tyrosine hydroxylase gene expression in human ventral mesencephalon: detection of tyrosine...
TRANSCRIPT

.lournal of Neuroscience Research 25569475 ( 1990)
Rapid Communication Tyrosine Hydroxylase Gene Expression in Human Ventral Mesencephalon: Detection of Tyrosine Hydroxylase Messenger RNA in Neurites S. Dumas, F. Javoy-Agid, E. Hirsch, Y . Agid, and ,J. Mallet Laboratoirc dc Neurohiolngie Cellulaire. Ccntrc National dc la Recherche Scicntifiquc. Gif-sur-Yvette (S.D. . J .M. ) . and Laboratoire dt: Medecine Experimentale. INSERM Ll B Y . Hhpital de la Salp6triere. Paris (F.J . -A, . E .H. . Y . A . ) . France
The distribution of both tyrosine hydroxylase (TH) antigen and messenger RNA was investigated in the ventral mesencephalon from human brain using a "S-labeled human TH cDNA probe and a polyclonal TH antibody. A hybridization signal was detected not only in the perikaryon but also in the neurites, most probably dendrites, of immunoreactive neurons. In cell bodies, the level of TH mRNA varies, revealing a regional heterogeneity. In the substantia nigra of patients with Parkinson's disease the number of labeled cells was markedly reduced as compared to controls.
Key words: dendrites, Parkinson's disease, TH
INTRODUCTION
Neuronal cells can be conveniently classified in terms of the neurotransmitter-synthesizing enzymes that they contain. Appropriate antibodies stain not only cell bodies but also axons and nerve terminals, where the enzymes are transported and where the bulk of neuro- transmitter synthesis takes place. However, in some in- stances. intensive transport of the enzyme along axons may significantly deplete it in cell bodies, a situation that has been circumvented by blocking axonal flow with colchicine (Mugnaini and Oertel, 1985). A more satis- factory means of overcoming this problem is to perform in situ hybridization with the corresponding cDNA probes; messenger RNAs are concentrated in the soma and have, so far, not been detected in nerve terminals. Such a restricted distribution facilitates the analysis of mRNA levels, as well as of their modulation in response to stimuli, and allows a further classification of neurons (Chesselet and Robbins, 1989).
We have previously reported the direct visualiza- tion of the TH gene activity and the modulation induced by reserpine treatment on rat brain sections in the locus coeruleus. the ventral tegmental area, and the substantia nigra (Berod et a]. , 1987). Similarly, Schalling and col- laborators ( 1989), using oligonucleotide probes, have re- cently shown a rapid increase in TH mRNA level in human sympathetic ganglia after electrical stimulation. In the present study, as a first step toward exploring the physiopathology of neurodegenerative disorders, such as Parkinson's disease (PD), we analyzed the expression of human TH mRNA in the substantia nigra of postmortem brain samples. Using conditions that allow detection of TH mRNA and protein on the same section, our data show the presence of TH mRNA in neurites of some dopaminergic neurons of the ventral mesencephalon. These neurites are likely to correspond to dendrites.
MATERIALS AND METHODS Tissue Preparation
Tissue samples were obtained postmortem from the brains of four individuals suffering from no known neu- rologic disorders (from 67 to 92 years old, with post- mortem delay between 4 and 7 hr) and from the brains of two patients deceased with clinically and histopatholog- ically diagnosed PD (6 1 and 68 years old, with postmor- tem delay of 6 and 18 hr. respectively). Brain tissues
Received December 30, 1989; accepted February 7, 1990.
Address reprint requests to Dr. Jacques Mallet, Lahoratoire de Neu- robiologie Cellulaire. Centre National de la Recherche Scientifique, 91 198 Gif-sur-Yvette Cedex. France.
0 1990 Wiley-Liss, Inc.

,570 Dumas et al.
were obtained within 2 hr after autopsy (postmortcm de- lay). The brainstem was isolated. and tissue blocks. 1 cni thick, containing the substantia nigra and the ventral tegniental area were dissected and fixed by immersion for 3 days at 4°C in 0. I M sodiuni phosphate buffer. pH 7.4. containing 4% paraformaldehyde and 15% saturated picric acid. Tiswes were then exposed successively at 4°C to ;I serics of wash solutions composed of 0. 1 M sodium phosphate buffer containing consecutively OC%, 5 % , 10%. 15% , and 20% sucrosc for 12 hr each (Gray- biel et al.. 1987) before being rapidly frozen in dry ice powder o r in isopentane cooled t o -70°C in nitrogen vapour. Sections ( I S pin) were cut with a cryostat. mounted o n gelatinc-coated slides. and stored at - 70°C until use.
