ex-pression in breast tumors and/or breast tissue. Twoknown breast-specific genes, mammaglobin andGABA�, were also subjected to microarray analysis(Fig. 1) and found to have a relative increase inexpression in breast tumor and normal breast com-pared with other normal tissues. mRNA expressionof mammaglobin was previously described to beupregulated in proliferating breast tissue, includingbreast tumors [9–11]. In previous studies,GABA�showed overexpression in the uterus and, to somedegree, in prostate and lung [32], but no previousstudy indicated elevated levels in breast tumors.
Microarray is an effective method to evaluate
large numbers of genes, but because of its limitedsensitivity, it may not accurately determine the ab-solute tissue distribution of low-abundance genes ormay underestimate the degree of overexpression ofmore abundant genes because of signal saturation.For those genes showing overexpression by micro-array expression profiling, further analysis was per-formed using quantitative RT-PCR based on Taq-man probe detection, which has a greater dynamicrange of sensitivity. Several different panels of nor-mal and tumor tissues, distant metastases, and celllines were used for this purpose.
Interestingly, three of the novel genes,B311D,B533S, and B726P, showed identical breast tissueexpression profiles by quantitative PCR analysis.These genes were evaluated in quantitative PCR ontwo different panels consisting of (1) breast tumor,normal breast, and normal tissues, and (2) a panel ofbreast tumor metastases (primarily lymph nodes).Primers and probes used are listed in Table 1. Over-all, the expression profiles were similar, but levelsof expression were considerably different.B311Din general was expressed at lower levels thanB533S, and both were less thanB726P, but all threewere restricted to breast tissue (data not shown). Allthree sequences were used to search against theGenbank database. BothB311D and B533S se-quences contain different repetitive sequences, andan open reading frame (ORF) has not been identi-fied for either.B726P is a novel gene, with mRNAsplicing yielding several different putative ORFs(S.L. Harlocker, R. Jones, J.L. Mitcham, et al.,manuscript in preparation).
Quantitative PCR confirmed a differentialmRNA expression between normal breast tissueand breast tumors, with approximately 50% ofbreast tumors overexpressing these genes. Testedon a panel of distant metastases derived from 21different breast cancers, all three genes reacted with14 of 21 metastases and presented similar profiles.Interestingly, when tested on a prostate cancerpanel, all three genes identified the same 3 of 24prostate tumors, but at much lower expression lev-els than in breast. This group of genes also showedincreasing levels of expression as a function ofpathological stage of the tumor (data not shown)and is being further evaluated.
Complementary GenesMammaglobin, a homologue of a rabbit uteroglo-
bin and the rat steroid-binding protein subunit C3, is
Gene Complementation in Breast Cancer • Houghton et al. 83

84 Molecular Diagnosis Vol. 6 No. 2 June 2001

a highly glycosylated low-molecular-weight protein[9–11]. In contrast to its homologs, mammaglobinhas been reported to be breast specific, and overex-pression has been described in breast tumor biopsyspecimens (23%) and primary and metastatic breasttumors (∼ 75%), with reports of the detection ofmammaglobin mRNA expression in 91% of lymphnodes from patients with metastatic breast cancer[14,15]. However, more rigorous analysis of mam-maglobin gene expression by microarray (Fig. 1)and quantitative PCR using panels of tumors andnormal tissues indicates expression at significantlevels in skin and salivary gland and much lowerlevels in esophagus and trachea (Table 2).B511S, apreviously unknown gene identified using cDNAlibrary subtraction, has a slightly different profile ofreactivity on breast tumors and normal breast tissuecompared with mammaglobin, yet it reacted with asimilar subset of normal tissues as mammaglobin.B511S by prediction of protein sorting signals andlocalization sites in amino acid sequences analysisis indicated to have an ORF of 90 aa and be asecreted protein, as is the case for mammaglobin.The latter detected 14 of 24 distant metastatic breasttumors and 31 of 42 breast tumors and showed ten-fold overexpression in tumors and metastases com-pared with normal breast tissue. There was at least
300-fold overexpression of mammaglobin in nor-mal breast tissue versus other negative normal tis-sues and tumors tested.B511S detected 33 of 42breast tumors and 14 of 24 distant metastases. Acombination ofB511S with mammaglobin wouldbe predicted to detect 38 of 42 breast tumors and 17of 24 metastatic lesions (Table 2). The quantitativelevels of expression ofB511S and mammaglobinwere also in similar ranges, in concordance with themicroarray profiles observed for these two genes(Fig. 1).
