vegetative and reproductive innovations of early land plants
TRANSCRIPT

Vegetative and reproductive innovationsof early land plants implications for a
uni ed phylogeny
Karen Sue Renzaglia1 R Joel Duiexcl1 Daniel L Nickrent1 and David J Garbary2
1Department of Plant Biology and Center for Systematic Biology Southern Illinois University CarbondaleIL 62901- 6509 USA (renzagliaplantsiuedu)
2Department of Biology St Francis Xavier University Antigonish Nova Scotia Canada B2G 2W5
As the oldest extant lineages of land plants bryophytes provide a living laboratory in which to evaluatemorphological adaptations associated with early land existence In this paper we examine reproductive andstructural innovations in the gametophyte and sporophyte generations of hornworts liverworts mossesand basal pteridophytes Reproductive features relating to spermatogenesis and the architecture of motilemale gametes are overviewed and evaluated from an evolutionary perspective Phylogenetic analyses of adata set derived from spermatogenesis and one derived from comprehensive morphogenetic data arecompared with a molecular analysis of nuclear and mitochondrial small subunit rDNA sequences
Although relatively small because of a reliance on water for sexual reproduction gametophytes of bryo-phytes are the most elaborate of those produced by any land plant Phenotypic variability ingametophytic habit ranges from leafy to thalloid forms with the greatest diversity exhibited by hepaticsAppendages including leaves slime papillae and hairs predominate in liverworts and mosses whilehornwort gametophytes are strictly thalloid with no organized external structures Internalization ofreproductive and vegetative structures within mucilage-centlled spaces is an adaptive strategy exhibited byhornworts The formative stages of gametangial development are similar in the three bryophyte groupswith the exception that in mosses apical growth is intercalated into early organogenesis a feature echoedin moss sporophyte ontogeny
A monosporangiate unbranched sporophyte typicentes bryophytes but developmental and structuralinnovations suggest the three bryophyte groups diverged prior to elaboration of this generation Sporo-phyte morphogenesis in hornworts involves non-synchronized sporogenesis and the continued elongationof the single sporangium features unique among archegoniates In hepatics elongation of the sporophyteseta and archegoniophore is rapid and requires instantaneous wall expandability and hydrostatic supportUnicellular spiralled elaters and capsule dehiscence through the formation of four regular valves areautapomorphies of liverworts Sporophytic sophistications in the moss clade include conducting tissuestomata an assimilative layer and an elaborate peristome for extended spore dispersal Characters suchas stomata and conducting cells that are shared among sporophytes of mosses hornworts and pterido-phytes are interpreted as parallelisms and not homologies
Our phylogenetic analysis of three diiexclerent data sets is the most comprehensive to date and points to asingle phylogenetic solution for the evolution of basal embryophytes Hornworts are supported as theearliest divergent embryophyte clade with a mossliverwort clade sister to tracheophytes Among pterido-phytes lycophytes are monophyletic and an assemblage containing ferns Equisetum and psilophytes issister to seed plants Congruence between morphological and molecular hypotheses indicates that thesedata sets are tracking the same phylogenetic signal and reinforces our phylogenetic conclusions Itappears that total evidence approaches are valuable in resolving ancient radiations such as those charac-terizing the evolution of early embryophytes More information on land plant phylogeny can be found athttpwwwsciencesiuedulandplantsindexhtml
Keywords bryophytes embryophytes gametophyte land plant phylogeny morphogenesisspermatogenesis
1 INTRODUCTION
The early evolution of land plants was characterized byhigh rates of morphological and reproductive innovations
that were fuelled by stochastic genetic changes and culledby natural selection (Niklas 1997 Bateman et al 1998)Poised at the extremity of an uninhabited landscapetransitional streptophytes (green plants) experienced aburst of diversicentcation that resulted in radiation into andexploitation of a variety of new terrestrial sites Repeated
Phil Trans R Soc Lond B (2000) 355 769^793 769 copy 2000 The Royal Society
Present address Department of Biology The University of AkronAkron OH 44325-3908 USA
and simultaneous patterns of extensive morphologicaldiversicentcation followed by widespread decimationpresumably characterized these early stages of land colon-ization (Gould 1989 Kenrick amp Crane 1997a) Of themagnitude of morphological experiments that wereattempted only limited fragments have persisted throughthe millennia including the three lineages of bryophytes(Stewart amp Rothwell 1993 Taylor amp Taylor 1993)
Given the time since divergence the depauperate fossilrecord and the vastness of the newly inhabited landscapeit is not surprising that the early phylogenetic history ofland plants remains one of the major unresolved problemsin evolutionary biology Axiomatic to current evolu-tionary thought is the concept that embryophytes aremonophyletic that bryophytes represent the basal cladesand that Charophyceae especially Coleochaetales andCharales are the closest living algal relatives of arche-goniates (McCourt 1995) Beyond these accepted maximsthere is no consensus as to interrelationships among bryo-phytes and basal pteridophytes Indeed virtually everyconceivable hypothesis regarding bryophyte phylogenyhas been proposed from paraphyletic in a variety ofbranching orders to hornworts basal and mosses andliverworts monophyletic to a completely monophyleticbryophyte assemblage (Bopp amp Capesius 1996 Capesiusamp Bopp 1997 Bremer et al 1987 Garbary et al 1993Garbary amp Renzaglia 1998 Hedderson et al 1998 Lewiset al 1997 Kenrick amp Crane 1997a Malek et al 1996Mishler amp Churchill 1984 1985 Mishler et al 1994 Duiexclamp Nickrent 1999 Nishiyama amp Kato 1999) Clearly newdata or perhaps new insights on analysing existing dataare required to resolve this phylogenetic dilemma
By nature of their antiquity liverworts hornwortsmosses and basal pteridophytes represent relics of a oncemore diverse poundora and as the oldest living remnants ofearly land colonization these organisms provide a livinglaboratory in which to examine early morphologicaladaptations to existence on land In this paper weexplore morphological and developmental features of bryo-phytes and basal pteridophytes at the cell tissue organand whole organism levels Whenever possible evidence ispresented from more informative basal representatives ofthe major clades This comparative approach is designedto identify successful phenotypic innovations of both thegametophyte and sporophyte which enabled these plantsto optimize vegetative and reproductive activities in aterrestrial setting In addition to illuminating phyloge-netic acurrennities contemplation of morphological andmorphogenetic strategies provides valuable insight intothe direction and sequence of character transformation
This review is not intended to be a comprehensiveoverview of morphology in bryophytes but rather the goalis to explore answers to the following questions related toevolutionary adaptations exhibited by early plant life onland What are the vegetative and reproductive innova-tions that enabled the three bryophyte lineages to persist How do these organisms cope with the constraints ofterrestrial existence especially desiccation and gravityAnd centnally what can be gleaned from careful scrutiny ofextant basal embryophytes about morphological char-acter evolution including divergences convergencesparallelisms and homologies These questions provide thecontext for comprehensive phylogenetic analyses in which
we review and update two previously published morpho-logical data sets (Garbary et al 1993 Garbary amp Renza-glia 1998) and a molecular data set that combinessequences of both nuclear and mitochondrial smallsubunit (SSU) rDNA
2 CELL STRUCTURE
Among embryophytes there is remarkable consistencyin the ultrastructure of parenchyma cells The `typicalrsquoliving photosynthetic cell contains a large central vacuolea peripheral nucleus mitochondria endoplasmic reti-culum and lenticular chloroplasts equipped with granaand scattered starch (Gunning amp Steer 1996) The bryo-phytes and lycophytes exhibit the greatest diversity inorganellar complement and centne structure and thesecellular features provide important clues as to evolu-tionary relationships (Duckett amp Renzaglia 1988 Brownamp Lemmon 1993) Solitary plastids (monoplastidy) arefound in most vegetative cells of hornworts and charac-terize mitotic cells of representative taxa andor tissues(especially spermatogenous tissue) of all bryophytes andlycophytes (Brown amp Lemmon 1990a 1993) Likewisemonoplastidic meiosis (discussed below) is distributedamong representatives of all these clades Based on thewidespread occurrence of monoplastidy in charophyceanalgae and the exclusive distribution of this condition amongbasal embryophytes it is safe to conclude that monoplastidyis plesiomorphic (Brown amp Lemmon 1990a Graham1993) To ensure distribution of a plastid into daughtercells the division cycle of plastids must be tightly linkedto nuclear division in monoplastidic cell lineages Plastidsare intimately associated with microtubule-organizingcentres (MTOCs) and the two form the focal points forproduction of spindle microtubules
In liverworts only spermatogenous cells are monoplas-tidic and vegetative cells are polyplastidic During themitotic cycle well-decentned electron-dense aggregationsthe so-called polar organizers organize the spindlesThese structures also characterize the monoplastidicdividing cells in spermatogenous tissue of liverwortsEndoplasmic reticulum within the polar organizer isconnected to the nuclear envelope and in this regardresembles the nuclear envelope-based centrosome ofmosses and tracheophytes (Vaughn amp Harper 1998) Latespermatogenous cells of all archegoniates have centriolarcentrosomes and this enables direct comparison withsimilar algal MTOCs (see below)
Examination of the internal structure of hornwort plas-tids provides evidence of further retention of algalfeatures Pyrenoids are found in most anthocerote taxaand as in algae these electron-dense intrachloroplastbodies are the site of localization of RUBISCO (Vaughnet al 1990) Grana of anthocerotes lack end membranestypical of other embryophytes and as in Coleochaete thyla-koids traverse and dissect the pyrenoid into subunits(Vaughn et al 1992) Channel thylakoids found in algaebut not other land plants further point to these organellesas primitive in structure Physiological specializationsapparently are associated with this chloroplast micro-anatomy especially mechanisms concentrating CO2 andthese may have been benecentcial in a land habitat (Smithamp Gricurrenths 1986ab)
770 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Although chloroplast structure in hepatics is similar tothat of mosses and tracheophytes liverworts possess aunique and diagnostic single membrane-bound oil bodyOil bodies are not found in all liverworts but they arewidespread in basal taxa suggesting loss in more derivedlineages Originating in meristematic cells from endo-plasmic reticulum (Duckett amp Ligrone 1995) the oil bodyhas no counterpart in any other plant group The functionof these organelles is problematic as they apparently arenot used as a food reserve
3 VEGETATIVE GAMETOPHYTES
The phrase gametophyte dominantrsquo distinguishes bryo-phytes from all other embryophytes With the sexualphase as the prominent persistent generation modicentca-tions of the bryophyte gametophyte entailed adaptivewalks that optimized vegetative elaboration while simul-taneously facilitating water-dependent sexual reproduc-tion In this regard bryophytes and pteridophytes areamphibious ie they require uninterrupted access towater for reproductive success and continue with vegeta-tive activities during times of water deprivation Bryo-phytes diiexcler from pteridophytes in that it is thegametophyte alone that contends with these often-conpoundicting roles of sexual reproductive and vegetativepersistence and dispersal In pteridophytes the sporo-phyte is the phase responsible for production of biomassand colonization of new sites It is not surprising thereforethat from perusal of structural complexity in gameto-phytes among land plants two features are evident (i)this generation never attains great stature and (ii) bryo-phyte gametophytes are the most elaborate of the landplants Thus bryophytes provide an opportunity to eval-uate the structural strategies that enabled robust gameto-phytes to endure in an environment that presentedextreme selective pressures for a motility-based fertiliza-tion system We acknowledge that physiological mechan-isms for contending with life in the air includingelaborate biochemical pathways for desiccation toleranceand thermotolerance in many mosses and pteridophytesplayed a signicentcant role in plant evolution (Oliver ampWood 1997 Kenrick amp Crane 1997a Hanson et al 1999)However it is not within the scope of this review todiscuss these physiological strategies
Two fundamental growth forms are exhibited by bryo-phytes a poundattened prostrate thallus (hornworts andcomplex thalloid and simple thalloid hepatics) and anerect or creeping cylindrical leafy shoot (leafy liverwortsselected simple thalloid liverworts and mosses) Thesegrowth habits correspond to the optimal oblate spheroidand cylindrical life forms hypothesized for small multi-cellular terrestrial organisms in morphospace (Niklas1997) Compared with spherical growth forms that arefound in representative green algae dorsiventrallycompressed thalli reduce total surface area whileproviding maximum surface area to volume for gasexchange and light harvesting (Niklas 1997) Leafy formssegregate vegetative functions into specialized organspoundattened leaves maximize light capture and enhancephotosynthetic capacity while the central cylindrical stemenhances water conservation and enables exploratorygrowth including upright growth in some taxa Thalloid
genera rarely grow upright interestingly Hymenophytonand Symphyogyna two notable exceptions in the simplethalloid hepatics both possess well-developed internalwater-conducting systems (Ligrone et al this issue) Theerect habit facilitates exploitation of air for exchange ofgases and light trapping It also provides a means forelevating the attached sporophyte and thus promotingspore dispersal At issue for both the thalloid and leafygrowth forms is water conservation and protection ofvulnerable growing tissues and reproductive organs
A single apical cell is responsible for growth of bryo-phyte gametophytes Apical cell geometry may be one offour fundamental shapes wedge-shaped (cuneate) lens-shaped (lenticular) tetrahedral (pyramidal) or hemidis-coid (Crandall-Stotler 1986 Renzaglia 1982) Pyramidalsystems are typically associated with leafy habits lenti-cular systems with highly poundattened thalli often withthickened midrib and monostromatic wings (eg Metz-geria and Pallavicinia) and wedge-shaped and hemidiscoidapical cells are responsible for thalloid growth formsincluding those of complex thalloid liverworts and horn-worts In mosses pyramidal systems predominate withthe rare secondary derivation of a lenticular system(Crandall-Stotler 1984) Similarly the wedge-shapedapical initial is widespread in hornworts with a develop-mental and evolutionary transformation to hemidiscoid-based generative cells in Dendroceros (Renzaglia 1978)Hepatics are the most diverse in that all four apical cellshapes occur in various taxa and developmental modicentca-tions of cell geometry are commonplace Documentationexists for ontogenetic transformation of cell shape fromtetrahedral to lens-shaped from lens-shaped to tetra-hedral or wedge-shaped and from wedge-shaped tohemidiscoid (Fulford 1956 Renzaglia 1982 Renzaglia ampBartholomew 1985) Often these geometric transitions areassociated with a change in gross morphology andororientation of growth For example the transition fromjuvenile to mature habit is accompanied by a transforma-tion from lenticular to tetrahedral apical cell geometry incertain leafy hepatics (Fulford 1956) In Fossombronia atetrahedral apical cell forms immediately in the uprightsporeling and limited segmentation produces three rowsof leaves (Renzaglia amp Bartholomew 1985) Furtheroblique divisions convert the tetrahedral apical cell to alenticular cell and this change results in the maturethallus habit with two lateral leaf rows Concomitantwith this metamorphosis is a switch in orientation ofgrowth from vertical to horizontal The discussion aboveserves to illustrate that (i) conversion of apical cell fromone geometric shape to another is a cornerstone ofnormal developmental processes in bryophytes and isespecially widespread in liverworts (ii) changes in apicalcell shape are accompanied by changes in both growthform and thallusshoot orientation and (iii) because ofthe easersquo with which apical cell shape is modicented and theprofound inpounduence this modicentcation has on habit thisprocess alone may well have provided signicentcantmorphological variation for major evolutionary change
Crandall-Stotler (1980 1984 1986) clearly documenteda fundamental diiexclerence between most mosses and leafyliverworts (Jungermanniales) in segmentation of thepyramidal apical cell and subsequent leaf developmentSpiralled undivided leaves in mosses versus three rows of
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 771
Phil Trans R Soc Lond B (2000)
bicentd leaves are the rudiments of these two distinct onto-genetic patterns As concluded by Crandall-Stotler(1980) these underlying developmental discrepanciessupport the concept of independent origin of leaves inmosses and liverworts We further suggest that additionaldiiexclerences in developmental design provide evidence thatleaves or leaf-like structures (`leavesrsquo) have evolved atleast twice in mosses and perhaps multiple times in liver-worts Production of the dissected leaves (phyllids) ofTakakia is decidedly diiexclerent from leaf development inSphagnum and bryopsid mosses (Crandall-Stotler 1986)suggesting that leaf evolution in Takakia was independentof that in other mosses Because of diiexclerences in apicalorganization and the phyletic distance between taxa suchas Haplomitrium Treubia Fossombronia Noteroclada Sphaero-carpos and Blasia it is possible that `leaf rsquo acquisition ishomoplastic in all of these taxa
Leaves not only function as photosynthetic organs butthey also provide protection for the fragile growing tipsLeaf primordia overarch and surround the apical cell andimmediate derivatives Controlled production of stalkedmucilage-secreting hairs (slime papillae) accompaniesleaf development in mosses and liverworts and furtheraiexclords protection against damage and desiccation In theabsence of leaves abundant uniseriate to branched slimepapillae surround and protect the meristematic region ofsimple thalloid liverworts (Renzaglia 1982 Duckett et al1990) In addition to slime papillae poundattened multicel-lular scales with marginal papillae provide an alternativebut equally eiexclective protective structure in Blasia Cavicu-laria and complex thalloid liverworts
In contrast to mosses liverwort evolution has involvedselection towards dorsiventrality Schuster (1979 1984ab)suggested that the ancestral liverwort gametophyte was abranched leapoundess upright axis much like that of mostDevonian gametophytes but lacking conducting tissueand stomata (Remy et al 1993) In extant hepatics erectradially symmetrical shoots with three equal rows ofleaves are considered plesiomorphic while procumbentthalloid or leafy forms with reduced to absent underleavesare viewed as specialized Appression to the substratemaximizes contact with water sources and thus minimizespotential damage associated with desiccation In leafyliverworts prostrate habits have evolved as an adaptationto horizontal and vertical substrates In general incubousleaf insertions are associated with vertical substrates (egtree trunks) while succubous phenotypes predominate onhorizontal substrates
While erect radial habits are exhibited by relativelyfew basal taxa of hepatics isophyllous acrocarpscomprise approximately half of all mosses (Crum ampAnderson 1981) Evolutionary adaptations that haveenabled mosses to grow upright and persist in relativelydry environments include the acquisition of more exten-sive and specialized conducting tissues (Hebant 1977Ligrone et al this issue) the thickening of cell walls andaccumulation of polyphenolic compounds in these wallsand the exploitation of metabolic pathways that conferdesiccation and thermal tolerance Moreover growth ofmultiple individuals in dense mats and tufts eiexclectivelyfacilitates absorption and retention of water in manymosses Abundant uniseriate branched to scale-like para-phyllia and pseudoparaphyllia are found on branches of
some mosses and further serve to protect against desicca-tion and damage (Crum amp Anderson 1981)
In mosses branched centlamentous protonemata developupon spore germination and unlike sporelings of liver-worts and hornworts these juvenile stages have theability to produce and disperse multiple leafy shootsderived from a single spore This latter feature is particu-larly adaptive in acrocarps that do not produce creepingshoots and rarely branch
The exclusively thalloid hornworts do not produceleaves slime papillae scales or any organized externalappendage Indeed these plants perform virtually allvegetative and reproductive functions in undiiexclerentiatedparenchyma cells or within internal thallus chambers(Renzaglia 1978 Renzaglia amp Vaughn 2000) Perhapsone of the fundamental features that has been instru-mental in the survival of anthocerotes is the ability toproduce copious mucopolysaccharides (mucilage) invirtually any cell apparently in response to environ-mental cues Mucilages are carbohydrates that areconcerned with imbibition and retention of waterAlthough mosses and liverworts produce mucilage thissubstance with rare exceptions (eg the ventral surface ofthe thallus of Treubia and the underground axes of Calo-bryales) is produced only in slime papillae In hornwortsprotection of the growing notch is aiexclorded by mucilagesecretion from epidermal cells of young apical derivativesScattered internal cells contain mucilage in most taxaand large mucilage-centlled chambers are common in Antho-ceros and selected species of other genera During develop-ment antheridial chambers contain mucilage andmucilage surrounds the exterior of archegonial necksCapitalization on mucilage production and the ability tosequester this hydrophilic substance in internal chambersare integral processes that bestowed partial tolerance todrying conditions in these small thalloid taxa
A further evolutionary innovation that almost certainlycontributed to the vegetative and reproductive success ofhornworts (as measured by length of time in existence)was the ability to form schizogenous spaces Essential tothis process is the separation of adjacent cells at themiddle lamella followed by controlled division and expan-sion of cells around a chamber or canal In addition toexclusively mucilage-containing cavities hornwortssequester antheridia and Nostoc colonies in such chambers
This motif of internalization coupled with cell separa-tion and mucilage proliferation is embodied in thedevelopment of Nostoc colonies in anthocerotes (Renzaglia1978) This symbiotic relationship between the hornwortand a nitrogen-centxing bacterium has enabled these plantsto poundourish under conditions of low nitrogen The develop-ment of Nostoc colonies is ubiquitous in anthocerotes andbegins near the apical cell with the apparently randomseparation between two adjacent epidermal cells in theventral thallus The resulting mucilage cleft provides aport through which soil-dwelling cyanobacteria mayenter the thallus With maturation of the thallus a schizo-genous mucilage-centlled chamber forms internal to the cleftand expands with multiplication of cells of the NostocendosymbiontThallus cells elongate and grow among thecyanobacterium The mature colony may be rather exten-sive and often forms a mound that projects from theventral thallus
772 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
In hepatics Nostoc colonies are restricted to two sistergenera Blasia and Cavicularia (Renzaglia 1982) A cursorycomparison of Nostoc colony production in Blasia withthat in anthocerotes exemplicentes highly divergent develop-mental strategies for attaining functionally similar struc-tures in the two bryophyte groups In Blasia Nostoc ishoused in external `organsrsquo on the ventral thallus This so-called auricle is produced in a controlled fashion fromapical derivatives and originates as a mucilage hairwhich undergoes extensive elaboration Nostoc enters theauricle when it is a small dome-shaped structure andconcomitant growth of Nostoc and thallus intermixes cellsof the plant and prokaryote Continued auricle expansionresults in a massive external structure that supercentciallyresembles the Nostoc colony of hornworts
Having considered apical growth protective structuresand habit variability in the three bryophyte groupscomparisons with basal pteridophytes are in order Ingeneral gametophytes of pteridophytes are not as compli-cated or long-lived as those of bryophytes (Bierhorst 1971Giiexclord amp Foster 1988 Bell 1994 Whittier 1977 19811983 Whittier amp Webster 1986 Whittier amp Thomas1993) Although variability exists among the genera twofundamental growth forms predominate in basal tracheo-phytes (i) green epiterrestrial forms with irregularupright lamellae and (ii) thick poundeshy subterranean formswith a fungal symbiont The latter type exemplicented byHuperzia (centgure 1a) Lycopodium Diphasiastrum Phegmar-iurus Psilophytes (centgure 1b) Botrychium and Ophioglossumis generally considered ancestral These gametophytesmay persist for several years and they have the capacityto produce multiple sporophytes and form abundantregenerants The gametophytes of Phylloglossum (Whittieramp Braggins 1992) and Huperzia (A R Maden and D P
Whittier unpublished data) are non-photosynthetic whenyoung and convert to green forms when exposed to lightEpiterrestrial gametophytes of Palhinhaea LycopodiellaPseudolycopodiella and Equisetum typically survive for a singlegrowing season but in some instances may bear severalsporophytes (Bierhorst 1971 Duckett amp Duckett 1980)Apical growth is accomplished by a meristematic zone inEquisetum (Duckett 1970) and in lycophytes (centgure 1a) butinvolves a well-decentned tetrahedral apical cell in Psilotumand Tmesipteris (centgure 1bc) Protection of the growingregion and gametangia is accomplished by production ofglandular and non-glandular hairs rhizoids and limitedmucilage production (Whittier amp Peterson 1984) Oftenapices of subterranean gametophytes are not surroundedby protective structures (centgure 1bc) Although no leavesare produced by any tracheophyte gametophyte the capa-city to form abundant hairs and papillae is reminiscent ofmosses and liverworts However conducting tissue whichis commonplace in mosses and more restricted in occur-rence in liverworts is a developmental anomaly in gameto-phytes of extant pteridophytes (Bierhorst 1971)
Fungal associations are widespread in gametophytes ofpteridophytes liverworts and hornworts but are lackingin mosses (Duckett et al 1991) Fungal hyphae facilitateabsorption and translocation of minerals water andorganic molecules within the plant In nature hetero-trophic subterranean gametophytes of pteridophytes relyexclusively on endophytic fungi for nutrient uptakeFungal^plant symbioses were probably established in theprimitive archegoniates and have developed repeatedlyduring the evolution of land plants (Pirozynski ampMalloch 1975 Pirozynski 1981) For a discussion ofsymbiotic interactions among fungi and bryophytes seeRead et al (this issue)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 773
Phil Trans R Soc Lond B (2000)
Figure 1 Scanning electron micrographs of cultured pteridophyte gametophytes provided by Dean P Whittier (a) Younggametophyte of Huperzia lucidula Meristematic zone (M) in a groove that continues around laterally Rhizoids (R) originatefrom ventral derivatives while uniseriate hairs and gametangia (not produced yet) are formed on a dorsal crown Bar ˆ 01 mmMicrograph by Angel R Maden (bc) Tmesipteris lanceolata (b) Cylindrical gametophyte with abundant uniseriate hairs (H)archegonia (A) and rhizoids (R) The growing tip appears to be unprotected Bar ˆ 01 mm (c) Enlarged view of the growing tipshowing triangular pyramidal apical cell (AC) surrounded by packets of cells The prominent walls that outline cell packets(merophytes) represent original segmentation from the apical cell Bar ˆ 005 mm
4 REPRODUCTIVE GAMETOPHYTES
Selective pressures in a terrestrial environment forreproductive processes that require uninterrupted accessto water are indeed extreme The transmigration of algaeto land and the subsequent evolution of embryophytesnecessitated the evolution of multicellular sex organs(Niklas 1997) These organs not only served to protectvulnerable developing gametes but also the origin ofmulticellular female sex organs (archegonia) was a requi-site for embryo development a universal feature of landplants The evolution of gametangia in archegoniatesundoubtedly was a complex historical process thatrequired coordination of morphogenetic with physio-logical and ecological signals For example production ofsex organs must be timed appropriately and must occurrapidly in response to pounductuating seasonal conditionsGametangia must be strategically placed on the plant andprotective structures amply developed In gametophyte-dominant land-dwelling plants the compromise betweenoptimization of vegetative growth and the constraints ofsexual reproduction often resulted in the abandonment orreduced emphasis on genetic exchange (ie elaboration ofmechanisms for asexual reproduction and self-fertilization)Such strategies are also common in algal groups butevolved in response to diiexclerent selective pressures
In an attempt to understand adaptive strategies andcharacter evolution in the sexual phase of early arche-goniates we will briepoundy examine and compare (i) sexorgan ontogeny and (ii) spermatogenesis among basalbryophytes and seedless tracheophytes
Unlike those of pteridophytes gametangia of mossesand liverworts are stalked and extend from the epidermalsurface (Bold et al 1987) In leafy taxa gametangia areeither associated with leaves or leaves surround a terminal
774 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
hornworts
pteridophytes
(a)
( f ) (g) (h)
(b) (c) (d) (e)
(i) ( j )
mature neck
mature neck
neckventer
neck canal cellsventral canal cellegg
neck canal cellsventral canal cellegg
neck
Figure 2 Comparison of archegonial development in hornworts (a^e) and pteridophytes ( f^j ) (a) Longitudinal view and(b) cross-sectional view of archegonial initial traversed by three anticlinal walls that delimit a central axial cell and threeperipheral cells The axial cell gives rise to the neck canal cells ventral canal cell and egg while the peripheral cells form sterileouter layers (c) As seen in cross-section three additional segments in the peripheral cells form six rows of neck cells (d ) Lightmicrograph surface view of six rows of neck cells in Phaeoceros laevis (e) Longitudinal section of the sunken archegonium ofPhaeoceros laevis ( f ) Longitudinal view and (g) cross-sectional view of archegonial initial traversed by single periclinal wall thatdelimits an outer cell that forms the neck and an inner cell that gives rise to neck canal cells ventral canal cell and egg (h) Asseen in cross-section successive anticlinal divisions in the outer cell form four rows of neck cells (i) Scanning electron micrographof archegonial neck with four rows of cells in Psilotum nudum ( j ) Longitudinal section of sunken archegonium of Pteridiumaquilinum
Figure 3 Light micrograph of antheridium of Takakiaceratophylla Longitudinal section of anterior of developingelongated antheridium Cells are in packets arranged in tworows that represent the original biseriate centlament producedby left^right divisions from an apical cell The outline of theapical cell is visible as a triangular wall containing developingsperm cells at the antheridial tip Bar ˆ 100 m m
cluster of sex organs Abundant papillae hairs and para-physes are intermixed among gametangia and providefurther protection Thalloid liverworts have evolved avast array of structures and mechanisms for protectinggametangia including inrolled ventral branches (Metz-geria) secondarily sunken chambers (Pellia Noteroclada andcomplex thalloid liverworts) and lamellar outgrowths ofthe thallus (Renzaglia 1982) In contrast gametangia inhornworts are produced and maintained within theconcentnes of the thallus and as such supercentcially resemblethose of pteridophytes Gametangia of basal pterido-phytes are never stalked and are either entirely sunkenwithin the thallus or form conspicuous epidermal mounds(Bierhorst 1971)
To evaluate homology of gametangia it is critical toexamine cell division patterns and especially the initialformative division sequences during which the develop-mental fate of subsequent cell lineages is determined(Roux 1895 Niklas 1997) These primary divisions aregenetically controlled and provide the blueprint for organo-genesis At centrst glance there appears to be no similarityin archegonial ontogeny among liverworts hornworts andmosses Archegonial development in mosses uniquelyinvolves the initial production of an apical cell thatsegments left and right forming a biseriate centlament(Schocenteld 1985 Smith 1955) The archegonium properforms from the terminal cell in the centlament after theapical cell ceases its patterned mitotic divisions Hornwortand liverwort archegonia develop without elongation
from an apical cell In mosses and liverworts the arche-gonial initial elongates and divides above the epidermalsurface while in hornworts the developing archegoniumremains surrounded by thallus tissue This mosaic ofcharacters related to archegonial ontogeny providesevidence of divergent developmental pathways in thebryophyte clades However when the formative divisionsequences that establish the archegonium proper areexamined a fundamental pattern emerges that is diag-nostic of bryophytes (Schuster 1966) In all three groupsthe process involves three longitudinal divisions that forma central triangular axial cell surrounded by threeperipheral cells (centgure 2ab) The peripheral cells formthe neck and venter while the axial cell gives rise to theneck canal cells ventral canal cell and egg Further divi-sions in the peripheral cells typically result in a neck ofcentve or six cell rows (centgure 2cd) (Renzaglia 1982)
This division pattern is markedly diiexclerent from that ofpteridophytes in which the single epidermal initial centrstdivides transversely (periclinal division) The outer cellthen further divides to form the neck while the inner celldivides periclinally to produce neck canal cells ventralcanal cell and egg (centgure 2 fg j) (Bierhorst 1971 Kenrickamp Crane 1997a) With expansion the archegoniumprojects from the epidermal surface The neck invariablyconsists of four vertical rows of cells (centgure 2hi) Clearlythere are no parallels in development between the sunkenarchegonia of hornworts (centgure 2e) and similarembedded archegonia in pteridophytes (centgure 2 j)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 775
Phil Trans R Soc Lond B (2000)
hornworts
(a)
(c)
(b)
(d)
chamber roof
schizogenous space
antheridial initials
antheridial complex
pteridophytes
jacket
spermatogenous tissue
antheridium
Anthoceros
Ophioglossum
Figure 4 Comparison of antheridial development in hornworts (ab) and pteridophytes (cd) (a) In hornworts a periclinaldivision in a supercentcial cell delimits a subepidermal cell that gives rise to up to 25 antheridia and an epidermal cell that producesa chamber roof two cells thick A schizogenous space forms the cavity that houses the antheridia Antheridial development frominitials at the base of the cavity follows a developmental pathway similar to other bryophytes (b) Light micrograph of antheridialcomplex in Anthoceros agrestis (c) In pteridophytes a periclinal division in a supercentcial cell delimits a subepidermal cell that givesrise to the spermatogenous tissue and an epidermal cell that forms the sterile jacket (d) The single antheridium that developsfrom this process in Ophioglossum engelmannii
Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

