urethanes andsoil nitrificationaem.asm.org/content/1/6/282.full.pdf · urethanes andsoil...
TRANSCRIPT

Urethanes and Soil NitrificationJ. H. QUASTEL AND P. G. SCHOLEFIELD
From the Research Institute, Montreal General Hospital, Montreal, Canada
Received for publication May 15, 1953
Urethanes have marked physiological activity inplants and animals. Lefevre (1939) found that phenylurethane affects wheat germination, and subsequentlyTempleman and Sexton (1946) showed that a variety ofurethanes suppress the germination of wheat and oatswithout affecting charlock. The most active urethanes(for example, isopropylphenylcarbamate;- ethylphenyl-carbamate) are effective at a concentration of a fewparts per million. These arylcarbamic esters are knownto affect mitosis in cells and this fact led to an investi-gation of urethanes as possible inhibitors of tumorgrowth. Haddow and Sexton (1946) obtained positiveresults in animals with isopropylphenylcarbamate,ethylphenylcarbamate and ethyl urethane, though thelast is apparently inactive in plants. Ethyl urethane isused in the treatment of leukemia (Patterson et al.,1946), but in mice its administration gives rise to lungtumors (Larsen and Heston, 1945; Larsen, 1947). Theseeffects of urethane take place at much smaller concen-trations than those required to bring about its wellknown narcotic affects. Esterases are inhibited byurethanes (Stedman and Stedman, 1931, 1932), the in-hibition of choline esterase by such urethanes as eserineand prostigmine being well known. Schweitzer, Sted-man and Wright (1939) conclude that the urethanegrouping is essential for esterase inhibition.Meyerhof (1916) showed that urethanes inhibit the
respiration of nitrite oxidising organisms, the in-hibitory power increasing with increasing size of thealiphatic radicle; for example, methyl urethane, in-hibits the oxygen uptake of these organisms by 50per cent at a concentration of 0.3M, but isoamylurethane is equally effective at a concentration of0.022M. Ethyl urethane also inhibits respiration ofisolated pure cultures of Nitrosomonas by 42 per cent ata cocentration of 0.016M and gives 4 per cent inhibitionof Nitrobacter respiration at a concentration of 0.11M.The inhibitory effect of urethane was confirmed byLees and Quastel (1946), who found, however, a highersensitivity of the process of nitrification to ethylurethane in soil culture. The inhibitory effect of ethylurethane on soil nitrification was also shown to bereversible. Quastel and Scholefield (1949) pointed outthat while 0.01M ethyl urethane completely suppressesthe nitrification of 0.01m ammonium chloride in gardensoil for about twenty days, the subsequent rate ofnitrification proceeds normally. The final nitrate
282
nitrogen recovered, however, accounts for 80 per centof the total nitrogen of the ammonium chloride and ofthe ethyl urethane. Urethane perfused by itself, isultimately metabolized by soil organisms. Moreover,urethane at low concentrations (10-m) greatly inhibitsnitrate formation from ammonium ions on perfusionthrough a fresh soil, but at a concentration of3.6 X 10-m it inhibits by only 50 per cent the rate ofoxygen uptake due to ammonium ion oxidation in anenriched soil. At a concentration one tenth that re-quired to produce a definite inhibition of the rate ofoxygen uptake, ethyl urethane secures almost completeinhibition of proliferation of the ammonia oxidisingorganisms in soil. The conclusion was drawn (Quasteland Scholefield, 1949) that ethyl urethane, at low con-centrations, interferes with some aspect of ammoniametabolism essential for the development of theseorganisms.Although ethyl urethane at a concentration of
3 X 10-m inhibits the growth of the ammonia oxidisingorganisms, and at 36 X 10 4m inhibits by 50 per centtheir rate of ammonia oxidation, a concentration of2,000 X 10-4M is required to inhibit the respiration ofrat brain cortex of 50 per cent (Jowett, 1938) and aconcentration of 300 X 1074m inhibits guinea pig brainrespiration by only 20 per cent and does not affectyeast respiration at all (Quastel and Wheatley, 1932).
