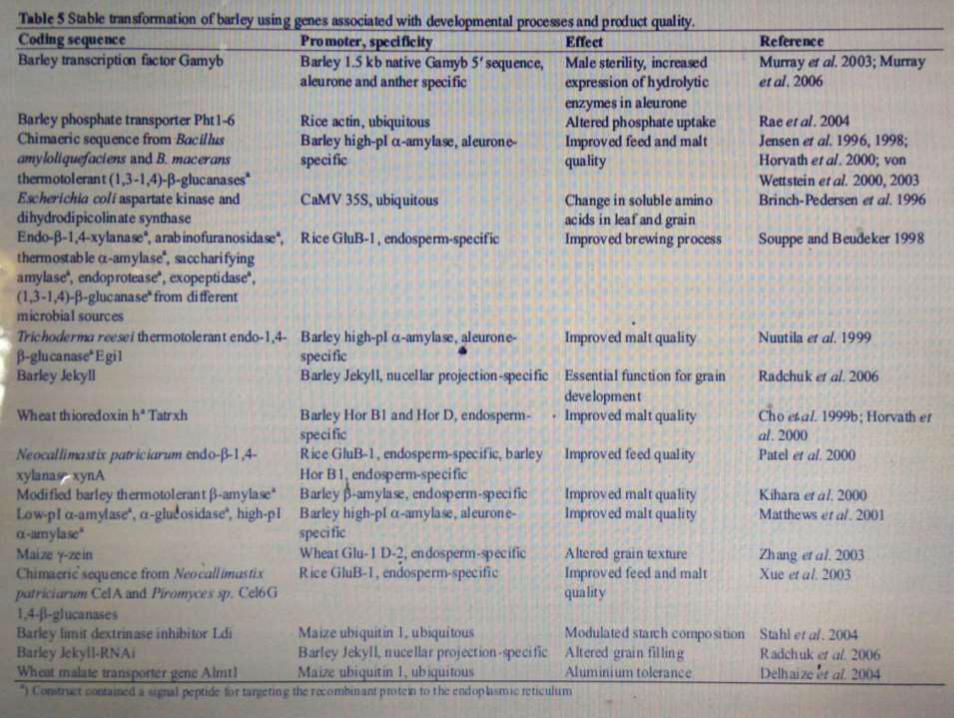
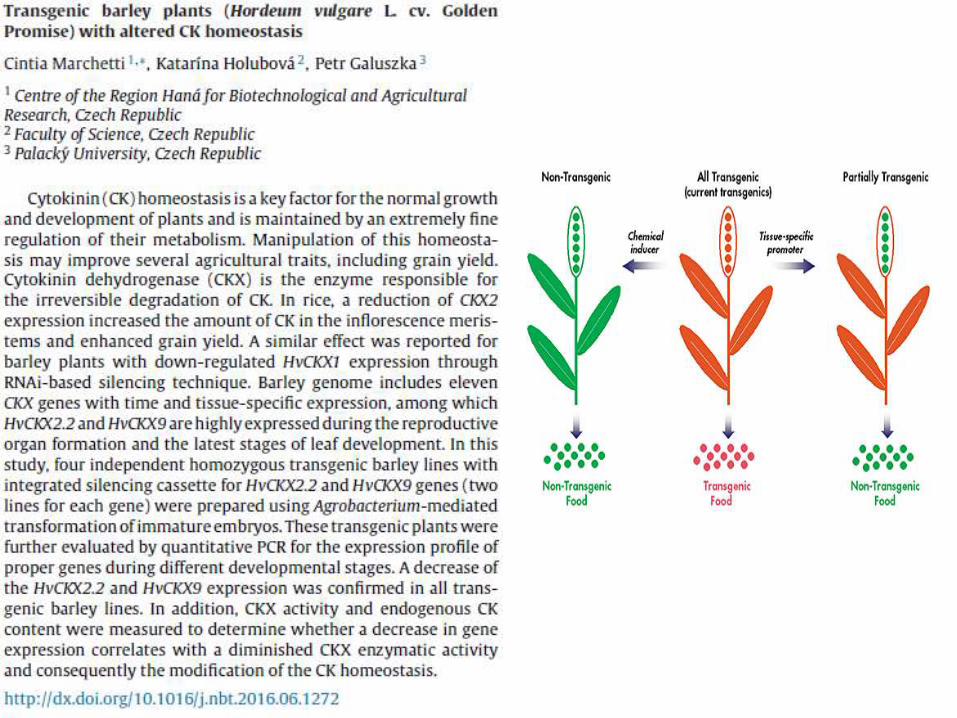
transgenic barley
TRANSCRIPT

SUBMITTED BY:ABHISHEK DAS
ADM. NO.- 09ABT/15

SCIENTIFIC CLASSIFICATION:
Kingdom Plantae
Angiosperms
Monocots
Commelinids
Order Poales
Family Poaceae
Subfamily Pooideae
Tribe Triticeae
Genus Hordeum
Species H. vulgare
BINOMIAL NAME:
Hordeum vulgare L.

TRANSGENIC BARLEY
One of the founder crops of agriculture.4th most important cereal grain worldwide.Used as malt in brewing & distilling industry, as an additive for
animal feed, and as a component of various food and bread for human consumption.
Progress in stable genetic transformation of it ensures a potential for improvement of its agronomic performance or use of barley in various biotechnological and industrial applications.
Used in molecular farming as a promising bioreactor adapted for production of human therapeutic proteins or animal vaccines.
Genetic resources of barley and its tools such as sequence data, microarrays, genetic maps & databases have been regenerated.

