very low density lipoproteins promote triglyceride...
TRANSCRIPT

Very Low Density Lipoproteins PromoteTriglyceride Accumulation in
Macrophages
Sandra R. Bates, Patricia L. Murphy, Zongchen Feng,Takemichi Kanazawa, and Godfrey S. Getz
Incubation of mouse peritoneal macrophages with very low density lipoproteins(VLDL) from normal rats or rhesus monkeys markedly increased the levels of intracel-lular trlglycerldes by 10- to 56-fold and was accompanied by the production of oil red0 positive vacuoles. The stimulation of triglyceride accumulation in macrophageswas time- and concentration-dependent and was specific for VLDL. Three possiblemechanisms for the VLDL-stlmulated triglyceride accumulation In macrophages wereexplored: receptor-mediated uptake, action of llpoprotein llpase, and phagocytosis.Macrophage uptake and degradation of 125l-monkey and rat VLDL demonstrated satu-rable and nonsaturable components. Uptake of 125I-VLDL could be inhibited by unla-beled normal VLDL, although hyperllpemlc VLDL was more effective. HDL did notcompete to a significant extent. Heparln released llpoprotein lipase-llke activity fromperitoneal macrophages. Addition of heparln with VLDL resulted in a greater, morerapid elevation in Intracellular triglycerides, which was partially Inhibited by albumin.Free fatty acid and Intrallpid also produced triglyceride accumulation In macro-phages. The data showed that all three of the mechanisms examined could contributeto the metabolism of VLDL by macrophages and cause the production of trlglycerlde-rlch cells with a "foamy" appearance, although the evidence suggested that the actionof llpoprotein llpase was probably the most important In this process.(Arteriosclerosis 4:103-114, March/April 1984)
Lipid deposition in the aorta and other major arter-ies is an important component of the atherogenic
process. The accumulation of lipids within cells of thearterial wall is evident as oil red O positive inclusionswhich give the cells a foamy appearance. These"foam" cells isolated from the aorta are enriched incholesterol;1"4 the triglyceride content of such cellshas not been reported. The diseased vessel itself isenriched with cholesterol esters with a modest in-crease in triglycerides.5 The accumulation of choles-
From the Departments of Pathology and Biochemistry, and theSpecialized Center of Research in Atherosclerosis, University ofChicago, Chicago, Illinois. Dr. Feng Is now at the Department ofBiochemistry, Wuhan Medical College, Wuhan, The Peoples Re-public of China. Dr. Kanazawa is at the Department of InternalMedicine, Hirosakl Medical University, Hlrosaki, Japan.
This investigation was supported in part by USPHS Grant HL-15062 (SCOR) and Grant HL-27005 from the National Heart,Lung and Blood Institute of the National Institutes of Health.
Address for reprints: Dr. Sandra R. Bates, University of Chica-go, Department of Pathology, Box 414, 950 East 59th Street,Chicago, Illinois 60637.
Received June 20, 1983; revision accepted December 20,1983.
terol esters is thought to give the cells of the arterywall their "foamy" appearance although high intra-cellular triglycerides can also produce "foamy" cellsvery similar in appearance.67
Ultrastructural studies have demonstrated thatfoam cells have smooth muscle cell and macro-phage characteristics.8"11 Recently, foam cells isolat-ed from atherosclerotic lesions induced by diet werefound to have several histological and functionalproperties specific for macrophages.1011 The lipidmetabolism of macrophages has been extensivelyinvestigated in the past,12 but recent attention hasfocused on their cholesterol metabolism. Studieshave demonstrated that macrophages have cell sur-face receptors specific for cholesterol ester—rich,beta-migrating very low density lipoproteins (P-VLDL) isolated from animals with diet-induced hy-perlipidemia.1314 p-VLDL were shown to promoteextensive cholesterol ester build-up in macro-phages.1314 In contrast, little is known regarding theinteraction of macrophages with triglyceride-richVLDL from normolipemic animals or the possible ef-fects of this lipoprotein fraction on their triglyceride
103
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

104 ARTERIOSCLEROSIS VOL 4, No 2, MARCH/APRIL 1984
metabolism. The synthesis and accumulation of tri-glyceride by other cell types has been studied in vitroby several laboratories.815"17 Human skin fibro-blasts,1516 bovine aortic endothelial cells,17 and ratpreadipocyte cultures6 showed increased triglycer-ide content when incubated in the presence of nor-mal VLDL, although to different extents. The presentstudy describes the accumulation of triglycerides inmouse peritoneal macrophages stimulated by nor-mal triglyceride-rich VLDL.
Methods
Tissue Culture
Unstimulated peritoneal macrophages were iso-lated from female Swiss-Webster mice according tothe method of Cohn and Benson18 using sterile phos-phate-buffered saline (PBS). The exudate was cen-trifuged, and the supernatant discarded. When con-taminating red blood cells were present, they werelysed by a brief exposure to distilled water dilutedimmediately with Dulbecco's Modified Eagle's medi-um (DMEM) (Grand Island Biological Company,Grand Island, New York). After recentrifugation, thecells were suspended in DMEM plus 10% heat-inac-tivated fetal calf serum (FCS) (K.C. Laboratory Sup-ply Company, Indianapolis, Indiana). The cells wereplated in 1.5 ml aliquots in Multiwell tissue cultureplates (Costar, 12 wells/plate). The amount of cellu-lar protein per well ranged from 50 to 150 ng (ap-proximately 2-5 x 106 cells/ml). After 2 hours ofincubation at 37°C in a 5% CO2 atmosphere, themedia containing nonadherent cells were removed,and the attached cells were washed twice with PBS.The cells were then incubated for 1 or 2 days in 1.5ml of DMEM plus 10% FCS. Finally, the macro-phages were analyzed or used for experiments. Atthe start of an experiment, the cells were washedtwice with PBS. One ml of test medium consisting ofDMEM plus additions was added per well and thecells were incubated for the appropriate time periodsat 37°C. At the end of an experiment, the cells werewashed three times with PBS and harvested by ex-posure to 0.1 N NaOH for 20 minutes at room tem-perature. This short treatment with NaOH did nothydrolyze cellular triglycerides. The dissolved ma-crophages were neutralized with concentrated ace-tic acid and extracted immediately using the methodof Bligh and Dyer.19
Macrophage-like cell lines J774A.1 (J774)20 andP388D1, interleukin-1 secretor (P388)21 were ob-tained from the Cell Distribution Center of the SalkInstitute, San Diego, California, and were maintainedin DMEM containing 10% FCS. The cell lines wereseeded so that the final protein concentration perwell was 0.2-0.5 mg.
Arterial medial smooth muscle cells were obtainedfrom outgrowths of explants from the thoracic aortaof Macaca mulatta monkeys (rhesus) as previouslydescribed,22-23 grown in Basal Medium of Eagle
(BME) with 5% calf serum, and used between the 4thand 6th passages (0.3-1.0 mg protein/flask). Experi-ments were carried out in 25 cm2 flask (Falcon) in avolume of 2.0 ml per flask and terminated as de-scribed for macrophages.