and simultaneous patterns of extensive morphologicaldiversicentcation followed by widespread decimationpresumably characterized these early stages of land colon-ization (Gould 1989 Kenrick amp Crane 1997a) Of themagnitude of morphological experiments that wereattempted only limited fragments have persisted throughthe millennia including the three lineages of bryophytes(Stewart amp Rothwell 1993 Taylor amp Taylor 1993)
Given the time since divergence the depauperate fossilrecord and the vastness of the newly inhabited landscapeit is not surprising that the early phylogenetic history ofland plants remains one of the major unresolved problemsin evolutionary biology Axiomatic to current evolu-tionary thought is the concept that embryophytes aremonophyletic that bryophytes represent the basal cladesand that Charophyceae especially Coleochaetales andCharales are the closest living algal relatives of arche-goniates (McCourt 1995) Beyond these accepted maximsthere is no consensus as to interrelationships among bryo-phytes and basal pteridophytes Indeed virtually everyconceivable hypothesis regarding bryophyte phylogenyhas been proposed from paraphyletic in a variety ofbranching orders to hornworts basal and mosses andliverworts monophyletic to a completely monophyleticbryophyte assemblage (Bopp amp Capesius 1996 Capesiusamp Bopp 1997 Bremer et al 1987 Garbary et al 1993Garbary amp Renzaglia 1998 Hedderson et al 1998 Lewiset al 1997 Kenrick amp Crane 1997a Malek et al 1996Mishler amp Churchill 1984 1985 Mishler et al 1994 Duiexclamp Nickrent 1999 Nishiyama amp Kato 1999) Clearly newdata or perhaps new insights on analysing existing dataare required to resolve this phylogenetic dilemma
By nature of their antiquity liverworts hornwortsmosses and basal pteridophytes represent relics of a oncemore diverse poundora and as the oldest living remnants ofearly land colonization these organisms provide a livinglaboratory in which to examine early morphologicaladaptations to existence on land In this paper weexplore morphological and developmental features of bryo-phytes and basal pteridophytes at the cell tissue organand whole organism levels Whenever possible evidence ispresented from more informative basal representatives ofthe major clades This comparative approach is designedto identify successful phenotypic innovations of both thegametophyte and sporophyte which enabled these plantsto optimize vegetative and reproductive activities in aterrestrial setting In addition to illuminating phyloge-netic acurrennities contemplation of morphological andmorphogenetic strategies provides valuable insight intothe direction and sequence of character transformation
This review is not intended to be a comprehensiveoverview of morphology in bryophytes but rather the goalis to explore answers to the following questions related toevolutionary adaptations exhibited by early plant life onland What are the vegetative and reproductive innova-tions that enabled the three bryophyte lineages to persist How do these organisms cope with the constraints ofterrestrial existence especially desiccation and gravityAnd centnally what can be gleaned from careful scrutiny ofextant basal embryophytes about morphological char-acter evolution including divergences convergencesparallelisms and homologies These questions provide thecontext for comprehensive phylogenetic analyses in which
we review and update two previously published morpho-logical data sets (Garbary et al 1993 Garbary amp Renza-glia 1998) and a molecular data set that combinessequences of both nuclear and mitochondrial smallsubunit (SSU) rDNA
2 CELL STRUCTURE
Among embryophytes there is remarkable consistencyin the ultrastructure of parenchyma cells The `typicalrsquoliving photosynthetic cell contains a large central vacuolea peripheral nucleus mitochondria endoplasmic reti-culum and lenticular chloroplasts equipped with granaand scattered starch (Gunning amp Steer 1996) The bryo-phytes and lycophytes exhibit the greatest diversity inorganellar complement and centne structure and thesecellular features provide important clues as to evolu-tionary relationships (Duckett amp Renzaglia 1988 Brownamp Lemmon 1993) Solitary plastids (monoplastidy) arefound in most vegetative cells of hornworts and charac-terize mitotic cells of representative taxa andor tissues(especially spermatogenous tissue) of all bryophytes andlycophytes (Brown amp Lemmon 1990a 1993) Likewisemonoplastidic meiosis (discussed below) is distributedamong representatives of all these clades Based on thewidespread occurrence of monoplastidy in charophyceanalgae and the exclusive distribution of this condition amongbasal embryophytes it is safe to conclude that monoplastidyis plesiomorphic (Brown amp Lemmon 1990a Graham1993) To ensure distribution of a plastid into daughtercells the division cycle of plastids must be tightly linkedto nuclear division in monoplastidic cell lineages Plastidsare intimately associated with microtubule-organizingcentres (MTOCs) and the two form the focal points forproduction of spindle microtubules
In liverworts only spermatogenous cells are monoplas-tidic and vegetative cells are polyplastidic During themitotic cycle well-decentned electron-dense aggregationsthe so-called polar organizers organize the spindlesThese structures also characterize the monoplastidicdividing cells in spermatogenous tissue of liverwortsEndoplasmic reticulum within the polar organizer isconnected to the nuclear envelope and in this regardresembles the nuclear envelope-based centrosome ofmosses and tracheophytes (Vaughn amp Harper 1998) Latespermatogenous cells of all archegoniates have centriolarcentrosomes and this enables direct comparison withsimilar algal MTOCs (see below)
Examination of the internal structure of hornwort plas-tids provides evidence of further retention of algalfeatures Pyrenoids are found in most anthocerote taxaand as in algae these electron-dense intrachloroplastbodies are the site of localization of RUBISCO (Vaughnet al 1990) Grana of anthocerotes lack end membranestypical of other embryophytes and as in Coleochaete thyla-koids traverse and dissect the pyrenoid into subunits(Vaughn et al 1992) Channel thylakoids found in algaebut not other land plants further point to these organellesas primitive in structure Physiological specializationsapparently are associated with this chloroplast micro-anatomy especially mechanisms concentrating CO2 andthese may have been benecentcial in a land habitat (Smithamp Gricurrenths 1986ab)
770 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Although chloroplast structure in hepatics is similar tothat of mosses and tracheophytes liverworts possess aunique and diagnostic single membrane-bound oil bodyOil bodies are not found in all liverworts but they arewidespread in basal taxa suggesting loss in more derivedlineages Originating in meristematic cells from endo-plasmic reticulum (Duckett amp Ligrone 1995) the oil bodyhas no counterpart in any other plant group The functionof these organelles is problematic as they apparently arenot used as a food reserve
3 VEGETATIVE GAMETOPHYTES
The phrase gametophyte dominantrsquo distinguishes bryo-phytes from all other embryophytes With the sexualphase as the prominent persistent generation modicentca-tions of the bryophyte gametophyte entailed adaptivewalks that optimized vegetative elaboration while simul-taneously facilitating water-dependent sexual reproduc-tion In this regard bryophytes and pteridophytes areamphibious ie they require uninterrupted access towater for reproductive success and continue with vegeta-tive activities during times of water deprivation Bryo-phytes diiexcler from pteridophytes in that it is thegametophyte alone that contends with these often-conpoundicting roles of sexual reproductive and vegetativepersistence and dispersal In pteridophytes the sporo-phyte is the phase responsible for production of biomassand colonization of new sites It is not surprising thereforethat from perusal of structural complexity in gameto-phytes among land plants two features are evident (i)this generation never attains great stature and (ii) bryo-phyte gametophytes are the most elaborate of the landplants Thus bryophytes provide an opportunity to eval-uate the structural strategies that enabled robust gameto-phytes to endure in an environment that presentedextreme selective pressures for a motility-based fertiliza-tion system We acknowledge that physiological mechan-isms for contending with life in the air includingelaborate biochemical pathways for desiccation toleranceand thermotolerance in many mosses and pteridophytesplayed a signicentcant role in plant evolution (Oliver ampWood 1997 Kenrick amp Crane 1997a Hanson et al 1999)However it is not within the scope of this review todiscuss these physiological strategies
Two fundamental growth forms are exhibited by bryo-phytes a poundattened prostrate thallus (hornworts andcomplex thalloid and simple thalloid hepatics) and anerect or creeping cylindrical leafy shoot (leafy liverwortsselected simple thalloid liverworts and mosses) Thesegrowth habits correspond to the optimal oblate spheroidand cylindrical life forms hypothesized for small multi-cellular terrestrial organisms in morphospace (Niklas1997) Compared with spherical growth forms that arefound in representative green algae dorsiventrallycompressed thalli reduce total surface area whileproviding maximum surface area to volume for gasexchange and light harvesting (Niklas 1997) Leafy formssegregate vegetative functions into specialized organspoundattened leaves maximize light capture and enhancephotosynthetic capacity while the central cylindrical stemenhances water conservation and enables exploratorygrowth including upright growth in some taxa Thalloid
genera rarely grow upright interestingly Hymenophytonand Symphyogyna two notable exceptions in the simplethalloid hepatics both possess well-developed internalwater-conducting systems (Ligrone et al this issue) Theerect habit facilitates exploitation of air for exchange ofgases and light trapping It also provides a means forelevating the attached sporophyte and thus promotingspore dispersal At issue for both the thalloid and leafygrowth forms is water conservation and protection ofvulnerable growing tissues and reproductive organs
A single apical cell is responsible for growth of bryo-phyte gametophytes Apical cell geometry may be one offour fundamental shapes wedge-shaped (cuneate) lens-shaped (lenticular) tetrahedral (pyramidal) or hemidis-coid (Crandall-Stotler 1986 Renzaglia 1982) Pyramidalsystems are typically associated with leafy habits lenti-cular systems with highly poundattened thalli often withthickened midrib and monostromatic wings (eg Metz-geria and Pallavicinia) and wedge-shaped and hemidiscoidapical cells are responsible for thalloid growth formsincluding those of complex thalloid liverworts and horn-worts In mosses pyramidal systems predominate withthe rare secondary derivation of a lenticular system(Crandall-Stotler 1984) Similarly the wedge-shapedapical initial is widespread in hornworts with a develop-mental and evolutionary transformation to hemidiscoid-based generative cells in Dendroceros (Renzaglia 1978)Hepatics are the most diverse in that all four apical cellshapes occur in various taxa and developmental modicentca-tions of cell geometry are commonplace Documentationexists for ontogenetic transformation of cell shape fromtetrahedral to lens-shaped from lens-shaped to tetra-hedral or wedge-shaped and from wedge-shaped tohemidiscoid (Fulford 1956 Renzaglia 1982 Renzaglia ampBartholomew 1985) Often these geometric transitions areassociated with a change in gross morphology andororientation of growth For example the transition fromjuvenile to mature habit is accompanied by a transforma-tion from lenticular to tetrahedral apical cell geometry incertain leafy hepatics (Fulford 1956) In Fossombronia atetrahedral apical cell forms immediately in the uprightsporeling and limited segmentation produces three rowsof leaves (Renzaglia amp Bartholomew 1985) Furtheroblique divisions convert the tetrahedral apical cell to alenticular cell and this change results in the maturethallus habit with two lateral leaf rows Concomitantwith this metamorphosis is a switch in orientation ofgrowth from vertical to horizontal The discussion aboveserves to illustrate that (i) conversion of apical cell fromone geometric shape to another is a cornerstone ofnormal developmental processes in bryophytes and isespecially widespread in liverworts (ii) changes in apicalcell shape are accompanied by changes in both growthform and thallusshoot orientation and (iii) because ofthe easersquo with which apical cell shape is modicented and theprofound inpounduence this modicentcation has on habit thisprocess alone may well have provided signicentcantmorphological variation for major evolutionary change
Crandall-Stotler (1980 1984 1986) clearly documenteda fundamental diiexclerence between most mosses and leafyliverworts (Jungermanniales) in segmentation of thepyramidal apical cell and subsequent leaf developmentSpiralled undivided leaves in mosses versus three rows of
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 771
Phil Trans R Soc Lond B (2000)
bicentd leaves are the rudiments of these two distinct onto-genetic patterns As concluded by Crandall-Stotler(1980) these underlying developmental discrepanciessupport the concept of independent origin of leaves inmosses and liverworts We further suggest that additionaldiiexclerences in developmental design provide evidence thatleaves or leaf-like structures (`leavesrsquo) have evolved atleast twice in mosses and perhaps multiple times in liver-worts Production of the dissected leaves (phyllids) ofTakakia is decidedly diiexclerent from leaf development inSphagnum and bryopsid mosses (Crandall-Stotler 1986)suggesting that leaf evolution in Takakia was independentof that in other mosses Because of diiexclerences in apicalorganization and the phyletic distance between taxa suchas Haplomitrium Treubia Fossombronia Noteroclada Sphaero-carpos and Blasia it is possible that `leaf rsquo acquisition ishomoplastic in all of these taxa
Leaves not only function as photosynthetic organs butthey also provide protection for the fragile growing tipsLeaf primordia overarch and surround the apical cell andimmediate derivatives Controlled production of stalkedmucilage-secreting hairs (slime papillae) accompaniesleaf development in mosses and liverworts and furtheraiexclords protection against damage and desiccation In theabsence of leaves abundant uniseriate to branched slimepapillae surround and protect the meristematic region ofsimple thalloid liverworts (Renzaglia 1982 Duckett et al1990) In addition to slime papillae poundattened multicel-lular scales with marginal papillae provide an alternativebut equally eiexclective protective structure in Blasia Cavicu-laria and complex thalloid liverworts
In contrast to mosses liverwort evolution has involvedselection towards dorsiventrality Schuster (1979 1984ab)suggested that the ancestral liverwort gametophyte was abranched leapoundess upright axis much like that of mostDevonian gametophytes but lacking conducting tissueand stomata (Remy et al 1993) In extant hepatics erectradially symmetrical shoots with three equal rows ofleaves are considered plesiomorphic while procumbentthalloid or leafy forms with reduced to absent underleavesare viewed as specialized Appression to the substratemaximizes contact with water sources and thus minimizespotential damage associated with desiccation In leafyliverworts prostrate habits have evolved as an adaptationto horizontal and vertical substrates In general incubousleaf insertions are associated with vertical substrates (egtree trunks) while succubous phenotypes predominate onhorizontal substrates
While erect radial habits are exhibited by relativelyfew basal taxa of hepatics isophyllous acrocarpscomprise approximately half of all mosses (Crum ampAnderson 1981) Evolutionary adaptations that haveenabled mosses to grow upright and persist in relativelydry environments include the acquisition of more exten-sive and specialized conducting tissues (Hebant 1977Ligrone et al this issue) the thickening of cell walls andaccumulation of polyphenolic compounds in these wallsand the exploitation of metabolic pathways that conferdesiccation and thermal tolerance Moreover growth ofmultiple individuals in dense mats and tufts eiexclectivelyfacilitates absorption and retention of water in manymosses Abundant uniseriate branched to scale-like para-phyllia and pseudoparaphyllia are found on branches of
some mosses and further serve to protect against desicca-tion and damage (Crum amp Anderson 1981)
In mosses branched centlamentous protonemata developupon spore germination and unlike sporelings of liver-worts and hornworts these juvenile stages have theability to produce and disperse multiple leafy shootsderived from a single spore This latter feature is particu-larly adaptive in acrocarps that do not produce creepingshoots and rarely branch
The exclusively thalloid hornworts do not produceleaves slime papillae scales or any organized externalappendage Indeed these plants perform virtually allvegetative and reproductive functions in undiiexclerentiatedparenchyma cells or within internal thallus chambers(Renzaglia 1978 Renzaglia amp Vaughn 2000) Perhapsone of the fundamental features that has been instru-mental in the survival of anthocerotes is the ability toproduce copious mucopolysaccharides (mucilage) invirtually any cell apparently in response to environ-mental cues Mucilages are carbohydrates that areconcerned with imbibition and retention of waterAlthough mosses and liverworts produce mucilage thissubstance with rare exceptions (eg the ventral surface ofthe thallus of Treubia and the underground axes of Calo-bryales) is produced only in slime papillae In hornwortsprotection of the growing notch is aiexclorded by mucilagesecretion from epidermal cells of young apical derivativesScattered internal cells contain mucilage in most taxaand large mucilage-centlled chambers are common in Antho-ceros and selected species of other genera During develop-ment antheridial chambers contain mucilage andmucilage surrounds the exterior of archegonial necksCapitalization on mucilage production and the ability tosequester this hydrophilic substance in internal chambersare integral processes that bestowed partial tolerance todrying conditions in these small thalloid taxa
A further evolutionary innovation that almost certainlycontributed to the vegetative and reproductive success ofhornworts (as measured by length of time in existence)was the ability to form schizogenous spaces Essential tothis process is the separation of adjacent cells at themiddle lamella followed by controlled division and expan-sion of cells around a chamber or canal In addition toexclusively mucilage-containing cavities hornwortssequester antheridia and Nostoc colonies in such chambers
This motif of internalization coupled with cell separa-tion and mucilage proliferation is embodied in thedevelopment of Nostoc colonies in anthocerotes (Renzaglia1978) This symbiotic relationship between the hornwortand a nitrogen-centxing bacterium has enabled these plantsto poundourish under conditions of low nitrogen The develop-ment of Nostoc colonies is ubiquitous in anthocerotes andbegins near the apical cell with the apparently randomseparation between two adjacent epidermal cells in theventral thallus The resulting mucilage cleft provides aport through which soil-dwelling cyanobacteria mayenter the thallus With maturation of the thallus a schizo-genous mucilage-centlled chamber forms internal to the cleftand expands with multiplication of cells of the NostocendosymbiontThallus cells elongate and grow among thecyanobacterium The mature colony may be rather exten-sive and often forms a mound that projects from theventral thallus
772 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
In hepatics Nostoc colonies are restricted to two sistergenera Blasia and Cavicularia (Renzaglia 1982) A cursorycomparison of Nostoc colony production in Blasia withthat in anthocerotes exemplicentes highly divergent develop-mental strategies for attaining functionally similar struc-tures in the two bryophyte groups In Blasia Nostoc ishoused in external `organsrsquo on the ventral thallus This so-called auricle is produced in a controlled fashion fromapical derivatives and originates as a mucilage hairwhich undergoes extensive elaboration Nostoc enters theauricle when it is a small dome-shaped structure andconcomitant growth of Nostoc and thallus intermixes cellsof the plant and prokaryote Continued auricle expansionresults in a massive external structure that supercentciallyresembles the Nostoc colony of hornworts
Having considered apical growth protective structuresand habit variability in the three bryophyte groupscomparisons with basal pteridophytes are in order Ingeneral gametophytes of pteridophytes are not as compli-cated or long-lived as those of bryophytes (Bierhorst 1971Giiexclord amp Foster 1988 Bell 1994 Whittier 1977 19811983 Whittier amp Webster 1986 Whittier amp Thomas1993) Although variability exists among the genera twofundamental growth forms predominate in basal tracheo-phytes (i) green epiterrestrial forms with irregularupright lamellae and (ii) thick poundeshy subterranean formswith a fungal symbiont The latter type exemplicented byHuperzia (centgure 1a) Lycopodium Diphasiastrum Phegmar-iurus Psilophytes (centgure 1b) Botrychium and Ophioglossumis generally considered ancestral These gametophytesmay persist for several years and they have the capacityto produce multiple sporophytes and form abundantregenerants The gametophytes of Phylloglossum (Whittieramp Braggins 1992) and Huperzia (A R Maden and D P
Whittier unpublished data) are non-photosynthetic whenyoung and convert to green forms when exposed to lightEpiterrestrial gametophytes of Palhinhaea LycopodiellaPseudolycopodiella and Equisetum typically survive for a singlegrowing season but in some instances may bear severalsporophytes (Bierhorst 1971 Duckett amp Duckett 1980)Apical growth is accomplished by a meristematic zone inEquisetum (Duckett 1970) and in lycophytes (centgure 1a) butinvolves a well-decentned tetrahedral apical cell in Psilotumand Tmesipteris (centgure 1bc) Protection of the growingregion and gametangia is accomplished by production ofglandular and non-glandular hairs rhizoids and limitedmucilage production (Whittier amp Peterson 1984) Oftenapices of subterranean gametophytes are not surroundedby protective structures (centgure 1bc) Although no leavesare produced by any tracheophyte gametophyte the capa-city to form abundant hairs and papillae is reminiscent ofmosses and liverworts However conducting tissue whichis commonplace in mosses and more restricted in occur-rence in liverworts is a developmental anomaly in gameto-phytes of extant pteridophytes (Bierhorst 1971)
Fungal associations are widespread in gametophytes ofpteridophytes liverworts and hornworts but are lackingin mosses (Duckett et al 1991) Fungal hyphae facilitateabsorption and translocation of minerals water andorganic molecules within the plant In nature hetero-trophic subterranean gametophytes of pteridophytes relyexclusively on endophytic fungi for nutrient uptakeFungal^plant symbioses were probably established in theprimitive archegoniates and have developed repeatedlyduring the evolution of land plants (Pirozynski ampMalloch 1975 Pirozynski 1981) For a discussion ofsymbiotic interactions among fungi and bryophytes seeRead et al (this issue)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 773
Phil Trans R Soc Lond B (2000)
Figure 1 Scanning electron micrographs of cultured pteridophyte gametophytes provided by Dean P Whittier (a) Younggametophyte of Huperzia lucidula Meristematic zone (M) in a groove that continues around laterally Rhizoids (R) originatefrom ventral derivatives while uniseriate hairs and gametangia (not produced yet) are formed on a dorsal crown Bar ˆ 01 mmMicrograph by Angel R Maden (bc) Tmesipteris lanceolata (b) Cylindrical gametophyte with abundant uniseriate hairs (H)archegonia (A) and rhizoids (R) The growing tip appears to be unprotected Bar ˆ 01 mm (c) Enlarged view of the growing tipshowing triangular pyramidal apical cell (AC) surrounded by packets of cells The prominent walls that outline cell packets(merophytes) represent original segmentation from the apical cell Bar ˆ 005 mm
4 REPRODUCTIVE GAMETOPHYTES
Selective pressures in a terrestrial environment forreproductive processes that require uninterrupted accessto water are indeed extreme The transmigration of algaeto land and the subsequent evolution of embryophytesnecessitated the evolution of multicellular sex organs(Niklas 1997) These organs not only served to protectvulnerable developing gametes but also the origin ofmulticellular female sex organs (archegonia) was a requi-site for embryo development a universal feature of landplants The evolution of gametangia in archegoniatesundoubtedly was a complex historical process thatrequired coordination of morphogenetic with physio-logical and ecological signals For example production ofsex organs must be timed appropriately and must occurrapidly in response to pounductuating seasonal conditionsGametangia must be strategically placed on the plant andprotective structures amply developed In gametophyte-dominant land-dwelling plants the compromise betweenoptimization of vegetative growth and the constraints ofsexual reproduction often resulted in the abandonment orreduced emphasis on genetic exchange (ie elaboration ofmechanisms for asexual reproduction and self-fertilization)Such strategies are also common in algal groups butevolved in response to diiexclerent selective pressures
In an attempt to understand adaptive strategies andcharacter evolution in the sexual phase of early arche-goniates we will briepoundy examine and compare (i) sexorgan ontogeny and (ii) spermatogenesis among basalbryophytes and seedless tracheophytes
Unlike those of pteridophytes gametangia of mossesand liverworts are stalked and extend from the epidermalsurface (Bold et al 1987) In leafy taxa gametangia areeither associated with leaves or leaves surround a terminal
774 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
hornworts
pteridophytes
(a)
( f ) (g) (h)
(b) (c) (d) (e)
(i) ( j )
mature neck
mature neck
neckventer
neck canal cellsventral canal cellegg
neck canal cellsventral canal cellegg
neck
Figure 2 Comparison of archegonial development in hornworts (a^e) and pteridophytes ( f^j ) (a) Longitudinal view and(b) cross-sectional view of archegonial initial traversed by three anticlinal walls that delimit a central axial cell and threeperipheral cells The axial cell gives rise to the neck canal cells ventral canal cell and egg while the peripheral cells form sterileouter layers (c) As seen in cross-section three additional segments in the peripheral cells form six rows of neck cells (d ) Lightmicrograph surface view of six rows of neck cells in Phaeoceros laevis (e) Longitudinal section of the sunken archegonium ofPhaeoceros laevis ( f ) Longitudinal view and (g) cross-sectional view of archegonial initial traversed by single periclinal wall thatdelimits an outer cell that forms the neck and an inner cell that gives rise to neck canal cells ventral canal cell and egg (h) Asseen in cross-section successive anticlinal divisions in the outer cell form four rows of neck cells (i) Scanning electron micrographof archegonial neck with four rows of cells in Psilotum nudum ( j ) Longitudinal section of sunken archegonium of Pteridiumaquilinum
Figure 3 Light micrograph of antheridium of Takakiaceratophylla Longitudinal section of anterior of developingelongated antheridium Cells are in packets arranged in tworows that represent the original biseriate centlament producedby left^right divisions from an apical cell The outline of theapical cell is visible as a triangular wall containing developingsperm cells at the antheridial tip Bar ˆ 100 m m
cluster of sex organs Abundant papillae hairs and para-physes are intermixed among gametangia and providefurther protection Thalloid liverworts have evolved avast array of structures and mechanisms for protectinggametangia including inrolled ventral branches (Metz-geria) secondarily sunken chambers (Pellia Noteroclada andcomplex thalloid liverworts) and lamellar outgrowths ofthe thallus (Renzaglia 1982) In contrast gametangia inhornworts are produced and maintained within theconcentnes of the thallus and as such supercentcially resemblethose of pteridophytes Gametangia of basal pterido-phytes are never stalked and are either entirely sunkenwithin the thallus or form conspicuous epidermal mounds(Bierhorst 1971)
To evaluate homology of gametangia it is critical toexamine cell division patterns and especially the initialformative division sequences during which the develop-mental fate of subsequent cell lineages is determined(Roux 1895 Niklas 1997) These primary divisions aregenetically controlled and provide the blueprint for organo-genesis At centrst glance there appears to be no similarityin archegonial ontogeny among liverworts hornworts andmosses Archegonial development in mosses uniquelyinvolves the initial production of an apical cell thatsegments left and right forming a biseriate centlament(Schocenteld 1985 Smith 1955) The archegonium properforms from the terminal cell in the centlament after theapical cell ceases its patterned mitotic divisions Hornwortand liverwort archegonia develop without elongation
from an apical cell In mosses and liverworts the arche-gonial initial elongates and divides above the epidermalsurface while in hornworts the developing archegoniumremains surrounded by thallus tissue This mosaic ofcharacters related to archegonial ontogeny providesevidence of divergent developmental pathways in thebryophyte clades However when the formative divisionsequences that establish the archegonium proper areexamined a fundamental pattern emerges that is diag-nostic of bryophytes (Schuster 1966) In all three groupsthe process involves three longitudinal divisions that forma central triangular axial cell surrounded by threeperipheral cells (centgure 2ab) The peripheral cells formthe neck and venter while the axial cell gives rise to theneck canal cells ventral canal cell and egg Further divi-sions in the peripheral cells typically result in a neck ofcentve or six cell rows (centgure 2cd) (Renzaglia 1982)
This division pattern is markedly diiexclerent from that ofpteridophytes in which the single epidermal initial centrstdivides transversely (periclinal division) The outer cellthen further divides to form the neck while the inner celldivides periclinally to produce neck canal cells ventralcanal cell and egg (centgure 2 fg j) (Bierhorst 1971 Kenrickamp Crane 1997a) With expansion the archegoniumprojects from the epidermal surface The neck invariablyconsists of four vertical rows of cells (centgure 2hi) Clearlythere are no parallels in development between the sunkenarchegonia of hornworts (centgure 2e) and similarembedded archegonia in pteridophytes (centgure 2 j)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 775
Phil Trans R Soc Lond B (2000)
hornworts
(a)
(c)
(b)
(d)
chamber roof
schizogenous space
antheridial initials
antheridial complex
pteridophytes
jacket
spermatogenous tissue
antheridium
Anthoceros
Ophioglossum
Figure 4 Comparison of antheridial development in hornworts (ab) and pteridophytes (cd) (a) In hornworts a periclinaldivision in a supercentcial cell delimits a subepidermal cell that gives rise to up to 25 antheridia and an epidermal cell that producesa chamber roof two cells thick A schizogenous space forms the cavity that houses the antheridia Antheridial development frominitials at the base of the cavity follows a developmental pathway similar to other bryophytes (b) Light micrograph of antheridialcomplex in Anthoceros agrestis (c) In pteridophytes a periclinal division in a supercentcial cell delimits a subepidermal cell that givesrise to the spermatogenous tissue and an epidermal cell that forms the sterile jacket (d) The single antheridium that developsfrom this process in Ophioglossum engelmannii
Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