Probe Preparation A 1.8 kb cDNA insert (HTH2: Grirna et al..
1987). containing the cotnplete coding and 3' untrans- lated sequence of human HTH2 mRNA was used i n these experiments. The insert. prepared by restriction endonucleasc digestion of plasmid DNA. was purified by electroclution after agarosc gel electrophoresis and labeled by nich-translation (Rigby et al.. 1977) with is S - s u bs t i t u t ccl ade nos i ne and c y t id i ne 5 ' ( u- t h i o ) - triphosphate (Amersham) to a specific activity of 6-9 x 10' cpmipg. The average size o f the nick-translated l'ragments wxs about 150 base pairs. As a control dopaniine-P-hydroxylase cDNA ( L ~ I I I O U ~ O L I X ct a]. . 1987) w;is nick-translated and ~tsed as described tor the TH probe
Hybridization Procedure Hybridization was perfcmiied according to Berod ct
al. ( 1987 ). with some modifications. Sections were prc- hybridized for I hr at room temperature in a ao lu t ion containing 4 x SSC ( 1 X SSC - 0.15 M sodium chlo- ridciO.015 M sodiuni citrate. pH 7.0) and I x Dcn- hardt's solution ( I x Denhartlt's solution = 0.02% FicolliO.02% polyvinyIpyrrolidone/0.03'%~ bovine serum albumin). The slides were then clehydrated through graded alcohols and allowed to clt-y. About 0.1 ng of nick-translated probe in 30 p.1 of hybridiLation solution (50% forrnamide. 4 x SSC. 1 x Denhardt's solution, I % sarcosyl. 100 mM dithiothrcitol. 0. 1 M potassium phosphate. pH 7.4. 100 ng of' pBR322. 100 ng of Es- c~ho-ic~hic/ coli tRNA and I00 ng o f denaturated tierrins sperm DNA) was boiled for I0 min, applied to each section. and sealed under a coverslip. Hybridization w;is allowed to proceed for 18 hr at 32°C. Coverslips were then gently rcmoved. and the sections were washed at 42°C twice f o r 30 min i n I x SSC and twice for 30 min in 0.1 x SSC.
Competition experiments were perlhrnietl o n sev-
eral ;id.jricent sections. Sections were hybridized a s de- scribed above except that a 100-fold excess of gcl-puri- fied nonradioactive TH cDNA was added to the hybridization medium.
Immunohistochemical Procedure Alter posthybridization washes. the sections were
fixed i n a 4% solution of parsfhrmaldehyde in 0. 1 M phosphate buffer. pH 7.4. and then treated by inimuno- histochemislry with TH antibodies (from Eugene Tech International). The immunological procedure was per- formed using the ABC kit Vecstastain from Vector Lab- oratories and the procedure o f Riche et al. (1989). Incu- bation with TH antibodies was performed for 72 hr at 4°C.
Analysis of Autoradiograms Autoradiograms of tissue were generated by appo-
sition o f "S-labeled sections to X-ray films (p rnax film; Amersham) or by dipping i n ;I I : I mixture of NTB? Kodak emulaion and I X SSC maintained at 42°C. Aftet- appropriate times o f exposure ( 2 days and 3-5 wcehs. respectively). films and emulsions were developed. Tis- sue sections that had not been subjected to the immuno- logical procedure were stained with cresyl violet.
Optical Observations
dcr classical conditions (bright-field view). Slides were exumined using a Leitz microscope un-
RESULTS
Frontal sections through the dopaminergic areas of hit man mew nceidialon we re hybridized with -"S- I abe led TH probe a s described in Materials and Methods. Auto- raclio~raphy with X-ray film i n contact with the sections revealed an intense signal i n the ventral tegmental area and in the substantia nigra (Fig. l a ) . No signal was ob- served when an excess of unlabeled cDNA was atlcled to the hybridization medium o r when a labeled cDNA probe encoding human clopamine-P-liyclroxyla~e was used (Fig. Ib).
