validatie van de verschillende muismodellen voor auto...
TRANSCRIPT

Validatie van de verschillendemuismodellen voor
auto-immune hepatitis
Laura Vanden Daele
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Isabelle Colle
Vakgroep Inwendige Ziekten
Academiejaar 2012-2013


Validatie van de verschillendemuismodellen voor
auto-immune hepatitis
Laura Vanden Daele
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Isabelle Colle
Vakgroep Inwendige Ziekten
Academiejaar 2012-2013

“De auteur en de promotor geven de toelating deze masterproef voor consultatie
beschikbaar te stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk
ander gebruik valt onder de beperkingen van het auteursrecht, in het bijzonder
met betrekking tot de verplichting uitdrukkelijk de bron te vermelden bij het
aanhalen van resultaten uit deze masterproef.”
Datum
Laura Vanden Daele Isabelle Colle

Voorwoord
Eindelijk, mijn masterproef is af! De weg naar de eindmeet was niet altijd even eenvoudig,
maar uiteindelijk ben ik er geraakt, maar niet zonder de hulp van anderen.
Eerst en vooral wil ik mijn promotor Isabelle Colle bedanken voor de hulp bij mijn
masterproef, vooral voor het vertrouwen en de steun.
Ik zou ook graag mijn begeleider Muhammed Yüksel bedanken voor de inhoudelijke hulp,
ook al was dit niet zo eenvoudig nadat hij naar Yale vertrok.
Verder ben ik Eliene Bogaerts en Stephanie Coulon ongelofelijk dankbaar! Toen Mo naar
Yale vertrok, zorgden zij voor de beste vervanging die ik kon wensen. Eliene stond altijd
klaar, zowel om mee te gaan naar de VUB, om te helpen bij een experiment, of om mijn
teksten na te lezen. Ook Stephanie was altijd paraat; ik ben haar veel dank verschuldigd voor
de hulp bij de analyse van mijn resultaten en het vele keren nalezen van mijn masterproef.
Beiden deden dit met een ongelofelijk enthousiasme en slaagden erin mij weer moed te geven
wanneer mijn masterproef niet zo goed vlotte.
Daarnaast zou ik ook nog graag Sandra Van Lint van de VUB bedanken voor de hulp bij onze
experimenten en het vrijmaken van haar tijd hiervoor, en Martijn de Jong voor het aanleren
van verschillende labotechnieken.
Ik wil ook nog mijn medemasterproefstudent Sander Lefere bedanken voor de hulp en de
steun, zo was ik toch niet de enige masterproefstudent met een begeleider in Amerika en 11
muizen in Brussel.
Tenslotte wil ik nog mijn broer, mijn ouders en mijn vrienden bedanken voor hun steun, niet
enkel tijdens deze masterproef, maar ook tijdens de afgelopen 5 jaar. Dank u, om meer in mij
te geloven dan ik zelf deed, en mij te helpen moedig te zijn tijdens moeilijke momenten.

Inhoudstafel
Samenvatting.............................................................................................................................. 1
1. Inleiding .............................................................................................................................. 2
1.1. De lever ...................................................................................................................... 2
1.1.1. Anatomie ............................................................................................................ 2
1.1.2. Fysiologie ........................................................................................................... 4
1.2. Autoimmune hepatitis ................................................................................................ 5
1.2.1. Definitie.............................................................................................................. 5
1.2.2. Epidemiologie .................................................................................................... 5
1.2.3. Pathogenese ........................................................................................................ 5
1.2.3.1. Genetica.......................................................................................................... 5
1.2.3.2. Immunopathogenese....................................................................................... 6
1.2.4. Klinische presentatie en diagnose ...................................................................... 8
1.2.5. Behandeling...................................................................................................... 11
1.2.6. Leverfibrose en hypoxie................................................................................... 12
1.2.7. Muismodellen voor AIH .................................................................................. 13
1.3. Probleemstelling....................................................................................................... 14
2. Materialen en methoden.................................................................................................... 15
2.1. Keuze van het muismodel ........................................................................................ 15
2.2. Inductie van auto-reactieve T-cellen ........................................................................ 15
2.2.1. Immunisatie met CYP2D6 ............................................................................... 15
2.2.2. In vivo cytotoxic T lymphocyte assay (CTL) .................................................. 16
2.3. Validatie van het muismodel na 3 maanden............................................................. 20
2.3.1. Extra immunisatie met CYP2D6...................................................................... 20
2.3.2. Bepaling antilichamen en leverwaarden .......................................................... 20
2.3.3. Sacrificatie........................................................................................................ 20
2.3.4. Paraffinecoupes ................................................................................................ 21

2.4. Histologie ................................................................................................................. 21
2.4.1. Haematoxyline en eosine (H&E) kleuring op de lever .................................... 21
2.4.2. Sirius Red kleuring op de lever ........................................................................ 22
2.5. Immunohistochemische kleuringen.......................................................................... 22
2.5.1. HIF-1α kleuring op de lever ............................................................................. 22
2.5.2. Glut1 kleuring op de lever................................................................................ 23
2.5.3. F4/80 kleuring op de lever ............................................................................... 24
2.6. Histopathologie ........................................................................................................ 25
2.7. Kwantitatieve Real Time PCR ................................................................................. 25
2.8. Statistiek ................................................................................................................... 26
3. Resultaten.......................................................................................................................... 27
3.1. Inductie van auto-reactieve T-cellen ........................................................................ 27
3.2. Validatie van het muismodel na 3 maanden............................................................. 27
3.2.1. Algemene karakteristieken ............................................................................... 27
3.2.2. Lever histologie................................................................................................ 28
3.2.3. Analyse van inflammatoire merkers ................................................................ 29
3.2.4. Analyse van hypoxische merkers ..................................................................... 32
3.2.4.1. Analyse van Hifs en Pdhs............................................................................. 32
3.2.4.2. Analyse van Glut1, Pfk en Vegfa................................................................. 34
4. Discussie ........................................................................................................................... 37
5. Algemeen besluit .............................................................................................................. 44
6. Referenties ........................................................................................................................ 46

1
Samenvatting
Inleiding: Auto-immune hepatitis (AIH) is een zeldzame inflammatoire leverziekte, die
progressief en chronisch is. De behandeling bestaat uit het toedienen van immunosuppressiva,
maar is vaak niet succesvol. Het ontwikkelen van een muismodel dat representatief is voor
AIH bij de mens zou kunnen helpen om de pathofysiologie te ontrafelen en zo een goede
behandeling te ontwikkelen. In deze masterproef werd geprobeerd een muismodel voor AIH
te ontwikkelen en dit te valideren.
Materialen en methoden: vijf C57BL/6 muizen werden in de inguinale lymfeknoop
geïnjecteerd met mRNA van CYP2D6. De ontwikkeling van AIH werd vergeleken met zes
controle muizen. Dit werd geëvalueerd door een cytotoxic T lymphocyte assay (CTL) om te
kijken of er auto-reactieve T-cellen tegen CYP2D6 aanwezig waren. Verder werden ALT,
AST en auto-antilichamen in het serum onderzocht. Na 3 maanden werden de muizen
gesacrificeerd en werden de levers onderzocht op inflammatie aan de hand van
haematoxyline-eosine kleuringen en op fibrose aan de hand van Sirius Red kleuringen.
Daarnaast werd inflammatie in de lever ook onderzocht aan de hand van de Tnf-mRNA
expressie door een qPCR en een F4/80 kleuring. De hypoxie in de lever werd ook
geanalyseerd. Dit gebeurde door de qPCR-analyse van de mRNA-expressie van Hif1a, Hif2a,
Phd1, Phd2, Phd3, Glut1, Pfk en Vegfa en de mRNA-expressie van Hif1a en de
eiwitexpressie van Glut1 op immunohistochemische kleuringen.
Resultaten: De CTL leverde geen interpreteerbare resultaten op en na 3 maanden kon er geen
AIH vastgesteld worden bij de behandelde muizen. Er werden geen auto-antilichamen
gedetecteerd en de waarden van ALT en AST waren niet gestegen ten opzichte van de
controle groep. Op histologisch niveau werden er geen pathologische veranderingen
waargenomen. De analysen wezen op minder inflammatie en hypoxie in de CYP2D6 muizen
dan bij de controle muizen. Er werd wel haarverlies op de rug gezien bij CYP2D6 muizen en
niet bij de controle muizen.
Discussie: Het herhalen van het experiment met een goed werkende CTL zou kunnen
aantonen of deze manier van immunisatie een goede manier is om autoreactieve T-cellen te
induceren. Indien dit zo zou zijn, zou de deletie van regulatoire T-cellen een eventueel punt
van verbetering kunnen zijn. De hogere inflammatie en hypoxie in de controle muizen zou
kunnen te wijten zijn aan statistische fouten of aan een Mouse hepatitis virus (MHV) infectie.
Het haarverlies op de rug heeft geen aangetoond verband met AIH en zou misschien aan
barbering kunnen te wijten zijn, maar meer waarschijnlijk ligt dit aan een nog onbekende
oorzaak.

2
Inleiding
1.1. De lever
1.1.1. Anatomie
De lever is de grootste klier in het lichaam. Het gewicht varieert tussen 1200 tot 1700 gram
bij een volwassene. Ze bevindt zich in de abdominale holte, bovenaan rechts onder het
diafragma. Een kwart van de lever bevindt zich links van de mediaanlijn en de rest bevindt
zich rechts ervan. De vorm van de lever is wigvormig en heeft twee oppervlakken, de convexe
facies diaphragmatica en de vlakke facies visceralis. De lever is bedekt door het visceraal
peritoneum, behalve ter hoogte van de area nuda. Hier slaat het visceraal peritoneum om in
het pariëtaal peritoneum dat zich tegen het diafragma bevindt.
Uitwendig is de lever verdeeld in de linker- en rechterlob, door het ligamentum falciforme aan
het voorste deel van het facies diaphragmatica, en door de fissuur, waarin het ligamentum
venosum ligt, aan het facies visceralis. Het ligamentum falciforme is een dubbelgevouwen
peritoneum. Onderaan is dit ligament vrij en verbreedt het, dit wordt het ligamentum teres
hepatis genoemd.
Langs de viserale zijde zijn er nog twee andere lobben, rechts van de fissuur. Bovenaan
bevindt zich de lobus caudatus en onderaan de lobus quadratus. Deze twee lobben zijn
verbonden via de leverpoort. Langs hier bereiken de autonome zenuwen, die van de plexus
coeliacus komen, de lever. Hier komen ook de arteria hepatis, die de lever van zuurstofrijk
bloed voorziet, en de vena portae hepatis, met nutriëntenrijk bloed dat afkomstig is van de
maag, darmen en milt, binnen. De ductus hepaticus communis treedt hier naar buiten en voert
de gal naar de galblaas die zich rechts onderaan, naast de lobus quadratus, bevindt. Tenslotte
rechts bovenaan, naast de lobus caudatus, ligt de vena hepatica inferior die het bloed terug
stuurt naar het lichaam (zie figuur 1) (1-3).

3
Figuur 1: vooraanzicht (boven) en achteraankant (onder) van de lever (http://www.edoctoronline.com/medical-atlas.asp?c=4&id=20713)
Op microscopisch niveau bestaat de lever uit meer dan een miljoen leverlobuli, met een
diameter van ongeveer 0,5 – 2mm. Deze zijn hexagonaal en bestaan allemaal uit hepatocyten.
Tussen platen van hepatocyten lopen leversinusoïden waarin bloed stroomt. In het midden van
een leverlobulus bevindt zich een centrale vene. In de hoeken van de leverlobuli ligt de
portale triade. Dit is een bindweefselkapsel met daarin een takje van de arteria hepatica, de
venae portae en een galkanaaltje. Er lopen ook lymfevaten in deze portale triade. De gal die
door de hepatocyten wordt gemaakt gaat centrifugaal, via canaliculi, naar het kanaaltje van
Hering, dat uitmondt in het galkanaal in de portale triade (zie figuur 2). Het bloed stroomt
centripetaal vanuit de arteria hepatica en venae portae in de portale triade, en verder via de
leversinusoïden naar de centrale vene. De centrale venen monden uiteindelijk uit in de vena
cava inferior.
Figuur 2: leverlobulus (links) met V= centrale vene, T = portale triade en C = bindweefsel en portale triade(rechts) met PV= vena porta, A= arteria hepatica, B = galkanaaltje, L = lymfevat en S= leversinusoïden (4)

4
De leversinusoïden zijn bloedvaten die omringd worden door endotheelcellen met een
venster. De endotheelcellen zelf sluiten niet aan op elkaar en er is geen basale membraan
aanwezig. Tussen de leversinusoïden en de hepatocyten bevindt zich de ruimte van Disse.
Hierdoor komen de hepatocyten gemakkelijk in contact met het bloed, wat belangrijk is voor
de functie van de hepatocyten. In de leversinusoïden, vast aan de endotheelcellen, bevinden
zich nog cellen van een ander type, namelijk Kupffer cellen of macrofagen van de lever. Deze
cellen ruimen biologisch debris en toxische partikels op. Tenslotte is er nog een laatste type
cel in de lever, in de ruimte van Disse: de stellaatcellen, die vitamine A opslaan (2, 4, 5).
1.1.2. Fysiologie
De lever vervult veel functies. Om te binnen speelt ze een belangrijke rol in het koolhydraten-
metabolisme, ze regelt de bloedglucosespiegel. Wanneer er te veel glucose in het bloed is,
zullen de lever en de spieren via de glycogenese glucose omzetten in glycogeen, dat
intracellulair opgeslagen wordt. Wanneer de glucosespiegel in het bloed te laag is, kan het
glycogeen via de glycogenolyse terug in glucose omgezet worden. Glucose kan ook gevormd
worden uit lactaat, aminozuren of glycerol via de gluconeogenese. Verder zorgt de lever er
ook voor dat galactose en fructose in glucose worden omgezet.
In het aminozuurmetabolisme komt de lever tussen bij de afbraak van aminozuren. Er vindt
deaminatie van de aminozuren plaats en er wordt ureum gevormd. De lever maakt verder alle
plasma-eiwitten aan, behalve immunoglobulinen.
Vetten worden door de lever omgezet in vetzuren. De lever maakt ook cholesterol, lipo-
proteïnen en fosfolipiden.
De lever produceert gal die bestaat uit water, elektrolieten en organische moleculen
waaronder galpigmenten, galzouten, fosfolipiden en cholesterol. De gal gaat naar de dunne
darm, eventueel na opslag in de galblaas, en zorgt daar voor de vertering en absorptie van
vetten en vetoplosbare vitaminen. Bilirubine en cholesterol, die in de gal zitten, worden
hierdoor uitgescheiden.
Een laatste functie van de lever is het omzetten van lichaamsvreemde stoffen, zoals
geneesmiddelen en toxines. Dit gebeurt door middel van fase I en fase II reacties in de lever
(2, 4, 6, 7).