CURRENT STATUS ON BARLEY TRANSFORMATION TECHNOLOGIES
1) Gene transfer techniques.2) Targets 3) Progeny stabilization4) Recent trials for improvement of agricultural traits &
performance of barley5) Especially in relation to increased biotic and abiotic stress
tolerance.6) Potential use of barley grain as a protein production platform.
Barley represents a promising tool for both agricultural &
biotechnological transgenic approaches, and is considered an
ancient but rediscovered crop as a model industrial platform
for molecular farming.

Varieties of barley which are hardly amenable for genetic transformation due to low tissue culture response were stably transformed by optimizing transformation parameters for each genotype separately according to their tissue culture response. Scutelia of immature zygotic embryos were transformed with the plasmid pAHC25 carrying the selectable marker gene bar and the reporter gene uidA. Gene delivery was optimized using the uidA-expression as transient marker for the transformation efficiency and taking into account the maintenance of the regeneration capacity after the bombardment.

The grain of the self-pollinating diploid barley species offers two modes of producing recombinant enzymes or other proteins. One uses the promoters of genes with aleurone-specific expression during germination and the signal peptide code for export of theprotein into the endosperm. The other uses promoters of the structural genes for storage proteins deposited in the developing endosperm. Production of a protein-engineered thermotolerant (1,3–1, 4)-b-glucanase with the D hordein gene (Hor3–1) promoterduring endosperm development was analyzed in transgenic plants with four different constructs.
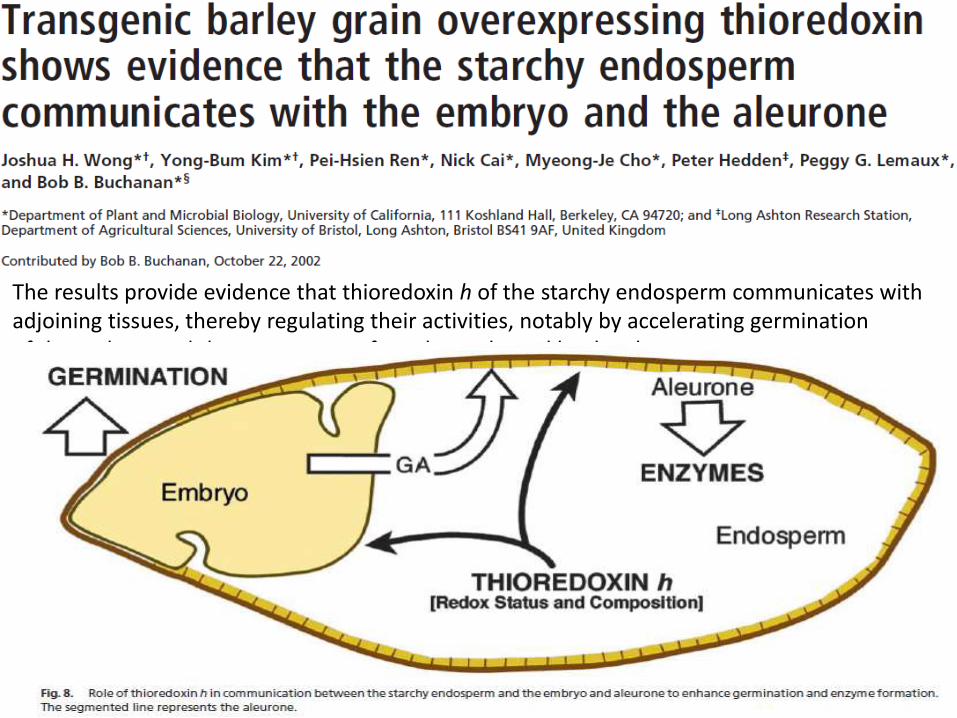
The results provide evidence that thioredoxin h of the starchy endosperm communicates with adjoining tissues, thereby regulating their activities, notably by accelerating germinationof the embryo and the appearance of-amylase released by the aleurone.