Llpoproteins
Normal lipoproteins were isolated from the plasmaof male Sprague-Dawley rats (King) fed Purina ratchow or from rhesus monkeys fed monkey chow.Normal VLDL isolated from rats or monkeys hadsimilar chemical compositions. The rat VLDL was55% triglyceride, 11% protein, 25% phospholipid,5% total cholesterol, and 4% free fatty acid while themonkey VLDL was 52% triglyceride, 11% protein,21% phospholipid, 12% total cholesterol, and 5%free fatty acid. The apoprotein content, however,was different. The rat VLDL had both high molecularweight (Mr 335,000) apoprotein B (apo B-100) andlower molecular weight (M, 240,000) apoprotein B(apo B-48) with significant apoprotein E levels, whilethe monkey VLDL had predominantly apo B-100 andlittle apoprotein E. Human VLDL was isolated fromthe plasma of fasting normal healthy volunteers. Hy-perlipemic VLDL was obtained from a rhesus mon-key fed a Purina monkey chow diet supplementedwith 2% cholesterol and 25% coconut oil for 2 years(mean serum total cholesterol value = 950 mg/dl).The hyperiipemic VLDL was beta-migrating and cho-lesterol ester-rich. The plasma was centrifuged at25,000 rpm for 20 minutes in a SW 27 rotor to re-move chylomicrons. VLDL (d < 1.006), low densitylipoproteins (LDL) (d = 1.019-1.050) and high den-sity lipoproteins (HDL) (d = 1.063-1.21) were isolat-ed according to the method of Havel et al.24 withmodifications as described by Scanu and col-leagues.2526 Then 10 ;u,M phenylmethylsulfonyl flu-oride (PMSF) was added to the plasma before lipo-protein isolation to inhibit serine proteases. Alllipoproteins were washed by recentrifugation andchecked for purity by agarose electrophoresis. Thelipoproteins were dialyzed against PBS and steril-ized by filtration through a 0.45 /zm Millex-HA filter(Millipore) before use. Lipoprotein-deficient serum(LDS) was obtained at a density greater than 1.21from normal monkey plasma, dialyzed, heated at56°C for 30 minutes and filtered. All lipoproteins wereused within 2 weeks of preparation.
Protein was determined according to the methodof Lowry27 and phospholipids according to the proce-dure of Bartlett.28 The total cholesterol and triglycer-ide content of the VLDL was analyzed with a Techni-con Autoanalyzer II.29 Fatty acid-free bovine serumalbumin was purchased from Sigma, and Intralipidwas purchased from Cutter Laboratory.
Assays
Triglyceride and free fatty acid were quantitatedusing the method of Marsh and Weinstein30 as modi-fied by Kritchevsky et al.31 Briefly, the extracted lipids
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

VLDL INCREASES MACROPHAGE TRIGLYCERIDES Bates et al. 105
were separated on silica gel G glass thin-layer chro-matography plates (Fisher Scientific Company,Itasca, Illinois) and developed with petroleum ether/ethyl ether/acetic acid (75:25:1). The separated li-pids were charred with sulfuric acid and were ana-lyzed spectrophotometrically. The internal standardwas 4-uC cholesterol (56 Ci/mol, Amersham/Searle)added during the extraction, and the results werecorrected for recovery (mean recovery = 80%).Gas-liquid chromatographic techniques were usedto determine the free and total cholesterol content ofthe cells and media in Table 6.a The esterified cho-lesterol was calculated by subtraction of the freecholesterol from the total cholesterol values and re-presents the cholesterol in the cholesterol ester. So-dium oleate complexed to bovine serum albuminwas prepared according to the procedure of St. Clairet al.32
The secretion of lipoprotein lipase-like activity bycells was measured by incubating cells in DMEMwith or without heparin (10 lU/ml, 71 /xg/ml) for 5hours (J774) or 24 hours (peritoneal macrophages).The media were removed, centrifuged to remove anyfloating cells, and assayed immediately. Media incu-bated without cells served as the control. Lipoproteinlipase-like activity was assayed using 3H-triolein ac-cording to the method described by Nilsson-Ehle andSchotz.33
Interaction of 12SI-VLDL and Macrophages
VLDL were iodinated by the iodine monochloridemethod described by Karlin et al.34 as previously out-lined.35 Of the 125I lipoprotein, 99% was TCA-precipi-table; less than 20% of the label was in lipids. Deter-minations of the uptake (binding plus incorporation)and degradation of 125I-VLDL were done as de-scribed previously35 using the method of Brown andGoldstein.36 The macrophages were incubated for 5hours at 37°C in Multiwell tissue culture plates (Cos-tar, 24 wells/plate) with 0.5 ml of media containing125I-VLDL and 2.5 mg/ml LDS. At the end of the incu-bation, the media were removed and the macro-phages were washed three times with cold PBS con-taining 0.2% albumin and three times with PBSalone. We used 0.2 N NaOH to harvest the macro-phages and took aliquots for protein determinationand measurements of 125l-radioactivity (= uptake,binding plus incorporation). The media were ana-lyzed for degraded VLDL as previously described35
using the procedures of Brown and Goldstein.36 Thedata were corrected for binding or degradation whichoccurred in the absence of cells.
ResultsMouse peritoneal macrophages incubated in me-
dium with 10% FCS contained very little triglyceride(8.2 ± 4.4 /ng triglyceride/mg cell protein). Incuba-tion in medium alone for 48 hours did not change thetriglyceride content of the cells (Table 1). After expo-
Table 1. Effect of Normal VLDL on the TriglycerideContent of Mouse Peritoneal Macrophages
Experi-ment
1234
Medium(a)
(^g triglyceride/mg
12.14.56.3
14.2
Medium +VLDL
(b)cell protein)
130.097.4
355.4200.7
b/a(fold
stimulation)
10.721.656.414.1
Macrophages were incubated for 48 hours in mediumalone or with normal rat VLDL (300 fig triglyceride/ml).
sure to normal VLDL (300 /^g triglyceride/ml) for 48hours, the cells contained many large luminescent oilred O positive vacuoles which were not visible in thecontrol cells, and as shown in Table 1, the triglycer-ide content of the macrophages increased approxi-mately 10- to 56-fold. Thus, under similar experimen-tal conditions, the macrophages accumulated aslittle as 97 ng triglyceride/mg cell protein or as muchas 355 fig triglyceride/mg cell protein. Although thequantity of triglyceride varied between preparationsof macrophages, within each experiment the dupli-cates were usually within 10% of each other. Thereason for these differences between experiments isnot clear, but preliminary results indicate that a por-tion of this variation may be attributable to the num-ber of cells per unit area since, at low cell numbersper well, triglyceride accumulation was inversely pro-portional to the amount of cellular protein. The age ofthe VLDL preparation, the state of the macrophagesand other unknown factors probably also contributedto the observed variation. The free fatty acid contentof cells incubated in 10% FCS was 21.2 ± 11.7 ^glmg cell protein. After 48 hours in medium alone or inmedium with added VLDL, the cells contained 16.8± 12.7 pg free fatty acid or 33.0 ± 18.0/ig free fattyacid/mg cell protein, respectively, indicating only aminor effect of VLDL on the free fatty acid content ofthe macrophages.
In contrast to macrophages, monkey arterialsmooth muscle cells showed a more limited incre-ment in triglyceride levels upon exposure to VLDL.Control cells incubated in LDS contained 19.6 ± 6.9/xg triglyceride/mg cell protein (n = 6), while cellsexposed to VLDL (200 ^g triglyceride/ml) for 48hours had 48.1 ± 11.8 ^g triglyceride/mg cell pro-tein (n = 10).