Genes that complemented mammaglobin’s ex-pression profile, i.e., were shown to express in tu-mors that mammaglobin did not, are listed in Table3. B305D was highly overexpressed in breast tu-mors, prostate tumors, normal prostate tissue, andtestis compared with normal tissues, including nor-mal breast tissue. Different splice variants ofB305D have been identified; forms A and C werethe most abundant. All forms tested have similartissue profiles in quantitative PCR [33]. A and Cforms of B305D contain ORFs of 320 and 384amino acids, respectively.B305D C-form is pre-dicted by prediction of protein sorting signals andlocalization sites in amino acid sequences to be atype II membrane protein that contains a series ofankyrin repeats. A known gene shown to be
Table 2. Normalized Distribution of Mammaglobin and B511S mRNA in Various Tissues
TissueMean Copies
Mammaglobin/ng�-Actin*PCRPositive
Mean CopiesB511S/ng �-actin*
PCRPositive
PCR Positive(mammaglobin/B511S)
Breast tumors 1,233.88 ± 3,612.74 31/42 1,800.40 ± 3,893.24 33/42 38/42Breast tumor metastases 1,912.54 ± 4,625.85 14/24 3,329.50 ± 10,820.71 14/24 17/24Benign breast disorders 121.87 ± 78.63 3/3 524.66 ± 609.43 2/3 3/3Normal breast 114.19 ± 94.40 11/11 517.64 ± 376.83 8/9 11/11Breast reduction 231.50 ± 276.68 2/3 482.54 ± 680.28 1/2 2/3Other tumors 0.13 ± 0.65 1/39 24.17 ± 36.00 5/23Salivary gland 435.65 ± 705.11 2/3 45,766.61 ± 44,342.43 3/3Skin 415.74 ± 376.14 7/9 7,039.05 ± 7,774.24 9/9Esophagus 4.45 ± 3.86 2/3 1.02 ± 0.14 0/3Bronchia 0.16 0/1 84.44 ± 53.31 2/2Other normal tissues 0.33 ± 1.07 0/85 5.49 ± 10.65 3/75
Quantitative mRNA levels for mammaglobin andB511S in different tissues.*Mean ± SD.
<
Fig. 1. Microarray characterization of breast genes. Breast cDNA clones were PCR amplified, attached onto glass slides, and probedwith Cy3-labeled probe 1 and Cy5-labeled probe 2. The glass slides were then scanned. Pseudo colors represent the fluorescenceintensity that correlates with hybridization intensity; each successive color represents a twofold change in fluorescence. Microarrayanalysis of mammaglobin (MAM),B511S, B533S, B305D, B311D (breast chip 1),B726P, andGABA� (breast chip 2). *Metastaticbreast tumor.†Human breast tumor grown in SCID mouse.‡Human mammary epithelial cell (HMEC).¶Peripheral-blood mono-nuclear cell.