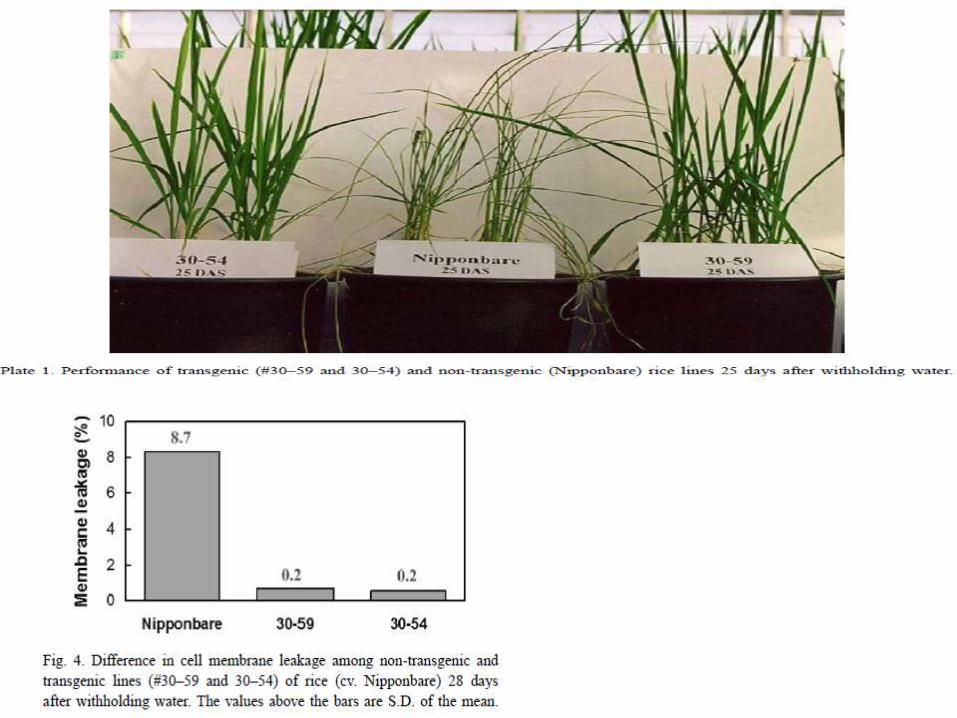
Understanding the mechanism of stress tolerance in these transgenic plants under agronomically realistic stress conditions would further hasten breeding for drought resistance in rice. In this study, transgenic rice lines expressing the barley HVA1 gene were tested under prolonged drought stress cycle to understand the mechanism of dehydration tolerance. Transgenic plants maintained higher leaf relative water content (RWC) and showed lesser reduction in plant growth under drought stress as compared to non-transgenic (NT) plants.production of HVA1 proteins might have helped in better performance of transgenic rice plants by protecting cell membrane from injury under drought stress.

HvGAMYB is a transcription factor protein, which is part of the gibberellin (GA) signal transduction pathway in barley aleurone cells. It activates GA-regulated genes to cause synthesis of alpha-amylase and other hydrolytic enzymes. In aleurone cells, HvGAMYB is up-regulated by GA prior to hydrolytic enzyme gene expression.

Stable genetic transformation represents the gold standard approach to the detailed elucidation of plant gene functions.The analysis of representative numbers of transgenic lines revealed that the obtained T-DNA copy numbers are typically low, the generative transmission of the recombinant DNA is in accordance with the Mendelian rules and the vast majority of the primary transgenics produce progeny that expresses the respective transgene product.
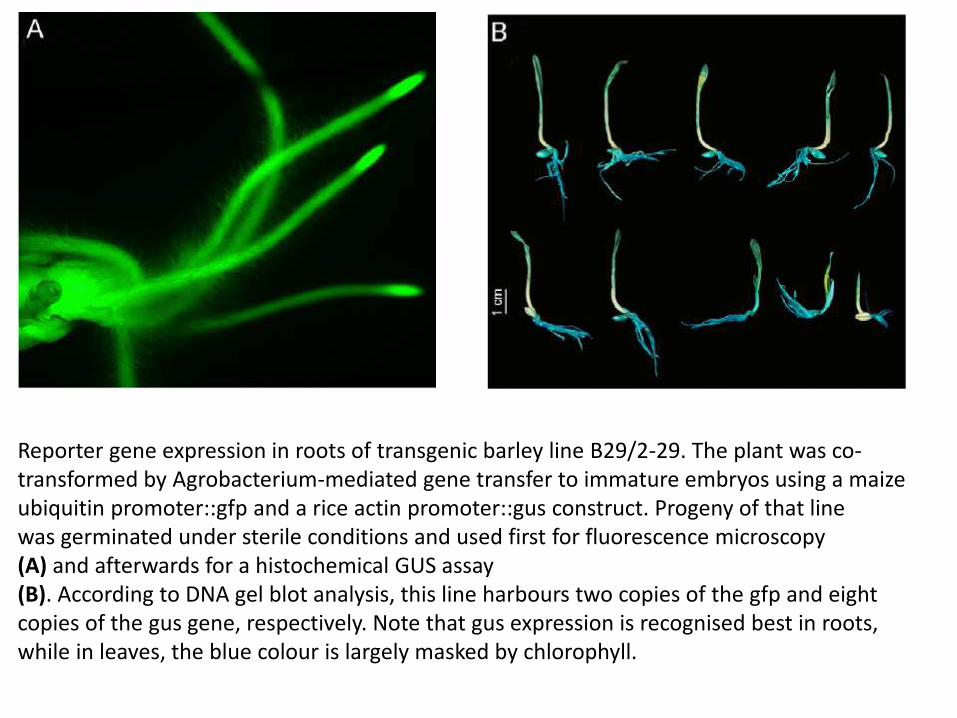
Reporter gene expression in roots of transgenic barley line B29/2-29. The plant was co-transformed by Agrobacterium-mediated gene transfer to immature embryos using a maize ubiquitin promoter::gfp and a rice actin promoter::gus construct. Progeny of that linewas germinated under sterile conditions and used first for fluorescence microscopy (A) and afterwards for a histochemical GUS assay (B). According to DNA gel blot analysis, this line harbours two copies of the gfp and eight copies of the gus gene, respectively. Note that gus expression is recognised best in roots, while in leaves, the blue colour is largely masked by chlorophyll.