Quastel and Scholefield (1951) conclude from theresults obtained on perfusing mixtures of ammoniumchloride and ethyl urethane through soils that nitrifi-cation of such mixtures takes place only when organismscapable of breaking down the urethane have developedin the soil.
It is the purpose of this paper to describe the effectsof various urethanes on soil nitrification and to showthat, in the presence of ethyl urethane, organisms de-velop in soils which possess an enzyme capable ofattacking ethyl urethane probably with the formation ofethanol (cf. Skipper et al., 1951).
METHODSThe technique adopted has been fully described else-
where (Quastel and Scholefield, 1951) and therefore willnot be given in detail. Briefly, the method of investi-gation was that of continuous perfusion in the dark at21 C of a neutral solution containing ammonium chlo-ride and/or ethyl urethane through a column of 30 g
on May 12, 2018 by guest
http://aem.asm
.org/D
ownloaded from

URETHANES AND SOIL NITRIFICATION
air-dried crumbed soil (2-4 mm crumbs) under con-uditions of optimal aeration and water saturation but-with no waterlogging. The rates of conversion of Am-monium ions to nitrate were measured, the methods ofanalysis of samples from the soil perfusion apparatusbeing those previously described (Quastel and Schole-field, 1951).
Manometric studies were carried out with the con-ventional Warburg manometric apparatus using soilsenriched or saturated with nitrifying organisms or withorganisms capable of oxidising ethyl urethane. Theprocess of adapting such soils for manometric investi-gations has been described in detail (Qua"tel andScholefield, 1951).
TABLE 1. Effects of varying urethanes (0.0038 M),on rates of nitrification of ammonium chloride
(0.01 M)TIME IN DAYS
URETHANE USED IN PRESENCE OF O.01M ESTIMATOEODNH4CIt jA~~~g/ILL OF LAG PERIODNH4C1t ~~~~NITRATE-
NITROGZN
None.......................... 9 0*Isopropyl carbanilate C6H5NH.CO*OCH(CH)2 ................. 41 32
Ethyl-N-butyl carbamate C4H9-NH.CO-OC2H5 .................. 38 29
Ethyl carbamate NH2 COOC2HW. 20 11Ethyl carbanilate CGH5NH CO0OC2H5.......................... 15.5 6.5
Ethyl - N - isopropyl carbamate(CH3)2CH-NH*CO*OC2H, ....... 15 6
*Ethyl-2-cyclohexylcyclohexyl car-bamate C6H11 C6Ho- NH*CO-OC2H0 .......................... 12 3
*Octyl-cyclohexyl carbamateCsH11NH.CO*OC8H7 .................. 9 0
*Octyl-N-phenyl-carbamate C6H5,NH.CO*OC8H17 ................. 9 0
* used in suspension.t 30 g soil at 21 C.
RESULTS
Effects of Urethanes on Nitrification of AmmoniumIons in Soils
A series of urethanes R NH - CO* OR', including ethylurethane NH2 CO OC2H-, was perfused in the presence
of 0.01M NH4Cl through soil. Their effects on the lengthof the lag period before nitrification of the ammoniumion commenced was determined from the period requiredto produce 70 usg/ml nitrate-nitrogen. Typical results areshown in table 1.The most active compounds in the series were ethyl-
N-butyl carbamate and isopropyl-carbanilate. Eventhough the latter was used as a 3.3 X 10-3M suspensionit produced a thirty-two-day lag period before nitrifi-cation of ammonium chloride began. Ethyl carbamate(urethane) caused an eleven-day lag period at this
concentration. The presence of a phenyl group incarbanilates is not itself responsible for high toxicity, asethyl carbanilate produced a shorter lag period (6.5days) than that due to ethyl carbamate. Ethyl N-isopropyl carbamate produced a 6-day lag period com-pared with the 32 days obtained with isopropyl car-banilate. It seems, therefore, that the ability of thesubstituted carbamates to inhibit nitrification is a func-tion of both substituents, R and R1. A substituted ure-thane having a high inhibitory effect on nitrification ofammonium ions in soil is eserine which is an ester ofN-methyl carbamic acid. Typical results with eserineare shown in table 2. It proves to bB at least ten timesas effective as ethyl urethane in inhibiting ammoniumion oxidation in a fresh garden soil.
