transfer of murine intracisternal a particle phenotype in chloramphenicol-resistant cytoplasts
TRANSCRIPT

Cell, Vol . 9, 3 83-391, November 1976, Copyright O1976 by MIT
Transfer of Murine Intracisternal A ParticlePhenotype in Chloramphenicol-Resistant Cytoplasts
Harry L. Malech and Nelson A . WivelLaboratory of Cell BiologyNational Cancer InstituteBethesda, Maryland 20014
Summary
Murine intracisternal A particles have a number ofproperties which are common to known RNA tumorviruses, but horizontal transmission has not beenpreviously demonstrated. The apparent absence ofinfectivity may be related to the failure of theseparticles to be released from cisternae of endo-plasmic reticulum . Previous biological studiesusing isolated, purified A particles have been com-promised by the fact that the isolation procedurerequires small amounts of nonionic detergent .Using some techniques of somatic cell hybrid-
ization, we have assessed the capacity for A parti-cle genome transfer from positive to negative cells .Since it has been previously shown that somechloramphenicol-resistant cell lines can transferthis resistance in the cytoplasm, we have used thischaracteristic as a marker for cytoplasmic frag-ments. Mouse cells containing A particles weremutagenized, and clones resistant to chloram-phenicol were selected ; by enucleating these cellsand fusing the resultant cytoplasts to each of tworecipient mouse cell lines negative for A particles,it is possible to identify clones of cells known tobe the product of a fusion event between a cyto-plast and a whole cell (cybrids). Under these con-ditions, intracisternal A particles appear in the cy-brid clones as a phenotypic trait that has not beensegregated over at least 60-80 cell generations .
Introduction
Mouse intracisternal A particles (IAP) are virus-likeparticles that bud from the endoplasmic reticuluminto the cisternae within both normal and neoplasticmurine cells (Bernhard, 1960 ; Wivel and Smith,1971). Morphologically similar particles have beenseen in guinea pig leukemic cells (Feldman andGross, 1970 ; Fong and Hsuing, 1976) and in aviancells infected with Rous sarcoma virus (de Giuliet al., 1975 ; Maul and Lewandowski, 1975), butany relationship to murine IAP has not beenestablished .
IAP release from murine cells has not been de-monstrated in either in vivo or in vitro systems (Kuffet al ., 1972), but they can be isolated from cell frac-tions as intact particles that are spherical in shapewith a thin outer shell and thicker hollow inner shell(Malech and Wivel, 1976) . Biochemical analysisshows that they contain a 70S RNA and a specific
reverse transcriptase (Yang and Wivel, 1973, 1974),and on this basis have been included in a classifica-tion of retraviruses (Dalton et al ., 1975) . Despitethese characteristics, horizontal transmission of IAPhas not been demonstrated . Murine cell lines nega-tive for IAP have been cultivated with whole cells,cell fractions, and isolated IAP from IAP-producinglines without successful demonstration of IAP pro-duction in the negative cells (Kuff et al ., 1972 ;Minna, Lueders, and Kuff, (1974) .
Studies of somatic cell hybrids of IAP-positiveand -negative lines show IAP expression to be adominant genetic trait (Minna et al ., 1974 ; Yot-suyanagi and Ephrussi, 1974) . Data based on ex-periments with mouse-hamster somatic cell hybridssuggest that the persistence of IAP and C type parti-cles depends upon the retention of different mousechromosomes (Yotsuyanagi and Ephrussi, 1974) .Although isolated IAP have not been shown to
be capable of infection, it is possible that the prob-lem is one of entry and uncoating within the recipi-ent cell . Since presently available isolation proce-dures involve the use of low concentrations ofTriton X-100 (Kuff et al ., 1972), it is probable thatthe infective capacity of the particles is compro-mised . The absence of IAP activity in cell homoge-nates may be related to the release of lytic enzymesduring homogenization. All the previous studiessuggest that a minimum manipulation of IAP-con-taining cells offers the optimum chance for preserv-ing and demonstrating biological activity . The pres-ence of 70S RNA and reverse transcriptase withinthe mature particle suggests that the capability forinfection might be manifested if the IAP compo-nents at various stages of maturation could be intro-duced directly into a recipient cell . The most directway of introducing the potentially infective materialinvolves fusing the cytoplasmic contents of an IAP-producing cell into an. IAP-negative cell and isolat-ing a resultant clone of cells to search for IAPproduction .
Recently, the necessary techniques for carryingout such experiments have been developed . Cellscan be enucleated with high efficiency to yield via-ble cytoplasmic fragments called cytoplasts, whichcontain all cellular components except the nucleus(Prescott, Myerson, and Wallace 1972 ; Wise andPrescott, 1973) . These cytoplasts can be fused towhole cells to yield a hybridization product in whichthe nucleus and chromosome content is entirely ofthe whole cell parent, but the cytoplasm is a hybridof both parent cells . These cells have been termedcybrids (Poste and Reeve, 1972 ; Bunn, Wallace,and Eisenstadt, 1974 ; Wallace, Bunn, and Eisen-stadt, 1974, 1975) .
Some mutant mammalian cells selected for resis-tance to chloramphenicol (CAP) carry a resistance

Cell384
marker which can be transferred in the cytoplasmby fusion of resistant cytoplasts to sensitive wholecells (Bunn et al ., 1974; Wallace et al ., 1974, 1975) .It has been suggested that the resistance may becoded in the mitochondrial DNA . Using other mu-tant markers along with CAP resistance, it is possi-ble to select clones of cells known to be derivedfrom a fusion event between a cytoplast and awhole cell (Bunn et al ., 1974 ; Wallace et al., 1974,1975) .