Although chloroplast structure in hepatics is similar tothat of mosses and tracheophytes liverworts possess aunique and diagnostic single membrane-bound oil bodyOil bodies are not found in all liverworts but they arewidespread in basal taxa suggesting loss in more derivedlineages Originating in meristematic cells from endo-plasmic reticulum (Duckett amp Ligrone 1995) the oil bodyhas no counterpart in any other plant group The functionof these organelles is problematic as they apparently arenot used as a food reserve
3 VEGETATIVE GAMETOPHYTES
The phrase gametophyte dominantrsquo distinguishes bryo-phytes from all other embryophytes With the sexualphase as the prominent persistent generation modicentca-tions of the bryophyte gametophyte entailed adaptivewalks that optimized vegetative elaboration while simul-taneously facilitating water-dependent sexual reproduc-tion In this regard bryophytes and pteridophytes areamphibious ie they require uninterrupted access towater for reproductive success and continue with vegeta-tive activities during times of water deprivation Bryo-phytes diiexcler from pteridophytes in that it is thegametophyte alone that contends with these often-conpoundicting roles of sexual reproductive and vegetativepersistence and dispersal In pteridophytes the sporo-phyte is the phase responsible for production of biomassand colonization of new sites It is not surprising thereforethat from perusal of structural complexity in gameto-phytes among land plants two features are evident (i)this generation never attains great stature and (ii) bryo-phyte gametophytes are the most elaborate of the landplants Thus bryophytes provide an opportunity to eval-uate the structural strategies that enabled robust gameto-phytes to endure in an environment that presentedextreme selective pressures for a motility-based fertiliza-tion system We acknowledge that physiological mechan-isms for contending with life in the air includingelaborate biochemical pathways for desiccation toleranceand thermotolerance in many mosses and pteridophytesplayed a signicentcant role in plant evolution (Oliver ampWood 1997 Kenrick amp Crane 1997a Hanson et al 1999)However it is not within the scope of this review todiscuss these physiological strategies
Two fundamental growth forms are exhibited by bryo-phytes a poundattened prostrate thallus (hornworts andcomplex thalloid and simple thalloid hepatics) and anerect or creeping cylindrical leafy shoot (leafy liverwortsselected simple thalloid liverworts and mosses) Thesegrowth habits correspond to the optimal oblate spheroidand cylindrical life forms hypothesized for small multi-cellular terrestrial organisms in morphospace (Niklas1997) Compared with spherical growth forms that arefound in representative green algae dorsiventrallycompressed thalli reduce total surface area whileproviding maximum surface area to volume for gasexchange and light harvesting (Niklas 1997) Leafy formssegregate vegetative functions into specialized organspoundattened leaves maximize light capture and enhancephotosynthetic capacity while the central cylindrical stemenhances water conservation and enables exploratorygrowth including upright growth in some taxa Thalloid
genera rarely grow upright interestingly Hymenophytonand Symphyogyna two notable exceptions in the simplethalloid hepatics both possess well-developed internalwater-conducting systems (Ligrone et al this issue) Theerect habit facilitates exploitation of air for exchange ofgases and light trapping It also provides a means forelevating the attached sporophyte and thus promotingspore dispersal At issue for both the thalloid and leafygrowth forms is water conservation and protection ofvulnerable growing tissues and reproductive organs
A single apical cell is responsible for growth of bryo-phyte gametophytes Apical cell geometry may be one offour fundamental shapes wedge-shaped (cuneate) lens-shaped (lenticular) tetrahedral (pyramidal) or hemidis-coid (Crandall-Stotler 1986 Renzaglia 1982) Pyramidalsystems are typically associated with leafy habits lenti-cular systems with highly poundattened thalli often withthickened midrib and monostromatic wings (eg Metz-geria and Pallavicinia) and wedge-shaped and hemidiscoidapical cells are responsible for thalloid growth formsincluding those of complex thalloid liverworts and horn-worts In mosses pyramidal systems predominate withthe rare secondary derivation of a lenticular system(Crandall-Stotler 1984) Similarly the wedge-shapedapical initial is widespread in hornworts with a develop-mental and evolutionary transformation to hemidiscoid-based generative cells in Dendroceros (Renzaglia 1978)Hepatics are the most diverse in that all four apical cellshapes occur in various taxa and developmental modicentca-tions of cell geometry are commonplace Documentationexists for ontogenetic transformation of cell shape fromtetrahedral to lens-shaped from lens-shaped to tetra-hedral or wedge-shaped and from wedge-shaped tohemidiscoid (Fulford 1956 Renzaglia 1982 Renzaglia ampBartholomew 1985) Often these geometric transitions areassociated with a change in gross morphology andororientation of growth For example the transition fromjuvenile to mature habit is accompanied by a transforma-tion from lenticular to tetrahedral apical cell geometry incertain leafy hepatics (Fulford 1956) In Fossombronia atetrahedral apical cell forms immediately in the uprightsporeling and limited segmentation produces three rowsof leaves (Renzaglia amp Bartholomew 1985) Furtheroblique divisions convert the tetrahedral apical cell to alenticular cell and this change results in the maturethallus habit with two lateral leaf rows Concomitantwith this metamorphosis is a switch in orientation ofgrowth from vertical to horizontal The discussion aboveserves to illustrate that (i) conversion of apical cell fromone geometric shape to another is a cornerstone ofnormal developmental processes in bryophytes and isespecially widespread in liverworts (ii) changes in apicalcell shape are accompanied by changes in both growthform and thallusshoot orientation and (iii) because ofthe easersquo with which apical cell shape is modicented and theprofound inpounduence this modicentcation has on habit thisprocess alone may well have provided signicentcantmorphological variation for major evolutionary change
Crandall-Stotler (1980 1984 1986) clearly documenteda fundamental diiexclerence between most mosses and leafyliverworts (Jungermanniales) in segmentation of thepyramidal apical cell and subsequent leaf developmentSpiralled undivided leaves in mosses versus three rows of
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 771
Phil Trans R Soc Lond B (2000)
bicentd leaves are the rudiments of these two distinct onto-genetic patterns As concluded by Crandall-Stotler(1980) these underlying developmental discrepanciessupport the concept of independent origin of leaves inmosses and liverworts We further suggest that additionaldiiexclerences in developmental design provide evidence thatleaves or leaf-like structures (`leavesrsquo) have evolved atleast twice in mosses and perhaps multiple times in liver-worts Production of the dissected leaves (phyllids) ofTakakia is decidedly diiexclerent from leaf development inSphagnum and bryopsid mosses (Crandall-Stotler 1986)suggesting that leaf evolution in Takakia was independentof that in other mosses Because of diiexclerences in apicalorganization and the phyletic distance between taxa suchas Haplomitrium Treubia Fossombronia Noteroclada Sphaero-carpos and Blasia it is possible that `leaf rsquo acquisition ishomoplastic in all of these taxa
Leaves not only function as photosynthetic organs butthey also provide protection for the fragile growing tipsLeaf primordia overarch and surround the apical cell andimmediate derivatives Controlled production of stalkedmucilage-secreting hairs (slime papillae) accompaniesleaf development in mosses and liverworts and furtheraiexclords protection against damage and desiccation In theabsence of leaves abundant uniseriate to branched slimepapillae surround and protect the meristematic region ofsimple thalloid liverworts (Renzaglia 1982 Duckett et al1990) In addition to slime papillae poundattened multicel-lular scales with marginal papillae provide an alternativebut equally eiexclective protective structure in Blasia Cavicu-laria and complex thalloid liverworts
In contrast to mosses liverwort evolution has involvedselection towards dorsiventrality Schuster (1979 1984ab)suggested that the ancestral liverwort gametophyte was abranched leapoundess upright axis much like that of mostDevonian gametophytes but lacking conducting tissueand stomata (Remy et al 1993) In extant hepatics erectradially symmetrical shoots with three equal rows ofleaves are considered plesiomorphic while procumbentthalloid or leafy forms with reduced to absent underleavesare viewed as specialized Appression to the substratemaximizes contact with water sources and thus minimizespotential damage associated with desiccation In leafyliverworts prostrate habits have evolved as an adaptationto horizontal and vertical substrates In general incubousleaf insertions are associated with vertical substrates (egtree trunks) while succubous phenotypes predominate onhorizontal substrates
While erect radial habits are exhibited by relativelyfew basal taxa of hepatics isophyllous acrocarpscomprise approximately half of all mosses (Crum ampAnderson 1981) Evolutionary adaptations that haveenabled mosses to grow upright and persist in relativelydry environments include the acquisition of more exten-sive and specialized conducting tissues (Hebant 1977Ligrone et al this issue) the thickening of cell walls andaccumulation of polyphenolic compounds in these wallsand the exploitation of metabolic pathways that conferdesiccation and thermal tolerance Moreover growth ofmultiple individuals in dense mats and tufts eiexclectivelyfacilitates absorption and retention of water in manymosses Abundant uniseriate branched to scale-like para-phyllia and pseudoparaphyllia are found on branches of
some mosses and further serve to protect against desicca-tion and damage (Crum amp Anderson 1981)
In mosses branched centlamentous protonemata developupon spore germination and unlike sporelings of liver-worts and hornworts these juvenile stages have theability to produce and disperse multiple leafy shootsderived from a single spore This latter feature is particu-larly adaptive in acrocarps that do not produce creepingshoots and rarely branch
The exclusively thalloid hornworts do not produceleaves slime papillae scales or any organized externalappendage Indeed these plants perform virtually allvegetative and reproductive functions in undiiexclerentiatedparenchyma cells or within internal thallus chambers(Renzaglia 1978 Renzaglia amp Vaughn 2000) Perhapsone of the fundamental features that has been instru-mental in the survival of anthocerotes is the ability toproduce copious mucopolysaccharides (mucilage) invirtually any cell apparently in response to environ-mental cues Mucilages are carbohydrates that areconcerned with imbibition and retention of waterAlthough mosses and liverworts produce mucilage thissubstance with rare exceptions (eg the ventral surface ofthe thallus of Treubia and the underground axes of Calo-bryales) is produced only in slime papillae In hornwortsprotection of the growing notch is aiexclorded by mucilagesecretion from epidermal cells of young apical derivativesScattered internal cells contain mucilage in most taxaand large mucilage-centlled chambers are common in Antho-ceros and selected species of other genera During develop-ment antheridial chambers contain mucilage andmucilage surrounds the exterior of archegonial necksCapitalization on mucilage production and the ability tosequester this hydrophilic substance in internal chambersare integral processes that bestowed partial tolerance todrying conditions in these small thalloid taxa
A further evolutionary innovation that almost certainlycontributed to the vegetative and reproductive success ofhornworts (as measured by length of time in existence)was the ability to form schizogenous spaces Essential tothis process is the separation of adjacent cells at themiddle lamella followed by controlled division and expan-sion of cells around a chamber or canal In addition toexclusively mucilage-containing cavities hornwortssequester antheridia and Nostoc colonies in such chambers
This motif of internalization coupled with cell separa-tion and mucilage proliferation is embodied in thedevelopment of Nostoc colonies in anthocerotes (Renzaglia1978) This symbiotic relationship between the hornwortand a nitrogen-centxing bacterium has enabled these plantsto poundourish under conditions of low nitrogen The develop-ment of Nostoc colonies is ubiquitous in anthocerotes andbegins near the apical cell with the apparently randomseparation between two adjacent epidermal cells in theventral thallus The resulting mucilage cleft provides aport through which soil-dwelling cyanobacteria mayenter the thallus With maturation of the thallus a schizo-genous mucilage-centlled chamber forms internal to the cleftand expands with multiplication of cells of the NostocendosymbiontThallus cells elongate and grow among thecyanobacterium The mature colony may be rather exten-sive and often forms a mound that projects from theventral thallus
772 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
In hepatics Nostoc colonies are restricted to two sistergenera Blasia and Cavicularia (Renzaglia 1982) A cursorycomparison of Nostoc colony production in Blasia withthat in anthocerotes exemplicentes highly divergent develop-mental strategies for attaining functionally similar struc-tures in the two bryophyte groups In Blasia Nostoc ishoused in external `organsrsquo on the ventral thallus This so-called auricle is produced in a controlled fashion fromapical derivatives and originates as a mucilage hairwhich undergoes extensive elaboration Nostoc enters theauricle when it is a small dome-shaped structure andconcomitant growth of Nostoc and thallus intermixes cellsof the plant and prokaryote Continued auricle expansionresults in a massive external structure that supercentciallyresembles the Nostoc colony of hornworts
Having considered apical growth protective structuresand habit variability in the three bryophyte groupscomparisons with basal pteridophytes are in order Ingeneral gametophytes of pteridophytes are not as compli-cated or long-lived as those of bryophytes (Bierhorst 1971Giiexclord amp Foster 1988 Bell 1994 Whittier 1977 19811983 Whittier amp Webster 1986 Whittier amp Thomas1993) Although variability exists among the genera twofundamental growth forms predominate in basal tracheo-phytes (i) green epiterrestrial forms with irregularupright lamellae and (ii) thick poundeshy subterranean formswith a fungal symbiont The latter type exemplicented byHuperzia (centgure 1a) Lycopodium Diphasiastrum Phegmar-iurus Psilophytes (centgure 1b) Botrychium and Ophioglossumis generally considered ancestral These gametophytesmay persist for several years and they have the capacityto produce multiple sporophytes and form abundantregenerants The gametophytes of Phylloglossum (Whittieramp Braggins 1992) and Huperzia (A R Maden and D P
Whittier unpublished data) are non-photosynthetic whenyoung and convert to green forms when exposed to lightEpiterrestrial gametophytes of Palhinhaea LycopodiellaPseudolycopodiella and Equisetum typically survive for a singlegrowing season but in some instances may bear severalsporophytes (Bierhorst 1971 Duckett amp Duckett 1980)Apical growth is accomplished by a meristematic zone inEquisetum (Duckett 1970) and in lycophytes (centgure 1a) butinvolves a well-decentned tetrahedral apical cell in Psilotumand Tmesipteris (centgure 1bc) Protection of the growingregion and gametangia is accomplished by production ofglandular and non-glandular hairs rhizoids and limitedmucilage production (Whittier amp Peterson 1984) Oftenapices of subterranean gametophytes are not surroundedby protective structures (centgure 1bc) Although no leavesare produced by any tracheophyte gametophyte the capa-city to form abundant hairs and papillae is reminiscent ofmosses and liverworts However conducting tissue whichis commonplace in mosses and more restricted in occur-rence in liverworts is a developmental anomaly in gameto-phytes of extant pteridophytes (Bierhorst 1971)
Fungal associations are widespread in gametophytes ofpteridophytes liverworts and hornworts but are lackingin mosses (Duckett et al 1991) Fungal hyphae facilitateabsorption and translocation of minerals water andorganic molecules within the plant In nature hetero-trophic subterranean gametophytes of pteridophytes relyexclusively on endophytic fungi for nutrient uptakeFungal^plant symbioses were probably established in theprimitive archegoniates and have developed repeatedlyduring the evolution of land plants (Pirozynski ampMalloch 1975 Pirozynski 1981) For a discussion ofsymbiotic interactions among fungi and bryophytes seeRead et al (this issue)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 773
Phil Trans R Soc Lond B (2000)
Figure 1 Scanning electron micrographs of cultured pteridophyte gametophytes provided by Dean P Whittier (a) Younggametophyte of Huperzia lucidula Meristematic zone (M) in a groove that continues around laterally Rhizoids (R) originatefrom ventral derivatives while uniseriate hairs and gametangia (not produced yet) are formed on a dorsal crown Bar ˆ 01 mmMicrograph by Angel R Maden (bc) Tmesipteris lanceolata (b) Cylindrical gametophyte with abundant uniseriate hairs (H)archegonia (A) and rhizoids (R) The growing tip appears to be unprotected Bar ˆ 01 mm (c) Enlarged view of the growing tipshowing triangular pyramidal apical cell (AC) surrounded by packets of cells The prominent walls that outline cell packets(merophytes) represent original segmentation from the apical cell Bar ˆ 005 mm
4 REPRODUCTIVE GAMETOPHYTES
Selective pressures in a terrestrial environment forreproductive processes that require uninterrupted accessto water are indeed extreme The transmigration of algaeto land and the subsequent evolution of embryophytesnecessitated the evolution of multicellular sex organs(Niklas 1997) These organs not only served to protectvulnerable developing gametes but also the origin ofmulticellular female sex organs (archegonia) was a requi-site for embryo development a universal feature of landplants The evolution of gametangia in archegoniatesundoubtedly was a complex historical process thatrequired coordination of morphogenetic with physio-logical and ecological signals For example production ofsex organs must be timed appropriately and must occurrapidly in response to pounductuating seasonal conditionsGametangia must be strategically placed on the plant andprotective structures amply developed In gametophyte-dominant land-dwelling plants the compromise betweenoptimization of vegetative growth and the constraints ofsexual reproduction often resulted in the abandonment orreduced emphasis on genetic exchange (ie elaboration ofmechanisms for asexual reproduction and self-fertilization)Such strategies are also common in algal groups butevolved in response to diiexclerent selective pressures
In an attempt to understand adaptive strategies andcharacter evolution in the sexual phase of early arche-goniates we will briepoundy examine and compare (i) sexorgan ontogeny and (ii) spermatogenesis among basalbryophytes and seedless tracheophytes
Unlike those of pteridophytes gametangia of mossesand liverworts are stalked and extend from the epidermalsurface (Bold et al 1987) In leafy taxa gametangia areeither associated with leaves or leaves surround a terminal
774 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
hornworts
pteridophytes
(a)
( f ) (g) (h)
(b) (c) (d) (e)
(i) ( j )
mature neck
mature neck
neckventer
neck canal cellsventral canal cellegg
neck canal cellsventral canal cellegg
neck
Figure 2 Comparison of archegonial development in hornworts (a^e) and pteridophytes ( f^j ) (a) Longitudinal view and(b) cross-sectional view of archegonial initial traversed by three anticlinal walls that delimit a central axial cell and threeperipheral cells The axial cell gives rise to the neck canal cells ventral canal cell and egg while the peripheral cells form sterileouter layers (c) As seen in cross-section three additional segments in the peripheral cells form six rows of neck cells (d ) Lightmicrograph surface view of six rows of neck cells in Phaeoceros laevis (e) Longitudinal section of the sunken archegonium ofPhaeoceros laevis ( f ) Longitudinal view and (g) cross-sectional view of archegonial initial traversed by single periclinal wall thatdelimits an outer cell that forms the neck and an inner cell that gives rise to neck canal cells ventral canal cell and egg (h) Asseen in cross-section successive anticlinal divisions in the outer cell form four rows of neck cells (i) Scanning electron micrographof archegonial neck with four rows of cells in Psilotum nudum ( j ) Longitudinal section of sunken archegonium of Pteridiumaquilinum
Figure 3 Light micrograph of antheridium of Takakiaceratophylla Longitudinal section of anterior of developingelongated antheridium Cells are in packets arranged in tworows that represent the original biseriate centlament producedby left^right divisions from an apical cell The outline of theapical cell is visible as a triangular wall containing developingsperm cells at the antheridial tip Bar ˆ 100 m m
cluster of sex organs Abundant papillae hairs and para-physes are intermixed among gametangia and providefurther protection Thalloid liverworts have evolved avast array of structures and mechanisms for protectinggametangia including inrolled ventral branches (Metz-geria) secondarily sunken chambers (Pellia Noteroclada andcomplex thalloid liverworts) and lamellar outgrowths ofthe thallus (Renzaglia 1982) In contrast gametangia inhornworts are produced and maintained within theconcentnes of the thallus and as such supercentcially resemblethose of pteridophytes Gametangia of basal pterido-phytes are never stalked and are either entirely sunkenwithin the thallus or form conspicuous epidermal mounds(Bierhorst 1971)
To evaluate homology of gametangia it is critical toexamine cell division patterns and especially the initialformative division sequences during which the develop-mental fate of subsequent cell lineages is determined(Roux 1895 Niklas 1997) These primary divisions aregenetically controlled and provide the blueprint for organo-genesis At centrst glance there appears to be no similarityin archegonial ontogeny among liverworts hornworts andmosses Archegonial development in mosses uniquelyinvolves the initial production of an apical cell thatsegments left and right forming a biseriate centlament(Schocenteld 1985 Smith 1955) The archegonium properforms from the terminal cell in the centlament after theapical cell ceases its patterned mitotic divisions Hornwortand liverwort archegonia develop without elongation
from an apical cell In mosses and liverworts the arche-gonial initial elongates and divides above the epidermalsurface while in hornworts the developing archegoniumremains surrounded by thallus tissue This mosaic ofcharacters related to archegonial ontogeny providesevidence of divergent developmental pathways in thebryophyte clades However when the formative divisionsequences that establish the archegonium proper areexamined a fundamental pattern emerges that is diag-nostic of bryophytes (Schuster 1966) In all three groupsthe process involves three longitudinal divisions that forma central triangular axial cell surrounded by threeperipheral cells (centgure 2ab) The peripheral cells formthe neck and venter while the axial cell gives rise to theneck canal cells ventral canal cell and egg Further divi-sions in the peripheral cells typically result in a neck ofcentve or six cell rows (centgure 2cd) (Renzaglia 1982)
This division pattern is markedly diiexclerent from that ofpteridophytes in which the single epidermal initial centrstdivides transversely (periclinal division) The outer cellthen further divides to form the neck while the inner celldivides periclinally to produce neck canal cells ventralcanal cell and egg (centgure 2 fg j) (Bierhorst 1971 Kenrickamp Crane 1997a) With expansion the archegoniumprojects from the epidermal surface The neck invariablyconsists of four vertical rows of cells (centgure 2hi) Clearlythere are no parallels in development between the sunkenarchegonia of hornworts (centgure 2e) and similarembedded archegonia in pteridophytes (centgure 2 j)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 775
Phil Trans R Soc Lond B (2000)
hornworts
(a)
(c)
(b)
(d)
chamber roof
schizogenous space
antheridial initials
antheridial complex
pteridophytes
jacket
spermatogenous tissue
antheridium
Anthoceros
Ophioglossum
Figure 4 Comparison of antheridial development in hornworts (ab) and pteridophytes (cd) (a) In hornworts a periclinaldivision in a supercentcial cell delimits a subepidermal cell that gives rise to up to 25 antheridia and an epidermal cell that producesa chamber roof two cells thick A schizogenous space forms the cavity that houses the antheridia Antheridial development frominitials at the base of the cavity follows a developmental pathway similar to other bryophytes (b) Light micrograph of antheridialcomplex in Anthoceros agrestis (c) In pteridophytes a periclinal division in a supercentcial cell delimits a subepidermal cell that givesrise to the spermatogenous tissue and an epidermal cell that forms the sterile jacket (d) The single antheridium that developsfrom this process in Ophioglossum engelmannii
Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

