tyr394 and tyr505 are autophosphorylated in recombinant lck protein-tyrosine kinase expressed in...
TRANSCRIPT

Eur. J. Biochem. 224, 589-596 (1994) 0 FEBS 1994
Tyr394 and Tyr505 are autophosphorylated in recombinant Lck protein-tyrosine kinase expressed in Escherichia coli Pascale JULLIEN', CCcile BOUGERET', Luc CAMOIN2, Monique BODEUS', HervC DURAND', James P. DISANT03, Siegmund FISCHER' and Richard BENAROUS'
Institut National de la SantC et de la Recherche MCdicale Unit6 332, Institut Cochin de GCnCtique MolCculaire, UniversitC Paris RenC Descartes, France Centre National de la Recherche Scientifique UPR 0415, Institut Cochin de GCnCtique Moltculaire, Paris, France Institut National de la SantC et de la Recherche MCdicale Unit6 132, HBpital Necker-Enfants Malades, Paris, France
(Received April WJune 8, 1994) - EJB 94 0501/1
The activity of the Src family protein-tyrosine kinase ~ 5 6 " ~ is regulated by phosphorylation and dephosphorylation of two critical tyrosine residues Tyr394 and Tyr505. Tyr394 is autophosphory- lated after p56Ick activation, whereas phosphorylation of Tyr505 is believed to be due to p50Ak which negatively modulates ~ 5 6 " ~ activity.
To determine whether Tyr505 could be autophosphorylated, we used the prokaryotic glutathione S-transferase expression system to express wild-type Lck, the mutants [Y394F]Lck and [Y505F]Lck, a kinase-deficient ~ 5 6 " ~ with a mutation of the ATP-binding site [K273E]Lck and a double mutant [Y394F, Y505FILck. We studied the kinase activities and the patterns of autophosphorylation for tyrosine residues in these mutants and wild-type Lck both in vivo and in vitro.
Wild-type Lck, [Y505F]Lck and [Y394F]Lck were phosphorylated on tyrosine. Both the kinase- deficient mutant[K273E]Lck and the double mutant [Y394F, Y505FILck did not react with mo- noclonal anti-phosphotyrosine antibody [anti-Y(P) mAb], thus providing evidence that (a) the bacte- rial strains used lacked intrinsic protein-tyrosine kinase activities, and therefore tyrosine phosphory- lations of wild-type Lck, [Y505F]Lck and [Y394F]Lck are due to autophosphorylation occurring in vivo in bacteria, and (b) that p56"* can only be autophosphorylated on two tyrosine residues, namely Tyr394 and Tyr505. Phosphopeptide mapping analysis confirmed that ~ 5 6 ' " ~ can undergo autophos- phorylation on these two tyrosine residues. We propose that autophosphorylation at Tyr505 of ~ 5 6 " ~ may represent an accessory mechanism for the down-regulation of the tyrosine kinase activity of ~56 ' '~ .
Ten distinct members of the src gene family have been cloned and the encoded protein-tyrosine kinases have been characterized (Mustelin, 1994). These include Src, Yes, Fgr, Fyn, Lyn, Lck, Blk, Hck, Yrk and Rak. The protein-tyrosine kinase ~56"' has been found to be exclusively expressed in cells of the lymphoid lineage, predominantly in T lympho- cytes (Fischer et al., 1984; Voronova et al., 1985; Marth et al., 1985; Koga et al., 1986; Trevillyan et al., 1986) and in several human leukaemic T-cell lines (Jurkat, KE37, JM, CEM) (Fischer et al., 1984; Trevillyan et al., 1986). ~ 5 6 " ~ shares structural features with the other members of the Src family, namely (a) a myristate group on the N-terminal gly- cine residue that is involved in membrane binding (Resh, 1990), (b) Src-homology 2 and Src-homology 3 domains
Correspondence to R. Benarous, Institut National de la SantC et de la Recherche Mtdicale Unit6 332, 22 rue MCchain, F-75014, Paris, France
Abbreviations. GST, glutathione S-transferase; GST-Lck, GST- Lck fusion proteins ; anti-Y(P) mAb, anti-phosphotyrosine monoclo- nal antibody; SH2 or SH3, Src-homology 2 or Src-homology 3 do- mains ; IPTG, isopropyl-thio-P-D-galactoside ; PhMeSO,F, phenyl- methylsulfonyl fluoride; [Y394F]Lck, mutant with tyrosine at posi- tion 394 replaced by phenylalanine; [Y505F]Lck, mutant with tyro- sine at position 505 replaced by phenylalanine; [Y394F, Y505F]Lck, mutant with tyrosine residues at positions 394 and 505 replaced by phenylalanine residues; [K273E]Lck, mutant with lysine at position 273 replaced by glutamic acid.
Enzyme. Protein-tyrosine kinase(s) (EC 2.7.1.112).
(SH2 and SH3), and (c) a highly conserved catalytic domain that contains the ATP-binding site and the kinase activity.
~ 5 6 ' ' ~ is physically and functionally associated with at least four important transmembrane T-cell proteins, CD4 (Rudd et al., 1988), CD8 (Veillette et al., 1988), CD2 (Carmo et al., 1993) and the 75-kDa P-chain of the interleukin-2 re- ceptor (Hatakyama et al., 1991). Via these molecules, ~ 5 6 " ~ is presumably involved in T-cell development and prolifera- tion and in the regulation of signal transduction (Bolen and Veillette, 1989; Mustelin and Altman, 1989; Sefton, 1991).
Further characterization of the processes regulating the enzymic function of ~ 5 6 ' " ~ is critical for a better understand- ing of the role of this enzyme in normal cellular physiology. Previous studies have shown that a key mechanism of regula- tion for Src family kinases is phosphorylation and dephos- phorylation of specific tyrosine residues located in the cata- lytic domain which are conserved in all Src family members. Tyr394 is the site of autophosphorylation in vitro in p56lck (Casnellie et al., 1983; Luo and Sefton, 1990), and is phos- phorylated at a low level in vivo. Replacement of Tyr394 by phenylalanine prevents ~56" ' activation (Abraham and Veil- lette, 1990; Veillette and Fourmel, 1990), implying that Tyr394 phosphorylation may up-regulate the enzymic activ- ity of ~ 5 6 " ~ .