5
1.2. Auto-immune hepatitis
1.2.1. Definitie
Auto-immune hepatitis (AIH) is een progressief chronische inflammatoire leverziekte. Deze
ziekte wordt gekenmerkt door de aanwezigheid van autoantilichamen,
hypergammaglobulinemie, verhoogde waarden van transaminasen en interface hepatitis op
histologisch niveau. Naargelang welke autoantilichamen er aanwezig zijn, wordt AIH
onderverdeeld in 2 types. Bij type 1 kunnen er de volgende autoantilichamen aanwezig zijn:
het anti-nuclear antibody (ANA) en/of het anti-smooth muscle antibody (SMA). Bij AIH type
2 kunnen het anti-liver kidney microsomal antibody type 1 (anti-LKM-1) en anti-liver cytosol
type 1 (anti-LC-1) aanwezig zijn (8-10).
1.2.2. Epidemiologie
AIH is een zeldzame ziekte, waardoor er niet veel epidemiologische data beschikbaar zijn. De
ziekte komt over heel de wereld voor, bij alle rassen en zowel bij kinderen als bij volwassenen
(11). De incidentie en de prevalentie zouden respectievelijk tussen de 1 en 2 per 100000 en de
10 en 20 per 100000 variëren. Het komt meer voor bij vrouwen met een ratio vrouw/man van
ongeveer 3:1. De gemiddelde leeftijd waarop de incidentie van AIH voorkomt bevindt zich
tussen de 35 en 40 jaar. AIH type 2 komt voor op een jongere leeftijd in vergelijking met type
1 en dit type vertegenwoordigt ongeveer 80% van alle AIH gevallen (11, 12). Het voorkomen
van AIH is geassocieerd met het hebben van andere auto-immuunziekten (20%) en het
voorkomen van andere auto-immuunziekten in de familie (40%) (9, 11). Voorbeelden van
auto-immuunziekten waarmee AIH geassocieerd is zijn: auto-immune thyroiditis, diabetes
type 1, inflammatoire darmziekten, rheumatoide arthritis, vitiligo en coeliakie (11).
1.2.3. Pathogenese
1.2.3.1.Genetica
De primaire oorzaak van AIH is onbekend. Wel wordt er verondersteld dat er een verband is
tussen genetica en het al dan niet krijgen van AIH. Het grootste verband wordt gezien bij
Major Histocompatibility Complex (MHC) klasse II genen, meer specifiek bij de DRB1
allelen van het Human Leukocyte Antigen (HLA) gen op de korte arm van chromosoom 6. De
rol van MHC moleculen is het presenteren van peptiden aan en het activeren van CD4/8+ T-
cellen (13).
Er bestaan momenteel 3 soorten modellen gebaseerd op de aanwezigheid van een bepaald
aminozuur op een bepaalde positie in het DRB1 allel dat geassocieerd is met AIH type 1. Het

6
eerste model veronderstelt dat er een associatie is tussen lysine (K) of arginine (R) op plaats
71 in de sequentie van 6 aminozuren: LLEQ-K/R-R van het HLA-DRB1 eiwitcomplex. Dit
werd gezien in blanke Europese en Noord-Amerikaanse patiënten met de genotypes
DRB1*0301 en DRB1*0401, waarbij het aminozuur op plaats 71 lysine is. Bij patiënten uit
Japan, Argentinië en Mexico met de genotypes DRB1*0405 en DRB1*0404 bevindt er zich
een arginine op plaats 71. Het tweede model is gebaseerd op het voorkomen van valine (V) of
glycine (G) op plaats 86 van het van het HLA-DRB1 eiwitcomplex. Deze associatie met AIH
type 1 werd gezien bij patiënten in Argentine en Brazilië. Tenslotte is er het laatste model
gebaseerd op patiënten in Japan, waarbij histidine (H) op positie 13 met AIH type 1
geassocieerd wordt.
Het ontwikkelen van AIH type 2 wordt geassocieerd met genotypes DRB1*0701 en
DRB1*0301. Patiënten met genotypes DRB1*0701 hebben een agressievere ziekte en een
slechtere prognose.
Wanneer er naar de associatie tussen HLA klasse II allelen en AIH wordt gekeken, blijken
genotypes DQB1*02, DQB1*0603, DRB1*0405 en DRB1*1301 geassocieerd te zijn met
AIH. Genotypes DRB1*1302 en DQB1*0301 blijken juist bescherming te geven.
Naast de coderende MHC genen, zouden er ook 126 non-coderende genen voor HLA zijn die
in verband met AIH kunnen gebracht worden. 60 van deze genen hebben een rol in de
regulatie van inflammatie of zijn geassocieerd met andere auto-immune ziekten.
Buiten de MHC genen worden polymorfismen van het cytotoxic T-lymphocyte associated
antigen 4 (CTLA-4), geassocieerd met AIH type 1, waarbij in exon 1 op plaats 49 adenine
door guanine vervangen is. CTLA-4 is een adhesie molecule die de perifere T-cel immune
respons controleert om overactivatie te voorkomen. Eveneens met AIH type 1 geassocieerd is
een polymorfisme op 308 van de promotor regio van TNF-α. Met AIH type 2 zijn mutaties in
de 14 exonen van het AIRE1 gen geassocieerd. AIRE1 is een transcriptie factor die betrokken
is in de clonale deletie van zelf-reactieve T-cellen. Homozygote mutaties in dat gen zijn de
oorzaak van autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED),
20% van deze patiënten hebben ook AIH type 2 (13, 14).
1.2.3.2.Immunopathogenese
De reden voor het ontstaan van AIH is tot op heden nog steeds niet gekend. Vermoedelijk is
er naast de genetische aanleg nog een trigger nodig zodat AIH zal ontstaan. Mogelijke triggers
uit de omgeving kunnen virussen en geneesmiddelen zijn. Voorbeelden van virussen zijn het
mazelenvirus, het Epstein-Barr virus, het cytomegalovirus (CMV) en hepatitis virussen.

7
Voorbeelden van geneesmiddelen zijn antibiotica, zoals Minocycline en Oxafloxacine en
statines. Ook zou er mogelijk een associatie kunnen zijn met het gebruik van anti-tumor
necrosis factor agents, zoals Adalimumab en Infliximab (10). De genetische aanleg samen
met een bepaalde trigger zal er voor zorgen dat de auto-immuniteit de overhand krijgt waarbij
tolerogene mechanismen niet meer toereikend zijn. Dit defect wordt vermoedelijk veroorzaakt
door een te kort aan regulatoire T-cellen (Tregs). Deze cellen controleren of er auto-reactieve
T-cellen zijn. Hoe minder Tregs er aanwezig zijn, hoe hoger de niveau’s van de antilichamen
zijn bij AIH, wat verbonden is met de ernst van de ziekte. Ook blijkt uit functionele studies
dat Tregs niet meer in staat zijn om de proliferatie van CD-4+ en CD8+ cellen, die betrokken
zijn in de auto-immune aanval op de lever, en hun interferon gamma (IFN-γ) productie te
reguleren. Daarnaast zijn de T-cellen ook defect om cytokines te produceren bij hun
doelwitcellen, die er normaal voor zorgen dat de functie van de Tregs in stand wordt
gehouden (9, 13).
Een ander mechanisme dat tot auto-immuniteit kan leiden is ‘molecular mimicry’. Bij
molecular mimicry worden er antilichamen / specifieke T-cellen tegen een bepaald virus
gevormd, maar het antilichaam / T-cel vertoont kruisreactiviteit en zal ook een eigen antigen
aanvallen. Dit verband is aangetoond tussen enerzijds antilichamen tegen het hepatitis C
virus, CMV, het herpes simplex virus en anderzijds tegen CYP2D6, waartegen er
autoantilichamen bij AIH worden gevormd (9, 13, 15).
Antigenen van de lever worden door antigen-presenterende cellen (APC), zoals dendritische
cellen, marcrofagen en B lymfocyten, gepresenteerd aan een ongedifferentieerde T-helper cel
(Th0 cel). Het antigen op de APC wordt omgeven door de HLA klasse I/II molecule en de
binding tussen de APC en de Th0 cel wordt geïnduceerd door de interactie van CD28 op de
Th0 cel en CD80 op APC.
Naargelang er meer IL-12 of IL-4 in de omgeving aanwezig is, zal de Th0 differentiëren in
een Th1 cel of een Th2 cel. Wanneer er veel IL-12 aanwezig is, zal de Th0 cel een Th1 cel
worden. Deze cel zal IL-2 en IFN-γ produceren, wat de cytotoxische T-cellen stimuleert, die
dan de leverschade zullen veroorzaken. De Th1 cellen activeren ook de macrofagen, die IL-1
en TNF-α secreteren. Verder zorgen de Th1 cellen ook voor een verhoogde expressie van
HLA I moleculen op APC en HLA II moleculen op hepatocyten. Normaal expresseren
hepatocyten dit niet, maar nu kunnen ze direct aan de Th1 cel het antigen presenteren.

8
Als er in de omgeving veel IL-4 aanwezig is zal de Th0 cel differentiëren in een Th2 cel.
Deze cellen produceren vooral IL-4, IL-10 en IL-13. Deze cytokines zorgen ervoor dat B-
cellen in plasmacellen veranderen, die dan autoantilichamen produceren tegen het antigen.
De leverschade wordt dus veroorzaakt door enerzijds de cytokines die de Th1 cellen
produceren, de macrofagen en de cytotoxische T-cellen, en anderzijds de natural killer (NK)
cellen die op de autoantilichamen zullen binden (9, 13).
1.2.4. Klinische presentatie en diagnose
AIH kan zich op verschillende manieren presenteren. Er kan een acuut ziektebeeld optreden
met aspecifieke symptomen, zoals abdominale pijn, misselijkheid, braken en anorexie,
gevolgd door geelzucht, bleke stoelgang en donkere urine. Het ziektebeeld kan ook minder
opvallend ontstaan, met toenemende vermoeidheid, hoofdpijn, anorexie, amenorroe,
gewichtsverlies en terugkerende geelzucht. Later worden de complicaties van portale
hypertensie duidelijk, zoals vb. het overgeven van bloed door de ontstane slokdarmvarices.
Ten slotte is het ook mogelijk dat patiënten volledig asymptomatisch blijven (9).
De diagnose wordt gesteld op basis van de aanwezigheid van autoantilichamen, die bij AIH
aanwezig zijn, verhoogde niveau’s van transaminasen en immunoglobuline G (IgG), en
interface hepatitis (9, 16).
Interface hepatitis is een histologisch kenmerk dat zich presenteert als inflammatie rond de
portale triades, te zien als verschillende niet strak afgelijnde parenchymale infiltraties van
immuuncellen. Er bevinden zich hier lymfocyten, plasmacellen en macrofagen, die de
hepatocyten aanvallen. De hepatocyten zullen een zwelling en pycnotische necrose vertonen.
Verder zal er, behalve bij milde vormen van AIH, ook fibrose te zien zijn. Interface hepatitis
komt echter niet alleen voor bij AIH, maar ook bij andere leveraandoeningen (9).

9
Figuur 3: interface hepatitis (13)
Alanine amino transferase (ALT) en aspartaat amino transferase (AST) zijn 2 enzymen die in
de lever voorkomen. Bij leverschade zijn deze enzymen in het serum verhoogd. Om de
diagnose van AIH te stellen en deze van primaire biliaire cirrose te onderscheiden, wordt de
verhouding van alkaline fosfatase (ALP) op ALT of AST gemeten (9).
De autoantilichamen worden gedetecteerd door middel van indirecte immunofluorescentie.
Naargelang welke autoantilichamen er aanwezig zijn, wordt AIH in 2 types onderverdeeld.
Bij type 1 komen de autoantilichamen antinuclear antibody (ANA) en/ of anti-smooth muscle
antibody (SMA) voor. Bij type 2 komen de autoantilichamen anti-liver-kidney microsome
type1 (anti-LMK1) en/ of anti-liver cytosol type 1 (anti-LC-1) voor (zie 1.21.). Naast deze
standaard autoantichamen kunnen ook perinucleair antineutrophil cytoplasmic antibodies
(pANCA) en anti-soluble liver antigen antibodies (anti-SLA) voorkomen. Deze
autoantilichamen kunnen wijzen op AIH in afwezigheid van de standaard autoantilichamen
(16, 17).
Bij indirecte immunofluorescentie kleurt ANA de lever, nier en maag. Het bindt aan de kern
en geeft meestal een homogeen patroon, of soms een ruw of fijn gespikkeld patroon. Het is tot
nu toe nog onbekend waarmee het precies in de kern interageert.
SMA wordt eveneens gedetecteerd op de lever, nier en maag. SMA kleurt de wand van de
arteries in de lever. De maag kleurt de muscularis mucosa en de lamina propria. In de nier
zullen bij AIH de vezels, de glomeruli en eventueel de tubuli aangekleurd zijn. Wanneer dit
allemaal aangekleurd is, komt dit overeen met het microfilamenten patroon. Maar eveneens
zoals bij ANA is de precieze interactie van SMA nog niet gekend (16, 17).

10
Anti-LKM-1 kleurt het cytoplasma van de levercellen en het 3de proximale deel van de
niertubuli. Bij anti-LKM-1 is dit sterk gekleurd, in tegenstelling tot het anti-mitochondrial
antibody (AMA) dat dit zwak aankleurt. AMA kleurt daarnaast ook de pariëtale cellen van de
maag aan. AMA is aanwezig bij primaire biliaire cirrose. Anti-LKM-1 is gericht tegen
CYP2D6, een enzym in de lever dat deel uitmaakt van het cytochroom P450-systeem. Dit
systeem zorgt er voor dat er aan farmaca een polaire groep wordt gekoppeld zodat farmaca
meer hydrofiel worden en kunnen worden geëlimineerd uit het lichaam. Dit gebeurt door een
fase 1 reactie die zowel een oxidatie, een reductie of een hydrolyse kan zijn. Maar in het
cytochroom P450-systeem gebeuren er enkel oxidaties (18). Naast anti-LKM-1 komen er nog
varianten op dit autoantilichaam voor bij AIH geassocieerd met APECED; anti-LM reageert
met CYP1A2 en anti-LKM-1 like met CYP2A6(17).
Anti-LC-1 wordt ook gedetecteerd door indirecte immunofluerescentie op de lever, nier en
maag. Het kleurt het cytoplasma van de levercellen, doordat het bindt met formimino-
transferase cyclodeaminase (FTCD) (16, 17).
Anti-SLA bindt met UGA serina transfer RNA (tRNA)-associated protein. Dit
autoantilichaam kan zowel bij AIH type 1 en type 2 gevonden worden, alsook bij AIH zonder
typebepaling door afwezigheid van ANA, SMA en anti-LKM-1, en AIH/sclerosing
cholangistis overlap syndroom. Wanneer Anti-SLA wordt gedetecteerd heeft dit een hoge
specificiteit voor AIH (16, 17).
pANCA geeft het cytoplasma van neutrofielen een perinuclear patroon, het bindt met
myeloperoxidase. In AIH type 1 komt dit autoantilichaam frequent voor, maar het bindt ook
met periferale nucleaire membraan bestanddelen, waardoor dit autoantilichaam atypisch is.
Toch kan de detectie van deze autoantilichamen op AIH wijzen indien andere
autoantilichamen afwezig zijn (16, 17).
Meestal is de diagnose van AIH duidelijk. Bij twijfel kan het diagnostisch scoring systeem
van de International Autoimmune Hepatitis Group worden gebruikt die de diagnose stelt van
zekere of waarschijnlijke AIH door aan de kenmerken van de verschillende parameters, die
bij de diagnose van AIH belangrijk zijn, punten te geven (16, 17).