Gene Complementation in Breast Cancer • Houghton et al. 85

complementary withB305D in breast tumors wasGABA�. This gene is a member of the GABAA
receptor family that encodes a protein that has 30%to 40% amino acid homology with other familymembers and has been shown by northern blotanalysis to be expressed in lung, thymus, and pros-tate at low levels and highly overexpressed in uterus[32]. Its expression in breast tissue has not beenpreviously described. This tissue expression profileis in contrast to other GABAA receptors that typi-cally have appreciable expression in neuronal tis-sues. An additional observation was that tissue ex-pression profiling of this gene showed it wasoverexpressed in breast tumors [36] in an inverserelationship to theB305D gene (Table 3).GABA�detected 15 of 25 tumors and six of 21 metastases,including four tumors and five metastases missedby mammaglobin. Conversely,B305D detected 13of 25 breast tumors and eight of 21 metastases,again including three tumors and two metastasesmissed by mammaglobin. A combination of justB305D andGABA� is predicted to identify 22 of 25breast tumors and 14 of 21 metastases. The combi-nation ofB305D andGABA� with mammaglobin indetecting breast metastases is shown in Table 3 andFig. 2. This combination detected 20 of 21 breastmetastases, as well as 25 of 25 breast tumors evalu-ated on the same panels for all three genes. The onebreast metastasis just less than the cutoff value forthese three genes was positive forB726P (tumorB523A; Fig. 2).
Detection of Circulating BreastCancer Cells in Peripheral Blood
To evaluate the presence of circulating tumorcells, we adopted an immunocapture method to firstenrich for epithelial cells before RT-PCR analysis.Immunomagnetic beads coated with monoclonalantibodies specific to two glycoproteins on the sur-face of epithelial cells were used for this purpose[25]. Compared with direct isolation of poly A+
RNA from blood, such an enrichment procedureincreased the sensitivity of detection (∼ 100-fold),listed in Table 4. MB415 cells were spiked intowhole blood at various concentrations, then RNAwas extracted by using either epithelial cell enrich-ment or direct isolation from blood. MammaglobinmRNA using enrichment procedures was detectableat much lower levels than when direct isolation wasused. Whole-blood samples from patients withmetatastic breast cancer were subsequently treatedwith the immunomagnetic beads, polyA+ RNA wasthen isolated, and cDNA was made and run in quan-titative PCR using two gene-specific primers (Table1) to mammaglobin and a fluorescent probe (Taq-man) [23]. As observed in breast cancer tissues,complementation was also seen in the detection ofcirculating tumor cells derived from breast cancers.Again, mammaglobin PCR detected circulating tu-mor cells in a high percentage of blood samples,albeit at low levels, from metastatic breast cancer(20 of 32 samples) compared with normal bloodsamples (Table 5). Several other genes tested todate could further increase this detection rate, in-
Table 4. Extraction of Mammaglobin-Positive Cells(MB415) Spiked Into Whole Blood and Detection
by Real-Time PCR
MB415 Cells/mLBlood
Epithelial CellExtraction
(Poly A+ RNA)*Direct Extraction(Poly A+ RNA)*
100,000 54,303.2 58,527.110,000 45,761.9 925.91,000 15,421.2 61.6
100 368.0 5.110 282.0 1.11 110.2 00 0 0
MB415 cells were spiked into whole blood (0 to 100,000)cells/mL then extracted using epithelial cell enrichment (Dy-nal) or direct extraction (Roche-Boehringer-Mannheim). Inboth cases, poly A+ RNA was extracted, cDNA was prepared,and real-time PCR (Taqman) was performed using mammaglo-bin-specific primers (Table 1).
*In copies of mammaglobin per nanograms of�-actin.