Treatnient of the tissue sections with an emulsion showed that most labeled cells of the substantia nigra contained ticuromclanin (Fig. Ic). a characteristic o f cat- echolaminergic cells from the substantia nigra (Bogerts. 1981: Gaspar ct al.. 1983). No hybridization signal ia observed in the control experiments (Fig. Id) .
Incubation of the tissue sections with TH antibod- ies following the hybridization procedure allowed visu- alization on the same section of both TH mRNA and TH antigen. As is shown in Figures 2 and 3, a similar pattern ot' distribution was obtained with the TH antibody and TH cDNA probe. Cells that did not contain neuromcla-
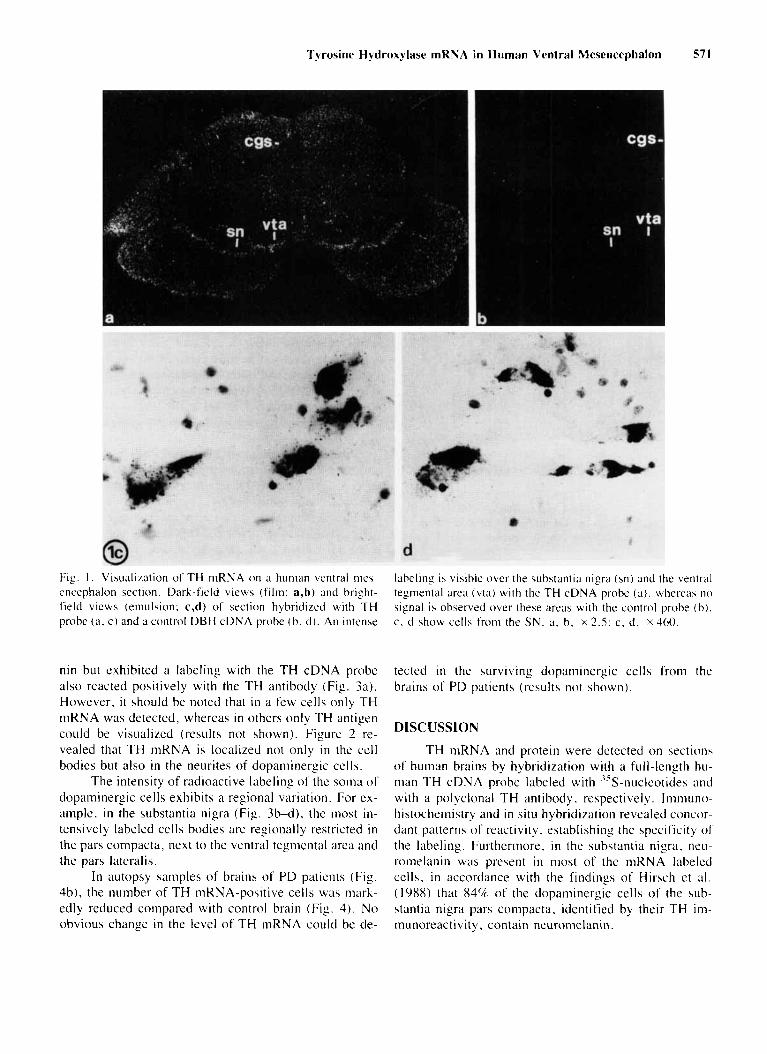
Tj rosine Hydroxglase mRNA in Human Ventral Mesencephalon 571
Fig. I . Visualization of TH mRNA on a hummi ventral nies- cncephalon section. Dark-field views (film; a,b) and bright- field vicws (emulsion: c,d) o f section hybridized with TH probe (a. c ) and a control DBH cDNA probe (b. (1). An intense
labeling is vihible over the substantia nigra (sn) and the ventt-id tegmental area (vta) w i t h the TH cDNA probe (a) . whereas no signal is observed over these ;mils with the control probe ( b ) . c. d show cel ls from the SN. ;I, b. x 2.5: c. d. ~ 4 6 0 .
n i n but exhibited a labeling with the TH cDNA probc also reacted positively with the TH antibody (Fig. 3a). However. i t should be noted that in a few cells only TH m R K A was detected, whereas i n others on ly TH antigen could be visualized (results not shown). Figure 2 re- vealed that TH mRNA is localized not only in the cell bodies but also in the neurites of dopaminergic cells.
