university of groningen heat resistance of bacillus spores … · 2016-05-23 · the formation of a...
TRANSCRIPT

University of Groningen
Heat resistance of Bacillus sporesBerendsen, Erwin Mathijs
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2016
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Berendsen, E. M. (2016). Heat resistance of Bacillus spores: Natural variation and genomic adaptation.[Groningen]: Rijksuniversiteit Groningen.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 29-04-2020

General introduction
Wells-Bennik MHJ, Eijlander RT, den Besten HMW, Berendsen EM, Warda AK, Krawczyk AO, Nierop Groot MN, Xiao Y, Zwietering MH, Kuipers OP, Abee T (2016). Bacterial spores in food: survival, emergence, and outgrowth. Annual Review of Food Science and Technology 7:457-482.
Chapter 1
Erwin M. Berendsen
Partly based on the review published as:

8
Chapter 1
EndosporesBacterial spores are widely present in nature. The resistance properties of spores allow for survival against environmental insults. A very important property of spores is their ability to withstand high temperatures. The level of resistance of spores to heating can vary between different spore forming species, but can also vary between strains. The mechanisms underlying strain specific variation in heat resistance of spores are not well understood. Understanding which factors have a major impact on heat resistance of spores can have important implications for their control in food and in applications to improve health. The focus of this thesis is on establishing variation in heat resistance of spores that exists between strains, and on the identification of genomic determinants that underlie this variation.
Spore formers from the Clostridiales and Bacillales orders have the extraordinary ability to enter sporulation as an adaptive strategy to survive conditions encountered in their natural habitat, for instance in soil, aquatic environments or in the gut of insects and animals (23, 49, 69, 77). This complex regulatory process transforms the bacterial cell into a dormant endospore (3, 38, 50, 51, 108), a state in which it can reside for undefined periods of time. The isolation of spores have been reported for a strain of Bacillus sphaericus from 25 to 40 million year old Dominican amber (22), and a Bacillus sp. 2-9-3 from a salt crystal of 250 million years old (118). Although these reports were met with scepticism about potential external contamination, they illustrate the potential extent of longevity of spores. Dormant endospores are resistant to environmental stress conditions including heat, salinity, acidity, radiation, oxygen and/or water depletion or low availability of nutrients (71, 77, 103).
The sporulation process is induced by high cell densities and nutrient limitation, and has been extensively studies in Bacillus subtilis strain 168, which is a gram positive model organism (38, 51). The availability of the genome sequence of B. subtilis 168 and the ability to genetically amend this strain greatly facilitated the progress to understand sporulation and spore resistance mechanisms (11, 55). The sporulation process is a unique developmental pathway that is fundamentally different from binary fission, and involves differentiation, intercellular signaling and programmed cell death among others (39).
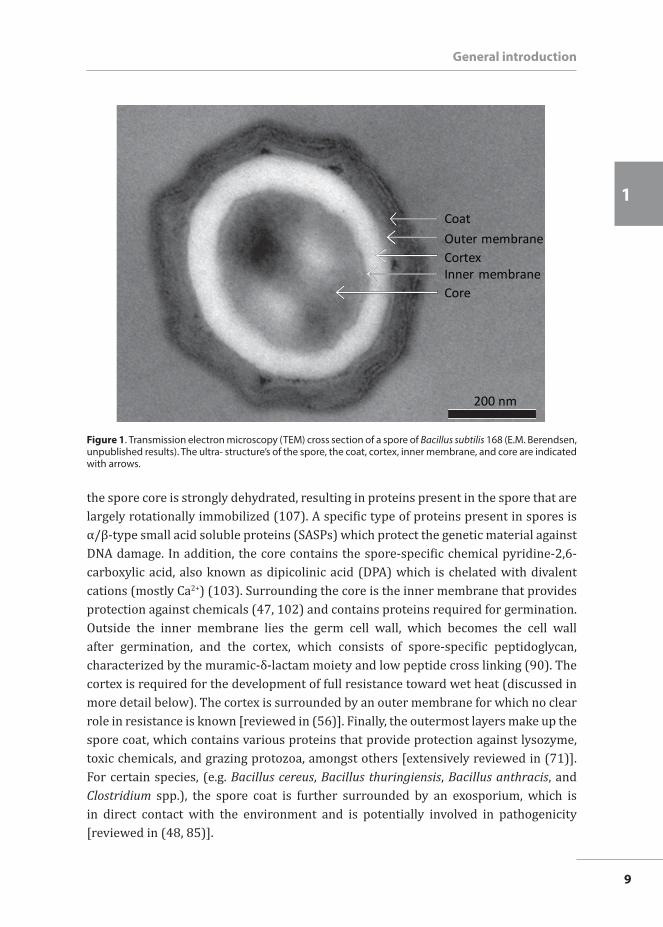
The composition of a spore Bacterial spores have various layered structures (Figure 1) that provide resistance against environmental insults and thus against several food processing conditions. The way in which different structures contribute to spore resistance has been extensively reviewed elsewhere (56, 103) and will be only briefly discussed here. First and foremost,
General introduction
9
1
the spore core is strongly dehydrated, resulting in proteins present in the spore that are largely rotationally immobilized (107). A specific type of proteins present in spores is α/β-type small acid soluble proteins (SASPs) which protect the genetic material against DNA damage. In addition, the core contains the spore-specific chemical pyridine-2,6-carboxylic acid, also known as dipicolinic acid (DPA) which is chelated with divalent cations (mostly Ca2+) (103). Surrounding the core is the inner membrane that provides protection against chemicals (47, 102) and contains proteins required for germination. Outside the inner membrane lies the germ cell wall, which becomes the cell wall after germination, and the cortex, which consists of spore-specific peptidoglycan, characterized by the muramic-δ-lactam moiety and low peptide cross linking (90). The cortex is required for the development of full resistance toward wet heat (discussed in more detail below). The cortex is surrounded by an outer membrane for which no clear role in resistance is known [reviewed in (56)]. Finally, the outermost layers make up the spore coat, which contains various proteins that provide protection against lysozyme, toxic chemicals, and grazing protozoa, amongst others [extensively reviewed in (71)]. For certain species, (e.g. Bacillus cereus, Bacillus thuringiensis, Bacillus anthracis, and Clostridium spp.), the spore coat is further surrounded by an exosporium, which is in direct contact with the environment and is potentially involved in pathogenicity [reviewed in (48, 85)].
Cortex
CoreInner membrane
Outer membraneCoat
Cortex
CoreInner membrane
Outer membraneCoat
200 nm
Figure 1. Transmission electron microscopy (TEM) cross section of a spore of Bacillus subtilis 168 (E.M. Berendsen, unpublished results). The ultra- structure’s of the spore, the coat, cortex, inner membrane, and core are indicated with arrows.

10
Chapter 1
Sporulation and germination The spore properties, that are at the basis of resistance and dormancy, are largely determined by the sporulation process. To exit the dormant state, spores undergo a germination process. Proteins that are required for germination are already produced during the sporulation process. The sporulation and germination process will be discussed below.
The sporulation process
The formation of a bacterial endospore is the result of a complex regulatory process that has been extensively reviewed elsewhere (3, 38, 50, 51, 108). The sporulation process involves a series of steps that ultimately lead to the formation of a dormant spore. The point at which a vegetative cell of B. subtilis enters the sporulation process is determined by nutrient limitation and high cell densities. The key sporulation regulator is Spo0A, which requires to be phosphorylated and to form a dimer for initiation of sporulation (19, 59). In B. subtilis the phosphorylation of Spo0A is mediated via Spo0F and Spo0B and involves several histidine kinases KinA, KinB, KinC, KinD, and KinE, which all respond to different environmental stimuli (39). The phosphorylation of Spo0A is indirect, therefore this system is referred to as the phosphorelay system (19). Upon reaching a certain threshold level of phosporylated Spo0A (Spo0A~P) in the cell, an auto stimulatory positive feedback loop is established, initiating sporulation by the expression of multiple genes (27).
The sporulation process is tightly controlled by a gene regulatory network, which has been extensively reviewed elsewhere, and therefore will only be discussed briefly here (38, 51). The spore is formed within the vegetative cell (mother cell) and after lysis of this cell is released into the environment. At the basis of the sporulation process are the RNA polymerase sigma factors that change the promoter specificity of the Bacillus RNA polymerase. A strict temporal regulation between the forespore and the mother cell compartments is mediated by the sporulation specific sigma factors σF, σE, σG and σK (38, 51). The sigma factors σF and σG are forespore specific, whereas σE and σK are mother cell specific (38, 51). The sporulation process can be divided in seven different stages as presented in Figure 2 (51). At stage 0, the cell is committed to sporulation and the DNA becomes more dense, followed by axial filament formation at stage I, during which genes are regulated by σA and σH. During stage II, an asymmetric division of the vegetative cell takes place, and a septum divides the vegetative cell into a smaller forespore compartment and a larger mother cell compartment. The gene regulation during stage II is mediated by σE in the mother cell and by σF in the forespore. The forespore is subsequently engulfed by the membrane of the mother cell during stage III, while gene expression is regulated by σG and σK, in the forespore and mother cell,