The ability of other lipoproteins to augment theintracellular triglyceride content of macrophageswas examined. The addition of monkey LDL or HDLto the culture medium elevated the intracellulartriglyceride content of macrophages but not to theextent seen with VLDL (Figure 1). Thus, at compara-ble triglyceride concentrations, other lipoproteinclasses were much less effective than VLDL in stim-ulating triglyceride accumulation in macrophages.
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from
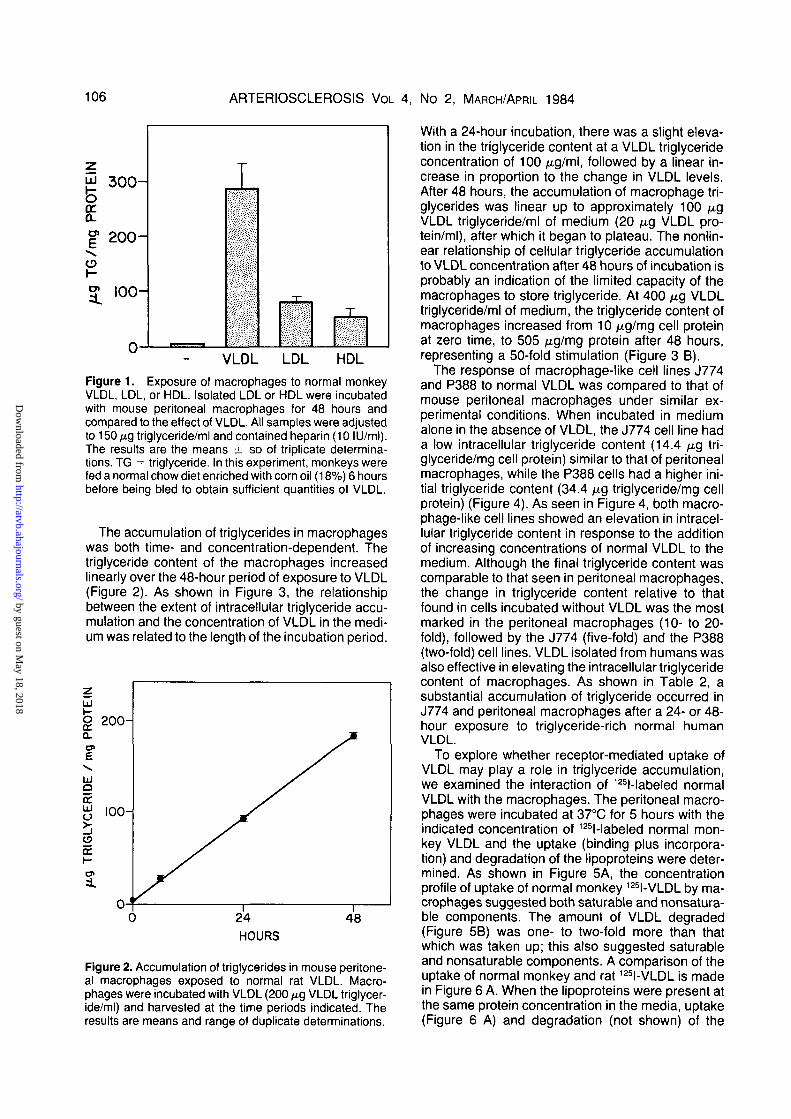
106 ARTERIOSCLEROSIS VOL 4, No 2, MARCH/APRIL 1984
300-
PRO
o>E\<Dh-O>
=L
200
I00
-
I
-j-
T
VLDL LDL HDL
Figure 1. Exposure of macrophages to normal monkeyVLDL, LDL, or HDL. Isolated LDL or HDL were incubatedwith mouse peritoneal macrophages for 48 hours andcompared to the effect of VLDL. All samples were adjustedto 150/xgtriglyceride/ml and contained heparin (10 ILJ/ml).The results are the means ± SD of triplicate determina-tions. TG = triglyceride. In this experiment, monkeys werefed a normal chow diet enriched with corn oil (18%) 6 hoursbefore being bled to obtain sufficient quantities of VLDL.
The accumulation of triglycerides in macrophageswas both time- and concentration-dependent. Thetriglyceride content of the macrophages increasedlinearly over the 48-hour period of exposure to VLDL(Figure 2). As shown in Figure 3, the relationshipbetween the extent of intracellular triglyceride accu-mulation and the concentration of VLDL in the medi-um was related to the length of the incubation period.
HOURS
Figure 2. Accumulation of triglycerides in mouse peritone-al macrophages exposed to normal rat VLDL. Macro-phages were incubated with VLDL (200 /ng VLDL triglycer-ide/ml) and harvested at the time periods indicated. Theresults are means and range of duplicate determinations.
With a 24-hour incubation, there was a slight eleva-tion in the triglyceride content at a VLDL triglycerideconcentration of 100 /xg/ml, followed by a linear in-crease in proportion to the change in VLDL levels.After 48 hours, the accumulation of macrophage tri-glycerides was linear up to approximately 100 /ugVLDL triglyceride/ml of medium (20 /xg VLDL pro-tein/ml), after which it began to plateau. The nonlin-ear relationship of cellular triglyceride accumulationto VLDL concentration after 48 hours of incubation isprobably an indication of the limited capacity of themacrophages to store triglyceride. At 400 /u,g VLDLtriglyceride/ml of medium, the triglyceride content ofmacrophages increased from 10 //.g/mg cell proteinat zero time, to 505 /ng/mg protein after 48 hours,representing a 50-fold stimulation (Figure 3 B).
The response of macrophage-like cell lines J774and P388 to normal VLDL was compared to that ofmouse peritoneal macrophages under similar ex-perimental conditions. When incubated in mediumalone in the absence of VLDL, the J774 cell line hada low intracellular triglyceride content (14.4 /u.g tri-glyceride/mg cell protein) similar to that of peritonealmacrophages, while the P388 cells had a higher ini-tial triglyceride content (34.4 /ug triglyceride/mg cellprotein) (Figure 4). As seen in Figure 4, both macro-phage-like cell lines showed an elevation in intracel-lular triglyceride content in response to the additionof increasing concentrations of normal VLDL to themedium. Although the final triglyceride content wascomparable to that seen in peritoneal macrophages,the change in triglyceride content relative to thatfound in cells incubated without VLDL was the mostmarked in the peritoneal macrophages (10- to 20-fold), followed by the J774 (five-fold) and the P388(two-fold) cell lines. VLDL isolated from humans wasalso effective in elevating the intracellular triglyceridecontent of macrophages. As shown in Table 2, asubstantial accumulation of triglyceride occurred inJ774 and peritoneal macrophages after a 24- or 48-hour exposure to triglyceride-rich normal humanVLDL.