11
1.2.5. Behandeling
Na het stellen van de diagnose moeten alle patiënten met AIH behandeld worden, behalve
asymptomatische patiënten met milde interface hepatitis, waarbij de voordelen van de
behandeling niet opwegen tegen de bijwerkingen ervan. Deze patiënten moeten wel worden
opgevolgd om na te gaan of hun symptomen niet verergeren. De behandeling van AIH heeft
als doel de niveau’s van ALT, AST en IgG weer te normaliseren en de inflammatie in de lever
te verminderen. Daarbij zullen de symptomen verbeteren en wordt de overleving verlengd
(19).
De standaardbehandeling bestaat uit het toedienen van Prednisolone of Prednisone.
Prednisone wordt in de lever omgezet tot Prednisolone, maar dit kan niet goed doorgaan bij
patiënten met levercirrose. Prednisolone is een corticosteroïd dat immunosuppressief werkt.
Prednisone heeft echter ook veel bijwerkingen, zoals het ontwikkelen van het syndroom van
Cushing. Hierbij verandert de vetverdeling, er wordt meer vet ter hoogte van de buik
opgeslagen, terwijl de ledematen vermageren, patiënten krijgen een ronder gezicht,
gewichtstoename, acne,… Ernstigere bijwerkingen zijn osteoporose, diabetes, hypertensie,
cataract en psychoses (19, 20).
Vaak wordt Prednisolone gecombineerd met Azathioprine. Azathioprine is ook een
immunosuppressor die de nucleïnezuursynthese in T- en B-lymfocyten inhibeert. De ‘Practice
guidelines for the diagnosis and treatment of AIH’ van de American Association for the Study
of Liver Diseases(AASLD) raadt aan om met een dosis van 30 mg Prednisolone per dag te
starten. Daarbij wordt er ook Azathioprine toegediend in een dosis van 1 mg/kg per dag. Over
een periode van 2 tot 3 maanden wordt de Prednisolone afgebouwd tot 10mg/dag als de ALT
en AST niveau’s beginnen te dalen (19).
Wanneer Prednisolone te veel bijwerkingen geeft, kan er in de plaats 9 mg Budesonide per
dag gegeven worden, een 2de generatie corticosteroïd dat ook immunosuppressief werkt. Dit
kan echter niet gebruikt worden wanneer er al levercirrose aanwezig is, omdat bij deze
patiënten Budesonide het first pass effect niet kan ondergaan waardoor de concentratie van
Budesonide niet genoeg zal dalen en er zich zo te veel bijwerkingen zullen ontwikkelen (20,
21).
Azathioprine kan ook ernstige bijwerkingen geven, waaronder beenmerg depressie. De
actieve metabolieten van Azathioprine worden afgebroken door thiopurine methyltransferase.
Sommige mensen zijn homozygoot voor het lage activiteit allel van dit gen, en zullen

12
bijgevolg Azathioprine trager afbreken en meer bijwerkingen hebben. Bij deze pariënten kan
er Mycophenolate in de plaats van Azathioprine worden toegediend, een immunosuppressor
die inwerkt op de purine synthese bij T- en B-lymfocyten (19-21).
Er bestaat discussie over hoelang de behandeling moet worden aangehouden. De AASLD
raadt aan om gedurende 2 jaar 5-10 mg Prednisoline per dag en 1 mg Azathioprine /kg per
dag te geven en minstens nog 1 jaar na de normalisatie van de ALT en AST waarden (19).
20% van de patiënten met AIH type 1 zou succesvol blijvend kunnen stoppen met de
behandeling, terwijl dit zelden is bij AIH type 2 (9).
Ondanks de bestaande medicamenteuze behandeling van AIH is toch soms een
levertransplantatie nodig. Dit is nodig bij patiënten waarbij fulminant leverfalen voorkomt.
Dit is het geval bij 10-20% van de kinderen met AIH, of wanneer er te laat met de
behandeling werd gestart of indien er al cirrose aanwezig was op het moment van de
diagnose. De 5-jaar overleving is 80-90%, maar bij ongeveer 1/3 van de getransplanteerde
patiënten zal AIH weer terugkeren. Er wordt aangeraden om de patiënten te behandelen met
steroïden die immuunsupressief werken (9, 10, 22).
1.2.6. Leverfibrose en hypoxie
De vorming van fibrose in de lever is een reactie op de inflammatie die ontstaat door de auto-
immune aanval op de lever. De initiatie van deze reactie begint met het activeren van de
stellaatcellen in de lever, die de functie om vitamine A op te slaan verliezen. De genexpressie
en de fenotypes van deze cellen veranderen door paracriene stimulatie van inflammatoire
cellen, beschadigde hepatocyten en veranderingen in de extracellulaire matrix. De
geactiveerde stellaatcellen zullen prolifereren en chemotaxisch worden. Cytokines die
geproduceerd werden bij de inflammatie zullen de stellaatcellen aantrekken. Onder invloed
van TGF-β1 zullen de stellaatcellen zullen ook meer matrix produceren, onder andere
collageen I, fibronectine en proteoglycanen. Daarnaast zullen de stellaatcellen meer
contractiliteit vertonen, waardoor de portale weerstand zal verhogen. De stellaatcellen
produceren ook cytokines die inflammatie cellen aantrekken (23, 24).
Bij leverfibrose is het evenwicht tussen het aanmaken en afbreken van de matrix verstoort. De
levermatrix wordt eerst vervangen door een litteken matrix. Deze matrix kan worden
afgebroken door matrix-metalloproteinases (MMPs). Deze enzymen worden geproduceerd
door de stellaatcellen, maar worden geïnhibeerd door tissue inhibitors of metalloproteinases
(TIMPs), eveneens door stellaatcellen geproduceerd. Daarnaast is TIMP-1 anti-apoptotisch
tegenover stellaatcellen. Daardoor zorgen deze TIMPs er niet enkel voor dat de matrix minder

13
wordt afgebroken, maar ook dat geactiveerde stellaatcellen niet kunnen vernietigd worden. Er
zal dus meer en meer matrix, met onder andere collageen, aanwezig zijn in de lever. Cirrose is
het eindstadium was leverfibrose, het leverparenchym zal zodanig veranderd zijn dat er
nodule vorming is en de leverfunctie veranderd is (23, 24).
Door schade in de lever ontstaat er ook hypoxie. Hypoxia inducible factors (Hifs) reageren op
een te kort aan zuurstof. Hifs zijn transcriptiefactoren die bestaan uit een α- en een β-subunit.
Er zijn 3 α-subunits: Hif1α, Hif2α en Hif3α en 1 β-subunit: Hif1β. Onder normale
omstandigheden worden de α-subunits van Hif gehydroxyleerd door prolyl-hydroxylases
(Phd1, Phd2 en Phd3) en afgebroken, zodat ze niet kunnen binden met de β-subunit van Hif.
Onder hypoxische omstandigheden zullen deze hydroxyleringen niet doorgaan en kunnen de
α-subunits wel binden met de β-subunit. Deze actieve Hifs migreren vervolgens naar de kern
en binden op hypoxia responsive elements (HRE) in de promotor regio van bepaalde genen.
Dit is het geval bij de Hif1α en de Hif2α subunit, de functie van Hif3α is tot nu toe nog
onbekend. De genen die door Hif1α gereguleerd worden zijn genen die te maken hebben met
erythropoëse en het ijzermetabolisme, angiogenese en het glucose metabolisme (25, 26).
Voorbeelden van deze genen zijn Vascular endothelial growth factor (VEGF),
Phosphofructokinase (PFK) en Glucose transporter 1 (Glut1) (25, 27).
Verder blijkt ook dat Hif1α een regulator van fibrose is. Hif1a reguleert genen die een rol
spelen in de activatie van stellaatcellen, chemokinereceptoren 1 en 5 (CCR-1 en CCR-5). Ook
de PDGF-B in de Kupffer-cellen, die zo de Stellaatcellen mee activeren, worden door Hif1a
opgereguleerd. Verder worden ook de angiogenese en de collageensynthese, door meer
expressie van de IL-13 receptor α1, door Hif1a gereguleerd (28-30).
1.2.7. Muismodellen voor AIH
In de eerste soort muismodellen voor AIH werd er experimentele AIH (EAH) geïnduceerd.
Dit werd gedaan door allogene leverhomogenaten met Complete Freud’s adjuvant (CFA) in
muizen te injecteren. In de lever ontstond er periportale infiltraten en necrosis van de
hepatocyten. Gelijkaardige resultaten werden bekomen door SMA met leverhomogenaten en
lysaten van Klebsiela pneumonia te injecteren. Bij het injecteren van leverhomogenaten met
CFA bleek het histologisch beeld op AIH te lijken, ook ALT steeg, maar er werden geen
antilichamen gevormd die bij AIH in de mens voorkomen.

14
De tweede mogelijkheid die werd uitgeprobeerd was het cytokine model. Hier werd een
transgeen muismodel gemaakt waarbij IFN-γ tot expressie kwam. Dit leidde tot verhoogde
transaminasen waarden en inflammatie en necrose in de lever.
Verder werden er verschillende andere transgene muismodellen ontwikkeld. Hierbij werden
zelf-neoantigenen tot expressie gebracht in de hoop via kruisreactiviteit de lever aan te vallen.
Vele pogingen faalden echter door de inductie van tolerantie. Om toch hepatitis te
veroorzaken was het noodzakelijk om inflammatoire factoren tot expressie te brengen of de
muizen te infecteren met een pathogeen. De hepatitis was echter maar tijdelijk en daarnaast
werden CD8+ T-cellen geactiveerd, terwijl bij AIH vooral CD4+ T-cellen geactiveerd zijn
(31, 32).
Een andere aanpak was het injecteren van dendritische cellen (DC) geladen met
gedifferenteerde hepatocellulaire carcinoom cellen in C57BL/6 muizen, gevolgd door
intraperitonale injectie met IL-12. Dit leidde tot inflammatie zoals gezien bij AIH (33).
Een beter muismodel voor AIH is het murine model van AIH type 2. C57BL/6 muizen
werden geïnfecteerd met plasmiden met humaan CYP2D6 en humaan FTCD. De muizen
ontwikkelden AIH met inflammatie, necrose, anti-LKM-1 en anti-LC-1, verhoogde waarden
van ALT, maar geen fibrose (34). Het meest succesvolle muismodel tot nu toe lijkt het model
waarbij FVB/N muizen geïnfecteerd werden met een Adenovirus dat het humaan CYP2D6 tot
expressie bracht. De muizen ontwikkelden een meer blijvende AIH met anti-LKM-1 en
subcapsulaire fibrose. De fibrose evolueerde wel niet zoals bij de mens naar cirrose (35).
1.3. Probleemstelling
De huidige muismodellen van AIH zijn niet representatief genoeg. Deze hebben geen of
beperkte fibrose, wat niet representatief is voor AIH bij de mens. Er was ook geen evolutie
naar cirrose in de dieren die in leven bleven. Daarom werd er geprobeerd om een nieuw
muismodel voor AIH te ontwikkelen, om zo de pathofysiologie van AIH te ontrafelen en een
therapie te ontwikkelen.

15
2. Materialen en methoden
2.1. Keuze van het muismodel
In de literatuur is er nog geen beschrijving gevonden van een muismodel dat het volledige
spectrum van de pathofysiologie van AIH in patiënten kan nabootsen. In de literatuur zijn er
wel al verschillende, al dan niet genetisch gemodificeerde, muismodellen beschreven. Deze
muismodellen werden ontwikkeld aan de hand van C57BL/6, 129/Sv en BALB/c muizen.
Voor onze experimenten hebben we besloten om te werken met vrouwelijke C57BL/6 muizen
van 8 weken oud.
2.2. Inductie van auto-reactieve T-cellen
2.2.1. Immunisatie met CYP2D6
Om auto-reactieve T-cellen tegen CYP2D6 in muizen te induceren, werden 5 muizen
geïmmuniseerd met CYP2D6. Twee muizen, die als positieve controle dienen, werden
geïmmuniseerd met Ovalbumine, wat tot de inductie van auto-reactieve T-cellen leidt. De
negatieve controle bestaat uit 2 muizen geïmmuniseerd met truncated nerve growth factor
receptor (tNGFR), waarvan aangetoond is dat dit geen auto-reactieve T-cellen doet ontstaan
(zie tabel 1).
Alle 9 muizen werden eerst intraperitoneaal verdoofd met een ketamine / xylazine oplossing,
daarna werden de muizen geschoren met een scheermesje rondom het gebied waar beide
inguinale lymfeknopen liggen. De inguinale lymfeknoop is een blauwzwarte plek iets boven
de knie, waar bloedvaten samenkomen. Ter hoogte van deze lymfeknoop werd een kleine
incisie in de huid gemaakt. Daarna werd deze incisie verder opengeknipt. De inguinale
lymfeknoop werd samen met het omliggende vetweefsel vastgenomen. De 5 muizen uit de
CYP2D6-groep werden in deze lymfeknopen geïnjecteerd met 10µg CYP2D6 mRNA en
Trimix. De Trimix bestaat uit een mix van 3 mRNA’s: CD40 ligand (CD40L), constitutief
actieve toll like receptor 4 (TLR4) en CD70, ligand van CD27. Dit dient om het
immuunsysteem extra te activeren, zodat er mogelijks autoreactieve T-cellen tegen CYP2D6
ontstaan. De 2 muizen uit de positieve groep werden geïnjecteerd met Ovalbumine mRNA en
de Trimix, en de 2 muizen uit de negatieve groep werden geïnjecteerd met tNGFR mRNA en
de Trimix. De mRNA’s werden opgelost in Ringerlactaat. In totaal werd er een volume van
10µl in beide inguinale lymfeknopen geïnjecteerd. Vervolgens werden de incisies weer
dichtgenaaid.

16
Tabel 1: Tijdslijn experimentTijdslijn experiment
Inductie auto-reactieve T-cellen Validatie muismodel
dag 1 dag 4 dag 9 dag 10 na 3 maandna 3 maanden 10 dagen
5 CYP2D6muizen
injectieCYP2D6/Trimix
in beidelymfeknopen
injectieCYP2D6/Trimix
in beidelymfeknopen
injectiesgeladen
splenocytenanalyseFACS
injectieCYP2D6/Trimix
in 1lymfeknoop sacrificatie
2 positievecontrolemuizen
injectieOvalbumine/Trimix
in beidelymfeknopen
injectieOvalbumine/Trimix
in beidelymfeknopen
injectiesgeladen
splenocytenanalyseFACS - sacrificatie
2 negatievecontrolemuizen
injectietNGFR/Trimix
in beidelymfeknopen
injectietNGFR/Trimix
in beidelymfeknopen
injectiesgeladen
splenocytenanalyseFACS - sacrificatie
2 naïevemuizen - -
injectiesgeladen
splenocytenanalyseFACS - sacrificatie
2.2.2. In vivo cytotoxic T lymphocyte assay (CTL)
Door middel van de CTL werd gekeken of er auto-reactieve T-cellen tegen CYP2D6
aanwezig waren door de uitgevoerde immunisatie. Dit werd getest door splenocyten in de
muizen te injecteren die geladen waren met peptiden van het CYP2D6 eiwit. Daarnaast
werden ook splenocyten geïnjecteerd die niet geladen zijn met peptiden. Wanneer er auto-
reactieve T-cellen aanwezig zijn, zouden de splenocyten geladen met peptiden van CYP2D6
vernietigd worden. De splenocyten zonder peptiden zouden niet vernietigd worden door de
auto-reactieve T-cellen (zie tabel 2).