Table 3. mRNA Complementation of Mammaglobin With Other Genes
MammaglobinPositive
Mammaglobin Negative
B305D GABA� B726PB305D +GABA�
B305D +GABA� + B726P
Breast metastases 13/21 2/8 5/8 3/8 7/8 8/8Breast tumors 18/25 3/7 4/7 5/7 7/7 7/7
86 Molecular Diagnosis Vol. 6 No. 2 June 2001
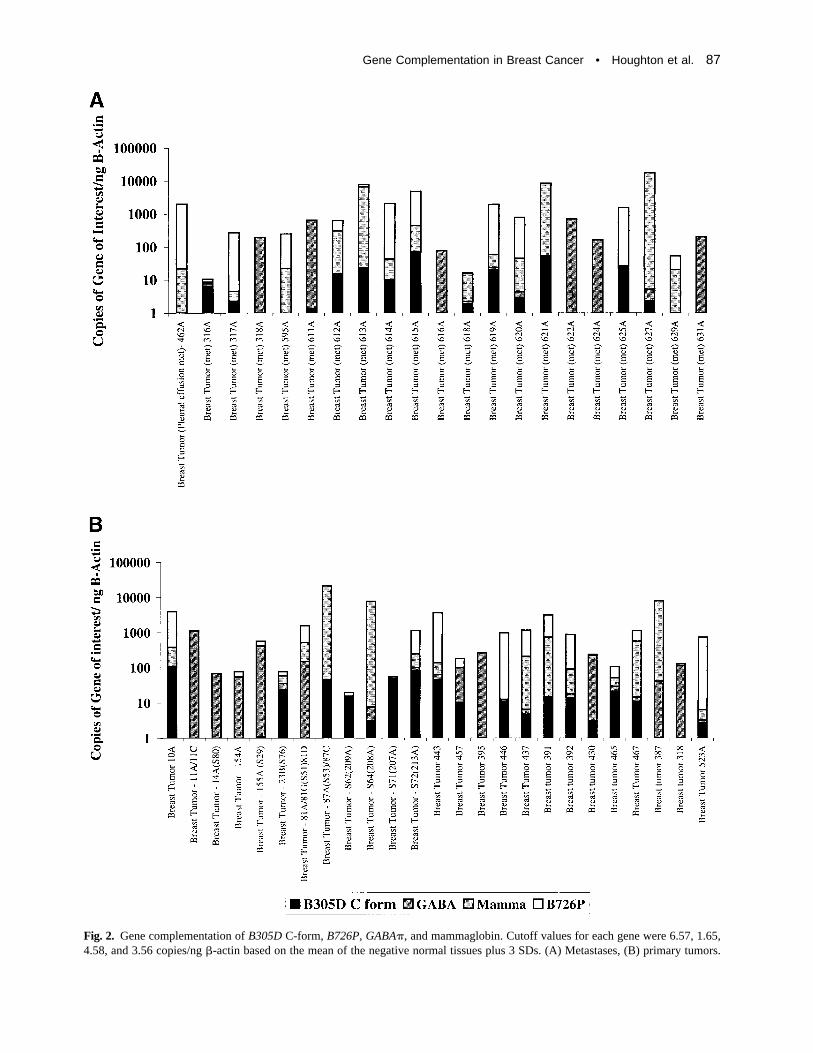
Fig. 2. Gene complementation ofB305D C-form, B726P, GABA�, and mammaglobin. Cutoff values for each gene were 6.57, 1.65,4.58, and 3.56 copies/ng�-actin based on the mean of the negative normal tissues plus 3 SDs. (A) Metastases, (B) primary tumors.