To explore whether receptor-mediated uptake ofVLDL may play a role in triglyceride accumulation,we examined the interaction of 125l-labeled normalVLDL with the macrophages. The peritoneal macro-phages were incubated at 37°C for 5 hours with theindicated concentration of 125l-labeled normal mon-key VLDL and the uptake (binding plus incorpora-tion) and degradation of the lipoproteins were deter-mined. As shown in Figure 5A, the concentrationprofile of uptake of normal monkey 125I-VLDL by ma-crophages suggested both saturable and nonsatura-ble components. The amount of VLDL degraded(Figure 5B) was one- to two-fold more than thatwhich was taken up; this also suggested saturableand nonsaturable components. A comparison of theuptake of normal monkey and rat 125I-VLDL is madein Figure 6 A. When the lipoproteins were present atthe same protein concentration in the media, uptake(Figure 6 A) and degradation (not shown) of the
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

VLDL INCREASES MACROPHAGE TRIGLYCERIDES Bates et al. 107
500-
£ 400fe.
o
E
o
3 0 0 -
2 0 0 -
I00 -
24 HOURS
200
48 HOURS
300 400 0 100 200VLDL TRIGLYCERIDE /ml
B
300 400
Figure 3. Effect of increasing concentrations of VLDL on macrophage triglyceride con-tent. A. Macrophages were incubated with increasing concentrations of VLDL for 24 hours.o = Experiment 1. • = Experiment 2. B. Macrophages were incubated with increasingconcentrations of VLDL and harvested after 48 hours. The results are typical for two suchexperiments performed. TG = triglyceride. All experiments contained heparin (10 lU/ml).The results represent the mean and range of duplicate samples.
z£ 100-oor
o>E
IxJQ
8J 50"
O
Q:h-
u
PERITONEAL
A
//
// /B
i I
J774
/
y/
i i
P388
P B
y/
T/
/I
1 10 80 I60 0 80 160 0 80
VLDL TRIGLYCERIDE/mlI60
Figure 4. Comparison of mouse peritoneal macrophages to macrophage-like cell lines.Mouse peritoneal macrophages (Peritoneal) and two macrophage-like cell lines (J774 andP388) were incubated for 48 hours in increasing concentrations of normal rat VLDL. A and Brepresent two different experiments. The results are means of duplicate determinations.
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

108 ARTERIOSCLEROSIS VOL 4, No 2, MARCH/APRIL 1984
UPTAKE DEGRADATION
60 0 20
fj.q I 2 5 I - V L D L Protein/ml
B
40
Figure 5. Uptake (A) and degradation (B) of monkey 125I-VLDL by mouse peritonealmacrophages. The indicated concentration of monkey I-VLDL was added to peritonealmacrophages for 5 hours at 37°C. The data represent the results of four separate experi-ments. Most points are the means of duplicate cultures with a few single determinations.Degradation was not measured in all experiments.
c<v2Q.
E_i
VLD
i
n
_
1.6-
1.2-
0.8-
0.4-
0.0-
A.
J%
0
I25IVLDL Uptake
//
/
/
// O Monkey
M «Rat
1 1 1
B. Competition with RatL I 2 5 I VLDL
vN—-~-^\ \ • - ^ ^ Monkey\ \ ^ HDL
\ ^ J S L\<
Coconut OilVLDL
I 1
20 40 60 0 15 30I 2 5 I -VLDL Unlabeled Lipoprotein
-100
rol
G
-50 °H-o
• 0
(/Ltg Protein/ml)A B
Figure 6. Interaction of normal 125I-VLDL and macrophages. A. Increasing concentra-tions of normal 125I-VLDL from rhesus monkeys or rats were added to mouse peritonealmacrophages for 5 hours at 37°C. The data represent 125I-VLDL bound and incorporated bythe cells = uptake. The results are means of duplicate determinations. The data in this figurehave been published.55 B. Competition of unlabeled monkey HDL, normal rat VLDL, andcholesterol ester-rich, beta-migrating hyperlipemic VLDL from monkeys fed a diet of 25%coconut oil and 2% cholesterol with normal rat 125I-VLDL (10 /ug protein/ml) for uptake bymacrophages. The control value for the uptake of 125l-normal rat VLDL was 0.4 ;u.g VLDLprotein/mg cell protein. Lipoprotein-deficient serum (2.5 mg/ml) was present in all samples.
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

VLDL INCREASES MACROPHAGE TRIGLYCERIDES Bates et al. 109
Table 2. Effect of Human VLDL on the TriglycerideAccumulation in Macrophages
Experi-ment
Additionsto
medium
Triglyceride(/ig/mg
cell protein) No.1 None
VLDL
NoneVLDL
NoneVLDL
NoneVLDL
12.4102.6
3.755.7
0.0152.7
0.0148.5
22
24
24
23
Experiment 1. J774 cells, 300 /^g VLDL triglyceride/ml,2.5 mg albumin/ml, 48 hours of incubation. Experiments 2and 3. J774 cells, 200 /xg VLDL triglyceride/ml, 24 hours ofincubation. Experiment 4. Peritoneal macrophages, 200/u.g VLDL triglyceride/ml, 24 hours of incubation. No. =number of determinations.
Table 3. Heparln-Releasable Llpase Activity In Medi-um From Macrophage Cultures
Additions toculture medium
Cell typeAdditionsto assay None
Heparin(10 lU/ml)
J774* Serum 120 531Macrophagest — 8 167
Serum 15 463Serum + 0.5 3 34
M NaCI
Data represent the means of duplicate or triplicate deter-minations and are expressed as nmol oleic acid releasedin 60 minutes at 37°C per ml of culture medium. Rat serumheated 56°C for 30 minutes served as the source of apo-protein C-ll.
'Medium was added to the J774 macrophages for 5hours, removed, and assayed for lipase activity for a 60-minute time period.
fMedium was added to peritoneal macrophages for 24hours, removed and assayed for lipase activity for a 90-minute time period.
VLDL from the two species were virtually identical.The uptake was specific for VLDL in that the additionof excess unlabeled normal VLDL competed withitself for uptake by the macrophages (Figure 6 B) andmonkey HDL did not compete to a significant extent.However, the cholesterol ester-rich, beta-migratingVLDL isolated from monkeys fed a coconut oil pluscholesterol diet were much more effective than nor-mal VLDL in the inhibition of the uptake of labelednormal rat VLDL. Comparable results were obtainedwith 12Sl-monkey VLDL (data not shown).
The possible contribution of lipoprotein lipase totriglyceride accumulation was explored after it wasreported that J774, a macrophage-like cell line,37 hu-man monocyte macrophages,38'M and rabbit alveo-lar macrophages37'M secreted lipoprotein lipase. Todemonstrate that mouse peritoneal macrophagesalso produced a lipoprotein lipase-like enzyme, me-dium with or without heparin was added to J774 cellsand mouse peritoneal macrophages, removed andtested for lipase activity. The data in Table 3 providepartial evidence for the presence of heparin-releas-able lipase activity in medium from mouse peritonealmacrophages with characteristics comparable tothat produced by J774 cells. Furthermore, the hy-drolysis of triglyceride to free fatty acid stimulated bymedium from peritoneal macrophages was inhibited93% when 0.5 M NaCI was included in the assay.