17
Tabel 2: injectie splenocyten cytotoxic T lymphocyte assay (CTL)splenocyten met hoge
concentratie CFSE, geladen
met:
splenocyten met lage
concentratie CFSE, geladen
met:
5 CYP2D6 muizen 3 peptiden van CYP2D6 geen peptiden
2 positieve controle
muizen peptiden van Ovalbumine geen peptiden
2 negatieve controle
muizen geen peptiden geen peptiden
2 naïeve muizen 3 peptiden van CYP2D6 geen peptiden
De splenocyten, die nodig waren om in de muizen te injecteren, werden uit 3 muizen
geïsoleerd. De 3 muizen werden eerst gesacrificeerd door middel van cervicale dislocatie en
vervolgens werd de milt gepreleveerd. De milten werden fijngemalen op een 40µm nylon
filter. Onder de filter bevond zich een 6-well plaat met 5ml RBS lysis buffer. Daarna werden
de splenocyten overgebracht in een 50ml tube en werd er een ruime hoeveelheid phosphate
buffered saline (PBS) toegevoegd. Dit geheel werd gecentrifugeerd bij 5°C bij 1500 rpm.
Daarna werden de splenocyten geresuspendeerd in 10 ml RPMI+ (zie tabel 3) en werden de
cellen met een Bürker telkamer geteld.
Tabel 3: Samenstelling gebruikte celmedia
RPMI+40ml RMPI-1640 (Roswell Park Memorial Institute medium) + 2ml 5% FCI
+ 40µl 50µM β-mercaptoethanol
RPMI-10 40ml RMPI-1640 + 4ml 10% FCI
PBS/0,1% BSA 0,15g BSA(bovine serum albumin) + 150ml PBS(phosphate buffered saline)
De splenocyten werden heropgelost in RMPI+ zodat de uiteindelijke concentratie gelijk werd
aan 20.106 splenocyten/ml. De oplossing werd verdeeld over 2 tubes. De splenocyten in de
ene tube werden geladen met CYP2D6 peptiden en de splenocyten in de andere tube werden
niet geladen met peptiden.
Aan de ene tube werden 3 peptiden van het CYP2D6 eiwit toegevoegd, namelijk peptide 1:
394-402, peptide 2: 290-295 en peptide 3: 287-295, zodat de splenocyten geladen werden met
deze 3 peptiden. De peptiden werden toegevoegd zodat de concentratie in de tube 5µg/ml
bedroeg voor elk van de peptiden. Aan de andere tube werd niets toegevoegd. De incubatie

18
van de splenocyten vond plaats bij 37°C en 5% CO2 gedurende 1,5 u. Vervolgens werden de
splenocyten in beide tubes gecentrifugeerd gedurende 7 min. bij 1500 rpm en werd het
supernatans verwijderd. De splenocyten werden gewassen met RPMI-10 (zie tabel 3) door het
centrifugeren van de splenocyten bij 1500 rpm gedurende 7 min.
Met behulp van een fluorescence-activated cell sorter (FACS) werd bepaald hoeveel
splenocyten van beide populaties overbleven. Hiervoor werd er een fluorescente stof,
carboxyfluorescein succinimidyl ester (CFSE) aan de splenocyten toegevoegd. FACS is een
toestel dat gebruikt wordt om eigenschappen van cellen te identificeren. Dit gebeurt door een
monoclonaal antilichaam met een kleurstof aan de cellen te binden, in dit geval hier CFSE. De
cellen werden samen met een groot volume aan fysiologische zoutoplossing door een
spuitstuk gedreven, zodat er een fijne straal van cellen met vloeistof ontstond. Elke cel
passeerde afzonderlijk een laserstraal, die er voor zorgde dat de fluorescente kleurstof op de
cel geëxciteerd werd en fluoresceerde. De fluorescentie werd vervolgens waargenomen door
een sensitieve fotomultiplicator, zodat het gekend is welk monoclonaal antilichaam doorheen
de FACS passeerde. De fotomultiplicator nam daarnaast ook de grootte en granulariteit van de
cellen waar door middel van de verstrooiing van het licht. Daarna kwam de cel in de cell
sorter. De computer heeft ondertussen waargenomen welke eigenschappen deze cel heeft. Op
basis hiervan kan de computer een elektrische lading genereren juist op het moment dat de cel
met vloeistof voorbij de laserstraal passeert in een druppeltje, zodat enkel dit druppeltje
geladen worden. Vervolgens komen de mogelijks geladen druppeltjes die elk een cel bevatten,
voorbij twee tegengestelde geladen platen, en worden de cellen op basis van hun lading
gesorteerd (15).
In dit experiment werden alle splenocyten met CFSE geladen. Om daarna met de FACS te
weten of het om met peptiden geladen splenocyten ging of niet, werd het CFSE in
verschillende concentraties toegevoegd. De splenocyten die al geladen waren met peptiden
van het CYP2D6 eiwit werden ook geladen met een hoge concentratie aan CFSE. De
splenocyten die niet geladen waren met peptiden, kregen een lage concentratie aan CFSE (zie
tabel 2). Bij de analyse met de FACS op basis van de concentratie van CFSE kon bepaald
worden tot welke populatie de splenocyten behoorden. De FACS gaf een histogram weer dat
de concentratie aan CFSE uitzette tegenover het aantal cellen. Indien er auto-reactieve T-
cellen tegen het CYP2D6 aanwezig waren, dan zouden de splenocyten geladen met de
peptiden en de hoge concentratie aan CFSE verminderd zijn (zie figuur 4).

19
Figuur 4: resultaat van de FACS wanneer er auto-reactieve T-cellen aanwezig zijn
Vervolgens werden de splenocyten met CFSE geladen. Beide splenocyten populaties werden
elk opgelost in 5 ml PBS/0,1%BSA. Vervolgens werden er 2 verdunningen van CFSE
aangemaakt, de hoogste concentratie was 2,5µM en de laagste 0,25µM. Beide splenocyten
populaties werden geïncubeerd bij 37°C en 5% CO2 gedurende 10 min.
Daarna werden de splenocyten populaties gecentrifugeerd gedurende 7 min bij 1500 rpm en
werd het supernatans verwijderd. De splenocyten werden gewassen door 10 ml RPMI-10 toe
te voegen, te centrifugeren en het supernatans af te nemen. De splenocyten werden
heropgelost in 15 ml PBS en de viabele cellen werden met behulp van de Bürker telkamer
geteld. Gelijke volumes van beide splenocyten populaties werden gemengd en met de FACS
werd er nagegaan of er in de oplossing inderdaad evenveel splenocyten van beide populaties
aanwezig waren. Daarna werden de splenocyten weer 7 minuten gecentrifugeerd bij 1500 rpm
en werd het supernatans verwijderd. De splenocyten werden weer opgelost in PBS zodat het
aantal splenocyten dat in de muis werd geïnjecteerd tussen de 10.106 en de 20.106 splenocyten
lag. Per muis werd er 200 µl in de staartvene geïnjecteerd.
De 5 muizen die 5 dagen eerder voor de 2de keer geïmmuniseerd werden met CYP2D6
werden hiermee geïnjecteerd. Daarnaast werden er nog 2 naïeve muizen geïnjecteerd met deze
splenocyten. Bij de positieve controle, waarbij 2 muizen met Ovalbumine werden
geïmmuniseerd, werden deze geïnjecteerd met splenocyten bereid volgens hetzelfde protocol
als hierboven. De splenocyten werden hier evenwel niet geladen met peptiden van het
CYP2D6 eiwit, maar met het CD8 peptide SIINFEKL van Ovalbumine. Aangezien
Ovalbumine auto-reactieve T-cellen induceert, zullen de splenocyten met Ovalbumine
vernietigd worden. Bij de negatieve controle, waarbij 2 muizen met tNGFR werden
geïmmuniseerd, ontstaan er geen auto-reactieve cellen. De muizen werden geïnjecteerd met
splenocyten die zowel bij hoge als lage concentratie aan CFSE niet geladen waren met
peptiden. De splenocyten zullen bij de negatieve controle niet vernietigd worden (zie tabel 2).

20
Na 24 u. werd er van alle muizen ongeveer 12 druppels bloed via de staartvene afgenomen.
De rode bloedcellen werden gelyseerd met RBS lysis buffer, zodat enkel de splenocyten
overbleven. De splenocyten werden gewassen en daarna werden ze geanalyseerd met FACS.
2.3. Validatie van het muismodel na 3 maanden
2.3.1. Extra immunisatie met CYP2D6
De oorspronkelijke 5 muizen die met CYP2D6 werden geïmmuniseerd, werden 3 maanden
later nog 1 keer met CYP2D6 en Trimix geïnjecteerd. Dit gebeurde in 1 lymfeknoop en
volgens hetzelfde protocol als hierboven beschreven (zie tabel 1 en 2.2.1.).
2.3.2. Bepaling antilichamen en leverwaarden
Voor de bepaling van de ALT en AST waarden van de lever en autoantilichamen die bij AIH
voorkomen werd er 2 keer bloed afgenomen. Het bloed werd retro-orbitaal afgenomen via de
vena opthalamica. Na 15 min werd het bloed gecentrifugeerd gedurende 10 min op 12000 rpm
bij 4°C, en het serum werd afgenomen. De eerste keer werd er ongeveer 250µl bloed per muis
afgenomen voor de analyse van de specifieke antilichamen, AMA, p-ANCA, ANA en Anti-
LKM-1. Bij de tweede retro-orbitale bloedafname, 3 dagen later, werd er ongeveer 100µl
bloed afgenomen voor de analyse van ALT en AST waarden.
2.3.3. Sacrificatie
Alle 11 muizen werden na 3 maanden gesacrificeerd (zie tabel 1). Eerst werden de muizen
verdoofd door middel van een intraperitoneale injectie met 15µl xylazine-ketamine. De
muizen werden gewogen en extern geanalyseerd. Het bloed werd retro-orbitaal afgenomen en
daarna werden de muizen gesacrificeerd door een cervicale dislocatie. De lever werd
gepreleveerd en gewogen. Daarna werden er kleine stukjes van de lever gesneden en in een
cryotube met RNA later (Ambion Inc., België) bewaard. De rest van de lever werd in een
histologiecassette in formol bewaard.
Vervolgens werd de milt uit de muis gehaald en gewogen. De milt werd in een
histologiecassette geplaatst. Verder werden er ook een stukje van de maag, dunne darm, dikke
darm, hart, long en de nier in een histologiecassette geplaatst. De histologiecassettes werden
gedurende één nacht in formol bewaard.

21
2.3.4. Paraffinecoupes
Alle afgenomen organen werden ingebed om er daarna coupes voor kleuringen te kunnen van
maken. Het inbedden van de coupes in paraffine gebeurde op de dienst pathologie van het
Universitair Ziekenhuis Gent (UZ Gent). Ze werden eerst gefixeerd met verdunde formaline.
Hierna werden de stalen in een aantal baden met alcohol van toenemende sterkte (30-50-70-
96-99 %) gebracht om heel langzaam het water in de cellen te vervangen door zuivere alcohol
en de alcohol daarna te vervangen door een stof waarin vetten oplosbaar zijn, zoals het hier
gebruikte xyleen. Het resulterende blokje weefsel werd dan in paraffine ingebed en werd met
behulp van een microtoom in coupes van 5 µm gesneden. Er werd een rotatiemicrotoom
gebruikt (Leica Microsystems, Duitsland). De coupes werden gedurende één nacht bij 37°C in
een warme lucht oven geplaatst (Binder, Duitsland).
2.4. Histologie
2.4.1. Haematoxyline en eosine (H&E) kleuring op de lever
De Haematoxyline-eosine kleuring is een kleuring met 2 kleuren. Haemotoxyline is een
basische kleurstof die zure bestanddelen paars-blauw aankleurt, zoals de kern of andere
structuren die veel DNA of RNA bevatten en dus zuur zijn. Eosine daarentegen is een zure
kleurstof die basische bestanddelen roze-rood aankleurt. De meeste cytoplasmatische
bestanddelen zijn basisch, waardoor het cytoplasma met een H&E kleuring roze-rood
aankleurt.
Deze kleuring werd hier uitgevoerd op de lever, om eventuele inflammatie te kunnen
aantonen.
De paraffinecoupes werden gedeparaffineerd in xyleen (3 maal 5 minuten) (Klinipath, België)
en gerehydrateerd in dalende concentraties ethanol (99 %, 96 %, 70 %, 50 %; telkens 2 maal 5
minuten) (Klinipath, België) en gedurende 1 maal 5 minuten in gedemineraliseerd water of
aqua dest (AD). Vervolgens werden ze 5 minuten in een Mayer’s haematoxyline-oplossing
(Sigma, België) geplaatst, gevolgd door 3 seconden in een 1 % HCl oplossing (Merck,
Duitsland). Daarna werden de coupes 1 minuut in een 1 % eosine oplossing geplaatst (Merck,
Duitsland). Dehydratatie van de coupes gebeurde in AD gevolgd door een opeenvolgende
reeks van baden met stijgende concentraties ethanol (50 %, 70 %, 96 %, 99 %; telkens 2 maal
5 minuten) (Klinipath, België) en xyleen (3 maal 5 minuten) (Klinipath, België). Ten slotte
werden de coupes gemonteerd met Neutral Mounting Medium (D.P.X) (Klinipath, België) en
een dekglaasje (VWR International, België).

22
2.4.2. Sirius Red kleuring op de lever
Sirius Red is een kleurstof die collageen rood aankleurt, doordat de sulfonische zure groepen
van Sirius Red met de basische groepen van collageen binden. Deze kleuring werd hier
uitgevoerd op de lever om te kijken of er collageen aanwezig is en dus fibrose.
De paraffinecoupes werden gedeparaffineerd in xyleen (Klinipath, België) en gerehydrateerd
in dalende concentraties ethanol (99 %, 96 %, 70 %, 50 %; telkens 2 maal 5 minuten)
(Klinipath, België) en gedurende 1 maal 5 minuten in AD. Vervolgens werden de coupes voor
30 minuten in een 0,1 % Sirius Red oplossing (Sirius Red poeder (Klinipath, België) in
picrinezuur (Sigma, België)) geplaatst. Dehydratatie van de coupes gebeurde in AD gevolgd
door een opeenvolgende reeks van baden met stijgende concentraties ethanol (50 %, 70 %, 96
%, 99 %; telkens 2 maal 5 minuten) (Klinipath, België) en xyleen (3 x 5 minuten) (Klinipath,
België). Ten slotte werden de coupes gemonteerd met Neutral Mounting Medium (D.P.X)
(Klinipath, België) en een dekglaasje (VWR International, België).
2.5. Immunohistochemische kleuringen
2.5.1. HIF-1α kleuring op de lever
HIF-1α wordt meer tot expressie gebracht bij hypoxie en dus mogelijks fibrose. Daarom werd
er hier een kleuring tegen HIF-1α uitgevoerd op de lever om te kijken of er fibrose was.
De paraffinecoupes werden gedeparaffineerd in xyleen (Klinipath, België), 3 maal 5 minuten
en daarna gerehydrateerd in dalende concentraties ethanol (99%, 96%, telkens 2 maal 5
minuten en 20 maal onderdompelen in 70% en 50% ethanol) (Klinipath, België) en gedurende
1 maal 5 minuten in AD. Daarna werden de coupes eerst in kraantjeswater en dan in AD
gespoeld, telkens gedurende 5 minuten. Vervolgens volgde de ‘antigen retrieval’ stap. De
coupes werden 30 minuten ondergedompeld in een citraatbuffer bij 95°C. Na 15 minuten
afkoelen in een citraatbuffer werden de coupes gespoeld in PBS en PBS met 0,1% BSA (2
maal 5 minuten en 1 maal 5 minuten). De endogene peroxidasen werden geblokt door de
coupes geduren 10 minuten onder te dompelen in PBS met 3% waterstofperoxide (VWR
international, België). Daarna werden de coupes 3 maal 5 minuten gespoeld in PBS en 30
minuten in PBS met 1% BSA bij 37°C.
Daarna werden de coupes geïncubeerd met het primaire antilichaam, Santa Cruz antibody n°
SC-53546 HIF1α (Santa Cruz Biotechnology, Duitsland), dat 1/400 verdund was in PBS met
0,1% BSA. Per coupe werd er 300µl aangebracht en de incubatie duurde 1 uur bij 37°C.
Hierna werden de coupes 3 x 5 minuten gespoeld in PBS.