General introduction
11
1
respectively. Between the two membrane layers the spore cortex is built up, followed by the dehydration of the spore core (stage IV). During stage V, the SASPs are synthesized in the forespore. Additionally, DPA is synthesized in the mother cell by SpoVFA and SpoVFB and subsequently incorporated into the spore core by proteins encoded on the spoVA operon (111). Finally, a number of protein coat layers are formed, and after maturation of the spore the mother cell lyses, whereupon the spore is released into the environment. The spores can exit the dormant state via the process of germination, which will be discussed in the following section.
The germination process
Spores can exit dormancy via the process of germination followed by outgrowth into a vegetative cell. The germination of spores can be triggered by different means, namely by nutrients and by non-nutrients triggers. The germination process has been extensively reviewed (84, 101, 122) and will be briefly discussed here. The germination process consists of two different stages (Figure 2) (102). Prior to the first stage, the
Sporulation process Germination process
Stage VIIDormant spore
Activation
Stage ICation release DPA releasePartal core hydration
Stage IICortex hydrolysis Core hydration
OutgrowthResume metabolism SASP degradation
Stage IAxial filament formation
Stage IIAssymetric division
Stage IIIEngulfment
Stage IVCore dehydrationCortex synthesis
Stage VSASPs productionDPA incorporation Coat layers
Stage VICell lysis
Stage 0DNA more dense
σA σH
σE σF
σKσG
Figure 2. Overview of the sporulation and germination process of B. subtilis 168. The sporulation process can be divided in seven stages, indicated in the figure. At the onset stage of sporulation, asymmetric cell division occurs, leading to formation of a mother cell and a forespore compartment. This is followed by a commitment stage. After engulfment of the forespore, a maturation state allows for completion of sporulation. Upon completion of sporulation, the mother cell lyses and the spore is released into the environment. If conditions are favorable, germination may occur, that can be followed by outgrowth into a vegetative cell. The germination process is divided in an activation step, stage I, stage II and outgrowth.

12
Chapter 1
spore is activated to germinate. During the first stage, monovalent cations and Ca-DPA are released from the spore core, which in turn is partly rehydrated, thereby losing a phase bright appearance using phase contrast microscopic imaging. During the second stage of germination, the cortex is hydrolyzed by cortex lytic enzymes and the core is hydrated further. During the outgrowth stage, the metabolic activity is resumed, SASPs are degraded and the spore exits from the coat layers.
Nutrient triggers, like sugars, nucleosides, salts and amino acids can activate the germination receptors (GR) that are present in the inner membrane of the spore. For B. subtilis 168, the tricistronic operons gerA, gerK and gerB encode three distinct germination receptor complexes. Additionally, the genome of B. subtilis 168 contains two cryptic GR operons for which no role in germination has been established (6). The proteins GerAA and GerAC of the GerA complex, and GerBA of the GerB complex were shown to be localized to the inner membrane (52, 83). Additionally, there is GerD, which is required for germination and also localizes to the inner membrane (87, 88). The GRs, GerD, but not SpoVA proteins cluster together in the inner membrane and are called “germinosome” (101). Recently, an alternative pathway involved in spore germination was discovered, involving a serine/threonine kinase PrkC that is induced by peptidoglycan fragments of vegetative cells (104).
During germination, the release of Ca-DPA from the spore core is mediated by the SpoVA proteins (117). However, the signaling from the germination receptors to the SpoVA proteins is not known, and remains to be elucidated. The protein SpoVAC acts as a mechanosensitive channel and is required for the uptake and release of Ca-DPA (116). The protein SpoVAD specifically binds to Ca-DPA (62). During germination of B. subtilis, the spore cortex is hydrolyzed by two cortex lytic enzymes, namely CwlJ and SleB. Mutant strains lacking both sleB and cwlJ can proceed through the first stage of germination but are impaired in the second stage of germination. As a result, germinated spores do not succeed to grow out (16, 26). The proper localization of SleB is mediated by YpeB, while CwlJ requires the presence of GerQ (ywdL) for correct localization (16, 92).
Non-nutrient germination triggers can be exogenous DPA, dodecylamine, or the application of a high temperature or high hydrostatic pressure (HHP) (84). Exogenous DPA directly activates the cortex lytic enzyme CwlJ, resulting in degradation of the cortex. The surfactant dodecylamine directly triggers the release of DPA from the spore core (117), thereby initiating degradation of the cortex by the cortex lytic enzymes. HHP can be used to trigger germination via the GRs at pressures around 150 MPa, and by opening of the SpoVA channels at moderate pressures of 500MPa or higher (15, 93).

General introduction
13
1
Genomics of spore formers
Genomic information of the sporulation process is largely based on the model organism B. subtilis 168 (11, 38, 55, 78). Recent advances have been in made in genomics of bacterial spore formers, extending the genomic knowledge from B. subtilis 168 to other spore formers (30). For example, Galperin et al. (2012) made a thorough comparison of genomes of spore formers, aiming at identifying a minimal set of genes required for sporulation (41). This genomic analysis revealed that Bacilli and Clostridia share a core set of genes required for sporulation, but variation was seen between both classes with regard to gene presence and absence (41). A detailed comparison of 20 Bacillus spp. genomes showed that sporulation genes are divergent among the species, with specific gene sets for the B. subtilis group, specific genes missing in the B. cereus group, and genes missing in the group of thermomophilic spore formers (4). The sporulation genes involved in gene regulation like spo0A sigE, sigF, sigG, and sigK were found to belong to the core sporulation genes (4, 41). In addition, genes were conserved for which the encoded proteins have different roles during the sporulation process, such as the spore maturation proteins spmA and spmB, DPA synthesis genes spoVFA and spoVFB and the SASP sspI (4). A major divergence in sporulation genes was observed in sensory kinases, coat proteins, and SASPs (4). Similarly, a study by Earl et al. (2007) used a microarray based genomic hybridization approach (M-CGH) to assess seventeen strains of the B. subtilis group, and revealed that more than half of the variable genes in sporulation encoded proteins that are part of the spore coat (36). It should be noted that the genomic variations observed in the above mentioned studies were not linked to strain-specific phenotypic differences of spores.
Spores in foodSpore forming bacteria play an important role in food spoilage and foodborne disease, and food industries actively employ strategies to ensure adequate inactivation of spores and to control outgrowth. The resistance properties of spores lay at the heart of their ubiquitous presence in the environment, and as a consequence, it is inevitable that spores enter into the food chain. Given the robustness of spores, they are generally quite resistant to processing and preservation treatments used in food manufacturing (25, 106). Yet, spores can sense changes in their direct surroundings (e.g. the availability of nutrients), which can trigger the process of germination (101). This may occur in foods because these are rich in nutrients. Once germination is initiated, the spore can return to its vegetative cell form and once again start exponential cell division. If spore germination occurs in a final food product, followed by growth of vegetative cells (possibly even followed by sporulation), this can lead to food spoilage. In the case of foodborne pathogens, foodborne illness may occur upon intake of foods that contain