The intensity of radioactive labeling o f the soma o f dopaminergic cells exhibits a regional variation. For ex- ample. in the substantia nigra (Fig. 3b-d), the most in- tensively labeled cells bodies are regionally restricted in the pars compacta, next to the ventral tcgniental area and the pars lateralis.
In autopsy samples o f brains of PD patients (Fig. 4b) , the number of TH mRNA-positive cells was mark- edly reduced compared with control brain (Fig. 4) . No obvious change in the level of TH mRNA could be de-
tected i n the surviving dopaniinergic cells from the brains of PD patients (results not shown).
DISCUSSION
TH mRNA and protein were detected o n sections o f human brains by hybridization with a full-lcngth hu- nian TH cDNA probe labeled with 35S-nuclcotides and with a polyclonal TH antibody. respectively. Immuno- histochemistry and in situ hybridization revealed concor- dant patterns of reactivity. establishing the specificity of' the labeling. Furthermore. i n the substantia nigra, neu- romelanin wab present i n most of the mRNA labeled cella, in accordance with the findings of Hirsch et al. ( 19x8) that 84%' of the dopaniinergic cells o f the sub- stantia nigra pars compacta. identified by their TH im- munoreactivity, contain neuromelanin.
. .
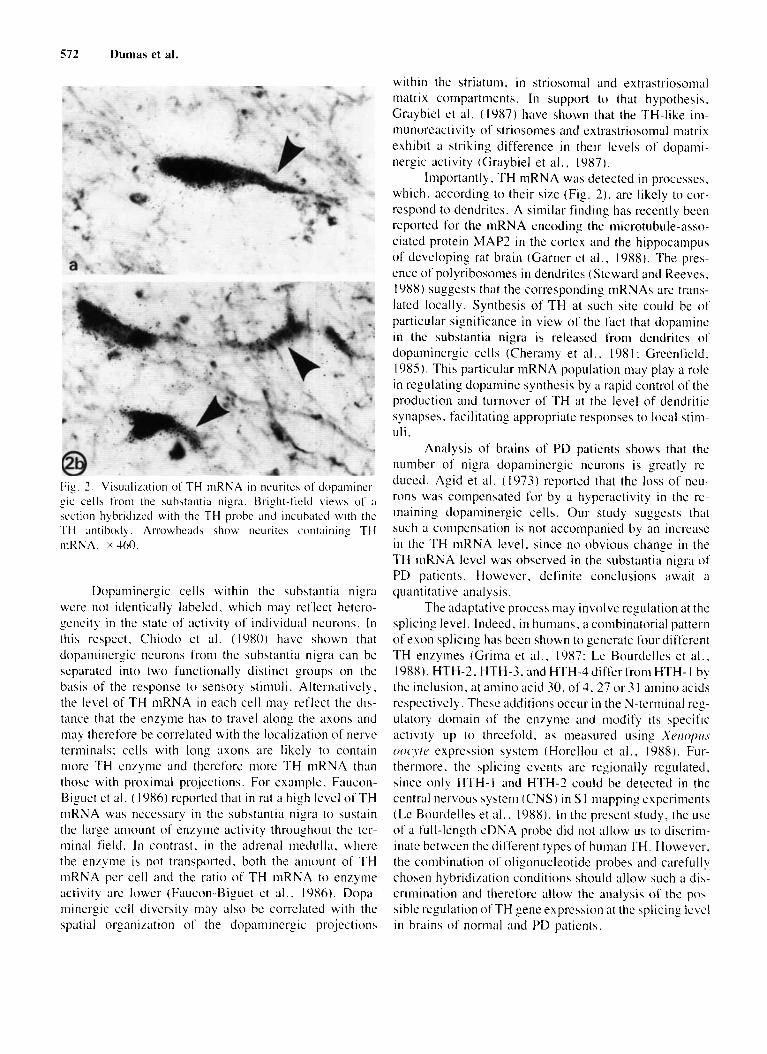
572 Dumas et al.
Fig. 7 . Visualization of TH rnRNA i n neurites o f dopaminrr- gic cell5 t'rom the substantia nigra. t3right-ficltl views of ;i
section hybridircd with thc TH probc ;rnd incubated with thc TH antibodq. Arrowheads show neuritc.; containing TH niKNA. x 460.