23
Vervolgens werden de coupes geïncubeerd met het secundair antilichaam Biotinylated link
Universal (LSAB+ system-HRP) (Dako Cytomation, VSA), gedurende 10 min. Daarna
werden de coupes 3 maal 5 minuten gespoeld met PBS. Vervolgens werden ze 10 minuten
geïncubeerd met Streptavidin-HRP (LSAB+ system-HRP) (Dako Cytomation, VSA) en
daarna 3 maal 5 minuten gespoeld met PBS. Op elke coupe werd 3,3’ diaminobenzidine
(DAB) (Dako Cytomation, VSA) aangebracht, gedurende 1 minuut, totdat de coupes
verkleurden. De kleuring werd gestopt door 5 minuten te spoelen in AD. Hierna werd een
tegenkleuring uitgevoerd met Mayer’s haematoxyline (Sigma, België) gedurende 3 minuten,
waarna de coupes 10 minuten in stromend water werden geplaatst. Tenslotte werden de
coupes weer gedehydrateerd door de coupes eerst 5 minuten onder te dompelen in AD,
vervolgens telkens 20 maal onder te dompelen in 50% en 70% ethanol, 2 maal 5 minuten in
96% ethanol, 2 maal 5 minuten 99% ethanol en 3 maal 5 minuten in xyleen. De coupes
werden gemonteerd met Neutral Mounting Medium (D.P.X) (Klinipath, België) en een
dekglaasje (VWR International, België).
2.5.2. Glut1 kleuring op de lever
Glut 1 is een merker voor hypoxie. Wanneer er hypoxie in de levercellen aanwezig is, zal er
meer Glut 1 aanwezig zijn.
De paraffinecoupes werden gedeparaffineerd in xyleen (3 maal 5 minuten) (Klinipath, België)
en gerehydrateerd in dalende concentratie ethanol (telkens 2 maal 5 minuten in 99% ethanol
en 96% ethanol en telkens 20 maal onderdompelen in 70% ethanol en 50% ethanol)
(Klinipath, België) en 1 maal 5 minuten in AD. Vervolgens volgde de ‘antigen retrieval’ stap.
De coupes werden 30 minuten ondergedompeld in een citraatbuffer bij 95°C en 15 minuten
afgekoeld in een citraatbuffer. Daarna werden de coupes 2 maal 5 minuten gespoeld in PBS
met 0,1% BSA. De endogene peroxidasen werden geblokt door de coupes 10 minuten onder
te dompelen in AD met 3% waterstofperoxidase. De coupes werden daarna 2 maal 5 minuten
gespoeld met PBS. Vervolgens werden de proteïnen geblokt door de coupes 15 minuten onder
te dompelen in proteinblock (Dako Cytomation, VSA).
Daarna werden de coupes met het primaire antilichaam, Glut 1 (Abcam, UK), geïncubeerd
gedurende 1,5 u. Het antilichaam was 1/400 verdund in PBS met 0,1% BSA en per coupe
werd er 300 µl aangebracht. Hierna werden de coupes 3 x 5 minuten gespoeld in PBS.
Vervolgens werd er op de coupes het secundair antilichaam aangebracht, Biotinylated link
Universal (LSAB+ system-HRP) (Dako Cytomation, VSA), gedurende 10 min. Daarna
werden de coupes 3 maal 5 minuten gespoeld met PBS. Vervolgens werden ze 10 minuten

24
geïncubeerd met Streptavidin-HRP (LSAB+ system-HRP) (Dako Cytomation, VSA) en
daarna 3 maal 5 minuten gespoeld met PBS. Op elke coupe werd DAB (Dako Cytomation,
VSA) aangebracht, gedurende 15 seconden, totdat de coupes verkleurden. De kleuring werd
gestopt door 5 minuten te spoelen in AD. Hierna werd een tegenkleuring uitgevoerd met
Mayer’s haematoxyline (Sigma, België) gedurende 3 minuten, waarna de coupes 10 minuten
in stromend water werden geplaatst. Tenslotte werden de coupes weer gedehydrateerd door de
coupes eerst 5 minuten onder te dompelen in AD, vervolgens telkens 20 maal onder te
dompelen in 50% en 70% ethanol, 2 maal 5 minuten in 96% ethanol, 2 maal 5 minuten 99%
ethanol en 3 maal 5 minuten in xyleen. De coupes werden gemonteerd met Neutral Mounting
Medium (D.P.X) (Klinipath, België) en een dekglaasje (VWR International, België).
2.5.3. F4/80 kleuring op de lever
F4/80 is een merker die gebruikt wordt om inflammatie aan te tonen. Deze kleuring werd dus
uitgevoerd om te kijken of er inflammatie in de lever aanwezig was.
Paraffinecoupes werden gedeparaffineerd in xyleen (3 maal 5 minuten) (Klinipath, België) en
gerehydrateerd in dalende concentraties ethanol (99 %, 96 %, 70 %, 50 %; telkens 2 maal 5
minuten) (Klinipath, België) en gedurende 1 maal 5 minuten in AD. Eerst volgde de ‘antigen
retrieval’ stap. De coupes werden 15 minuten in trypsine geplaatst bij een temperatuur van
37°C, gevolgd door een spoelbeurt met 0,1% BSA in PBS (3 maal 5 minuten). Nadien werden
ze 10 minuten in een citraatbuffer ondergedompeld en dit bij 95°C. Een afkoeling van 15
minuten werd gevolgd door opnieuw een spoelbeurt met 0,1% BSA in PBS (3 maal 5
minuten). De endogene peroxidasen werden geblokt door de coupes gedurende 15 minuten
onder te dompelen in 3 % waterstofperoxide (in PBS) (VWR international, België), te spoelen
met Tris buffered saline (TBS) gedurende 3 maal 5 minuten en daarna 30 minuten in een 1%
BSA oplossing bij 37°C.
Daarna werden de coupes geïncubeerd met een primair antilichaam, namelijk het rat-anti-
mouse F4/80 antilichaam (AbD serotec, Duitsland), 1/600 verdund in TBS. Hiervoor werd
telkens 300µl antilichaam per coupe aangebracht. De coupes bleven gedurende één nacht
staan bij kamertemperatuur.
De dag nadien werden de coupes 3 maal 5 minuten gespoeld in TBS en gedurende 10 minuten
geïncubeerd met een secundair antilichaam, Biotinylated link Universal (LSAB+ system-HRP)
(Dako Cytomation, VSA), gevolgd door 10 minuten Streptavidin-HRP (LSAB+ system-HRP)
(Dako Cytomation, VSA). Op elke coupe werd gedurende 1 minuut DAB (Dako Cytomation,
VSA) aangebracht. De reactie werd gestopt door de coupes 5 minuten in AD te brengen.

25
Hierna volgde een tegenkleuring met Mayer’s haematoxyline (Sigma, Belgïe) gedurende 3
minuten, waarna de coupes 10 minuten onder stromend kraantjeswater geplaatst werden.
Dehydratatie van de coupes gebeurde in AD gevolgd door een opeenvolgende reeks van
baden met stijgende concentraties ethanol (50 %, 70 %, 96 %, 99 %; telkens 2 maal 5 minuten)
(Klinipath, België) en xyleen (3 maal 5 minuten) (Klinipath, België). Ten slotte werden de
coupes gemonteerd met Neutral Mounting Medium (D.P.X) (Klinipath, België) en een
dekglaasje (VWR International, België).
2.6. Histopathologie
Alle histologische kleuringen (H&E en Sirius Red) werden bekeken om na te gaan of er AIH
ontstaan was. Immunohistochemische kleuringen werden gescoord aan de hand van de
intensiteit van de DAB aankleuring. Dit gebeurde met behulp van het softwaresysteem Cell B.
Alle coupes werden gescoord met behulp van een lichtmicroscoop (Olympus BX41, België).
Per coupe werden vijf regions of interest (ROI) geselecteerd.
2.7. Kwantitatieve Real Time PCR (qPCR)
Het RNA uit de levers werd geanalyseerd om na te gaan of er een afwijkende expressie was
van bepaalde genen tussen de CYP2D6 groep en de controle groep. Hiervoor werd RNA uit
20 à 30 mg lever geëxtraheerd volgens het bijgevoegde protocol van de kit (Qiagen, Venlo,
The Netherlands). Het bekomen RNA werd verdund naar een concentratie van 100 ng/µl.
Vervolgens werd er cDNA gemaakt vertrekkende van 1 µg RNA met behulp van de ‘iscript
cDNA Synthesis Kit’ en oligo(dT) primers, volgens het bijgevoegde protocol (Bio-Rad,
Nazareth, Belgium). Expressie van mHprt, mHmbs, mSdha, mGapdh, mTnf, mPhd1, mPhd2,
mPhd3, mGlut1, mVegfa, mPfk, mHif1α en mHif2α werden bepaald. Alle reacties werden in
duplo gepipeteerd. Per reactie werden 4µl Sensimix Cybergreen no Rox (Bioline,
Kampenhout, Belgium) en 1µl forward + reverse primer gebruikt. Vervolgens werd er aan
elke reactie 3µl cDNA toegevoegd. De gebruikte genen en primers kunnen terug gevonden
worden in Tabel 4.

26
Tabel 4: Sequenties van sense en antisense primers gebruikt voor de bepaling van mRNA expressie van
Hydroxymethylbilane synthase (mHmbs), hypoxanthine guanine phosphoribosyl transferase (mHprt),
glyceraldehyde 3-phosphate dehydrogenase (mGapdh), succinate dehydrogenase (mSdha), Tumor necrosis factor
(mTnf), Prolyl hydroxylase 1 (mPhd1), Prolyl hydroxylase 2 (mPhd2), Prolyl hydroxylase 3 (mPhd3), Glucose
transporter 1 (mGlut1), Vasculaire endothelial growth factor A (mVegfa), Phospofructokinase (mPfk), Hypoxia-
inducible factor 1a (mHif1a) en Hypoxia-inducible factor 2a (mHif2a).
Forward primer (5’ 3’) Reverse primer (5’ 3’)
mHmbs AAGGGCTTTTCTGAGGCACC AGTTGCCCATCTTTCATCACTG
mHprt GTTAAGCAGTACAGCCCCAAA AGGGCATATCCAACAACAAAC TT
mGapdh CATGGCCTTCCGTGTTCCTA GCGGCACGTCAGATCCA
mSdha CTTGAATGAGGCTGACTGTG ATCACATAAGCTGGTCCTGT
mTnf CATCTTCTCAAAATTCGAGTGACAA TGGGAGTAGACAAGGTACAACCC
mPhd1 GAAGTGGGGCGGGCGTCTTC TGGTCCCCACGAATGCTGCG
mPhd2 AACGTAAAGGACGGGCGGGC AGGTCGGTCGAAGCGCGAGA
mPhd3 CAGACCGCAGGAATCCACAT TTCAGCATCGAAGTACCAGACAGT
mGlut1 GCT TAT GGG CTT CTC CAA ACT GT GAC ACC TCT CCC ACA TAC
mVegfa ACTCGGATGCCGACACGGGA CCTGGCCTTGCTTGCTCCCC
mPfk GCCGGCTCAGTGAGACAAG TGGCACCTTCAGCAACAATG
mHif1a ACT CCT AAC TTT TCC CAG CCT TGA CCA TAT CGC TAT CCA CAT CA
mHif2a GTGACATGATCTTTCTGTCGGAA CGCAAGGATGAGTGAAGTCAAA
2.8. Statistiek
De data werden beschreven als het gemiddelde ± de standaard deviatie. Om de significante
verschillen tussen de controlemuizen en de AIH geïnduceerde muizen na te gaan, werd er in
SPSS (Statistical Package for the Social Sciences, SPSS inc, VSA) ofwel een ongepaarde t-
test voor normaal verdeelde waarden, ofwel een Mann Whitney U test voor niet normaal
verdeelde waarden uitgevoerd. Een p-waarde < 0,05 werd beschouwd als een statistisch
significant verschil.