TABLE 2. Effects of eserine at various concentrationson nitrification of 0.01 M NH4Cl
NITiLATE-NITROGEN IN
CNETOFPERFPUSATE* pg/IuL PENFUSATECONTENTS OF PERFUS AFTER 13 DAYSPERFUSION
0.01 M NHCl ............................ 740.01 M NH4Cl + 0.001 per cent Eserine... 650.01 M NH4Cl + 0.003 per cent Eserine... 510.01 M NH4Cl + 0.01 per cent Eserine.... 330.01 M NH4C1 + 0.03 per cent Eserine.... 190.01 M NH4Cl + 0.05 per cent Eserine.... 17
* 30 g fresh soil at 21 C.
TABLE 3. Effect of CMU(ClC6H4HNH.CON(CH3)2)on nitrite formation from ammonium chloride in
fresh soil
CONTENTS OF PERFUSATE* pAg NITRITE FORMED/IL
3.3 X 10-3MCMU....................... 4102 M NH4C1 ............................ 2810-2M NH4Cl + 3.3 x 10-3 M CMU ......
* 10- M NaC0l present throughout. 10 days at 21 C; 30 gsoil.
Comparison of Inhibition of Nitrification with HerbicidalEffects of Substituted Urethanes
Urethanes such as isopropyl carbanilate and ethylcarbanilate, having high herbicidal activities, havemarked inhibitory effects on the nitrification of am-monium ions in soil. On the other hand, a powerfulinhibitor of soil nitrification, ethyl urethane, has butlittle herbicidal activity. There is, therefore, no com-plete parallel between the inhibition of nitrification andthe herbicidal action of the urethanes. There may be,however, some connection between these two propertiesof the urethanes, since the most active of them asherbicides tend to be the most active inhibitors ofnitrification. The substituted ureide known as CMU(Cl.C.H4 NH-C00N(CH3)2), which is a powerfulherbicide, is also a powerful inhibitor of soil nitrification.Typical results with CMU are shown in table 3.
,283
on May 12, 2018 by guest
http://aem.asm
.org/D
ownloaded from

J. H. QUASTEL AND P. G. SCHOLEFIELD
Nitrate Formation from UrethanesEthyl urethane and other carbamic esters are con-
verted to nitrate in fresh soil after lengthy lag periods.It has already been shown (Quastel and Scholefield,1949) that urethane may be so converted, and theresults given in figure 1 show the effects of perfusing soil
120
80
NO' - N/miI
0 2 4DAYS
FIG. 1. The nitrification of 0.01 M ethyl urethane by enrichedsoils. Each value is the average of the results of twelve experi-ments carried out at the same time.
possibly due to the action of heterotrophic organisms inutilizing the liberated ammonia or its oxidation prod-ucts. The final rate of nitrification is quite constantduring successive perfusions and, once established, maybe used for kinetic studies.The effects of perfusing substituted urethanes alone
and in admixture with ethyl urethane over soils en-riched by successive perfusions of ethyl urethane areshown in table 4. None of the substituted urethanesstudied was immediately converted to nitrate at a rapidrate by the ethyl urethane enriched soil. Thusorganismswhich develop in soil in response to the presence ofethyl urethane and which areabletoattack it, giv,ingriseto nitrate, are not immediately capable of conkvertingto nitrate the other urethanes investigated. On the otherhand the presence of various urethanes (for example,isopropyl N-phenyl carbamate, ethyl-N-butyl car-bamate, and ethyl-N-isopropyl carbamate) greatlyinhibits the rate of nitrate fornation from ethylurethane (table 4).The mechanism of this inhibition is not yet estab-
lished. The inhibitory urethane may act by competingwith ethyl urethane for the enzyme responsible for theinitial attack on this substance, or it may act by in-hibiting the nitrification of ammonium ions liberated on
TABLE 4. Effects on rates of nitrate formation of perfusing substituted urethanes (R .NH. COOR1) together with ethylurethane(NH2. COOC2H6) through a soil,t enriched with organisms capable of converting ethyl urethane to nitrate.