14
Chapter 1
spores of pathogenic species that may germinate and grow in the gut, or upon intake of foods in which spores have already germinated and grown to high numbers. In the latter case, foodborne illness may be due to intake of toxin produced in the food (food poisoning) or intake of the pathogen that subsequently forms toxins in the gut, leading to diarrhea (foodborne infection) (8, 37, 65, 106).
Although the impact of pathogenic spore formers is apparent, non-pathogenic species also pose major challenges to the food industry. A plethora of spore formers – sometimes with very distinct characteristics with respect to spore resistance and/or requirements for growth – may be responsible for product defects or spoilage in a variety of food products, leading to substantial economic losses in the food chain and leading to substantial food waste (25, 91, 106).
Selective pressure for high-level heat resistant spores
High heat treatments are applied to produce commercially sterile foods with a long shelf life, such as sterilized canned foods (e.g. meats, fish, and vegetables), and ultra high temperature (UHT) treated liquids (e.g. juices and milk). In canned, bottled, or vacuum packed products with a pH high enough to allow for growth of C. botulinum, heat treatments must exceed 3 minutes at 121°C (or the equivalent) to inactivate spores of this organism (7). Highly heat resistant spores can survive such heat treatments, and depending on the product characteristics and storage temperatures during shelf life, different species may cause spoilage (65) (Figure 3). These may include Bacillus spp. and Paenibacillus spp. (20, 53, 96, 97), but also obligate thermophilic species that require temperatures greater than 45°C for growth, such as Geobacillus species (9, 20, 33). High-level heat resistant spores of the mesophilic spore formers of the B. subtilis group, which are focussed on in the work described in this thesis, are found in various food products, such as milk, bread, herbs and spices, soups, sauces and cocoa amongst others (63, 74, 81, 109, 110, 112). These mesophilic species generally have the ability to grow at temperatures up to 55°C, and for certain strains their spores are not heat inactivated by a heat treatment of 30 minutes at 100°C (81). The application of heating may introduce further heterogeneity in the potential of surviving spores to germination and outgrowth in food matrices (10, 37, 114). Anaerobic spore formers producing highly heat resistant spores may also play a role in spoilage of commercially sterile foods; the most heat resistant spores described to date are produced by Moorella thermoacetica (21) that belongs to the class Clostridia (phylum Firmicutes), which is naturally associated with anaerobic warm waters. The spores can easily survive sterilization, with a reported decimal reduction value (D-value) of 111 minutes at 121°C (21).

General introduction
15
1
Wet heat inactivation of spores
The composition of spores provides resistance against conditions that are commonly used in the food industry to reduce bacterial loads, such as heat, acid, salt and oxidising agents (see also (71, 103)). As indicated above, an important process to inactivate microbes is the application of heat.
Exact mechanisms that lead to inactivation of spores after application of wet heat are still not clear. When spore populations of B. subtilis were exposed to heat, two types of spores were recovered using buoyant density gradient centrifugation, namely spores that lost the internal DPA and suffered protein denaturation and a second fraction that consisted of spores did not lose internal DPA (28). Inactivated spores that still contained internal DPA were able to germinate, but they were impaired in outgrowth. This suggests that DPA release takes place after the heat inactivation and that proteins involved in outgrowth and resuming of metabolic activity are denatured (28). The denaturation of proteins during wet heat treatment was confirmed in spores of B. cereus and B. megaterium in which the outgrowth was also impaired (29). Later research indicated that proteins involved in germination, including the cortex lytic enzyme CwlJ, are damaged by wet heat treatment, leading to prolonged lag times of germination (119). Another suggested mode of action of wet heat is reduction of the ability to release CaDPA from the spore core due to protein damage.
Natural diversity
Selective pressuree.g. heating
Surviving spores Inactivated sporesInactivated cells
Figure 3. Overview of natural biodiversity of spore forming bacteria represented with different spores and cells. The application of a selective pressure (e.g. heating) selects for those spores that are able to survive the specific treatment. Studying these survivors might provide insight in adaptive mechanisms that allow for better survival of these spores compared to others.

16
Chapter 1
Wet heat application remains the most common treatment applied in the food industry to decrease bacterial spore loads from food ingredients and products. Quantification of spore resistance against wet heat and mechanisms that play a role in this resistance are further discussed in the next sections.
Recovery of spores
The resistance of spores towards wet heat treatment can be quantified by assessment of the surviving fraction expressed in colony forming units (CFU) obtained by plate counting. This quantification of spore survival and viability after heat treatments greatly depends on the ability of spores to germinate and grow out to a colony forming unit. This includes the recovery of spores that survive inactivation treatments but that sustained damage, requiring proper cultivation conditions to avoid underestimation of viable spore numbers after such treatments (57, 120)
The inactivation data obtained can be fitted using different models to accurately describe the inactivation curve (68). A common way to express the inactivation kinetics is the D-value, which is the time required to achieve a decimal reduction of the micro-organisms. In addition, the z-value is used, which is the increase in temperature required to achieve one extra decimal reduction. Alternatively, other methods can be employed to determine resistance properties of spores, that do not depend on CFU formation, like flow cytometry (70), measuring DPA release (54) and Raman spectroscopy (45). These techniques can provide more insights in individual spore behavior and heterogeneity in spore populations (37).
Known factors influencing spore heat resistance
Environmental factors influencing wet heat resistance of spores
It is well known that environmental factors during sporulation can influence properties of spores including the wet heat resistance (1, 58, 76). The temperature during the sporulation process has been reported to play a role in the wet heat resistance of the spores that were formed. Melly et al. (2002) reported that increased sporulation temperatures resulted in increased wet heat resistance of B. subtilis spores, which correlated with a lower core water content and a higher degree of muramic acid cross-linking in the cortex (72). Spores of B. subtilis prepared at 45°C survived heating at 90°C without showing any inactivation, whereas spores of the same strain prepared at 22°C showed three log reduction in viable spore counts (72). Similarly, the sporulation temperature was important for the final heat resistance properties of spores of B. weihenstephanensis, where a ten-fold increase spore wet heat resistance was observed when the sporulation temperature was increased from 5°C to 30°C (12). An increase

General introduction
17
1
in sporulation temperature from 20°C to 45°C also resulted in increased wet heat resistance (six-fold) of spores of B. licheniformis (12). Other reports describe increased wet heat resistance of B. subtilis and B. megaterium spores upon heat shock (45°C or 48°C for 30 minutes) during sporulation (73, 75, 100). It is not clear how this increase in resistance is mediated; heat shock proteins appear not to be involved in increased spore heat resistance despite their presence (73).
The presence of cations in the growth medium (in particular calcium, potassium, magnesium and manganese) can also contribute to an increased wet heat resistance of spores (24, 80). For spores of B. subtilis A163 prepared on sporulation medium with added salts (Ca2+
, Mn2+, K+, Mg2+), no inactivation of viable spore count was observed after heating at 114°C for 4 minutes, whereas for spores prepared without these salts in the sporulation medium, five log reduction in viable spore counts was observed (24). Interestingly, such effects may be species-specific, as Ghosh et al. (2011) observed an increased wet heat resistance of B. megaterium spores with higher concentrations of manganese in the sporulation medium (43), whereas Granger et al. (2011) did not see this effect for spores of B. subtilis (46).
To achieve maximum wet heat resistance, spores require a maturation period after release from the sporulating cell (95). The presence of divalent cations, like Ca2+, is required for this maturation; addition of the chelator EDTA abolished the maturation and thereby acquisition of maximal wet heat resistance (95). This maturation and subsequent increase in wet heat resistance is most likely mediated by chemical cross linking of proteins in the outer coat layer and crust (2).
In general, conditions encountered during sporulation in the natural habitat of spore formers undoubtedly play a role in resistance properties of spores that occur in foods and ingredients (49, 63, 115). However, often these conditions are not known or are hard to mimic in a laboratory setting. For example, spores of B. subtilis prepared on sporulation medium with agar were three fold more resistant than spores prepared in the same liquid medium (94). When preparing spores in a laboratory, conditions used to prepare spores (e.g. medium type, incubation temperatures, liquid broth or agar surface) can greatly influence spore wet heat resistance. This is a factor to be reckoned with, and such conditions may require optimization when studying the mechanisms that determine the wet heat resistance properties of spores.
Intrinsic factors influencing wet heat resistance of spores
Besides the effect of environmental conditions during sporulation on the final heat resistance properties, certain gene products are known to influence heat resistance properties of spores. Establishing the role of such genes relies mostly on the construction

