uva-dare (digital academic repository) neogene climate … · carvalho, m. j. garcia, o....
TRANSCRIPT

UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl)
UvA-DARE (Digital Academic Repository)
Neogene climate evolution in Amazonia and the Brazilian Northeast
Hoorn, M.C.; Bernardes-de-Oliveira, M.E.C.; Dino, R.; Garcia, M.J.; Antonioli, L.; da CostaCasado, F.; Hooghiemstra, H.Published in:Paleontologia: Cenários de vida. - Volume 5: Paleoclimas
Link to publication
Citation for published version (APA):Hoorn, C., Bernardes-de-Oliveira, M. E. C., Dino, R., Garcia, M. J., Antonioli, L., da Costa Casado, F., &Hooghiemstra, H. (2014). Neogene climate evolution in Amazonia and the Brazilian Northeast. In I. de SouzaCarvalho, M. J. Garcia, O. Strohschoen, & C. Cunha Lana (Eds.), Paleontologia: Cenários de vida. - Volume 5:Paleoclimas (pp. 277-310). Rio de Janeiro: Editora Interciência.
General rightsIt is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s),other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons).
Disclaimer/Complaints regulationsIf you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, statingyour reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Askthe Library: http://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam,The Netherlands. You will be contacted as soon as possible.
Download date: 13 Nov 2018

ISBN Paleontologia: Cenários de Vida – Paleoclimas
NEOGENE CLIMATE EVOLUTION IN AMAZONIA AND THE BRAZILIAN NORTHEAST
EVOLUÇÃO CLIMÁTICA NEÓGENA DA AMAZÔNIA E NORDESTE BRASILEIROCarina Hoorn1, Mary E.C. Bernardes-de-Oliveira2,3, Rodolfo Dino4,5, Maria Judite Garcia2,6, Luzia Antonioli4, Fabio da Costa Casado6 & Henry Hooghiemstra1
1University of Amsterdam, Paleoecology and Landscape Ecology, Institute for Biodiversity and Ecosystem Dynamics, P.O. BOX 94248 / 1090 GE, Amsterdam, The Netherlands 2Universidade Guarulhos, CEPPE/UnG, Praça Tereza Cristina 1, Centro, 07023-070 Guarulhos, SP, Brazil 3Universidade de São Paulo, Instituto de Geociências, rua do Lago 562, Cidade universitária, 05508-080, São Paulo, Brazil.4CENPES-Petrobras, Avenida Horácio Macedo, 950. 20550-900 Ilha do Fundão, Rio de Janeiro, RJ, Brazil 5Universidade do Estado do Rio de Janeiro, IG, Rua São Francisco Xavier, 524, Maracanã, 20550-900 Rio de Janeiro, RJ, Brazil6Universidade Guarulhos, Laboratório de Geociências, Universidade Guarulhos, Praça Tereza Cristina 1, Centro, 07023-070, Guarulhos, SP, BrazilE-mails: [email protected], [email protected], [email protected], [email protected], [email protected], [email protected], [email protected]
ABSTRACT
Climate change follows from the interaction between global atmospheric and oceanic processes with regional processes. In this chapter we review which factors determined climate evolution in Amazonia and the Brazilian Northeast and present a recompilation of Neogene palynological and paleobotanical records. These records suggest generally warm and humid conditions throughout this region during the Neogene. The uplift of the Andes, in the western half of South America, created a pattern of intense orographic precipitation in Western Amazonia that derived from humid air masses of Atlantic origin. During this time interval uplift of the Borborema Plateau, in the northern Brazilian Highlands, also formed an orographic barrier that allowed the development of a lush forest on the Atlantic side (Mata Atlantica), while rain shadow on the western flank gave place to the semi-arid Caatinga vegetation. The arid conditions in the Brazilian Northeast were further exacerbated by the evolution of the oceanic current system in the Atlantic, a system that was also responsible for the creation of the Namib Desert. The palynological and paleobotanical data illustrate the long history of Amazonia’s tropical forest, but also the influence of the Andes formation on this region. The limited paleobotanical remains from the Brazilian Northeast tentatively point at longstanding humid conditions that, in the latter part of the Neogene transformed into the predominantly arid conditions that characterize the area at present.Keywords: climate, Miocene, Pliocene, plant macrofossils, palynomorphs
RESUMO
As mudanças climáticas resultam da interação global entre os processos atmosféricos e oceânicos que afetam os processos regionais. Neste capítulo são revistos os fatores que determinaram a

280 Paleontologia: Cenários de Vida – Paleoclimas
evolução do clima na Amazônia e no Nordeste brasileiro, além de apresentar uma recompilação dos registros palinológicos e paleobotânicos. Tais registros sugerem condições quentes e úmidas, para essas regiões, durante o Neógeno. O soerguimento dos Andes, na metade ocidental da América do Sul, criou um padrão de intensa precipitação orográfica, na Amazônia Ocidental, provocado pelas de massas de ar úmidas de origem atlântica. Durante este intervalo de tempo, o soerguimento do Planalto da Borborema, no norte brasileiro mais elevado, também formou uma barreira orográfica que permitiu o desenvolvimento da exuberante floresta no lado do Atlântico (Mata Atlântica), enquanto a ausência de chuva no flanco ocidental deu lugar ao semi-árido com a vegetação de Caatinga. As condições áridas foram ainda mais acentuadas pela evolução das correntes oceânicas do sistema Atlântico, que também foi responsável pela criação do deserto da Namíbia. Os dados palinológicos e paleobotânicos ilustram a longa história da floresta tropical da Amazônia, assim como a influência que recebeu pela elevação dos Andes. Os registros paleobotânicos do Nordeste brasileiro são escassos e indicam, pontualmente, longos períodos de clima umido que, na última parte do Neógeno, transformaram-se em condições predominantemente áridas que caracterizam a área até o momento.Palavras chave: climate, Mioceno, Plioceno, macrofitofósseis, palinomorfos
1. INTRODUCTION
Amazonia and the Brazilian Northeast are two areas of great climatic contrast. Although they are geographically close and both situated at -or near- the Equator, Amazonia is lush and densely forested, while the Brazilian Northeast is among the driest parts of the country. The present climatic conditions and the discrepancies in this region are determined by an interplay of geology, topography and atmospheric and oceanic circulation patterns. But, when and how were they established? How did global climatic change play a role in the evolution of this region? And what can palynological and paleobotanical evidence tell us about the climatic history of the region? These are the questions that we address in this chapter. We will do this by providing: a) an overview of timing and interplay of climate and geological processes that led to the present geographic configuration; b) present an inventory of the palynological and paleobotanical data from Amazonia and the Brazilian Northeast; c) make some inferences and recommendations based on this, and d) discuss the relation between the regional and global climate during the Neogene.
2. THE INTERPLAY OF GEOLOGICAL, OCEANIC AND ATMOSPHERIC PROCESSES
Modern climates in Amazonia and northeastern Brazil are dictated by their equatorial latitude and location in the causeway of the Intertropical Convergence Zone (ITCZ) (Figures 1 and 2). Other important factors that influence the climate in this region are orographic barriers such as the Andes and the Brazilian Highlands.

281Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra
Figure 1. A. Topographic map of Brazil (from NASA) with vegetation units of Amazonia and Northeast Brazil (modified after Pestana, 1994) (see legend in Fig. 2a); B. Geological map of Amazonia and Northeast Brazil shows the main geological units consiting of the (I) intracratonic Paleozoic Basins of the (II) Brazilian and (III) Guiana Shield and (IV) the coastal escarpment formed by the Borborema Plateau (modified after http://0.tqn.com/d/geology/1/0/t/I/brazilmap.gif)
The Andean Cordillera in the west reaches up to ca. 7000 m elevation and the presence of this mountain range plays a key role in the landscape and climate evolution of its wider periphery (Marengo et al., 2004; Vera et al., 2006; Veblen et al., 2007; Garreaud et al., 2009; Sepulchre et al., 2010). The meeting of the humid Atlantic air masses with the Andes barrier results in particularly high precipitation rates on the eastern flank of the Andes and in western Amazonia (Figure 2b). This configuration, and the high volumes of sediment and water discharge from the Andes, make

282 Paleontologia: Cenários de Vida – Paleoclimas
Amazonia the largest drainage basin in the world that contributes 20% of the total fresh water input into the world’s oceans (Goulding et al., 2003).
On the eastern side of the continent the northernmost Brazilian Highlands reach up to ca. 1200 m elevation and, albeit less dramatic, also form an orographic barrier to air masses entering from the Atlantic (Gottsberger & Silberbauer-Gottsberger, 2006) (Figures 1 and 2). This coastal escarpment prompts increased precipitation rates that enable the subsistence of the lush Atlantic rainforest or ‘Mata Atlantica’. However, the rain shadow, together with unpredictable interannual climatic variations, cause prolonged droughts in the landward flank which result in a gradual transition from dry (agreste) towards semi-desertic conditions (sertão) that give way to the Caatinga vegetation (Archibold, 1995; Behling et al., 2000).
Figure 2. A. Vegetation map of Amazonia and Northeast Brazil and oceanic currents (modified after Pestana, 1994); B. Precipitation patterns in Amazonia and Northeast Brazil (modified after Pestana, 1994).

283
Other crucial factors that determine the climatic evolution in this region are: a) periodic variations at orbital time scales caused by the changing position of the Earth around the Sun, b) interannual climatic variations caused by ocean currents, c) El Niño Southern Oscillation (ENSO), and d) the migration of the overland ITCZ and the South Atlantic Convergence Zone (SACZ) (Garreaud & Aceituno, 2007). These cyclic variations play a critical role in sea surface temperatures (SSTs), which in turn determine the length of the dry season and precipitation patterns on land (i.e. Behling et al., 2000; Kaandorp et al., 2005; Vonhof & Kaandorp, 2010) and are cause of the extreme droughts in the Brazilian Northeast (Gottsberger & Silberbauer-Gottsberger, 2006).
3. CENOZOIC EVOLUTION OF AMAZONIA AND THE BRAZILIAN NORTHEAST
During the Paleogene (65.5 – 23 Ma) global temperatures were relatively high and latitudinal variations small (Figure 3). Northern South America was characterized by a diverse rainforest ( Jaramillo et al., 2006; Jaramillo & Cardenas, 2013), which possibly extended into what is now Amazonia (Hoorn et al., 2010). During this time, and most of the Neogene, South America was a large island that gradually migrated northwards until colliding with the Caribbean Plate complex. In parallel, the reorganization of the Pacific plates caused uplift of the Andes and changed the topography and regional climate (Garzione et al., 2008; Shephard et al., 2010). Onset of uplift was dated as Oligocene (Parra et al., 2009) and last evidence of lowland conditions in Eastern Andes between Oligocene and early Miocene (Ochoa et al., 2012). This marked transition towards the modern relief and climate system.
Climate models coincide with this regional geological framework and since middle Miocene times an effective orographic barrier changed regional atmospheric patterns (Barnes et al., 2012; Poulsen et al., 2010; Insel et al., 2010) increasing precipitation and run-off with the modern precipitation regime in place around the middle Miocene (Kaandorp et al., 2005) (Figure 3).
The transition from a low to a high Andes extended well beyond the Andes itself and affected sedimentation patterns across South America even reaching the Atlantic coast (Dobson et al., 2001; Figueiredo et al., 2009). During this time a wetland system gradually extended over Western Amazonia, which during the late Miocene merged into the fluvial Amazon River system (Figueiredo et al., 2009). Climate, geography, sedimentary environments and biota in northern South America all were influenced by Andean uplift (Gentry, 1982; Antonelli et al., 2009; Hoorn et al., 2010), while global temperatures gradually declined (Zachos et al., 2008) and climatic change promoted erosion (Harris & Mix, 2002).
Eastern Amazonia and the Brazilian Northeast have a different geological history and form part of the Brazilian Shield, which is composed of ancient crystalline rocks. Geographically, the Brazilian Northeast is part of Plateau of Borborema (reaching up to 800 m) and the Chapada Diamantina (reaching up to 1200 m) that constitute the northern part of the Brazilian Highlands. These highlands -or Planalto Central- are thought to have a complex exhumation history that evolved in 4 phases since the opening of the Southern Atlantic (Cretaceous), with the youngest uplift dated as Miocene, coinciding with uplift in the Andes (Morais-Neto et al., 2009; Jepsen et al., 2012).
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

284 Paleontologia: Cenários de Vida – Paleoclimas
Figure 3. The global climate curve (from Zachos et al., 2008) compared with the main tectonic and oceanic events in Amazonia and Northeast Brazil (modified after Gottsberger & Silberbauer-Gottsberger, 2006).
The very different geological histories of Eastern and Western Amazonia also are reflected in edaphic processes and, indirectly, in plant composition. On young and unstable strata of Andean origin different soil types are formed than on the crystalline rocks of the shield where prolonged weathering rates are common (Horbe et al., 2004, 2007; Gottsberger & Silberbauer-Gottsberger, 2006; Young et al., 2007).
The regional climate history should be seen in the wider context of global climate change (see Figure 3; Zachos et al., 2008). During the early and middle Miocene (23 – 16 Ma) temperatures are relatively high, albeit lower than in the early part of the Paleogene. Global cooling and sea level fall set in during the late Miocene and act as prelude to the Quaternary ice ages.