27
3. Resultaten
3.1. Inductie van auto-reactieve T-cellen
De analyse van de splenocyten met FACS, om na te gaan of er auto-reactieve T-cellen
aanwezig waren, gaf geen interpreteerbare resultaten. Naargelang er auto-reactieve T-cellen
aanwezig waren of niet, zouden er 1 of 2 pieken moeten te zien zijn. Wanneer er geen auto-
reactieve T-cellen waren, zouden er dus 2 pieken te zien zijn. De ene piek van de splenocyten
zonder peptiden geladen met een lage concentratie aan CFSE en de andere van de splenocyten
met peptiden geladen met een hoge concentratie aan CFSE. Wanneer er wel auto-reactieve T-
cellen aanwezig zouden zijn, zouden de splenocyten met peptiden en een hoge concentratie
aan CFSE vernietigd worden door de auto-reactieve T-cellen, zodat er maar 1 piek zou te zien
zijn.
Bij de negatieve controle met tNGFR, zouden er normaal 2 pieken te zien moeten zijn. Maar
er werden geen 2 pieken gezien, zelfs niet 1 piek. Bij de positieve controle met Ovalbumine
werd verwacht 1 piek te zien, maar deze werd niet gezien. Er waren ook geen 2 pieken te zien.
Bij de CYP2D6 groep tenslotte werd er ook geen enkele duidelijke piek gezien. Deze
resultaten waren dus niet interpreteerbaar met het gevolg dat het onmogelijk is te besluiten of
er auto-reactieve T-cellen aanwezig waren.
3.2. Validatie van het muismodel na 3 maanden
3.2.1. Algemene karakteristieken
Op het eerste zicht werd er uitwendig geen verschil tussen de CYP2D6 groep en de controle
groep vastgesteld. Behalve dat er bij 4 van de 5 CYP2D6 muizen haarverlies op de rug was,
terwijl dit bij geen enkele van de controle muizen het geval was. Bij de ene CYP2D6 muis
zonder haarverlies op de rug, was er wel wat haarverlies op de buik. Het lichaamsgewicht van
de CYP2D6 muizen was iets minder dan de muizen uit de controle groep, maar dit verschil
was niet significant (zie tabel 5). Het relatief gewicht van de lever was in de beide groepen
nagenoeg gelijk en dus ook niet significant verschillend. Het relatief gewicht van de milt bij
de CYP2D6 groep was hoger dan bij de controle groep, maar eveneens niet significant.
Uit de serumanalyse bleek ALT verhoogd te zijn bij de controle groep ten opzichte van de
CYP2D6 groep, maar dit verschil was niet significant. AST daarentegen was lichtjes
verhoogd bij de CYP2D6 groep, maar dit verschil was eveneens niet significant.
Uit de analyse van de antilichamen, bleken er geen antilichamen tegen CYP2D6, anti-LKM1,
aanwezig zijn. Dit zowel bij de CYP2D6 groep als bij de controle groep. pANCA werd wel
bij beide groepen gedetecteerd, net zoals AMA in de glomerulen. ANA was bij 1 van de 5

28
CYP2D6 muizen en 2 van de 6 controle muizen duidelijk afwezig. In de andere gevallen was
dit waarschijnlijk ook negatief, maar toch werd er een lichte aankleuring gezien, waaruit niet
duidelijk te besluiten valt of het antilichaam aanwezig was in het serum of niet.
Tabel 5: gemiddelde ± standaarddeviatie van algemene karakteristieken bij de CYP2D6 muizen en de controlemuizen en de p-waarden
CYP2D6 Controle p-waarde
Lichaamsgewicht (g) 22,0 ± 0,15 23,5 ± 0,67 0,074
Relatief gewicht lever 0,052 ± 0,002 0,051 ± 0,001 0,925
Relatief gewicht milt 0,0058 ± 0,0005 0,0051 ± 0,0003 0,26
ALT (U/l) 44,25 ± 10,10 50,25 ± 10,21 0,666
AST (U/l) 61,63 ± 9,73 60,17 ± 26,97 0,923
Anti-LKM1 Negatief Negatief -
3.2.2. Lever histologie
Er werd een H&E kleuring op de levers uitgevoerd om te kijken of er inflammatie aanwezig
was en een Sirius Red kleuring om te kijken of er fibrose vorming was.
In figuur 5A. en 5B. is de H&E kleuring op respectievelijk de lever van een controle muis en
een CYP2D6 muis te zien. Op beide foto’s is de lever te zien met duidelijke portale triades: de
vena porta, het galkanaaltje en de minder goed zichtbare arterie. De kernen van de
hepatocyten zijn paars aangekleurd en het cytoplasma roze. Rondom de portale triades is er
geen inflammatie te zien en ook verder in het leverparenchym is geen inflammatie te
bemerken. Ook op de andere coupes van de levers van de andere muizen werd hetzelfde beeld
gezien. Nergens was er inflammatie te zien, en dus ook geen interface hepatitis.
In figuur 5C. is een Sirius Red kleuring op de lever van een controle muis te zien en in figuur
5D. één van een CYP2D6 muis. Het cytoplasma is lichtrood aangekleurd en het donkerder
rood kleurt het collageen aan. Rondom de vena porta en het galkanaaltje is op beide foto’s
duidelijk meer aankleuring en bevindt er zich dus collageen. Verder is er op deze kleuringen
soms een meer donkerrode aankleuring in het leverparenchym, maar dit is heel beperkt en
gelijk in beide groepen muizen. Bijgevolg werden deze coupes niet gescoord. Er was dus in
de lever van beide groepen bijna geen collageen aanwezig, en dus geen fibrosevorming.

29
Figuur 5 – H&E en Sirius Red kleuring – A. H&E kleuring lever controle muis. B. H&E kleuring CYP2D6muis. C. Sirius Red kleuring lever controle muis. D. Sirius Red kleuring lever CYP2D6 muis. (Schaalbalk =100µm)
3.2.3. Analyse van inflammatoire merkers
Om een idee van de aanwezige inflammatie te krijgen werd de genexpressie van Tnf op
mRNA niveau in de lever bekeken door middel van qPCR. mTnf stijgt wanneer er
inflammatie aanwezig is. Uit de resultaten van de qPCR bleek dat de expressie van mTnf
hoger lag bij de controle muizen dan bij de CYP2D6 muizen. Dit verschil was echter niet
significant (p-waarde = 0,295) (zie figuur 6).

30
Figuur 6: Gemiddelde relatieve expressie van mTnf bij de controle muizen en de CYP2D6 muizen (p-waarde =0,295)
Om een beeld van de inflammatie in de lever te krijgen werd F4/80 als inflammatoire merker
gebruikt. Deze bindt aan de macrofagen en kleurt met DAB bruin, zoals in Figuur 7 te zien is.
De kernen zijn paars gekleurd door het haematoxyline en het cytoplasma is lichter paars
gekleurd. Verder zijn de portale triades op de coupes te zien met de vena porta, het
galkanaaltje en de arteria hepatica. Figuur 7A. is een levercoupe van een controle muis en
figuur 7B. is er één van een CYP2D6 muis. Op beide foto’s is een vergelijkbaar aantal
macrofagen te zien, regelmatig verspreid in de lever, zonder plaatsen met hoge concentraties
aan macrofagen. Op de levercoupes van de overige muizen werd een gelijkaardig beeld
gezien.

31
Figuur 7 – F4/80 kleuring – A. F4/80 kleuring lever controle muis. B. F4/80 kleuring CYP2D6 muis.
(Schaalbalk = 100µm)
Per coupe werden er 5 ROI geselecteerd om zo de coupes te scoren aan de hand van de
intensiteit van de DAB aankleuring. Uit deze scoring bleek dat de gemiddelde aankleuring
van DAB, en dus F4/80, hoger was bij de controle groep dan bij de CYP2D6 groep. Dit
verschil was significant (p-waarde = 0,047) (zie figuur 8).
Figuur 8: Gemiddelde F4/80 aankleuring bij de controle muizen en de CYP2D6 muizen (p-waarde = 0,047)
32
3.2.4. Analyse van hypoxische merkers
3.2.4.1.Analyse van Hifs en Pdhs
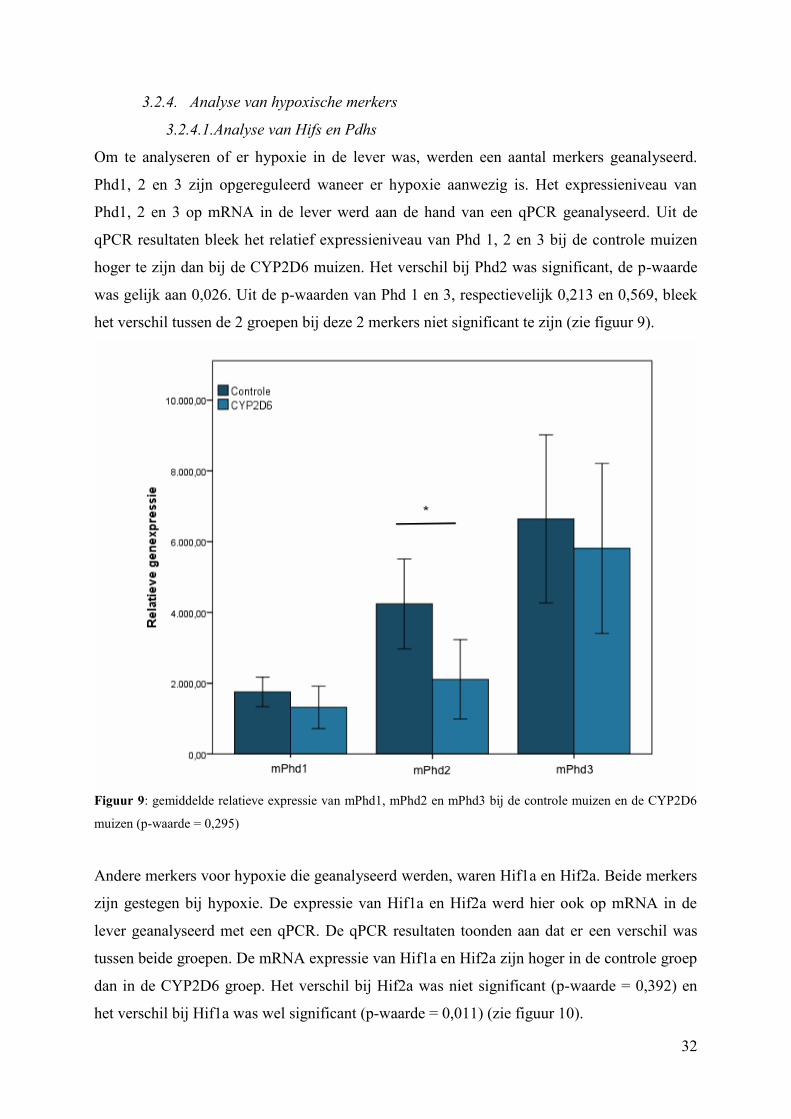
Om te analyseren of er hypoxie in de lever was, werden een aantal merkers geanalyseerd.
Phd1, 2 en 3 zijn opgereguleerd waneer er hypoxie aanwezig is. Het expressieniveau van
Phd1, 2 en 3 op mRNA in de lever werd aan de hand van een qPCR geanalyseerd. Uit de
qPCR resultaten bleek het relatief expressieniveau van Phd 1, 2 en 3 bij de controle muizen
hoger te zijn dan bij de CYP2D6 muizen. Het verschil bij Phd2 was significant, de p-waarde
was gelijk aan 0,026. Uit de p-waarden van Phd 1 en 3, respectievelijk 0,213 en 0,569, bleek
het verschil tussen de 2 groepen bij deze 2 merkers niet significant te zijn (zie figuur 9).
Figuur 9: gemiddelde relatieve expressie van mPhd1, mPhd2 en mPhd3 bij de controle muizen en de CYP2D6
muizen (p-waarde = 0,295)
Andere merkers voor hypoxie die geanalyseerd werden, waren Hif1a en Hif2a. Beide merkers
zijn gestegen bij hypoxie. De expressie van Hif1a en Hif2a werd hier ook op mRNA in de
lever geanalyseerd met een qPCR. De qPCR resultaten toonden aan dat er een verschil was
tussen beide groepen. De mRNA expressie van Hif1a en Hif2a zijn hoger in de controle groep
dan in de CYP2D6 groep. Het verschil bij Hif2a was niet significant (p-waarde = 0,392) en
het verschil bij Hif1a was wel significant (p-waarde = 0,011) (zie figuur 10).

33
Figuur 10: Gemiddelde relatieve expressie van mHif1a en mHif2a bij de controle muizen en de CYP2D6
muizen (p-waarde mHifa = 0,011 en p-waarde mHif2a = 0,392)
Hif1a werd ook gevisualiseerd op de levercoupes. Met DAB kleurden de cellen met hypoxie
bruin aan. Dit is te zien op de levercoupes bij een controle muis in figuur 11A. en bij een
CYP2D6 muis in figuur 11B. De kernen van de hepatocyten zijn paars aangekleurd door de
haematoxyline en het cytoplasma is lichter paars gekleurd. Hier en daar is er een hepatocyt
met bruin cytoplasma te zien, dit zijn hypoxische cellen. Verder is op de coupes een portale
triade te zien met duidelijke een vena porta, galkanaaltje en arteria hepatica.
De aankleuring van Hif1a is redelijk beperkt, ook op al de andere levercoupes van de
CYP2D6 en de controle muizen. Daarom werd besloten om geen scoring voor de Hif1a
kleuring uit te voeren.

34
Figuur 11 – HIF1a kleuring – A. HIF1a kleuring lever controle muis. B. HIF1a kleuring CYP2D6 muis.
(Schaalbalk = 100µm)
3.2.4.2.Analyse van Glut1, Pfk en Vegfa
De laatste hypoxische merkers die geanalyseerd werden, waren Glut1, Pfk en Vegfa. Al deze
merkers komen meer tot expressie bij hypoxie. De expressie ervan werd ook op mRNA-
niveau met een qPCR geanalyseerd. Uit de resultaten bleken zowel mGlut1, mPfk als mVegfa
hoger te zijn bij de controle muizen dan bij de CYP2D6 muizen. Al deze verschillen waren
significant, de p-waarden van Glut1, mPfk en mVegfa waren respectievelijk 0,015; 0,029 en
0,006 (zie figuur 12).

35
Figuur 12: Gemiddelde relatieve expressie van mGlut1, mPfk en mVegfa bij de controle muizen en de CYP2D6
muizen (p-waarde mGlut1 = 0,015, p-waarde mPfk = 0,029 en p-waarde mVegfa = 0,006)
De expressie van Glut1 op eiwitniveau in de lever werd met een kleuring gevisualiseerd.
Glut1 komt aan de plasmamembraan van de hepatocyten tot expressie en zal met behulp van
DAB bruin aankleuren. In figuur 13A. is de kleuring op de lever van een controle muis te zien
en in figuur 13B. op de lever van een CYP2D6 muis. Centraal op de foto’s is weer de portale
triade te zien met de duidelijke zichtbare vena porta, het galkanaaltje en de arteria hepatica.
De kernen van de hepatocyten zijn blauw-paars aangekleurd door de haematoxyline. Het
cytoplasma is lichter paars gekleurd, maar daarnaast ook bruin door DAB. De
plasmamembraan is iets donkerder bruin gekleurd, maar dit verschil is moeilijk te zien omdat
de DAB kleuring niet op tijd werd gestopt zodat alles sterker aangekleurd is. Op de andere
coupes is een gelijkaardig beeld te zien.