RATE OBTAINED RATE OBTAINED WITH RATZ OBTAINZD WITH (3) EXPRESSEDR RWITH ETHYL R-NH.COOR' (2) AS PERCENTA E MIXTuRz OF ETHYL AS PERCENTAGEURETHANE ALONEt AWNEt OF (1) URETEANE + URETHANE
(1) (2) R*NH*COOR' (3)t 0F ()
H C2H6 1.405 *1.465 1041.016 *0.996 98
C6H,Hll C6H,o C2Hr 1.598 0.076 51.669 1.696 102
C065 (CH,)2CH 1.882 0.037 2 - -1.185 0.052 4
(CH3)2CH C2H5 0.956 0.053 5 - -1.243 0.762 61
C6H,j CsH0 7 1.405 0.045 3 - -1.511 1.728 114
CXH5 CsH17 2.032 0.091 4 _1.357 1.323 98
C4H9 C2Hs 1.203 0.033 31.364 - 0.025 2
dT5 C2H5 1.950 **>0.65 >33 /1.473 _***>0.85 '*58
* i.e. Value found on reperfusion. ** Lag period = 6 days. *** Lag period = 6.5 days.t 30 g soil. Duration of experiment 8 days at 21 C.t Rates expressed as ;&g NO'3-N formed per ml perfusate per hour.
with ethyl urethane, after several previous perfusionswith this substance. For two days nitrate formationproceeds slowly; it then increases rapidly and finallyproceeds at a constant rate. This type of result is notconfined to ethyl urethane but is typical of the course ofnitrate formation from most organic nitrogen com-pounds that have been studied. The initial low rate is
breakdown of ethyl urethane. If the first explanation iscorrect, then the enzyme first attacking ethyl urethanepossesses a high degree of specificity. If the secondexplanation is correct, it is apparent that nitrifyingorganisms may retain their ability to oxidise ammoniumions in presence of excess of ethyl urethane but not inpresence of the other urethanes investigated. Thus an
284
-7-
on May 12, 2018 by guest
http://aem.asm
.org/D
ownloaded from
URETHANES AND SOIL NITRIFICATION
acquired resistance to ethyl urethane would not implyan acquired resistance to other urethanes.
Course of Breakdown of Ethyl Urethane bySoil Microorganisms
The breakdown of ethyl urethane, on perfusionthrough soil, is a biological process. This is shown bythe fact that the lengthy lag period preceding theconversion of urethane to nitrate is shortened by theintroduction into a fresh soil of a small quantity of asoil capable of rapid breakdown of ethyl urethane. Theeffects on the rates of nitrate formation from ethylurethane of such inoculation with varying quantities ofenriched soil are shown in figure 2, where the logarithmof nitrate-N production on perfusion of ethyl urethane
2.00 7 ~~~/1.60
log(piNO t*%n)
DAYSFIG. 2. The effect of size of the inoculum of enriched soil
on the rate of nitrate formation from 0.O1m ethyl urethane ina fresh soil.
(o 1.0 g Inoculum* 2.0 g Inoculum0 4.0 g InoculumA No inoculum, showing values obtained at 26th and 28th
days.
through soil is plotted against time. It will be seen thatthere is a linear relationship, a result to be expected ifthe nitrate formation is due to proliferation of micro-organisms (Quastel and Scholefield, 1951). Further, theinitial rates of nitrate formation are increased when thesize of the inoculum of enriched soils is increased and thevarious lines in figure 2 are parallel to each other. Thelast fact indicates that the rate of proliferation of thenitrifying organisms is a constant under the experi-mental conditions.