285
4. AMAZONIA, EVIDENCE OF AN ANCIENT RAINFOREST AND A WARM AND HUMID CLIMATE
The geological history and the interaction of bedrock, soil and climate all influenced the composition and evolution of flora and fauna in Amazonia and the Brazilian Northeast. Although Neogene paleobotanical records of terrestrial plants are geographically sparse and fragmented they give us an insight into past vegetation and climate.
Palynological and paleobotanical remains are preserved in sedimentary basins such as the Acre foreland basin, the intracratonic Solimões and Amazonas basins (also known as Paleozoic sedimentary basins, see Wanderley Filho et al., 2010), the Marajo graben system (Rossetti et al., 2013) and the Foz do Amazonas basin (Figueiredo et al., 2009) (Figure 1a). The sedimentary infill of these basins consists of hundreds of meters of sand, clay and lignite of Neogene age. The subsurface and outcrop data points in Amazonia and the Brazilian Northeast are shown in Table 1 and Figure 4. Nearest living relatives of the plant fossils and their habitat are listed in Tables 2 - 6, and all pollen and spores recorded in Brazilian Amazonia are listed in Table 7; for ecology see Jaramillo et al. (2010) and for taxonomic references see the paleobiology database (http://paleodb.org/). A selection of characteristic pollen and spores and paleobotanical remains are shown in Figures 5 and 6. Works up to 2010 were reviewed by Jaramillo et al. (2010), but there have been some additions since then and which are reviewed below.
Table 1. Coordinates of all fóssil localities and relevant references.
N° LOCALITY STATE COORDINATES AUTHORS FOSSILS
1 Igarapé Waterfall AC 70°54’00,00’’W 08°58’60,00’’S Duarte & Japiassú (1971) Plant remains
Wood
2 Ouro Preto River ( Juruá River tributary ) AC 72°49’40,59’’W
08°23’47,24’’S Duarte & Japiassú (1971) Plant remainsWood
3 Rio Branco ( Juruá-Mirim River tributary) AC 73°13’35,83’’W
08°14’57,43’’S Duarte & Japiassú (1971) Plant remainsWood
4 Cruzeiro do Sul County, upstream Porto Peters AC 72°39’49,61’’W
07°38’53,07’’S
Hollick & Berry (1924); Berry (1935, 1937);
Maury (1937);Duarte (1970);
Duarte & Japiassú (1971);Machado et al. (2012).
LeafsWood
5 Sena Madureira(Purus River) AC 68°39’05,73’’W
09°04’44,35’’S Duarte & Japiassú (1971); Wood
6 Canutama County,Santa Rosa AM 64°24’51,92’’W
06°33’16,67’’S Duarte & Japiassú (1971); Wood
7 Lábrea County AM 64°48’8,46’’W 07°14’27,80’’S
Oliveira & Leonardos (1943);Duarte & Japiassú (1971); Leafs
8 São Paulo de Olivença County, Içá River AC 68°46’44,92’’W
03°21’15,21’’S Duarte & Japiassú (1971); Wood
9 Santo Antonio do Içá County, Ipiranga AC 67°54’42,78’’W
03°06’44,07’’SDolianitti (1948);
Duarte & Japiassú (1971) Caraceas
10 Capanema County PA 47°10’50,61’’W 01°10’16,61’’S Duarte & Japiassú (1971) Foliar print-outs
11 Irituia County(Ipixuna Formation) PA 47°26’40,12’’W
01°45’40,90’’SJapiassú (1969);
Duarte & Japiassú (1971) Leafs
12 São Luiz County(Anil District ) MA 44°14’03,01’’W
02°32’53,18’’S Duarte & Japiassú (1971) Wood
13 Humberto de Campos County MA 43°27’46,66’’W 02°35’11,70’’S Duarte & Japiassú (1971) Foliar print-outs
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

286 Paleontologia: Cenários de Vida – Paleoclimas
14 Nova Iorque Bed MA 44°03’00,00’’W 06°45’56,00’’S
Duarte & Japiassú (1971); Lima (1991);Cristalli (1997);
Cristalli & Bernardes-de-Oliveira (1998);
Anzótegui & Cristalli (2000);Dino et al. (2006).
PalynomorphsLeafs
15 Aracaju County (Barreiras Formation) SE 36°56’15,81’’W
10°42’36,71’’SMussa (1958);
Duarte & Japiassú (1971) Wood
16
Juazeiro County - Icó Farm;- Iço Lake;
- Parteiro Farm
BA 40°27’38,8’’W 09°24’59,13’’S
Guimarães (1964);Duarte & Japiassú (1971)
Plant remains Wood
17 Pedrão / Irará County BA 38°43’27,41’’W 12°05’15,47’’S
Berry (1924);Berry (1935);
Duarte & Japiassú (1971)Leafs
18 Alagoinhas – Aramari County BA 38°29’50,69’’W 12°04’34,01’’S
Hollick & Berry (1924);Duarte & Japiassú (1971) Leafs
19 Maraú County BA 39°02’10,81’’W 14°06’30,16’’S
Berry (1935);Hollick & Berry (1924);
Duarte & Japiassú (1971)Leafs
20 Maraú(Barretos / Camamu / João Branco) BA 39°06’56,73’’W
13°57’05,38’’S Duarte & Japiassú (1971) Foliar print-outsWood
21 Bom Jesus da Lapa County, Lapa BA 43°23’38,01’’W 13°14’26,74’’S
Oliveira (1937);Duarte & Japiassú (1971) Foliar print-outs
22 Solimões Formation(IAS 33 AM Well) AC 71°33’00,00’’W
05°15’00,00’’S Leite (2006) Palynomorphs
23 Solimões Formation(IAS 4a AM Well) AC 70°55’00,00’’W
04°23’00,00’’S Hoorn (1993) Palynomorphs
24 Novo Remanso Formation(Amazonas Basin / NS) 01) AM 60°32’02,00’’W
03°18’28,00’’S Dino et al. (2012) Palynomorphs
25 Novo Remanso Formation(Amazonas Basin / NS 05) AM 60°25’39,00’’W
03°17’58,00’’S Dino et al. (2012) Palynomorphs
26 Barreiras Formation PA 48°20’26,00’’W 01°03’37,00’’S Behling & Costa (2004) Palynomorphs
27 Outeiro Island(Barreiras Formation) PA 48°27’44,32’’W
01°15’09,09’’S
Leite (1997);Leite (2004);Leite (2006);
Arai et al. (1994); Truckenbrodt et al. (1994);
Dutra et al. (2001).
Palynomorphs Wood
28 Mosqueiro(Barreiras Formation) PA 48°28’21,07’’W
01°09’38,59’’S
Leite (1997);Leite (2004);Leite (2006);
Arai et al. (1994); Leite et al. (1997 a,b);
Truckenbrodt et al. (1994).
Palynomorphs
29 Atalaia(Barreiras and Pirabas formations) PA 48°17’13,73’’W
01°26’04,07’’S
Leite (1997);Leite (2004);Leite (2006);
Arai et al. (1994); Leite et al. (1997 a,b);
Truckenbrodt et al. (1994).
Palynomorphs
30 Capanema(Pirabas Formation) PA 47°10’50,61’’W
01°10’16,61’’S
Leite (1997);Leite (2004);Leite (2006);
Arai et al. (1994); Leite et al. (1997 a,b);
Truckenbrodt et al. (1994).
Palynomorphs
31 Amazon River, mouth PA 48°16’00,68’’W 03°18’14,62’’N Carvalho (1996) Palynomorphs
32 (IAS 51 AM Well) AC 69°02’00,00’’W 01°51’00,00’’S Leandro (2012) Palynomorphs

287
33 (IAS 52 AM Well) AC 69°29’00,00’’W 02°47’00,00’’S Leandro (2012) Palynomorphs
34 (IAS 27 AM Well) AC 67°55’00,00’’W 04°17’00,00’’S
Silva (2008);Silva-Caminha (2010); Leandro (2012) Palynomorphs
35 (IAS 19 AM Well) AC 69°10’00,00’’W 04°33’00,00’’S
Silva (2008);Silva-Caminha (2010); Leandro (2012) Palynomorphs
36 Feijó County AC 70°21’25.52”W 08°10’01.94”S Machado et al. (2012). Wood
37 Santa Rosa dos Purus County (Purus River) AC 70°29’34,00’’W
09°25’28,00’’S Machado et al. (2012). Wood
38 Niterói County AC 67°48’46,00’’W 10°04’14,00’’S Machado et al. (2012). Wood
39 Niterói County AC 67°48’47,20’’W 10°08’07,00’’S Mussa et al. (2002). Wood
40 Niterói County AC 67°48’46,00’’W10°08’07,00’’S Latrubesse et al. (2007) Palynomorphs
41 Rio Doce, outcrop AC 69°35’46.95”W10°57’48,51”S Latrubesse et al. (2007) Palynomorphs
42 IAS 32 AM Well AC 69°03’00,00’’W 01°52’00,00’’S Latrubesse et al. (2007) Palynomorphs
43 Coari County(P9 outcrop) AC 63°38’43,62’’W
3°54’22.61”S Silveira & Nogueira (2007). Palynomorphs
44 Coari County(P1 outcrop) AC 63°01’08,20’’W
04°03’52,68’’S Silveira & Nogueira (2007). Palynomorphs
45 Coari County(P2 outcrop) AC 63°03’25,36’’W
04°05’30,83’’S Silveira & Nogueira (2007). Palynomorphs
46 Coari County(P11 outcrop) AC 63°08’20,32’’W
03°59’49,67’’S Silveira & Nogueira (2007). Palynomorphs
47 Coari County(P12 Outcrop)
ACAC 63°06’31,52’’W
04°06’10,49’’S Silveira & Nogueira (2007). Palynomorphs
48 Coari County(P13 Outcrop) AC 63°08’58,26’’W
04°06’40,35’’S Silveira & Nogueira (2007). Palynomorphs
49 Capanema County(B17 Mine) PA 47°13’00,00’’W
1°10’00,00’’S Aguilera et al. (2013). Palynomorphs
50 Eirunepé village(Well and Outcrops) AC 69°46’11.70”W
06°33’55,50”S
Duarte & Japiassu (1971); Del’Arco et al. (1977);
Gross et al. (2011);Paz et al. (in press).
Foliar print-outsWood
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

288 Paleontologia: Cenários de Vida – Paleoclimas
Figure 4. Locations of all fossil sites reported in this chapter. Coordinates and authors are listed in Table 1.