bicentd leaves are the rudiments of these two distinct onto-genetic patterns As concluded by Crandall-Stotler(1980) these underlying developmental discrepanciessupport the concept of independent origin of leaves inmosses and liverworts We further suggest that additionaldiiexclerences in developmental design provide evidence thatleaves or leaf-like structures (`leavesrsquo) have evolved atleast twice in mosses and perhaps multiple times in liver-worts Production of the dissected leaves (phyllids) ofTakakia is decidedly diiexclerent from leaf development inSphagnum and bryopsid mosses (Crandall-Stotler 1986)suggesting that leaf evolution in Takakia was independentof that in other mosses Because of diiexclerences in apicalorganization and the phyletic distance between taxa suchas Haplomitrium Treubia Fossombronia Noteroclada Sphaero-carpos and Blasia it is possible that `leaf rsquo acquisition ishomoplastic in all of these taxa
Leaves not only function as photosynthetic organs butthey also provide protection for the fragile growing tipsLeaf primordia overarch and surround the apical cell andimmediate derivatives Controlled production of stalkedmucilage-secreting hairs (slime papillae) accompaniesleaf development in mosses and liverworts and furtheraiexclords protection against damage and desiccation In theabsence of leaves abundant uniseriate to branched slimepapillae surround and protect the meristematic region ofsimple thalloid liverworts (Renzaglia 1982 Duckett et al1990) In addition to slime papillae poundattened multicel-lular scales with marginal papillae provide an alternativebut equally eiexclective protective structure in Blasia Cavicu-laria and complex thalloid liverworts
In contrast to mosses liverwort evolution has involvedselection towards dorsiventrality Schuster (1979 1984ab)suggested that the ancestral liverwort gametophyte was abranched leapoundess upright axis much like that of mostDevonian gametophytes but lacking conducting tissueand stomata (Remy et al 1993) In extant hepatics erectradially symmetrical shoots with three equal rows ofleaves are considered plesiomorphic while procumbentthalloid or leafy forms with reduced to absent underleavesare viewed as specialized Appression to the substratemaximizes contact with water sources and thus minimizespotential damage associated with desiccation In leafyliverworts prostrate habits have evolved as an adaptationto horizontal and vertical substrates In general incubousleaf insertions are associated with vertical substrates (egtree trunks) while succubous phenotypes predominate onhorizontal substrates
While erect radial habits are exhibited by relativelyfew basal taxa of hepatics isophyllous acrocarpscomprise approximately half of all mosses (Crum ampAnderson 1981) Evolutionary adaptations that haveenabled mosses to grow upright and persist in relativelydry environments include the acquisition of more exten-sive and specialized conducting tissues (Hebant 1977Ligrone et al this issue) the thickening of cell walls andaccumulation of polyphenolic compounds in these wallsand the exploitation of metabolic pathways that conferdesiccation and thermal tolerance Moreover growth ofmultiple individuals in dense mats and tufts eiexclectivelyfacilitates absorption and retention of water in manymosses Abundant uniseriate branched to scale-like para-phyllia and pseudoparaphyllia are found on branches of
some mosses and further serve to protect against desicca-tion and damage (Crum amp Anderson 1981)
In mosses branched centlamentous protonemata developupon spore germination and unlike sporelings of liver-worts and hornworts these juvenile stages have theability to produce and disperse multiple leafy shootsderived from a single spore This latter feature is particu-larly adaptive in acrocarps that do not produce creepingshoots and rarely branch
The exclusively thalloid hornworts do not produceleaves slime papillae scales or any organized externalappendage Indeed these plants perform virtually allvegetative and reproductive functions in undiiexclerentiatedparenchyma cells or within internal thallus chambers(Renzaglia 1978 Renzaglia amp Vaughn 2000) Perhapsone of the fundamental features that has been instru-mental in the survival of anthocerotes is the ability toproduce copious mucopolysaccharides (mucilage) invirtually any cell apparently in response to environ-mental cues Mucilages are carbohydrates that areconcerned with imbibition and retention of waterAlthough mosses and liverworts produce mucilage thissubstance with rare exceptions (eg the ventral surface ofthe thallus of Treubia and the underground axes of Calo-bryales) is produced only in slime papillae In hornwortsprotection of the growing notch is aiexclorded by mucilagesecretion from epidermal cells of young apical derivativesScattered internal cells contain mucilage in most taxaand large mucilage-centlled chambers are common in Antho-ceros and selected species of other genera During develop-ment antheridial chambers contain mucilage andmucilage surrounds the exterior of archegonial necksCapitalization on mucilage production and the ability tosequester this hydrophilic substance in internal chambersare integral processes that bestowed partial tolerance todrying conditions in these small thalloid taxa
A further evolutionary innovation that almost certainlycontributed to the vegetative and reproductive success ofhornworts (as measured by length of time in existence)was the ability to form schizogenous spaces Essential tothis process is the separation of adjacent cells at themiddle lamella followed by controlled division and expan-sion of cells around a chamber or canal In addition toexclusively mucilage-containing cavities hornwortssequester antheridia and Nostoc colonies in such chambers
This motif of internalization coupled with cell separa-tion and mucilage proliferation is embodied in thedevelopment of Nostoc colonies in anthocerotes (Renzaglia1978) This symbiotic relationship between the hornwortand a nitrogen-centxing bacterium has enabled these plantsto poundourish under conditions of low nitrogen The develop-ment of Nostoc colonies is ubiquitous in anthocerotes andbegins near the apical cell with the apparently randomseparation between two adjacent epidermal cells in theventral thallus The resulting mucilage cleft provides aport through which soil-dwelling cyanobacteria mayenter the thallus With maturation of the thallus a schizo-genous mucilage-centlled chamber forms internal to the cleftand expands with multiplication of cells of the NostocendosymbiontThallus cells elongate and grow among thecyanobacterium The mature colony may be rather exten-sive and often forms a mound that projects from theventral thallus
772 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
In hepatics Nostoc colonies are restricted to two sistergenera Blasia and Cavicularia (Renzaglia 1982) A cursorycomparison of Nostoc colony production in Blasia withthat in anthocerotes exemplicentes highly divergent develop-mental strategies for attaining functionally similar struc-tures in the two bryophyte groups In Blasia Nostoc ishoused in external `organsrsquo on the ventral thallus This so-called auricle is produced in a controlled fashion fromapical derivatives and originates as a mucilage hairwhich undergoes extensive elaboration Nostoc enters theauricle when it is a small dome-shaped structure andconcomitant growth of Nostoc and thallus intermixes cellsof the plant and prokaryote Continued auricle expansionresults in a massive external structure that supercentciallyresembles the Nostoc colony of hornworts
Having considered apical growth protective structuresand habit variability in the three bryophyte groupscomparisons with basal pteridophytes are in order Ingeneral gametophytes of pteridophytes are not as compli-cated or long-lived as those of bryophytes (Bierhorst 1971Giiexclord amp Foster 1988 Bell 1994 Whittier 1977 19811983 Whittier amp Webster 1986 Whittier amp Thomas1993) Although variability exists among the genera twofundamental growth forms predominate in basal tracheo-phytes (i) green epiterrestrial forms with irregularupright lamellae and (ii) thick poundeshy subterranean formswith a fungal symbiont The latter type exemplicented byHuperzia (centgure 1a) Lycopodium Diphasiastrum Phegmar-iurus Psilophytes (centgure 1b) Botrychium and Ophioglossumis generally considered ancestral These gametophytesmay persist for several years and they have the capacityto produce multiple sporophytes and form abundantregenerants The gametophytes of Phylloglossum (Whittieramp Braggins 1992) and Huperzia (A R Maden and D P
Whittier unpublished data) are non-photosynthetic whenyoung and convert to green forms when exposed to lightEpiterrestrial gametophytes of Palhinhaea LycopodiellaPseudolycopodiella and Equisetum typically survive for a singlegrowing season but in some instances may bear severalsporophytes (Bierhorst 1971 Duckett amp Duckett 1980)Apical growth is accomplished by a meristematic zone inEquisetum (Duckett 1970) and in lycophytes (centgure 1a) butinvolves a well-decentned tetrahedral apical cell in Psilotumand Tmesipteris (centgure 1bc) Protection of the growingregion and gametangia is accomplished by production ofglandular and non-glandular hairs rhizoids and limitedmucilage production (Whittier amp Peterson 1984) Oftenapices of subterranean gametophytes are not surroundedby protective structures (centgure 1bc) Although no leavesare produced by any tracheophyte gametophyte the capa-city to form abundant hairs and papillae is reminiscent ofmosses and liverworts However conducting tissue whichis commonplace in mosses and more restricted in occur-rence in liverworts is a developmental anomaly in gameto-phytes of extant pteridophytes (Bierhorst 1971)
Fungal associations are widespread in gametophytes ofpteridophytes liverworts and hornworts but are lackingin mosses (Duckett et al 1991) Fungal hyphae facilitateabsorption and translocation of minerals water andorganic molecules within the plant In nature hetero-trophic subterranean gametophytes of pteridophytes relyexclusively on endophytic fungi for nutrient uptakeFungal^plant symbioses were probably established in theprimitive archegoniates and have developed repeatedlyduring the evolution of land plants (Pirozynski ampMalloch 1975 Pirozynski 1981) For a discussion ofsymbiotic interactions among fungi and bryophytes seeRead et al (this issue)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 773
Phil Trans R Soc Lond B (2000)
Figure 1 Scanning electron micrographs of cultured pteridophyte gametophytes provided by Dean P Whittier (a) Younggametophyte of Huperzia lucidula Meristematic zone (M) in a groove that continues around laterally Rhizoids (R) originatefrom ventral derivatives while uniseriate hairs and gametangia (not produced yet) are formed on a dorsal crown Bar ˆ 01 mmMicrograph by Angel R Maden (bc) Tmesipteris lanceolata (b) Cylindrical gametophyte with abundant uniseriate hairs (H)archegonia (A) and rhizoids (R) The growing tip appears to be unprotected Bar ˆ 01 mm (c) Enlarged view of the growing tipshowing triangular pyramidal apical cell (AC) surrounded by packets of cells The prominent walls that outline cell packets(merophytes) represent original segmentation from the apical cell Bar ˆ 005 mm
4 REPRODUCTIVE GAMETOPHYTES
Selective pressures in a terrestrial environment forreproductive processes that require uninterrupted accessto water are indeed extreme The transmigration of algaeto land and the subsequent evolution of embryophytesnecessitated the evolution of multicellular sex organs(Niklas 1997) These organs not only served to protectvulnerable developing gametes but also the origin ofmulticellular female sex organs (archegonia) was a requi-site for embryo development a universal feature of landplants The evolution of gametangia in archegoniatesundoubtedly was a complex historical process thatrequired coordination of morphogenetic with physio-logical and ecological signals For example production ofsex organs must be timed appropriately and must occurrapidly in response to pounductuating seasonal conditionsGametangia must be strategically placed on the plant andprotective structures amply developed In gametophyte-dominant land-dwelling plants the compromise betweenoptimization of vegetative growth and the constraints ofsexual reproduction often resulted in the abandonment orreduced emphasis on genetic exchange (ie elaboration ofmechanisms for asexual reproduction and self-fertilization)Such strategies are also common in algal groups butevolved in response to diiexclerent selective pressures
In an attempt to understand adaptive strategies andcharacter evolution in the sexual phase of early arche-goniates we will briepoundy examine and compare (i) sexorgan ontogeny and (ii) spermatogenesis among basalbryophytes and seedless tracheophytes
Unlike those of pteridophytes gametangia of mossesand liverworts are stalked and extend from the epidermalsurface (Bold et al 1987) In leafy taxa gametangia areeither associated with leaves or leaves surround a terminal
774 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
hornworts
pteridophytes
(a)
( f ) (g) (h)
(b) (c) (d) (e)
(i) ( j )
mature neck
mature neck
neckventer
neck canal cellsventral canal cellegg
neck canal cellsventral canal cellegg
neck
Figure 2 Comparison of archegonial development in hornworts (a^e) and pteridophytes ( f^j ) (a) Longitudinal view and(b) cross-sectional view of archegonial initial traversed by three anticlinal walls that delimit a central axial cell and threeperipheral cells The axial cell gives rise to the neck canal cells ventral canal cell and egg while the peripheral cells form sterileouter layers (c) As seen in cross-section three additional segments in the peripheral cells form six rows of neck cells (d ) Lightmicrograph surface view of six rows of neck cells in Phaeoceros laevis (e) Longitudinal section of the sunken archegonium ofPhaeoceros laevis ( f ) Longitudinal view and (g) cross-sectional view of archegonial initial traversed by single periclinal wall thatdelimits an outer cell that forms the neck and an inner cell that gives rise to neck canal cells ventral canal cell and egg (h) Asseen in cross-section successive anticlinal divisions in the outer cell form four rows of neck cells (i) Scanning electron micrographof archegonial neck with four rows of cells in Psilotum nudum ( j ) Longitudinal section of sunken archegonium of Pteridiumaquilinum
Figure 3 Light micrograph of antheridium of Takakiaceratophylla Longitudinal section of anterior of developingelongated antheridium Cells are in packets arranged in tworows that represent the original biseriate centlament producedby left^right divisions from an apical cell The outline of theapical cell is visible as a triangular wall containing developingsperm cells at the antheridial tip Bar ˆ 100 m m
cluster of sex organs Abundant papillae hairs and para-physes are intermixed among gametangia and providefurther protection Thalloid liverworts have evolved avast array of structures and mechanisms for protectinggametangia including inrolled ventral branches (Metz-geria) secondarily sunken chambers (Pellia Noteroclada andcomplex thalloid liverworts) and lamellar outgrowths ofthe thallus (Renzaglia 1982) In contrast gametangia inhornworts are produced and maintained within theconcentnes of the thallus and as such supercentcially resemblethose of pteridophytes Gametangia of basal pterido-phytes are never stalked and are either entirely sunkenwithin the thallus or form conspicuous epidermal mounds(Bierhorst 1971)
To evaluate homology of gametangia it is critical toexamine cell division patterns and especially the initialformative division sequences during which the develop-mental fate of subsequent cell lineages is determined(Roux 1895 Niklas 1997) These primary divisions aregenetically controlled and provide the blueprint for organo-genesis At centrst glance there appears to be no similarityin archegonial ontogeny among liverworts hornworts andmosses Archegonial development in mosses uniquelyinvolves the initial production of an apical cell thatsegments left and right forming a biseriate centlament(Schocenteld 1985 Smith 1955) The archegonium properforms from the terminal cell in the centlament after theapical cell ceases its patterned mitotic divisions Hornwortand liverwort archegonia develop without elongation
from an apical cell In mosses and liverworts the arche-gonial initial elongates and divides above the epidermalsurface while in hornworts the developing archegoniumremains surrounded by thallus tissue This mosaic ofcharacters related to archegonial ontogeny providesevidence of divergent developmental pathways in thebryophyte clades However when the formative divisionsequences that establish the archegonium proper areexamined a fundamental pattern emerges that is diag-nostic of bryophytes (Schuster 1966) In all three groupsthe process involves three longitudinal divisions that forma central triangular axial cell surrounded by threeperipheral cells (centgure 2ab) The peripheral cells formthe neck and venter while the axial cell gives rise to theneck canal cells ventral canal cell and egg Further divi-sions in the peripheral cells typically result in a neck ofcentve or six cell rows (centgure 2cd) (Renzaglia 1982)
This division pattern is markedly diiexclerent from that ofpteridophytes in which the single epidermal initial centrstdivides transversely (periclinal division) The outer cellthen further divides to form the neck while the inner celldivides periclinally to produce neck canal cells ventralcanal cell and egg (centgure 2 fg j) (Bierhorst 1971 Kenrickamp Crane 1997a) With expansion the archegoniumprojects from the epidermal surface The neck invariablyconsists of four vertical rows of cells (centgure 2hi) Clearlythere are no parallels in development between the sunkenarchegonia of hornworts (centgure 2e) and similarembedded archegonia in pteridophytes (centgure 2 j)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 775
Phil Trans R Soc Lond B (2000)
hornworts
(a)
(c)
(b)
(d)
chamber roof
schizogenous space
antheridial initials
antheridial complex
pteridophytes
jacket
spermatogenous tissue
antheridium
Anthoceros
Ophioglossum
Figure 4 Comparison of antheridial development in hornworts (ab) and pteridophytes (cd) (a) In hornworts a periclinaldivision in a supercentcial cell delimits a subepidermal cell that gives rise to up to 25 antheridia and an epidermal cell that producesa chamber roof two cells thick A schizogenous space forms the cavity that houses the antheridia Antheridial development frominitials at the base of the cavity follows a developmental pathway similar to other bryophytes (b) Light micrograph of antheridialcomplex in Anthoceros agrestis (c) In pteridophytes a periclinal division in a supercentcial cell delimits a subepidermal cell that givesrise to the spermatogenous tissue and an epidermal cell that forms the sterile jacket (d) The single antheridium that developsfrom this process in Ophioglossum engelmannii
Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