In this study, an IAP-positive mouse line was mu-tagenized and clones were selected that were resis-tant to CAP. Those CAP-resistant clones shownto be capable of transmitting CAP resistance in thecytoplasm were used as a source of IAP . By fusingcytoplasts from these clones to IAP-negative murinelines, it was possible to observe production of IAPas a permanent phenotypic trait in some resultantcybrid clones .
Results
Cell Line DerivationAll IAP-positive lines are derived from MT-29240, anuncloned, epithelioid, highly contact-inhibited lineadapted to tissue culture from a transplantablemammary tumor which originally arose spontane-ously in a female Balb/c mouse . The tumor was ob-tained from G . Vlahakis .
MT-29240 and all its derivative clones containlarge numbers of IAP per cell when examined byelectron microscopy . Intracytoplasmic A particles,B type, and C type particles are not seen .
Prior to all mutant selections, subconfluent cul-tures were mutagenized with 0 .5 mM ethyl methane-sulfonate for 18 hr resulting in 40% cell viability,as measured by Trypan blue . After a rinse in saline,the cultures are allowed to recover in new mediafor 24 hr prior to seeding into selective media .MT-0 is a ouabain (OUA)-resistant clone ob-
tained by growing MT-29240 in increasing concen-trations of OUA and then cloning in 6 mM OUA (K+in all media is 6 meq) . As reported for this type ofmutant, the OUA resistance of MT-0 shows co-dominant inheritance in somatic cell hybrids and isa stable trait even after 3 months of culture in non-selective media (Baker et al ., 1974) .MT-OC-1 and MT-OC-10 are OUA- and CAP-
resistant. They are derived from MT-O by growingMT-0 in CAP, 50 µg/ml, in near confluent cultureswith frequent splitting for 2 weeks . The high celldensity seems to protect cells partially from the tox-icity of CAP . Then the cells are seeded into several90 mm petri dishes at 1000 cells per dish in CAP .A few rapidly growing colonies were cloned, surviv-ing dispersion into CAP, 50 µg/ml. MT-OC-1 and
MT-OC-10 were subsequently shown to be capableof transmitting CAP resistance cytoplasmically .
MT-06TG-10 is derived from MT-0 by growing inincreasing concentrations of 6-thioguanine (6TG)and cloning in 6TG, 100 pg/ml . This line is killedin hypoxanthine, aminopterine, thymidine (HAT) se-lective media (Littlefield, 1964), but is OUA- and6TG-resistant .MT-BU-8 is derived from MT-29240 by growing
in increasing concentrations of 5-bromodeoxyuri-dine (BrdUrd) and cloning in BrdUrd, 100 µg/ml .This line is killed in HAT medium .
Two IAP-negative lines were used as recipientsof cytoplast fusions : 3T3-4EF and 116C . The cell line3T3-4EF is a subclone (from Dr . H . G . Coon) derivedfrom clone 3T3-4E (Matsuya and Green, 1969), aline of Swiss mouse origin . The line 3T3-4E is nega-tive for IAP antigens by complement-fixation assay,and has been used as a negative line in a previoussomatic cell hybridization study of IAP antigen ex-pression (Minna et al ., 1974) . Careful electron mi-croscopic examination of 3T3-4EF reveals a rare Ctype viral bud at the plasma membrane, but no IAP .3T3-4EF is BrdUrd-resistant and killed in HAT .
116C is a clone of feral mouse origin previouslyshown to be negative for C type viruses (Hartleyand Rowe, 1975) . This line is negative for IAP anti-gen by diffusion gel analysis, using techniques pre-viously described (Kuff et al ., 1972), and on repeatelectron microscopic examination is negative forany virus-like particles . This line probably lacks Fv-1gene function and thus efficiently replicates N andB tropic variants of MuLV (Hartley and Rowe, 1975) .This line carries no mutant markers and will growin HAT medium .
Cybrid and Hybrid Clone SelectionInitial experiments demonstrating the ability ofMT-OC-1 and MT-OC-10 to transfer CAP resistancecytoplasmically were performed by fusing cyto-plasts of these lines with MT-BU-8 and selecting inmedia containing BrdUrd, 50 µg/ml, and CAP, 50µg/ml. As a control, MT-BU-8 was enucleated andfused to whole cells of itself . Experiments 1-3 ofTable 1 summarize the results of these crosses .
CAP resistance is transferred at an efficiency of18-65 colonies per 10 5 cells plated . The differencesfrom one experiment to another probably representrandom differences in fusion efficiency .Several cybrid colonies were cloned from each
experiment with 80% survival on dispersal into CAPmedia. All clones are CAP- and BrdUrd-resistant,but are killed in OUA, 1 mM, or HAT, thus resem-bling the MT-BU-8 parent except for CAP resis-tance . OUA resistance, which is presumably codedin the MT-OC-1 or MT-OC-10 nucleus, and which

Transfer of Intracisternal A Particle Phenotype3 8 5
Table 1 . Results of Fusion Experiments
Experiment Type of Fusion-
1
enMT-OC-1 x MT-BU-8enMT-BU-8 x MT-BU-8
2 enMT-OC-1 x MT-BU-8enMT-OC-10 x MT-BU-8enMT-BU-8 x MT-BU-8
3 enMT-OC-10 x MT-BU-8enMT-BU-8 x MT-BU-8enMT-OC-10
4
enMT-OC-10 x 3T3-4EFenMT-BU-8 x 3T3-4EF
5
enMT-OC-10 x 3T3-4EFenMT-BU-8 x 3T3-4EF
6
MT-06TG-10 x 3T3-4EF
7
enA-MT-BU-Al x 116C-enMT-BU-8
x 116C
8
A-MT-BU-AI x 116C(1200 : 3 x 105)
usually shows co-dominant inheritance in somaticcell hybrids (Baker et al ., 1974), is not passed intothe cybrids . In experiments not shown, MT-06TG-10was fused to MT-BU-8 and selected in HAT . Al-though OUA resistance was not selected for, all hy-brid clones are resistant to OUA, 3 mM, thus dem-onstrating the dominant mode of inheritance of theCILIA marker .