36
Figuur 13 – Glut1kleuring – A. Glut1 kleuring lever controle muis. B. Glut1 CYP2D6 muis. (Schaalbalk =
100µm)
Per coupe werden er 5 ROI geselecteerd om zo aan de hand van de DAB aankleuring de
coupes te scoren. Uit deze scoring bleek dat er een significant verschil was in aankleuring
tussen de controle groep en de CYP2D6 groep. De CYP2D6 groep had gemiddeld een hogere
aankleuring van Glut1 vergeleken met de controle groep (p-waarde = 0,047) (zie figuur 14).
Dit in tegenstelling tot de expressie van Glut1 op mRNA-niveau waarbij de controle groep
een significant hogere expressie van mGlut1 had dan de CYP2D6 groep.
Figuur 14: Gemiddelde GLUT1 aankleuring bij de controle muizen en de CYP2D6 muizen (p-waarde = 0,047)

37
4. Discussie
Het doel van deze masterproef was te onderzoeken of het intranodaal injecteren van CYP2D6
mRNA in C57BL/6 muizen AIH kan induceren en of er een beeld van AIH ontstaat dat
representatief is voor AIH bij de mens. AIH is een zeldzame chronische inflammatoire
leverziekte, die toe tot nu toe geen optimale behandeling heeft (8-10). Door een representatief
muismodel voor AIH te ontwikkelen kan de pathofysiologie van AIH verder ontrafeld worden
en kan er een geschikte therapie ontwikkeld worden.
Het muismodel werd op de aanwezigheid van autoantilichamen geanalyseerd. Deze analyse
wees niet op de ontwikkeling van AIH. Er was geen anti-LKM-1 in het serum aanwezig.
Nochtans had dit bijna zeker moeten aanwezig zijn indien de strategie voor de ontwikkeling
van AIH voldeed. De muizen werden immers geïmmuniseerd met CYP2D6 mRNA, met de
bedoeling dat hiertegen auto-immuniteit ontstond en dus vorming van antilichamen tegen
CYP2D6 of Anti-LKM-1 (16, 17).
De andere geanalyseerde antilichamen: ANA, pANCA en AMA, kunnen de afwezigheid van
AIH noch bevestigingen noch ontkennen. De aanwezigheid van ANA zou op de AIH type 1
kunnen wijzen. Dit antilichaam was zeker afwezig bij 1 van de 5 CYP2D6 muizen. Bij de 4
andere muizen is er twijfel door een lichte aankleuring door de indirecte immunofluorescentie.
Maar aangezien deze lichte aankleuring ook bij 4 van 6 controle muizen gezien werd, wees dit
eerder in de richting van de afwezigheid van ANF en AIH (16, 17).
Dezelfde twijfel ontstond bij de analyse van AMA en pANCA. Hoewel AMA vooral
voorkomt bij primaire biliaire cirrose, kan het ook bij AIH voorkomen. pANCA komt bij AIH
voor, maar ook bij andere auto-immuun ziekten (16, 17). Zowel bij de CYP2D6 muizen als
bij de controle muizen was er twijfel of AMA en pANCA aanwezig waren. Aangezien de
resultaten van beide groepen gelijkaardig waren, is het aannemelijk om te besluiten dat de
antilichamen niet aanwezig waren. Indien er antilichamen die op AIH kunnen wijzen
aanwezig waren, dan zou er vermoedelijk een verschil tussen de CYP2D6 groep en de
controle groep geweest zijn, maar dit werd niet gezien.
De analyse van de transaminasen waarden wees ook op de afwezigheid van AIH. De waarden
van ALT en AST zouden bij AIH verhoogd moeten zijn en dit was niet het geval (9).
Verhoogde transaminasen waarden wijzen op leverschade. De ALT-waarden in de controle
groep waren zelfs hoger dan die bij de CYP2D6 groep, maar dit was wel niet significant.

38
Interface hepatitis is ook een typisch kenmerk van AIH (9) en dit werd ook niet gezien op de
levercoupes. De lever van de CYP2D6 muizen zag er gezond uit, zonder inflammatie, wat dus
eveneens wijst op de afwezigheid van AIH.
De afwezigheid van inflammatie werd bevestigd door de resultaten van de qPCR op mTnf in
de lever. Het expressie-niveau van mTnf was lager bij de CYP2D6 muizen dan bij de controle
muizen. Het verschil was wel niet significant. Het verschil tussen de F4/80 was echter wel
significant, met meer aankleuring bij de controle groep. Dit wijst ook op de afwezigheid van
AIH bij de CYP2D6 muizen, maar tegen alle verwachtingen in heeft de controle groep
significant meer inflammatie. Wanneer er inflammatie in de lever aanwezig is, zullen de
transaminasen waarden ook gestegen zijn. Dit verband kan ook gezien worden in deze
resultaten. Er was meer inflammatie aanwezig bij de controle groep en er was een niet-
significante, maar toch een duidelijke stijging in ALT-waarden. Bij de AST-waarden is deze
stijging echter niet zichtbaar.
Dus, tot zo ver lijkt niets in de CYP2D6 groep in de richting van AIH te wijzen. Er was wel
een niet-significant verminderd lichaamsgewicht bij de CYP2D6 muizen, wat op een ziekte
zoals AIH zou kunnen wijzen. Maar dit komt niet overeen met de overige geanalyseerde
parameters.
Er was echter wel één opvallend verschil tussen de CYP2D6 groep en de controle groep, het
haarverlies op de rug. Dit werd enkel bij de CYP2D6 groep vastgesteld. Uit de literatuur blijkt
er geen vastgesteld verband te zijn tussen haarverlies en AIH, met uitzondering van een
casestudie bij een kind en een konijn, waar dit verband wel werd naar voorgeschoven (36, 37).
Een andere studie onderzocht ook dit verband maar wijt dit aan een zinktekort en niet aan
AIH (38). Ook op eerder ontwikkelde muismodellen voor AIH werd er nergens iets
beschreven over het feit dat de muizen haarverlies op de rug krijgen. Dit wordt ook niet
vermeld bij AIH-muismodellen bij C57BL/6 muizen (34).
De meeste aannemelijke oorzaak van het haarverlies is ‘barbering’. Barbering is een gedrag
van muizen waarbij snorharen en haren van andere muizen en van zichzelf worden geplukt.
Dit zou een dominant gedrag zijn en wijzen op de sociale hiërarchie. Dit wordt vaak gezien
bij laboratoriummuizen. Naar gelang de genetische achtergrond van de muizen, komt
barbering vaker voor of niet. Het is beschreven dat barbering bij C57BL/6 muizen vaak
voorkomt. Er wordt vooral gezien dat er haarverlies ter hoogte van de snuit, nek en hoofd
voorkomt ten gevolge van barbering (39-41). Ondanks de genetische voorgeschiktheid voor
barbering bij C57BL6 muizen, doen niet alle muizen dit. Bij de controle groep zal er
waarschijnlijk geen enkele ‘barber’ muis aanwezig geweest zijn en bij de CYP2D6 groep wel.

39
Er zijn echter wel 2 argumenten tegen het feit dat het haarverlies op de rug te wijten zou zijn
aan barbering. Bij de CYP2D6 muizen was er enkel een kalende plek op de rug, en niet op de
typische plaatsen van barbering. Bovendien hadden 2 controle muizen in dezelfde kooi als de
CYP2D6 muizen geen haarverlies, terwijl bij barbering vaak alle muizen in een kooi door één
muis aangetast zijn. Het haarverlies op de rug kan dus ook veroorzaakt zijn door een tot nu
toe nog onbekende, eventueel auto-immune, oorzaak.
De levers van de muizen werd ook geanalyseerd op fibrosevorming, maar noch bij de
CYP2D6 muizen als bij de controle muizen werd op de Sirius Red kleuring fibrose gezien.
Dit beeld klopt met de vorige besproken parameters. Wanneer er geen inflammatie in de lever
is, en geen verhoogde transaminasen-waarden en geen autoantilichamen aanwezig zijn, is het
logisch dat er geen evolutie naar fibrose in de lever is. Alhoewel er bij de controle groep wel
inflammatie aanwezig was, werd ook hier geen fibrosevorming waargenomen.
Tot hier toe leek er helemaal geen ontwikkeling van AIH te zijn. Toch werd er verder
onderzocht of er mogelijk geen hypoxie in de lever was. Leverinflammatie kan namelijk
zorgen voor de ontwikkeling van hypoxie en fibrose (25).
Bij de evaluatie van hypoxie in de lever werden er tussen beide groepen significant
verschillen gezien. De expressie van Hif1a, Phd2, Glut1, Vegfa en Pfk op mRNA-niveau
waren hoger in de controle groep.
De significante verschillen gezien op mRNA hebben een logisch verband met elkaar.
Wanneer er hypoxie aanwezig is, zullen Hifs opgereguleerd zijn. Dit is zowel bij Hif1a en
Hif2a te zien, maar enkel Hif1a was significant. Bij hypoxie zijn ook de Phd’s gestegen en dit
werd ook gezien, maar enkel Phd2 was significant gestegen. Hieruit volgt dat de genen die
gereguleerd worden door Hifs ook opgereguleerd moeten zijn, en dit was ook het geval bij de
3 geanalyseerde genen: Glut1, Vegfa en Pfk (26, 27).
De analyse van de Glut1-expressie op eiwit-niveau door de immunohistochemische kleuring
was echter groter in de CYP2D6 groep. Dit lijkt onlogisch, want op mRNA-niveau werd het
omgekeerde gezien. Vermoedelijk was de expressie van Glut1 ook verhoogd in de controle
groep en is het omgekeerde resultaat te wijten aan een te donkergekleurde Glut1 kleuring.
Niet enkel Glut1 werd door DAB donkerbruin aangekleurd, maar ook het cytoplasma. Wat het
vergelijken van de Glut1 aankleuring moeilijk maakte.

40
Een aanpassing van het protocol van de immunohistochemische kleuring van Glut1 zou dan
ook een goed idee zijn. Vermoedelijk is het verkorten van de incubatie met DAB, die zorgt
voor de bruinverkleuring, al voldoende. De coupes zouden maximum 10 seconden mogen
incuberen en de incubatie moet onmiddellijk gestopt worden met gedestilleerd water wanneer
de bruinkleuring zichtbaar begint te worden (42).
Uit de evaluatie van de hierboven vermelde analysen, blijkt dus duidelijk dat de CYP2D6
muizen op 3 maanden geen ontwikkeling van AIH hebben. Er schort dus iets aan het protocol
van dit experiment.
De keuze van het muismodel was gebaseerd op de literatuur. De genetische achtergrond, met
de aan- of afwezigheid van bepaalde MHC genen, speelt duidelijk een rol bij het ontwikkelen
van AIH en de genetische achtergrond van het muismodel is dus ook belangrijk. De keuze
werd vooral gebaseerd op een artikel waarin het ontwikkelen van AIH bij C57BL/6, 129/Sv
en BALB/c muizen werd onderzocht. Daaruit bleek dat C57BL/6 muizen het meest geschikt
waren voor het ontwikkelen van AIH (43). De keuze van het muismodel zal dus
waarschijnlijk niet gezorgd hebben voor de negatieve resultaten in de experimenten hier. Het
zal waarschijnlijk aan de manier van inductie liggen.
Wat er mogelijkerwijze is misgegaan bij de inductie en wat hieraan zou kunnen verbeterd
worden is echter moeilijk te bepalen. Indien de CTL wel zou gewerkt hebben zou dit wel
duidelijk geweest zijn. Als de CTL had aangetoond dat er geen cytotoxische T-cellen
aanwezig waren, dan wees dit uit dat de intranodale injectie met CYP2D6 mRNA geen goede
manier was om AIH te induceren.
De eventuele afwezigheid van cytotoxische T-cellen zou ook vals negatief kunnen geweest
zijn, aangezien de aanwezigheid van deze cellen werd getest met 3 peptiden van CYP2D6. Er
werd gekeken of deze peptiden door de eventueel gevormde cytotoxische T-cellen vernietigd
werden. Deze peptiden werden gekozen op basis van predictieprogramma’s (Syfpeithi,
Rankpep en Immune epitope) die bekeken welke peptiden van het humaan CYP2D6 het beste
binden met de MHC I van muizen (44). De MHC I genen verschillen immers van die van de
mens. De scores van peptiden die de beste binding hadden, verschilden tussen de
verschillende predictieprogramma’s. De 3 peptiden die het meest voorkwamen en gemiddeld
de beste score hadden werden gekozen en gebruikt voor de immunisatie. Deze peptiden
kwamen niet overeen met peptiden die als epitopen van CYP2D6 bij de mens beschreven zijn,
terwijl er in de literatuur studies beschreven zijn die er in slaagden om AIH te induceren aan
de hand van CYP2D6 epitopen van de mens (45).

41
Daarenboven bleken de predictieprogramma’s niet zo betrouwbaar (44) en zou in de toekomst,
bij het herhalen van de CTL van dit experiment, rekening moeten gehouden worden dat een
eventueel negatief resultaat kan gelegen zijn aan de peptiden die gebruikt worden om dit te
controleren.
Een eventueel punt van verbetering van dit experiment zou de deletie van Tregs kunnen zijn.
In de literatuur is beschreven dat in AIH er minder Tregs aanwezig zijn en hoe minder er
aanwezig zijn, hoe ernstiger de ziekte. De Tregs controleren het immuunsysteem; indien de
Tregs dus disfunctioneel zouden worden, zou dit misschien de inductie van AIH
vergemakkelijken (9, 13, 46).
Tenslotte blijft nog de vraag waarom de controle muizen meer inflammatie en hypoxie in de
lever vertoonden dan de CYP2D6 muizen. De F4/80 kleuring toonde significant meer
inflammatie bij de controle muizen aan en de analyseerde genen op mRNA-niveau toonden
hypoxie aan. Het lijkt eerder dat de controle muizen AIH hebben ontwikkeld dan de CYP2D6
muizen, wat onlogisch overkomt.
Een eerst mogelijke verklaring hiervoor zou kunnen zijn dat de significante verschillen die
zijn aangetoond er niet echt zijn. Bij de uitgevoerde t-testen wordt vertrokken van een
nulhypothese, dat de gemiddelden van de gemeten parameters gelijk zijn aan elkaar. Indien dit
niet zo is, wordt de nulhypothese verworpen. Er is echter een kans dat de nulhypothese
verworpen wordt, ook al is ze eigenlijk waar is. Dit is de type I fout, de kans dat deze fout
voorkomt is gelijk aan het significantieniveau, hier 5%. Door het significantieniveau te
verlagen zou deze type I fout minder voorkomen. Dan komt echter wel de type II fout meer
voor, dat is wanneer de nulhypothese niet verworpen wordt als deze niet waar was. Om de
kans op een type II fout te verminderen, en dus bij gevolg ook de type I fout, zouden de
groepen van het experiment vergroot moeten worden (47). Om dus te controleren dat de
resultaten inderdaad aan de type I fouten te wijten zijn, zou het experiment herhaald moeten
worden met grotere groepen. Er is echter wel een argument dat deze verklaring in twijfel trekt.
Indien al deze significante resultaten immers aan type I fouten te wijten zouden zijn, zou er
ook verwacht worden dat de significante verschillen willekeurig zouden zijn. Maar alle
significante resultaten, op de Glut1 kleuring na, hebben een logisch verband met elkaar. Al de
resultaten wijzen in een de richting van inflammatie en hypoxie in de lever en zelfs als naar de
trend van de niet significante resultaten wordt gekeken, is hetzelfde logisch verband aanwezig.