In hepatics Nostoc colonies are restricted to two sistergenera Blasia and Cavicularia (Renzaglia 1982) A cursorycomparison of Nostoc colony production in Blasia withthat in anthocerotes exemplicentes highly divergent develop-mental strategies for attaining functionally similar struc-tures in the two bryophyte groups In Blasia Nostoc ishoused in external `organsrsquo on the ventral thallus This so-called auricle is produced in a controlled fashion fromapical derivatives and originates as a mucilage hairwhich undergoes extensive elaboration Nostoc enters theauricle when it is a small dome-shaped structure andconcomitant growth of Nostoc and thallus intermixes cellsof the plant and prokaryote Continued auricle expansionresults in a massive external structure that supercentciallyresembles the Nostoc colony of hornworts
Having considered apical growth protective structuresand habit variability in the three bryophyte groupscomparisons with basal pteridophytes are in order Ingeneral gametophytes of pteridophytes are not as compli-cated or long-lived as those of bryophytes (Bierhorst 1971Giiexclord amp Foster 1988 Bell 1994 Whittier 1977 19811983 Whittier amp Webster 1986 Whittier amp Thomas1993) Although variability exists among the genera twofundamental growth forms predominate in basal tracheo-phytes (i) green epiterrestrial forms with irregularupright lamellae and (ii) thick poundeshy subterranean formswith a fungal symbiont The latter type exemplicented byHuperzia (centgure 1a) Lycopodium Diphasiastrum Phegmar-iurus Psilophytes (centgure 1b) Botrychium and Ophioglossumis generally considered ancestral These gametophytesmay persist for several years and they have the capacityto produce multiple sporophytes and form abundantregenerants The gametophytes of Phylloglossum (Whittieramp Braggins 1992) and Huperzia (A R Maden and D P
Whittier unpublished data) are non-photosynthetic whenyoung and convert to green forms when exposed to lightEpiterrestrial gametophytes of Palhinhaea LycopodiellaPseudolycopodiella and Equisetum typically survive for a singlegrowing season but in some instances may bear severalsporophytes (Bierhorst 1971 Duckett amp Duckett 1980)Apical growth is accomplished by a meristematic zone inEquisetum (Duckett 1970) and in lycophytes (centgure 1a) butinvolves a well-decentned tetrahedral apical cell in Psilotumand Tmesipteris (centgure 1bc) Protection of the growingregion and gametangia is accomplished by production ofglandular and non-glandular hairs rhizoids and limitedmucilage production (Whittier amp Peterson 1984) Oftenapices of subterranean gametophytes are not surroundedby protective structures (centgure 1bc) Although no leavesare produced by any tracheophyte gametophyte the capa-city to form abundant hairs and papillae is reminiscent ofmosses and liverworts However conducting tissue whichis commonplace in mosses and more restricted in occur-rence in liverworts is a developmental anomaly in gameto-phytes of extant pteridophytes (Bierhorst 1971)
Fungal associations are widespread in gametophytes ofpteridophytes liverworts and hornworts but are lackingin mosses (Duckett et al 1991) Fungal hyphae facilitateabsorption and translocation of minerals water andorganic molecules within the plant In nature hetero-trophic subterranean gametophytes of pteridophytes relyexclusively on endophytic fungi for nutrient uptakeFungal^plant symbioses were probably established in theprimitive archegoniates and have developed repeatedlyduring the evolution of land plants (Pirozynski ampMalloch 1975 Pirozynski 1981) For a discussion ofsymbiotic interactions among fungi and bryophytes seeRead et al (this issue)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 773
Phil Trans R Soc Lond B (2000)
Figure 1 Scanning electron micrographs of cultured pteridophyte gametophytes provided by Dean P Whittier (a) Younggametophyte of Huperzia lucidula Meristematic zone (M) in a groove that continues around laterally Rhizoids (R) originatefrom ventral derivatives while uniseriate hairs and gametangia (not produced yet) are formed on a dorsal crown Bar ˆ 01 mmMicrograph by Angel R Maden (bc) Tmesipteris lanceolata (b) Cylindrical gametophyte with abundant uniseriate hairs (H)archegonia (A) and rhizoids (R) The growing tip appears to be unprotected Bar ˆ 01 mm (c) Enlarged view of the growing tipshowing triangular pyramidal apical cell (AC) surrounded by packets of cells The prominent walls that outline cell packets(merophytes) represent original segmentation from the apical cell Bar ˆ 005 mm
4 REPRODUCTIVE GAMETOPHYTES
Selective pressures in a terrestrial environment forreproductive processes that require uninterrupted accessto water are indeed extreme The transmigration of algaeto land and the subsequent evolution of embryophytesnecessitated the evolution of multicellular sex organs(Niklas 1997) These organs not only served to protectvulnerable developing gametes but also the origin ofmulticellular female sex organs (archegonia) was a requi-site for embryo development a universal feature of landplants The evolution of gametangia in archegoniatesundoubtedly was a complex historical process thatrequired coordination of morphogenetic with physio-logical and ecological signals For example production ofsex organs must be timed appropriately and must occurrapidly in response to pounductuating seasonal conditionsGametangia must be strategically placed on the plant andprotective structures amply developed In gametophyte-dominant land-dwelling plants the compromise betweenoptimization of vegetative growth and the constraints ofsexual reproduction often resulted in the abandonment orreduced emphasis on genetic exchange (ie elaboration ofmechanisms for asexual reproduction and self-fertilization)Such strategies are also common in algal groups butevolved in response to diiexclerent selective pressures
In an attempt to understand adaptive strategies andcharacter evolution in the sexual phase of early arche-goniates we will briepoundy examine and compare (i) sexorgan ontogeny and (ii) spermatogenesis among basalbryophytes and seedless tracheophytes
Unlike those of pteridophytes gametangia of mossesand liverworts are stalked and extend from the epidermalsurface (Bold et al 1987) In leafy taxa gametangia areeither associated with leaves or leaves surround a terminal
774 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
hornworts
pteridophytes
(a)
( f ) (g) (h)
(b) (c) (d) (e)
(i) ( j )
mature neck
mature neck
neckventer
neck canal cellsventral canal cellegg
neck canal cellsventral canal cellegg
neck
Figure 2 Comparison of archegonial development in hornworts (a^e) and pteridophytes ( f^j ) (a) Longitudinal view and(b) cross-sectional view of archegonial initial traversed by three anticlinal walls that delimit a central axial cell and threeperipheral cells The axial cell gives rise to the neck canal cells ventral canal cell and egg while the peripheral cells form sterileouter layers (c) As seen in cross-section three additional segments in the peripheral cells form six rows of neck cells (d ) Lightmicrograph surface view of six rows of neck cells in Phaeoceros laevis (e) Longitudinal section of the sunken archegonium ofPhaeoceros laevis ( f ) Longitudinal view and (g) cross-sectional view of archegonial initial traversed by single periclinal wall thatdelimits an outer cell that forms the neck and an inner cell that gives rise to neck canal cells ventral canal cell and egg (h) Asseen in cross-section successive anticlinal divisions in the outer cell form four rows of neck cells (i) Scanning electron micrographof archegonial neck with four rows of cells in Psilotum nudum ( j ) Longitudinal section of sunken archegonium of Pteridiumaquilinum
Figure 3 Light micrograph of antheridium of Takakiaceratophylla Longitudinal section of anterior of developingelongated antheridium Cells are in packets arranged in tworows that represent the original biseriate centlament producedby left^right divisions from an apical cell The outline of theapical cell is visible as a triangular wall containing developingsperm cells at the antheridial tip Bar ˆ 100 m m
cluster of sex organs Abundant papillae hairs and para-physes are intermixed among gametangia and providefurther protection Thalloid liverworts have evolved avast array of structures and mechanisms for protectinggametangia including inrolled ventral branches (Metz-geria) secondarily sunken chambers (Pellia Noteroclada andcomplex thalloid liverworts) and lamellar outgrowths ofthe thallus (Renzaglia 1982) In contrast gametangia inhornworts are produced and maintained within theconcentnes of the thallus and as such supercentcially resemblethose of pteridophytes Gametangia of basal pterido-phytes are never stalked and are either entirely sunkenwithin the thallus or form conspicuous epidermal mounds(Bierhorst 1971)
To evaluate homology of gametangia it is critical toexamine cell division patterns and especially the initialformative division sequences during which the develop-mental fate of subsequent cell lineages is determined(Roux 1895 Niklas 1997) These primary divisions aregenetically controlled and provide the blueprint for organo-genesis At centrst glance there appears to be no similarityin archegonial ontogeny among liverworts hornworts andmosses Archegonial development in mosses uniquelyinvolves the initial production of an apical cell thatsegments left and right forming a biseriate centlament(Schocenteld 1985 Smith 1955) The archegonium properforms from the terminal cell in the centlament after theapical cell ceases its patterned mitotic divisions Hornwortand liverwort archegonia develop without elongation
from an apical cell In mosses and liverworts the arche-gonial initial elongates and divides above the epidermalsurface while in hornworts the developing archegoniumremains surrounded by thallus tissue This mosaic ofcharacters related to archegonial ontogeny providesevidence of divergent developmental pathways in thebryophyte clades However when the formative divisionsequences that establish the archegonium proper areexamined a fundamental pattern emerges that is diag-nostic of bryophytes (Schuster 1966) In all three groupsthe process involves three longitudinal divisions that forma central triangular axial cell surrounded by threeperipheral cells (centgure 2ab) The peripheral cells formthe neck and venter while the axial cell gives rise to theneck canal cells ventral canal cell and egg Further divi-sions in the peripheral cells typically result in a neck ofcentve or six cell rows (centgure 2cd) (Renzaglia 1982)
This division pattern is markedly diiexclerent from that ofpteridophytes in which the single epidermal initial centrstdivides transversely (periclinal division) The outer cellthen further divides to form the neck while the inner celldivides periclinally to produce neck canal cells ventralcanal cell and egg (centgure 2 fg j) (Bierhorst 1971 Kenrickamp Crane 1997a) With expansion the archegoniumprojects from the epidermal surface The neck invariablyconsists of four vertical rows of cells (centgure 2hi) Clearlythere are no parallels in development between the sunkenarchegonia of hornworts (centgure 2e) and similarembedded archegonia in pteridophytes (centgure 2 j)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 775
Phil Trans R Soc Lond B (2000)
hornworts
(a)
(c)
(b)
(d)
chamber roof
schizogenous space
antheridial initials
antheridial complex
pteridophytes
jacket
spermatogenous tissue
antheridium
Anthoceros
Ophioglossum
Figure 4 Comparison of antheridial development in hornworts (ab) and pteridophytes (cd) (a) In hornworts a periclinaldivision in a supercentcial cell delimits a subepidermal cell that gives rise to up to 25 antheridia and an epidermal cell that producesa chamber roof two cells thick A schizogenous space forms the cavity that houses the antheridia Antheridial development frominitials at the base of the cavity follows a developmental pathway similar to other bryophytes (b) Light micrograph of antheridialcomplex in Anthoceros agrestis (c) In pteridophytes a periclinal division in a supercentcial cell delimits a subepidermal cell that givesrise to the spermatogenous tissue and an epidermal cell that forms the sterile jacket (d) The single antheridium that developsfrom this process in Ophioglossum engelmannii
Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

4 REPRODUCTIVE GAMETOPHYTES
Selective pressures in a terrestrial environment forreproductive processes that require uninterrupted accessto water are indeed extreme The transmigration of algaeto land and the subsequent evolution of embryophytesnecessitated the evolution of multicellular sex organs(Niklas 1997) These organs not only served to protectvulnerable developing gametes but also the origin ofmulticellular female sex organs (archegonia) was a requi-site for embryo development a universal feature of landplants The evolution of gametangia in archegoniatesundoubtedly was a complex historical process thatrequired coordination of morphogenetic with physio-logical and ecological signals For example production ofsex organs must be timed appropriately and must occurrapidly in response to pounductuating seasonal conditionsGametangia must be strategically placed on the plant andprotective structures amply developed In gametophyte-dominant land-dwelling plants the compromise betweenoptimization of vegetative growth and the constraints ofsexual reproduction often resulted in the abandonment orreduced emphasis on genetic exchange (ie elaboration ofmechanisms for asexual reproduction and self-fertilization)Such strategies are also common in algal groups butevolved in response to diiexclerent selective pressures
In an attempt to understand adaptive strategies andcharacter evolution in the sexual phase of early arche-goniates we will briepoundy examine and compare (i) sexorgan ontogeny and (ii) spermatogenesis among basalbryophytes and seedless tracheophytes
Unlike those of pteridophytes gametangia of mossesand liverworts are stalked and extend from the epidermalsurface (Bold et al 1987) In leafy taxa gametangia areeither associated with leaves or leaves surround a terminal
774 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
hornworts
pteridophytes
(a)
( f ) (g) (h)
(b) (c) (d) (e)
(i) ( j )
mature neck
mature neck
neckventer
neck canal cellsventral canal cellegg
neck canal cellsventral canal cellegg
neck
Figure 2 Comparison of archegonial development in hornworts (a^e) and pteridophytes ( f^j ) (a) Longitudinal view and(b) cross-sectional view of archegonial initial traversed by three anticlinal walls that delimit a central axial cell and threeperipheral cells The axial cell gives rise to the neck canal cells ventral canal cell and egg while the peripheral cells form sterileouter layers (c) As seen in cross-section three additional segments in the peripheral cells form six rows of neck cells (d ) Lightmicrograph surface view of six rows of neck cells in Phaeoceros laevis (e) Longitudinal section of the sunken archegonium ofPhaeoceros laevis ( f ) Longitudinal view and (g) cross-sectional view of archegonial initial traversed by single periclinal wall thatdelimits an outer cell that forms the neck and an inner cell that gives rise to neck canal cells ventral canal cell and egg (h) Asseen in cross-section successive anticlinal divisions in the outer cell form four rows of neck cells (i) Scanning electron micrographof archegonial neck with four rows of cells in Psilotum nudum ( j ) Longitudinal section of sunken archegonium of Pteridiumaquilinum
Figure 3 Light micrograph of antheridium of Takakiaceratophylla Longitudinal section of anterior of developingelongated antheridium Cells are in packets arranged in tworows that represent the original biseriate centlament producedby left^right divisions from an apical cell The outline of theapical cell is visible as a triangular wall containing developingsperm cells at the antheridial tip Bar ˆ 100 m m
cluster of sex organs Abundant papillae hairs and para-physes are intermixed among gametangia and providefurther protection Thalloid liverworts have evolved avast array of structures and mechanisms for protectinggametangia including inrolled ventral branches (Metz-geria) secondarily sunken chambers (Pellia Noteroclada andcomplex thalloid liverworts) and lamellar outgrowths ofthe thallus (Renzaglia 1982) In contrast gametangia inhornworts are produced and maintained within theconcentnes of the thallus and as such supercentcially resemblethose of pteridophytes Gametangia of basal pterido-phytes are never stalked and are either entirely sunkenwithin the thallus or form conspicuous epidermal mounds(Bierhorst 1971)
To evaluate homology of gametangia it is critical toexamine cell division patterns and especially the initialformative division sequences during which the develop-mental fate of subsequent cell lineages is determined(Roux 1895 Niklas 1997) These primary divisions aregenetically controlled and provide the blueprint for organo-genesis At centrst glance there appears to be no similarityin archegonial ontogeny among liverworts hornworts andmosses Archegonial development in mosses uniquelyinvolves the initial production of an apical cell thatsegments left and right forming a biseriate centlament(Schocenteld 1985 Smith 1955) The archegonium properforms from the terminal cell in the centlament after theapical cell ceases its patterned mitotic divisions Hornwortand liverwort archegonia develop without elongation
from an apical cell In mosses and liverworts the arche-gonial initial elongates and divides above the epidermalsurface while in hornworts the developing archegoniumremains surrounded by thallus tissue This mosaic ofcharacters related to archegonial ontogeny providesevidence of divergent developmental pathways in thebryophyte clades However when the formative divisionsequences that establish the archegonium proper areexamined a fundamental pattern emerges that is diag-nostic of bryophytes (Schuster 1966) In all three groupsthe process involves three longitudinal divisions that forma central triangular axial cell surrounded by threeperipheral cells (centgure 2ab) The peripheral cells formthe neck and venter while the axial cell gives rise to theneck canal cells ventral canal cell and egg Further divi-sions in the peripheral cells typically result in a neck ofcentve or six cell rows (centgure 2cd) (Renzaglia 1982)
This division pattern is markedly diiexclerent from that ofpteridophytes in which the single epidermal initial centrstdivides transversely (periclinal division) The outer cellthen further divides to form the neck while the inner celldivides periclinally to produce neck canal cells ventralcanal cell and egg (centgure 2 fg j) (Bierhorst 1971 Kenrickamp Crane 1997a) With expansion the archegoniumprojects from the epidermal surface The neck invariablyconsists of four vertical rows of cells (centgure 2hi) Clearlythere are no parallels in development between the sunkenarchegonia of hornworts (centgure 2e) and similarembedded archegonia in pteridophytes (centgure 2 j)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 775
Phil Trans R Soc Lond B (2000)
hornworts
(a)
(c)
(b)
(d)
chamber roof
schizogenous space
antheridial initials
antheridial complex
pteridophytes
jacket
spermatogenous tissue
antheridium
Anthoceros
Ophioglossum
Figure 4 Comparison of antheridial development in hornworts (ab) and pteridophytes (cd) (a) In hornworts a periclinaldivision in a supercentcial cell delimits a subepidermal cell that gives rise to up to 25 antheridia and an epidermal cell that producesa chamber roof two cells thick A schizogenous space forms the cavity that houses the antheridia Antheridial development frominitials at the base of the cavity follows a developmental pathway similar to other bryophytes (b) Light micrograph of antheridialcomplex in Anthoceros agrestis (c) In pteridophytes a periclinal division in a supercentcial cell delimits a subepidermal cell that givesrise to the spermatogenous tissue and an epidermal cell that forms the sterile jacket (d) The single antheridium that developsfrom this process in Ophioglossum engelmannii
Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

cluster of sex organs Abundant papillae hairs and para-physes are intermixed among gametangia and providefurther protection Thalloid liverworts have evolved avast array of structures and mechanisms for protectinggametangia including inrolled ventral branches (Metz-geria) secondarily sunken chambers (Pellia Noteroclada andcomplex thalloid liverworts) and lamellar outgrowths ofthe thallus (Renzaglia 1982) In contrast gametangia inhornworts are produced and maintained within theconcentnes of the thallus and as such supercentcially resemblethose of pteridophytes Gametangia of basal pterido-phytes are never stalked and are either entirely sunkenwithin the thallus or form conspicuous epidermal mounds(Bierhorst 1971)
To evaluate homology of gametangia it is critical toexamine cell division patterns and especially the initialformative division sequences during which the develop-mental fate of subsequent cell lineages is determined(Roux 1895 Niklas 1997) These primary divisions aregenetically controlled and provide the blueprint for organo-genesis At centrst glance there appears to be no similarityin archegonial ontogeny among liverworts hornworts andmosses Archegonial development in mosses uniquelyinvolves the initial production of an apical cell thatsegments left and right forming a biseriate centlament(Schocenteld 1985 Smith 1955) The archegonium properforms from the terminal cell in the centlament after theapical cell ceases its patterned mitotic divisions Hornwortand liverwort archegonia develop without elongation
from an apical cell In mosses and liverworts the arche-gonial initial elongates and divides above the epidermalsurface while in hornworts the developing archegoniumremains surrounded by thallus tissue This mosaic ofcharacters related to archegonial ontogeny providesevidence of divergent developmental pathways in thebryophyte clades However when the formative divisionsequences that establish the archegonium proper areexamined a fundamental pattern emerges that is diag-nostic of bryophytes (Schuster 1966) In all three groupsthe process involves three longitudinal divisions that forma central triangular axial cell surrounded by threeperipheral cells (centgure 2ab) The peripheral cells formthe neck and venter while the axial cell gives rise to theneck canal cells ventral canal cell and egg Further divi-sions in the peripheral cells typically result in a neck ofcentve or six cell rows (centgure 2cd) (Renzaglia 1982)
This division pattern is markedly diiexclerent from that ofpteridophytes in which the single epidermal initial centrstdivides transversely (periclinal division) The outer cellthen further divides to form the neck while the inner celldivides periclinally to produce neck canal cells ventralcanal cell and egg (centgure 2 fg j) (Bierhorst 1971 Kenrickamp Crane 1997a) With expansion the archegoniumprojects from the epidermal surface The neck invariablyconsists of four vertical rows of cells (centgure 2hi) Clearlythere are no parallels in development between the sunkenarchegonia of hornworts (centgure 2e) and similarembedded archegonia in pteridophytes (centgure 2 j)
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 775
Phil Trans R Soc Lond B (2000)
hornworts
(a)
(c)
(b)
(d)
chamber roof
schizogenous space
antheridial initials
antheridial complex
pteridophytes
jacket
spermatogenous tissue
antheridium
Anthoceros
Ophioglossum
Figure 4 Comparison of antheridial development in hornworts (ab) and pteridophytes (cd) (a) In hornworts a periclinaldivision in a supercentcial cell delimits a subepidermal cell that gives rise to up to 25 antheridia and an epidermal cell that producesa chamber roof two cells thick A schizogenous space forms the cavity that houses the antheridia Antheridial development frominitials at the base of the cavity follows a developmental pathway similar to other bryophytes (b) Light micrograph of antheridialcomplex in Anthoceros agrestis (c) In pteridophytes a periclinal division in a supercentcial cell delimits a subepidermal cell that givesrise to the spermatogenous tissue and an epidermal cell that forms the sterile jacket (d) The single antheridium that developsfrom this process in Ophioglossum engelmannii
Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

Bryophyte antheridia contain well-developed stalksand globose to elongated antheridial bodies As in arche-gonial development antheridial ontogeny in mosses ispeculiar in that an apical cell is responsible for earlyelongation (Smith 1955) This feature alone ie antheri-dial development involving an apical cell enables theidenticentcation of Takakia as a moss (centgure 3) (Smith ampDavison 1993) The genesis of hornwort antheridia withinan internal thallus chamber has been cited as a funda-mental departure in morphogenetic design from all otherarchegoniates (Crandall-Stotler 1980 Bold et al 1987)However with closer scrutiny a single underlying patternof development is manifest in antheridial development ofmosses hornworts and liverworts In all three groups theantheridial initial elongates and similar division cyclesform either two primary spermatogones with foursurrounding jacket initials (mosses and leafysimple thal-loid liverworts) or four primary spermatogones with eightperipheral jacket initials (hornworts and complex thalloidliverworts) These similarities must represent plesio-morphies of the land plant clade The diiexclerence in horn-worts is that the antheridial initial is located at the baseof the schizogenous antheridial chamber and not at thethallus surface as in other bryophytes Apparently inhornworts an evolutionary shift in developmental poten-tial has occurred from epidermal (layer surrounding theexternal surface) to epithelial (layer surrounding aninternal space) cells both of which are surface cells thatenclose tissue The designation of hornwort antheridia asendogenous refers only to the location of developmentand not to an inherently diiexclerent developmentalpathway
An initial periclinal division in a supercentcial cell inhornworts mimics early antheridial development in pteri-dophytes (cf centgure 4ac) However unlike in pterido-phytes where this initial division gives rise to a singleantheridium (see below) in hornworts extensive celldivisions in the resulting two cells produce an entireantheridial complex consisting of a sunken chamber withoverlying two-layered roof and up to 25 enclosed anther-idia (centgure 4ab)
Antheridial development is essentially identical toarchegonial development in pteridophytes (centgure 4c)(Bierhorst 1971) The process begins with a transversedivision in an epidermal cell The outer cell gives rise to asterile jacket and the inner cell forms the spermatogenoustissue So similar are these division patterns in the twosex organs of pteridophytes that it is virtually impossibleto diiexclerentiate antheridia from archegonia in early stagesof organogenesis Moreover in protandrous lycophyteswhen the transition between male and female sex organsoccurs it is possible to centnd bisexual gametangia thatcontain developing sperm in the `neck regionrsquo and an eggcell at the base (centgure 5) (K S Renzaglia and D PWhittier unpublished data)
5 SPERMATOGENESIS
Sperm cells of archegoniates are propitiouslyconstructed for optimal swimming ecurrenciency in a terres-trial environment As the most complicated cellsproduced by archegoniates motile sperm cells provide awealth of developmental and phylogenetic data
(Renzaglia amp Duckett 1988 1991 Garbary et al 1993)During spermatogenesis undiiexclerentiated parenchymacells are progressively transformed into streamlinedcoiled cells containing minimal organelles The processinvolves the origin and development of an elaborate loco-motory apparatus a structure that enables unparalleledcomparisons with motile algal cells A unifying feature ofmotile gametes of archegoniates is the de novo origin ofcentrioles in late spermatogenous tissue (Vaughn ampHarper 1998 Vaughn amp Renzaglia 1998) In plants withbipoundagellated sperm cells centrioles originate as bicen-trioles structures that are composed of two centriolesattached end to end The bicentriole is the land plantanalogue of the orthogonal centriolar pair that is typicalof protists and animals In multipoundagellated gametescentrioles originate as more complicated spherical orga-nelles known as blepharoplasts In addition to origin anddevelopment of the locomotory apparatus histogenesis ofmale sex organs provides comparative data on cyto-plasmic phenomena such as plastid behaviour andcellular polarity Numbers of organelles and their spatialarrangements are established during the proliferativedivisions of organogenesis and the nascent spermatid isorganized for expeditious diiexclerentiation into a motile cell(Renzaglia amp Duckett 1987 Bernhard amp Renzaglia 1995Renzaglia et al 1994) The process of spermatogenesis and
776 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 5 Longitudinal section of Phylloglossum drummondiibisexual gametangium In the region of the maturegametophyte where antheridial production shifts toarchegonial production bisexual gametangia are commonThis gametangium has developing sperm cells (SC) in the`neck regionrsquo that overlie a mature egg cell (E) Bar ˆ 20 m m
its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