As shown in Figure 7, the addition of heparin (10lU/ml) with VLDL (150 fj.g triglyceride/ml) resulted ina more pronounced increase of triglyceride in macro-phages, culminating in a 1.5-fold greater cellular tri-glyceride content in macrophages incubated for 48hours with VLDL in the presence of heparin than withVLDL alone (Figure 7). VLDL from either monkeys orrats was equally effective in promoting triglycerideaccumulation, which was similarly stimulated when
heparin was present in the medium (Figure 8). Asdemonstrated in Figure 7, the presence of albumininhibited the increase in intracellular triglycerides for6 hours but did not prevent its eventual accumula-tion. Table 4 summarizes the results of several ex-periments where macrophages were incubated with
HOURS
Figure 7. Effect of heparin on the triglyceride content ofmacrophages. Normal rat VLDL (150 ^g triglyceride/ml)and 10 mg/ml of bovine serum albumin were added tomouse peritoneal macrophages for the indicated time per-iods. To one group of cultures, heparin was added (10 IU/ml). The data represent results from a typical experiment ofthe three performed and are the mean and range of dupli-cate determinations.
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

110 ARTERIOSCLEROSIS VOL 4, No 2, MARCH/APRIL 1984
400-
300-
o8: 2oo-]
IOO-
+HEPARIN
DMEM
-HEPARIN
MONKEY RATVLDL
+ HEPARIN
MONKEY RATVLDL
Table 5. Oleic Acid Stimulation of the IntracellularTriglyceride Content of Macrophages
Figure 8. Effect of VLDL from rhesus monkeys or rats onthe triglyceride levels in macrophages in the absence orpresence of heparin. Normal monkey and rat VLDL (300tig triglyceride/ml) were added to mouse peritoneal macro-phages for 48 hours with or without heparin (10 IU/ml). Themonkeys were fasted before being bled while the rats werenot fasted. TG = triglyceride. The results are means ± SDof triplicate determinations.
rat VLDL with and without heparin in the presence orabsence of albumin. The effect of albumin was vari-able and ranged from a slight effect (Experiment 1) to50% suppression of triglyceride accumulation (Ex-periment 3). In all cases, however, heparin stimulat-ed triglyceride accumulation.
The hydrolysis of VLDL triglycerides by lipoproteinlipase would result in the release of free fatty acidswhich could then be taken up by the cell and reesteri-fied into triglycerides. In experiments using 200 /u.gVLDL triglyceride/ml of medium, more than 50% of
Table 4. Effect of Albumin and Heparin on Triglycer-ide Accumulation in Macrophages Incubated withVLDL
Experi-ment
1234
- Albumin
- Hep-arin
97.5200.7214.9
67.3
+ Hep-arin
137.6350.8309.6172.1
+ Albumin
- Hep-arin
82.6
77.845.1
+ Hep-arin
166.1
163.1136.1
Data are means of duplicate determinations. Albumin =bovine serum albumin, 10 mg/ml; heparin at 10 IU/ml. Experiments 1 and 2. Rat VLDL (300 ^g triglyceride/ml), 48-hour incubation; peritoneal macrophages. Ex-periment 3. Rat VLDL (200 ng triglyceride/ml), 24-hourincubation; peritoneal macrophages. Experiment 4. RatVLDL (300 ,ug triglyceride/ml), 48-hour incubation; J774cell line.
Experi-ment
1
2
3
Macro-phage
J774
J774
Peritoneal
Oleateconcen-tration(Mg/ml)
100—
100—
125
Cell tri-glyceride(jug/mg
cellprotein)
23.0150.4
9.3123.4
0203.3
Cells were incubated for 24-hours with oleate com-plexed to bovine serum albumin at a 6.8 to 1.0 molar ratio.
the triglyceride was no longer present in the mediumafter a 48-hour incubation, indicating that, theoreti-cally, 100 ju,g of fatty acid was available to the cells ifall of the VLDL triglycerides were hydrolyzed via theaction of lipoprotein lipase. The data in Table 5 showthat such levels of fatty acids added to the mediumwere capable of causing triglyceride levels to in-crease in the macrophages. After a 24-hour incuba-tion, 100 jug/ml of oleate produced a substantial in-crement in cellular triglycerides in both J774 andperitoneal macrophages.
Changes in the cholesterol content of the macro-phages during incubation with VLDL were deter-mined. As shown in Table 6, exposure of J774 mac-rophages to normal rat VLDL resulted in an increasein intracellular triglycerides but did not elevate thecholesterol ester content of the cells. After 24 hoursof incubation, the free cholesterol content of the ma-crophages increased slightly while the cholesterolester content was actually reduced from control val-ues in all three experiments. The increase in thecholesterol content of macrophages produced by hy-perlipemic monkey VLDL which has been reportedelsewhere14 is confirmed here and used for com-parative purposes.
The role of phagocytosis was examined using In-tralipid, a triglyceride-phospholipid micelle mixturecontaining no apoproteins such as apo C-ll, which isrequired as an obligatory factor for lipoprotein lipase.Since incubations were done in the absence of se-rum, the Intralipid should not be a substrate for lipo-protein lipase, although other lipases may hydrolyzethe triglycerides. When incubated with the two ma-crophage cell lines (Table 7), Intralipid caused a 17-fold stimulation in the triglyceride content of J774and a threefold increase in that of P388 macro-phages. Mouse peritoneal macrophages were alsoenriched with triglyceride after incubation with Intrali-pid (data not shown). This was not due to a nonspeci-fic adsorption of Intralipid to the cell surface since anexperiment using P388 demonstrated that there wasno triglyceride associated with the cells after a 2-hour incubation period.
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

VLDL INCREASES MACROPHAGE TRIGLYCERIDES Bates et al. 111
Table 6. Triglycerlde and Cholesterol Content of J774 Macrophages Exposed to Normollpemicor Hyperlipemic VLDL
Experi-ment
1
2
3
Additionsto medium
NoneVLDL (normolipemic)VLDL (hyperlipemic)AlbuminVLDL (normolipemic)AlbuminVLDL (normolipemic)VLDL (hyperlipemic)
Triglyceride(/xq lipid/mqcell protein)
054.36.2
10.649.46.1
156.79.0
Free
6.99.1
36.58.0
14.19.1
12.728.0
Cholesterollipid/mg cell protein)
Ester
4.10.4
31.11.40.02.41.0
39.3
Total
11.09.5
67.69.4
14.111.513.767.3
No.
23323222
J774 cells were exposed to normolipemic rat VLDL (200 ^g triglyceride/ml, 12 /xg total cholesterol/ml)or hyperlipemic monkey VLDL (31 /xg triglyceride/ml, 257 /xg total cholesterol/ml) for 24 hours. Albumin =bovine serum albumin (10 mg/ml). No. = number of determinations
Table 7. Effect of Intralipld on Triglycerlde Accumu-lation in J774 and P388 Macrophages
Cells
J774P388
Additions to medium(/xg triglyceride/mg cell protein)
None
10.5±3.259.9 ±12.5
Intralipid
174.8 + 35.5152.5 ±34.6
Cells were exposed to medium alone or medium con-taining Intralipid (300 /xg triglyceride/ml) for 48 hours. Theresults are the mean ± S.D. of four separate cultures andare representative of the six experiments performed.