H. vulgare genome is available already for couple of years;however its functional annotation is still incomplete.Here we present whole transcriptome response, using extended annotation, to severe drought stress and subsequent re-watering in wild-type barley plants in stemelongation phase of growth. Up- and down-regulated genes fall into distinct GO (gene ontology) categories and these enriched by stress and revitalization are highlighted. Transcriptomic data were evaluated separately for root and aerial tissues.

Rpg1 is a valuable resistance gene that has protected barley from serious yield losses due to stem rust (Puccinia graminis f. sp. tritici [Pgt])Stem rust susceptibility and failure to respond to avirulence factors by the high-copy Rpg1 transgenic lines G04-287 and G04-288 are probably due to the failure to degrade the RPG1 protein.
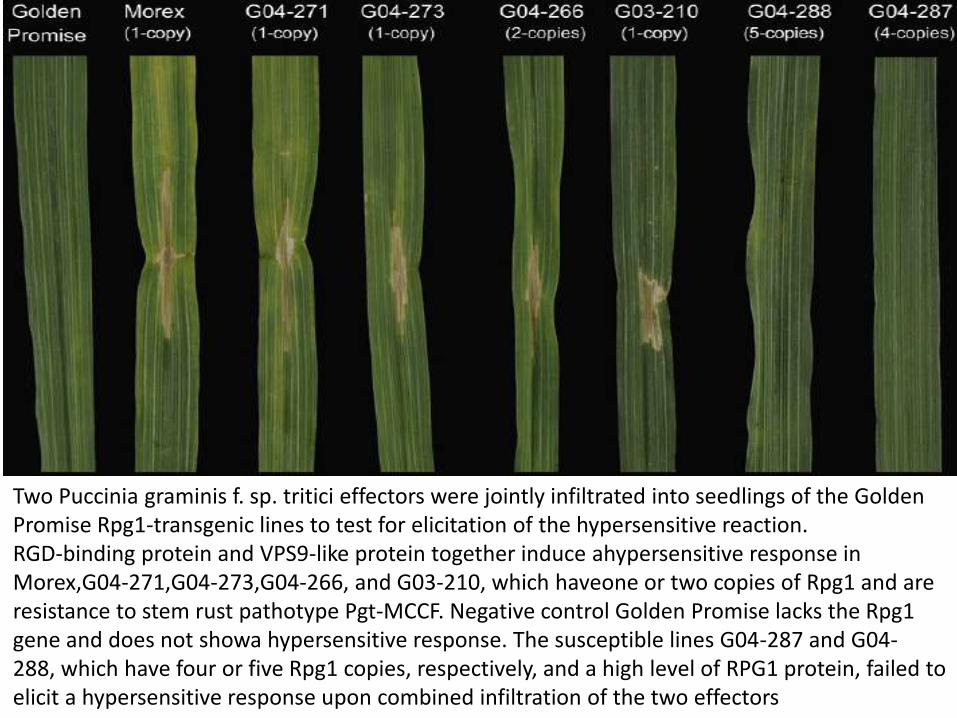
Two Puccinia graminis f. sp. tritici effectors were jointly infiltrated into seedlings of the Golden Promise Rpg1-transgenic lines to test for elicitation of the hypersensitive reaction.RGD-binding protein and VPS9-like protein together induce ahypersensitive response in Morex,G04-271,G04-273,G04-266, and G03-210, which haveone or two copies of Rpg1 and areresistance to stem rust pathotype Pgt-MCCF. Negative control Golden Promise lacks the Rpg1 gene and does not showa hypersensitive response. The susceptible lines G04-287 and G04-288, which have four or five Rpg1 copies, respectively, and a high level of RPG1 protein, failed to elicit a hypersensitive response upon combined infiltration of the two effectors

For many gene function studies, barley can be used as a model for wheat due to its highly efficient transformation rates and smaller, less complex genome. An ideal transformation system needs to be extremely efficient, simple to perform, inexpensive, genotype-independent, and give the required expression of the transgene.

Regeneration from immature embryos of barley cultivar Golden Promise.
Transformation of (A) Optic, (B) Oxbridge, (C) Tipple, and(D) Golden Promise showing the development of transformedcallus on plates containing hygromycin as the selective agent.(E, F) Callus development without transformation: (E) GoldenPromise, (F) Maythorpe.