its bearing on plant phylogenetics will be reviewed else-where (Renzaglia amp Garbary 2000) Here we considergeneral features of the mature spermatozoid especially ofbipoundagellated cells and evaluate the selective processesthat have acted to bring about these phenotypes
Bipoundagellated sperm cells are produced by charophyceanalgae (except Zygnematales) bryophytes and most lyco-phytes (centgures 6 and 7) while all other tracheophyteswith motile sperm produce multipoundagellated cells (centgures8 and 9) The mature bipoundagellated cell in bryophytes is ahelical cylinder with an anteriorly positioned locomotory
apparatus and four organelles an anterior mitochon-drion a compacted central nucleus and a posteriorplastid with an associated mitochondrion (centgure 6b^d )In addition to poundagella and basal bodies the locomotoryapparatus consists of a lamellar strip and a narrow bandof microtubules (the so-called spline) which extendaround the cell providing a framework for positioning oforganelles The lamellar strip is composed of centrin acalcium-binding contractile protein and as such functionsas an MTOC for the spline microtubules and in posi-tioning the poundagella (Vaughn et al 1993) At cellular
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 777
Phil Trans R Soc Lond B (2000)
Figure 6 Reconstructions of mature bipoundagellated spermatozoids of Charales and bryophytes Note sinistral coiling of all butPhaeoceros (b) which exhibits a right-handed coil Colour coding red (pink) ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange ˆ lamellar strip green ˆ plastid (a) Chara vulgaris Adapted fromDuncan et al (1997) (b) Phaeoceros laevis Adapted from Carothers amp Duckett (1980) and Renzaglia amp Duckett (1989) (c) Blasiapusilla Adapted from Renzaglia amp Duckett (1987) (d) Aulacomnium palustre Data derived from Bernhard amp Renzaglia (1985)Drawing by H Dee Gates
maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

maturity the lamellar strip regresses to a dense rim alongthe leading edge of the cell (centgure 6b^d )
Among the algal outgroups to land plants only Char-ales possess coiled spermatozoids (centgure 6a) In male
gametes of Chara and Nitella the same basic organizationas in bryophytes is exhibited the primary diiexclerence is inthe number of organelles Approximately 30 mitochondriaare aligned at the cell anterior and up to six starch-centlled
778 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 7 Reconstructions of mature bipoundagellated spermatozoids of lycophytes Colour coding red (pink) ˆ poundagella and basalbodies blue ˆ nucleus brown ˆ mitochondria yellow ˆ spline microtubules purple ˆ lamellar strip green ˆ plastid grey ˆ extra-neous cytoplasm (a) Selaginella kraussiana Adapted from Renzaglia et al (1998) (b) Lycopodium obscurum Adapted from Maden etal (1996) (c) Palhinhaea cernua Adapted from Robbins amp Carothers (1978) (d) Lycopodiella lateralis Adapted from Maden et al(1997)
plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

plastids are associated with scattered mitochondria at thecell terminus Gametes of Chara never possess a lamellarstrip but those of Nitella and Coleochaete possess thiscomponent of the locomotory apparatus In contrast tobryophytes in which the plates of the lamellar strip areoriented at 458 to the longitudinal axis of spline micro-tubules in algal cells this angle is consistently 908(Graham 1993)
Mature spermatozoids of hornworts mosses and liver-worts are immediately distinguished from each other bymajor architectural diiexclerences Hornwort sperm cells areextremely small (ca 30 m m in diameter) and bilaterallysymmetrical (centgure 6b) The two basal bodies insert theparallel poundagella into the cell anterior at approximately thesame level Unlike spermatozoids of all other archegoni-ates which are sinistrally coiled hornwort gametes exhibita dextral coil In contrast the bilateral spermatozoids ofliverworts and mosses are asymmetrical (centgure 6cd) Inboth groups of bryophytes the two basal bodies are mark-edly diiexclerent in length structure and insertion into thecell As a consequence the poundagella are staggered in theiremergence from the cell When viewed from the cell ante-rior the basal body that is positioned further forwards issituated on the right and the more posterior basal body ison the left side of the cell The anterior basal body in bothmosses and liverworts contains dorsal microtubule tripletsthat extend in front of the basal body proper Similarly inboth groups the posterior basal body exhibits a unique yetconsistent structure with ventral microtubule triplet exten-sions accounting for most of its length The strikingcommonalities in construction of this complex locomotoryapparatus including basal bodies and poundagella stronglysupport monophyly of liverworts and mosses (Renzaglia ampDuckett 1991 Garbary et al 1993) The primary diiexclerencesin moss compared with liverwort sperm cells are theoccurrence of a stray microtubule and the position ofthe plastid and associated posterior mitochondrion alongthe inner nuclear surface (centgure 6d) and not at the cellterminus as in hepatics (centgure 6c)
Lycophyte spermatozoids are structurally diverseprobably more so than in any other land plant group(centgures 7 and 8) (Maden et al 1996 1997 Renzaglia et al1998 1999) Within Lycopodiaceae gametes may be bi-poundagellated and coiled (centgure 7b) multipoundagellated andcoiled (centgure 8a) or bipoundagellated with a more ovoidoutline (centgure 7cd) The lamellar strip persists in themature gamete and the angle between lamellar strip andspline ranges from 458 to 908 (Maden et al 1996 1997)Compared with bryophyte spermatozoids those of lycop-sids contain more cytoplasm including numerous smallmitochondria at the cell posterior Plastid number variesfrom one to centve Plastids are either positioned on theinner nuclear surface as in the more coiled gametes(centgure 7b) or aggregated at the cell posterior in ovoidcells (centgure 7cd) Apparent evolutionary trends withinthe family include an increase in cell length and diameterand a decrease in coiling (Maden et al 1997) Flagella areslightly staggered in the smaller coiled sperm cells ofHuperzia and Lycopodium and more widely separated in themore specialized ovoid cells of Palhinhaea and LycopodiellaThese ideas are contrary to those of Robbins amp Carothers(1978) who speculated that an ovoid cell with widelyspaced poundagella is the primitive condition in archegoniates
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 779
Phil Trans R Soc Lond B (2000)
Figure 8 Transmission electron micrographs of multi-poundagellated male gametes of lycophytes (a) Phylloglossumdrummondii This spermatozoid contains approximately 20poundagella (F) aligned in a coil over the multilayered structure(MLS) at the cell anterior The nucleus (N) is broad slightlycoiled and contains uncondensed inclusions Numerous mito-chondria (MI) and a plastid (P) with large starch grains arepositioned at the rear of the cell Bar ˆ 10 m m (b) Isoelaquo tesbolanderi Approximately 20 poundagella (F) are distributed alongthe length of this narrow and coiled spermatozoid Thenucleus (N) is a compacted narrow cylinder and it overlapswith two mitochondria (MI) An inconspicuous posteriorplastid (P) lacks starch Upon motility the large centralmass of cytoplasm (CM) is shed Bar ˆ 10 m m Unpublishedmicrograph by Gayleen Cochran
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)
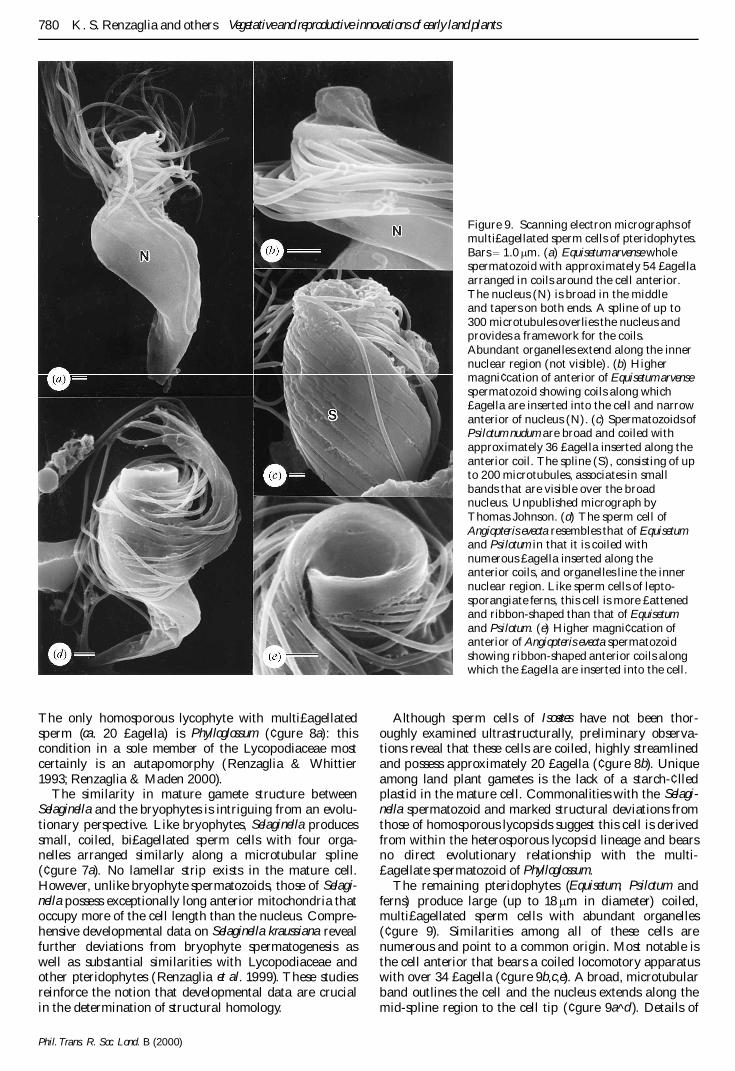
The only homosporous lycophyte with multipoundagellatedsperm (ca 20 poundagella) is Phylloglossum (centgure 8a) thiscondition in a sole member of the Lycopodiaceae mostcertainly is an autapomorphy (Renzaglia amp Whittier1993 Renzaglia amp Maden 2000)
The similarity in mature gamete structure betweenSelaginella and the bryophytes is intriguing from an evolu-tionary perspective Like bryophytes Selaginella producessmall coiled bipoundagellated sperm cells with four orga-nelles arranged similarly along a microtubular spline(centgure 7a) No lamellar strip exists in the mature cellHowever unlike bryophyte spermatozoids those of Selagi-nella possess exceptionally long anterior mitochondria thatoccupy more of the cell length than the nucleus Compre-hensive developmental data on Selaginella kraussiana revealfurther deviations from bryophyte spermatogenesis aswell as substantial similarities with Lycopodiaceae andother pteridophytes (Renzaglia et al 1999) These studiesreinforce the notion that developmental data are crucialin the determination of structural homology
Although sperm cells of Isoelaquo tes have not been thor-oughly examined ultrastructurally preliminary observa-tions reveal that these cells are coiled highly streamlinedand possess approximately 20 poundagella (centgure 8b) Uniqueamong land plant gametes is the lack of a starch-centlledplastid in the mature cell Commonalities with the Selagi-nella spermatozoid and marked structural deviations fromthose of homosporous lycopsids suggest this cell is derivedfrom within the heterosporous lycopsid lineage and bearsno direct evolutionary relationship with the multi-poundagellate spermatozoid of Phylloglossum
The remaining pteridophytes (Equisetum Psilotum andferns) produce large (up to 18 m m in diameter) coiledmultipoundagellated sperm cells with abundant organelles(centgure 9) Similarities among all of these cells arenumerous and point to a common origin Most notable isthe cell anterior that bears a coiled locomotory apparatuswith over 34 poundagella (centgure 9bce) A broad microtubularband outlines the cell and the nucleus extends along themid-spline region to the cell tip (centgure 9a^d ) Details of
780 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 9 Scanning electron micrographs ofmultipoundagellated sperm cells of pteridophytesBars ˆ 10 m m (a) Equisetum arvense wholespermatozoid with approximately 54 poundagellaarranged in coils around the cell anteriorThe nucleus (N) is broad in the middleand tapers on both ends A spline of up to300 microtubules overlies the nucleus andprovides a framework for the coilsAbundant organelles extend along the innernuclear region (not visible) (b) Highermagnicentcation of anterior of Equisetum arvensespermatozoid showing coils along whichpoundagella are inserted into the cell and narrowanterior of nucleus (N) (c) Spermatozoids ofPsilotum nudum are broad and coiled withapproximately 36 poundagella inserted along theanterior coil The spline (S) consisting of upto 200 microtubules associates in smallbands that are visible over the broadnucleus Unpublished micrograph byThomas Johnson (d) The sperm cell ofAngiopteris evecta resembles that of Equisetumand Psilotum in that it is coiled withnumerous poundagella inserted along theanterior coils and organelles line the innernuclear region Like sperm cells of lepto-sporangiate ferns this cell is more poundattenedand ribbon-shaped than that of Equisetumand Psilotum (e) Higher magnicentcation ofanterior of Angiopteris evecta spermatozoidshowing ribbon-shaped anterior coils alongwhich the poundagella are inserted into the cell
spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

spermatozoid structure in these plants will be presentedelsewhere (Renzaglia amp Garbary 2000)
All spermatozoids of land plants exhibit coiling eitherexternal in cell architecture or internal in arrangement oforganelles Coiling is the direct result of cellular elongationassociated with extension of the single band of spline micro-tubules within the constraints of a spherical cell iecompaction and elongation necessarily parallel the circularboundaries of the cellThe complicated layoutof land plantsperm cells most certainly provides a hydrodynamicallysound architecture Cellular elongation or streamliningreduces excess baggage and perhaps even more impor-tantly is instrumental in movement of the spermatozoidthrough the narrow tube of the archegonial neck Theconvergence in structure of archegonia and the egg appa-ratus of Charales may explain similarities in sperm cellshape between Charales and basal land plants and mayaccount for the marked diiexclerences between these cellsand those of Coleochaetales
In bryophytes and perhaps some lycophytes spermcells have remained relatively small and simple The lowhaploid DNA contents of these plants are correlated withsperm size and are repoundected in small cell size (Renzagliaet al 1995) Among bryophytes hornworts have the smal-lest genome size and the smallest sperm cell Thesymmetric placement of poundagella at the anterior is aneiexclective organization to move an extremely small cellthrough water With greater DNA content and increase incell size it would follow that asymmetry and staggeringof poundagellar insertion may be necessary to facilitate move-ment Data from sperm cell structure suggest that horn-worts diverged from other bryophytes before poundagellarstaggering evolved and that mosses and liverwortsacquired staggering from a common ancestor with largerspermatozoids than those of the hornwort^embryophyteprogenitor If cells are symmetrical as in hornworts thedirection of coiling aiexclords no selective advantage and isfree to change without consequence to swimming perform-ance Once asymmetry was established as in all otherland plants the diiexclerent constructions of the right andleft cellular halves may have had `locked inrsquo genetic deter-mination of coiling direction Based on these ideas theprototypic spermatozoid of archegoniates was probably aminute bipoundagellated cell with minimal sinistral coilingObservations of remarkably well-preserved sperm cells inDevonian gametophytes strengthen the speculation thatprimitive land plants produced small coiled gametes(Remy et al 1993 Duncan et al 1997)
6 SPOROPHYTES
Liverworts hornworts and mosses are readily distin-guished from polysporangiate tracheophytes by theirunbranched monosporangiate sporophytes that are nutri-tionally dependent on the persistent photosyntheticgametophyte Given the likely origin of the sporophytethrough a delay in meiosis (ie antithetic theory) thelack of elaboration of the sporophyte in bryophytes isviewed as a plesiomorphy among embryophytes Indeedcontemporary evidence both molecular and morpho-logical supports the basal position of bryophytes in landplant phylogeny (Graham 1993) However re-evaluationof sporophyte structure suggests that the interpretation of
bryophyte sporophytes as less complex than those of pter-idophytes is an oversimplicentcation In pteridophytes inaddition to facilitating spore production and dispersalthe sporophyte is responsible for vegetative growthincluding maximum light harvesting and mechanicalsupport In bryophytes where the gametophyte is thevegetative phase the sole function of the sporophyte is toproduce and disperse spores In eiexclect each bryophytesporophyte is a solitary sporangium with or without astalk that is designed to make and release spores and assuch has undergone virtually no selection for vegetativegrowth It is not surprising then that bryophyte sporo-phytes although they are unbranched and lack leavesproduce sporangia that are the most complicated of anyproduced by land plants Lines of specialization have ledto elaborate spore production and dispersal mechanismssuch as the peristome of true mosses cellular elaters ofliverworts and the highly complicated sporangium ofhornworts that continues to elongate throughout thegrowing season No parallels of such intricate sporangialcomplexity are found in any tracheophyte for indeedthese roles have been taken over by the vegetative sporo-phytic tissues not the sporangium itself
This new look at sporophyte complexity in bryophytesmay be extended further in an evaluation of reproductivepotential in sporophytes of embryophytes Moss liver-wort and hornwort sporophytes are limited in theircapacity for spore production because individually theybear only a single sporangium However when thetotality of sporophytes that are potentially produced by asingle gametophyte is considered a very diiexclerent inter-pretation emerges During their lifetime and even withina single growing season the typical sexual bryophytegametophyte generates a multitude of sporophytes Ingametophytes of `promiscuousrsquo bryophytes it is con-ceivable that the total number of spores produced by the`populationrsquo of sporophytes on a single gametophyte mayexceed that of a single pteridophyte sporophyte during agrowing season Moreover unlike the multiple sporangiaof a pteridophyte sporophyte (generated by a singlegametophyte) the numerous sporophytes produced by asolitary bryophyte gametophyte are potentially productsof independent fertilizations Given outcrossing eachsporophyte is genetically unique In bryophytes evolu-tion has maximized both sexual reproduction and theconsequential production of genetically diversesporangia while in tracheophytes vegetative growth isresponsible for increased development of geneticallysimilar sporangia
Among tracheophytes each gametophyte of hetero-sporous forms has the potential to produce only a solitarysporophyte Basal homosporous tracheophytes in con-trast often have the capacity to produce several sporo-phytes and longer-lived subterranean gametophytes mayyield multiple sporophytes (Bierhorst 1971 A R Madenunpublished data) Subterranean gametophytes are dicurren-cult to centnd in nature and therefore they have been in-sucurrenciently examined As a result the extent ofsporophyte production by a single gametophyte in basalpteridophytes remains unknown Nevertheless it can beconcluded that the ability to generate multiple sporo-phytes signicentcantly increases reproductive potential inland plants with persistent gametophytes
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 781
Phil Trans R Soc Lond B (2000)
Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

Because selection for individual sporophytes of bryo-phytes has acted to enhance the manufacture anddispersal of spores it is not surprising that a number ofdiverse mechanisms for elevating the sporangium haveevolved To expose spores to sucurrencient wind currentssporangia must be raised above the gametophyte surfaceSome of these strategies for sporangial elevation involveextension of gametophytic tissue (archegoniophore inMarchantiales and pseudopodium in Sphagnum andAndreaeales) while most involve growth of sporophytictissue In those with gametophytic elevation of sporangiathe seta remains short and the sporangium relativelysimple eg no conducting tissue is present in Sphagnum orAndreaeales Elaboration of sporophytic tissue entailseither the production of a seta (true mosses Takakia leafyand simple thalloid hepatics) or the development of abasal meristem (hornworts)
In hepatics seta extension is rapid and involves theextension of turgid cells Mechanical strength is providedby hydrostatic pressure against the cell wall from withinthe large central vacuole Elongation of the archegonio-phore and development of protective structures aroundthe developing sporophyte also entail rapid wall expand-ability and extension of ephemeral organs an innovationthat is common in a variety of hepatic tissues Thestrategy in these plants is for `instantaneousrsquo and eiexclectivedispersal of spores This process is facilitated by elatersspecialized elongated sterile cells with spiralled wallthickenings With an ephemeral elongated mature sporo-phyte it follows that there would be no selection forelaboration of conducting tissue photosynthetic regionsand stomata in liverworts Such tissues and cells are adap-tations benecentcial to sporophytes such as those of mossesand hornworts where sporophyte extension is gradual andbrought about by cell division followed by gradual elon-gation Mechanical strength and movement of materialsespecially upwards to the region where spores diiexcler-entiate are crucial to the success of sporophytes thatpersist through the growing season and disperse sporesover an extended period Hence selective pressures in atleast some of these lineages have favoured the retention ofsporophytic innovations related to increased strength(cells with thickened walls) photosynthetic function(stomata and assimilative regions) and enhanced trans-port of nutrients (conducting tissue) Sporophytes of hepa-tics require a comparable period of time to mature butthis process occurs within the concentnes of protectivegametophytic tissues which provide a continuous supply ofwater and nutrients
Not insignicentcant in eiexclecting spore dispersal is the factthat many bryophytes grow on vertical substrates In suchsituations the entire plant is elevated above ground leveland is exposed to wind currents that facilitate maximumspore dispersal Sporophytes of epiphytic mosses forexample are often less elongated than those of grounddwelling taxa Thus location of habitat must be factoredinto the equation for successful spore dissemination
Diiexclerent strategies of sporophyte elevation are comple-mented by divergent morphogenetic designs in embryoand sporophyte growth and diiexclerentiation Salientfeatures of sporophyte development in bryophyte lineagesare considered below The hornwort zygote is dissectedcentrst by a vertical wall not a transverse partition as in
mosses and liverworts The endothecium in hornwortsand Sphagnum forms a columella only and is not responsiblefor producing sporogenous tissue as in other bryophytesAs in gametangial ontogeny apical cell involvement isechoed in the formative phase of embryogenesis in mossesLater in development before diiexclerentiation of the capsulean intercalary meristem typically forms and brings aboutfurther elongation of the sporophyte (Roth 1969) In bothmosses and hornworts photosynthetic capability in thesporophyte is partially responsible for continued growthAn assimilative layer extends the length of the hornwortsporophyte In mosses the photosynthetic region extendsalong the entire capsule and includes internal specializa-tion of aerenchyma In contrast the hepatic sporophyte iscontained within protective gametophytic cells until sporeshave matured Thus gametophytic nurturing occursthrough the life of the hepatic sporophyte
Taken as a whole the fundamental diiexclerences indevelopment structure and spore dispersal strategiessuggest divergence of the three groups of bryophytes priorto specialization of mechanisms for elongation andcontinued growth With this scenario the prototypicbryophyte sporophyte would be a small mass of cells witha fertile internal region as predicted by Graham (1993)Mishler amp Churchill (1984) and Hemsley (1994) Becausea placenta with specialized cells at the sporophytegametophyte interface is ubiquitous in embryophytes(Ligrone et al 1993) this specialized absorptive regionwas probably in place in the early stages of sporophytediversicentcation The occurrence of transfer cells in game-tophyte tissue surrounding the zygote of Coleochaetesupports this supposition Thus certain Charophyceaewere preadapted for embryo evolution (Graham 1993) Asan unelongated mass of cells the ancestral sporophytewould not have acquired structures necessary forsustained growth eg localized meristems photosyntheticzones conducting tissue and stomata In particularstomata assimilative regions and conducting tissues areadaptive specializations that enable persistence of an elon-gated sporophyte through the growing season Thereforethe homology of such structures among basal land plantsrequires evaluation
Stomata occur in Sphagnum and the true moss lineage(but not basal taxa such as Takakia and Andreaeales) andin two of the centve widely recognized genera of hornwortsIt should be noted that Megaceros the hornwort genuswith the largest sporophytes does not possess stomata Inmany mosses instead of distinct guard cells stomateontogeny uniquely produces a binucleate cell with acentral pore formed by incomplete septum formationduring cytokinesis (Sack amp Paollilo 1985) Separation ofadjacent cells along the middle lamella is commonplace inhornwort gametophytes It would follow then that thegenetic potential for producing two adjacent cells thatseparate from each other ie stomata is inherent in theseplants Since this innovation is restricted in occurrence toonly one lineage of anthocerotes (Phaeoceros^Anthoceros)stomata probably evolved independently in mosses andhornworts This interpretation is reasonable given thesimplicity of stomatal design and the apparent lack ofdiurnal cycles in bryophyte stomata (Paton amp Pearce1957) Certainly much more complicated structures andprocesses have evolved multiple times in embryophytes
782 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