Discussion
The present study demonstrated that normalVLDL produced a marked increase in the triglyceridecontent of mouse peritoneal macrophages and cer-tain macrophage-like cell lines. The stimulation oftriglyceride accumulation in macrophages was spe-cific for VLDL, since low and high density lipopro-teins caused only slight changes. The effect of VLDLwas both time- and concentration-dependent. Incontrast to macrophages, the intracellular triglycer-ide content of monkey smooth muscle cells was onlymoderately affected by VLDL. A slight elevation incellular triglyceride concentrations has been ob-served in human skin fibroblasts15'16 and bovine aor-tic endothelial cells17 after exposure to VLDL at lev-els comparable to those used in this study, whilefibroblasts derived from rat foreskin and lung werenot affected by VLDL.6 If hypertriglyceridemic humanserum15 or very high concentrations of VLDL (800 ^gVLDL triglyceride/ml) were used, substantial eleva-tions in intracellular triglycerides could be producedin human skin fibroblasts. VLDL isolated from eithertriglyceride-rich or normal human serum producedcomparable, but limited, changes in triglyceride lev-els in skin fibroblasts.16 The most pronounced re-sponse to normal VLDL was that seen in preadipo-
cytes derived from rat epididymal fat pads, whichaccumulated intracellular triglyceride levels of 2 mg/mg cell protein under specific conditions.6
Three mechanisms which might play a role in theobserved accumulation of triglycerides in macro-phages were explored in this study. They includeuptake of VLDL and its associated triglyceride viaspecific cell surface receptors; action of lipoproteinlipase on VLDL triglyceride in the media resulting inthe uptake of the liberated fatty acids by the cells andtheir incorporation into cellular triglycerides; andphagocytosis of intact triglyceride-rich VLDL. The re-sults gave suggestive evidence that all three mecha-nisms could have contributed to the elevation inmacrophage triglyceride levels. Further detailedstudies are in progress to elucidate the relative im-portance of each to the process.
The interaction of VLDL with cell surface receptorswas examined using 125l-labeled VLDL. Mouse peri-toneal marophages possess few LDL cell surfacereceptors40 but do have high affinity receptors specif-ic for both hypertriglyceridemic VLDL7 and p-VLDL,the beta-migrating cholesterol ester-rich lipoproteinsfrom cholesterol-fed animals.1314 Partial evidencefor the specific interaction of normal VLDL with mac-rophage cell surface receptors is presented in thisreport. Macrophages appeared to take up and de-grade VLDL through both saturable and nonsatura-ble processes. In addition, preliminary results indi-cated that increasing concentrations of unlabeledmonkey LDL had only a slight effect on the uptake ofrat 125I-VLDL HDL did not interfere with the uptake ofVLDL, but p-VLDL produced a marked inhibition ofthe uptake of normal triglyceride-rich VLDL. Thus,the two types of VLDL particles may have differentaffinities for the same receptor, or, alternatively, thepresence of p-VLDL may affect the uptake of normalVLDL by the macrophages. It is of interest that, al-though the VLDL from rats and monkeys containdifferent complements of apoproteins B-100, B-48and E, they both interact with the macrophage recep-tors in a similar fashion, raising questions about the
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

112 ARTERIOSCLEROSIS VOL 4, No 2, MARCH/APRIL 1984
identity of the ligand. Evidence from other laborato-ries has indicated that many cell types are capable ofincorporating either intact VLDL particles or compo-nents of VLDL. 12sl-labeled VLDL has been shown tobe taken up and degraded by smooth muscle cells,41
lymphocytes,42 and fibroblasts.1543 The radioactivetriglycerides of labeled VLDL were incorporated andmetabolized by endothelial cells,17 L cells,44 and Ehr-lich ascites cells45 without significant hydrolysis.
Several lines of evidence indicate that the pre-dominant mechanism through which triglyceridesaccumulate in macrophages is through the action oflipoprotein lipase. First, various types of macro-phages secrete lipoprotein lipase37"39 and, in thepresent study, mouse peritoneal macrophages wereshown to produce a hepan'n-releasable lipase thatwas largely inhibited by 0.5M NaCI.
Second, the addition of heparin with VLDL result-ed in a faster, more substantial increase in the intra-cellular triglyceride content of macrophages thanseen with VLDL alone. In studies using preadipocytecultures, heparin caused the release of lipoproteinlipase into the culture media.48'47 The cells then syn-thesized more lipase, resulting in a net increase inthe total quantity of lipase per culture which resultedin a greater triglyceride accumulation in the pre-adipocytes incubated with VLDL and heparin ascompared to those not exposed to heparin.48 Theheparin-stimulated accumulation of triglycerides inmacrophages probably operates through mecha-nisms similar to those described for preadipocytes.
Third, bovine serum albumin, which might act as asink for the released fatty acids, was able to tempo-rarily inhibit the elevation in triglyceride levels inmacrophages.
Fourth, free fatty acids that would be generatedfrom the hydrolysis of VLDL triglycerides by lipopro-tein lipase, were themselves able to produce triglyc-eride accumulation in macrophages.
Fifth, since the cholesterol ester-rich, triglyceride-poor p-VLDL was very effective in blocking the up-take of normal 125I-VLDL, it should have been able toprevent the intracellular triglyceride increase stimu-lated by normal VLDL, if the latter were operatingsolely through a receptor-mediated process. Howev-er, preliminary experiments indicated that co-incuba-tion of cells with normal VLDL and p-VLDL produceda level of intracellular triglyceride accumulation simi-lar to that which would have been expected in thepresence of normal VLDL alone.
Sixth, the VLDL from rats and monkeys containdifferent amounts of apoproteins B-100, B-48 and E,yet both VLDL produced equivalent levels of intracel-lular triglycerides in macrophages, suggesting thatthe triglyceride accumulation might be independentof these apoproteins. Lipoprotein lipase would onlyrequire the presence of apoprotein C-ll.
Seventh, the increase in intracellular triglyceridelevels produced by rat VLDL was not accompaniedby an elevation in the cholesterol content of the cells,suggesting that intact particles may not be taken up
by the macrophages. However, the cholesterol es-ters could have been hydrolyzed and released by thecells.
Finally, a 24-hour incubation of macrophages withincreasing concentrations of VLDL did not show thesaturation kinetics for triglyceride accumulation ex-pected of a receptor-mediated process. Thus, weconclude that the lipoprotein lipase produced bymacrophages is probably of major importance intheir accumulation of intracellular triglyceride.
Gianturco et al.7 proposed that cell surface recep-tors specific for hypertriglyceridemic VLDL were re-sponsible both for the accumulation of approximately400 fxg triglyceride/mg cell protein after a 24-hourincubation and for the appearance of oil red 0 stain-able droplets in mouse peritoneal macrophages.While some triglyceride uptake may be receptor-me-diated, our results with normal VLDL suggest that thebulk of the triglyceride accumulation in the macro-phages occurs through the action of lipoprotein li-pase. In the work reported by Gianturco et al., normalVLDL did not produce triglyceride accumulation,while the present studies demonstrated a markedincrease in intracellular levels of triglyceride uponincubation of macrophages with VLDL from normalrats, monkeys, and humans. The extent of triglycer-ide accumulation is sensitive to many experimentalconditions that could well account for the differencesbetween our results and those of Gianturco et al.7
In the work presented here the lipoproteins werefrom rats, monkeys, and humans, while the macro-phages were from mice. Since the VLDL from allthree sources produced similar enrichments in mac-rophage cellular triglycerides, it appears that spe-cies differences probably do not play a major role.However, the effect of mouse VLDL was not exam-ined and the possibility that species differences be-tween lipoproteins and cells may have affected theresults cannot be entirely ruled out.