18
Chapter 1
of deletion mutants. For example, mutations in the small acid soluble proteins (SASPs) are known to influence the final heat resistance properties of spores. In studies in which the small acid soluble protein encoded by ssp4 was removed from C. perfringens SM101, the decimal reduction times of spores at 100°C (D100°C) were reduced from 59.1 to 8.7 min, which is approximately a seven-fold reduction (60). In accordance with these findings, deletion of the α/β SASPs from B. subtilis led to a reduction of the D90°C of spores from 18 to 2.5 minutes, which is a seven-fold reduction (103). Deletion of certain genes can have a direct impact on the sporulation efficiency and on spore structures. For example, deletion of stoA from B. subtilis strongly impairs cortex synthesis, resulting in reduced sporulation efficiency and increasing the heat sensitivity of spores. Similarly, mutations in dacB, spmA and spmB which are involved in cortex synthesis, resulted in spores with up to eight-fold lower heat resistance levels than those of the parent strain (89). A complication of using mutants with deletions in genes that play a role in spore formation to subsequently study the wet heat resistance properties of spores, is that the composition of the spores might be severely affected. Additionally, the sporulation process itself might be blocked if genes required for the completion of sporulation are mutated, e.g. the mutations in the spoVA operon (111).
Apart from the above mentioned genetic factors, it is known that small subsets of spores that do not respond to known germination triggers (also termed superdormant spores) display an elevated wet heat resistance, which is likely caused by lower core water content than spores that respond normally to germination triggers (44). This can greatly influence the response of spores to environmental triggers. Likewise, spores of the industrial isolate B. subtilis A163 as well as B. sporothermodurans spores showed an increased wet heat resistance linked to increased dormancy when compared with the laboratory strain B. subtilis 168 (54). These observations demonstrate a strong correlation between spore heat resistance and dormancy. The consequence may be that spores with the highest level of heat resistance are the hardest to retrieve on cultivation media as a result of their impaired germination, possibly also leading to underestimation of their true numbers in foods.
Understanding variation in wet heat resistance of spores
Strain-specific variation in wet heat resistances of spores
Spores encountered in food ingredients and foods may have a large natural diversity, including variation in heat resistance properties. The optimal growth temperature of species often correlates with thermal resistance of their spores, with thermophilic species generally producing spores with higher thermoresistance than mesophilic species, which in turn produce spores with higher heat resistance than psychrotrophic

General introduction
19
1
species (42, 77, 121). However, this is not always the case. While marked differences in spore heat resistance are found between different species, remarkable difference can also be observed between different strains of the same species. For example, for B. cereus, strain variation was demonstrated to be a main variability factor for spore heat resistance (67). The same conclusion was drawn based on a meta analysis of heat resistances of spores of B. cereus (113). Within the B. subtilis group, large strain specific variation in heat resistance of spores has been reported in various studies (54, 63, 81). For spores of B. sporothermodurans, the heat resistances of spores varied between strains from a D100°C of 14 minutes to 800 minutes (97). Similarly, differences in heat resistance of spores were observed for strains of the anaerobic spore formers C. perfringens (60, 61, 82, 123) and C. botulinum (86).
In general, it is not clear how the strain specific differences in heat resistances of spores are mediated. Furthermore, as the mechanisms underlying these differences in resistance of spores are not clear, it is not known whether these adaptations are similar or different between strains and species. The use of comparative genomics can contribute to an improved understanding of mechanisms that play a role in determining spore heat resistance properties of different strains and species.
Genomics approach to understand variation in wet heat resistance of spores
Recently, the use of omics technology, including whole genome sequencing, in food safety gained more attention (5, 14, 18). Whole genome sequencing of Bacillus spp. strains can likely provide new insights in mechanisms of high-level heat resistance of spores (5, 66). Recently, more than 90 B. subtilis genomes have been sequenced and published (31, 35, 40, 98, 99, 105, 124, 125). These genome sequences provide a good basis for comparative genomic studies. Linking genomic variations to strain specific differences in heat resistance of spores can provide insight in genomic adaptations underlying high-level heat resistance of spores. A potential driver of genomic adaptation is horizontal gene transfer, and genetic material can be transferred between species and strains (79). The transfer of genetic material can take place via different mechanisms, such as phage transduction, conjugation and natural competence. Other processes underlying genomic variation, are mutations, genome reshuffling, insertion sequences and transposons, amongst others.
An approach that is promising for the identification of genes responsible for high-level wet heat resistance of spores is a so-called gene-trait matching, as described by Bayanov et al. (13) (Figure 4). The requirements for gene-trait matching were described previously by Dutilh et al. (34). The correlation of presence or absence of genes with a phenotype does not necessarily mean that there is a causative effect. To establish the role

20
Chapter 1
of a certain gene that is correlated with a phenotype, deletion or complementation of the specific gene is required. The gene-trait matching approach focuses on the presence and absence of genes in relation to phenotypes. More subtle changes in the genome, such as single nucleotide polymorphisms (SNP) or differences at transcriptional levels or translational levels will not easily be found using this approach. Alternatively, a transcriptome-trait matching could be employed, possibly revealing genes that are correlated to a given phenotype, based on different expression profiles (17, 32).
B. subtilis 168 is a well-studied model organism and provides a good basis for genomic comparisons with other B. subtilis strains that produce spores with different wet heat resistance properties. For example, Brul et al. (2011) performed a genomic comparison between B. subtilis 168 and B. subtilis A163 that produces spores with higher heat resistance properties, and DNA-DNA hybridization experiments revealed three genes, namely, xlyB, spsA, and wapA with stronger hybridization for B. subtilis A163 (18). It was not confirmed whether these genes play are role in the increased wet heat resistance of B. subtilis A163 spores (18). Using whole genome sequencing of B. subtilis strains M1
Single species- different strains
Phenotypic comparison
Genome comparison(Transcriptome comparison)
Gene unique for phenotype 1
Gene-trait matching(Transcriptome-trait matching)
Phenotype 1 Phenotype 2
A B C D EA B D E FA B C D E FA C D E F
Gene D
A B C D F
A B C EA B C E FA C E FA B C E FA B C E F
Figure 4. Gene-trait matching approach: theoretical example. Within a single species multiple strains are present. Upon testing a specific phenotype (e.g. heat resistance of spores) these strains may display different phenotypic behavior. Genome comparison reveals strain specific differences in genome content. Using a gene trait matching approach, in which phenotypic variation is matched with genotypic variation, genes correlating with a specific phenotype can be discovered. In this example gene D is always present in strains displaying phenotype 1, and always absent in strains with phenotype 2. Alternatively to genome comparison and gene-trait matching, transcriptome comparison and transcriptome can be used for identification of genes unique for a phenotype.

General introduction
21
1
and M112, which both produce spores with high heat resistance, and comparison with genes present in the B. subtilis 168 genome, Lima (64) reported genes uniquely present in strains M1 and M112, however no clear role for these genes in the increased resistance was hypothesized nor demonstrated (64). The above mentioned approaches could result in an understanding of high-level heat resistance of spores. The understanding of molecular processes underlying differences in strain specific resistance properties of spores will aid a better detection of highly heat resistant spore formers and allow for improved control of these spores.
Thesis outlineIn this thesis, variation in heat resistance of spores within the B. subtilis group has been investigated. Following detailed phenotypic analyses of spores (which showed significant differences between strains) and combining these data with genome analyses of the respective strains, breakthrough insights were obtained, uncovering genes that determine high-level heat resistance of spores and their germination potential.
To assess the variation in heat resistances of spores, spore heat inactivation kinetics were determined for fourteen strains of the B. subtilis group in Chapter 2. With respect to heat resistance of spores, two groups could be distinguished for the 14 strains of B. subtilis group. It was not known how this variation in heat resistance between strains within a species was mediated. To unravel the underlying cause of this difference in heat resistance of spores between B. subtilis strains, a gene trait-matching approach was used. This revealed a Tn1546 transposon that was correlated with the increased heat resistance of spores, as described in Chapter 3. It was demonstrated that the Tn1546 transposon is directly responsible for increased heat resistance of spores: transfer of the transposon resulted in increased heat resistance of spores. Additionally, genome mining for the spoVA operon in other spore forming Bacillus sp. was performed. The occurrence of different spoVA operons in 103 spore forming Bacillus sp. was studied, revealing the same genomic element in strains of other species, including certain strains of B. amyloliquefaciens and B. licheniformis. Spores of various strains of these species, with and without the element, were evaluated for high-level heat resistance in Chapter 4, confirming the role of Tn1546 transposon and spoVA operon in these species as well. Spores of highly heat resistant spore formers often have unpredictable germination behavior and heat resistance properties in foods. These properties were assessed for spores of four B. thermoamylovorans strains in Chapter 5. Overall, these strains showed very limited germination on standard rich media due to poor nutrient-induced germination, leading to strong underestimations of the viable spore numbers and the spore heat resistances. The impact of the findings from Chapters 2, 3, 4, and 5 is discussed in Chapter 6 together with perspectives for the food industry.