MANOMETRIC RESULTS
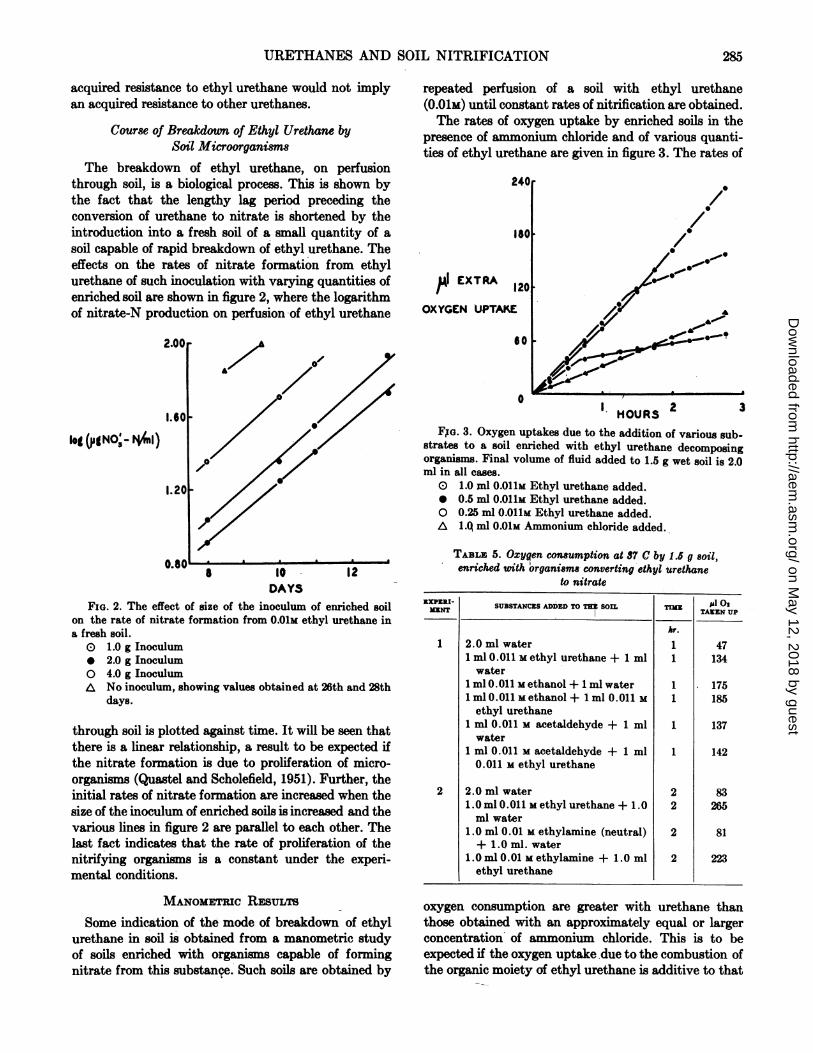
Some indication of the mode of breakdown of ethylurethane in soil is obtained from a manometric studyof soils enriched with organisms capable of formingnitrate from this substanqe. Such soils are obtained by
a
repeated perfusion of a soil with ethyl urethane(0.01M) until constant rates of nitrification are obtained.The rates of oxygen uptake by enriched soils in the
presence of ammonium chloride and of various quanti-ties of ethyl urethane are given in figure 3. The rates of
EXTRA
OXYGEN UPTAIKE
v 1I, 2 3HOURS
FXo. 3. Oxygen uptakes due to the addition of various sub-strates to a soil enriched with ethyl urethane decomposingorganisms. Final volume of fluid added to 1.5 g wet soil is 2.0ml in all cases.O 1.0 inl 0.011m Ethyl urethane added.* 0.5 ml 0.O11M Ethyl urethane added.O 0.25 ml 0.O11M Ethyl urethane added.A LQ ml 0.01M Ammonium chloride added.