Dopaminergic cells within the substantia nigra "ere not identically labeled. which may reflect hetcro- geneity in the state of activity of individual neurons. In this respect. Chiodo et al. (1980) have shown that dopaniinergic neurons f'rom the substantia nigra can be separated into two functionally distinct groups on the basis o f the response t o sensory stimuli. Alternativclq, the level of TH niRNA in each cell may reflect the dis- tance that the enzyme has to travel along the iixons and may therefore be correlated with the loculization 01' nerve terminals: cells with long axons are likely to contain inore TH enzyme and therefore niore TH mRNA than those with proximal pro.jections. For example. Faucon- Biguet et al. (1986) reported that in rat a high level o f T H mRNA was necessary in the substantia nigra to sustain the large anicxtnt of enzyme activity throuyhout the ter- minal field. I n contrast. in the adrenal medulla, where the enzyme is not transported. both the amount of TH mRNA per cell and the ratio of TH mRNA t o enzyme activity are lower (Faticon-Biguet et al.. 19x6). Dopa- minergic cell diversity may also be correlated with the spatial organization 01' the dopaniinergic projections
within the striatum. in striosonial and extrastriosonial matrix compartments. I n support to that hypothesis. Craybiel et al. (1987) have shown that the 'TH-like i n - munoreactivity of striosomes and extrastriosomal matrix exhibit a striking difference in their levels of dopami- nergic activity (Graybiel et al.. 1987).
Importantly, TH mKNA was detected in processes. which, according to their size (Fig. 21, are likely to cor- respond to dendrites. A similar finding has recently been reported for the mRNA encodiny the microtubule-asso- ciated protein MAP? in the cortex and the hippocampus of developing rat brain (Garner et a l . , 1988). The pres- ence of polyribosomes in dendrites (Steward and Reeves, 1988) suggests that the corresponding tnRNAs are trans- lated locally. Synthesis o f TH at such site could be o f particular significance in view of the fact that dopamine in the substantia nigra is released from dendrites o f dopaminergic cells (Cheramy et a l . , I98 I : Greenfield. 1985). This particular niRNA population may play a role in regulating dopamine synthesis by a rapid control of the production and turnover of TH at the level of dendritic synapses. facilitating appropriate responses to local s t i n - uli .
Analysis of brains of PD patients shows that the number of nigra dopaniinergic ncurons is greatly rc- duccd. Agid et al . (1973) reported that the loss of ncu- rons was cornpensated for by a hyperactivity i n the rc- maining dopaminergic cells. Our study suggests that such a compensation is not accompanied by an increase i n the TH mRNA level. since n o obvious change in the TH niRNA level was observed in the substantia nigra o f PD patients. However. definite conclusions await ;I
quantitative analysis. The adaptative process may involve regulation at the
splicing level. Indeed. in humans. ;i combinatorial pattern of exon splicing has been shown to generate four different TH enzymes (Grima et al., 19x7; Le Bourdelles et al.. 1988). HTH-2. HTH-3. and HTH-3diflerfrom HTH- I by the inclusion. at amino acid 30. of3. 27 or 3 I amino acids respectively. These additions occur in the N-terniinal reg- ulatory domain of the enzyme atid modify its specific activity up to threefold. as measured using X P I I O ~ I I . ~ ooi'\/o expression system (Horellou et al.. 1988). Fur- thermore. the splicing events are regionally rcgulated. since o n l y HTH-I and HTH-7 could be detected in the central nervous system (CNS) in S 1 mapping experiments (Lc Bourdelles et a l . . 1988). In the present study, the use o f a full-length cDNA probc did not allow us to discrim- inate between the difl'erent types of human TH. However. the combination of oligonucleotide probes and carefully chosen hybridization conditions should allow such a dis- crimination and therefore allow the analysis o f the p o h -
sible regulation ofTH gene expression at the splicing l e ~ e l in brains o f normal and I'D patienls.