289
Table 2. Habitat and ecology of the main taxa that were reported in the Cruzeiro do Sul County – Porto Peters, Juruá river valley (Point 4).
4. CRUZEIRO DO SUL COUNTY – PORTO PETERS, RIO JURUÁ RIVER VALLEY (Duarte, 1970)
EARLY MIOCENE HABIT HABITATFABACEAECassia
Machaerium
Tree 4 – 8m Deciduous, heliophytic, hygrophytes
Tree 8 – 14m.Semideciduous, heliophytic, xerophytes
Amazon region (AP and AM), common in the lower forest of the Amazonian riparian forest, flood plains and wetlands, capoeiras, clay soils wet or flooded
MA to PR, broadleaf semideciduous. Forest of the Paraná basin and in the pluvial Atlantic forest.
URTICACEAECoussapoa
Trees latescent (8 – 15m).Tree top, evergreen, heliophytics, hygrophytes
Pluvial Atlantic forest; coastal plain wood. Exclusive of Atlantic pluvial forest, in open areas or border of the jungle, along the watercourses.
LAURACEAEPersea
MespilodaphneOcoteaEndlichera
Bushes and trees (4 – 12m)Round and thin top, heliophytic, hygrophytes
Trees 10 – 25m.Semideciduous, heliophytic, xerophytes
TreesEvergreen, ciophyte, hygrophytes
South and southeast Brazil, ciliar woods and wet flood-plains, mainly in higher regions and in swampy soils.
RJ, SP, MG – Cerrado, high fields of the Mantiqueira Hill and Mata dos Pinhais, Pr to RS. Draw off soils.
HERNANDIACEAESparattanthelium
Bushes 3 – 5m de high, climbing plantsSemideciduous, heliophytic, hygrophytes
CE to RJ, pluvial Atlantic forest, Restinga and board. Arboreous Restinga, NE coast.
EUPHORBIACEAESapium
Trees 6 – 12m.Semideciduous, globous shape, branches near the ground, heliophytic
MG, GO, MT, MS to RS; semideciduous woods from Paraná basin and Uruguay, argillaceous ground, rich in organic matter, wet floodplains and river banks.
VOCHYSIACEAEVochysia
Trees of 7 – 24mEvergreen, dense globous shape, heliophytic, hygro-phytes, xerophytes
Rainforest Atlantic slope and coastal plain SP and RJ and Central Brazil; in savannah 800 m - 1000 m altitude. Well-drained slope land
BIGNONIACEAEBignonites Bushes or trees 4 – 10m Coastal wetlands MT, GO, MG, AM, BA, PE, southern
MA, PI, PA, in Cerrado.
Table 3. Habitat and ecology of the main taxa that were reported in the Pirabas Formation, Capanema County (Point 10).
10. PIRABAS FORMATION – CAPANEMA, PA (DUARTE, 1972, 2004)
EARLY MIOCENE HABIT HABITATANNONACEAEGuatteria Evergreen, heliophytic to ciophytic, xerophytes Amazon rainforest - firm ground
LAURACEAEEndlicheria Evergreen, ciophytic, hygrophytes Rainforest of the Atlantic hillside
DILLENIACEAEDavilla Liana Mainly Cerrado, central Brazil
BONNETIACEAEBonnetia Bushes Amazon rainforest: Archytaea
Coastal dunes from Bahia: BonnetiaCARYOCARACEAECaryocar
TreesEvergreen, heliophytic, hygrophytes; riparian forest Amazon region (AM, PA, AP)
CRYSOBALANACEAEHirtella
Trees or bushesSemideciduous, heliophytic to ciophytic, xerophyte
Margins of savannahs of the Amazon or Central Brazil; Riparian forest, capoeiras, land slopes
PUTRANJIVACEAE Drypetes
TreesEvergreen, ciophytic or heliophytic, xerophyte Atlantic rainforest of the coast of Bahia on high slopes
RUTACEAEHostia
Bushes or treesEvergreen, heliophytic, xerophyte
Atlantic rainforest and slope; RJ and MG semidecidu-ous altitude forest
MELIACEAETrichilia
TreesEvergreen, esciophytic, hygrophyte
Atlantic rainforest (200-600m); early slopes - soil moist and deciduous to semi-deciduous forests.
SAPINDACEAESerjania______________________Sapindus
Lianas_________________________________Evergreen, heliophytic or semideciduous
Borders of tropical and subtropical forests moist to semi-arid, north to southern Brazil_______________________________Amazon floodplains of low and semi-deciduous broad-leaf forest
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra
290 Paleontologia: Cenários de Vida – Paleoclimas
MALVACEAEApeiba
Herbs of treesEvergreen, heliophytic
The Amazon region until MG and SP. Rainforest to semi-deciduous broadleaf forest
MYRTACEAEMyrcia Evergreen, heliophytic, ciophytic, hygrophytes Rainforest on the Atlantic slope of hillside land
RHIZOPHORACEAECassipourea
Bushes or treesEvergreen
Amazon. Forests, plains, wetlands, swamps in tropical and subtropical areas of flooding.
EBENACEAEDiospyros
Evergreen, semideciduous and deciduous, helio-phytic, hygrophyte to xerophytes
Cerrado, semideciduous broadleaf forest in central Brazil, northeast and south
RUBIACEAEFaramea Herbs, bushes, ciophytic Cerrado, riparian rainforest, rupestrian fields, montane
and high montane Atlantic rainforest RAPATACEAERapatea Aquatic or swampy herbs Associated with the border of creeks in the Amazon.
Table 4. Habitat and ecology of the main taxa that were reported in the Nova Iorque Bed, Cenozoic coverage of the Parnaíba Basin - Nova York Village - Margin of the Parnaíba River (Point 14).
14. NOVA IORQUE BED , CENOZOIC COVERAGE OF THE PARNAÍBA BASIN - NOVA YORK VILLAGE (MA) - MARGIN OF THE PARNAÍBA RIVER. Cristalli (1997); Cristalli & Bernardes de Oliveira (1998) e Anzotegui & Cristalli (2000).
EARLY MIOCENE HABIT HABITATLAURACEAELaurophyllum sp.
Trees or shrubsEvergreen plants
Tropical and subtropical. Rainforests of the Americas and Asia. Common in the Amazon and Atlantic forest.
MALVACEAE Mostly herbs, bushes, lianas or trees Pantropical - cosmopolitanEBENACEAEcf. Diospyros sp. orLisfocarpa sp.
Woody plants arboreal or shrub of 4 – 7m high Savannahs from MA, PI, CE to MS and PR.
FABACEAECaesalpinoideae cf. Caesalpinoideae sp.
Trees of 8 – 30 mSemideciduous, heliophytic, hygrophytes or dry lands, humid lowlands
Typical of the inside of the forest, occurs in the pluvial Atlantic forest.
FABACEAECaesalpinoideae cf. Cassia sp.
Trees of 8 – 20 mUsually deciduous, heliophytic
Araucarian forest and Atlantic forest, in the Amazon region, solid land forest.
FABACEAEFaboideaecf. Dalbergia sp.
Trees de 4 – 16mEvergreen, heliophytics or ciophytes,. Occurs in deep valleys, clay and moist soils.
Atlantic slope rainforest and in the pine forests woods.
FABACEAEFaboideaecf. Cassavalia sp.
Leguminose, climbing plants
Pantropical with 60 species, 30 of which in tropical America, subtropical NE. In Brazil occurs in the N, NE and central-west regions and in the Amazonia, Caatinga and Mata Atlântica biomes.
MELIACEAEcf. Cedrela sp. Bushes or trees
Pantropical. Tropical America from N to S. Well-drained habitats, from lowland rainforests or mountains in decidual forests
SMILACACEAEcf. Smilax sp. Herbaceous or woody lianas Tropical
MELASTOMATACEAEMiconia sp.
or
Meriania sp.
Trees of 15 – 22mEvergreen, heliophytic, single leaves, coriaceous and glabrous.
Trees or bushes, single leaves rarely verticilated.
Pluvial Atlantic forest in wet soils. From BA until SC.
Exclusively from the S and SE Brazil and pluvial Atlantic forest From RJ to SC – low lands until high fields .
MYRTACEAE Trees or bushes, single leaves rarely verticilated. Pantropical or subtropical, concentrated in Neotropical and Australia. Atlantic forest and Restinga.
RUTACEAE ? Bushes or trees Pantropical. Seasonal forests
CLUSIACEAE Bushes or treesPantropical. Floodplain forests. Disperses around Brazil: Cerrados, Restingas, etc.
Nova Iorque Beds (MA): humid tropical climate - evergreen leaves vegetation, with entire margins leaves and acuminate apex.

291
Table 5. Habitat and ecology of the main taxa that were reported in the Pedrão / Irará County (Point 17).
17. PEDRÃO/IRARÁ COUNTY – PLIOCENE, Berry (1924, 1935).
FAMILY AND GENERA HABIT HABITAT
POLYGONACEAECoccoloba
Trees, shrubs, herbs or lianasHeliophytic, hygrophytes
Floodplain forest and flooded woods in the Amazonian pluvial forest or latifoliated riparian wood, semideciduous of the central-western Brazil
Table 6. Habitat and ecology of the main taxa that were reported in the Mocambo / Ouriçanga / Aramari / Maraú County (Point 19).
19. MOCAMBO / OURIÇANGA / ARAMARI / MARAÚ COUNTY – PLIOCENE (Berry, 1935)
FAMILY AND GENERA HABIT HABITAT
CANELLACEAECanella Trees, rarely bushes Neotropical. No occurrence in Brazil, only
Cinnamodendron.
CLUSIACEAEGarcinia (=Rheedia)
High trees, evergreen, heliophytic, hygrophytes. Requiring humid soils.
Typically from Amazonia and Mata Atlantica from sea level until 1500m altitude in the ombrophyllous dense forest.
4.1 Acre Basin
Latrubesse and colleagues (2007) first documented the palynological content of strata exposed along the Acre River at Barranco da Elizete, Murici, Niteroi and Patos (40-41, Figure 4). The authors concluded that the age of these deposits is late Miocene and the environment was characterized by freshwater lakes and swamps with grasslands and gallery forests and fluctuating water levels implying a tropical to tropical to wet-dry tropical seasonal climate.
Berry (1935, 1937), Maury (1937) and Duarte (1970), all studied plant macrofossils from Cruzeiro do Sul (Alto Juruá), Sena Madureira and Rio Branco (Acre State) (4-5, Figure 4) from sediments of estimated Pliocene age, and with occurrences of Fabaceae, Urticaceae, Lauraceae, Rosaceae, Sapindaceae, Vochysiaceae, Hernandiaceae and Bignoniaceae. Berry (1937) further observed plant fragments from clay beds at Tgaraje Agua Fuente, a tributary of the Rio Moa and refers to these strata as Pebas Beds. He reports the following families: Coussapoa (Moraceae), Cassia aguaytiensis (Caesalpiniaceae), Machaerium prematicum (Papilionaceae), Vochysia acuminatafolia (Vochysiaceae), Sapium pliocenicum (Euphorbiaceae), Sparattanthelium pliocenicum (Hernandiaceae), Mespilodaphne acrensis and Persea amoneana (Lauraceae), Bignonites mauryae (Bignoniaceae), Endlichera rhamnoides (Rubiaceae). These taxa, and the species illustrated in Duarte’s (1970) paper, are all suggestive of warm and wet tropical conditions.
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

292 Paleontologia: Cenários de Vida – Paleoclimas
Figure 5. Characteristic pollen and spores from the Solimões Formation (Neogene Amazonia). 1- Crassoretitriletes vanraadshoovenii; 2- Deltoidospora adriennis; 3- Magnastriatites grandiosus; 4- Perimonoletes sp.; 5- Verrucatosporites ssp.; 6- Polypodiaceiosporites potoniei; 7- Echitriletes muellerii; 8- Mauritiidites franciscoi; 9- Grimsdalea magnaclavata; 10- Echiperiporites stelae; 11- Fenestrites spinosus; 12- Echitricolporites spinosus; 13- Echitricolporites maristellae; 14- Retitriporites dubiosus; 15- Margocolporites aff. vanwijhei; 16- Crototricolpites sp.; 17- Crassiectoapertites colombianus; 18- Psilastephanoporites tesseroporus; 19- Perisyncolporites pokornyi; 20- Retistephanoporites crassiannulatus; 21- Bombacacidites bellus; 22- Bombacacidites baculatus; 23- Multimarginites vanderhammeni; 24- Perfotricolpites digitatus; 25- Retimonocolpites maximus; 26- Multiareolites formosus; 27- Rhoipites guianensis; 28- Psilatricolporites caribbiensis; 29- Jandufouria seamrogiformis; 30- Polyadopollenites mariae (modified from Leite, 2006).
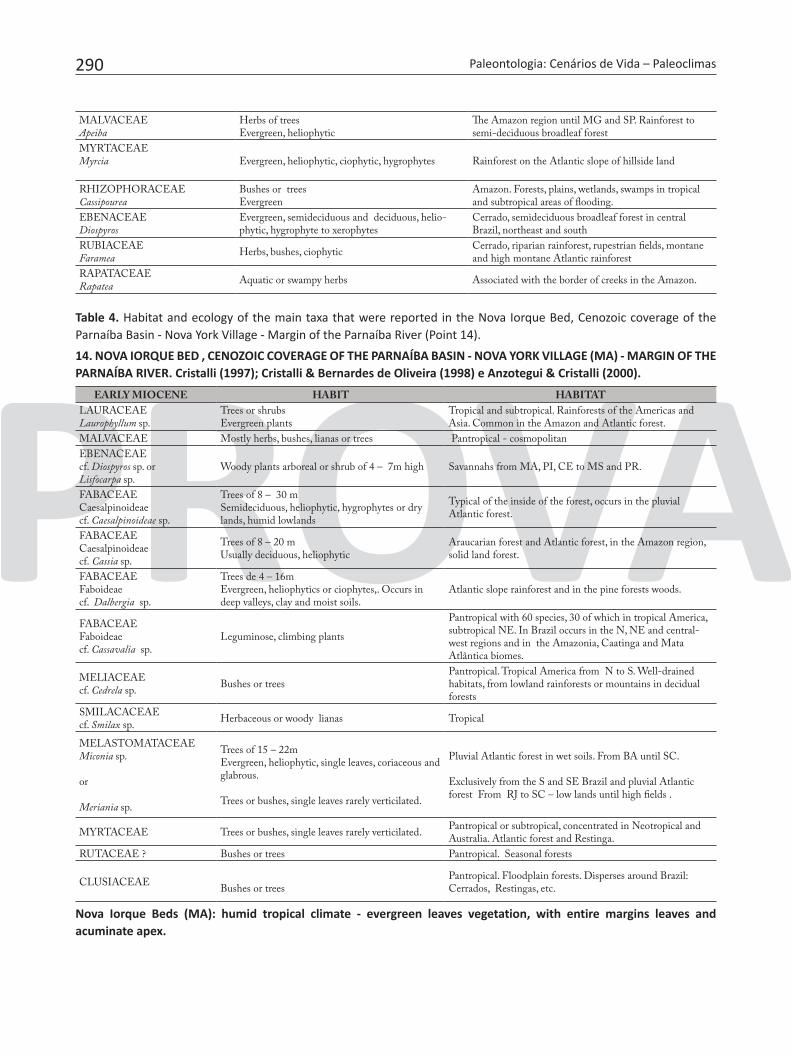
293
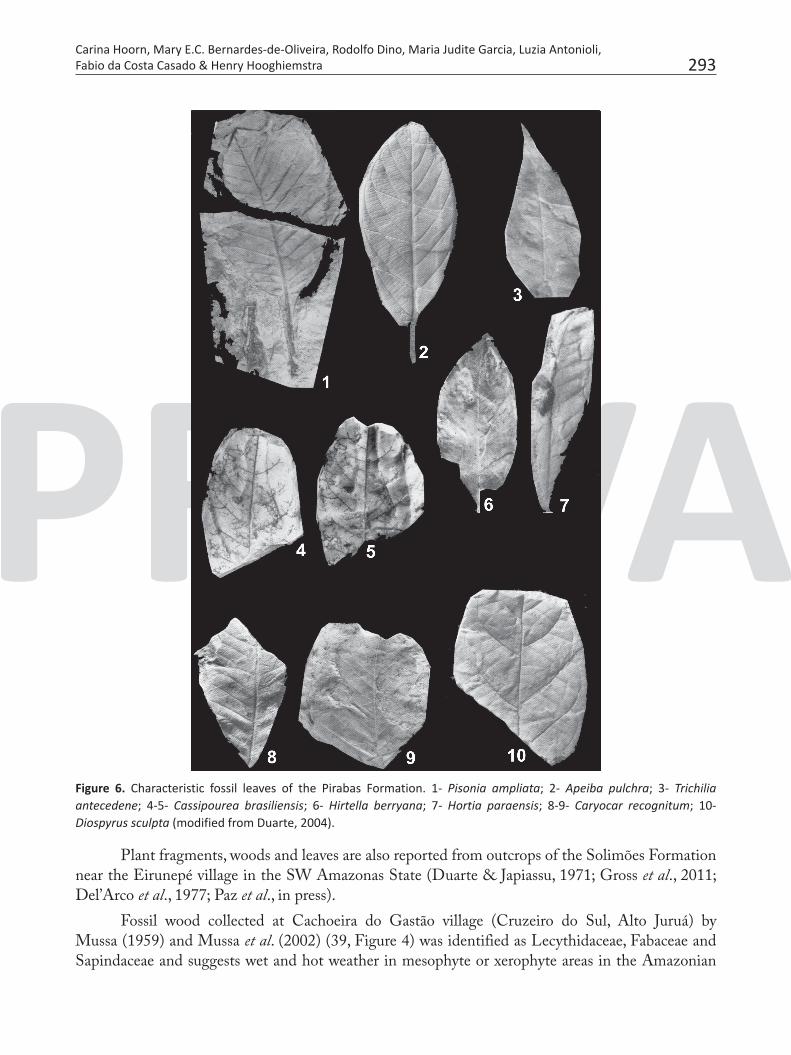
Figure 6. Characteristic fossil leaves of the Pirabas Formation. 1- Pisonia ampliata; 2- Apeiba pulchra; 3- Trichilia antecedene; 4-5- Cassipourea brasiliensis; 6- Hirtella berryana; 7- Hortia paraensis; 8-9- Caryocar recognitum; 10- Diospyrus sculpta (modified from Duarte, 2004).
Plant fragments, woods and leaves are also reported from outcrops of the Solimões Formation near the Eirunepé village in the SW Amazonas State (Duarte & Japiassu, 1971; Gross et al., 2011; Del’Arco et al., 1977; Paz et al., in press).
Fossil wood collected at Cachoeira do Gastão village (Cruzeiro do Sul, Alto Juruá) by Mussa (1959) and Mussa et al. (2002) (39, Figure 4) was identified as Lecythidaceae, Fabaceae and Sapindaceae and suggests wet and hot weather in mesophyte or xerophyte areas in the Amazonian
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