secondary tissues and heterospory are two excellentexamples (Bateman et al 1998 Niklas 1997) Bateman ampDiMichele (1994) concluded that heterospory evolved aminimum of 11 times during embryophyte history
Among bryophytes conducting tissues are relativelywidespread in gametophytes but restricted in distributionwithin sporophytes to mosses a feature that suggests acurren-nities with tracheophytes However because of develop-mental and ultrastructural divergences contemporaryinvestigations have called into question the homology ofmoss and tracheophyte conducting cells and have resultedin the rejection of this presumption (see Ligrone et althis issue and references therein) Most features of sievecells the food-transporting cells of pteridophytes whichare shared with leptoids the food-conducting cells ofmosses are found exclusively in the Polytrichales a rela-tively derived moss taxon The fundamental cytologicaldesign involving an endoplasmic microtubule system isshared by mosses and liverworts and is absent in tracheo-phytes Similarly the concept of homology betweenhydroids water-conducting cells of mosses and tracheidswater-conducting cells of tracheophytes has been severelychallenged Basal moss clades except Takakia are devoidof conducting cells and those of Takakia more closelyresemble water-conducting cells of the liverwort Haplomi-trium than they do those of the true moss lineage Clearlyconducting-cell homology among bryophytes andtracheophytes requires further evaluation and can nolonger be regarded as a working precept
The primitive method of sporangial dehiscence is theformation of a single suture Among archegoniates thiscondition is present in all basal clades The process mayinvolve a specialized line of cells as in Haplomitrium(Bartholomew-Began 1991) or it may occur betweenseemingly undiiexclerentiated cells as in Takakia and Huperzia(Renzaglia et al 1997 A R Maden unpublished data)Hornwort sporophytes typically dehisce along two sutureson either side of the sporangium while capsules of moreadvanced liverworts split into four valves and those ofmosses produce an operculum with an underlying elaborateperistome
Spores and sporogenesis are fundamentally conservedin embryophytes and provide a wealth of phylogeneticallyinformative characters (Brown amp Lemmon 1988 1990bGarbary amp Renzaglia 1998) For example monoplastidicmeiosis is an ancestral condition that characterizes Coleo-chaetales hornworts mosses and primarily basal taxa ofliverworts lycophytes and ferns (Renzaglia et al 1994Brown amp Lemmon 1997) Associated with monoplastidyin archegoniates but not in green algae is a unique quad-ripolar microtubule system (QMS) that is organized atthe plastids and predicts the polarity of the two meioticdivisions (Brown amp Lemon 1997) The occurrence of aQMS in both monoplastidic and more-derived polyplas-tidic meiocytes of liverworts and the putative loss of thismicrotubule assemblage in tracheophytes provides a well-decentned cytomorphogenetic transformation series in landplants (Brown amp Lemmon 1997)
The spore wall (sporoderm) of embryophytes showscontinuity in structure typically with a pecto-cellulosicinner layer (intine) and an outer sculptured layer (exine)which is impregnated with sporopollenin (Blackmore ampBarnes 1987) As a protective coating around the single cell
contained within the sporoderm was a necessary adapta-tion for survival and dispersal of airborne spores Intineand exine are derived from the cytoplasm and aredeposited in a centripetal fashion Tripartite lamellae aremajor components of the exine in most mosses liverwortsand tracheophytes but apparently are lacking in horn-worts and Andreaeales (Brown amp Lemmon 1988 Renza-glia amp Vaughn 2000) A primexine is laid down duringmeiotic cytokinesis in liverworts and hornworts and thislayer provides a template within which wall layers aresuccessively deposited Premeiotic patterning of wall sculp-turing is restricted to certain liverworts (HaplomitriumPallavicinia and Nowellia) (Brown et al 1986) with a morecomprehensive survey this feature may be informative indecentning clades within hepatics Wall-sculpturing patternsinvolving deposition of a perine from extrasporal tissue areshared by mosses and pteridophytes (Blackmore amp Barnes1987) A true perine is lacking in hornworts and liverwortswhere outer exine layers are responsible for wall ornamen-tation In hornworts a dense layer is deposited late in devel-opment (Renzaglia amp Vaughn 2000)This `pseudoperinersquo isdiiexclerentiated from true perine because it is derived fromthe spore mother cell wall and not the tapetum
Elaters consisting of entire cells are restricted to liver-worts and hornworts These cellular entities are theproduct of controlled mitotic divisions and thus the ratioof spores to elaters often is centxed within a taxon Ingeneral the ratio of spores to elaters is greater than fourto one in hepatics mitotic divisions occur in sporogenouscells after the elater mother cell is delimited In compar-ison elaters of hornworts are laid down in tiers that alter-nate with spore-generating layers Typically four sporesare produced per elater but in some instances mitoticdivisions in the elater mother cell decrease the spore-to-elater ratio (Schuster 1984c) The lack of specialized wallthickenings in most taxa and the tendency for adjacentelaters to adhere to one another has led to the conceptthat the `pseudoelaterrsquo of hornworts is multicellular Thecurious occurrence of elaters with spiralled wall thicken-ings in Megaceros and Dendroceros suggests homology ofthese cells with morphologically similar elaters of hepa-tics However because of the variability in elater struc-ture in hornworts and the existence of spiralled elaters inpresumably derived taxa (Megaceros and Dendroceros) theresemblance of elaters of certain hornworts and liverwortsis most realistically interpreted as a convergence Furtherdevelopmental studies are required to test this hypothesis
7 PHYLOGENY INTERPRETED FROM MALE
GAMETOGENESIS
A comprehensive data set on spermatogenesis was devel-oped to address relationships among pteridophytes andbryophytes (Garbary et al 1993) The original data setconsisted of 90 characters Accumulation of additionaldevelopmental and experimental data from other organ-isms has resulted in a deeper understanding of characterhomology and the original 90-character data set has nowbeen reduced to 72 more informative characters Thusdata from male gametogenesis may provide the single bestset of morphological characters in terms of homologyRecent eiexclorts to increase taxon sampling and to completedata on critical pteridophyte and bryophyte genera have
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 783
Phil Trans R Soc Lond B (2000)
enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

enabled greater resolution of relationships especiallywithin lycophytes Of special importance were the archi-tectural and developmental data for Selaginella kraussiana(Renzaglia et al 1998) and S australiensis (Renzaglia et al1999) previous data were derived mainly from the maturesperm structure (Robert 1974) The 1993 analysis resultedin a highly intuitive cladogram with bryophytes monophy-letic and with mosses and liverworts forming a singleclade The only apparent anomaly was the placement ofSelaginella on the bryophyte rather than the tracheophyteclade Subsequent cladistic analyses of this sperm data setresolve the anomalous position of Selaginella (Maden et al1997 this paper) and place it with other lycophytes
The current maximum parsimony (MP) analysis of 21archegoniates using 72 characters produces three equallyparsimonious trees that have the same basic topology(centgure 10) as those published previously (except for themore appropriate position here of Selaginella with thelycophytes) in which there is a dichotomy between bryo-phytes and tracheophytes The monophyly of bryophytesreceives weak bootstrap (BS) support (68 BS decayindex of 2) whereas the tracheophyte clade is stronglysupported (97 BS decay index of 5) Within bryophytesall three groups are monophyletic and the moss and horn-wort clades are strongly supported There is only weaksupport for the moss plus liverwort clade (BS 57 decayindex of 1) however three fundamental characterssupport this clade ie dimorphic basal body structurethe presence of a spline aperture and the location of thestellate pattern in the poundagellum
Lycophytes form a clade sister to a monophyletic assem-blage containing ferns Equisetum Psilotum and seed plantsThe clade of heterosporous taxa (Isoelaquo tes and Selaginella) isstrongly supported (BS 94 decay index of 4) whereasits sister group containing the remaining homosporouslycophytes receives weaker support (BS 60 decayindex of 1) This analysis of lycophytes is congruent with
those based on rbcL (WikstrIcircm amp Kenrick 1997 Korall etal 1999) except for the placement of Phylloglossum in aclade with Lycopodium not Huperzia However relation-ships among these three homosporous taxa are poorlyresolved based on spermatogenesis Because there is enor-mous variation in male gamete structure within lyco-phytes it would be useful to expand the data set toinclude all generic segregates of lycophytes (Ogravellgaard1987) and to further sample subgenera within Selaginella
The placement of Equisetum in the fern lineage is contro-versial (Kenrick amp Crane 1997ab) however the centrstunequivocal evidence that supported the hypothesis thathorsetails are reduced ferns came from male gametogen-esis characters (Bierhorst 1971) This hypothesis later foundsupport in an analysis combining rbcL and morphologicalcharacters (Pryer et al 1995) as well as an analysis ofmtSSU rDNA sequences (Duiexcl amp Nickrent 1999) A sisterrelationship between Psilotum and eusporangiate ferns issupported by a number of molecular studies (Manhart1995 Pryer et al 1995 Pahnke et al 1996 Malek et al 1996Wolf 1997 Wolf et al 1998 Hedderson et al 1998)
Manual adjustments on tree topology were conductedusing MacClade (Maddison amp Maddison 1992) to studythe eiexclect on tree length (TL) Only two other hypotheseswere reasonably consistent with these dataTree length wasonly two steps longer (TL ˆ 166) when centgure 10 wasconstrained to match the results from the overall morpho-logical data set (centgure 11) and the molecular data set(centgure 12) both of which resolve hornworts as sister to allother embryophytes The embryophyte clade minus horn-worts is supported by a character associated with the stag-gering of the basal bodies a fundamental feature of motilecell organization (Renzaglia amp Duckett 1991 see discus-sion in frac12 5) An additional single step (ie to TL ˆ 167) wasrequired to make mosses sister to tracheophytes whileretaining hornworts as the basal lineage The mosstracheophyte clade is supported by two characters the centrst
784 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 10 One of three trees of length 164(CI ˆ 078 RI ˆ 088) obtained from a maximumparsimony analysis (implemented with PAUP 40)of characters derived from spermatogenesisNumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch
involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

involves spline growth (in association with nuclearshaping) and the second involves the direction of nuclearshaping Other tree topologies had even less support andwhen the liverworts were placed as the outgroup to theremaining land plants (ie the Mishler amp Churchill (1984)hypothesis) the cladogram was centve steps longer than themost parsimonious tree (TL ˆ 169) For this arrangementthere were no characters that supported the cladeconsisting of all embryophytes minus liverworts
The lack of congruence between the tree topologybased on spermatogenesis (centgure 10) and that based onoverall morphology (centgure 11) may be due to inherentproblems in the spermatogenesis data set First becauseof convergences in cell architecture between spermato-zoids of Charales and archegoniates Chara may not be anideal choice for an outgroup However spermatogenesisin Coleochaete another potential charophyte outgroup isinsucurrenciently known hence many characters cannot becoded for this genus Moreover based on the limited dataavailable spermatozoids of Coleochaete lack the presumedancestral features of archegoniate sperm cells Forexample no coiling has been described in coleochaeta-lean spermatozoids One could argue that the selectivepressures for motile cells of land plants are decidedlydiiexclerent from those confronting aquatic algae and thusevolution of these cells in archegoniates was rapid anddirectional Moreover multicellular gametangia and theproblems related to negotiating passage through anarchegonial neck provided additional selective forces Asnoted above similarities in architecture of charalean andbryophyte spermatozoids may repoundect similar structure ofthe oogonium with its crown cells and the archegoniumwith its neck cells and the common constraints of move-ment through a narrow channel
Given the results of our successive MP analyses of thespermatogenesis data set we regard the occurrence ofbryophyte monophyly as an artefact However we areconvinced that more detailed analyses of sperm cell ultra-structure in a wide range of charophycean algae will
provide abundant data for a robust reanalysis and willenable recentned evaluation of preadaptations in these cellsthat led to the architecture of sperm cells of basalembryophytes
Our MP analyses have dealt only with land plants withpoundagellated sperm The comparison of poundagellated and non-poundagellated sperm by Southworth amp Cresti (1997) suggeststhat analogous features may be present in non-poundagellatedseed plants and that there are some remnants of structuralhomology between the two architectures However theremay be too few characters on which to attempt a phylo-genetic analysis across the transition between these spermtypes The absence of comparable studies of spermato-genesis in gymnosperms and basal angiosperms is also animpediment to a critical analysis of non-poundagellated spermevolution
8 PHYLOGENY INTERPRETED FROM
MORPHOLOGY AND DEVELOPMENT
A comprehensive data set incorporating morphologicaldevelopmental and ultrastructural features was compiledto approach questions relating to the position of the threebryophyte groups within the global phylogeny of plants(Garbary amp Renzaglia 1998) Specicentc characters to resolverelationships among groups of tracheophytes generallywere not included Of the 125 characters 62 were codedfor the gametophyte generation and 63 were coded for thesporophyte generationTo curtail overweighting of charac-ters related to spermatogenesis only 11 informativefeatures of male gametogenesis were included To helpensure character homology features of gametophytes wereconsidered independent of sporophytes
The analysis presented here of 22 ingroup taxa repre-sents a slightly revised character and organism matrixrelative to Garbary amp Renzaglia (1998) A single mostparsimonious tree results from MP analysis of themorphological data (centgure 11) This tree is highly resolvedas shown by the fact that 13 of the 20 total ingroup clades
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 785
Phil Trans R Soc Lond B (2000)
Figure 11 The single most parsimonious tree oflength 264 (CI ˆ 070 RI ˆ 084) obtained froma maximum parsimony analysis (implementedwith PAUP 40) of 125 developmental andmorphological characters Numbers above eachbranch represent the number of characterssupporting the branch Bootstrap percentagevalues (100 replications) followed by decay indicesare given below the branch
receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

receive BS support of 70 or greater Strong BS supportexists for the basal position of hornworts the monophylyof hornworts mosses and liverworts the mossliverwortclade tracheophytes and ferns Among these primaryclades only the separation of lycophytes from othertracheophytes received weak support (51 BS decayindex of 1) The remaining weakly supported nodes existmainly within the moss and liverwort clades Despitediiexclerent taxon sampling this phylogram is generallycongruent with the analysis of spermatogenous characters(centgure 10) with the following three exceptions (i) theposition of hornworts as sister to the remaining landplants (ii) the relative positions of Psilotum and Equisetumand (iii) the resolution of relationships within the mossclade The two primary features of this cladogram are theposition of hornworts basal to all other land plants andthe position of mosses and liverworts as monophyleticand sister to tracheophytes
The characters supporting the individual clades arelargely discussed by Garbary amp Renzaglia (1998) andwill not be repeated here The primary branching withinland plants (ie embryophytes minus hornworts) issupported by four synapomorphies (i) the change from adorsiventral to an axial gametophyte (ii) the presence ofpoundagellar staggering (iii) the presence of poundavonoids and(iv) the occurrence of grana end membranes The posi-tions of Megaceros and Takakia are of interest as these taxalack stomata and they are the apparent basal clades ofboth hornworts and mosses respectively This providesstrong support for our contention that stomata may haveevolved several times within land plants A similar treetopology is supported by our molecular analysis (below)with respect to the position of Takakia but not the rela-tionships within hornworts
Manipulations of the cladogram in centgure 11 usingMacClade do not provide even weak support for any
other phylogenetic hypotheses Thus forcing bryophytemonophyly (ie the spermatogenesis hypothesis) producesa cladogram three steps longer (TL ˆ 267) makingmosses sister to tracheophytes adds four steps (TL ˆ 268)reproducing the Mishler amp Churchill (1984) hypothesis(ie liverworts sister to remaining embryophytes) wasseven steps longer (TL ˆ 271) Thus the total morphologytree supports the primary features of the tree fromspermatogenesis Based on the nature of the characterssupporting the primary clades we consider the cladogramin centgure 11 to be an accurate representation of relation-ships at the base of the land plant clade It is alsocongruent with previously published trees based on mole-cular data (see frac12 9)
9 PHYLOGENY INTERPRETED FROM NUCLEAR
AND MITOCHONDRIAL SSU rDNA
To test hypotheses based on morphology and to eval-uate morphological character evolution we conductedanalyses of 22 land plants using a data set combiningsequences from nuclear and mitochondrial SSU rDNAThis MP analysis includes previously published sequencesin addition to new sequences including Psilotum Megacerosaenigmaticus Polytrichum and Cycas for mitochondrial SSUrDNA and Megaceros tosanus for nuclear SSU rDNA (seeDuiexcl amp Nickrent (1999) for details on methods) Mito-chondrial SSU rDNA sequences were not available forSpirogyra Chara Coleochaete and Selaginella so these taxawere scored as `missingrsquo in the data matrix The singletree resulting from a heuristic search of 3647 characters(562 informative) shows the same basic topology as thetotal morphology tree (centgure 12) The tree is well-resolvedwith 14 of the total 21 ingroup nodes receiving BS supportof 70 or more Strong BS support exists for the mono-phyly of hornworts mosses liverworts and ferns but
786 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Figure 12 The single MP tree of length 2543(CI ˆ 067 RI ˆ 053) resulting from a combinedanalysis of nuclear and mitochondrial SSU rDNANumbers above each branch represent the numberof characters supporting the branch Bootstrappercentage values (100 replications) followed bydecay indices are given below the branch Nodes Aand B received BS support of 55 and 61respectively when the outgroup Prototheca wasremoved Taxa with an asterisk () lack mitochon-drial SSU rDNA sequences
unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

unlike the morphological tree (centgure 11) only weaksupport is obtained for the basal position of hornwortsamong land plants (BS 43) a moss^liverwort clade (BS36) and tracheophytes (BS 45) It is of interest thatBS support for the tracheophyte clade (A in centgure 12)increases to 55 and the lycophyte clade (B in centgure 12)increases to 61 when the distant outgroup Prototheca isremoved from the analysis Support for these and otherrelationships including the basal position of hornworts isgreatly strengthened by the addition of mitochondrialSSU rDNA sequences of algal outgroups that are moreclosely related to embryophytes than Prototheca (eg Chara)(C L Parkinson and J D Palmer unpublished data)This tree topology is also congruent with the maximumlikelihood tree derived from the mitochondrial SSUrDNA data alone (Duiexcl amp Nickrent 1999) Additionalanalyses utilizing more genes and greater taxon samplingpoint to even stronger support for both the basal positionof the hornworts and the sister relationship of liverwortsand mosses (D L Nickrent R J Duiexcl C L Parkinsonand J D Palmer unpublished data)
Despite poor bootstrap support for some critical basalclades in the present analysis rearrangements of majorbranches in centgure 12 to match other proposed topologiesresult in even weaker support For example a cladogramcongruent with the Mishler amp Churchill (1984) hypoth-esis is six steps longer than the MP tree and a monophy-letic bryophyte topology is eight steps longer It hasbecome apparent that the phylogenetic signal present innuclear SSU rDNA is low especially for deeper nodes onthe land plant tree (Soltis et al 1999) thus this partitioncontributes a signicentcant amount of homoplasy to acombined data set
10 CONGRUENCE OF MORPHOLOGICAL AND
MOLECULAR PHYLOGENIES
The three separate data sets are largely congruent andpoint to a single topology for the relationships of the majorgroups of extant embryophytes The key features of thisphylogenetic hypothesis are as follows (i) that hornwortsliverworts mosses lycophytes and the remaining pterido-phytes and seed plants are each monophyletic (ii) thathornworts are sister to the remaining land plants (iii) thatmosses plus liverworts are monophyletic and (iv) that thismossliverwort clade is sister to another comprisingtracheophytes Of these features it is the basal position ofthe hornworts and the sister group relationship betweenmosses and liverworts that seem to be most debated
Because of their universal occurrence SSU rDNAgenes have been utilized extensively in phylogeneticstudies of the living world and specicentcally nuclear SSUrDNA sequences are informative in reconstructing plantalgal and protist phylogenies (Soltis amp Soltis 1998 Soltiset al 1999) Our primary conclusions are congruent withnuSSU rDNA phylogenies (eg Hedderson et al 1998Soltis et al 1999) although it should be noted that someanalyses of this gene have given radically diiexclerent results(eg Bopp amp Capesius 1996 Capesius amp Bopp 1997)Previous molecular analyses have also supported thebasic tree topologies shown in centgures 11 and 12 Malek etal (1996) derived a sister group relationship betweenmosses and liverworts by using sequences of the mito-
chondrial cox3 gene Steinhauser et al (1999) and Beckertet al (2000) arrived at the same conclusion based on thejoint possession of a group I intron and a phylogeneticanalysis of mitochondrial nad5 sequences A recent studyemploying centve genes from the chloroplast genome (butwith limited taxon sampling) also derived the same basictree topology (Nishiyama amp Kato 1999) Although anumber of molecular analyses and biochemical charactershave produced conclusions that are not consistent withour tree (reviewed by Qiu amp Palmer 1999) these wereoften compromised by small numbers of taxa short genes(or incomplete sequences) saturated third positions inprotein-coding genes rate heterogeneity and problematicanalyses (eg Van de Peer et al 1990 Waters et al 1992Manhart 1994 Mishler et al 1994 Bopp amp Capesius1996 Lewis et al 1997) Thus we do not consider themcompelling refutations of our primary conclusions Thedistribution of mitochondrial introns discussed by Qiu etal (1998) supports liverworts as the basalmost land plantlineage and is thus inconsistent with our conclusion Itshould be noted that their data set has a high level ofhomoplasy as several lineages higher in the cladogramalso possess the same character state as liverworts Thusthe absence of these three introns can be explained asarising from a loss in the common ancestor to liverwortsGiven our limited understanding of mitochondrialgenome evolution in plants we suggest that phylogeneticrelationships based on mitochondrial introns be inferredwith caution
11 INFERENCES ABOUT LAND PLANT EVOLUTION
It is premature to enumerate detailed speculations onmorphological transformation series in bryophytes Anumber of hypotheses have been laid out in the abovediscussion and these will require rigorous testing as robustmolecular and total evidence analyses on more taxabecome available In this section we make limited infer-ences about character evolution based on the consensustopology outlined in 10
Monosporangiate sporophytes and continuity in game-tophyte features such as apical organization and gametan-gial development support bryophytes as basal land plantlineagesThalloid gametophytes occur in hornworts and assuch are best viewed as plesiomorphic among land plantsEarly patterns in sporeling development suggest that anaxial erect habit is ancestral to the prostrate growth formof many mature liverwort and pteridophyte gametophytes(Whittier amp Braggins 1992 Bartholomew-Began 1991Bartholomew amp Crandall-Stotler 1986) Leafy buds thatoriginate from moss protonemata are also upright Thusthe mossliverwort progenitor was probably a branchedupright leapoundess axis Lateral appendages`leavesrsquo and theprocumbent habit apparently evolved a number of inde-pendent times in gametophytes of mosses and liverwortsParallel acquisition of `leavesrsquo in such taxa as Takakiatrue mosses Haplomitrium Phyllothallia Fossombronia andJungermanniales is supported by divergent develop-mental strategies as well as our phylogenetic analyses
Sporophyte reduction has occurred in all three bryo-phyte lineages and in our analyses is best illustrated bythe hornwort Notothylas Both the molecular and morpho-logical trees support the hypotheses that the highly
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 787
Phil Trans R Soc Lond B (2000)
simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

simplicented sporophyte of Notothylas is derived from themore complicated sporophytic structure that typicentes thegroup This concept was promoted by traditionalmorphologists (Schuster 1984c 1992 Proskauer 1960Lang 1907 Campbell 1895 Bartlett 1928) but has beencalled into question by more contemporary neontologists(Mishler amp Churchill 1985 Graham 1993) Notothylasoccurs in disturbed sites that presumably select for rapidgeneration time and reduced complexity of both lifecycle phases In general the rate of genomic evolutionand consequently phenotypic change is greater in smallorganisms that grow and reproduce rapidly a phenom-enon that in turn is often correlated with low genomesize (Niklas 1997 John amp Miklos 1988 Renzaglia et al1995) Thus the interpretation of small ephemeral taxasuch as Notothylas among anthocerotes and Riccia amonghepatics as derived is consistent with life history
phenomena and the seasonal instability experienced bythese plants
Homology of stomata is assumed in many morpholo-gical phylogenies (Kenrick amp Crane 1997ab Mishler ampChurchill 1984 1985) Developmental physiological andmorphological diiexclerences among stomata of hornwortsmosses and tracheophytes as well as the occurrence ofthese structures in specialized and restricted mosses andhornworts suggest that reappraisal of stomate homology iswarranted Interpretation of independent acquisition ofstomata in three lineages is equally parsimonious with theview that stomata evolved once in embryophytes andwere lost in liverworts three hornwort genera and basalmosses The speculation that bryophyte groups divergedprior to elongation and elaboration of sporophytessupports the contention that stomata are homoplastic inthese plants there would be no selective forces favouring
788 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Blasia Selaginella
Lycopodium
Psilotum
Angiopteris
EquisetumChara
Phaeoceros
Aulacomnium
Haplomitrium
Figure 13 Sperm cell architecture mapped onto a cladogram based on total morphology and nuclear plus mitochondrial SSUrDNA sequences See frac12 11 for explanation of the centgure Colour coding red ˆ poundagella and basal bodies blue ˆ nucleusbrown ˆ mitochondria yellow ˆ spline microtubules orange or purple ˆ lamellar strip green ˆ plastid grey ˆ extraneouscytoplasm
stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

stomate evolution in the unexpanded ephemeral ances-tral sporophyte Modern comprehensive comparativestudies are required to further test stomate homologyamong bryophytes and basal pteridophytes
Ultrastructural data on food-conducting cells support amoss^liverwort clade and provide refutation of the preceptthat leptoids in mosses are homologous to sieve cells oftracheophytes Because water-transporting cells probablyevolved at least three times in liverworts and twice inmosses (Ligrone et al this issue) care must be exercised ininterpreting homologyof these cells among taxa
Scrutiny of sperm cell structure and evaluation ofevolutionary trends based on molecular and morpholo-gical trees reveals examples of parallelisms divergencesand convergences among archegoniates (centgures 13 and14) The plesiomorphic architecture of plant spermato-zoids is a slightly coiled bipoundagellated cell Presumablyincreased compaction and coiling occurred independentlyin several lineages including the Charales mosses liver-worts heterosporous lycophytes and ferns (centgure 13)Hornwort sperm cells remained small and diverged fromother bryophytes before poundagellar staggering evolved An
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 789
Phil Trans R Soc Lond B (2000)
Phylloglossum
Lycopodium
Huperzia
Lycopodiella
Palhinhaea Selaginella
Isoetes
Figure 14 Lycophyte sperm cell architecture mapped onto a cladogram derived from spermatogenesis and rbcL sequences(WikstrIcircm amp Kenrick 1997) Multipoundagellated spermatozoids have evolved two independent times in lycophytes once in theheterosporous Isoelaquo tes and once in the homosporous Phylloglossum Within Lycopodiaceae bipoundagellated sperm cells of Lycopodiumand Huperzia are minute and coiled while those of Lycopodiella and Palhinhaea are more ovoid
autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