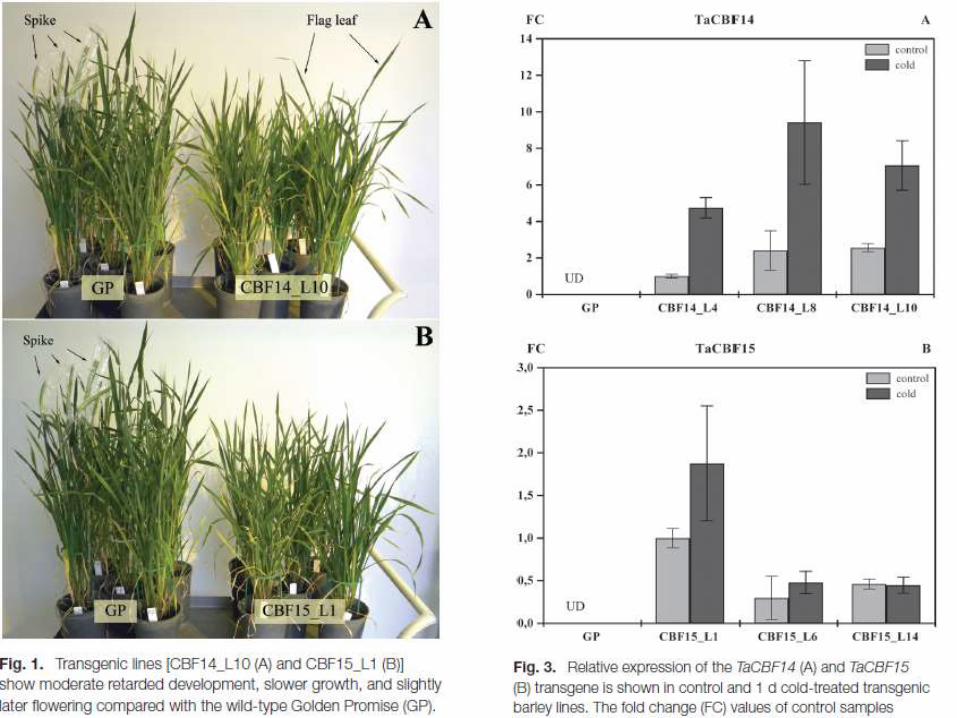
The aim of the present work was the functional analysis of two wheat CBF transcription factors, namely TaCBF14 and TaCBF15, shown by previous experiments to play a role in the development of frost tolerance. These genes were isolated from winter wheat and then transformed into spring barley, after which the effect of the transgenes on low temperature stress tolerance was examined.Gene expression studies showed that HvCOR14b, HvDHN5, and HvDHN8 genes were up-regulated by TaCBF14 and TaCBF15. Beyond that, transgenic lines exhibited moderate retarded development, slower growth, and minor late flowering compared with the wild type, with enhanced transcript level of the gibberellin catabolic HvGA2ox5 gene.

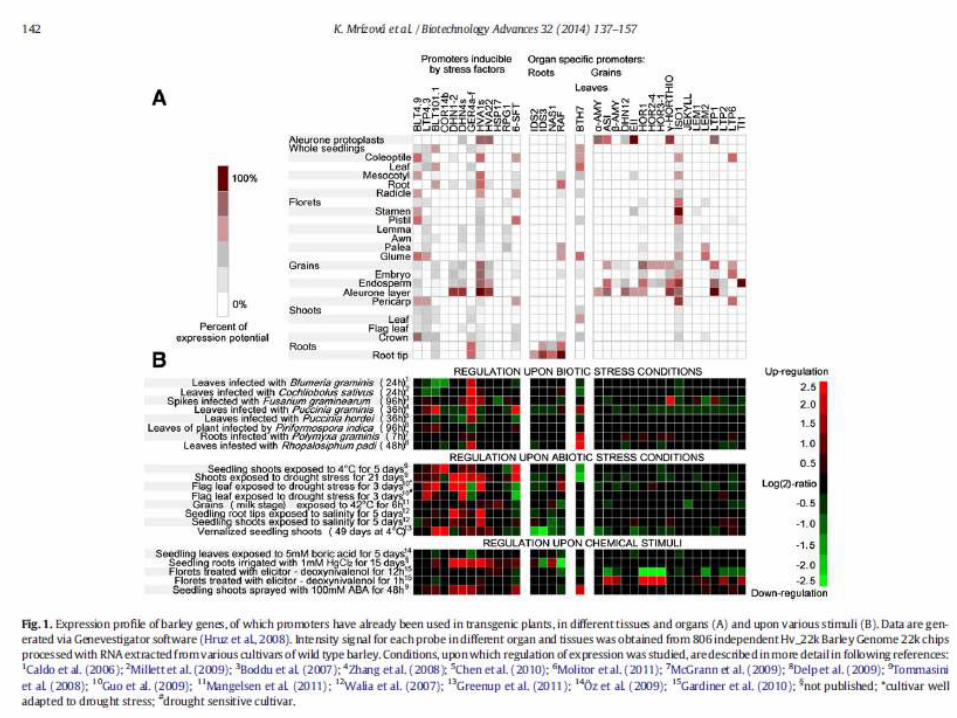

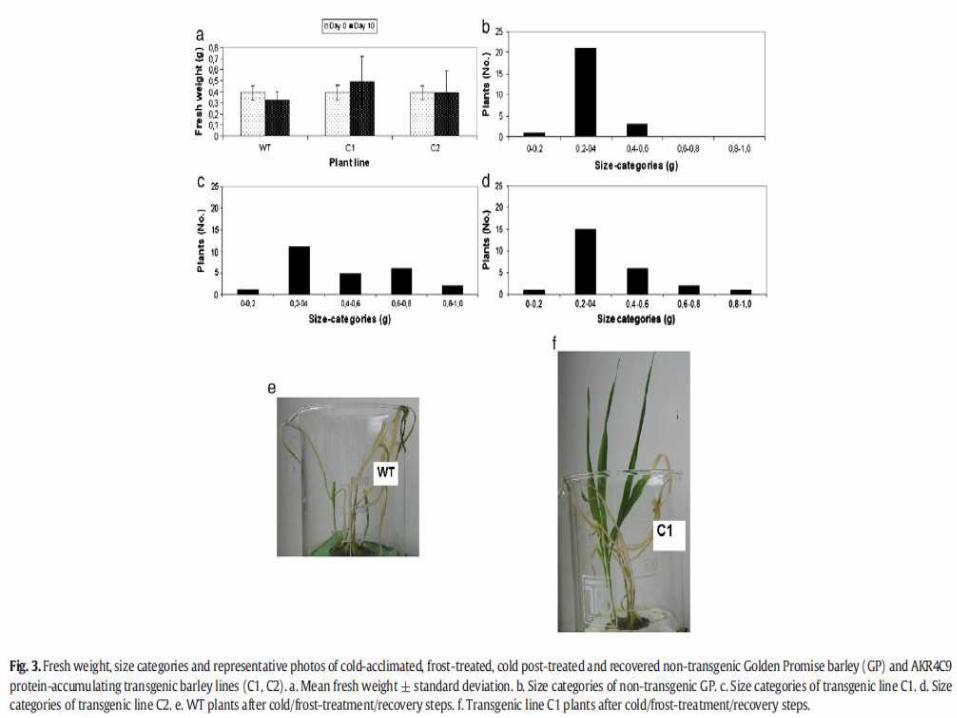
Aldo-keto reductase (AKR) enzymes contribute to reactive aldehyde detoxifying capacity and to osmotic stress protection of various plant species