22
Chapter 1
References
1. Abbas, A. A., S. Planchon, M. Jobin, and P. Schmitt. 2014. Absence of oxygen affects the capacity to sporulate and the spore properties of Bacillus cereus. Food Microbiology 42:122-131.
2. Abhyankar, W., R. Pandey, A. Ter Beek, S. Brul, L. J. de Koning, and C. G. de Koster. 2015. Reinforcement of Bacillus subtilis spores by cross-linking of outer coat proteins during maturation. Food Microbiology 45, Part A:54-62.
3. Al-Hinai, M. A., S. W. Jones, and E. T. Papoutsakis. 2015. The Clostridium sporulation programs: Diversity and preservation of endospore differentiation. Microbiology and Molecular Biology Reviews 79:19-37.
4. Alcaraz, L., G. Moreno-Hagelsieb, L. Eguiarte, V. Souza, L. Herrera-Estrella, and G. Olmedo. 2010. Understanding the evolutionary relationships and major traits of Bacillus through comparative genomics. BMC Genomics 11:332.
5. Alkema, W., J. Boekhorst, M. Wels, and S. A. van Hijum. 2015. Microbial bioinformatics for food safety and production. Briefings in Bioinformatics.
6. Alzahrani, O. M., and A. Moir. 2014. Spore germination and germinant receptor genes in wild strains of Bacillus subtilis. Journal of Applied Microbiology 117:741-9.
7. Anderson, N. M., J. W. Larkin, M. B. Cole, G. E. Skinner, R. C. Whiting, L. G. M. Gorris, A. Rodriguez, R. Buchanan, C. M. Stewart, J. H. Hanlin, L. Keener, and P. A. Hall. 2011. Food safety objective approach for controlling Clostridium botulinum growth and toxin production in commercially sterile foods. Journal of Food Protection 74:1956-1989.
8. Andersson, A., U. Rönner, and P. E. Granum. 1995. What problems does the food industry have with the spore-forming pathogens Bacillus cereus and Clostridium perfringens? International Journal of Food Microbiology 28:145-155.
9. Andre, S., F. Zuber, and F. Remize. 2013. Thermophilic spore-forming bacteria isolated from spoiled canned food and their heat resistance. Results of a French ten-year survey. International Journal of Food Microbiology 165:134-43.
10. Augustin, J.-C. 2011. Challenges in risk assessment and predictive microbiology of foodborne spore-forming bacteria. Food Microbiology 28:209-213.
11. Barbe, V., S. Cruveiller, F. Kunst, P. Lenoble, G. Meurice, A. Sekowska, D. Vallenet, T. Wang, I. Moszer, C. Medigue, and A. Danchin. 2009. From a consortium sequence to a unified sequence: the Bacillus subtilis 168 reference genome a decade later. Microbiology 155:1758-75.
12. Baril, E., L. Coroller, O. Couvert, I. Leguérinel, F. Postollec, C. Boulais, F. Carlin, and P. Mafart. 2012. Modeling heat resistance of Bacillus weihenstephanensis and Bacillus licheniformis spores as function of sporulation temperature and pH. Food Microbiology 30:29-36.
13. Bayjanov, J. R., D. Molenaar, V. Tzeneva, R. J. Siezen, and S. A. van Hijum. 2012. PhenoLink--a web-tool for linking phenotype to ~omics data for bacteria: application to gene-trait matching for Lactobacillus plantarum strains. BMC Genomics 13:170.
14. Bergholz, T. M., A. I. Moreno Switt, and M. Wiedmann. 2014. Omics approaches in food safety: fulfilling the promise? Trends in Microbiology 22:275-281.
15. Black, E. P., K. Koziol-Dube, D. Guan, J. Wei, B. Setlow, D. E. Cortezzo, D. G. Hoover, and P. Setlow. 2005. Factors influencing germination of Bacillus subtilis spores via activation of nutrient receptors by high pressure. Applied and Environmental Microbiology 71:5879-5887.
16. Boland, F. M., A. Atrih, H. Chirakkal, S. J. Foster, and A. Moir. 2000. Complete spore-cortex hydrolysis during germination of Bacillus subtilis 168 requires SleB and YpeB. Microbiology 146:57-64.
17. Bron, P. A., M. Wels, R. S. Bongers, H. van Bokhorst-van de Veen, A. Wiersma, L. Overmars, M. L. Marco, and M. Kleerebezem. 2012. Transcriptomes reveal genetic signatures underlying physiological variations imposed by different fermentation conditions in Lactobacillus plantarum. PLoS ONE 7:e38720.
18. Brul, S., J. van Beilen, M. Caspers, A. O’Brien, C. de Koster, S. Oomes, J. Smelt, R. Kort, and A. Ter Beek. 2011. Challenges and advances in systems biology analysis of Bacillus spore physiology; molecular differences between an extreme heat resistant spore forming Bacillus subtilis food isolate and a laboratory strain. Food Microbiology 28:221-227.

General introduction
23
1
19. Burbulys, D., K. A. Trach, and J. A. Hoch. 1991. Initiation of sporulation in B. subtilis is controlled by a multicomponent phosphorelay. Cell 64:545-552.
20. Burgess, S. A., D. Lindsay, and S. H. Flint. 2010. Thermophilic bacilli and their importance in dairy processing. International Journal of Food Microbiology 144:215-225.
21. Byrer, D. E., F. A. Rainey, and J. Wiegel. 2000. Novel strains of Moorella thermoacetica form unusually heat-resistant spores. Archives of Microbiology 174:334-339.
22. Cano, R., and M. Borucki. 1995. Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber. Science 268:1060-1064.
23. Carlin, F. 2011. Origin of bacterial spores contaminating foods. Food Microbiology 28:177-82.24. Cazemier, A. E., S. F. M. Wagenaars, and P. F. Ter Steeg. 2001. Effect of sporulation and recovery
medium on the heat resistance and amount of injury of spores from spoilage bacilli. Journal of Applied Microbiology 90:761-770.
25. Checinska, A., A. Paszczynski, and M. Burbank. 2015. Bacillus and other spore-forming genera: Variations in responses and mechanisms for survival. Annual Review of Food Science and Technology 6:351-369.
26. Chirakkal, H., M. O’Rourke, A. Atrih, S. J. Foster, and A. Moir. 2002. Analysis of spore cortex lytic enzymes and related proteins in Bacillus subtilis endospore germination. Microbiology 148:2383-2392.
27. Chung, J. D., G. Stephanopoulos, K. Ireton, and A. D. Grossman. 1994. Gene expression in single cells of Bacillus subtilis: evidence that a threshold mechanism controls the initiation of sporulation. Journal of Bacteriology 176:1977-1984.
28. Coleman, W. H., D. Chen, Y.-q. Li, A. E. Cowan, and P. Setlow. 2007. How moist heat kills spores of Bacillus subtilis. Journal of Bacteriology 189:8458-8466.
29. Coleman, W. H., P. Zhang, Y. q. Li, and P. Setlow. 2010. Mechanism of killing of spores of Bacillus cereus and Bacillus megaterium by wet heat. Letters in Applied Microbiology 50:507-514.
30. de Hoon, M. J. L., P. Eichenberger, and D. Vitkup. 2010. Hierarchical Evolution of the Bacterial Sporulation Network. Current biology : CB 20:R735-R745.
31. Deng, Y., Y. Zhu, P. Wang, L. Zhu, J. Zheng, R. Li, L. Ruan, D. Peng, and M. Sun. 2011. Complete genome sequence of Bacillus subtilis BSn5, an endophytic bacterium of Amorphophallus konjac with antimicrobial activity for the plant pathogen Erwinia carotovora subsp. carotovora. Journal of Bacteriology 193:2070-1.
32. Dijkstra, A., W. Alkema, M. Starrenburg, J. Hugenholtz, S. van Hijum, and P. Bron. 2014. Fermentation-induced variation in heat and oxidative stress phenotypes of Lactococcus lactis MG1363 reveals transcriptome signatures for robustness. Microbial Cell Factories 13:148.
33. Durand, L., S. Planchon, M.-H. Guinebretiere, F. Carlin, and F. Remize. 2015. Genotypic and phenotypic characterization of foodborne Geobacillus stearothermophilus. Food Microbiology 45, Part A:103-110.
34. Dutilh, B. E., L. Backus, R. A. Edwards, M. Wels, J. R. Bayjanov, and S. A. F. T. van Hijum. 2013. Explaining microbial phenotypes on a genomic scale: GWAS for microbes. Briefings in Functional Genomics 12:366-380.
35. Earl, A. M., M. Eppinger, W. F. Fricke, M. J. Rosovitz, D. A. Rasko, S. Daugherty, R. Losick, R. Kolter, and J. Ravel. 2012. Whole-genome sequences of Bacillus subtilis and close relatives. Journal of Bacteriology 194:2378-9.
36. Earl, A. M., R. Losick, and R. Kolter. 2007. Bacillus subtilis genome diversity. Journal of Bacteriology 189:1163-1170.
37. Eijlander, R. T., T. Abee, and O. P. Kuipers. 2011. Bacterial spores in food: how phenotypic variability complicates prediction of spore properties and bacterial behavior. Current Opinion in Biotechnology 22:180-186.
38. Eijlander, R. T., A. de Jong, A. O. Krawczyk, S. Holsappel, and O. P. Kuipers. 2014. SporeWeb: an interactive journey through the complete sporulation cycle of Bacillus subtilis. Nucleic Acids Research 42:D685-91.
39. Errington, J. 2003. Regulation of endospore formation in Bacillus subtilis. Nature Reviews Microbiology 1:117-126.
40. Fan, L., S. Bo, H. Chen, W. Ye, K. Kleinschmidt, H. I. Baumann, J. F. Imhoff, M. Kleine, and D. Cai. 2011. Genome sequence of Bacillus subtilis subsp. spizizenii gtP20b, isolated from the Indian ocean.