autapomorphy of hornworts is the reverse in direction-ality of coiling which as noted above was probablyinconsequential to the hydrodynamics of this minute cellWith resolution of a moss^liverwort clade the detailedsimilarities in locomotory apparatus microstructurebetween these two groups are interpreted as homologousFlagellar staggering involving dimorphic basal bodiesand an aperture in the spline were features of the proto-typic moss^liverwort sperm cell that further specializedafter historical separation of the two lineages (Renzagliaet al 1995)
Multipoundagellated sperm cells evolved within three sepa-rate lineages of pteridophytes in Phylloglossum a homo-sporous lycophyte (Renzaglia amp Maden 2000) in Isoelaquo tesa heterosporous lycophyte and in the Equisetum^Psilotum^fern^seed plant lineage (centgures 13 and 14) Additionalspecializations within the lycophytes include decreasedcoiling and increased poundagellar stagger in taxa with sub-terranean gametophytes and extreme streamlining inheterosporous taxa Commonalities in sperm cell devel-opment and organization provide strong support for afern^Equisetum^Psilotum clade an assemblage that hasbeen proposed by other contemporary analyses (Kenrickamp Crane 1997ab Duiexcl amp Nickrent 1999)
12 CONCLUSIONS AND PROSPECTS
Morphological and molecular data have nowconverged to provide resolution of the problem of bryo-phyte phylogeny in the context of the evolution of basalland plants Phylogenetic analyses of total evidence twomorphological and one molecular data set providesupport for hornworts as the basalmost clade of extantterrestrial organisms Furthermore there is reasonablesupport for the recognition of mosses and liverworts assister groups Although these conclusions are contrary tosome current interpretations we feel that they providethe best explanations of character homology and adaptiveradiation within the three bryophyte groups and betweenthese clades and tracheophytes
The ongoing dicurrenculties in resolving precise relation-ships among basal land plants are in part due to theirrapid radiation in a virtually uninhabited ancient land-scape and in part to the limitations of data sets Given thelimited number of surviving lineages in basal embryo-phytes the huge gaps between charophycean algae andland plants and between bryophytes and lycophytes(among extant organisms) morphological homology mayremain dicurrencult to resolve Conpoundicting data may repoundectthe almost simultaneous radiation of the bryophytelineages and considerable convergence based on commonevolutionary potential Resolution of these problemsrequires clear phylogenetic hypotheses and the willingnessto analyse homology based on developmental and ultra-structural features and not on supercentcial functional simi-larities of mature structures or organs Thus we arguethat the homologies in stomatal and transport cell charac-ters that were previously used to support the ancestralliverwort hypothesis are untenable
Molecular data are rapidly accumulating and we arecurrently analysing sequences of multiple genes from allthree plant genomes (ie nucleus mitochondrion andchloroplast) sampled from all appropriate extant lineages
From these data and from combined analyses withmorphological data a more resolved phylogeny shouldemerge that will probably mirror our general conclusionsSuch congruence will allow the phylogenetic debate toproceed beyond that of branching patterns so that plantbiologists can investigate equally interesting questions inevolutionary biology including rates and patterns ofevolutionary change With robust phylogenies and theaccumulation of new morphogenetic and ultrastructuraldata on key taxa we will be in a position to make moreaccurate and precise inferences about structural homolo-gies in basal land plants
This work would not have been possible without technical andconceptual contributions of present and previous students colla-borators and colleagues who have enabled the development ofour ideas In particular we note the involvement of J G Duck-ett D P Whittier K C Vaughn A R Maden R LigroneS J Schmitt H D Gates K D McFarland D K SmithH A Crum and L E Graham This work was supported bygrants to KSR DLN and DJG from the National ScienceFoundation USA (DEB-9207626 MCB-9808752 and DEB-9527735) and the Natural Sciences and Engineering ResearchCouncil Canada We thank J B Bozzola at the Micro-Imagingand Analysis Center SIUC and Thomas Johnson for technicalassistance in preparing the manuscript
REFERENCES
Bartholomew S E amp Crandall-Stotler B J 1986 The sporelingontogeny of Monoclea gottschei subsp elongata Bryologist 97244^252
Bartholomew-Began S E 1991 A morphogenetic re-evaluationof Haplomitrium Nees (Hepatophyta) Bryophytorum BibliothecaBand 41 Berlin J Cramer
Bartlett E M 1928 The comparative study of the developmentof the sporophyte in the Anthocerotaceae with special refer-ence to the genus Anthoceros Ann Bot 42 409^430
Bateman R M amp DiMichele W A 1994 Heterospory themost iterative key innovation in the history of the plantkingdom Biol Rev 69 345^417
Bateman R M Crane P R DiMichele W A Kenrick P Ramp Rowe N P 1998 Early evolution of land plants phylogenyphysiology and ecology of the primary terrestrial radiationA Rev Ecol Syst 29 263^292
Beckert S Steinhauser S Muhle H amp Knoop V 2000 Amolecular phylogeny of bryophytes based on nucleotidesequences of the mitochondrial sequences of the mitochon-drial nad5 gene Pl Syst Evol 218 179^192
Bell P R 1994 Green plants their origin and diversity PortlandOR Dioscorides Press
Bernhard D L amp Renzaglia K S 1995 Spermiogenesis in themoss Aulacomnium palustre Bryologist 98 52^70
Bierhorst D W 1971 Morphology of vascular plants New YorkMacMillan
Blackmore S amp Barnes S H 1987 Embryophyte spore wallsorigin development and homologies Cladistics 3 185^195
Bold H C Alexopoulos C J amp Delevoryas T 1987Morphology of plants and fungi 5th edn NewYork MacMillan
Bopp M amp Capesius I 1996 New aspects of bryophytetaxonomy provided by molecular approach Bot Acta 1091^5
Bremer K Humphries C J Mishler B D amp Churchill S P1987 On cladistic relationships in green plants Taxon 36339^349
Brown R C amp Lemmon B E 1988 Sporogenesis in bryo-phytes Adv Bryol 3 159^223
790 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

Brown R C amp Lemmon B E 1990a Monoplastidic cell divi-sion in lower land plants Am J Bot 77 559^571
Brown R C amp Lemmon B E 1990b Sporogenesis in bryo-phytes In Microspores evolution and ontogeny (ed S Blackmoreamp RV Knox) pp 55^94 London Academic Press
Brown R C amp Lemmon B E 1993 Diversity of cell divisionin simple land plants holds clues to evolution of the mitoticand cytokinetic apparatus in higher plants Mem Torrey BotClub 25 45^62
Brown R C amp Lemmon B E 1997 The quadripolar micro-tubule system in lower land plants J Plant Res 110 93^106
Brown R C Lemmon B E amp Renzaglia K S 1986Sporocytic control of spore wall patterns in liverworts Am JBot 73 593^596
Campbell D H 1895 The structure and development of mosses andferns (Archegoniatae) NewYork Macmillan
Capesius I amp Bopp M 1997 New classicentcation of liverwortsbased on molecular and morphological data Pl Syst Evol207 87^97
Carothers Z B amp Duckett J G 1980 The bryophyte sperma-tozoid a source of new phylogenetic information Bull TorreyBot Club 107 281^297
Crandall-Stotler B J 1980 Morphogenetic designs and atheory of bryophyte origins and divergence BioScience 30580^585
Crandall-Stotler B J 1984 Musci hepatics and anthocerotesoumlan essay on analogues In New manual of bryology vol 2 (edR M Schuster) pp1093^1129 Nichinan Japan HattoriBotanical Laboratory
Crandall-Stotler B J 1986 Morphogenesis developmentalanatomy and bryophyte phylogenetics contraindications ofmonophyly J Bryol 14 1^23
Crum H A amp Anderson L E 1981 Mosses of eastern NorthAmerica vols I^II NewYork Columbia University Press
Duckett J G 1970 Sexual behaviour of the genus Equisetumsubgenus Equisetum Bot J Linn Soc 63 327^352
Duckett J G amp Duckett A R 1980 Reproductive biology andpopulation dynamics of wild gametophytes of Equisetum BotJ Linn Soc 79 205^210
Duckett J G amp Ligrone R 1995 The formation of catenatefoliar gemmae and the origin of oil bodies in the liverwortOdotoschisma denudatum (Mart) Dum (Jungermanniales) alight and electron microscope study Ann Bot 76 405^419
Duckett J G amp Renzaglia K S 1988 Cell and molecularbiology of bryophytes ultimate limits to the resolution ofphylogenetic problems Bot J Linn Soc 98 225^246
Duckett J G Renzaglia K S amp Pell K 1990 Desiccationcauses the proliferation of multicellular hairs but not mucilagepapillae in Cryptothallus mirabilis a correlated light and elec-tron microscope study Can J Bot 68 697^706
Duckett J G Renzaglia K S amp Pell K 1991 A light andelectron microscope study of rhizoid^ascomycete associationsand poundagelliform axes in British hepatics with observations onthe eiexclects of the fungi on host morphology and comparisonswith mycorrhizae in the Ericaceae New Phytol 118 233^257
Duiexcl J R amp Nickrent D L 1999 Phylogenetic relationshipsamong land plants using mitochondrial small-subunit rDNAsequences Am J Bot 86 372^386
Duncan T M Renzaglia K S amp Garbary D J 1997Ultrastructure and phylogeny of the spermatozoid of Charavulgaris (Charophyceae) Pl Syst Evol 204 125^140
Fulford M 1956 The young stages of the leafy Hepaticae aresume Phytomorphology 6 199^235
Garbary D J amp Renzaglia K S 1998 Bryophyte phylogenyand the evolution of land plants evidence from developmentand ultrastructure In Bryology for the twenty-centrst century (edJ W Bates N W Ashton amp J G Duckett) pp 45^63 LeedsThe British Bryological Society and Maney Publishing
Garbary D J Renzaglia K S amp Duckett J G 1993 Thephylogeny of land plants a cladistic analysis based on malegametogenesis Pl Syst Evol 188 237^269
Giiexclord E M amp Foster A S 1988 Morphology and evolution ofvascular plants 3rd edn NewYork Freeman
Gould S J 1989 Wonderful lifeoumlthe Burgess Shale and the nature ofhistory NewYork W W Norton amp Co
Graham L E 1993 Origin of land plants NewYork JohnWileyGunning B E amp Steer M W 1996 Plant cell biology structure and
function Sudbury MA Jones and Bartlett PublishersHanson D T Swanson S Graham L E amp Sharkey T D
1999 Evolutionary signicentcance of isoprene emission frommosses Am J Bot 86 634^639
Hebant C 1977 The conducting tissues of bryophytesBryophytorum Bibliotheca vol 10Vaduz Cramer
Hedderson T A Chapman R amp Cox C J 1998 Bryophytesand the origins and diversicentcation of land plants new evidencefrom molecules In Bryology for the twenty-centrst century (ed J WBates N W Ashton amp J G Duckett) pp 65^77 Leeds TheBritish Bryological Society and Maney Publishing
Hemsley A R 1994 The origin of the land plant sporophytean interpolational scenario Biol Rev 69 263^273
John B amp Miklos G 1988 The eukaryote genome in development andevolution London Allen and Unwin
Kenrick P amp Crane P R 1997a The origin and early diversicentcationof land plants a cladistic study Washington DC SmithsonianInstitution Press
Kenrick P amp Crane P R 1997b The origin and early evolutionof plants on land Nature 389 33
Korall P Kenrick P amp Therrien J P 1999 Phylogeny ofSelaginellaceae evaluation of genericsubgeneric relationshipsbased on rbcL gene sequences Int J Plant Sci 160 585^594
LangW H 1907 On the sporogonium of Notothylas Ann Bot 21201^210
Lewis L A Mishler B D amp Vilgalys R 1997 Phylogeneticrelationships of the liverworts (Hepaticae) a basal embryo-phyte lineage inferred from nucleotide sequence data of thechloroplast gene rbcL Mol Phyl Evol 7 377^393
Ligrone R Duckett J G amp Renzaglia K S 1993 Thegametophyte sporophyte junction in land plants Adv Bot Res19 231^317
McCourt R M 1995 Green algal phylogeny Trends Evol Ecol10 159^163
Maddison W P amp Maddison D R 1992 MacClade analysis ofphylogeny and character evolution 30 Sunderland MA SinauerAssociates
Maden A R Renzaglia K S amp Whittier D P 1996Ultrastructure of the spermatozoid of Lycopodiella obscurum(Lycopodiaceae) Am J Bot 83 419^429
Maden A R Whittier D P Garbary D J amp RenzagliaK S 1997 Ultrastructure of the spermatozoid of Lycopodiellalateralis (RBr) B Ogravellgaard (Lycopodiaceae) Can J Bot 751728^1738
Malek O LIgravettig K Hiesel R Brennicke A amp Knoop V1996 RNA editing in bryophytes and a molecular phylogenyof land plants EMBOJ 15 1403^1411
Manhart J R 1994 Phylogenetic analysis of green plant rbcLsequences Mol Phyl Evol 3 114^127
Manhart J R 1995 Chloroplast 16S rDNA sequences andphylogenetic relationships of fern allies and ferns Am Fern J85 182^192
Mishler B D amp Churchill S P 1984 A cladistic approach tothe phylogeny of `bryophytesrsquo Brittonia 36 406^424
Mishler B D amp Churchill S P 1985 Transition to a land poundoraphylogenetic relationships of the green algae and bryophytesCladistics 1 305^328
Mishler B D Lewis L A Buchheim M A RenzagliaK S Garbary D J Delwiche C F Zechman F W
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 791
Phil Trans R Soc Lond B (2000)
Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

Kantz T S amp Chapman R L 1994 Phylogenetic relation-ships of the green algaersquo and `bryophytesrsquo Ann Mo Bot Gdn8 451^483
Niklas K J 1997 The evolutionary biology of plants University ofChicago Press
NishiyamaT amp Kato M 1999 Molecular phylogenetic analysisamong bryophytes and tracheophytes based on combineddata of plastid coded genes and the 18S rRNA gene Mol BiolEvol 16 1027^1036
Ogravellgaard B 1987 A revised classicentcation of the LycopodiaceaeOpera Botan 92 153^178
Oliver M J amp Wood A J 1997 Desiccation tolerance inmosses In Stress-inducible processes in higher eukaryotic cells (ed TKoval) pp1^26 NewYork Plenum Press
Pahnke J GoremykinV BobrovaV Troitsky A Antonov Aamp Martin W 1996 Utility of rDNA internal transcribedspacer sequences from the inverted repeat of chloroplast DNAin pteridophyte molecular phylogenetics In Pteridology inperspective (ed J M Camus M Gibby amp R J Johns) KewRoyal Botanic Gardens
Paton J A amp Pearce J V 1957 The occurrence structure andfunctions of the stomata of British bryophytesTrans Br BryolSoc 3 228^259
Pirozynski K A 1981 Interactions between fungi and plantsthrough the ages Can J Bot 59 1824^1827
Pirozynski K A amp Malloch D W 1975 The origin of landplants a matter of mycotropism Biosystems 6 153^164
Proskauer J 1960 Studies on Anthocerotales VI On spiralthickenings in the columella and its bearing on phylogenyPhytomorphology 10 1^19
Pryer K M Smith A R amp Skog J E 1995 Phylogeneticrelationships of extant ferns based on evidence frommorphology and rbcL sequences Am FernJ 85 205^282
Qiu Y L amp Palmer J D 1999 Phylogeny of early land plantsinsights from genes and genomesTrends Pl Sci 4 26^30
Qiu Y L Cho Y Cox J C amp Palmer J D 1998 The gain ofthree mitochondrial introns identicentes liverworts as the earliestland plants Nature 394 671^674
Remy W Gensel P G amp Hass H 1993 The gametophytegeneration of some early Devonian land plants Int J Pl Sci54 35^58
Renzaglia K S 1978 A comparative morphology and develop-mental anatomy of the Anthocerotophyta J Hattori Bot Lab44 31^90
Renzaglia K S 1982 A comparative developmental investiga-tion of the gametophyte generation in the Metzgeriales(Hepatophyta) Bryophytorum Bibliotheca vol24 Vaduz JCramer
Renzaglia K S amp Bartholomew S E 1985 Sporeling develop-ment in Fossombronia cristula Aust with special reference toapical organization and growth Bryologist 88 337^343
Renzaglia K S amp Duckett J G 1987 Spermatogenesis ofBlasia pusilla from antheridial initial through mature sperma-tozoid Bryologist 90 468^501
Renzaglia K S amp Duckett J G 1988 Diiexclerent develop-mental processes underlie similar spermatozoid architecturein mosses liverworts and hornworts J Hattori Bot Lab 64219^236
Renzaglia K S amp Duckett J G 1989 Ultrastructural studiesof spermatogenesis in the Anthocerotophyta V Nuclearmetamorphosis and the posterior mitochondrion of Notothylas orbicu-laris and Phaeoceros laevis Protoplasma 151 137^150
Renzaglia K S amp Duckett J G 1991 Towards an under-standing of the diiexclerence between the blepharoplasts ofmosses and liverworts and comparisons with hornwortsbipoundagellate lycopods and charophytes a numerical analysisNew Phytol 117 187^208
Renzaglia K S amp Garbary D J 2000 Motile male gametes ofland plants diversity development and evolution Crit Rev PlSci (In the press)
Renzaglia K S amp Maden A R 2000 Microtubule organizingcenters and the origin of centrioles during spermatogenesis inthe pteridophyte Phylloglossum Micros ResTech 49 (In the press)
Renzaglia K S amp Vaughn K C 2000 Anatomy developmentand classicentcation of hornworts In The biology of bryophytes (edJ Shaw amp B Gocurrennet) pp 1^19 Cambridge University Press
Renzaglia K S amp Whittier D P 1993 Spermatozoids ofPhylloglossum drummondii are multipoundagellated Am J Bot 80111 (Suppl)
Renzaglia K S Maden A R Duckett J G amp WhittierD P 1994 Monoplastidy in spermatogenesis of Lycopodiumobscurum Can J Bot 72 1436^1444
Renzaglia K S Rasch E M amp Pike L M 1995 Estimates ofnuclear DNA content in bryophyte sperm cells phylogeneticconsiderations Am J Bot 82 18^25
Renzaglia K S McFarland K D amp Smith D K 1997Anatomy and ultrastructure of the sporophyte of Takakia cera-tophylla (Bryophyta) Am J Bot 84 1337^1350
Renzaglia K S Dengate S B amp Bernhard D L 1998Architecture of the spermatozoid of Selaginella australiensisAm FernJ 88 1^16
Renzaglia K S Bernhard D L amp Garbary D J 1999Developmental ultrastructure of the male gamete ofSelaginella Int J Pl Sci 160 14^28
Robbins R R amp Carothers Z B 1978 Spermatogenesis inLycopodium the mature spermatozoid Am J Bot 65 433^440
Robert D 1974 Etude ultrastructurale de la spermiogenesenotamment de la diiexclerentiation de lrsquoappareil nucleaire chezle Selaginella kraussiana (Kunze) A Br Annales des SciencesNaturalles Botanique Paris Serie 12 15 65^118
Roth D 1969 Embryo und Embryotheca bei den LaubmoosenEine histogenetische und morphologische UntersuchungBiblioth Bot 129 1^49 Pls1^9
Roux W 1895 Gesammelte Abhandlungen zur Entwicklungsmechanikder Organismen Leipzig Engelmann
Sack F D amp Paollilo D J Jr 1985 Incomplete cytokinesis inFunaria stomata Am J Bot 72 1325^1333
Schocenteld W B 1985 Introduction to bryology NewYork MacmillanSchuster R M 1966 The Hepaticae and Anthocerotae of North
America east of the hundredth meridian vol 1 New YorkColumbia University Press
Schuster R M 1979 The phylogeny of the Hepaticae InBryophyte systematics (ed G C S Clarke amp J G Duckett)pp 41^82 London Academic Press
Schuster R M 1984a Comparative anatomy and morphologyof Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp760^891 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984b Evolution phylogeny and classicentcation ofthe Hepaticae In New manual of bryology vol 2 (ed R MSchuster) pp 892^1070 Nichinan Japan Hattori BotanicalLaboratory
Schuster R M 1984c Morphology phylogeny and classicentcationof the Anthocerotae In New manual of bryology vol 2 (edR M Schuster) pp1071^1092 Nichinan Japan HattoriBotanical Laboratory
Schuster R M 1992 The Hepaticae and Anthocerotae of NorthAmerica volV Chicago Field Museum of Natural History
Smith D K amp Davison P G 1993 Antheridia and sporophytesin Takakia ceratophylla (Mitt) Grolle evidence for reclassicentca-tion among the mosses J Hattori Bot Lab 73 263^271
Smith E C amp Gricurrenths H 1986a The occurrence of the chlor-oplast pyrenoid is correlated with the activity of a CO2-concentrating mechanism and carbon isotope discriminationin lichens and bryophytes Planta 198 6^16
792 K S Renzaglia and others Vegetative and reproductive innovations of early land plants
PhilTrans R Soc Lond B (2000)
Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)

Smith E C amp Gricurrenths H 1986b A pyrenoid-based carbonconcentrating mechanism is present in terrestrial bryophytesof the class Anthocerotae Planta 200 203^212
Smith G M 1955 Cryptogamic botany II Bryophytes and pterido-phytes 2nd edn NewYork McGraw-Hill
Soltis D E amp Soltis P S 1998 Choosing an approach and anappropriate gene for phylogenetic analysis In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp1^42 Boston MA Kluwer AcademicPublishers
Soltis P S Soltis D E Wolf P G Nickrent D L Chaw S-M amp Chapman R L 1999 The phylogeny of land plantsinferred from 18S rDNA sequences pushing the limits ofrDNA signal Mol Biol Evol 16 1774^1784
Southworth D amp Cresti M 1997 Comparisons of poundagellatedand nonpoundagellated sperm in plants Am J Bot 841301^1311
Steinhauser S Beckert S Capesius I Malek O amp KnoopV1999 Plant mitochondrial RNA editing J Mol Evol 48303^312
Stewart W N amp Rothwell G W 1993 Paleobotany and the evolu-tion of plants 2nd edn Cambridge University Press
Taylor T N amp Taylor E L 1993 The biology and evolution of fossilplants Englewood Cliiexcls NJ Prentice-Hall
van de Peer Y De Baere R Cauwenberghsm J amp WachterR 1990 Evolution of green plants and their relationships withother photosynthetic eukaryotes as deduced from 5S ribo-somal RNA sequences Pl Syst Evol 170 85^96
Vaughn K C amp Harper J D 1998 Microtubule organizingcenters and nucleating sites in land plants Int Rev Cytol 18175^149
Vaughn K C amp Renzaglia K S 1998 Origin of bicentrioles inanthocerote spermatogenous cells In Bryology for the twenty-centrstcentury (ed J W Bates N W Ashton amp J G Duckett)pp189^203 Leeds The British Bryological Society andManey Publishing
Vaughn K C Campbell E O Hasegawa J Owen H A ampRenzaglia K S 1990 The pyrenoid is the site of ribulose 1^5-bisphosphate carboxylaseoxygenase accumulation in thehornwort (Bryophyta Anthocerotae) chloroplast Protoplasma156 117^129
Vaughn K C Ligrone R Owen H A Hasegawa JCampbell E O Renzaglia K S amp Monge-Najera J1992 The anthocerote chloroplast a review New Phytol 120169^190
Vaughn K C Sherman T D amp Renzaglia K S 1993 Acentrin homologue is a component of the multilayeredstructure in bryophytes and pteridophytes Protoplasma 17558^66
Waters D A Buchheim M A Dewey R A amp ChapmanR L 1992 Preliminary inferences of the phylogeny of bryo-phytes from nuclear-encoded ribosomal RNA sequences AmJ Bot 79 459^466
Whittier D P 1977 Gametophytes of Lycopodium obscurum asgrown in axenic culture Can J Bot 55 563^567
Whittier D P 1981 Gametophytes of Lycopodium digitatum(formerly L complanatum var poundabelliforme) as grown in axenicculture Bot Gaz 142 519^524
Whittier D P 1983 Gametophytes of Ophioglossum engelmanniiCan J Bot 61 2369^2373
Whittier D P amp Braggins J E 1992 The young gametophyteof Phylloglossum (Lycopodiaceae) Ann Mo Bot Gdn 79730^736
Whittier D P amp Peterson R L 1984 Gametophytes ofBotrychium lunarioides and their mucilage-coated rhizoids CanJ Bot 62 2854^2860
Whittier D P amp Thomas R D 1993 Gametophytes and youngsporophytes of Botrychium jenmanii in axenic culture Int J PlSci 154 68^74
Whittier D P amp WebsterT R 1986 Gametophytes of Lycopodiumlucidulum from axenic culture Am FernJ 76 48^55
WikstrIcircm N amp Kenrick P 1997 Phylogeny of Lycopodiaceae(Lycopsida) and the relationships of Phylloglossum drummondiiKunze based on rbcL sequences Int J Pl Sci 158 862^871
Wolf P G 1997 Evaluation of atpB nucleotide sequences forphylogenetic studies of ferns and other pteridophytes Am JBot 84 1429^1440
Wolf P G Pryer K M Smith A R amp Hasebe M 1998Phylogenetic studies of extant pteridophytes In Molecularsystematics of plants II DNA sequencing (ed D E Soltis P SSoltis amp J J Doyle) pp 540^561 Boston MA KluwerAcademic Publishers
Vegetative and reproductive innovations of early land plants K S Renzaglia and others 793
Phil Trans R Soc Lond B (2000)