In experiment 2, there were a few colonies in thecontrol dishes. None of the ten small colonies sur-vived cloning, suggesting that they were only par-tially resistant . There was no growth of colonieswhen cytoplasts alone were plated in BrdUrd-CAPmedia without fusion .The IAP-negative line, 3T3-4EF, was used as the
whole cell recipient of a fusion to cytoplasts fromMT-OC-10. Experiment 4 resulted in 74 CAP-resis-tant cybrid colonies per 105 cells plated (see Table 1) .Experiment 5 resulted in recovery of only 18 CAP-resistant cybrid colonies per 10 5 cells plated . As inexperiments 1-3, considerable variation in the rateof cybrid recovery is seen from experiment to exper-iment. As a control, MT-BU-8 cytoplasts were fusedto 3T3-4EF . No colonies were seen in the controlto experiment 5, but in experiment 4, there werenine colonies or two per 10 5 cells plated . All thesecolonies were small and none survived cloning .However, 90% of cybrid colonies from the experi-
-en means enucleated cytoplasts, with enucleation efficiency of 99 .8% . The ratio of parental cells was 1 :1 .b3-5 x 105 cells total plated in each fusion (includes cytoplasts and whole cells) .=CAP 50 µg/ml, BrdUrd 50 i g/ml .d(n) is the number of clones tested .-The clone, A-MT-BU-Al, is one of the five clones derived from experiment 1 .
mental dishes survived cloning . Seven clones fromexperiment 4 and eleven from experiment 5 weresubjected to further analysis . All these had 3T3-4EFmorphology and growth pattern, were resistant toBrdUrd and CAP, and were killed by OUA, 1 mM,or by HAT .
In experiment 6, 3T3-4EF and MT-06TG-10 werefused without enucleation at a ratio of 1 :1 . Theywere plated in HAT, and nine colonies per 10 5 cellsplated were recovered . Five colonies were clonedin HAT. All the clones consist of large cells that areintermediate in morphology between 3T3-4EF andMT-06TG-10. All the clones are able to grow inOUA, 3 mM, even though the trait had not beenselected for in the cross .
In the last set of experiments, 116C whole cellswere used as the recipient of a CAP-resistant cyto-plast fusion . Since 116C does not contain any mutantmarkers, it was necessary to devise a selective pro-tocol that does not require the derivation of mutantclones of 116C. The cybrid clone A-MT-BU-Al is aCAP- and BrdUrd-resistant line cloned from experi-ment 1 (see Table 1), and is used as an IAP-positivecytoplast donor. This line is killed in HAT, presum-ably because it lacks thymidine kinase . 116C willgrow in HAT because it does not lack any of theenzymes necessary for utilization of exogenouspurines and pyrimidines. Since it is possible to enu-
Resistant Coloniesper 105 Cells Platedb Selective Media=
Sensitivity (s) orResistance (r) toOther Compounds
ClonesPositive for IAPper Number Examined
65 CAP-BrdUrd OUA' 1 mM, Hat, , (5)d 3/30 CAP-BrdUrd
19 CAP-BrdUrd QUA' 1 mM, HAT', (8)25 CAP-BrdUrd OUA' 1 mM, HAT', (8)2 CAP-BrdUrd
18 CAP-BrdUrd QUA' 1 mM, HAT , , (10)0 CAP-B rdUrd0 CAP-BrdUrd
74 CAP-BrdUrd OUA' 1 mM, HATs, (7) 6/72 CAP-BrdUrd
18 CAP-BrdUrd QUA' 1 mM, HAT', (11) 9/110 CAP-B rd U rd
9 HAT OUA' 3 mM, (5) 5/5
27 CAP-HAT 3/70 CAP-HAT
<1 CAP-HAT 1 /1

Cell386
cleate A-MT-BU-AI at 99.8% efficiency, the numberof residual nucleated cells available for fusion with116C is sharply limited . In experiment 7, 116C wasfused to enucleated A-MT-BU-Al and selected inHAT plus CAP. Colonies could grow either if theyare derived from a cytoplast fused to 116C or if theyare derived from a whole cell A-MT-BU-Al fused to116C. However, the ratio of fusions of cyto-plast x 116C/whole cell x 116C should be in therange of 500/1 .Experiment 7 yielded 27 colonies per 10 5 cells
plated. Of nine colonies cloned, seven survived . Allclones exhibit the morphology and growth patternof 116C . No colonies grew in the control fusion be-tween MT-BU-8 cytoplasts and 116C .In experiment 8, 3 x 10 5 116C cells were fused
with 1200 A-MT-BU-AI whole cells in suspension .This is about twice the approximate ratio of A-MT-BU-Al whole cells estimated to be contaminating thecytoplast fusion in experiment 7 . When selected inHAT-CAP media, only two colonies total or fewerthan one colony per 10 5 cells plated were seen .Only one of the two colonies survived cloning . Thisclone has a morphology intermediate between 116Cand A-MT-BU-Al .
IAP Production in Cybrid and Hybrid ClonesCybrid and hybrid clones derived from experiments4-8 were examined by electron microscopy for thepresence of IAP at about 30 cell generations afterfusion and again at about 60 cell generations.Clones negative for IAP remain negative, and posi-tive clones continue to produce IAP at approxi-mately the same level (Table 1) .