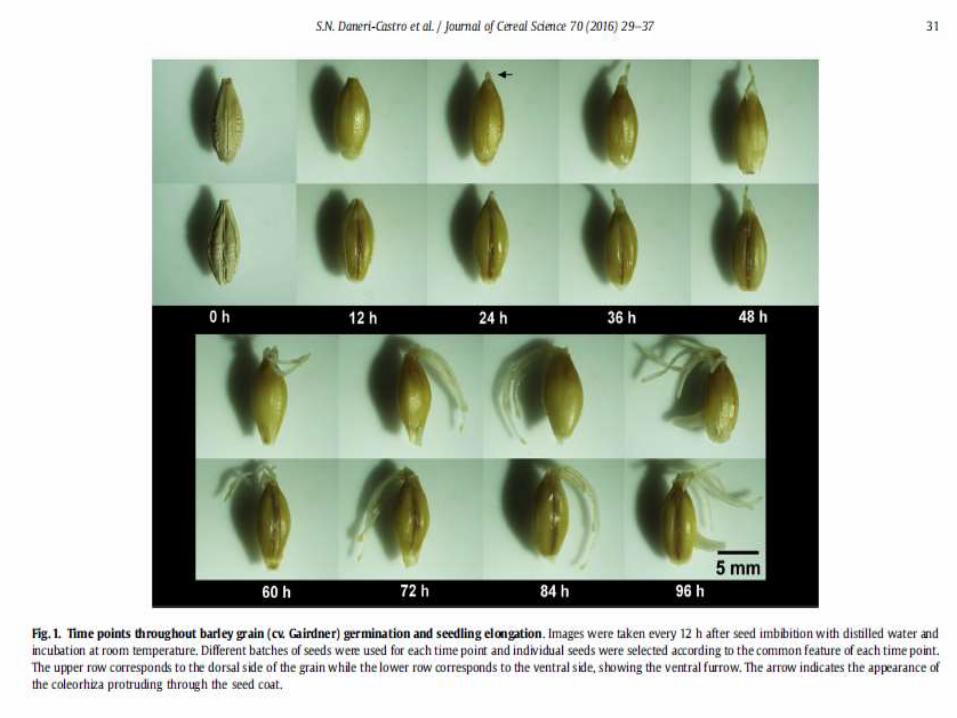
Germination of barley grain is central to the malting industry and is a valuable model for cereal grain germination.To maximize the benefits of the ‘omics’ revolution to the malting industry, there is a need to integrate these data, taking into account barley variety, time, tissue, and specific physiological processes.