Another tyrosine residue, Tyr505, located at a conserved C-terminal site, is extensively phosphorylated in vivo (Am- reim and Sefton, 1988; Marth et al., 1988; Abraham and

590
Veillette, 1990; Abraham et al., 1991). Phosphorylation of Tyr505 down-regulates the enzymic activity of ~56"' (Kmie- cik and Shalloway, 1987; Piwnica-Worms et al., 1987; Am- reim and Sefton, 1988; Marth et al., 1988; Abraham and Veillette, 1990; Abraham et al., 1991). Substitution of Tyr505 by phenylalanine results in constitutively active ~ 5 6 " ~ , capable of transforming rodent fibroblasts (Amreim and Sefton, 1988; Marth et al., 1988). Recently, the protein- tyrosine kinase responsible for phosphorylating the C-termi- nal Tyr527 of ~ 6 0 - ' ~ ' , designated ~ 5 0 ' " ~ (C-terminal Src ki- nase), has been purified and cloned from rat brain (Okada and Nakagawa, 1989; Nada et al., 1991). p 5 0 k was shown in vitro to phosphorylate specifically the negative regulatory sites of p60-"" (Okada and Nakagawa, 1989), ~ 5 6 ' ~ " and ~ 5 9 ~ " (Okada et al., 1991). Bergman et al. showed that ~ 5 0 ' ~ phosphorylates ~ 5 6 " ~ in vitro at Tyr505 and thereby down- regulates its catalytic activity (Bergman et al., 1992). Several groups have found that PSOC'~ expressed in yeast is able to phosphorylate Tyr527 in p60-'" thereby inhibiting its kinase activity in vivo (Superti-Furga et al., 1993; Okada et al., 1993; Murphy et al., 1993).
In the present study, using several mutant forms of ~ 5 6 ' ' ~ expressed in bacteria, we have investigated the possibility that ~56"" could be autophosphorylated on other tyrosine residues besides the well-known autophosphorylation site, Tyr394. We show that ~ 5 6 ' ' ~ is capable of autophosphorylat- ing two tyrosine residues, Tyr505 and Tyr394.
MATERIALS AND METHODS Generation of recombinant pGEX-2T-Lck plasmids
A 1.8-kb cDNA fragment containing the entire human Lck-coding sequence (kindly provided by Roger Perlmutter, University of Washington, Seattle, WA, USA), obtained after NcollHindIII digestion, was inserted using recombinant DNA technology into the bacterial expression vector pGEX- 2T (Pharmacia) at the EcoRI site, in frame with the glutathi- one S-transferase gene. In order to verify the insertion, double-stranded DNA sequencing was carried out using a Sequenase version 2.0 kit (United States Biochemical) following the protocol provided by the manufacturer.
Standard site-directed mutagenesis in M13 phage was performed to generate the [Y394F]Lck and [Y505F]Lck mu- tants (mutants where tyrosine residues at positions 394 and 505 have been replaced by phenylalanine, unpublished re- sults). pGEX-F394 Lck and pGEX-F505 Lck were generated by substituting, respectively, the CluI-EugI or BsmI-StuI mutated fragments in place of the corresponding fragment from normal Lck cDNA, giving rise to the glutathione S- transferase (GST)- [Y394F]Lck fusion protein which is mu- tated at the positive regulatory site and to GST- [Y505F]Lck fusion protein which is mutated at the negative regulatory site. The double mutant [Y394F, Y505FILck was generated by substituting in the [Y505F]Lck mutant the ClaI-EugI fragment from the [Y394F]Lck mutant which contains the Phe394 mutated residue in place of Tyr394.
The murine kinase-deficient [K273E]Lck cDNA was kindly provided by Roger Perlmutter and was inserted into the pGEX-2T vector. This mutant is devoid of any tyrosine kinase activity as a result of a point mutation in the ATP- binding site. In order to distinguish this inactive form of mu- rine ~ 5 6 ' ' ~ from the active forms, we inserted the mutant as a BumHl -Hind111 fragment corresponding to the Lck-coding region deleted from the first 33 amino acid residues giving
rise to a fusion protein 3.3 kDa smaller than the normal 83- kDa GST-Lck fusion protein.
Expression and purification of GST-Lck
The expression and purification of GST-Lck fusion pro- teins and non-recombinant GST protein were performed essentially as described by Bougeret et al. (1993).
Escherichia coli cultures (DH 5a), transformed with either pGEX-2T or recombinant pGEX-2T-Lck, were first grown overnight at 37 "C with agitation in Luria-Bertani medium containing 50 pg . ml-' ampicillin (Sigma), diluted 1: lO in the same medium and incubated at 30°C. Fudon protein expression was induced at 30°C by adding 0.1 mM isopropyl-thio-P-D-galactoside (IPTG, Clontech Laborato- ries) and growth was continued at 25°C for 5 h. Bacteria were harvested, suspended at a 1 : 10 dilution in 50 mM Tris/ HC1, pH 7.4, 1 mM EDTA, 100 mM sodium chloride, 10% glycerol and 1 % Nonidet P-40 containing protease inhibitors [ 1 mM phenylmethylsulphonyl fluoride (PhMeSO,F), 2 % aprotinin, 2 pg . ml-' leupeptin, 2 pg . ml-' pepstatin] and reducing agent (1 mM dithiothreitol), and lysed on ice by adding 0.5 mg . ml-' lysozyme (Boehringer) for 1 h at 3°C. This was followed by sonication and centrifugation at 10000 g for 15 min at 4°C. The supernatant (containing sol- uble protein) was loaded onto a glutathione-agarose (Sigma) affinity column. The column was washed with 5 column volumes 1 mM sodium chloride and ten column vol- umes of 1.5 M sodium chloride, 100 mM disodium hydro- genphosphate (NaClP,). Bound proteins were eluted with 1OmM reduced glutathione (Sigma) in 50mM Tris/HCl, pH 8. Eluted proteins were stored at -80°C.
Proteins were analysed by SDSPAGE, and visualized by Coomassie-brilliant-blue staining and immunoblotting.
Immunoblotting
Immunoblot analyses were performed as described by Towbin et al. (1979). Proteins were fractionated by SDS/ PAGE, transferred electrophoretically to nitrocellulose niem- branes (Schleicher & Schull, BA 85) and probed with mo- noclonal mouse anti-phosphotyrosine antibody [anti-Y(P) mAb] (4G-10, UBI) and an ant i -~56"~ mAb generated against a synthetic peptide corresponding to amino acids at positions 38 -62. Detection of immunoreactive polypeptides was per- formed using lZ51-protein A (Amersham) and subsequent autoradiography.