294 Paleontologia: Cenários de Vida – Paleoclimas
rainforest. Duarte & Japiassú (1971) (6-8, Figure 4) refer to petrified woods and leaf compressions of Pliocene age at the localities of Canutama, Lábrea and São Paulo de Olivença. However, there is no taxonomic details for these plant remains.
Machado et al. (2012) (4, Figure 4) also collected wood at outcrops of the Solimões Formation along the Juruá, Acre, Furnaia, Purus and Moa rivers and identified them as Fabaceae (Caesalpinioideae), Lythraceae, and Myrtaceae. The specimens show anatomical features that indicate predominantly hot and humid climate with seasonal fluctuations in water availability and a dry season. It is worth mentioning that some of the large angiosperm fossils that were collected from outcrops of the Solimões Formation situated along the Branco River are still awaiting identification and form part of the paleobotanical collection at Guarulhos University (Brazil).
4.2 Solimões Basin
The Neogene strata in the Solimões and Amazonas basins are known as Pebas Formation (Peru) and Solimões Formation (Maia et al., 1977; Hoorn 1993; Leguizamon-Vega et al., 2006; Wanderley Filho et al., 2010). A series of drill cores taken by Companhia do Pesquisas e Recursos Minerais (CPRM) recovered hundreds of meters of Neogene sediments which formed the basis of several palynological studies (Hoorn, 1993; Leite, 2006; Silva, 2004, 2008; Latrubesse et al., 2007; Silva-Caminha et al., 2010; Leandro, 2012) (22-23, 32-35, 42, Figure 4) in which hundreds of pollen taxa were identified (Table 7) that are illustrative for the diversity of the Neogene rainforest.
Table 7. Pollen and spores recorded in the Neogene sediments of Brazilian Amazonia.
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
POLLEN
Alnipollenites verus x x x
Anacolosidites cf. luteoides x
Araucariacidites sp. x
Arecipites perfectus x x
A. polaris x
A. regio x
Bombacacidites spp. x x x x x x x
B. araracuarensis x
B. baculatus x x x x x x
B. baumfalki x x x
B. bellus x x x x x
B. brevis x
B. ciriloensis x
“B. eckfeldensis” x
B. fossulatus x x
B. muinaneorum x x
B. nacimentoensis x
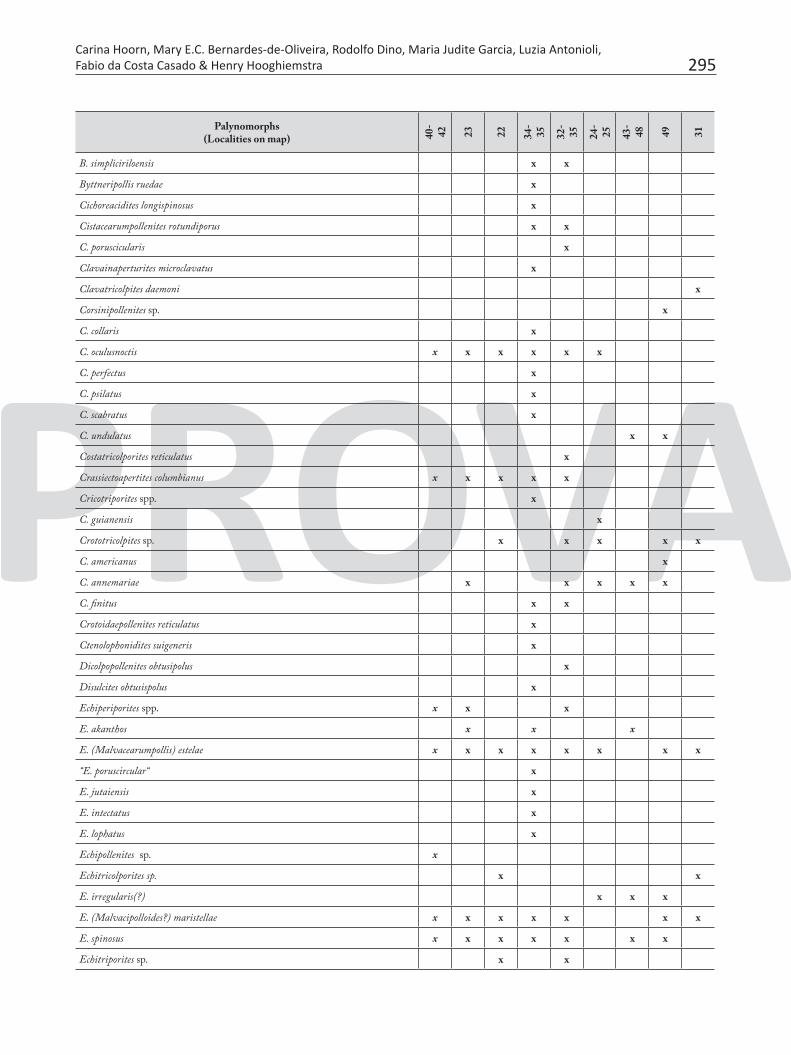
295
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
B. simpliciriloensis x x
Byttneripollis ruedae x
Cichoreacidites longispinosus x
Cistacearumpollenites rotundiporus x x
C. poruscicularis x
Clavainaperturites microclavatus x
Clavatricolpites daemoni x
Corsinipollenites sp. x
C. collaris x
C. oculusnoctis x x x x x x
C. perfectus x
C. psilatus x
C. scabratus x
C. undulatus x x
Costatricolporites reticulatus x
Crassiectoapertites columbianus x x x x x
Cricotriporites spp. x
C. guianensis x
Crototricolpites sp. x x x x x
C. americanus x
C. annemariae x x x x x
C. finitus x x
Crotoidaepollenites reticulatus x
Ctenolophonidites suigeneris x
Dicolpopollenites obtusipolus x
Disulcites obtusispolus x
Echiperiporites spp. x x x
E. akanthos x x x
E. (Malvacearumpollis) estelae x x x x x x x x
“E. poruscircular“ x
E. jutaiensis x
E. intectatus x
E. lophatus x
Echipollenites sp. x
Echitricolporites sp. x x
E. irregularis(?) x x x
E. (Malvacipolloides?) maristellae x x x x x x x
E. spinosus x x x x x x x
Echitriporites sp. x x
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra
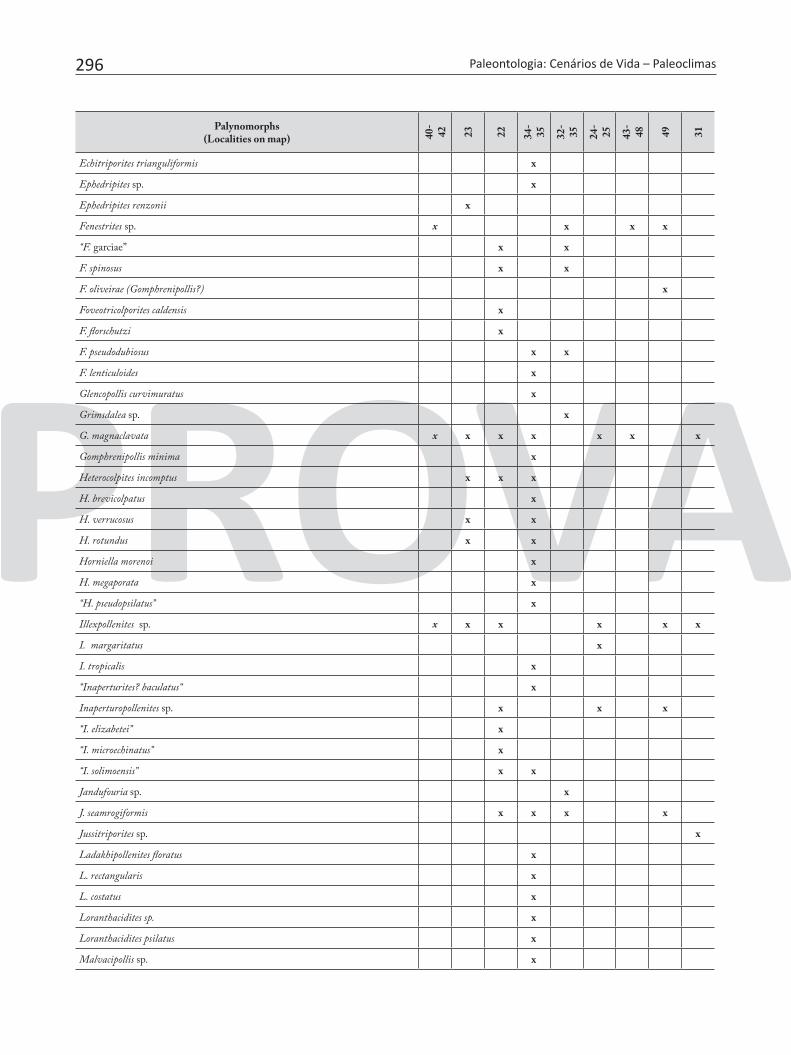
296 Paleontologia: Cenários de Vida – Paleoclimas
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
Echitriporites trianguliformis x
Ephedripites sp. x
Ephedripites renzonii x
Fenestrites sp. x x x x
“F. garciae” x x
F. spinosus x x
F. oliveirae (Gomphrenipollis?) x
Foveotricolporites caldensis x
F. florschutzi x
F. pseudodubiosus x x
F. lenticuloides x
Glencopollis curvimuratus x
Grimsdalea sp. x
G. magnaclavata x x x x x x x
Gomphrenipollis minima x
Heterocolpites incomptus x x x
H. brevicolpatus x
H. verrucosus x x
H. rotundus x x
Horniella morenoi x
H. megaporata x
“H. pseudopsilatus” x
Illexpollenites sp. x x x x x x
I. margaritatus x
I. tropicalis x
“Inaperturites? baculatus“ x
Inaperturopollenites sp. x x x
“I. elizabetei” x
“I. microechinatus” x
“I. solimoensis” x x
Jandufouria sp. x
J. seamrogiformis x x x x
Jussitriporites sp. x
Ladakhipollenites floratus x
L. rectangularis x
L. costatus x
Loranthacidites sp. x
Loranthacidites psilatus x
Malvacipollis sp. x
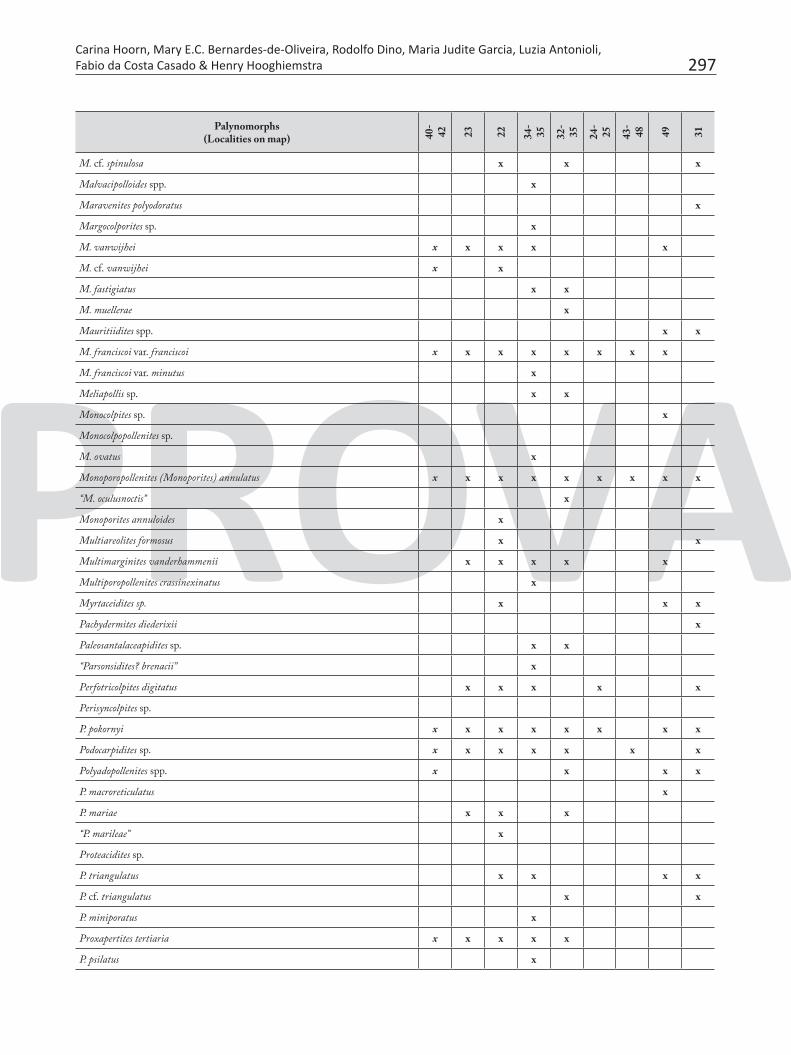
297
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
M. cf. spinulosa x x x
Malvacipolloides spp. x
Maravenites polyodoratus x
Margocolporites sp. x
M. vanwijhei x x x x x
M. cf. vanwijhei x x
M. fastigiatus x x
M. muellerae x
Mauritiidites spp. x x
M. franciscoi var. franciscoi x x x x x x x x
M. franciscoi var. minutus x
Meliapollis sp. x x
Monocolpites sp. x
Monocolpopollenites sp.
M. ovatus x
Monoporopollenites (Monoporites) annulatus x x x x x x x x x
“M. oculusnoctis” x
Monoporites annuloides x
Multiareolites formosus x x
Multimarginites vanderhammenii x x x x x
Multiporopollenites crassinexinatus x
Myrtaceidites sp. x x x
Pachydermites diederixii x
Paleosantalaceapidites sp. x x
“Parsonsidites? brenacii” x
Perfotricolpites digitatus x x x x x
Perisyncolpites sp.
P. pokornyi x x x x x x x x
Podocarpidites sp. x x x x x x x
Polyadopollenites spp. x x x x
P. macroreticulatus x
P. mariae x x x
“P. marileae” x
Proteacidites sp.
P. triangulatus x x x x
P. cf. triangulatus x x
P. miniporatus x
Proxapertites tertiaria x x x x x
P. psilatus x
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