24
Chapter 1
Journal of Bacteriology 193:1276-1277.41. Galperin, M. Y., S. L. Mekhedov, P. Puigbo, S. Smirnov, Y. I. Wolf, and D. J. Rigden. 2012. Genomic
determinants of sporulation in Bacilli and Clostridia: towards the minimal set of sporulation-specific genes. Environmental Microbiology 14:2870-90.
42. Gerhardt, P., and R. E. Marquis. 1989. Spore thermoresistance mechanisms, p. 17-63. In I. Smith, R. A. Slepecky, and P. Setlow (ed.), Regulation of prokaryotic development. American society for microbiology, Washington, D.C.
43. Ghosh, S., A. Ramirez-Peralta, E. Gaidamakova, P. Zhang, Y. Q. Li, M. J. Daly, and P. Setlow. 2011. Effects of Mn levels on resistance of Bacillus megaterium spores to heat, radiation and hydrogen peroxide. Journal of Applied Microbiology 111:663-670.
44. Ghosh, S., and P. Setlow. 2009. Isolation and characterization of superdormant spores of Bacillus species. Journal of Bacteriology 191:1787-1797.
45. Ghosh, S., P. Zhang, Y.-q. Li, and P. Setlow. 2009. Superdormant spores of Bacillus species have elevated wet-heat resistance and temperature requirements for heat activation. Journal of Bacteriology 191:5584-5591.
46. Granger, A. C., E. K. Gaidamakova, V. Y. Matrosova, M. J. Daly, and P. Setlow. 2011. Effects of Mn and Fe levels on Bacillus subtilis spore resistance and dffects of Mn2+, other divalent cations, orthophosphate, and dipicolinic acid on protein resistance to ionizing radiation. Applied and Environmental Microbiology 77:32-40.
47. Griffiths, K. K., and P. Setlow. 2009. Effects of modification of membrane lipid composition on Bacillus subtilis sporulation and spore properties. Journal of Applied Microbiology 106:2064-78.
48. Henriques, A. O., and J. Moran, Charles P. 2007. Structure, assembly, and function of the spore surface layers. Annual Review of Microbiology 61:555-588.
49. Heyndrickx, M. 2011. The importance of endospore-forming bacteria originating from soil for contamination of industrial food processing. Applied and Environmental Soil Science 2011:11.
50. Higgins, D., and J. Dworkin. 2012. Recent progress in Bacillus subtilis sporulation. FEMS Microbiology Reviews 36:131-148.
51. Hilbert, D. W., and P. J. Piggot. 2004. Compartmentalization of gene expression during Bacillus subtilis spore formation. Microbiology and Molecular Biology Reviews 68:234-262.
52. Hudson, K. D., B. M. Corfe, E. H. Kemp, I. M. Feavers, P. J. Coote, and A. Moir. 2001. Localization of GerAA and GerAC germination proteins in the Bacillus subtilis spore. Journal of Bacteriology 183:4317-4322.
53. Huemer, I. A., N. Klijn, H. W. J. Vogelsang, and L. P. M. Langeveld. 1998. Thermal death kinetics of spores of Bacillus sporothermodurans isolated from UHT milk. International Dairy Journal 8:851-855.
54. Kort, R., A. C. O’Brien, I. H. M. van Stokkum, S. J. C. M. Oomes, W. Crielaard, K. J. Hellingwerf, and S. Brul. 2005. Assessment of heat resistance of bacterial spores from food product isolates by fluorescence monitoring of dipicolinic acid release. Applied and Environmental Microbiology 71:3556-3564.
55. Kunst, F., N. Ogasawara, I. Moszer, A. M. Albertini, G. Alloni, V. Azevedo, M. G. Bertero, P. Bessieres, A. Bolotin, S. Borchert, R. Borriss, L. Boursier, A. Brans, M. Braun, S. C. Brignell, S. Bron, S. Brouillet, C. V. Bruschi, B. Caldwell, V. Capuano, N. M. Carter, S. K. Choi, J. J. Codani, I. F. Connerton, N. J. Cummings, R. A. Daniel, F. Denizot, K. M. Devine, A. Dusterhoft, S. D. Ehrlich, P. T. Emmerson, K. D. Entian, J. Errington, C. Fabret, E. Ferrari, D. Foulger, C. Fritz, M. Fujita, Y. Fujita, S. Fuma, A. Galizzi, N. Galleron, S. Y. Ghim, P. Glaser, A. Goffeau, E. J. Golightly, G. Grandi, G. Guiseppi, B. J. Guy, K. Haga, J. Haiech, C. R. Harwood, A. Henaut, H. Hilbert, S. Holsappel, S. Hosono, M. F. Hullo, M. Itaya, L. Jones, B. Joris, D. Karamata, Y. Kasahara, M. Klaerr-Blanchard, C. Klein, Y. Kobayashi, P. Koetter, G. Koningstein, S. Krogh, M. Kumano, K. Kurita, A. Lapidus, S. Lardinois, J. Lauber, V. Lazarevic, S. M. Lee, A. Levine, H. Liu, S. Masuda, C. Mauel, C. Medigue, N. Medina, R. P. Mellado, M. Mizuno, D. Moestl, S. Nakai, M. Noback, D. Noone, M. O’Reilly, K. Ogawa, A. Ogiwara, B. Oudega, S. H. Park, V. Parro, T. M. Pohl, D. Portetelle, S. Porwollik, A. M. Prescott, E. Presecan, P. Pujic, B. Purnelle, et al. 1997. The complete genome sequence of the Gram-positive bacterium Bacillus subtilis. Nature 390:249-256.
56. Leggett, M. J., G. McDonnell, S. P. Denyer, P. Setlow, and J. Y. Maillard. 2012. Bacterial spore structures and their protective role in biocide resistance. Journal of Applied Microbiology 113:485-