Gene Complementation in Breast Cancer • Houghton et al. 87
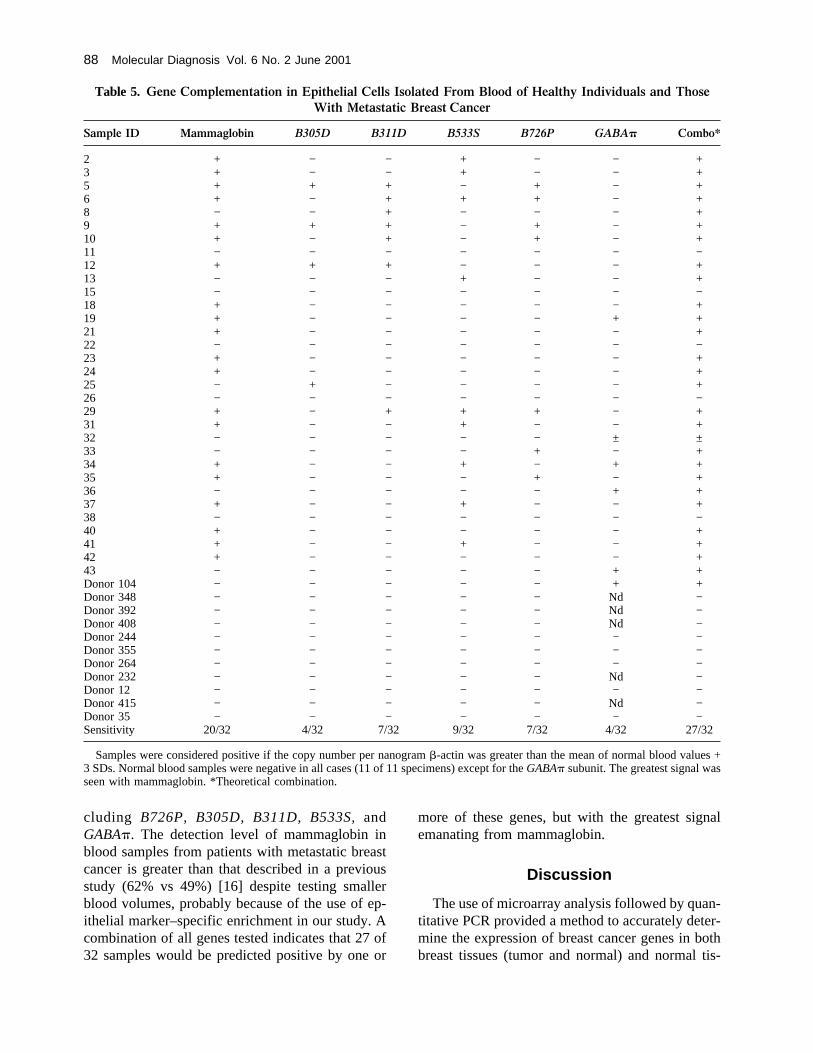
cluding B726P, B305D, B311D, B533S, andGABA�. The detection level of mammaglobin inblood samples from patients with metastatic breastcancer is greater than that described in a previousstudy (62% vs 49%) [16] despite testing smallerblood volumes, probably because of the use of ep-ithelial marker–specific enrichment in our study. Acombination of all genes tested indicates that 27 of32 samples would be predicted positive by one or
more of these genes, but with the greatest signalemanating from mammaglobin.
Discussion
The use of microarray analysis followed by quan-titative PCR provided a method to accurately deter-mine the expression of breast cancer genes in bothbreast tissues (tumor and normal) and normal tis-
Table 5. Gene Complementation in Epithelial Cells Isolated From Blood of Healthy Individuals and ThoseWith Metastatic Breast Cancer
Sample ID Mammaglobin B305D B311D B533S B726P GABA� Combo*
2 + − − + − − +3 + − − + − − +5 + + + − + − +6 + − + + + − +8 − − + − − − +9 + + + − + − +10 + − + − + − +11 − − − − − − −12 + + + − − − +13 − − − + − − +15 − − − − − − −18 + − − − − − +19 + − − − − + +21 + − − − − − +22 − − − − − − −23 + − − − − − +24 + − − − − − +25 − + − − − − +26 − − − − − − −29 + − + + + − +31 + − − + − − +32 − − − − − ± ±33 − − − − + − +34 + − − + − + +35 + − − − + − +36 − − − − − + +37 + − − + − − +38 − − − − − − −40 + − − − − − +41 + − − + − − +42 + − − − − − +43 − − − − − + +Donor 104 − − − − − + +Donor 348 − − − − − Nd −Donor 392 − − − − − Nd −Donor 408 − − − − − Nd −Donor 244 − − − − − − −Donor 355 − − − − − − −Donor 264 − − − − − − −Donor 232 − − − − − Nd −Donor 12 − − − − − − −Donor 415 − − − − − Nd −Donor 35 − − − − − − −Sensitivity 20/32 4/32 7/32 9/32 7/32 4/32 27/32
Samples were considered positive if the copy number per nanogram�-actin was greater than the mean of normal blood values +3 SDs. Normal blood samples were negative in all cases (11 of 11 specimens) except for theGABA� subunit. The greatest signal wasseen with mammaglobin. *Theoretical combination.