CASE STUDY OF
TRANSGENIC BARLEY

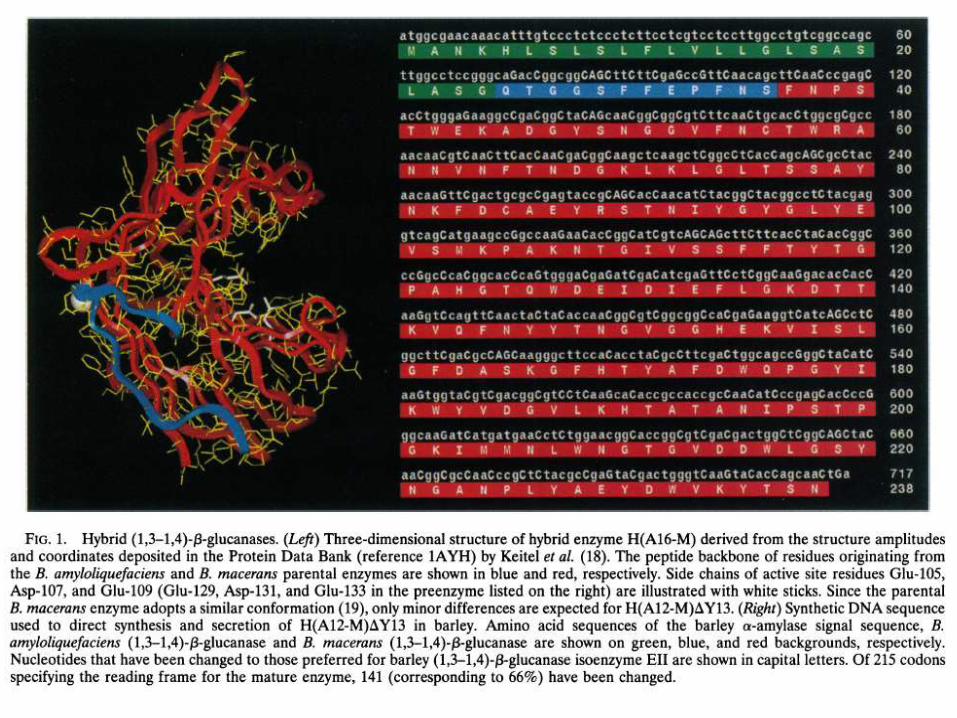
The codon usage of a hybrid bacterial gene encoding a thermostable (1,3-1,4)-13-glucanase was modified to match that of the barley (1,3-1,4)-p-glucanase isoenzymeEl gene. Both the modified and unmodified bacterial genes were fused to a DNA segment encoding the barley high-pI a-amylase signal peptide downstream of the barley (1,3-1,4)-,3-glucanase isoenzyme EII gene promoter. When introduced into barley aleuroneprotoplasts, the bacterial gene with adapted codon usage directed synthesis of heat stable (1,3- 1,4)-p8-glucanase, whereas activity of the heterologous enzyme was not detectable when protoplasts were transfected with the unmodified gene. In a different expression plasmid, the codon modified bacterial gene was cloned downstream of the barley high-pI a-amylase gene promoter and signal peptide coding region. This expression cassette was introduced into immature barley embryos together with plasmids carrying the barand the uidA genes. Green, fertile plants were regenerated and 75% of grains harvested from primary transformants synthesized thermostable (1,3-1,4)-p3-glucanase during germination. All three trans genes were detected in 17 progenies from a homozygous T1 plant.

EXPERIMENTAL PROCEDURES
Organisms and Materials. Grains ofHordeum vulgare L., cv. Himalaya (1985 harvest at Washington State University, Pullman, WA) were used for preparation of protoplasts, and immature embryos were isolated from cv. Golden Promise.Genomic DNA from cultivars Golden Promise and Carlsberg II was purified according to the procedure by Edwards et al. (26). Escherichia coli cells of strain DH5a (27) (Life Technologies,Grand Island, NY) were used for propagation of plasmids, which were purified using the Wizard System (Promega).Nucleotide sequence analysis was on an Applied Biosystems model 373A nucleotide sequencer.
Plasmid Constructions. The plasmid pEmuGN contains the uidA (29) gene encoding the ,3-glucuronidase and has been described (30). The plasmid pUBARN carries the bar gene (31). The plasmids pEII-aH(A12-M)AY13-N and pAMYaH(A12-M)AY13-N carry the unmodified bacterial (1,3-1,4)-f3-glucanase gene fused to the high-pI a-amylase signal peptide coding sequence under control of the promoters from barley (1,3-1,4)-j3-glucanase isoenzyme EII and high-pI a-amylase genes, respectively. These and the plasmids carrying the modified gene [pEII-aH(A12-M)AY13-GC-N and pAMYaH(A12-M)AY13-GC-N]