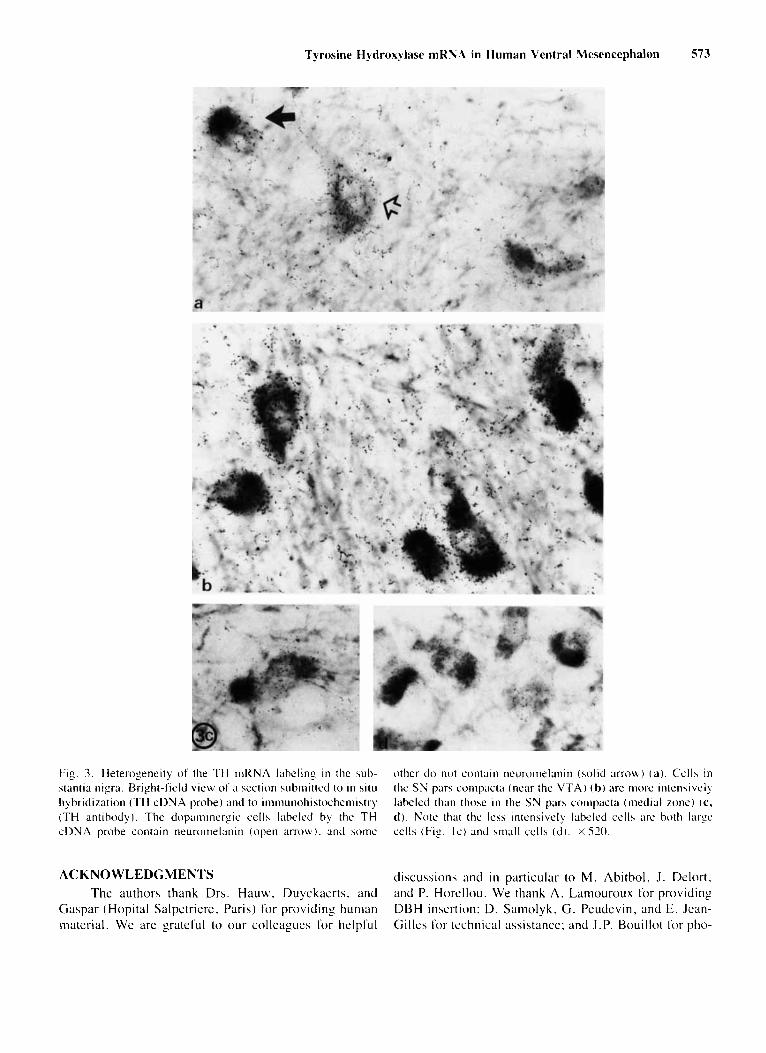
Tyrosine Hydroxylase mRNA in Human Ventral Mesencephalon 573
Fig. 3. Heterogeneity o f the TH mRNA labeling i n the sub- stantia nigra. Bright-field view of ;I section wbinitted to i n s i t i i hybridization (TH cDNA probe) antl to iiiiiiiuiioliiatochcmistry (TH antibody). The dopaminergic cells labeled by the TH c D N A probe contain neuroiiie1;inin (open a r r o w ) . antl acme
other do not contain neuronielanin (solid m o b ) ( a ) . Cclla i n the S N para coiiipact;i (near the V T A ) ( h ) are inow inknaively labeled than thwr in the SN pars compac(rr (iiieclial rone) ( c , d ) . Note that the leha intensively labeled cells are both large cells (Fig. Ic) and sinall cel ls ( d ) . X 520.