Phagocytosis of the intact VLDL particle may alsocontribute to triglyceride accumulation in macro-phages. The elevation in intracellular triglycerides inthe cell lines J774 and P388 observed after theirincubation with Intralipid suggests the possibility ofthe apoprotein-independent uptake of micellar parti-cles which also might occur with a VLDL particle.However, further studies are needed to substantiatethis point. Since the activator of lipoprotein lipase,apoprotein C-ll, was not added to medium containingIntralipid, it is unlikely that this enzyme was involvedin the triglyceride accumulation promoted by the arti-ficial micellar particles. On the other hand, the possi-bility that the macrophage secretes apo C-ll cannotbe excluded nor can the mediation of another secret-ed lipase be ruled out.
Zilversmit49 has proposed that lipoprotein lipaseplays an important contributory role in atherogenesisdue to its conversion of VLDL to remnant particles.Such products of VLDL catabolism have manifestedenhanced interactions with fibroblasts and smoothmuscle cells in culture when compared to the original
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

VLDL INCREASES MACROPHAGE TRIGLYCERIDES Bates et al. 113
triglyceride-rich lipoprotein particle.50"53 Althoughatherosclerotic plaques contain predominantly cho-lesterol rather than triglyceride, triglyceride accumu-lation may be a transient phenomenon in the evolu-tion of the atherosclerotic lesion. The remnantsremaining after the hydrolysis of triglycerides withinthe triglyceride-rich chylomicron and VLDL particlesmay contribute to the cholesterol of the lesion cells.Such a formulation puts added emphasis on the roleof triglyceride-rich lipoproteins. The recent re-ports54' 55 that macrophages, which are important cel-lular components of the evolving atheroscleroticplaque, produce lipoprotein lipase, add support tothe notion that these cells may play an important rolein the metabolism of VLDL by the arterial wall. Wepresent evidence that lipoprotein lipase not only con-tributes to macrophage metabolism of VLDL, but thatsuch interactions result in the production of a triglyc-eride-rich cell with oil red O positive inclusions and a"foamy" appearance.
AddendumAfter this manuscript had been submitted for publication, a re-
port appeared concerning the role of lipoprotein lipase in the me-tabolism of VLDL by macrophages which supported many of ourobservations.54
AcknowledgmentsThe authors are grateful for the continued encouragement and
support of Robert W. Wissler and to Beth A. Coughlin and Lynn M.Nyquist for excellent technical assistance. We thank JaymeBorensztajn for help with the lipoprotein lipase assay.
References1. Newman HAI, Murad TM, Geer JC. Foam cells of rabbit
atheromatous lesion. Identification and cholesterol uptake Inisolated cells. Lab Invest 1971;25:586-594
2. Shlo H, Haley NJ, Fowler S. Characterization of lipid-ladenaortic cells from cholesterol-fed rabbits. III. Intracellular local-ization of cholesterol and cholesteryl ester. Lab Invest 1979;41:160-167
3. Fowler S, Berberlan PA, Shlo H, Gokffischer S, WollnskyH. Characterization of cell populations isolated from aortas ofrhesus monkeys with experimental atherosclerosis. Circ Res1980;46:520-530
4. Haley NJ, Shlo H, Fowler S. Characterization of llpld-ladenaortic cells from cholesterol-fed rabbits. I. Resolution of aorticcell populations by metrizamide density gradient centrifuga-tion. Lab Invest 1977;37:287-296
5. Smith EB. The influence of age and atherosclerosis on thechemistry of aortic intimar Part 1. The lipids. J AtherosclerosisRes 1965;5:224-240
6. de la Llera M, Kempe J, DeMartinls F, Rothblat G. Very lowdensity lipoprotein stimulation of triglyceride accumulation inrat preadipocyte cultures. Biochim Biophys Acta 1978:529:359-364
7. Glarrturco SH, Bradley WA, Gotto AM, Monisatt JD,Peavy DL. Hypertriglyceridemic very low density lipoproteinsinduce triglyceride synthesis and accumulation In mouseperitoneal macrophages. J Clln Invest 1982:70:168-178
8. Stary HC, Strong J. The fine structure of nonatheroscleroticintimal thickening, of developing and regressing atheroscle-rotic lesions at the bifurcation of the left coronary artery. In:Day CE, ed. Atherosclerosis drug discovery. New York: Ple-num Press, 1976:89-108
9. Taylor K, Schaffner T, Wissler RW, Glagov S. Immunomor-phologic identification and characterization of cells derivedfrom experimental atherosclerotic lesions. Scan Electron Mi-crosc 1979:3:815-822
10. Schaffner T, Taylor K, Bartuccl EJ, Ftscher-Dzoga K, Bee-son JH, Glagov S, Wissler RW. Arterial foam cells withdistinctive immunomorphologic and histochemical features ofmacrophages. Am J Pathol 1980:100:57-80
11. Fowler S, Shlo H, Haley NJ. Characterization of lipid-ladenaortic cells from cholesterol-fed rabbits. IV. Investigation ofmacrophage-like properties of aortic cell populations. Lab In-vest 1979:41:372-378
12. Day A. Upid metabolism by macrophages and its relationshipto atherosclerosis. In Paoletti R, Kritchevsky D, eds. Ad-vances in lipid research, vol 5. New York: Academic Press1967:185-207
13. Goldstein JL, Ho YK, Brown MS, Innerarlty TL, MahleyRW. Cholesterol ester accumulation In macrophages reultingfrom receptor-mediated uptake and degradation of hypercho-lesterolemic canine 6-very low density lipoproteins. J BlolChem 1980:255:1839-1848
14. Mahley RW, Innerarlty TL, Brown MS, Ho YK, GoldsteinJL Cholesterol ester synthesis in macrophages: stimulationby S-very low density lipoproteins from cholesterol-fed ani-mals of several species. J Lipid Res 1980;21:970-980
15. Oram JF, Shafrlr E, Blerman EL. Triacylglycerol metabo-lism and triacylglycerol lipase activities of cultured humanskin fibroblasts. Biochim Biophys Acta 1980,619:214-227
16. de la Llera M, Rothblat G, Howard BV. Cell triacylgrycerolaccumulation from very low density lipoproteins isolated fromnormal and hypertrigryceridemic human sera. Biochim Bio-phys Acta 1979:574:414-422
17. Howard BV. Uptake of very low density lipoprotein triglycer-ide by bovine aortic endothelial cells in culture. J Lipid Re-search 1977:18:561-571
18. Conn ZA, Benson B. The differentiation of mononuclearphagocytes. J Exp Med 1965:121:153-169
19. Bllgh EG, Dyer WJ. A rapid method of total lipid extractionand purification. Can J Biochem Physiol 1959:37:911-917
20. Ralph P, Prlchard J. Cohn M. Reticulum cell sarcoma: aneffector cell in antibody-dependent cell-mediated immunity. JImmunol 1975:114:898-905
21. Koren HS, Handwerger BS, Wunderllch JR. Identificationof macrophage-like characteristics in a cultured murine tumorline. J Immunol 1975;114:894-897
22. Rscher-Dzoga K, Jones RM, Vessellnovltch D, WisslerRW. Ultrastructural and Immunohistochemical studies of pri-mary cultures of aortic medial cells. Exp Mol Pathol 1973;18:162-176