Transfection of Aleurone Protoplasts. Aleurone protoplasts were prepared (32) and transfected with 50 ,ug of plasmidDNA by PEG-mediated DNA uptake (33). Gibberellin A3 at a final concentration of 1 ,tM was included in all experiments and incubation was for 65 or 110 h at room temperature.Protoplasts were removed by centrifugation at 1000 x g for 5 min and the supernatant was assayed for (1,3-1,4)-,3-glucanase activity.
Plant Transformation, Selection, and Regeneration. Media used for tissue culture and plants were callus induction medium (CIM); plant growth medium (PGM), which is CIM without hormones added as described (34); as well as thehormone-free FHG medium (35). Plantlet induction medium (PIM) is CIM in which the auxin3,6-dichloro-o-anisic acid (Dicamba) is replaced by the cytokinin 6-benzylaminopurine.
Enzyme Assays.(1,3-1,4)-/-Glucanase activity was determined by the method of McCleary (36) using 200 ,ulof azobarley f3-glucan substrate. Analysis of protoplast supernatants was in 50 mM sodium acetate, pH 6.0/5 mM CaCl2 and incubation was at 56°C. Determination of bacterial (1,3-1,4)- P3-glucanase activity in extracts from germinating grains was in 50 mM BisTris (pH 7.4) at 65°C, and homologous barley (1,3-1,4)-43-glucanase activity was determined in 50 mMsodium acetate (pH 4.5) at 30°C.
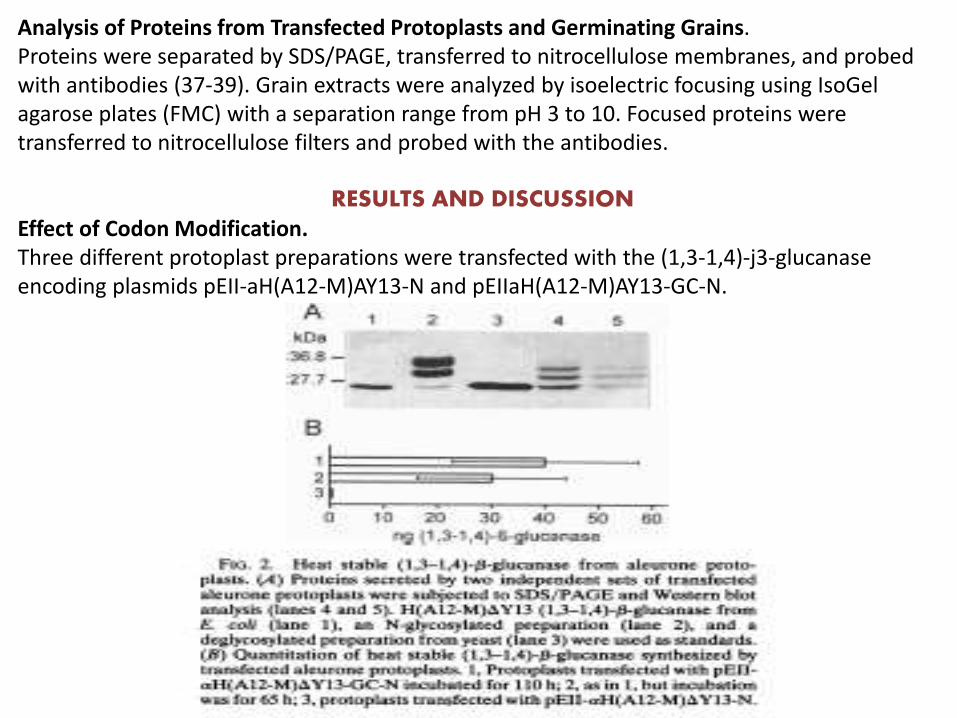
Analysis of Proteins from Transfected Protoplasts and Germinating Grains. Proteins were separated by SDS/PAGE, transferred to nitrocellulose membranes, and probed with antibodies (37-39). Grain extracts were analyzed by isoelectric focusing using IsoGelagarose plates (FMC) with a separation range from pH 3 to 10. Focused proteins were transferred to nitrocellulose filters and probed with the antibodies.
RESULTS AND DISCUSSIONEffect of Codon Modification. Three different protoplast preparations were transfected with the (1,3-1,4)-j3-glucanaseencoding plasmids pEII-aH(A12-M)AY13-N and pEIIaH(A12-M)AY13-GC-N.

Plant Transformation and Regeneration. The plasmid pAMY-aH(A12-M)AY13-GC-N encoding heat stable (1,3-1,4)-,3-glucanase was introduced into immature barley embryos together with plasmids carrying the bar and uidAgenes. In 10 experiments, 293 bisected and 45 whole embryos were bombarded. Twenty-two lines survived the selection procedure and the callus obtained grew well on bialaphoscontaining medium.
Analysis of Primary Transformants and Their Offspring.Analysis of the 14 primary transformants by PCR showed that all plants carried all three genes. Eight T, plants were obtained by germination of immature embryos from transgenic mother plant. Six of the offspring had inherited all three heterologous genes, while the other two did not carry any of them, indicating Mendelian segregation and linkage of the introduced genes.

Characterization of Heterologous (1,3-1,4)-f3-Glucanase.
Aliquots of single grain extracts were analysed by isoelectric focusing. Fig. 5A shows that heat stable (1,3-1,4)-p3-glucanase from a germinating grain of To plant 6.2.1 (lane 2) has the samepI value as a control sample of H(A12-M)AY13 (1,3-1,4)-/3- glucanase produced in E. coli (lane 1). Extract from a wild-type grain gives no activity zone. Fig. SB shows the reaction offocused proteins with antibodies. Soluble proteins from germinating grains of transgenic barley were separated by SDS/PAGE, transferred to nitrocellulose filters, and probed withantibodies. Fig. 5C shows four samples reacting with different intensities. This difference is in agreement with the activities measured using azobarley glucan as substrate. The mobility ofthe transgenic (1,3-1,4)-,3-glucanase is slightly decreased in comparison with the enzyme from E. coli.

Conclusion. In this plant breeding project, we endeavoured to produce barley plants that during steeping and germination express a (1,3-1,4)-f3-glucanase that survives the high temperatures used for kiln drying of green malt. This would allow the enzyme to act in the mash tun as do the thermostable a-amylases from barley, wheat, and rice. Such a heat stable (1,3-1,4)-f3-glucanase synthesized during germination might eliminate the requirement of complete endosperm wall depolymerization in the malting schedule and thereby provide new opportunities for the application of malting and mashing in production of conventional and novel biotechnological commodities.

ReferencesAchard P, Genschik P. 2009. Releasing the brakes of plant growth: how GAs shutdown DELLA proteins Journal of Experimental Botany 60, 1085–1092.
Achard P, Gong F, Cheminant S, Alioua M, Hedden P, Genschik P. 2008. The cold-inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. The Plant Cell 20, 2117–2129
Dewi K. 2006. The role of gibberellins in early growth regulation and dormancy breakage in barley (Hordeum vulgare L. ‘Himalaya’). PhD thesis, Australian National University, Canberra, Australia. Dhillon T, Pearce S, Stockinger EJ, Distelfeld A, Li C, Knox AK, Vashegyi I, Vágújfalvi A,
Galiba G, Dubcovsky J. 2010. Regulation of freezing tolerance and flowering in temperate cereals: the VRN-1 connection. Plant Physiology 153, 1846–1858.