TABLE 5. Oxygen consumption at 87 C by 1.5 g 80il,enriched with organisms converting ethyl urethane
to nitrateEXPEKI- SUBSTANCES ADDED To TEk SOI TIME IPI O__N_ I- TAKEN UP
1 2.0 ml water 1 471 ml 0.011 M ethyl urethane + 1 ml 1 134water
1 ml 0.011 methanol + 1 ml water 1 1751 ml 0.011 Methanol+ 1 ml 0.011M 1 185ethyl urethane
1 ml 0.011 M acetaldehyde + 1 ml 1 137water
1 ml 0.011 M acetaldehyde + 1 ml 1 1420.011 M ethyl urethane
2 2.0 ml water 2 831.0 ml 0.01 Methyl urethane + 1.0 2 265ml water
1.0 ml 0.01 M ethylamine (neutral) 2 81+ 1.0 ml. water
1.0 ml 0.01 Methylamine + 1.0 ml 2 223ethyl urethane
oxygen consumption are greater with urethane thanthose obtained with an approximately equal or largerconcentration of ammonium chloride. This is to beexpected if the oxygen uptake.due to the combustion ofthe organic moiety of ethyl urethane is additive to that
285r
on May 12, 2018 by guest
http://aem.asm
.org/D
ownloaded from

J. H. QUASTEL AND P. G.' SCHOLEFIELD
due to oxidation of liberated ammonia. The optimalrate of oxygen consumption with ethyl urethane is ob-tained with a concentration of 0.005M.
Using low concentrations of ethyl urethane it is pos-sible to estimate the amount of oxygen consumed forthe conversion to nitrate of 1 mole ethyl urethane.For example, the oxidation of 1.0 ml 0.OO1M ethylurethane required 68 ,ul oxygen. This corresponds to anuptake of 6 atoms oxygen per mole ethyl urethane andis in accordance with the following equation:
NH2'COOC2H6 + 302= CO2 + HNO3 + CH3COOH + H20
A likely method of breakdown of ethyl urethanewould be a preliminary hydrolysis to ethanol, ammoniaand carbon dioxide, followed by oxidation of ethanol toacetic acid and of ammonia to nitric acid. If this modeof breakdown takes place, then ethanol should be oxi-dised to acetic acid by a soil enriched with ethylurethane decomposing organisms. The results shown intable 5 indicate that ethanol is oxidised by such an en-riched soil at a greater rate than is ethyl urethane, andthis rate is not increased on admixture of ethyl urethanewith ethanol. Moreover, the oxygen consumption foundwith ethanol corresponds to two atoms oxygen per moleof ethanol or the amount required for oxidation toacetic acid. Addition of sodium acetate to an enrichedsoil leads to little or no increased rate of oxygen uptake.The addition of acetaldehyde, at concentrations
similar to those used with urethane and ethanol, alsoleads to an increased rate of oxygen uptake, the ratebeing about that obtained with urethane and is notincreased on addition of urethane (table 5). The amountof oxygen consumed amounts to one atom of oxygen permole acetaldehyde which again corresponds to aceticacid formation.Thus the rates and amounts of oxygen consumed by
ethanol and acetaldehyde in presence of soils enrichedwith organisms oxidising ethyl urethane are in accord-ance with the view that the urethane undergoes a pre-liminary hydrolysis to ethanol which is then oxidised toacetic acid.
It is unlikely that ethyl urethane first undergoes de-carboxylation to ethylamine, since the addition ofethylamine to an enriched soil does not lead to anyincreased rate of oxygen consumption (table 5).
ACKNOWLEDGMENTSWe are grateful to the Monsanto Chemical Company
for gifts of the various urethanes used in this investi-gation. We are also much indebted to the NationalCancer Institute of Canada for a grant-in-aid.