42
Een andere mogelijke verklaring voor het feit dat de controle muizen meer inflammatie en
hypoxie vertoonden dan de CYP2D6 muizen is het Mouse hepatitis virus (MHV). Gedurende
het experiment verbleven alle muizen in het animalarium van de Vrije Universiteit Brussel
(VUB), waar vermoedelijk het Mouse hepatitis virus aanwezig was. Het MHV is een ssRNA
virus van de Coronaviridae dat vaak bij laboratoriummuizen voorkomt. Dit virus kan
verschillende pathologieën bij muizen veroorzaken, waaronder hepatitis. Volwassen muizen,
zoals in dit experiment, zijn asymptomatisch. Maar toch kan door deze infectie milde enteritis
en colitis optreden, en daarnaast zijn er pathologische veranderingen in de hersenen en in de
lever (48, 49).
De Mouse hepatitis infectie zou dus verantwoordelijk kunnen zijn voor het beeld van de lever
in de controle muizen. Maar wat er aan deze redenering niet klopt is dat de CYP2D6 muizen
niet hetzelfde beeld in de lever hebben. Alle muizen waren in de VUB aan dezelfde
omstandigheden blootgesteld, bijgevolg dus ook aan de Mouse hepatitis infectie. Bijgevolg
zouden de analysen van de levers van de CYP2D6 muizen gelijkaardig moeten zijn aan die
van de controle muizen, maar dit is hier niet het geval. Alle omstandigheden waren dezelfde
op één iets na, de CYP2D6 muizen werden met CYP2D6 mRNA geïnjecteerd. Dit wijst in de
richting van de volgende mogelijke theorie: door de immunisatie met CYP2D6 mRNA zijn de
muizen misschien geïmmuniseerd tegen MHV.
Deze theorie is gesteund op een artikel waarin werd aangetoond dat C57BL/6 muizen
geïnfecteerd met het Mouse hepatitis virus A59 een auto-immune hepatitis-like ziekte
ontwikkelden (50). In dit artikel bleken de muizen, geïnfecteerd met het MHV, verschillende
symptomen van AIH te vertonen. Er waren verhoogde waarden van Ig en van transaminasen
aanwezig, infiltraten van lymfocyten en neutrofielen in de lever en autoantilichamen. Er
werden autoantilichamen tegen fumarylacetoacetate hydrolase gedetecteerd. Dit is een
oplosbaar enzyme aanwezig in het cytosol van de lever en de nieren, en het antilichaam
hiertegen wordt geïnduceerd door MHV. Daarnaast werden echter ook andere
autoantilichamen gedetecteerd tegen leverproteïnen. Tegen welke leverproteïnen er
autoantilichamen werden gevormd werd niet geanalyseerd.
Het vervolg van deze immunisatie-theorie is echter volledig gebaseerd op een
veronderstelling waarvoor geen bewijs in de literatuur werd teruggevonden. Stel dat door de
immunisatie met CYP2D6 mRNA toch autoantilichamen gevormd werden tegen een deel van
het CYP2D6 eiwit, of eventueel tegen een ander eiwit in de lever, en dat deze
autoantilichamen een kruisreactiviteit tegen de antigenen van MHV vertoonden. In het

43
hierboven vermelde artikel wordt er weliswaar het omgekeerde beschreven. MHV zal voor de
inductie van antilichamen zorgen tegen MHV en er worden autoantilichamen tegen
leverproteïnen gevormd en gedetecteerd. Daaruit zou kunnen besloten worden dat het
omgekeerde dus ook zou kunnen gebeuren. Dit is echter voorlopig speculatief, omdat het niet
zeker is of er auto-antilichamen door de immunisatie met CYP2D6 mRNA werden gevormd
en of ze kruisreactiviteit tegen MHV vertoonden. Maar stel dat het kan, zou het wel een
verklaring kunnen geven voor de hogere inflammatie en hypoxie in de controle muizen en
niet in de CYP2D6 muizen. Een manier om deze theorie te bevestigen zou het opnieuw
uitvoeren van al de analysen op controle C57BL/6 muizen zijn uit het animalarium van de
Universiteit Gent, waar er zeker geen MHV aanwezig is. Deze resultaten zouden dan met de
resultaten uit dit experiment hier moeten vergeleken worden en indien de resultaten
overeenkomen met die van de CYP2D6 groep en niet met de controle groep, dan zou deze
veronderstelling misschien kunnen kloppen.

44
5. Algemeen besluit
AIH is een zeldzame auto-immuunziekte van de lever waarbij er inflammatie ontstaat. De
ziekte is progressief en chronisch en heeft geen optimale behandeling. Daarnaast is ook nog
weinig gekend over de pathofysiologie van AIH. Daarom is er nood aan een representatief
muismodel om zo de pathofysiologie te ontrafelen en een optimale therapie te ontwikkelen.
In deze masterproef werd een muismodel voor AIH gevalideerd. De opzet van dit muismodel
was om door het intranodaal injecteren van CYP2D6 mRNA AIH te induceren.
Uit de validatie van het muismodel na 3 maanden bleek er echter geen ontwikkeling van AIH
te zijn. Er waren geen autoantilichamen tegen CYP2D6 aanwezig, ook andere
autoantilichamen waren vermoedelijk negatief. De transaminasenwaarden waren ook niet
gestegen. Er was wel duidelijk haarverlies op de rug bij de CYP2D6 muizen, dat afwezig was
bij de controle muizen. Maar dit is vrijwel zeker geen teken van de ontwikkeling van AIH.
Het haarverlies zou eventueel aan barbering liggen, maar meer waarschijnlijk ligt het aan een
nog onbekende en eventueel auto-immune oorzaak door de injectie met de CYP2D6 mRNA.
Op de leverkleuringen werd ook geen teken van AIH gezien, er werd geen interface hepatitis
op de H&E kleuringen gezien en geen fibrose vorming op de Sirius Red kleuringen.
Op de F4/80 kleuringen werd aangetoond dat de inflammatie in de CYP2D6 groep zelfs
minder was dan in de controle groep. De genexpressie van Hif1a, Phd2, Glut1, Pfk en Vegfa
op mRNA-niveau toonde aan dat de hypoxie in de lever minder was bij de CYP2D6 muizen
dan bij de controle muizen. De expressie van Glut1 op eiwit toonde echter het omgekeerde
aan, maar dit lag waarschijnlijk aan het protocol van de Glut1 kleuring. Een veel kortere
incubatie met DAB, dat voor de bruine aankleuring zorgt, zou de kleuring verbeteren en zou
de resultaten vermoedelijk doen overeenkomen met de bevindingen op mRNA-niveau.
Uit dit experiment kan dus met zekerheid worden besloten dat de poging om via dit
muismodel AIH te induceren niet gelukt is. Het merkwaardige in dit experiment is wel dat de
controle muizen meer inflammatie en hypoxie in de lever vertonen dan bij de CYP2D6
muizen. Dit zou misschien te wijten kunnen zijn aan het laag aantal gebruikte muizen in dit
experiment en een te hoog significantie-niveau, maar dit lijkt niet zo waarschijnlijk omdat de
resultaten een logisch verband met elkaar hebben. Een andere eventuele verklaring zou
kunnen zijn dat de CYP2D6 muizen door de injectie van het mRNA van CYP2D6 via
kruisreactiviteit geïmmuniseerd werden tegen MHV, dat in het animalarium van de VUB
aanwezig was. De controle muizen werden niet geïmmuniseerd en de inflammatie en hypoxie

45
in de lever zou dus misschien te wijten kunnen zijn aan een MHV-infectie. De analysen
herhalen op controle muizen zonder MHV zou hierover duidelijkheid kunnen brengen.
Dit muismodel heeft dus duidelijk nog aanpassingen nodig om een goed muismodel voor AIH
te worden. C57BL/6 muizen blijven een goede keuze wegens hun genetische
voorbeschiktheid. Of het niet ontstaan van AIH aan de manier van inductie lag, zou kunnen
opgehelderd worden door dit experiment opnieuw uit te voeren met een wel werkende CTL.
Dit zou kunnen aantonen of er nu wel of geen cytotoxische T-cellen tegen CYP2D6 aanwezig
waren. De depletie van Tregs zou mogelijkerwijze een punt van verbetering kunnen zijn.

46
Referenties1. Espeel M. Functionele Anatomie, Deel II. 2009:60.2. Marieb EN, Jon M, Wilgelm PB. Human Anatomy. Fifth ed: Pearson Benjamin Cummings; 2008.3. Feldman M, Friedman LS, Brandt LJ. Sleisenger and Fordtran's Gastrointestinal and Liver Disease. 9thed2010.4. Young B, Lowe JS, Stevens A, Heath JW. Wheater's Functional Histology. Fifth ed: Elsevier; 2006.5. Vral A, Mione S. Bijzondere weefselleer. 2de ed: Acco; 2010. 169 p.6. Geller DA, Goss JA, Tsung A. Chapter 31. Liver. Schwartz's Principles of Surgery. 9th ed: TheMcGraw-Hill Companies, Inc.; 2010.7. Barrett KE. Chapter 10. Functional Anatomy of the Liver and Biliary System. GastrointestinalPhysiology: The McGraw-Hill Companies, Inc.; 2006.8. McFarlane IG. Definition and classification of autoimmune hepatitis. Semin Liver Dis. 2002;22(4):317-24.9. Mieli-Vergani G, Heller S, Jara P, Vergani D, Chang MH, Fujisawa T, et al. Autoimmune Hepatitis. JPediatr Gastr Nutr. 2009;49(2):158-64.10. Gossard AA, Lindor KD. Autoimmune hepatitis: a review. J Gastroenterol. 2012;47(5):498-503.11. Lohse AW, Mieli-Vergani G. Autoimmune hepatitis. J Hepatol. 2011;55(1):171-82.12. Feld JJ, Heathcote EJ. Epidemiology of autoimmune liver disease. J Gastroen Hepatol.2003;18(10):1118-28.13. Longhi MS, Ma Y, Mieli-Vergani G, Vergani D. Aetiopathogenesis of autoimmune hepatitis. JAutoimmun. 2010;34(1):7-14.14. Oliveira LC, Porta G, Marin MLC, Bittencourt PL, Kalil J, Goldberg AC. Autoimmune hepatitis, HLAand extended haplotypes. Autoimmun Rev. 2011;10(4):189-93.15. Murphy KT, P.; Walport, M.;. Janeway's Immuno biology. 7th ed: Garland Science; 2008.16. Bogdanos DP, Mieli-Vergani G, Vergani D. Autoantibodies and their Antigens in AutoimmuneHepatitis. Semin Liver Dis. 2009;29(3):241-53.17. Vergani D, Longhi MS, Bogdanos DP, Ma Y, Mieli-Vergani G. Autoimmune hepatitis. SeminImmunopathol. 2009;31(3):421-35.18. Lefebvre RA. Farmacologie2011.19. Gleeson D, Heneghan MA. British Society of Gastroenterology (BSG) guidelines for management ofautoimmune hepatitis. Gut. 2011;60(12):1611-29.20. Obermayer-Straub P, Strassburg CP, Manns MP. Autoimmune hepatitis. J Hepatol. 2000;32:181-97.21. Manns MP, Czaja AJ, Gorham JD, Krawitt EL, Mieli-Vergani G, Vergani D, et al. Diagnosis andManagement of Autoimmune Hepatitis. Hepatology. 2010;51(6):2193-213.22. Tripathi D, Neuberger J. Autoimmune Hepatitis and Liver Transplantation: Indications, Results, andManagement of Recurrent Disease. Semin Liver Dis. 2009;29(3):286-96.23. Friedman SL. Liver fibrosis - from bench to bedside. J Hepatol. 2003;38:S38-S53.24. Friedman SL. Hepatic fibrosis-Overview. Toxicology. 2008;254(3):120-9.25. Rosmorduc O, Housset C. Hypoxia: A Link between Fibrogenesis, Angiogenesis, and Carcinogenesis inLiver Disease. Semin Liver Dis. 2010;30(3):258-70.26. Nath B, Szabo G. Hypoxia and hypoxia inducible factors: Diverse roles in liver diseases. Hepatology.2012;55(2):622-33.27. Zhang JZ, Behrooz A, Ismail-Beigi F. Regulation of glucose transport by hypoxia. Am J Kidney Dis.1999;34(1):189-202.28. Troeger JS, Schwabe RF. Hypoxia and hypoxia-inducible factor 1 alpha: potential links betweenangiogenesis and fibrogenesis in hepatic stellate cells. Liver Int. 2011;31(2):143-5.29. Copple BL, Bai S, Burgoon LD, Moon JO. Hypoxia-inducible factor-1 alpha regulates the expression ofgenes in hypoxic hepatic stellate cells important for collagen deposition and angiogenesis. Liver Int.2011;31(2):230-44.30. Moon JO, Welch TP, Gonzalez FJ, Copple BL. Reduced liver fibrosis in hypoxia-inducible factor-1alpha-deficient mice. Am J Physiol-Gastr L. 2009;296(3):G582-G92.31. Jaeckel E. Animal models of autoimmune hepatitis. Semin Liver Dis. 2002;22(4):325-37.32. Christen U, Hintermann E, Jaeckel E. New Animal Models for Autoimmune Hepatitis. Semin Liver Dis.2009;29(3):262-72.33. Tamaki S, Homma S, Enomoto Y, Komita H, Zeniya M, Ohno T, et al. Autoimmune hepaticinflammation by vaccination of mice with dendritic cells loaded with well-differentiated hepatocellularcarcinoma cells and administration of interleukin-12. Clin Immunol. 2005;117(3):280-93.34. Lapierre P, Djilali-Saiah I, Vitozzi S, Alvarez F. A murine model of type 2 autoimmune hepatitis:Xenoimmunization with human antigens. Hepatology. 2004;39(4):1066-74.

47
35. Holdener M, Hintermann E, Bayer M, Rhode A, Rodrigo E, Hintereder G, et al. Breaking tolerance tothe natural human liver autoantigen cytochrome P450 2D6 by virus infection. J Exp Med. 2008;205(6):1409-22.36. Chaves V, Paunier L, Berclaz R, Deleze G, Abuaf N, Belli DC. Anti-liver-kidney microsome antibody-positive autoimmune hepatitis associated with alopecia. J Pediatr Gastroenterol Nutr. 1991;12(2):288-90.37. Florizoone K, van der Luer R, van den Ingh T. Symmetrical alopecia, scaling and hepatitis in a rabbit.Vet Dermatol. 2007;18(3):161-4.38. Pereira Th C B-BMA, Hessel G. Alopecia in autoimmune hepatitis: association with zinc deficiency.Annals of Gastroentrology. 2010;23(1):71-3.39. Sarna JR, Dyck RH, Whishaw IQ. The Dalila effect: C57BL6 mice barber whiskers by plucking. BehavBrain Res. 2000;108(1):39-45.40. Garner JP, Dufour B, Gregg LE, Weisker SM, Mench JA. Social and husbandry factors affecting theprevalence and severity of barbering ('whisker trimming') by laboratory mice. Appl Anim Behav Sci. 2004;89(3-4):263-82.41. Kalueff AV, Minasyan A, Keisala T, Shah ZH, Tuohimaa P. Hair barbering in mice: Implications forneurobehavioural research. Behav Process. 2006;71(1):8-15.42. http://www.ihcworld.com.43. Lapierre P, Beland K, Djilali-Saiah I, Alvarez F. Type 2 autoimmune hepatitis murine model: theinfluence of genetic background in disease development. J Autoimmun. 2006;26(2):82-9.44. Lefere S. Validatie van een muismodel voor auto-immune hepatitis. Masterproef - Master of medicinein de geneeskunde. 2013.45. Hintermann E, Holdener M, Bayer M, Loges S, Pfeilschifter JM, Granier C, et al. Epitope spreading ofthe anti-CYP2D6 antibody response in patients with autoimmune hepatitis and in the CYP2D6 mouse model. JAutoimmun. 2011;37(3):242-53.46. Lapierre P, Beland K, Yang R, Alvarez F. Adoptive transfer of ex vivo expanded regulatory T cells inan autoimmune hepatitis murine model restores peripheral tolerance. Hepatology. 2013;57(1):217-27.47. Banerjee A, Chitnis UB, Jadhav SL, Bhawalkar JS, Chaudhury S. Hypothesis testing, type I and type IIerrors. Industrial psychiatry journal. 2009;18(2):127-31.48. Homberger FR. Enterotropic mouse hepatitis virus. Lab Anim. 1997;31(2):97-115.49. Baker DG. Natural pathogens of laboratory mice, rats, and rabbits and their effects on research. ClinMicrobiol Rev. 1998;11(2):231-+.50. Aparicio JL, Pena C, Retegui LA. Autoimmune hepatitis-like disease in C57BL/6 mice infected withmouse hepatitis virus A59. Int Immunopharmacol. 2011;11(10):1591-8.