298 Paleontologia: Cenários de Vida – Paleoclimas
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
Psilabrevitricolporites sp. x
“P. hoornii” x x
Psilabrevicolpites flexibilis x
Psiladiporites sp. x
P. redundantis x
P. minimus x x
Psilamonocolpites spp. x
P. amazonicus x
P. grandis x
P. nanus x x
P. medius x
P. rinconii x x
Psilaperiporites spp. x x x x
P. minimus x x
“P. elizabetei”
P. multiporatus x
Psilastephanocolporites spp. x x
P. fissilis x x x
P. cf. fissilis x
P. schneideri x
P. marinamensis x
Psilastephanoporites sp. x x x
P. herngreenii x x x
P. cf. stelatus x
P. tesseroporus x x x
Psilatricolpites spp. x
P. minutus x x
Psilatriporites spp. x
P. desilvae x x
P. corstanjei x
P. sarmientoi x x
Psilatricolporites spp. x x x x x
P. acerbus x x x
P. atalayensis x x
P. (Horniella) caribbiensis x x x x
P. costatus x
P. crassoexinatus x
P. crassus (Lanagiopollis crassa) x x
P. cyamus x

299
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
P. (Psilabrevitricolporites) devriesii x x x
P. divisus x
P. exiguus x
P. garzonii x
P. labiatus x x x
P. magniporatus x x
P. obesus x
P. (Ranunculacidites) operculatus x x x x x x x
P. (R.) cf. operculatus x x
P. pachydermatus x x
P. silvaticus x x
Psilatricolporites (Tetracolporopollenites) transversalis x x
P. (Psilabrevitricolporites) triangularis x x x x
P. vanus x x x
P. cf. varius x
P. venezuelanus x
Punctodiporites harrisii x
Retibrevitricolpites spp. x x
R. retibolus x
R. catatumbus x
R. cf. catatumbus x
R. yavarensis x x
Retibrevitricolporites spp. x
Retimonocolpites maximus x x
R. absyae x
Retistephanoporites sp. x
R. crassiannulatus x x x x
Retitricolpites spp. x x x
R. antonii x
R. amapaensis x x x x
R. colpiconstrictus x
R. conciliatus x
R. depressus x
R. florentinus x
R. perforatus x
R. simplex x x x
R. cf. simplex x
R. cf. grandis x
R. speciosus x
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra
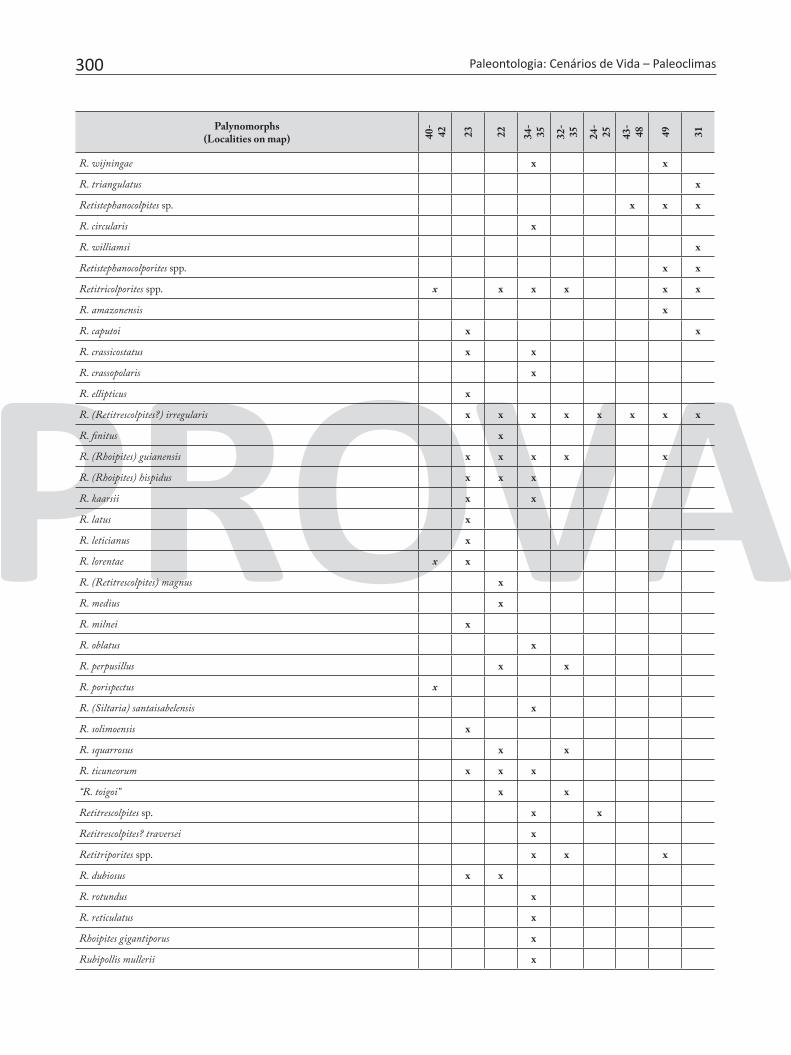
300 Paleontologia: Cenários de Vida – Paleoclimas
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
R. wijningae x x
R. triangulatus x
Retistephanocolpites sp. x x x
R. circularis x
R. williamsi x
Retistephanocolporites spp. x x
Retitricolporites spp. x x x x x x
R. amazonensis x
R. caputoi x x
R. crassicostatus x x
R. crassopolaris x
R. ellipticus x
R. (Retitrescolpites?) irregularis x x x x x x x x
R. finitus x
R. (Rhoipites) guianensis x x x x x
R. (Rhoipites) hispidus x x x
R. kaarsii x x
R. latus x
R. leticianus x
R. lorentae x x
R. (Retitrescolpites) magnus x
R. medius x
R. milnei x
R. oblatus x
R. perpusillus x x
R. porispectus x
R. (Siltaria) santaisabelensis x
R. solimoensis x
R. squarrosus x x
R. ticuneorum x x x
“R. toigoi” x x
Retitrescolpites sp. x x
Retitrescolpites? traversei x
Retitriporites spp. x x x
R. dubiosus x x
R. rotundus x
R. reticulatus x
Rhoipites gigantiporus x
Rubipollis mullerii x
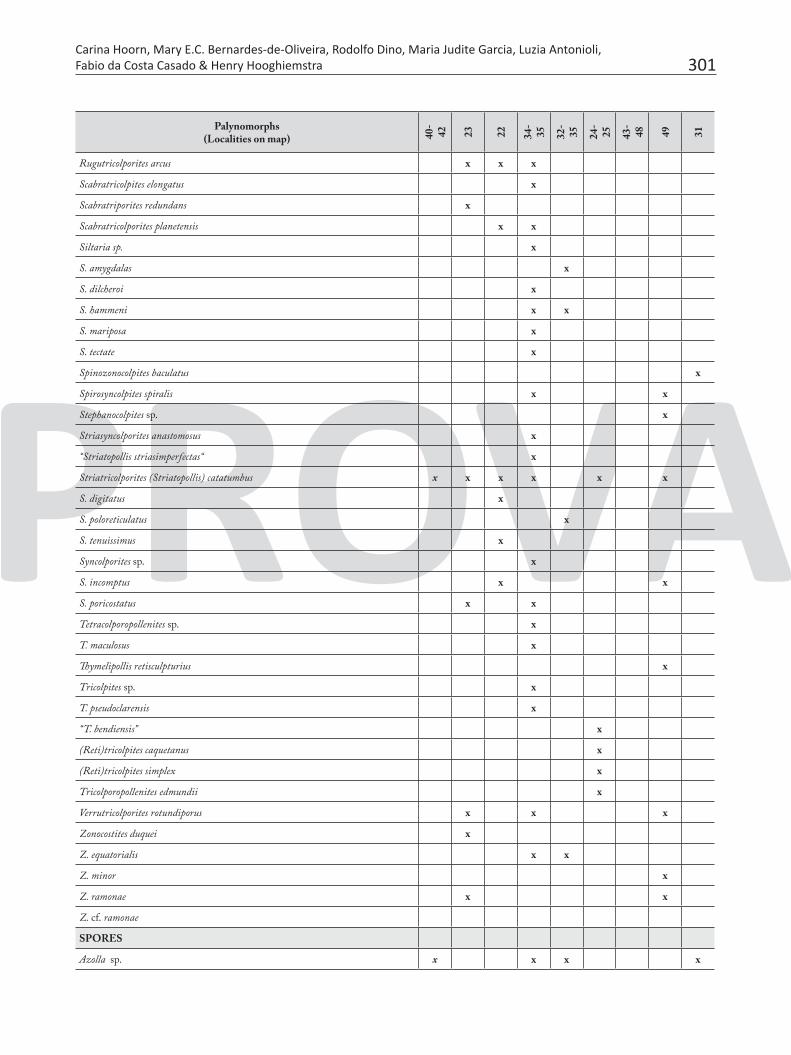
301
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
Rugutricolporites arcus x x x
Scabratricolpites elongatus x
Scabratriporites redundans x
Scabratricolporites planetensis x x
Siltaria sp. x
S. amygdalas x
S. dilcheroi x
S. hammeni x x
S. mariposa x
S. tectate x
Spinozonocolpites baculatus x
Spirosyncolpites spiralis x x
Stephanocolpites sp. x
Striasyncolporites anastomosus x
“Striatopollis striasimperfectas“ x
Striatricolporites (Striatopollis) catatumbus x x x x x x
S. digitatus x
S. poloreticulatus x
S. tenuissimus x
Syncolporites sp. x
S. incomptus x x
S. poricostatus x x
Tetracolporopollenites sp. x
T. maculosus x
Thymelipollis retisculpturius x
Tricolpites sp. x
T. pseudoclarensis x
“T. bendiensis” x
(Reti)tricolpites caquetanus x
(Reti)tricolpites simplex x
Tricolporopollenites edmundii x
Verrutricolporites rotundiporus x x x
Zonocostites duquei x
Z. equatorialis x x
Z. minor x
Z. ramonae x x
Z. cf. ramonae
SPORES
Azolla sp. x x x x
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