SUMMARY
The inhibitive effects of a series of urethanes on the
investigated. The most active compounds of the seriesare ethyl-N-butyl carbamate and isopropyl carbanilate.Eserine is also highly effective, being ten times more
active than ethyl urethane.The ureide, Cl * C6H4 * NH. CO- N(CH3)2, known as
CMU is a powerful inhibitor of soil nitrification.Although there is no strict parallelism between
herbicidal effects and inhibition of soil nitrification byurethanes, there is a tendency for the most powerfulherbicides to have the most vigorous inhibitive effectson nitrification in soil.Ethyl urethane and other carbamic esters are con-
verted to nitrate in fresh soils after lengthy lag periods.Organisms that develop in soil in response to the
presence of ethyl urethane and are capable of attackingit, are not immediately capable of attacking otherurethanes. The presence of a variety of urethanesinhibits the rate of nitrate formation from ethylurethane.Manometric studies of soils enriched with organisms
attacking ethyl urethane indicate that such organismsattack ethanol and acetaldehyde. The amounts ofoxygen consumed for the oxidation of ethyl urethane,alcohol and acetaldehyde are in accordance with theconclusion that acetic acid is the end product. Aceticacid is apparently not oxidised. Kinetic and stochio-metric evidence lead to the conclusion that ethylurethane is first broken down by soil organisms toethanol, ammonia and carbon dioxide, with subsequentformation of acetic and nitric acids.
REFERENCES
HADDOW, A., AND SEXTON, W. A. 1946 Influence of carbamicesters (urethanes) on experimental animal tumours.Nature, 147, 500-503.
JOWETT, M. 1938 The action of narcotics on brain respira-tion. J. Physiol., 92, 322-335.
LARSEN, C. D. 1947 Evaluation of the carcinogenicity ofa series of esters of carbamic acid. J. Natl. Cancer Inst.,8, 99-101.
LARSEN, C. D., AND HESTON, W. E. 1945 Induction of pul-monary tumours in mice by anaesthetic agents. CancerResearch, 5, 592.
LEES, H., AND QUASTEL, J. H. 1946 Biochemistry of nitrifi-cation in soil. Biochem. J., 40, 803-828.
LEFEVRE, J. 1939 Similarity of the cytologic action exertedby phenylurethan and colchicine on seedlings. Compt.rend., 208, 301-304.
MEYERHO?, 0. 1916 Untersuchungen uber den Atmungs-vorgang nitrifizierenden Bakterien. I. Die Atmung desNitratbildners. Pfluigers Arch. ges. Physiol., 164, 353-427.II. Beeinflussung der Atmung des Nitratbildners durch che-mische Substanzen. Ibid., 165, 229-284. III. Die Atmungdes Nitritbildners und ihre Beeinflussung durch chemischeSubstanzen. Ibid, 166, 240-280.
PATTERSON, E., HADDOW, A., THOMAS, I. A., AND WATKINSON,J. M. 1946 Leukemia treated with urethane comparedwith deep X-ray therapy. Lancet, 250, 677-682.
QUASTEL, J. H., AND SCHOLEFIELD, P. G. 1949 Influenceof organic compounds on nitrification in soil. Nature,
nitrification of ammonium ions in fresh soil have been
286
164, 1068-1072.
on May 12, 2018 by guest
http://aem.asm
.org/D
ownloaded from

HUMIDITY REQUIREMENTS FOR MOLD GROWTH
QUASTEL, J. H., AND SCHOLEFIELD, P. G. 1951 Biochemistryof nitrification in soil. Bacteriol. Revs., 15, 1-53.
QUASTEL, J. H., AND WHEATLEY, A. H. M. 1932 Narcosisand oxidations of the brain. Proc. Roy. Soc. (London)B., 112, 60-79.
SCHWEITZER, A., STEDMAN, E., AND WRIGHT, S. 1939 Centralaction of anticholinesterases. J. Physiol., 96, 302-336.
SKIPPER, H. E., BENNETr, L. L., JR., BRYAN, C. E., WHITE,L., JR., NEWTON, M. A., AND SIMPSON, L. 1951 Carba-mates in the chemotherapy of leukemia. VIII. Over-alltracer studies on carbonyl-labeled urethan, methylene-labeled urethan and methylene-labeled ethyl alcohol.Cancer Research, 11, 46-51.