ACKNOWLEDGMENTS The authors thank Drs. Hauw. Duyckaerts. and
Gaspar (Hopital Salpetriere. Paris) for providing human material. We are grateful to our colleagues for helpful
discussions and i n particular t o M . Abitbol, J. Delort. and P. Horellou. We thank A . Laniouroux for providing DBH insertion; D. Samolyk. G. Peudevin, and E. Jean- Gilles for technical assistance: and J.P. Bouillot for pho-
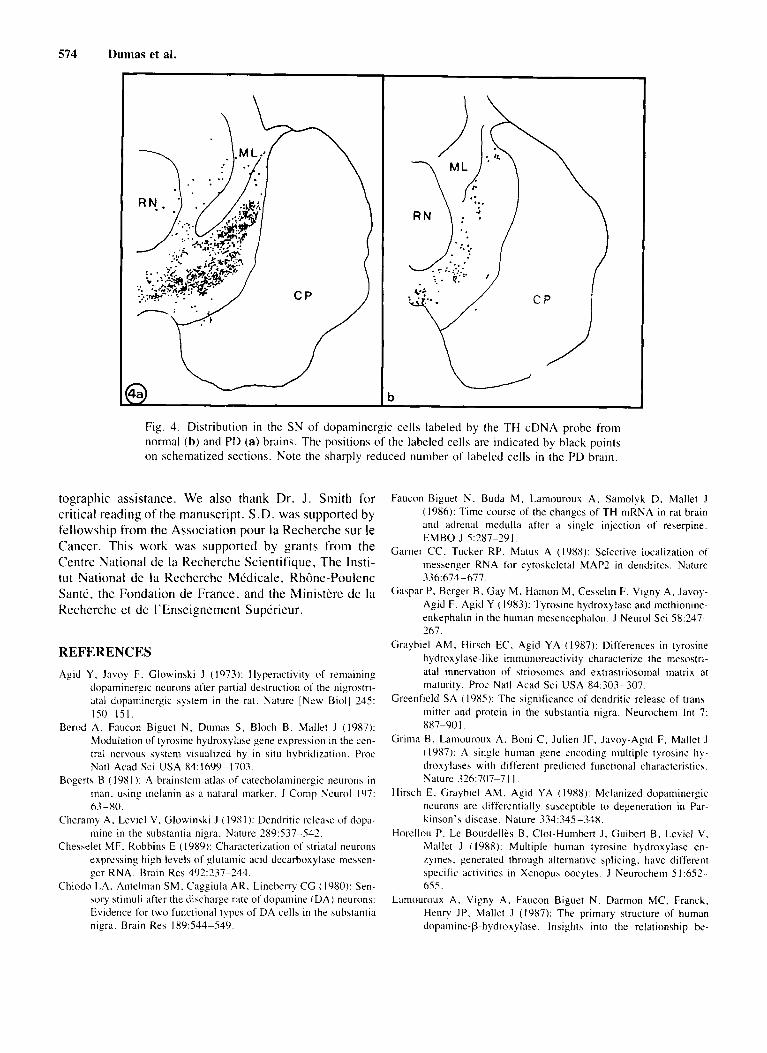
574 Dumas et al.
Fig. 4. Distribution in the S N of doparninergic cells labeled by the TH cDNA probe from norinal (b) and PD (a) brains. The positions of the labeled cells are indicated by black points on schematized sections. Note the sharply reduced number of labeled cells in the PD brain.
tographic assistance. We also thank Dr. J . Smith for critical reading of the manuscript. S.D. was supported by fellowship from the Association pour la Recherche sur le Canccr. This work was supported by grants from the Centre National de la Recherche Scientifique. The Insti- tu t National de la Recherche Medicale, Rh6ne-Poulenc Sante, the Fondation de France, and the Ministere de la Recherche et de I'Enseignement Superieur.
REFERENCES Agid Y , Javoy F. Glowinski J (1973): Hyperactivity of remaining
dopaminergic neurons alter partial destruction of the nigrostri- atal dopaminergic system in the rat. Nature [New Biol] 235: 150-15 I .
Berod A. Faucon Biguet N , Dumas S, Bloch B. Mallet J (19x7): Modulation of tyrosine hydroxylase gene expression in the cen- tral nervous system visualized by in situ hybridization. Proc Natl Acad Sci USA 84:1699-1703.
Bogerts B ( 198 1 ): A brainstem atlas of catecholaminergic neurons in man, using melanin as a natural marker. J Comp Ncurol 197: 63-80.
Cheraniy A. Leviel V. Glowinski J (19x1): Dendritic relea\e of dopa- mine in the substantia nigra. Nature 289:537-542.
Chesselet MF. Robbins E ( 19x9): Characterization of striatal neurons expressing high levels o f glutaniic acid dccarboxylaw inessen- ger RNA. Brain Res 492237-244.