88 Molecular Diagnosis Vol. 6 No. 2 June 2001

sues and assess their diagnostic and therapeutic po-tential. Five novel genes and two known genes wereevaluated using these techniques. Three of thesegenes,B311D, B533S, andB726P, showed concor-dant mRNA expression; collectively, data are con-sistent with coordinated expression of these threeloci at the level of transcription control. To date, allefforts to characterize full-length cDNAs for theseclones have indicated that they do not arise from thesame transcriptional unit, although recent genomicmapping studies are consistent with this explana-tion. All three genes showed differential expressionin breast tumors versus normal breast tissue, and thelevel of overexpression appeared related to thepathological stage of the tumor. In the case of mam-maglobin, expression was found in other tissuesapart from breast tissue. Expression was seen inskin, salivary gland, and, to a much lesser degree,trachea. Consistent with these data is the recentfinding that immunohistochemistry using mamma-globin-specific monoclonal antibodies showed re-activity with ductal tissue in skin (G. Fanger, et al.,manuscript in preparation). Of further interest isthat human tumors occasionally arise from epithe-lial cells of apoeccrine and salivary glands [37,38];these tumors are often histologically difficult to dis-tinguish from breast cancers. Thus, mammaglobinexpression may be a molecular remnant of the com-mon embryologic origin of breast tissue and sweatglands.
Expression ofGABA� in breast tumors was alsoa novel observation. Although the expression ofseveral genes complemented that seen with mam-maglobin, two genes in particular,B305D andGABA�, added to the diagnostic sensitivity ofmammaglobin detection (Table 3; Fig. 2). A com-bination of these three genes detected 45 of 46breast tumors and metastases (97.8%) evaluated. In-clusion of B726P enabled the detection of all 25breast tumors and 21 distant metastases (Table 3).Given that mammaglobin,B305D, GABA�, andB726P contain ORFs and may ultimately becomeT-cell vaccine targets, it may become important tomeasure levels of expression of these genes as partof a pretreatment assessment plan. This would besimilar to that currently done forher2/neu expres-sion and Herceptin (Genentech Inc, South San Fran-cisco, CA) therapy. Our ability to detect circulatingcancer cells in patients with known metastatic dis-ease, even in the absence of a detectable mass le-sion, is an example of the potential utility of this
approach in monitoring patients with breast cancer.This approach with further refinements may ulti-mately become a powerful tool in the diagnosis ofbreast cancer and monitoring residual disease.
Acknowledgment
Some tissue samples were provided by the Co-operative Human Tissue Network, which is fundedby the National Cancer Institute, and the NationalDisease Research Interchange.
Received October 31, 2000.Received in revised form December 21, 2000.
Accepted January 4, 2001.