STEDMAN, E., AND STEDMAN, E. 1931 Studies on the relation-ship between chemical constitution of physiological action.III. Inhibitory action of certain synthetic urethanes onactivity of liver esterase. Biochem. J., 25, 1147-1167.
STEDMAN, E., AND STEDMAN, E. 1932 Studies on the relation-ship between chemical constitution of physiological ac-tion. IV. Inhibitory action of certain synthetic urethaneson activity of esterases. Biochem. J., 26, 1214-1222.
TEMPLEMAN, W. G., AND SEXTON, W. A. 1946 Differentialeffect of synthetic growth substances and other com-pounds on plant species. II. Seed germination and earlygrowth responses to some aryl carbamic esters and relatedcompounds. Proc. Roy. Soc. (London) B. 133, 480-485.
Humidity Requirements for Mold GrowthS. S. BLOCK
Department of Chemical Engineering, University of Florida, Gainesville, Florida
Received for publication June 22, 1953
In the warm, humid areas of the United States and ofthe world, mold or mildew prevention ranks as a majorpest-control problem. Leather goods, clothing, foods,mattresses, painted walls, and even glass camera lensesare subject to the unsightly appearance, stain, damage,or musty odor associated with the growth of the lower-order plants belonging to the fungi and known popu-larly as mildew or mold.As has been mentioned previously in a discussion of
the mildew problem (Block, 1946), mold requires forgrowth and proliferation certain essential physical andchemical conditions. These include satisfactory tem-perature, adequate moisture, sufficient oxygen, properpH, and essential nutrients. Prevention of mold growthmay be effected by restricting any of the above require-ments. Mildew may be controlled also by physicalagents that destroy the organism, such as ultravioletradiation and heat sterilization, and by chemical poi-sons called fungicides. The specific conditions best de-termine the method to be employed to prevent moldgrowth. In certain applications the control of moisturethrough regulation of humidity has proved to be mostsatisfactory (Block, 1951). This applies to the preven-tionof mildew in home closets, rooms, storage spaces andshipping units.At the request of the British Public Record Office,
Groom and Panisset (1933) studied the conditionsnecessary for mildew to develop on materials used forbook covers. Working with Penicillium chrysogenum,they noted that the minimum relative humidity forer)ore germination on glass was 81 per cent and formildewing of leather and other book materials was72.8 per cent. Since it is obvious that mildew couldnot be present unless the mold spores could germinate,
Groom and Panisset suggest that the spores germinateat a lower relative humidity on book materials becauseof the presence of nutrients. Galloway (1935) reportedthat the minimum relative humidity permitting growthof molds varies from 75 to 95 per cent for different spe-cies; thus protection of materials is assured only if theatmosphere is below 75 per cent R.H. (relative hu-midity). Clayton (1942) found that some fungus sporesgerminate at 0 per cent humidity on glass, but thesefungi are associated with plant diseases and not mil-dewing of inanimate objects. A report of the U. S. Nat-ional Bureau of Standards (1947) states that at 85 percent R.H., or less, no mildew growth on leather oc-curred, but that at 95 per cent R.H. there was heavygrowth. Illman and Weatherburn (1947) made an ex-cellent study of the factors affecting the developmentof mold on various materials. They found no moldgrowth at 60 per cent R.H. and very little at 70 percent R.H., but increasing amounts from 80 to 100 percent R.H.To what extent does the moisture in the substrate
determine mold growth as apart from the relative hu-midity of the atmosphere? How are different mate-rials affected under the same physical conditions ofhumidity and temperature? Galloway (1935) offereddata to show that atmospheric moisture is more effectivethan moisture in the substrata for bringing about thegermination of mold spores. For information on thesecond question, Smith (1942) states, "If dry samples ofpure wool and pure cotton are exposed to the sameatmosphere the wool will take up approximately twiceas much moisture as the cotton and, leaving out of ac-count differences due to chemical composition, the twosamples will be approximately equally liable to mildew."
287
on May 12, 2018 by guest
http://aem.asm
.org/D
ownloaded from