In vitro autophosphorylation assays of GST-Lck fusion proteins
The reaction mixture (50 pl) contained 50 mM Tris/HCl, pH 7.4, 10 mM manganese chloride, 0.1 mM sodium \ma- date, 1 mM PhMeSO,F, 2% aprotinin, 10 pCi [y-'*P]ATP (5000 Ci . mmol-', Amersham) and usually 100 ng purified enzyme preparation. After 2 min at 25 "C, the reaction was terminated by adding 3 mM Tris/HCl, pH 6.8, 0.1% SDS, 0.5% glycerol, 0.25% 2-mercaptoethanol, 0.01 % bromophe- no1 blue (SDS sample buffer) and proteins were resolved by SDSPAGE. The gel was stained with Coomassie-brilliant blue and was subjected to autoradiography at -80°C on X- ray film (Kodak).

591
Protein-tyrosine kinase assay on the exogenous substrate enolase
The GST-Lck fusion proteins (100 ng) were incubated at 25°C in a reaction mixture (50 pl) containing 50 mM Tris/HCl, pH 7.4, 10 mh4 manganese chloride, 0.1 mM so- dium vanadate, 2% aprotinin, 1 mM PhMeSO,F, 10 pCi [y-32P]ATP (5000 Ci . mmol-', Amersham), 5 pM unlabelled ATP and 5 pg denatured enolase (Boehringer) (Cooper et al., 1984). After 2 min at 25"C, the reaction was stopped by add- ing SDS sample buffer and the proteins were resolved by SDSPAGE followed by autoradiography.
Phosphopeptide mapping of tyrosine-phosphorylated recombinant proteins Digestion with trypsin of gel slices containing in vitro labelled p56Ick
The phosphorylated proteins were isolated after SDS/ PAGE and the slices of the dried gel were rehydrated with 200 pl 50 mM Tns/HCl, pH 8.8, and exhaustively digested with trypsin (50 pg, Sigma) at 37°C for 18 h. Peptides were analyzed either by rerunning the gel slice overlaid with a buffer containing 125 mM Tris/HCl, pH 6.8, 6 M urea on 40% polyacrylamide gel, or by reverse-phase chromatogra- phy (see below).
Digestion with trypsin of nitrocellulose membrane containing in vitro labelled ~ 5 6 " ~
Alternatively, labelled protein was transferred to nitrocel- lulose membrane, visualized, excised and digested with tryp- sin using the method of Boyle et al. (1991).
Briefly, the piece of membrane corresponding to the pro- tein band was soaked in 0.5% polyvinylpyrrolidone (PVP- 360, Sigma) in 1OOmM acetic acid at 37°C for 30 min, washed briefly in water, and with fresh 50mM ammonium bicarbonate, pH 8. Digestion was achieved by incubating the piece of membrane in 200 p1 50 mM ammonium bicarbonate with 10 pg trypsin (modified sequencing grade, Boehringer) for 2 h at 37"C, followed by addition of another 10 pg tryp- sin for 2 h at 37°C. The yield of trypsin-generated peptides was approximately 80%. The soluble digestion protein was lyophylized, washed and phosphopeptides were analysed as mentioned above.
Phosphopeptide analysis
Phosphopeptides formed after trypsin treatment were concentrated to 10 p1, diluted with 140 p1 0.1% tnfluoro- acetic acid (sequenal grade from Pierce) and immediately applied to the column previously equilibrated with this solu- tion. Phosphopeptides were fractionated by reverse-phase chromatography on a Waters model 625 HPLC system at 22°C equipped with a 220 mmX2.1 mm (Aqaupore RP-300) C, column (DEAE). Elution was performed by applying a linear gradient from 35 -7% acetonitrile in 0.1 % trifluoroa- cetic acid within 80 min. Radioactivity in each fraction was analysed by Cerenkov counting in a scintillation counter (Wallac 1410; Pharmacia). Peptides of interest were sub- jected to two-dimensional phosphoamino acid analysis and one-dimensional phosphopeptide mapping.
Phosphoamino acid analysis
and treated with 100 pl constant-boiling 6 M hydrochloric acid (Sigma) for 1 h at 110°C. The phosphoamino acids were analysed on thin-layer cellulose plates (Schleicher & Schull) by two-dimensional electrophoresis at pH 1.9 in the first di- mension and at pH 3.5 in the second dimension (Hunter and Sefton, 1980), using an HTLE-7000 electrophoresis unit (CBS Scientific Co.). The phosphoamino acid spots were identified by spraying the sheet with a 0.25% ninhydrin solu- tion in acetone. The sheet was exposed to detect y-"P-la- belled phosphoamino acids.
Phosphopeptide sequencing HPLC-purified peptides were subjected to sequencing by
automated Edman degradation on an Applied Biosystems se- quencer, using the manufacturers operating procedures (Ed- man and Begg, 1967). Phosphopeptides were applied to pre- cycled filters which were coated with Polybrene (Biobrene from Applied Biosystem) to reduce peptide wash-out and to improve initial yields (Tarr al., 1978). Synthetic peptide, cor- responding to amino acids at positions 492 -509 from human ~ 5 6 ' " ~ , containing Tyr505 as a phosphorylated or non-phos- phorylated residue, was purchased from Chiron Mimotopes.
RESULTS Recombinant Lck expressed in bacteria is autophosphorylated on several tyrosine residues
Using the prokaryotic glutathione S-transferase expres- sion system, we have produced and purified various recombi- nant Lck proteins as GST fusion proteins (Smith and John- son, 1988), by insertion of the cDNAs into the bacterial ex- pression vector PGEX-2T. The p56"" constructs are shown in Fig. 1A.
The different GST -Lck fusion proteins expressed after induction by IPTG and purified on glutathione - agarose beads were analysed by immunoblotting with an anti-p56Ick mAb. All GST-Lck constructs, wild-type Lck, [Y394F]Lck, [Y505F]Lck, the double mutant [Y394F, Y505FILck and [K273E]Lck (Fig. 1B) reacted with the anti-p56lCk mAb.
To investigate whether the recombinant ~ 5 6 " ~ proteins produced in E. coli were phosphorylated on tyrosine resi- due(s), immunoblots were performed with anti-Y(P) mAb (Fig. 1C).
Only the ~ 5 6 " ~ constructs which possess catalytic kinase activities (wild-type Lck, [Y394F]Lck and [Y505F]Lck) re- acted with the anti-Y(P) mAb (Fig. 1C); the double mutant [Y394F, Y505FILck and the kinase-deficient mutant [K273E]Lck did not react with the anti-Y(P) mAb (Fig. 1C).