References
1. Donegan WL, Spratt JS: Cancer of the breast, 4th ed.Saunders, Philadelphia, 1995
2. Fendly BM, Winget M, Hudziak RM, Lipari MT,Napier MA, Ullrich A: Characterization of murinemonoclonal antibodies reactive to either the humanepidermal growth factor receptor orHER2/neu geneproduct. Cancer Res 1990;50:1550–1558
3. Baselga J, Norton L, Albanell J, Kim YM, Mendol-sohn J: Recombinant humanized anti-HER2 anti-body (Herceptin) enhances the antitumor activity ofpaclitaxel and doxorubicin against HER2/neu over-expressing human breast cancer xenografts. CancerRes 1998;58:2825–2831
4. Pegram MD, Lipton A, Hayes DF, et al.: Phase IIstudy of receptor-enhanced chemosensitivity usingrecombinant humanized anti-p185HER2/neu mono-clonal antibody plus cisplatin in patients with HER2/neu overexpressing metastatic breast cancer refrac-tory to chemotherapy treatment. J Clin Oncol 1998;16:2659–2671
5. Drebin JA, Link VC, Greene MI: Monoclonal anti-bodies specific for the neu oncogene product di-rectly mediate anti-tumor effects in vivo. Oncogene1988;2:387–394
6. Disis ML, Shiota FM, Cheever MA: Human HER-2/neu protein immunization circumvents toleranceto rat neu: A vaccine strategy for “self ” tumour an-tigens. Immunology 1998;93:192–199
7. Disis ML, Cheever MA: HER-2/neu oncogenic pro-tein: Issues in vaccine development. Crit Rev Im-munol 1998;18:37–45
8. Disis ML, Grabstein KH, Sleath PR, Cheever MA:Generation of immunity to the HER-2/neu oncoge-neic protein in patients with breast and ovarian can-cer using a peptide-based vaccine. Clin Cancer Res1999;5:1289–1297
Gene Complementation in Breast Cancer • Houghton et al. 89

9. Watson MA, Fleming TP: Mammaglobin, a mam-mary-specific member of the uteroglobin gene fam-ily, is overexpressed in human breast cancer. CancerRes 1996;56:860–865
10. Watson MA, Dintzis S, Darrow CM, et al.: Mam-maglobin expression in primary, metastatic, and oc-cult breast cancer. Cancer Res 1999;59:3028–3031
11. Watson MA, Darrow C, Zimonjic DB, Popescu NC,Fleming TP: Structure and transcriptional regulationof the human mammaglobin gene, a breast cancerassociated member of the uteroglobin gene familylocalized to chromosome 11q13. Oncogene 1998;16:817–824
12. Elston CW, Ellis IO: Pathological prognostic factorsin breast cancer. Histopathology 1991;19:403–410
13. Porter-Jordan K, Lippman ME: Overview of the bio-logic markers of breast cancer. Hematol Oncol ClinNorth Am 1994;8:73–100
14. Leygue E, Snell L, Dotzlaw H, et al.: Mammaglobin,a potential marker of breast cancer nodal metastasis.J Pathol 1999;189:28–33
15. Min JC, Tafra L, Verbanac KM: Identification ofsuperior markers for polymerase chain reaction de-tection of breast cancer metastases in sentinel lymphnodes. Cancer Res 1998;58:4581–4584
16. Zach O, Kasparu H, Krieger O, Hehenwarter W,Girschikofsky M, Lutz D: Detection of circulatingmammary carcinoma cells in the peripheral blood ofbreast cancer patients via a nested reverse transcrip-tase polymerase chain reaction assay for mamma-globin mRNA. J Clin Oncol 1999;17:2015–2019
17. Liang P, Pardee AB: Differential display of eukary-otic messenger RNA by means of polymerase chainreaction. Science 1993;257:967–971
18. Mou E, Miller H, Li J, Wand E, Chalifour L: Im-provements to the differential display method forgene analysis. Biochem Biophy Res Commun 1994;199:564–569
19. Hara T, Harada N, Mitsui H, Miura T, Ishizaka T,Miyajima A: Characterization of cell phenotype by anovel cDNA library subtraction system: Expressionof CD8 alpha in a mast cell derived interleukin-4dependent cell line. Blood 1994;84:189–199
20. Diatchenko L, Lau Y-FC, Campbell AP, et al.: Sup-pression subtractive hybridization: A method forgenerating differentially regulated or tissue-specificcDNA probes and libraries. Proc Natl Acad SciU S A 1996;93:6025–6030