302 Paleontologia: Cenários de Vida – Paleoclimas
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
Bacutriletes sp. x
Chomotriletes minor x x
Cicatricosisporites sp. x x x x x
Cingulatisporites sp. x
C. laevigatus x
C. rugulatus x
Clavatriletes sp. x
Concavisporites sp. x
“Concavissimisporites cingulatus” x
Crassoretitriletes sp. x
C. vanraadshoovenii x x x x x x x
Cyathidites sp. x
C. annulatus x x
Deltoidospora adriennis x x x x x x x x
Deltoidospora sp. x
Distaverrusporites margaritatus x x
Echinatisporis circularis x
E. muelleri x
Echitriletes sp. x x x
“Echitriletes clavatus” x
E. muellerii x x x x x
E. cf. muellerii x
“Foveomonoletes puntatus“ x
“Fossutriletes bollatus“ x
Foveotriletes ornatus x x
Hamulatisporites sp. x
Hydrosporis minor x
Kuylisporites waterbolkii x x x
Laevigatosporites sp. x x x
L. catanejensis x
L. tibuensis x
Leiotriletes adriennis
Leptolepidites sp. x
Magnastriatites sp. x
M. grandiosus x x x x x x x x
Matonisporites sp. x x x
M. mulleri x
Microfoveolatisporites sp. x x
Perinomonoletes sp. x x

303
Palynomorphs(Localities on map) 40
-42 23 22 34
-35 32
-35 24
-25 43
-48 49 31
Perotriletes sp. x
Polypodiaceoisporites spp. x x x
P. amazonensis x
P. potonie x x x x x
P. pseudopsilatus x
Polypodiisporites aff. specious x
P. scabraproximatus x
Polypodiisporites? planus x
Psilamonoletes spp.? x x
P. tibui
Psilatriletes sp. x x x x x
P. peruanus x x
P. lobatus x
Pteridaceoisporis gemmatus x
Retitriletes sp. x x
R. altimuratus x
Rugulatisporites sp. x
Rugutriletes circulatus x
Tuberositriletes? crassus x
Verrucatosporites spp. x x x x
V. (Polypodiisporites) usmensis x x x x x x
Verrucatotriletes sp. x
V. cf. bullatus x x
V. cf. baculatus x
V. etayoi x
Verrutriletes spp. x x x x
Verrumonoletes sp. x x
ALGAE
Pediastrum x x x
Ovoidites x x x
Botryococcus x x x
Leandro (2012) reports on study of two cores (1AS-51-AM and 1AS-52-AM) situated in the Jandaiatuba sub-basin (Solimões Basin, NW Amazonas State). Palynological zonations range from Psilodiporites minimus to Alnipollenites verus and suggest an age of early Miocene to Pleistocene. All authors coincide that tropical rainforest conditions existed in the area during the Neogene. Leandro further suggests that an increase in palynological diversity was observed during the Quaternary. However, it is not clear if sedimentary conditions or climate are responsible for this.
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

304 Paleontologia: Cenários de Vida – Paleoclimas
4.3 Amazonas Basin
Palynological and paleobotanical remains are rare in the Amazonas Basin. Dino et al. (2012) and Dino et al. (in prep.) report outcrops of the Novo Remanso Formation between the towns Manacapuru and Iranduba towns (24-25, Figure 4) and at the banks of the Uatumã River. They estimate the age of the sediments as middle Miocene and in these fluvial sediments they report a palynoflora containing spores of Lycopodiaceae, Selaginellaceae, Cyatheaceae, Gleicheniaceae, and Polypodiaceae, and pollen from Arecaceae (Mauritia and others), Aquifoliaceae, Onagraceae (Ludwigia; formerly Jussiaea), Euphorbiaceae, Malvaceae, Poaceae, Fabaceae and Malpighiaceae (among others). This assemblage is representative of a tropical rainforest that dwelled in hot and humid weather conditions. Noticeably, a pollen grain with as nearest living relative Araucaria was reported. Araucaria is now restricted to areas outside the humid tropical lowlands and known from forests of higher southern latitudes. Wood fossils of the Novo Remanso Formation were collected near the Manacapuru locality and are currently being analyzed by Pons.
Silveira & Nogueira (2007) also report on a series of late Miocene and Pleistocene outcrops along the Amazon River but make no paleoclimatic inferences. These outcrops are situated in the Coari region, west of Manaus and are about 20 m high each.
4.4 Marajó and the Foz do Amazonas Basin
In eastern Amazonia Rossetti & Goes (2004), Rossetti et al. (2013) and Aguilera et al. (2013) -among others- described the coastal carbonate and siliciclastic deposits of the Pirabas Formation (early Miocene) and the siliciclastic, fluvial-tidal deposits of the Barreiras Formation (middle Miocene) (27-30; 49, Figure 4). In Foz do Amazonas Basin, at the Atlantic coast, time equivalent units were described (Figueiredo et al., 2009). In an earlier study Carvalho (1996) documented the palynological potential of the Plio-Pleistocene section in this basin. Assemblages were assigned by quantifying biodiversity and he found that glacial period had higher abundance of montane taxa, whereas interglacial periods were richer in lowland taxa. Richards & Lowe (2002) add that late Pliocene sediments in the Foz do Amazonas have abundant Podocarpus, Alnipollenites verus, and charred Poaceae (grass) that are indicative of periods of mostly dry climate and frequent fires. They also suggest that high cuticle abundance may coincide with sea level lowstands and the possible existence of “El Niño” conditions in the pre-Quaternary.
The Miocene palynofloras of the Pirabas and Barreiras formations in the Bragantina region were studied by Arai et al. (1994), Truckenbrodt et al. (1994), Leite (1997, 2004), Leite et al.(1997 a,b) and Aguilera et al. (2013) (27-30, 49, Figure 4). Leite (1997, 2004) and Leite et al.(1997 a,b) described a coastal plain environment with mangroves that gradually changed into a beach ridge setting with a vegetation that grew under hot and humid climatic conditions.
In Eastern Amazonia (Pará, Maranhão, Amapá, Tocantins and Mato Grosso States), Neogene paleobotanical remains were found in the Miocene Pirabas Formation (Duarte, 1972, 2004). Duarte described outcrops in Caieira, municipality of Capanema (30, Figure 4) and an assemblage composed of Nyctaginaceae, Lauraceae, Dilleniaceae, Theaceae, Caryocaraceae, Euphorbiaceae, Rutaceae Chrysobalanaceae, Meliaceae, Sapindaceae, Malvaceae, Melastomataceae, Myrtaceae, Ebenaceae, Rubiaceae and Rhizophoraceae, Rapataceae. This assemblage is suggestive of a neotropical flora.

305
Some leaf morpho-adaptive features, such as the predominance of mesophyle type and acuminate apexes leaves further reinforce the idea of a humid environment in a tropical plain biome.
In Outeiro Island (27, Figure 4) Dutra et al. (2001) describe Bombacaceae plant macrofossils and palynomorphs were reported by Dutra et al. (2001), which are directly related to tropical wet climates, periodic flooding and sandy soils with mangrove growths.
5. THE BRAZILIAN NORTHEAST, EVIDENCE FOR A HUMID PAST
Unambiguous paleoclimatic data on the pre-Quaternary of the Brazilian Northeast are scarce. A Pliocene paleobotanical and palynological assemblage was found in Nova Iorque village (14, Figure 4), in the State of Maranhão – northeast Brazil. This location in no longer accessible and replaced by an artificial lake. The fossils were collected at this site in 1944, and deposited in the Departamento Nacional de Produção Mineral (DNPM). Cristalli (1997), Cristalli & Bernardes de Oliveira (1998) and Anzotegui & Cristalli (2000) identified Lauraceae, Malvaceae, Ebenaceae, Fabaceae, Meliaceae, Smilacaceae families and conclude that an Atlantic forest once occurred in the area. The palynoflora was studied by Lima (1991) and Dino et al. (2006, and in prep.) who confirmed the Pliocene age and the presence of pollen grains representatives of Arecaceae, Juglandaceae, Asteraceae, Poaceae, Onagraceae (Ludwigia; formerly Jussiaea), Euphorbiaceae, Aquifoliaceae, Compositae and Malpighiaceae families. This assemblage indicates tropical climatic conditions at the time of deposition.
In the State of Bahia, Pedrão region (17, Figure 4), Berry (1924, 1935) found Pliocene fossils of the Polygonaceae family. In Ouriçanguinhas and Alagoinhas villages, Hollick & Berry (1924) described macrofossils of Ascomycota, Cyatheaceae, Annonaceae, Lauraceae, Rutaceae, Moraceae, Nyctaginaceae, Chrysobalanaceae, Fabaceae, Meliaceae, Anacardiaceae, Aquifoliaceae, Celastraceae, Sapindaceae, Bombacaceae, Canellaceae, Clusiaceae, Myrtaceae, Melastomataceae, Myrsinaceae, Lecythidaceae, Zygophyllaceae, Styracaceae, Sapotaceae and Apocynaceae families.
At the locality of Maraú (19-20, Figure 4) these authors found fossils of the following families: Heliconiaceae, Polygonaceae, Clusiaceae, Chrysobalanaceae, Fabaceae, Humiriaceae, Rutaceae, Simaroubaceae, Vochysiaceae, Anacardiaceae, Sapindaceae, Myrtaceae and Sapotaceae. This taphoflora illustrates the composition of this Pliocene Atlantic forest (Atlantic ombrophilous dense forest) in a tropical everwet biome. Fossil wood of Combretaceae was found in Aracaju (15, Figure 4) by Mussa (1958) and indicates this family existed in Atlantic forest during the Miocene.
6. PALEOCLIMATE PROXIES
The palynological and paleobotanical data from Neogene deposits in Amazonia and the Brazilian Northeast all point at humid, warm tropical conditions across the region. However, currently there are no quantitative data available that can provide us with more precise estimates of temperature and precipitation rates for this time interval. Fossil plant remains can provide quantitative data on paleoclimate and the available collections should form a good basis for further research. Leaf morphology of plant fossils, for instance, form an excellent indicator of past temperature and precipitation as is shown in Kowalski (2002), but see Cardoso (2007) and http://clamp.ibcas.
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

306 Paleontologia: Cenários de Vida – Paleoclimas
ac.cn/. Both paleobotanical and palynological data can also be used in quantitative analysis such as coexistence approach, a method designed by Mossbrugger & Utescher (1997). This method should be well applicable to South American data, in particular if the paleo-database is extended with diagnostic South American taxa and their temperature and precipitation ranges (www.paleoflora.de).
In addition to palynological and paleobotanical data other proxies, such stable isotope analysis (carbonate and oxygen), can also provide important insights on the Amazonian climate. Kaandorp et al. (2005) used the isotopic composition of Miocene mollusks to reconstruct seasonal rainfall patterns in Western Amazonia. These authors found that seasonal migration of the ITCZ and the intensity of the hydrological cell was comparable to today and that climate conditions were in place to sustain a tropical rainforest comparable.
Molecular phylogenies also form an excellent source of information on plant evolutionary history and can be related to geological history and climate change. A study on montane and lowland taxa from the Andes and Amazonia suggest that high species diversity in western Amazonia is genetically bound to the pre-Quaternary uplift history of the Andes (Antonelli et al., 2009; Hoorn et al., 2010) rather than the sole result of periodic global climatic cooling during the Quaternary (following Haffer, 1969). Another study on molecular phylogenies suggests that central Brazilian forests played a bridging role between Amazonia and the Atlantic forest and that the latter differ substantially in composition from the Amazonian forest (Costa, 2003). Furthermore, molecular fossils – or biomarkers – in organic rich sediments (see Sachs et al., 2007) can also extend our knowledge on past temperature and precipitation regimes in this region.
7. DISCUSSION AND CONCLUSIONS
The palynological and paleobotanical evidence from Neogene deposits of Amazonia and the Brazilian Northeast all do coincide on humid tropical conditions. However, data from the arid regions with Caatinga vegetation are rare and in this respect the locality of Nova Iorque (nr. 14 in Figure 4) is of particular interest. The plant fossils from this location indicate that during the Pliocene an Atlantic forest type existed in an area currently populated by Caatinga. To get a grasp on the origin of this ecosystem we make inferences based on regional and global Cenozoic change.
The present climatic configuration of Amazonia and the Brazilian northeast dates back to the Oligocene and Miocene after continental drift caused a disjunction between Antarctica, Australia and South America leading to the geographical isolation of the Antarctic continent around 40 Ma. A circum-Antarctic Current developed causing also a thermic isolation of Antarctica, which led to the buildup of the Antarctic ice sheet (Bearman & Colling, 2002; but also see Dalziel et al. 2013). Cold waters were transported towards the equator along the western sides of the African continent (Benguela Current) and the South American continent (Humboldt Current). The cold offshore waters caused desert environments in the coastal areas: the Namib Desert and the Atacama Desert, respectively. These deserts are as old as the offshore cold ocean currents itself and this interconnected ocean-terrestrial system is characterized by a long stability, leading in the Namib Desert to geologically conserved endemics such as Welwitschia (Barnard, 1998).