In experiments 4 and 5, 6/7 and 9/11 clones arepositive for IAP in low titer. Thus when 3T3-4EF isthe recipient of an IAP-positive cytoplast fusion,about 83% of cybrid clones examined produce IAP .
In experiment 7, when 116C was the recipient ofIAP-positive cytoplast fusions, 3/7 clones were pos-itive for IAP .
It is important to note that the numbers of IAPseen in all the IAP-positive cybrid clones of experi-ments 4, 5, and 7 are low . The highest titer is oneIAP per three cells, and the lowest is one per twentycells in a single thin section . When an IAP is seen,it is always as a single particle with none other ap-parent in that cell section, as shown in Figure 1 ;this is distinctly different from the pattern of produc-tion in the mammary tumor parent clones, in whichevery cell section has 20-40 IAP .On the other hand, the five hybrid clones exam-
ined from experiment 6 (MT-06TG-10 x 3T3-4EF)are all positive for IAP production with every cellin a thin section containing from 10-20 IAP, asshown in Figure 1 . The one hybrid clone from ex-
periment 8 (A-MT-BU-Al x 116C) is similarly positivefor IAP in titers of 10-20 per cell section .
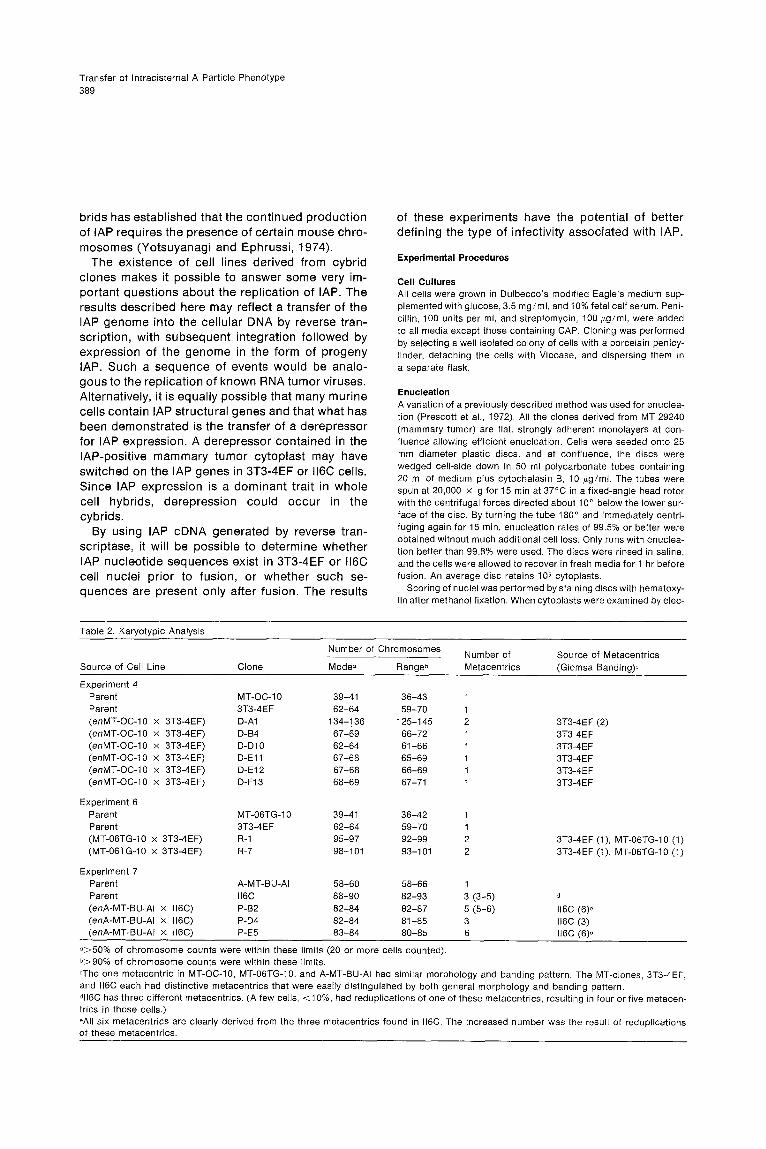
Karyotypic AnalysisOne basic assumption is that nuclear chromosomalgenes from the mammary tumor clones are notpassed into the cybrids . Although this assumptioncannot be proved absolutely, the phenotypic char-acteristics of the cybrids (morphology, growth pat-tern, and drug resistance pattern) suggest that thisis correct . To verify this assumption further, a partialkaryotypic analysis was done . Chromosome num-ber and the Giemsa banding pattern of markermetacentric chromosomes were determined .
All the mammary tumor clones contain one similarmetacentric marker chromosome whose morphol-ogy and banding pattern is distinct from the oneseen in 3T3-4EF and the three seen in 116C (seeFigure 2) .Both MT-OC-10 and MT-06TG-10 have a modal
number of 39-41 chromosomes, while 3T3-4EF hasa mode of 62-64 and a range of 59-70 chromo-somes (Table 2) . Six cybrid clones from experiment4 were examined . All six contain only the metacen-tric marker from the parent 3T3-4EF . The individualclones' modal chromosome numbers all fall withinthe range of the 3T3-4EF parent, except for oneclone. This clone, D-Al, has a chromosome numberabout twice that of 3T3-4EF and contains two identi-cal metacentric chromosomes matching that in3T3-4EF. It is possible that this line is derived froma fusion of a cytoplast with two cells of 3T3-4EF .It is of note that this line has the highest titers ofIAP of all the cybrid clones examined. However, noreadily apparent difference in chromosome numberdistinguishes clones with different titers of IAP .