21. Yang GP, Ross DT, Kuang WW, Brown PO, WeigelRJ: Combining SSH and cDNA microarrays forrapid identification of differentially expressedgenes. Nucleic Acids Res 1999;27:1517–1523
22. Shalon D, Smith SJ, Brown PO: A DNA microarraysystem for analyzing complex DNA samples using
two-color fluorescent probe hybridization. GenomeRes 1996;6:639–645
23. Holland PM, Abramson RD, Watson R, GelfandDH: Detection of specific polymerase chain reactionproduct by utilizing the 5� to 3� exonuclease activityof Thermus aquaticus DNA polymerase. Proc NatlAcad Sci U S A 1991;88:7276–7280
24. Schneeberger C, Spieser P, Kury F, Zeillinger R:Quantitative detection of reverse transcriptase-PCRproducts by means of a novel and sensitive DNAstain. PCR Methods Appl 1995;4:234–238
25. Momburg F, Moldenhauer G, Hammerling GJ, Mol-ler P: Immunohistochemical study of the expressionof a Mr 34,000 human epithelium-specific surfaceglycoprotein in normal and malignant tissues. Can-cer Res 1987;41:2883–2891
26. Naume B, Borgen E, Beiske K, et al.: Immunomag-netic techniques for the enrichment and detection ofisolated breast carcinoma cells in bone marrow andperipheral blood. J Hemother 1997;6:103–113
27. Naume B, Borgen E, Nesland JM, et al.: Increasedsensitivity for detection of micrometastases in bonemarrow/peripheral blood stem-cell products frombreast-cancer patients by negative immunomagneticseparation. Int J Cancer 1998;78:556–560
28. Martin VM, Siewert C, Scharl A, et al.: Immuno-magnetic enrichment of disseminated epithelial tu-mor cells from peripheral blood by MACS. Exp He-matol 1998;26:252–264
29. Hildebrandt M, Mapara MY, Korner IJ, Bargou RC,Moldenhauer G, Dorken B: Reverse transcriptase-polymerase chain reaction (RT-PCR)-controlled im-munomagnetic purging of breast cancer cells usingthe magnetic cell separation (MACS) system: A sen-sitive method for monitoring purging efficiency.Exp Hematol 1997;25:57–65
30. Eaton MC, Hardingham JE, Kotasek D, Dobrovic A:Immunobead RT-PCR: A sensitive method for thedetection of circulating tumor cells. Biotechniques1997;22:100–105
31. Brandt B, Griwatz C, Heidl S, Assmann G, ZankerKS: Detection of the metastatic potential of blood-borne and immunomagnetically enriched epithelialcells by quantitative erbB-2 RT-PCR. Clin Exp Me-tastases 1996;14:399–408
32. Hedblom E, Kirkness EF: A novel class of GABAA
receptor subunit in tissues of the reproductive sys-tem. J Biol Chem 1997;272:15346–15350
33. Houghton RL, Frudakis A, Schmidt C, et al.: Realtime PCR and microarray analysis of breast cancergenes identified using differential display tech-niques. Cancer Res 1999;40:32–33 (abstr 217)
90 Molecular Diagnosis Vol. 6 No. 2 June 2001

34. Jiang J, Xu J, Houghton RL, Silva S, Reed SG:Discovery of breast cancer antigens using subtractedcDNA libraries and cDNA microarrays. Proc AmAssoc Cancer Res 1999;40:32 (abstr 216)
35. Xu J, Stolk J, Silva S, Houghton RL, Reed SG:Identification of differentially expressed genes inhuman breast tumor using subtraction and microar-ray. Proc Am Assoc Cancer Res 1999;40:319 (abstr2115)
36. Molesh DA, Dillon DC, Schmidt J, et al.: Real timePCR analysis of breast cancer genes identified usingcDNA library subtraction and differential display
methods. Proc Am Assoc Cancer Res 2000;41:681(abstr 4330)
37. Wick MR, Ockner DM, Mills SE, Ritter JH, Swan-son PE: Homologous carcinomas of the breasts, skinand salivary glands. A histologic and histochemicalcomparison of ductal mammary carcinoma, ductalsweat gland carcinoma, and salivary duct carcinoma.Am J Clin Pathol 1998;109:75–84
38. Pfeifer JD, Barr RJ, Wick MR: Ectopic breast tissueand breast-like sweat gland metaplasias: An over-lapping spectrum of lesions. J Cutan Pathol 1999;26:190–196
Gene Complementation in Breast Cancer • Houghton et al. 91