307
On the basis of hundreds of Atlantic Ocean core tops reflecting the period since the last glacial maximum Pflaumann et al. (2003) developed paleogeographical maps of Atlantic Ocean conditions. One important conclusion was that the cold Benguela Current is crossing the South Atlantic Ocean and to touch the Brazilian coast around 10°S subsequently diverging into a northward- and a southward current (Figure 2a). These offshore cold waters caused in coastal eastern Brazil the development of a very dry biome i.e. the Caatinga. However, the coastal escarpment of the Brazilian Highland, created orographic rains overruling the effects of cold offshore waters. Therefore, south of ca. 8°S a narrow strip of wet Atlantic rainforest developed showing inland a steep gradient to the semi-desertic Caatinga vegetation. During the Neogene this paleogeographical and paleoceanographic setting formed a stable system explaining the similar age of Brazilian Caatinga and the Namib desert (see also the synthesis in Gottsberger & Silberbauer-Gottsberger, 2006). Late Pliocene global cooling might have further extended the size of these deserts (see e.g. Hartley and Chong, 2002).
Indeed, investigations in South American biogeography on non-mammal groups suggested that the Caatinga provided xeric refugia during mesic phases of the Pleistocene (Mares et al., 1985). This dynamic history between xeric and mesic environmental conditions at Pleistocene time scales explains why the Caatinga also harbours a mammal fauna that is more characteristic of a mesic tropical biome in terms of species composition as well as overall adaptations (Mares et al., 1985).
8. ACKNOWLEDGEMENTS
We kindly thank Fatima P.R. Leite for the use of selected photographic material that was first published in her PhD thesis in 2006.
CLIM-AMAZON (FP7 project N° 295091) is acknowledged for funding to the first author which made this collaborative work possible. The European Union is not liable for any use that may be made of the information contained therein.
9. REFERENCES
AGUILERA, O. ; GUIMARÃES, J.T.F. & MORAES-SANTOS, H. 2013. Neogene Eastern Amazon carbonate platform and the paleoenvironmental interpretation. Swiss Journal of Paleontology, in press. doi 10.1007/s13358-013-0051-5.
ANTONELLI, A. ; NYLANDER, J.A.A. ; PERSSON, C. & SANMARTIN I. 2009. Tracing the impact of the Andean uplift on Neotropical plant evolution. PNAS, 106, 9749-9754.
ANZÓTEGUI, L. M. & CRISTALLI, P. 2000. Primer Registro de Hojas de Malvaceae en el Neógeno de Argentina y Brasil. Ameghiniana, 37(2): 169-180.
ARAI, M.; TRUCKENBRODT, W.; NOGUEIRA, A.C.R.; GOES, A.M., & ROSSETTI, D.F. 1994. Novos dados sobre estratigrafia e ambiente deposicional dos sedimentos Barreiras, NE do Pará. In: SIMPOSIO DE GEOLOGIA DA AMAZONIA, IV, Belém, SBG, Boletim de Resumos., p. 185-187.
ARCHIBOLD, O.W. 1995. Ecology of world vegetation. Chapman and Hall, London, UK, 510 p.
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

308 Paleontologia: Cenários de Vida – Paleoclimas
BEARMAN, G. & COLLING, A. (eds.). 2002. Ocean circulation. The Open University, Milton Keynes & Butterworth-Heineman, 286 p.
BARNARD, P. (ed.). 1998. Biological diversity in Namibia, a country study. Namibian National Biodiversity Task Force, Windhoek, Namibia, 325 p.
BARNES, J. B.; EHLERS, T. A.; INSEL, N.; MCQUARRIE, N. & POULSEN, C. J. 2012. Linking orography, climate, and exhumation across the central Andes. Geology, 40: 1135-1138.
BEHLING, H.; ARZ, H.W.; PÄTZOLD, J. & WEFER, G. 2000. Late Quaternary vegetational and climate dynamics in northeastern Brazil, inferences from marine core GeoB 3104-1. Quaternary Science Reviews, 19(10): 981–994.
BEHLING, H. & COSTA, M.L. 2004. Mineralogy, geochemistry and palynology of late Tertiary mangrove deposits in the Barreiras Formation of Mosqueiro Island in northeastern Pará State, eastern Amazonia. Journal of South American Earth Sciences, 17: 285-295.
BERRY, E.W. 1924. Late Tertiary Flora from Bahia, Brazil. The Johns Hopkins University Studies in Geology, 5: 11-137.
BERRY, E.W. 1935. Tertiary Plants from Brazil. Proceedings American Philosophical Society, 75: 565-590.
BERRY, E.W. 1937. Late Tertiary Flora from the Territory of Acre, Brazil. The Johns Hopkins University Studies in Geology, 12: 81-90.
CARDOSO, N. 2007. Paleoecologia da flora de Catalão, paleolago cemitério, estado de Goiás. IG-UFRGS, Tese de Doutorado, 98 p.
CARVALHO, M.A. 1996. Estudo paleoecológico e paleoclimático, com base em palinologia, aplicado em sedimentos Pleistocênicos e Pliocênicos da Bacia da Foz do Amazonas. IG-UFRJ, Dissertação de Mestrado, Rio de Janeiro, 146 p.
COSTA, L. 2003. The historical bridge between the Amazon and the Atlantic Forest of Brazil: a study of molecular phytogeography with small mammals. Journal of Biogeography, 30(1): 71–86.
CRISTALLI, P. 1997. Tafoflora das Camadas Nova Iorque, Depósitos Neógenos do Rio Parnaíba, Município de Nova Iorque (MA), Brasil. IGc.USP. Dissertação de Mestrado, 157 p.
CRISTALLI, P. & BERNARDES-DE-OLIVEIRA, M.E.C. 1998. Tafoflora Neógena das Camadas Nova Iorque (MA-Brasil) e seu significado paleofitogeográfico. Acta Geológica Leopoldensia, 46-47: 55-67.
DALZIEL, I.W.D.; LAWVER, L.A.; PEARCE, J.A.; BARKER, P.F.; HASTIE, A.R.; BARFOD, D.N.; SCHENKE, H-F. & DAVIS, M.B. 2013. A potential barrier to deep Antarctic circumpolar flow until the late Miocene? Geology, doi: 10.1130/G34352.1
DEL’ARCO, J.O.; SANTOS, R.O.B.; RIVETTI, M.; ALVES, E.D.O.; FERNANDES, C. A.C. & SILVA, L.L. 1977. Geologia. In:. Projeto RADAMBRASIL. Folha SB.19-Juruá. DNPM, Rio de Janeiro, Levantamento de Recursos Naturais, 15:17-88

309
DINO, R.; GARCIA, M.J.; ANTONIOLI, L. & LIMA, M.R. 2006. Palinoflora das “Camadas Nova Iorque”, Registro Sedimentar do Plioceno na Bacia do Parnaíba (Maranhão). In: SIMPÓSIO DO CRETÁCEO DO BRASIL, 7 e SIMPÓSIO DO TERCIÁRIO DO BRASIL, 1, Serra Negra, Boletim de Resumos, p.42.
DINO, R.; GARCIA, M.J.; ANTONIOLI, L.; LIMA, M.R. & CASSAB, R. in prep.. Palinoflora das Camadas Nova Iorque, registro sedimentar do Plioceno na Bacia do Parnaíba (Maranhão).
DINO, R.; SOARES, E.A.A.; ANTONIOLI, L., RICCOMINI, C. & NOGUEIRA, A.C.R., 2012. Palynostratigraphy and sedimentary facies of Middle Miocene fluvial deposits of the Amazonas Basin, Brazil. Journal of South American Earth Sciences, 34: 61-80.
DINO, R.; SOARES, E.A.A. & ANTONIOLI, L. in prep. A Palynological and Sedimentological study on the Miocene outcrops (Uatumã and Jatapu Rivers) of Amazonas Basin – North
DOBSON, D.M.; DICKENS, G.R. & REA, D.K. 2001. Terrigenous sedimenton Ceara Rise: a Cenozoic record of South American orogeny and erosion. Paleogeography, Palaeoclimatology, Palaeoecology, 165: 215–229.
DOLIANITI, E. 1948. A Paleobotânica no Brasil. Rio de Janeiro, DNPM, Divisão de Geologia e Mineralogia, 123: 1-87.
DUARTE, L. 1970. Comentários sobre o “Status” Taxinômico de uma Coleção de Vegetais Fósseis do Acre. Anais da Academia Brasileira de Ciências, 42(3): 471-476.
DUARTE, L. 1972. Flórula da Formação Pirabas, Estado do Pará, Brasil. IGc.-USP, Tese de Doutorado, 156 p.
DUARTE, L. 2004. Paleoflórula. Cap. 6. In: ROSSETTI, D.F. & GÓES, A.M. (eds.). O Neógeno da Amazônia Oriental. Museu Emílio Goeldi. Coleção Friedrich Katzer. p. 169-198.
DUARTE, L. & JAPIASSÚ, A. M. S. 1971. Vegetais Meso e Cenozoicos do Brasil. Anais da Academia Brasileira de Ciências, 43(supl.): 433-443.
DUTRA, T. L.; ROSSETTI, D. F. & STRANZ, A. 2001. Bombacaceae Kuhnt., 1821 from Middle Miocene, Barreiras Formation (Depositional Sequence) in Pará State, Brazil. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 17, Rio Branco, AC, Boletim de Resumos, p.77.
FIGUEIREDO, J.P.; HOORN, C.; VAN DER VEN, P. & SOARES, E. 2009. Late Miocene onset of the Amazon River and the Amazon deep-sea fan: Evidence from the Foz do Amazonas Basin. Geology, 37(7): 619-622.
GARREAUD, R.D. & ACEITUNO, P. 2007. Atmospheric circulation and climate variability. In: VEBLEN, T.T.; YOUNG, K.R. & ORME, A.R. (eds.). 2007. The physical geography of South America. Oxford University Press, Oxford, UK, p. 45-59.
GARREAUD, R.D.; VUILLE, M.; COMPAGNUCCI, R. & MARENGO, J. 2009. Present-day South American climate. Palaeogeography, Palaeoclimatology, Palaeoecology, 281: 180–195.
GARZIONE, C.N.; HOKE, G.D.; LIBARKIN, J.C.; WITHERS, S.; MACFADDEN, B.; EILER, J.; GHOSH, P. & MULCH, A. 2008. Rise of the Andes. Science, 320(5881): 1304-1307.
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

310 Paleontologia: Cenários de Vida – Paleoclimas
GENTRY, A.H. 1982. Neotropical floristic diversity: phytogeographical connections between Central and South America, Pleistocene climatic fluctuations, or an accident of the Andean orogeny? Annals Missouri Botanical Garden, 69: 557–593.
GOTTSBERGER, G. & SILBERBAUER-GOTTSBERGER, I. 2006. Life in the Cerrado: a South American tropical seasonal ecosystem. Abteilung Systematische Botanik und Okologie und Botanischer Garten, Reta Verlag, Universität Ulm, Germany, 1: 277 p.
GOULDING, M.; BARTHEM, R. & FERREIRA, E. 2003. The Smithsonian atlas of the Amazon. Smithsonian Institution, Washington, USA, 255 p.
GROSS, M.; PILLER, W.E.; RAMOS, M.I. & PAZ, J.D.S. 2011. Late Miocene sedimentary environments in South-Western Amazonia (Solimões Formation; Brazil). Journal of South American Earth Sciences, 32: 169-181.
GUIMARÃES, D. 1964. Geologia do Brasil. Mem. Div. Fom. Prod. Mineral, DNPM, 1: 1-674.
HAFFER, J., 1969. Speciation in Amazonian Forest Birds. Science, 165: 131-137.
HARRIS, S.E. & A.C. MIX, 2002. Climate and tectonics drive continental erosion of tropical South America, 0-13 Ma. Geology, 30: 447-450.
HARTLEY, A.J. & CHONG, G. 2002. Late Pliocene age for the Atacama Desert: Implications for the desertification of western South America. Geology 30: 43-46.
HOLLICK, A. & BERRY, E. W. 1924. A Late Tertiary Flora from Bahia, Brazil. The Johns Hopkins University Studies in Geology, 5: 11-136.
HOORN, C. 1993. Marine incursions and the influence of Andean tectonics on the Miocene depositional history of northwestern Amazonia: Results of a palynostratigraphic study. Palaeogeography, Palaeoclimatology, Palaeoecology, 105: 267-309.
HOORN, C.; WESSELINGH, F.P.; TER STEEGE, H.; BERMUDEZ, M.A.; MORA, A.; SEVINK, J.; SANMARTÍN, I.; SANCHEZ-MESEGUER, A.; ANDERSON, C.L.; FIGUEIREDO, J.P.; JARAMILLO, C.; RIFF, D.; NEGRI, F.R.; HOOGHIEMSTRA, H.; LUNDBERG, J.; STADLER, T.; SARKINEN, T. & ANTONELLI, A. 2010. Amazonia through time: The effect of Andean uplift and climate change on landscape evolution and biodiversity. Science, 330: 927-931.
HORBE, A.M.C.; HORBE, M.A. & SUGUIO, K. 2004. Tropical Spodosols in northeastern Amazonas State, Brazil. Geoderma, 119: 55–68.
HORBE, A.M.C.; PAIVA, M.R.P.; MOTTA, M.B. & HORBE, M.A. 2007. Mineralogia e geoquímica dos perfis sobre sedimentos Neógenos e Quaternários da Bacia do Solimões na região de Coari – AM. Acta Amazonica, 37(1): 81-90.
INSEL, N.; POULSEN, C.J. & EHLERS, T.A. 2010. Influence of the Andes mountains on South American moisture transport, convection, and precipitation. Climate Dynamics, 35: 1477-1492.
JAPIASSU, A.M.S. 1969. Madeira Fóssil Humiriaceae de Iritúia. Estado do Pará. Boletim Emílio Goeldi-Geologia, 14: 1-12.
JARAMILLO, C.; RUEDA, M.J. & MORA, G. 2006. Cenozoic Plant Diversity in the Neotropics. Science, 311: 1893-1896.