The absence of reactivity of the kinase-deficient mutant [K273E]Lck with the anti-Y(P) mAb suggested that the bac- teria used in these experiments were devoid of any endoge- nous tyrosine kinase activity capable of phosphorylating ~ 5 6 " ~ . Therefore, we concluded that recognition of the dif- ferent recombinant proteins by the anti-Y(P) mAb was due to the autophosphorylation activity of these proteins.
Reactivity of [Y394F]Lck with the anti-Y(P) mAb indi- cated that ~ 5 6 ' " ~ was capable of undergoing autophosphoryla- tion in vivo in bacteria on additional tyrosine residues besides Tyr394, the well-known site of autophosphorylation. The non-reactivity of the double mutant [Y394F, Y505FILck with anti-Y(P) shows that the wild-type Lck phosphorylates itself onlv on these two residues, Tvr394 and Tvr505. However,
Phosphoamino acids were analysed by the method of Kamps and Sefton (1989). The HPLC fractions were dried
the-level of phosphorylation h [Y394F]Lck (Fig. 1C) was approximately 4-5 times lower than the phosphorylation of

592
A
N H 2 \ 4
myristic cD4,cD~ acid 9
ATP binding site binding site I 1-
C a t a l y t i c d o m a i n Regulatory d o m a i n
wild-type p 5 6 Lck
COOH NH2\
F 3 9 4 p56-Lck
y394 F 5 0 5 COOH N H 2 \ 4
F 5 0 5 ~ 5 6 - L c L
Y394 Y505 COOH
EZ73 p 5 6 - L ~ k
COOH N H 2 \ 4 F 3 9 4 F 5 0 5
F 3 9 4 I F 5 0 5 p56-LCk
GST-Lck, FPS ->
C GST-Lck, FPS ->
mAb a n t i - ~ 5 6 l c k
mAb anti- phosphotyrosine
1 2 3 4 5 Fig. 1. Construction and reactivities of GST-Lck fusion proteins with monoclonal antibodies anti-p56lck and anti-phosphotyrosine. (A) Constructs for the expression of intact and mutated Lck in bacteria. Wild-type or mutated Lck cDNAs were inserted in the pGEX-2T bacterial expression plasmid as indicated in the Materials and Methods section, to generate the different Lck recombinant proteins. Domains of ~ 5 6 " ~ are indicated together with the residues mutated in the various mutants. (B) GST-Lck protein, GST-[Y505F]Lck and GST- [Y394F]Lck mutants react with a monoclonal anti-phosphotyrosine antibody. GST-Lck fusion proteins (GST-Lck, FPs) were analysed by 10% SDSPAGE, electroblotted onto nitrocellulose filters and probed with either anti-p56"' mAb (B) or anti-Y(P) mAb (C). Detection of immunoreactive polypeptides was performed using '251-pr~tein A and subsequent autoradiography. 25 ng GST- [Y394F]Lck (lane l), 100 ng respectively GST-Lck (lane 2), GST-[Y505F]Lck (lane 3), GST-[Y394F, Y505FILck (lane 4), GST-[K273E]Lck (lane 5), were loaded on the gel; the positions of the fusion proteins are indicated (4).
wild-type Lck and [Y505F]Lck, which were both phosphory- lated with similar efficiencies.
Autophosphorylation and kinase activities of the GST-Lck fusion proteins in vitro
In order to confirm that the purified recombinant GST- Lck proteins were active kinases, in vitro phosphorylation
assays were performed (see Materials and Methods section). As shown in Fig. 2, the three recombinant proteins, wild-type GST-Lck, GST-[Y505F]Lck and GST-[Y394F]Lck are phosphorylated in vitro. The kinase-deficient mutant GST- [K273E]Lck and the double-mutant GST- [Y394F, Y505FILck were devoid of any activity (Fig. 2). Quantitative determination of the extent of phosphorylation obtained

593
GST-Lck,FPs ->
enolase -)
1 2 3 4 5 6 7 8 9 10 Fig.2. In vitro tyrosine kinase activity of different GST-Lck fusion proteins (GST-Lck, FPs). GST-[K273E]Lck (100 ng, lanes 1 and 9), GST-Lck (100 ng, lanes 2 and 6), GST-[Y505F]Lck (100 ng, lanes 3 and 7), GST-[Y394F]Lck (100 ng, lanes 4 and 8) and GST- [Y394F, Y505FILck (100 ng, lanes 5 and lo), were incubated in vitro with 10 FCi [y-'*P]ATP as described in the Materials and Methods section, without (lanes 1-5) or with (lanes 6-10) 5 pg of acid-denatured enolase, and were analysed by 10% polyacrylamide SDSPAGE followed by autoradiography.
showed that [Y505F]Lck and wild-type Lck were equally active, whereas [Y394F]Lck was approximately 4-5 times less active than wild-type Lck. These results are similar to those obtained with anti-Y(P) mAb and confirmed that ~ 5 6 ' " ~ can undergo in vitro autophosphorylation on two tyrosine residues.
To further test the tyrosine kinase activities of the GST- Lck fusion proteins, we used enolase as an exogenous sub- strate. The results are shown in Fig. 2. Except for the kinase- deficient mutant [K273E]Lck and the double mutant [Y394F, Y505F]Lck, the different GST-Lck fusion proteins, namely wild-type Lck, [Y505F]Lck and [Y394F]Lck were able to phosphorylate acid-denatured enolase, albeit to different ex- tents. The enzymic activities on enolase reflected the results obtained by autophosphorylation assays or immunoblotting with anti-Y(P) mAb. Namely, [Y394F]Lck was much less active than wild-type Lck and [Y505F]Lck. Interestingly, the [Y394F, Y505F]Lck, which has lost the ability to autopho- sphorylate, is also unable to phosphorylate enolase as an exogenous substrate, although its ATP-binding site is intact in terms of the amino acid sequence.