Two of the five hybrid clones from experiment 6were karyotyped. Both clones, R-1 and R-7, containthe marker metacentric chromosomes of both 3T3-4EF and MT-06TG-10 . The modal chromosomenumber in both clones is only slightly less than thatwhich would be expected by adding together thechromosome complement of both parent lines(Table 2) .
The three cybrid clones positive for IAP in experi-ment 7 were analyzed . None of the three containthe metacentric marker of A-MT-BU-Al . Althoughthe modal chromosome number of each is withinthe range seen for the 116C parent, two of the cybridclones have an increased number of metacentrics .116C has three different metacentric chromosomes .Giemsa banding clearly shows that four of the sixmetacentrics in clone P-B2 are derived from a redu-plication of one of the II6C metacentrics, while theother two chromosomes are the same as the othertwo from 116C . Clone P-D4 contains only three meta-
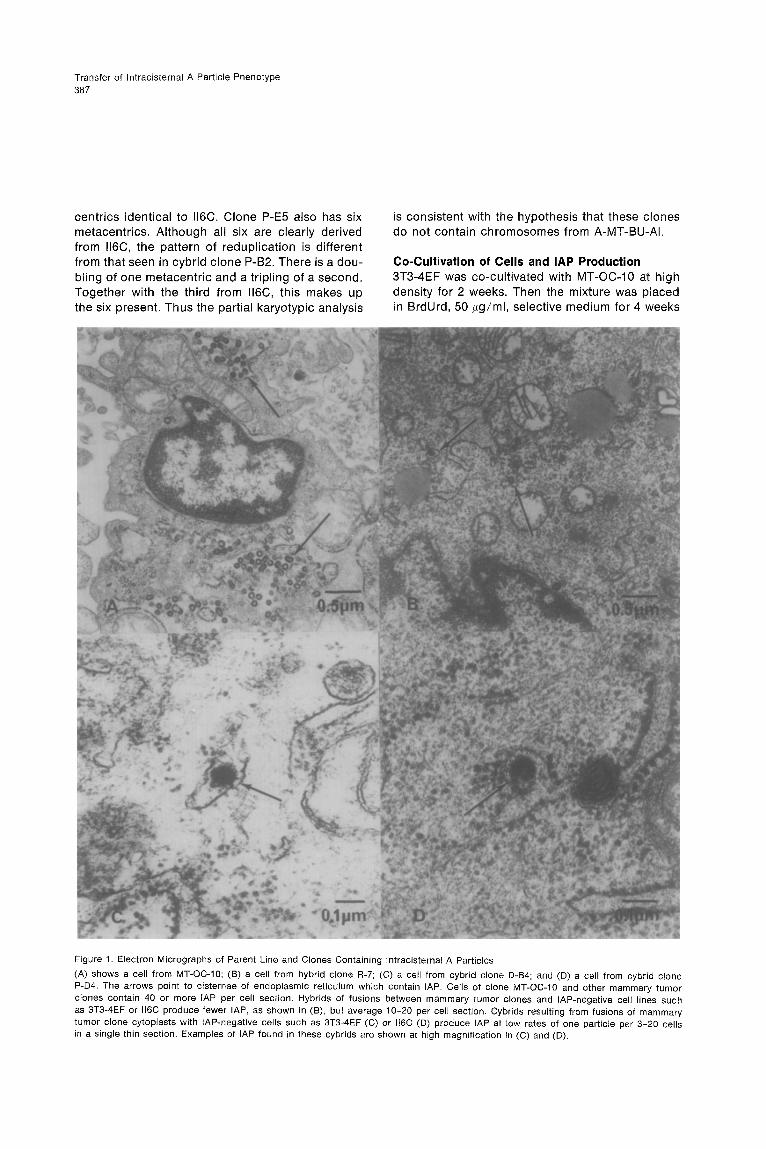
Transfer of lntracisternal A Particle Phenotype 387
centrics identical to 116C. Clone P-E5 also has six metacentrics. Although all six are clearly derived from ll6C, the pattern of reduplication is different from that seen in cybrid clone P-B2. There is a dou- bling of one metacentric and a tripling of a second. Together with the third from ll6C, this makes up the six present. Thus the partial karyotypic analysis
is consistent with the hypothesis that these clones do not contain chromosomes from A-MT-BU-AI.
Co-Cultivation of Cells and IAP Production 3T3-4EF was co-cultivated with MT-OC-10 at high density for 2 weeks. Then the mixture was placed in BrdUrd, 50 pg/ml, selective medium for 4 weeks
Figure 1. Electron Micrographs of Parent Line and Clones Containing lntracisternal A Particles
(A) shows a cell from MT-OC-10; (B) a cell from hybrid clone R-7; (C) a cell from cybrid clone D-B4; and (D) a cell from cybrid clone P-D4. The arrows point to cisternae of endoplasmic reticulum which contain IAP. Cells of clone MT-OC-10 and other mammary tumor clones contain 40 or more IAP per cell section. Hybrids of fusions between mammary tumor clones and IAP-negative cell lines such as 3T3-4EF or ll6C produce fewer IAP, as shown in (B), but average lo-20 per cell section. Cybrids resulting from fusions of mammary tumor clone cytoplasts with IAP-negative cells such as 3T3-4EF (C) or ll6C (0) produce IAP at low rates of one particle per 3-20 cells in a single thin section. Examples of IAP found in these cybrids are shown at high magnification in (C) and (D).