General introduction
25
1
498.57. Leguerinel, I., O. Couvert, and P. Mafart. 2000. Relationship between the apparent heat resistance
of Bacillus cereus spores and the pH and NaCl concentration of the recovery medium. International Journal of Food Microbiology 55:223-227.
58. Lenz, C. A., and R. F. Vogel. 2015. Differential effects of sporulation temperature on the high pressure resistance of Clostridium botulinum type E spores and the interconnection with sporulation medium cation contents. Food Microbiology 46:434-442.
59. Lewis, R. J., D. J. Scott, J. A. Brannigan, J. C. Ladds, M. A. Cervin, G. B. Spiegelman, J. G. Hoggett, I. Barák, and A. J. Wilkinson. 2002. Dimer formation and transcription activation in the sporulation response regulator Spo0A. Journal of Molecular Biology 316:235-245.
60. Li, J., and B. A. McClane. 2008. A novel small acid soluble protein variant is important for spore resistance of most Clostridium perfringens food poisoning isolates. PLoS Pathogens 4:e1000056.
61. Li, J., D. Paredes-Sabja, M. R. Sarker, and B. A. McClane. 2009. Further characterization of Clostridium perfringens small acid soluble protein-4 (Ssp4) properties and expression. PLoS ONE 4:e6249.
62. Li, Y., A. Davis, G. Korza, P. Zhang, Y. Q. Li, B. Setlow, P. Setlow, and B. Hao. 2012. Role of a SpoVA protein in dipicolinic acid uptake into developing spores of Bacillus subtilis. Journal of Bacteriology 194:1875-84.
63. Lima, L. J. R., H. J. Kamphuis, M. J. R. Nout, and M. H. Zwietering. 2011. Microbiota of cocoa powder with particular reference to aerobic thermoresistant spore-formers. Food Microbiology 28:573-582.
64. Lima, L. S. R. 2012. Microbial ecology of the cocoa chain: Quality aspects and insight into heat-resistant bacterial spores Wageningen University Wageningen.
65. Logan, N. A. 2012. Bacillus and relatives in foodborne illness. Journal of Applied Microbiology 112:417-429.
66. Loman, N. J., C. Constantinidou, J. Z. Chan, M. Halachev, M. Sergeant, C. W. Penn, E. R. Robinson, and M. J. Pallen. 2012. High-throughput bacterial genome sequencing: an embarrassment of choice, a world of opportunity. Nature Reviews Microbiology 10:599-606.
67. Luu-Thi, H., D. B. Khadka, and C. W. Michiels. 2014. Thermal inactivation parameters of spores from different phylogenetic groups of Bacillus cereus. International Journal of Food Microbiology 189:183-188.
68. Mafart, P., I. Leguérinel, O. Couvert, and L. Coroller. 2010. Quantification of spore resistance for assessment and optimization of heating processes: A never-ending story. Food Microbiology 27:568-572.
69. Markland, S. M., D. F. Farkas, K. E. Kniel, and D. G. Hoover. 2013. Pathogenic psychrotolerant sporeformers: an emerging challenge for low-temperature storage of minimally processed foods. Foodborne Pathogens and Disease 10:413-419.
70. Mathys, A., B. Chapman, M. Bull, V. Heinz, and D. Knorr. 2007. Flow cytometric assessment of Bacillus spore response to high pressure and heat. Innovative Food Science & Emerging Technologies 8:519-527.
71. McKenney, P. T., A. Driks, and P. Eichenberger. 2013. The Bacillus subtilis endospore: assembly and functions of the multilayered coat. Nature Reviews Microbiology 11:33-44.
72. Melly, E., P. C. Genest, M. E. Gilmore, S. Little, D. L. Popham, A. Driks, and P. Setlow. 2002. Analysis of the properties of spores of Bacillus subtilis prepared at different temperatures. Journal of Applied Microbiology 92:1105-1115.
73. Melly, E., and P. Setlow. 2001. Heat shock proteins do not influence wet heat resistance of Bacillus subtilis spores. Journal of Bacteriology 183:779-784.
74. Miller, R. A., D. J. Kent, M. J. Watterson, K. J. Boor, N. H. Martin, and M. Wiedmann. 2015. Spore populations among bulk tank raw milk and dairy powders are significantly different. Journal of Dairy Science 98:8492-8504.
75. Movahedi, S., and W. Waites. 2000. A two-dimensional protein gel electrophoresis study of the heat stress response of Bacillus subtilis cells during sporulation. Journal of Bacteriology 182:4758-4763.
76. Nguyen Thi Minh, H., A. Durand, P. Loison, J.-M. Perrier-Cornet, and P. Gervais. 2011. Effect of sporulation conditions on the resistance of Bacillus subtilis spores to heat and high pressure.

26
Chapter 1
Applied Microbiology and Biotechnology 90:1409-1417.77. Nicholson, W. L., N. Munakata, G. Horneck, H. J. Melosh, and P. Setlow. 2000. Resistance of
Bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiology and Molecular Biology Reviews 64:548-572.
78. Nicolas, P., U. Mäder, E. Dervyn, T. Rochat, A. Leduc, N. Pigeonneau, E. Bidnenko, E. Marchadier, M. Hoebeke, S. Aymerich, D. Becher, P. Bisicchia, E. Botella, O. Delumeau, G. Doherty, E. L. Denham, M. J. Fogg, V. Fromion, A. Goelzer, A. Hansen, E. Härtig, C. R. Harwood, G. Homuth, H. Jarmer, M. Jules, E. Klipp, L. Le Chat, F. Lecointe, P. Lewis, W. Liebermeister, A. March, R. A. T. Mars, P. Nannapaneni, D. Noone, S. Pohl, B. Rinn, F. Rügheimer, P. K. Sappa, F. Samson, M. Schaffer, B. Schwikowski, L. Steil, J. Stülke, T. Wiegert, K. M. Devine, A. J. Wilkinson, J. Maarten van Dijl, M. Hecker, U. Völker, P. Bessières, and P. Noirot. 2012. Condition-dependent transcriptome reveals high-level regulatory architecture in Bacillus subtilis. Science 335:1103-1106.
79. Ochman, H., J. G. Lawrence, and E. A. Groisman. 2000. Lateral gene transfer and the nature of bacterial innovation. Nature 405:299-304.
80. Oomes, S. J. C. M., and S. Brul. 2004. The effect of metal ions commonly present in food on gene expression of sporulating Bacillus subtilis cells in relation to spore wet heat resistance. Innovative Food Science & Emerging Technologies 5:307-316.
81. Oomes, S. J. C. M., A. C. M. van Zuijlen, J. O. Hehenkamp, H. Witsenboer, J. M. B. M. van der Vossen, and S. Brul. 2007. The characterisation of Bacillus spores occurring in the manufacturing of (low acid) canned products. International Journal of Food Microbiology 120:85-94.
82. Orsburn, B., S. B. Melville, and D. L. Popham. 2008. Factors contributing to heat resistance of Clostridium perfringens endospores. Applied and Environmental Microbiology 74:3328-3335.
83. Paidhungat, M., and P. Setlow. 2001. Localization of a germinant receptor protein (GerBA) to the inner membrane of Bacillus subtilis spores. Journal of Bacteriology 183:3982-3990.
84. Paredes-Sabja, D., P. Setlow, and M. R. Sarker. 2011. Germination of spores of Bacillales and Clostridiales species: mechanisms and proteins involved. Trends in Microbiology 19:85-94.
85. Paredes-Sabja, D., A. Shen, and J. A. Sorg. 2014. Clostridium difficile spore biology: sporulation, germination, and spore structural proteins. Trends in Microbiology 22:406-416.
86. Peck, M. W., S. C. Stringer, and A. T. Carter. 2011. Clostridium botulinum in the post-genomic era. Food Microbiology 28:183-191.
87. Pelczar, P. L., T. Igarashi, B. Setlow, and P. Setlow. 2007. Role of GerD in germination of Bacillus subtilis spores. Journal of Bacteriology 189:1090-1098.
88. Pelczar, P. L., and P. Setlow. 2008. Localization of the germination protein GerD to the inner membrane in Bacillus subtilis spores. Journal of Bacteriology 190:5635-5641.
89. Popham, D., B. Illades-Aguiar, and P. Setlow. 1995. The Bacillus subtilis dacB gene, encoding penicillin-binding protein 5*, is part of a three-gene operon required for proper spore cortex synthesis and spore core dehydration. Journal of Bacteriology 177:4721-4729.
90. Popham, D. L. 2002. Specialized peptidoglycan of the bacterial endospore: the inner wall of the lockbox. Cellular and Molecular Life Sciences 59:426-433.
91. Postollec, F., A. G. Mathot, M. Bernard, M. L. Divanac’h, S. Pavan, and D. Sohier. 2012. Tracking spore-forming bacteria in food: from natural biodiversity to selection by processes. International Journal of Food Microbiology 158:1-8.
92. Ragkousi, K., P. Eichenberger, C. van Ooij, and P. Setlow. 2003. Identification of a new gene essential for germination of Bacillus subtilis spores with Ca2+-dipicolinate. Journal of Bacteriology 185:2315-2329.
93. Reineke, K., A. Mathys, V. Heinz, and D. Knorr. 2013. Mechanisms of endospore inactivation under high pressure. Trends in Microbiology 21:296-304.
94. Rose, R., B. Setlow, A. Monroe, M. Mallozzi, A. Driks, and P. Setlow. 2007. Comparison of the properties of Bacillus subtilis spores made in liquid or on agar plates. Journal of Applied Microbiology 103:691-699.
95. Sanchez-Salas, J.-L., B. Setlow, P. Zhang, Y.-q. Li, and P. Setlow. 2011. Maturation of released spores is necessary for acquisition of full spore heat resistance during Bacillus subtilis sporulation. Applied and Environmental Microbiology 77:6746-6754.
96. Scheldeman, P., K. Goossens, M. Rodriguez-Diaz, A. Pil, J. Goris, L. Herman, P. De Vos, N. A.