Phosphopeptide mapping of tyrosine-phosphorylation sites in bacterially expressed Lck
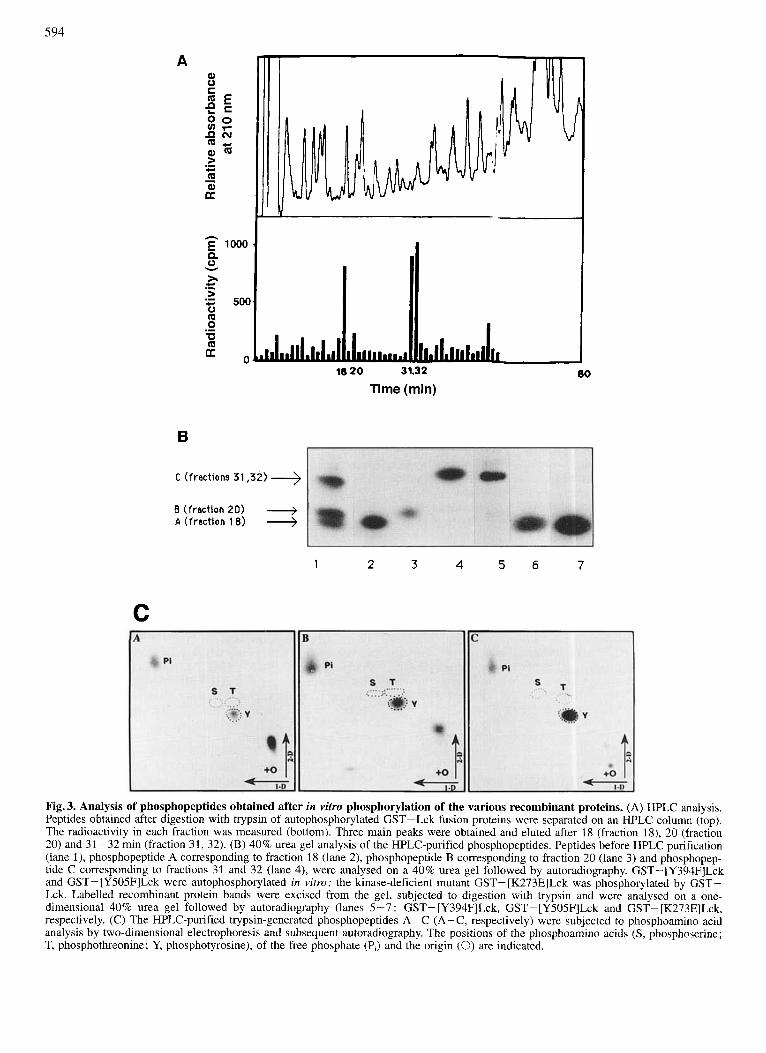
In order to determine which tyrosine residues were phos- phorylated upon ~ 5 6 ' ' ~ autophosphorylation, phosphopeptide mapping of trypsin-generated peptides was performed with each recombinant active ~ 5 6 ' " ~ protein following in vitro phosphorylation assays. Separation of labelled phospho- peptides after one-dimensional electrophoresis on a 40% acrylamideturea gel is shown in Fig. 3. The pattern obtained with GST-Lck revealed three phosphopeptides. The trypsin- generated phosphopeptides were further purified on a re- verse-phase HPLC column (see Materials and Methods sec- tion; Fig. 3A) and the isolated peptides were further resolved on a 40% acrylamidehrea gel (Fig. 3B). The HPLC fractions 18, 20 and 31/32 corresponded to the three phosphopeptides A, B and C, respectively.
We compared the pattern obtained by trypsin treatment of wild-type Lck with the patterns for mutants [Y394FlLck, [Y505F]Lck and the kinase-deficient mutant [K273E]Lck phosphorylated in trans by wild-type Lck. The phospho- peptide A (Fig. 3B) obtained from wild-type Lck and [Y505F]Lck corresponded to a peptide containing the autophosphorylation site, residue Tyr394. The phosphopep- tide C, obtained from wild-type Lck and [Y394F]Lck (Fig. 3), corresponded to a trypsin-generated peptide contain- ing the residue Tyr505. Therefore, we showed that besides
the known autophosphorylation site, Tyr394, ~ 5 6 ' ' ~ has at least one additional site of autophosphorylation, namely the negative regulatory site Ty505. Interestingly, the kinase-defi- cient mutant [K273E]Lck, phosphorylated by wild-type Lck, yielded only one phosphorylation site corresponding to phosphopeptide A which includes Tyr394. Phosphoamino acid analyses of the three phosphopeptides show that these peptides were phosphorylated only on tyrosine residues (Fig. 3C). Peptide B could correspond to the peptide contain- ing Tyr394 running as a doublet or to an other as yet uniden- tified phosphopeptide. The identity of the trypsin-generated peptides A and C as Tyr394-containing and Tyr505-contain- ing peptides, respectively, was further verified by protein se- quencing for peptide A and peptide C and migration of pep- tide C with the synthetic peptide containing TyrP505 (data not shown).
DISCUSSION We have produced various forms of recombinant ~ 5 6 ' " ~
in bacteria as GST-Lck fusion proteins and studied both their pattern of tyrosine phosphorylation and their catalytic activity. Five different Lck constructs were used in this study, three of which were active tyrosine kinases ; namely wild- type Lck, [Y394F]Lck and [Y505F]Lck mutants. Other con- structs were the kinase-deficient [K273E]Lck mutant which served as a substrate for wild-type Lck, and the inactive double-mutant [Y394F, Y505FILck. Immunoreactivity of the first three constructs with anti-Y(P) mAb allowed us to study tyrosine phosphorylation occurring in vivo in bacteria. The kinase-deficient mutant [K273E]Lck did not react with anti- Y(P) mAb, thereby demonstrating that, in the bacterial strains used in this study, no inherent tyrosine phosphoryla- tion was detectable. This indicates that any reactivity of the active Lck constructs with anti-Y(P) mAb was due to autophosphorylation occurring in vivo in bacteria. As [Y394F]Lck was autophosphorylated on tyrosine (although to a lesser extent than wild-type Lck or [Y505F]Lck) this demonstrates that besides the known site of autophosphoryla- tion, Tyr394, ~56'"" can be autophosphorylated on additional tyrosine residue(s). The double-mutant [Y394F, Y505FILck did not display any autophosphorylating activity. This obser- vation demonstrates that ~ 5 6 ' " ~ can autophosphorylate only two tyrosine residues namely Tyr394 and Tyr505. This is in agreement with results previously reported by Bergman et al. (1992). The recombinant p56lck proteins expressed in bacteria were enzymically active in vitro, exhibiting both autophos- phorylation activity and tyrosine kinase activity towards ex- ogenous substrates such as enolase. The [Y394F]Lck was 4-
594
A
h E 1000 Q 0
h > c 500 0 Q 0 0 Q
v
c .- .-
.-
a .