Chiodo LA. Antelman SM. Caggiula AR. Lineberry CG (1980): Sen- mry stimuli after the discharge rate of dop;umine ( D A ) neurons: Evidence ior two functional types of DA cells in the aubstantia nigra. Brain Res 189:544-539.
Faucon Biguet N. Buda M. Lamouroux A. Samolyk D. Mallet J ( 1986): Time course of the changes of TH mRNA in rat brain and adrenal medulla after a single injection of reserpine. EMBO J 5:?87-?9l.
Garner CC, Tucker RP. Matus A (1988): Selective localization of messenger RNA tor cytoskeletal MAP2 in dendrites. Nature 336:674-677.
Ga\par P. Berger B. Gay M. Hamon M, Cesselin F. Vigny A, Javoy- Agid F. Agid Y ( 19x3): Tyrosine hydroxylase and methionine- enkephalin i n the human mesencephalon. J Neurol Sci 58:247- 267.
Graybiel AM. Hirsch EC. Agid Y A (1987): Dilferences in tyrosine hydroxylase-like immunoreactivity characterize the mesostri- atal innervation of striosumes and extrastriosomal matrix at maturity. Proc Natl Acad Sci USA 84:303-307.
Greenfield SA ( 10x5): The significance of dendritic release of trans- mitter and protein i n the substantia nigra. Neurocheni Int 7: X87-90 I .
Grima B, Lamouroux A . Boni C. Julien JF. Javoy-Agid F. Mallet J ( 19x7): A .;inglc human gene encoding multiple tyrosine hy- droxylases with different predicted fiinctional characteristics. Nature 326:707-7 I I .
Hirsch E. Graybiel AM, Agid YA ( 1988): Mclanized dopaminergic neurons are differentially susceptible t o degeneration in Par- kinson's disease. Nature 334:345-348.
Horellou P, Le Bourdelles B, Clot-Humbert J. Guibert B. Leviel V. Mallet J ( 1988): Multiple human tyrosine hydroxylase en- ryines. generated through alternative splicing. have different specific activities i n Xenopus oocytes. J Neurochem 5 I :652- 655.
L L ~ I T I O U ~ O L I ~ A. Vigny A. Faucon Biguet N. Darmon MC, Franch. Henry JP. Mallet J (19x7): The primary structure of human dopamine-P-hydro~ylase. Insights into the relationship be-

Tyrosine Hydroxylase mRNA in Human Ventral Mesencephalon 575
tween the soluble and the membrane-bound torins o f the en- zyme.
Le Bourdelles B, Boularand S. Boni C. Horellou P. Dumas S. Grima B. Mallet J ( 1988): Analysis of the 5' region of human tyrosine hydroxylase gene: Combinatorial patterns olcxon splicing gcn- erate multiple regulated tyrosine hydroxylasc isoforma. J Neu- rochcni 50:988-991 .
Mugnaini E. Oertel WH (198.5): An atlas 0 1 the distribution of GABAergic neuron5 and terminals in the rat CNS ;IS revealed by C A D imiiiunohistochemistry. In Bjorhlund A. Hokfelt T (eds): "Handbook of Chemical Neuroanatomy. Vol 4: CABA and Neuropeptides in the CNS, Part I . * ' Amsterdam: Elsevier Science Puhlishera. pp 436-608.
Riche D. de Ponimery J . Menetrey D (1989): Neuropeptides and cat-
echolamines in efferent projections of the nuclei 01 thc solitary tract in the rat. J Coinp Neurol (in press).
Rigby PWJ. Dieckinann M. Rhodes C, Berg P (1Y77): Labcling de- oxyribonucleic acid to high specific activity i n vitro by nick translation with DNA polymerase I . J Mol Biol l13:237-1-51.
Schalling M , Stiep PE, Lindquist C. Goldstein M . Hokfelt T lY89): Rapid increasc in enzyme and peptide niRNA in sy mpathetlc ganglia after electrical stimulation i n humans. Proc Nat l Acad Sci USA X6:4302-4305.
Stcwarcl 0, Reeves T,M ( 1988): Protein-synthetic niachincry beneath postsynaptic sites on CNS neurons: Association between poly- ribosomes and other organelles at the hynaptic site. J Neurusci 8: 176-184.