General introduction
27
1
Logan, and M. Heyndrickx. 2004. Paenibacillus lactis sp. nov., isolated from raw and heat-treated milk. International Journal of Systematic and Evolutionary Microbiology 54:885-891.
97. Scheldeman, P., L. Herman, S. Foster, and M. Heyndrickx. 2006. Bacillus sporothermodurans and other highly heat-resistant spore formers in milk. Journal of Applied Microbiology 101:542-555.
98. Schroeder, J. W., and L. A. Simmons. 2013. Complete genome sequence of Bacillus subtilis strain PY79. Genome Announcements 1.
99. Schyns, G., C. R. Serra, T. Lapointe, J. B. Pereira-Leal, S. Potot, P. Fickers, J. B. Perkins, M. Wyss, and A. O. Henriques. 2013. Genome of a gut strain of Bacillus subtilis. Genome Announcements 1.
100. Sedlak, M., V. Vinter, J. Adamec, J. Vohradsky, Z. Voburka, and J. Chaloupka. 1993. Heat shock applied early in sporulation affects heat resistance of Bacillus megaterium spores. Journal of Bacteriology 175:8049-8052.
101. Setlow, P. 2014. Germination of spores of Bacillus species: What we know and do not know. Journal of Bacteriology 196:1297-1305.
102. Setlow, P. 2003. Spore germination. Current Opinion in Microbiology 6:550-556.103. Setlow, P. 2006. Spores of Bacillus subtilis: their resistance to and killing by radiation, heat and
chemicals. Journal of Applied Microbiology 101:514-525.104. Shah, I. M., M.-H. Laaberki, D. L. Popham, and J. Dworkin. 2008. A eukaryotic-like Ser/Thr kinase
signals bacteria to exit dormancy in response to peptidoglycan fragments. Cell 135:486-496.105. Smith, J. L., J. M. Goldberg, and A. D. Grossman. 2014. Complete genome sequences of Bacillus
subtilis subsp. subtilis laboratory strains JH642 (AG174) and AG1839. Genome Announcements 2.106. Stecchini, M. L., M. Del Torre, and P. Polese. 2013. Survival strategies of Bacillus spores in food.
Indian Journal of Experimental Biology 51:905-9.107. Sunde, E. P., P. Setlow, L. Hederstedt, and B. Halle. 2009. The physical state of water in bacterial
spores. Proceedings of the National Academy of Sciences 106:19334-19339.108. Talukdar, P. K., V. Olguín-Araneda, M. Alnoman, D. Paredes-Sabja, and M. R. Sarker. 2015.
Updates on the sporulation process in Clostridium species. Research in Microbiology 166:225-235.109. te Giffel, M. C., R. R. Beumer, S. Leijendekkers, and F. M. Rombouts. 1996. Incidence of Bacillus
cereus and Bacillus subtilis in foods in the Netherlands. Food Microbiology 13:53-58.110. te Giffel, M. C., A. Wagendorp, A. Herrewegh, and F. Driehuis. 2002. Bacterial spores in silage
and raw milk. Antonie van Leeuwenhoek 81:625-630.111. Tovar-Rojo, F., M. Chander, B. Setlow, and P. Setlow. 2002. The products of the spoVA operon are
involved in dipicolinic acid uptake into developing spores of Bacillus subtilis. Journal of Bacteriology 184:584-587.
112. Valerio, F., P. De Bellis, M. Di Biase, S. L. Lonigro, B. Giussani, A. Visconti, P. Lavermicocca, and A. Sisto. 2012. Diversity of spore-forming bacteria and identification of Bacillus amyloliquefaciens as a species frequently associated with the ropy spoilage of bread. International Journal of Food Microbiology 156:278-85.
113. van Asselt, E. D., and M. H. Zwietering. 2006. A systematic approach to determine global thermal inactivation parameters for various food pathogens. International Journal of Food Microbiology 107:73-82.
114. van Melis, C. C. J., H. M. W. den Besten, M. N. Nierop Groot, and T. Abee. 2014. Quantification of the impact of single and multiple mild stresses on outgrowth heterogeneity of Bacillus cereus spores. International Journal of Food Microbiology 177:57-62.
115. van Zuijlen, A., P. M. Periago, A. Amézquita, A. Palop, S. Brul, and P. S. Fernández. 2010. Characterization of Bacillus sporothermodurans IC4 spores; putative indicator microorganism for optimisation of thermal processes in food sterilisation. Food Research International 43:1895-1901.
116. Velasquez, J., G. Schuurman-Wolters, J. P. Birkner, T. Abee, and B. Poolman. 2014. Bacillus subtilis spore protein SpoVAC functions as a mechanosensitive channel. Molecular Microbiology 92:813-23.
117. Vepachedu, V. R., and P. Setlow. 2007. Role of SpoVA proteins in release of dipicolinic acid during germination of Bacillus subtilis spores triggered by dodecylamine or lysozyme. Journal of Bacteriology 189:1565-1572.
118. Vreeland, R. H., W. D. Rosenzweig, and D. W. Powers. 2000. Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal. Nature 407:897-900.

28
Chapter 1
119. Wang, G., P. Zhang, P. Setlow, and Y.-q. Li. 2011. Kinetics of germination of wet-heat-treated individual spores of Bacillus species, monitored by raman spectroscopy and differential interference contrast microscopy. Applied and Environmental Microbiology 77:3368-3379.
120. Warda, A. K., H. M. W. den Besten, N. Sha, T. Abee, and M. N. Nierop Groot. 2015. Influence of food matrix on outgrowth heterogeneity of heat damaged Bacillus cereus spores. International Journal of Food Microbiology 201:27-34.
121. Warth, A. D. 1978. Relationship between the heat resistance of spores and the optimum and maximum growth temperatures of Bacillus species. Journal of Bacteriology 134:699-705.
122. Xiao, Y., C. Francke, T. Abee, and M. H. J. Wells-Bennik. 2011. Clostridial spore germination versus bacilli: Genome mining and current insights. Food Microbiology 28:266-274.
123. Xiao, Y., A. Wagendorp, T. Abee, and M. H. J. Wells-Bennik. 2015. Differential outgrowth potential of Clostridium perfringens food-borne isolates with various cpe-genotypes in vacuum-packed ground beef during storage at 12°C. International Journal of Food Microbiology 194:40-45.
124. Yeo, I. C., N. K. Lee, and Y. T. Hahm. 2012. Genome sequencing of Bacillus subtilis SC-8, antagonistic to the Bacillus cereus group, isolated from traditional Korean fermented-soybean food. Journal of Bacteriology 194:536-7.
125. Zeigler, D. R. 2011. The genome sequence of Bacillus subtilis subsp. spizizenii W23: insights into speciation within the B. subtilis complex and into the history of B. subtilis genetics. Microbiology 157:2033-41.