B
1
18 20 31.32
Time (min)
C ( f rac t ions 31,32)
B ( f r a c t i o n 20) + A ( f r a c t i o n 18)
2 3 4 5 6
80
7
C
Fig. 3. Analysis of phosphopeptides obtained after in vitro phosphorylation of the various recombinant proteins. (A) HPLC analysis. Peptides obtained after digestion with trypsin of autophosphorylated GST-Lck fusion proteins were separated on an HPLC column (top). The radioactivity in each fraction was measured (bottom). Three main peaks were obtained and eluted after 18 (fraction 18), 20 (fraction 20) and 31 -32 min (fraction 31, 32). (B) 40% urea gel analysis of the HPLC-purified phosphopeptides. Peptides before HPLC purification (lane l), phosphopeptide A corresponding to fraction 18 (lane 2), phosphopeptide B corresponding to fraction 20 (lane 3) and phosphopep- tide C corresponding to fractions 31 and 32 (lane 4), were analysed on a 40% urea gel followed by autoradiography. GST-[Y394F]Lck and GST- [Y505F]Lck were autophosphorylated in vitro; the kinase-deficient mutant GST- [K273E]Lck was phosphorylated by GST- Lck. Labelled recombinant protein bands were excised from the gel, subjected to digestion with trypsin and were analysed on a one- dimensional 40% urea gel followed by autoradiography (lanes 5-7 : GST- [Y394F]Lck, GST-[Y505F]Lck and GST- [K273E]Lck, respectively. (C) The HPLC-purified trypsin-generated phosphopeptides A-C (A-C, respectively) were subjected to pbosphoamino acid analysis by two-dimensional electrophoresis and subsequent autoradiography. The positions of the phosphoamino acids (S, phosphoserine; T, phosphothreonine; Y, phosphotyrosine), of the free phosphate (P,) and the origin (0) are indicated.

595
5 times less active than wild-type Lck and [Y505F]Lck. The double mutant [Y394F, Y505FILck had no detectable kinase activity.
Autophosphorylation of ~ 5 6 ' " ~ on Tyr505 was also ob- served when p56lrk was overexpressed in eukaryotic cells such as Sf9 cells (Jullien, P., Benarous, R. and Fischer, S., unpublished results). Such Tyr505 autophosphorylation could be due to the high level of Lck expression reached in these different systems. Overexpression of ~ 5 6 " ~ could favor ~ 5 6 " ~ dimerization. In bacteria, homodimeric interactions between ~ 5 6 ' ~ ' ' fusion proteins through their GST moiety could also occur. However, by contrast with active recombinant Lck proteins, which can undergo autophosphorylation on both Tyr394 and Tyr505, the [K273E]Lck kinase-deficient mutant is phosphorylated by wild-type Lck only on residue Tyr394 and not on Tyr505. Hence, wild-type Lck is unable to phos- phorylate in trans the kinase-deficient mutant on residue Tyr.505, although both proteins are fused to GST and could dimerize through their GST moiety. This observation is com- patible with an intramolecular mechanism for the autophos- phorylation of ~ 5 6 " ~ at residue Tyr505. However, it is also possible that in kinase-deficient [K273E]Lck, residue Tyr505 occupies an unfavorable conformation such that it cannot be phosphorylated in trans by wild-type Lck.
Phosphopeptide mapping performed after in vitro autophosphorylation of the three active constructs confirmed that ~ 5 6 ' ' ~ is indeed capable of autophosphorylation on one additional tyrosine residue besides Tyr394, the negative regu- latory site, Tyr505. The identity of the trypsin-generated phosphopeptides was confirmed by sequencing and by com- parison with digest patterns after trypsin treatment of ~ 5 6 " ~ described earlier (Danielian et al., 1989; Casnellie et al., 1982; Casnellie, 1987; Marth et al., 1988).
Our results are in agreement with those reported by MacAuley et al. (1993) on the ability of p60'-"" to be auto- phosphorylated on Tyr527 in Saccharomyces cerevisiae. The authors suggested that autophosphorylation could provide a possible mechanism by which the partially transforming and non-transforming mutants are phosphorylated at Tyr527. Bra- godia et al. (1991) also found that p60"-"" isolated from mi- totic cells had lower levels of phosphorylation at Tyr527 if this p60 ST' was a lunase-inactive mutant rather than wild- type p60'-'". These authors claim that p60 7Tc kinase activity is required to maintain high stoichiometry of phosphorylation during mitosis, presumably by autophosphorylation. Hence, the autophosphorylation on the C-terminal regulatory tyro- sine site might be a general characteristic of all protein-tyro- sine kinases of the Src family and might have implications for the control of the catalytic activity of these tyrosine ki- nases.
Recent results on the phenotypes of mice lacking the csk gene have shown convincingly that ~ 5 0 " ~ is a major negative regulator of Src-like kinases in vivo. However, other data show that p 5 0 k might not be the only enzyme that phospho- rylates the C-terminal negative regulatory tyrosine residue of Src family kinases in vivo. Recently, Klages et al. (1994) and Bennet et al. (1994) have cloned cDNAs coding for Csk-like protein-tyrosine kinases, Ctk and Matk, from mouse brain and from human megakaryocytes, respectively. Ctk is capa- ble of phosphorylating ~ 5 6 " ~ at its regulatory site (Klages et al., 1994).
Immato and Soriano (1993) have shown that in cell lines established from embryonic mice lacking p5Wk, p60 'r is still phosphorylated to a low level on Tyr527. The authors propose two possible mechanisms, (a) phosphorylation by
a protein-tyrosine kinase different from p5Pk , TPKIIB for instance (Marin et al., 1991), or (b) autophosphorylation. The ability of ~ 5 6 ' ' ~ , p60 ST' and maybe all kinases of the Src family, to auto-control their catalytic activity through tyro- sine phosphorylation at the C-terminus may represent an ac- cessory mechanism to keep the Src family protein-tyrosine kinases inactive. It is possible that several mechanisms may co-operate in order to maintain the tight control of catalytic activity of the Src family tyrosine kinases.
The autophosphorylation of p56"" on Tyr505 reported in this study could be attributed to the high level of lck expres- sion in bacteria. Although such high levels are rarely attain- able in eukaryotic cells, one can speculate that under some condition such cells acting via receptor activation can induce a temporary and locally elevated concentration of ~ 5 6 " ~ in membranous structures, as has indeed been shown by immu- nofluorescense (Marie-Cardine et al., 1992). Such an event could result in Tyr505 autophosphorylation.