Cdl 388
Figure 2. Giemsa Banding Pattern of the Metacentric Marker Chromosomes
Metacentric chromosomes found in each of the parent cell lines. All the mammary tumor clones used, MT-OC-10, MT-OGTG-IO, and A-MT-BU-AI, contain a metacentric chromosome with the pattern indicated as mammary tumor (MT) clones.
while maintaining the cells in exponential growth. At the end of this selection, aliquots of cells (106) were tested for HAT or OUA, 1 mM, resistance. There was no colony growth in either selective agent. These 3T3-4EF cells were examined in the electron microscope and no IAP were seen.
ll6C were co-cultivated with A-MT-BU-AI for 2 weeks and then selected in HAT for 4 weeks. No IAP were detected in these cells; 3T3-4EF and 116C were each exposed to Sendai virus and 5 days of CAP treatment and allowed to recover for 2 weeks; again no IAP were seen.
Although these results cannot exclude the pres- ence of an extremely low number of IAP, not detect- able in a random survey, they are reminiscent of previously reported experiments in which A particle antigen could not be detected in originally negative cells after a similar co-cultivation protocol (Minna et al., 1974).
Discussion
The development of techniques for the purification of IAP has proved very useful in the study of these particles. There are sufficient data to indicate that
IAP have several characteristics of RNA tumor viruses, but the absence of demonstrable biological activity has precluded the definitive classification of such particles as viruses. The presence of IAP in primary oocytes, their disappearance during matu- ration, and their reappearance in embryos suggest that vertical transmission can take place (Biczysko et al., 1973; Calarco and Szollosi, 1973; Chase and Piko, 1973), but horizontal transmission has not been observed.
The current studies were designed to take advan- tage of the fact that cell fusion provides the most intimate contact between two different cell types, particularly in view of the previous failure of simple co-cultivation experiments to produce detectable horizontal transmission of IAP. In actuality, co-culti- vation by itself could be presumed to fail for at least two reasons, the apparent inability of IAP to be re- leased from a host cell, and the low rate of sponta- neous fusion in the absence of Sendai virus.
For the purposes of the current experiments, it is particularly useful that some CAP-resistant mu- tants have been shown to transfer such resistance in the cytoplasm (Bunn et al., 1974; Wallace et al., 1974, 1975). By developing CAP-resistant, IAP- containing cells, we are thus assured of having “marked” cytoplasts available for the fusion stud- ies. In addition, we have identified another system in which there is some cytoplasmic control of gene function. The cell lines derived from the cybrid clones all resemble the lap-negative cells with re- spectto morphology, growth pattern, drug resistance (except CAP resistance), and chromosome number. In all the 3T3-4EF cybrid clones examined, the number and Giemsa banding pattern of a marker metacentric chromosome are within the range seen in the parent 3T3-4EF. Even in the ll6C cybrids that have an increased number of metacentrics, all of these can be traced by banding pattern to redupli- cation of metacentrics already present in the ll6C parent. Given the high rate of enucleation in mam- mary tumor cells, the use of selective media, and the chromosome studies, it appears improbable that there is any transfer of mammary tumor chro- mosomal material into the cybrids.
At the same time, the low titer pattern of IAP pro- duction in the positive cybrids is distinctly different from the high titer pattern seen in either the mam- mary tumor clones or the whole cell hybrids. This may reflect the fact that there is an absence of a nuclear gene or genes which are required for the replication of IAP. Previous studies have shown a dominant mode of inheritance for A particle antigen when positive mouse cells are fused with negative mouse cells (Minna et al., 1974). Further, the analy- sis of appropriate hamster X mouse somatic cell hy-
Transfer of Intracisternal A Particle Phenotype389
brids has established that the continued productionof IAP requires the presence of certain mouse chro-mosomes (Yotsuyanagi and Ephrussi, 1974) .The existence of cell lines derived from cybrid
clones makes it possible to answer some very im-portant questions about the replication of IAP . Theresults described here may reflect a transfer of theIAP genome into the cellular DNA by reverse tran-scription, with subsequent integration followed byexpression of the genome in the form of progenyIAP . Such a sequence of events would be analo-gous to the replication of known RNA tumor viruses .Alternatively, it is equally possible that many murinecells contain IAP structural genes and that what hasbeen demonstrated is the transfer of a derepressorfor IAP expression . A derepressor contained in theIAP-positive mammary tumor cytoplast may haveswitched on the IAP genes in 3T3-4EF or 116C cells .Since IAP expression is a dominant trait in wholecell hybrids, derepression could occur in thecybrids .By using IAP cDNA generated by reverse tran-
scriptase, it will be possible to determine whetherIAP nucleotide sequences exist in 3T3-4EF or 116Ccell nuclei prior to fusion, or whether such se-quences are present only after fusion . The results
Table 2 . Karyotypic Analysis
of these experiments have the potential of betterdefining the type of infectivity associated with IAP .
Experimental Procedures
Cell CulturesAll cells were grown in Dulbecco's modified Eagle's medium sup-plemented with glucose, 3 .5 mg/ml, and 10% fetal calf serum . Peni-cillin, 100 units per ml, and streptomycin, 100 i g/ml, were addedto all media except those containing CAP . Cloning was performedby selecting a well isolated colony of cells with a porcelain penicy-linder, detaching the cells with Viocase, and dispersing them ina separate flask .
EnucleationA variation of a previously described method was used for enuclea-tion (Prescott et al ., 1972) . All the clones derived from MT 29240(mammary tumor) are flat, strongly adherent monolayers at con-fluence allowing efficient enucleation . Cells were seeded onto 25mm diameter plastic discs, and at confluence, the discs werewedged cell-side down in 50 ml polycarbonate tubes containing20 ml of medium plus cytochalasin B, 10 pg/ml . The tubes werespun at 20,000 x g for 15 min at 37°C in a fixed-angle head rotorwith the centrifugal forces directed about 10° below the lower sur-face of the disc . By turning the tube 180° and immediately centri-fuging again for 15 min, enucleation rates of 99 .5% or better wereobtained without much additional cell loss . Only runs with enuclea-tion better than 99 .8% were used . The discs were rinsed in saline,and the cells were allowed to recover in fresh media for 1 hr beforefusion . An average disc retains 105 cytoplasts .