311
JARAMILLO, C. & CARDENAS, A. 2013. Global warming and neotropical rainforests: a historical perspective. Annual Review of Earth and Planetary Sciences. In press.
JARAMILLO, C.; HOORN, C.; SILVA, S.A.F.; LEITE, F.; HERRERA, F.; QUIROZ, L.; DINO, R. & ANTONIOLI, L. 2010. The origin of the modern Amazon rainforest: implications of the palynological and paleobotanical record. In: Amazonia, landscape and species evolution, HOORN, C. & WESSELINGH, F.P. (eds.). Wiley-Blackwell, p. 317-334.
JEPSEN, P.; BONOW, J.M.; GREEN, P.F.; COBBOLD, P.R.; CHIOSSI, D.; LILLETVEIT, R.; MAGNAVITA, L.P. & PEDREIRA, A. 2012. Episodic burial and exhumation in NE Brazil after opening of the South Atlantic. GSA Bulletin, 124(5/6): 800–816.
KAANDORP, R.J.G.; VONHOF, H.B.; WESSELINGH, F.P.; ROMERO-PITTMAN, L.; KROON, D. & VAN HINTE, J.E. 2005. Seasonal Amazonian rainfall variation in the Miocene Climate Optimum. Palaeogeography, Palaeoclimatology, Palaeoecology, 221: 1-6.
KOWALSKI, E.A. 2002. Mean annual temperature estimation based on leaf morphology: a test from tropical South America. Palaeogeography, Palaeoclimatology, Palaeoecology, 188: 141–165.
LATRUBESSE, E.M.; SILVA, S.A.F.; COZZUOL, M. & ABSY, M.L. 2007. Late Miocene continental sedimentation in Southwestern Amazônia and its regional significance: biotic and geological evidence. Journal of South American Earth Sciences, 23: 61-80.
LEANDRO, L.M. 2012. Análises palinoestratigráficas da Formação Solimões na porção noroeste do Alto Solimões, Amazonas, Brasil. Universidade do Vale do Rio dos Sinos, Dissertação de Mestrado,109 p.
LEGUIZAMON VEJA, A.M.; NOGUEIRA, A.C.R.; MAPES, R.W. & COLEMAN, D.S. 2006. A late Miocene delta-lacustrine system in the eastern Solimões Basin: Prelude to the modern Amazon River. In: ANNUAL MEETING, Philadelphia, Geological Society of America, Abstracts, 38(7): 144.
LEITE, F.P.R. 1997. Palinofloras Neógenas da Formação Pirabas e Grupo Barreiras, Área Litorânea Nordeste do Estado do Pará, Brasil. Instituto de Geociências, Universidade de São Paulo. Dissertação de Mestrado. 102 p.
LEITE, F.P.R. 2004. Palinologia- Cap. 2. In: ROSSETTI, D. F. & GÓES, A. M. (eds). Neógeno da Amazônia Oriental, Museu Goeldi, Estado do Para, Brazil, p. 55-90.
LEITE, F.P.R. 2006. Palinologia da Formação Solimões, Neógeno da Bacia do Solimões, Estado do Amazonas: Implicações paleoambientais e bioestratigráficas. Universidade de Brasília, Tese de Doutorado, 138 p.
LEITE, F.P.R.; BERNARDES-DE-OLIVEIRA, M.E.C.; DE OLIVEIRA, P.E.; SILVESTRE-CAPELATO, M.S.; ARAI, M. & TRUCKENBRODT, W. 1997 a. Palinofloras miocenas da Formação Pirabas e Grupo Barreiras na região Bragantina, Estado do Pará Brasil. Revista Universidade Guarulhos, Geociências, 2(esp.): 128-140.
LEITE, F.P.R.; BERNARDES-DE-OLIVEIRA, M.E.C.; ARAI, M. & TRUCKENBRODT, W. 1997 b. Palinoestratigrafia da Formação Pirabas e Grupo Barreiras, Mioceno do Nordeste do Estado do Pará, Brasil. Revista Universidade Guarulhos, Geociências, 2(esp.): 141-147.
LIMA, M.R. 1991. Estudo Palinológico das “Camadas Nova Iorque” Terciário do Estado do Maranhão. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 12, São Paulo, Boletim de Resumos..., p.45.
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

312 Paleontologia: Cenários de Vida – Paleoclimas
MAIA, R.; GODOY, H.; YAMAGUTI, H.; DE MOURA, P.; COSTA, F.; DE HOLANDA, M. & COSTA, J. 1977. Projeto de Carvão no Alto Solimões. CPRM-DNPM, 137 p.
MACHADO, L. G.; SCHEEL-YBERT, R.; BOLZON, R. T.; CARVALHO, M. A. & CARVALHO, I. S. 2012. Lenhos fósseis do Neógeno da Bacia do Acre, Formação Solimões: contexto paleoambiental. Revista Brasileira de Geociências, 42(1): 67-80.
MARENGO, J.A.; SOARES, W.R.; SAULO, C. & NICOLINI, M. 2004. Climatology of the Low-Level Jet East of the Andes as Derived from the NCEP–NCAR Reanalyses: Characteristics and Temporal Variability. Journal of Climate, 17(12): 2261-2280.
MARES, M.A.; WILLIG, M.R. & LACHER JR., T.E. 1985. The Brazilian Caatinga in South American zoogeography: tropical mammals in a dry region. Journal of Biogeography, 12: 57-69.
MAURY, C.J. 1937. Argilas Fossilíferas do Território do Acre. Boletim do Serviço Geológico e Mineralógico, 77: 1-29.
MORAIS-NETO, J.M.; HEGART, K.A.; KARNER, G.D. & ALKMIM, F.F. 2009. Timing and mechanisms for the generation and modification of the anomalous topography of the Borborema Province, northeastern Brazil. Marine and Petroleum Geology, 26: 1070-1086.
MOSBRUGGER, V. & UTESCHER, T. 1997. The coexistence approach - a method for quantitative reconstructions of Tertiary terrestrial paleoclimate data using plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 134: 61 86.
MUSSA, D. 1958. Dicotiledônio Fóssil da Formação Barreiras, Estado de Sergipe. Boletim do DNPM, 181: 1-23.
MUSSA, D. 1959. Contribuição à Paleoanatomia Vegetal II. Madeira Fósseis do Territorio do Acre (Alto Jurua) Brasil. Boletim do DNPM, 195: 1-54.
MUSSA D.; BERNARDES-DE-OLIVEIRA, M.E.; MUNE, S.E.; DE SIQUEIRA, E.; CASTRO-FERNANDES, M.C. de & CARMO D.A. 2002. Lenho de Dicotiledônea da Formação Solimões, Neógeno da Bacia do Acre, Brasil. Paleontologia em Destaque, 40: 16-17.
OCHOA, D.; HOORN, C.; JARAMILLO, C.; BAYONA, G.; PARRA, M. & DE LA PARRA, F. 2012. The final phase of tropical lowland conditions in the axial zone of the Eastern Cordillera of Colombia: Evidence from three palynological records. Journal of South American Earth Sciences, 39: 157-169.
OLIVEIRA, A.I. & LEONARDOS, O.H. 1943. Geologia do Brasil. 2ª ed. Rio de Janeiro. 782p.
OLIVEIRA, E.P. 1937. Estado actual da Paleontologia Brasileira. Mineração e Metalurgia, 7: 11-12.
PARRA, M.; MORA, A.; JARAMILLO, C.; STRECKER, M.R.; SOBEL, E.R.; QUIROZ, L.; RUEDA, M. & TORRES, V. 2009. Orogenic wedge advance in the northern Andes: Evidence from the Oligocene-Miocene sedimentary record of the Medina Basin, Eastern Cordillera, Colombia GSA Bulletin, 121(5/6): 780-800.
PAZ, J.D.S.; RAMOS, M.I.F.; MORAES-SANTOS, H.M. & SILVA, S., in press. Depósitos fluviais da Formação Solimões, Bacia do Solimões, Eirunepé (AM), Brasil. Latin Am Eirunepé rican. Journal of Sedimentology and Basin Analysis.

313
PESTANA, F. (Coord.) 1994. Atlas Geográfico Mundial. São Paulo: Folha. 184p.
PFLAUMANN, U.; SARNTHEIN, M.; CHAPMAN, M.; D’ABREU, L.; FUNNELL, B.; HUELS, M.; KIEFER, T.; MASLIN, M.; SCHULZ, H.; SWALLOW, J.; VAN KREVELD, S.; VAUTRAVERS, M.; VOGELSANG, E. & WEINELT M. 2003. Glacial North Atlantic: sea-surface conditions reconstructed by GLAMAP 2000. Paleoceanography, 18(3), 1065, doi:10.1029/2002PA000774.
POULSEN, C.J.; EHLERS, T.A. & INSEL, I. 2010. Onset of Convective Rainfall During Gradual Late Miocene Rise of the Central Andes. Science, 328: 490-493.
RICHARDS, K. & LOWE, S. 2002. Plio-Pleistocene palynostratigraphy of the Amazon Fan, offshore Brazil: Insights into vegetation history, palaeo-climate & sequence stratigraphy. Does fluvial discharge style influence petroleum reservoir distribution? Poster. In: THE MICROPALAEONTOLOGICAL SOCIETY (TMS) AND AMERICAN ASSOCIATION OF STRATIGRAPHIC PALYNOLOGISTS (AASP), London (UK) September 2002.
ROSSETTI, D.F. & GOES, A.M. 2004. O Neógeno da Amazônia Oriental. Museu Goeldi, Estado do Pará, 222p.
ROSSETTI, D.F.; BEZERRA, F.H.R. & LANDIM DOMINGUEZ, J.M. 2013. Late Oligocene-Miocene transgressions along the equatorial and eastern margins of Brazil. Earth Science Reviews (in press) doi: 10.1016/j.earscirev.2013.04.005
SACHS, J.P., PAHNKE, K., SMITTENBERG, R., ZHANG, Z. 2007. Biomarker Indicators of Past Climate, in Encyclopedia of Quaternary Science (ed. Elias, S.), Elsevier, Amsterdam.
SEPULCHRE, P.; SLOAN, L.C. & FLUTEAU, F. 2010. Modelling the response of Amazonian climate to the uplift of the Andean mountain range. In: HOORN, C. & WESSELINGH, F.P. (eds.). Amazonia, landscape and species evolution. Wiley-Blackwell, p. 211-222.
SHEPHARD, G. E.; MULLER, R. D.; LIU, L. & GURNIS, M. 2010. Miocene drainage reversal of the Amazon River driven by plate–mantle interaction. Nature Geoscience, 3(12): 870-875.
SILVA, S.A.F. 2004. Palinologia do Neógeno da Amazônia Sul Ocidental, Brasil. Universidade Federal do Amazonas. UFAM, Dissertação de Mestrado, 105 p.
SILVA, S.A.F. 2008. Palinologia do Neógeno da Bacia do Alto Solimões, Amazônia Ocidental, Brasil: Aspectos Sistemáticos, Bioestratigráficos e Paleoecológicos. Universidade Federal do Amazonas, UFAM, Tese de Doutorado, 190 p.
SILVA-CAMINHA, S.A.F.; JARAMILLO, C.A. & ABSY, M.L. 2010. Neogene palynology of the Solimões Basin, Brazilian Amazonia. Paleontographica Abt B, 283: 1-67.
SILVEIRA, R.R. & NOGUEIRA, A.C.R. 2007. Cronoestratigrafia de Depósitos Miocenos da Bacia do Solimões e sua correlação com os depositos da Amazônia. In: CONGRESSO DA ABEQUA, 2007, Belém. Boletim de Resumos, CDRom.
TRUCKENBRODT, W.; ARAI, M.; NOGUEIRA, A.C.R. & GOES, A.M. 1994. Conteúdo fossilífero e estruturas sedimentares nos sedimentos Barreiras, NE do Pará. In: CONGRESSO BRASILEIRO DE GEOLOGIA, 38, Balneário Camboriu, SC. Boletim de Resumos Expandidos,3: 210-211.
Carina Hoorn, Mary E.C. Bernardes-de-Oliveira, Rodolfo Dino, Maria Judite Garcia, Luzia Antonioli, Fabio da Costa Casado & Henry Hooghiemstra

314 Paleontologia: Cenários de Vida – Paleoclimas
VEBLEN, T.T.; YOUNG, K.R. & ORME, A.R. (eds.). 2007. The physical geography of South America. Oxford University Press, Oxford, UK, 361 p.
VERA, C.; HIGGINS, W.; AMADOR, J.; AMBRIZZI, T.; GARREAUD, R.; GOCHIS, D.; GUTZLER, D.; LETTENMAIER, D.; MARENGO, J.; MECHOSO, C.R.; NOGUES-PAEGLE, J.; SILVA DIAS, P. L. & ZHANGl, C. 2006. Toward a Unified View of the American Monsoon Systems. Journal of Climate, 19: 4977-5000.
VONHOF, H.B. & KAANDORP, R.J.G. 2010. Climate variation in Amazonia during the Neogene and the Quaternary. In: HOORN, C. & WESSELINGH, F.P. (eds.). Amazonia, landscape and species evolution. Wiley-Blackwell, p. 201-210.
WANDERLEY FILHO, J.R.; EIRAS J.F.; DA CRUZ CUNHA P.R. & VAN DER VEN P.H., 2010. The Paleozoic Solimões and Amazonas basins and the Acre foreland basin of Brazil. In: HOORN, C. & WESSELINGH, F.P. (eds.). Amazonia, landscape and species evolution. Wiley-Blackwell, p. 29-36.
YOUNG, K.R.; BERRY, P.E. & VEBLEN, T.T. 2007. Flora and vegetation. In: VEBLEN, T.T.; YOUNG, K.R. & ORME, A.R. (eds.). The physical geography of South America. Oxford University Press, Oxford, UK, p. 91-100.
ZACHOS, J.C.; DICKENS, G.R. & ZEEBE, R.E. 2008. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature, 451: 279-83.