We thank Roger Perlmutter for the gift of Lck cDNAs, Heidi Mohn and Margaret Frame for critical reading of the manuscript. This study was supported in part by grants from the Agence Natio- nale de Recherches sur le SIDA (ANRS, 711 loo), from the Associa- tion pour la Recherche contre le Cancer (ARC, 6878), from the Ligue Nationale contre le Cancer, ComitC de Pans (RS 7/93), and from the Fondation pour la Recherche Mkdicale (FRM). This study is part of P. Jullien's Ph.D. thesis at the UniversitC Paris 11 (France). P. J. is a student fellow of the Ligue Nationale contre le Cancer.
REFERENCES Abraham, N., Miceli, M. C., Pames, J. R. & Veillette, A. (1991)
Enhancement of T cell responsiveness by the lymphocyte spe- cific tyrosine protein kinase p561ck, Nature 350, 62-66.
Abraham, N. & Veillette, A. (1990) Activation of p561ck through mutation of a regulatory carboxy-terminal tyrosine residue re- quires intact site of autophosphorylation and myristylation, Mol. Cell. Biol. 10, 5197' 5206.
Amreim, K. & Sefton, B. M. (1988) Regulatory of a site of tyrosine phosphorylation in the lymphocyte-specific tyrosine protein ki- nase, p561ck, reveals its oncogenic potential in fibroblast, Proc. Natl Acad. Sci. USA 85,4247-4251.
Bennett, B. D., Cowley, S., Jiang, S., London, R., Deng, B., Gra- barek, J., Groopman, J . E., Goeddel, D. V. & Avraham, H. (1994) Identification and characterization of a novel tyrosine kinase from megakaryocytes, Biochemistry J. 269, 1068 - 1074.
Bergman, M., Mustelin, T., Oetken, C., Partanen, J., Flint, N. A,, Amreim, K. E., Autero, M., Bum, P. & Atilo, K. (1992) The human p50csk tyrosine kinase phosphorylates p561ck at Tyr-505 and down regulates its catalytic activity, EMBO J. 11, 2919- 2924.
Bolen, J. 8. & Veillette, A. (1989) A function for the lck protoonco- gene, Trends. Biochem. Sci. 14, 404-407.
Bougeret, C., Rothhut, B., Jullien, P., Fischer, S. & Benarous, R. (1993) Recombinant Csk expressed in Escherichia coli is auto- phosphorylated on tyrosine residue(s), Oncogene 8, 1241 - 1247.
Boyle, W. J., Van Der Geer, P. & Hunter, T. (1991) Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates, Methods Enzymol. 201,
Bragodia, S., Chackalaparampil, I., Kmiecik, T. E. & Shalloway, D. (1991) Altered tyrosine 527 phosphorylation and mitotic activa- tion of p60 c-src, Nature 349, 172-175.
Carmo, A. M., Mason, D. W. & Beyers, A. D. (1993) Physical asso- ciation of the cytoplasmic domain of CD2 with the tyrosine ~ 5 6 " ~ and ~ 5 9 ~ " , Eur. J. Immunol. 23, 2196-2201.
Casnellie, J., Harrison, M., Hellstrom, K. & Krebs, E. (1983) A lymphoma cell line expressing elevated levels of tyrosine phos- phorylation, J. Biol. Chem. 258, 10738- 10742.
110-149.

596
Casnellie, J. E. (1987) Sites of in vivo phosphorylation of the T cell tyrosine protein kinase in LSTRA cells and their alteration by tumor-promoting phorbol esters, J. Biol. Chem. 262, 9859- 9864.
Casnellie, J. E., Harrison, M. L., Pike, L. J., Hellstrom, K. E. & Krebs, E. J. (1982) Phosphorylation of synthetic peptides by a tyrosine kinase from the particulate fraction of a lymphoma cell line, Proc. Natl Acad. Sci. USA 79, 282-286.
Cooper, J. A., Esch, F. S., Taylor, S. S. & Hunter, T. (1984) Phos- phorylation sites in enolase and lactate dehydrogenase utilized by tyrosine protein kinases in vivo and in vitro, J. Biol. Chem. 2.59, 7835-7841.
Danielian, S., Fagard, R., Alcover, A., Acuto, 0. & Fischer, S. (1989) The lymphocyte-specific protein kinase ~ 5 6 ' ~ ~ is hyper- phosphorylated on serine and tyrosine residues within minutes after activation via T cell receptor or CD2, Eul: J. Immunol. 19,
Edman, P. & Begg, G. (1967) A protein sequenator, Eul: J Biochem.
Fischer, S., Fagard, R., Gacon, G., Genetet, N., Piau, J. P. & Blaineau, C. (1 984) Stimulation of tyrosine phosphorylation in lectin-treated human lymphocytes, Biochem. Biophys. Res. Com- mun. 124, 682-689.
Hatakyama, M., Kono, T., Kobayashi, N., Kawahara, A,, Levin, N. D., Pelmutter, R. M. & Taniguchi, T. (1991) Interaction of the IL-2 receptor with the src-family kinase p561ck. Identification of novel intermolecular association, Science 252, 1523 - 1528.
Hunter, T. & Sefton, B. M. (1980) Transforming production of rous sarcoma virus phosphorylates tyrosine, Proc. Natl Acad. Sci.
Immato, A. & Soriano, P. (1993) Disruption of csk gene, encoding a negative regulator of src family tyrosine kinases, leads to neu- ral tube defects and embryonic lethality in mice, Cell 73, 1117- 1124.
Kamps, M. P. & Sefton, B. M. (1989) Acid and base hydrolysis of phosphoproteins bound to immobilon facilitates analysis of phosphoaminoacids in gel-fractionated proteins, Anal. Biochem. 176, 22-27.
Klages, S., Adam, D., Class, K., Fargnoli, J., Bolen, J. B. & Penhal- low, R. C. (1994) Ctk: a protein-tyrosine kinase related to Csk that defines an enzyme family, Proc. Natl Acad. Sci. USA 91,
Kmiecik, T. E. & Shalloway, D. (1987) Activation and suppression of pp60c-src transforming ability by mutation of its primary site of tyrosine phosphorylation, Cell 49, 65-73.
Koga, Y., Caccia, N., Toyonoga, B., Spolski, R., Yanagi, Y., Yoshi- kai, Y. & Mak, T. W. (1986) A human T cell-specific cDNA clone (YT16) encodes a protein with extensive homology to a family of protein-tyrosine kinase, EUK J. Immunol. 16, 1643- 1646.