Scoring of nuclei was performed by staining discs with hematoxy-lin after methanol fixation . When cytoplasts were examined by elec-
°>50% of chromosome counts were within these limits (20 or more cells counted) .b>90% of chromosome counts were within these limits .=The one metacentric in MT-OC-10, MT-06TG-10, and A-MT-BU-Al had similar morphology and banding pattern . The MT-clones, 3T3-4EF,and 116C each had distinctive metacentrics that were easily distinguished by both general morphology and banding pattern .dll6C has three different metacentrics . (A few cells, <10%, had reduplications of one of these metacentrics, resulting in four or five metacen-trics in those cells .)-All six metacentrics are clearly derived from the three metacentrics found in 116C . The increased number was the result of reduplicationsof these metacentrics .
Source of Cell Line Clone
Number of Chromosomes Number ofMetacentrics
Source of Metacentrics(Giemsa Banding)°Mode- Rangeb
Experiment 4Parent MT-OC-1 0 39-41 36-43 1Parent 3T3-4EF 62-64 59-70 1(enMT-OC-10 x 3T3-4EF) D-A1 134-136 125-145 2 3T3-4EF (2)(enMT-OC-10 x 3T3-4EF) D-B4 67-69 66-72 1 3T3-4EF(enMT-OC-10 x 3T3-4EF) D-D10 62-64 61-66 1 3T3-4EF(enMT-OC-10 x 3T3-4EF) D-E11 67-68 65-69 1 3T3-4 E F(enMT-OC-10 x 3T3-4EF) D-E12 67-68 66-69 1 3T3-4EF(enMT-OC-10 x 3T3-4EF) D-F13 68-69 67-71 1 3T3-4EF
Experiment 6Parent MT-06TG-10 39-41 36-42 1Parent 3T3-4EF 62-64 59-70 1(MT-06TG-10 x 3T3-4EF) R-1 95-97 92-99 2 3T3-4EF (1), MT-06TG-10 (1)(MT-06TG-10 x 3T3-4EF) R-7 98-101 93-101 2 3T3-4EF (1), MT-06TG-10 (1)
Experiment 7Parent A-MT-BU-Al 58-60 58-66 1Parent 116C 88-90 82-93 3 (3-5) d(enA-MT-BU-Al x 116C) P-B2 82-84 82-87 5 (5-6) 116C (6)e(enA-MT-BU-Al x 116C) P-D4 82-84 81-85 3 116C (3)(enA-MT-BU-Al x 116C) P-E5 83-84 80-85 6 116C (6)e

Cell390
tron microscopy, all usual cell organelles were present plus largenumbers of IAP, but no nuclei were seen .
FusionSendai virus (/3-propiolactone-inactivated), provided by Dr . H . G .Coon, was used to induce cell fusion . Fusion of cytoplasts withwhole cells was performed without removing cytoplasts from thediscs . Sendai virus (0 .2 ml at 4000 HAU/ml) was added to thediscs in a dry petri dish on ice . After 10 min, 0 .2 ml of a suspensionof whole cells (5 x 105 cells per ml) were added and incubatedfor another 10 min . The discs were transferred to a 37'C humidifiedincubator . At 1 hr, 5 ml of medium were added, and at 3 hr, thediscs were rinsed in saline and placed in CAP, 50 jig/ml, media .Fusions of whole cells not requiring enucleation were performedin suspension (Bunn et al ., 1974) .
Selection of CybridsAt 24 hr after fusion, the cytoplast-whole cell fusion products wereremoved from the discs with Viocase . The contents of three discs,averaging about 5 x 105 cells (including surviving cytoplasts),were plated in 90 mm petri dishes in CAP media at 5000 cellsper dish . This low plating density is essential to assure a cleanselection, since at higher plating densities, patches of confluentcells occur in the plates before CAP can exert its lethal effects .The expression of CAP sensitivity appears to require 6-8 cell divi-sions in exponential growth . After that time, growth ceases andsensitive cells are slowly eliminated.At 5 days after plating, a selective agent to which the recipient
whole cell is resistant and to which the cytoplast donor line issensitive is added to the CAP media . This results in growth onlyof clones resulting from a cytoplast-whole cell fusion . The rationalefor this selective procedure has been previously reported (Bunnet al ., 1974 ; Wallace et al ., 1974, 1975) .Many small colonies are seen at 10 days after plating in both
the control (cytoplasts from CAP-sensitive line) and experimental(CAP-resistant cytoplasts) . By 15 days, most of the colonies donot have mitotic cells and are beginning to thin out . By 20 days,a few large viable growing colonies are seen only in those cultureswhere CAP-resistant cytoplasts were used . These colonies can beclearly distinguished from the smaller thinning and dying ones .These large growing colonies show 80% survival when cloned intoCAP media. The thinning colonies invariably do not survive at-tempts to clone .
KaryotypingRapidly growing cells were arrested in metaphase with 0 .1 µg/mlof colchicine, After swelling in hypotonic buffer for 12 min, theywere fixed overnight in 3 :1 methanol :acetic acid and spread onglass slides. After drying, they were prepared for Giemsa bandinganalysis (Sumner, Evans, and Buckland, 1973) .
Electron MicroscopyThe techniques for electron microscopy have been described pre-viously (Wivel and Smith, 1971) . Thin sections were examined ina Hitachi HU 11E electron microscope . At least 200 cells from dif-ferent portions of the pellet were examined in every cell line whensearching for IAP . In those lines negative for IAP, two observerseach examined 300 cells .