Luo, K. & Sefton B. M. (1990) Cross-linking of T-cell surface mole- cules CD4 and CD8 stimulates phosphorylation of the Ick tyro- sine protein kinase at the autophosphorylation site, Mol. Cell. Biol. 10, 5305-5313.
MacAuley, A,, Okada, M., Nada, S., Nakagawa, H. & Cooper, J. A. (1993) Phosphorylation of src-mutants at Tyr527 in fibroblasts does not correlate with in vitro phosphorylation by Csk, Onco- gene 8, 117-124.
Marie-Cardine, A,, Maridonneau-Parini, I., Ferrer, M., Danielian, S., Rothhut, B., Fagard, R., Dautry-Varsat, A. & Fischer, S. (1992) The lymphocyte-specific tyrosine protein kinase p561ck is endo- cytosed in jurkat cells stimulated via CD2, J. Zmmunol. 148, 3879-3884.
Marin, O., Donella-Deana, A., Brunati, A. M., Fischer, S. & Pinna, L. A. (1991) Peptides reproducing the phosphoacceptor sites of pp69c-src as substrates for TPK-IIB, a splenic tyrosine kinase devoid of autophosphorylation, J. Biol. Chem. 266, 17798- 17806.
Marth, J. D., Peet, R., Krebs, E. G. & Perlmutter, R. M. (1985) A lymphocyte-specific protein tyrosine kinase gene is arranged and overexpressed in the murine cell lymphoma LSTRA, Cell 43, 393-404.
2183-2189.
I , 80-91.
USA 77, 1311-1315.
2597 -2601.
Marth, J. D., Cooper, J. A,, King, C. S., Ziegler, S. F., Tinker, D. A,, Overell, R. W., Krebs, E. G. & Perlmutter, R. M. (1988) Neoplastic transformation induced by an activated lymphocyte- specific protein tyrosine kinase (pp561ck), Mol. Cell. Biol. 8, 540-550.
Murphy, S. M., Bergman, M. & Morgan, D. 0. (1993) Suppression of c-Src activity by C-terminal Src kinases involves the c.-Src SH2 and SH3 domains. Analysis with Saccharomyces cereviyiae, Mol. Cell. Biol. 13, 5290-5300.
Mustelin, T. (1994) Src family tyrosine kinases in leukocytes. Mo- lecular Biology Intelligence Unit, R. G. Landes Co.
Mustelin, T. & Altman, M. (1989) Do CD4 and CD8 control T-cell activation via a specific tyrosine protein kinase? Immunology Today 15, 189-192.
Nada, S., Okada, M., MacAuley, A,, Cooper, J. A. & Nakagawa, H. (1991) Cloning of a complementary DNA for a protein-tyrosine kinase that specifically phosphorylates a negative regulatory site of p60c-src, Nature 351, 69-72.
Okada, M., Howell, B. W., Broome, M. A. & Cooper, J. A. (1 993) Deletion of the SH3 domain of Src interferes with regulation by the phosphorylated carboxyl-terminal tyrosine, J. Biol. C'hem. 268, 18070- 18075.
Okada, M., Nada, S., Yamanashi, Y., Yamamoto, T. & Nakagawa, H. (1991) CSK: a protein tyrosine kinase involved in regulation of src family kinase, J. Biol. Chem. 266, 24249-24252.
Okada, M. & Nakagawa, H. (1989) A protein tyrosine kinase in- volved in regulation of pp60src function, J. Biol. Chem. 264,
Piwnica-Worms, H., Saunders, K. B., Roberts, T. M., Smith, A. E. & Cheng, S. H. (1987) Tyrosine phosphorylation regulates the bio- chemical and biological properties of pp6Oc-src, Cell 49, 75-82.
Resh, M. D. (1990) Membrane interactions of pp6Ov-src, a model for myristylated protein kinase, Oncogene 5 , 1437- 1444.
Rudd, C. E., Trevillyan, J. M., Dasgypta, J. D., Wong, L. L. & Schossman, S. F. (1988) The CD4 receptor is complexed in de- tergent lysates to a protein tyrosine kinase (pp58) from human T lymphocytes, Proc. Natl Acad. Sci. USA 8.5, 5190-5194.
Sefton, B. M. (1991) The Ick tyrosine protein kinase, Oncogme 6,
Smith, D. B. & Johnson, K. S. (1988) Single step purification of polypeptides expressed in Escherichia coli as fusion with gluta- thione S-transferase, Gene (Amst.) 67, 31 -40.
Superti-Furga, G., Fumagalli, S., Koegl, M., Courtneidge, S. A. & Draetta, G. (1993) Csk inhibition of c-Src activity requires both the SH2 and SH3 domains of Src, EMBO J. 12, 262552634,
Tarr, G. E., Beecher, J. F., Bell, M. & McKean, D. J. (1978) Po- lyquaternary amines prevent peptide loss from sequenators, Anal. Biochem. 8.5, 126.
Towbin, H., Staehelin, T. & Gordon, G. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets. Procedure and some applications, Proc. Natl Acad. Sci. USA 76,4350-4354.
Trevillyan, J. M., Lin, Y., Chen, S. J., Phillips, C. A. & Linna, T. J. (1 986) Identification of the human T-lymphocyte protein-tyro- sine kinase by peptide-specific antibodies, Biochem. Biophys. Res. Commun. 888, 286-295.
Veillette, A., Bookman, M. A., Horack, E. M. & Bolen, J. B. (1988) The CD4 and CD8 T cell surface antigens are associated with the internal membrane tyrosine protein kinase p561ck, Cell 5.5, 301-308.
Veillette, A., Bolen, J. B. & Bookman, M. A. (1989) Alterations in tyrosine phosphorylation induced by antibody mediated cross- linking of the CD4 receptor of T lymphocytes, Mol. Cell. Biol. 9,4441 -4446.
Veillette, A. & Fourmel, M. (1990) The CD4 associated tyrosine protein kinase p561ck is positively regulated through its site of autophosphorylation, Oncogene 5, 1455 - 1462.
Voronova, A. F., Buss, J. E., Patschinsky, T., Hunter, T. & Sefton, B. (1 985) Characterization of the protein apparently responsible for the elevated tyrosine protein kinase activity in LSTRA cells, Mol. Cell. Biol. 4, 2705-2712.
20886 - 20 893.
683-686.