Received July 2, 1976 ; revised August 5, 1976
References
Baker, R . M ., Brunette, D . M ., Mankovitz, R ., Thompson, L . H .,Whitmore, G . F., Siminovitch, L., and Till, J . E . (1974) . Ouabain-Re-sistant mutants of mouse and hamster cells in culture . Cell 1, 9-21 .Bernhard, W. (1960) . The detection and study of tumor viruseswith the electron microscope . Cancer Res . 20, 712-727 .
Biczysko, W., Pienkowski, M ., Solter, D ., and Koprowski, H . (1973) .Virus particles in early mouse embryos . J . Nat . Cancer Inst . 51,1041-1050 .Bunn, C . L., Wallace, D . C ., and Eisenstadt, J . M . (1974) . Cytoplas-mic inheritance of chloramphenicol resistance in mouse tissue cul-ture cells. Proc . Nat . Acad . Sci . USA 71, 1681-1685 .Calarco, P ., and Szollosi, D . (1973) . Intracisternal A particles inova and preimplantation stages of the mouse . Nature New Biol .243, 91-93 .Chase, D ., and Piko, L. (1973) . Expression of A- and C- type parti-cles in early mouse embryos . J . Nat . Cancer Inst . 51, 1971-1975 .Dalton, A . J ., Melnick, J . L ., Bauer, H ., Beaudreau, G ., Bentvelzen,P ., Bolognesi, D ., Gallo, R ., Graffi, A ., Haguenau, F ., Huebner, R .,Todaro, G ., and Heine, U . (1975) . The case for a family of reversetranscriptase viruses : retraviridae . Intervirology 4, 201-206 .de Giuli, C ., Hanafusa, H ., Kawai, L ., Dales, L ., Chen, J ., and Hsu,K . (1975) . Relationship between A-type and C-type particles in cellsinfected by Rous sarcoma virus . Proc . Nat . Acad . Sci . USA 72,3706-3710 .
Feldman, D ., and Gross, L. (1970) . Electron microscopic study ofguinea pig leukemia virus . Cancer Res . 30, 2702-2711 .Fong, C . K . Y ., and Hsuing, G . D . (1976) . Oncornavirus of guineapigs : I . Morphology and distribution in normal and leukemic guineapig cells. Virology 70, 385-398 .Hartley, J . W., and Rowe, W . P . (1975) . Clonal cell lines from aferal mouse embryo which lack host-range restrictions for murineleukemia viruses . Virology 65, 128-134 .Kuff, E . L ., Lueders, K . K ., Ozer, H . L ., and Wivel, N . A . (1972) .Some structural and antigenic properties of intracisternal A parti-cles occurring in mouse tumors . Proc . Nat . Acad . Sci . USA 69,218-222 .Littlefield, J . W. (1964) . Selection of hybrids from matings of fibro-blasts in vitro and their presumed recombinants . Science 145,709-710 .Malech, H . L., and Wivel, N . A . (1976) . Properties of murine intracis-ternal A particles: electron microscopic appearance after criticalpoint drying and platinum shadowing . Virology 69, 802-809 .Matsuya, Y ., and Green, H . (1969) . Somatic cell hybrid betweenthe established human line D 98 (presumptive HeLa) and 3T3 .Science 163, 697-698 .
Maul, G ., and Lewandowski, L. (1975) . Detection of type A oncorna-virus-like structures in chicken embryo cells after infection withRous sarcoma virus . J . Virol . 16, 1071-1074 .Minna, J . D ., Lueders, K . K ., and Kuff, E . L . (1974) . Expressionof genes for intracisternal A-particle antigen in somatic cell hybrids .J . Nat . Cancer Inst . 52, 1211-1217 .Poste, G ., and Reeve, P . (1972) . Enucleation of mammalian cellsby cytochalasin B : II, Formation of hybrid cells and heterokaryonsby fusion of anucleate and nucleated cells . Exp . Cell Res . 73, 287-294 .Prescott, D . M ., Myerson, D ., and Wallace, J . (1972) . Enucleationof mammalian cells with cytochalasin B . Exp . Cell Res . 71, 480-485 .Sumner, A . T., Evans, H . J ., and Buckland, R . A . (1973) . Mechanisminvolved in the banding of chromosomes with Quinacrine and Giem-sa . Exp . Cell Res. 81, 214-222 .Wallace, D . C ., Bunn, C . L ., and Eisenstadt, J . M . (1974) . Cytoplas-mic transfer of chloramphenicol resistance between human cells .Fed . Proc . 33, 1520 .Wallace, D . C ., Bunn, C . L ., and Eisenstadt, J . M . (1975) . Cytoplas-mic transfer of chloramphenicol resistance in human tissue culturecells. J . Cell Biol . 67, 174-188 .Wise, G . E ., and Prescott, D . M . (1973) . Ultrastructure of enucleat-ed mammalian cells in culture . Exp. Cell Res . 81, 65-72 .

Transfer of Intracisternal A Particle Phenotype3 9 1
Wivel, N . A., and Smith, G . H . (1971) . Distribution of intracisternalA-particles in a variety of normal and neoplastic mouse tissues .Int . J . Cancer 7, 167-175.Yang, S . S ., and Wivel, N . A . (1973) . Analysis of high-molecular-weight ribonucleic acid associated with intracisternal A particles .J . Virol . 11, 287-298 .
Yang, S . S ., and Wivel, N . A. (1974) . Characterization of an endog-enous RNA-dependent DNA polymerase associated with murineintracisternal A particles . J . Virol . 13, 712-720 .Yotsuyanagi, Y ., and Ephrussi, B . (1974) . Behavior of three typesof ribovirus-like particles in segregating hamster x mouse somatichybrids. Proc . Nat . Acad . Sci . USA 71, 4575-4578 .