23. Bates SR, Wissler RW. Effect of hyperiipemic serum oncholesterol accumulation in monkey aortic medial cells. Bio-chim Biophys Acta 1976;450:78-88
24. Havel RJ, Eder HA, Bragdon JH. The distribution andchemical composition of ultracentrifugalry-separated lipopro-teins in human serum. J Clin Invest 1955;34:1345-1353
25. Scanu AM, Edelsteln C, Vltello L, Jones R, Wissler RW.The serum high density lipoproteins of Macacus rhesus. Part1. Isolation, composition, and properties. J Biol Chem, 1973;248:7648-7652
26. Fleas GM, Wissler RW, Scanu AM. Study of abnormal plas-ma low-density lipoproteins In rhesus monkeys with diet-in-duced hyperiipldemia. Biochemistry 1976;15:5799-5804
27. Lowry OH, Rosebrough NJ, Farr LA, Randall RJ. Proteinmeasurement with the Folin phenol reagent. J Biol Chem1951:193:265-275
28. Bartlett GR. Phosphorus assay In column chromatography. JBiol Chem 1959:234:466-468
29. Lipid Research Clinics Program. Manual of Laboratory Op-erations vol 1. Upid and lipoprotein analysis DHEW no. (NIH)
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

114 ARTERIOSCLEROSIS VOL 4, No 2, MARCH/APRIL 1984
75-628. Bethesda, Maryland: National Institutes of Health1974:38
30. Marsh JB, Welnsteln DB. Simple charring method for deter-mination of lipids. J Lipid Res 1966;7:574-576
31. Krltchevsky D, Davidson LM, Kim HK, Malhotra S. Quanti-tation of serum lipids by a simple TLC-charring method. ClinChem Ada 1973;46:63-68
32. St. Clalr RW, Smith BP, Wood LL. Stimulation of cholesterolesterification in rhesus monkey smooth muscle cells. CircRes 1977;40:166-173
33. Nllsaon-Ehle P, Schotz MC. A stable, radioactive substrateemulsion for assay of lipoprotein lipase. J Lipid Res 1976;17:536-541
34. Kartln JB, Juhn DJ, Floss G, Scanu AM, Rubensteln AH.Measurement of rhesus monkey (Macaca mulatta) apolipo-protein B in serum by radioimmunoassay. Comparison ofimmunoreactlvities of rhesus and human low density lipopro-teins. J Lipid Res 1978; 19:197-206
35. Elsele B, Bates SR, Wlssler RW. Interaction of low densitylipoproteins from normal and hyperlipemic rhesus monkeyswith arterial smooth muscle cells in culture. Atherosclerosis1980;36:9-24
36. Brown MS, Goldstein JL. Analysis of a mutant strain ofhuman fibroblasts with a defect in the Internalizatlon of recep-tor bound low density lipoprotein. Cell 1976;9:663-674
37. Khoo JC, Mahoney EM, Wltztum JL Secretion of lipopro-tein lipase by macrophages in culture. J Biol Chem 1981 ;256:7105-7107
38. Mahoney EJ, Khoo JC, Steinberg D. Lipoprotein lipase se-cretion by human monocytes and rabbit alveolar macro-phages in culture. Proc Natl Acad Sci USA 1982;79:1639-1642
39. Chart A, Iverius P, Brunzell JD. Lipoprotein lipase secretionby human monocyte-derived macrophages. J Clin Invest1982;69:49O-493
40. Goldstein JL, Yo YK, Basu SK, Brown MS. Binding site onmacrophages that mediates uptake and degradation of ace-tylated low density lipoprotein producing massive cholesteroldeposition. Proc Natl Acad Sci 1979;76:333-337
41. Albers JJ, Blerman EL. The influence of lipoprotein compo-sition on binding, uptake and degradation of different lipopro-tein fractions by cultured human artenal smooth muscle cells.Artery 1976;2:337-348
42. Poyser A, Nestel PJ. Metabolism of very low density lipopro-teins by human mononuclear cells. Artery 1979;6:122-143
43. Glanturco SH, Brown FB, Gotto AM, Bradley WA. Recep-
tor-mediated uptake of hypertriglyceridemic very low densitylipoproteins by normal human fibroblasts. J Lipid Res 1982;23:984-993
44. Bailey JM, Howard BV, Tlllman SF. Lipid metabolism incultured cells. XI. Utilization of serum triglycerides. J BiolChem 1973;248:1240-1247
45. Brennenman DE, Spector AA. Utilization of ascites plasmavery low density lipoprotein triglycerides by Ehrlich cells. JLipid Res 1974;15:309-316
46. Rothblat GH, DeMartlnls FD. Release of lipoprotein lipasefrom rat adipose tissue cells grown in culture. BiochemBiophys Res Comm 1977;78:45-50
47. Gllck JM, Rothblat GH. Effects of metabolic inhibitors on thesynthesis and release of lipoprotein lipase in cultured cellsderived from the stromal-vascular fraction of rat adipose tis-sue. Biochim Biophys Acta 1980;618:163-172
48. de la Llera M, Gllck JM, Rothblat G. Mechanism of triglycer-ide accumulation in rat preadipocyte cultures exposed to verylow density lipoprotein. J Lipid Res 1981;22:245-253
49. Zllversmlt DB. Role of triglyceride-rich lipoproteins in athero-genesis. Ann NY Acad Sci 1976;275:138-144
50. Schonfeld G, Patsch W, Pileger B, Wltztum JL, WeldmanSW. Lipolysis produces changes in the immunoreactivity andcell reactivity of very low density lipoproteins. J Clin Invest1979;64:1288-1297
51. Floren C, Albers JJ, Blerman EL. Uptake of chylomicronremnants causes cholesterol accumulation in cultured humanarterial smooth muscle cells. Biochim Biophys Acta 1981;663:336-349
52. Blerman EL, Elsenberg S, Stein O, Stein Y. Very low densi-ty lipoprotein "remnant" particles: uptake by aortic smoothmuscle cells in culture. Biochim Biophys Acta 1973;329:163-169
53. Catapano AL, Glanturco SH, Klnnunen PKJ, Elsenberg S,Gotto AM, Smith LC. Suppression of 3-hydroxy-3-methyl-glutaryl-CoA reductase by low density lipoproteins producedin vitro by lipoprotein lipase action on nonsuppressive verylow density lipoproteins. J Biol Chem 1982;254:1007-1009
54. Llndqvlst P, Ostlund-Llndqvlst A, Wltztum JL, SteinbergD, Little JA. The role of lipoprotein lipase in the metabolism oftriglyceride-rich lipoproteins by macrophages. J Biol Chem1983;258:9O86-9092
55. Feng Z, Bates SR, Getz GS, Wlssler RW. The receptor-mediated accumulation of triglyceride in macrophage ex-posed to very low density lipoproteins. Acta Acad Med Wuhan1983;3:8-15
Index Terms: very low density lipoprotein • macrophage • triglyceride • atherosclerosis • lipoprotein lipase
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from

S R Bates, P L Murphy, Z C Feng, T Kanazawa and G S GetzVery low density lipoproteins promote triglyceride accumulation in macrophages.
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1984 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleArteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.4.2.1031984;4:103-114Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/4/2/103World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document.Permissions and Rights Question and AnswerFurther information about this process is available in theis being requested is located, click Request Permissions in the middle column of the Web page under Services.Clearance Center, not the Editorial Office. Once the online version of the published article for which permission
can be obtained via RightsLink, a service of the CopyrightArteriosclerosis, Thrombosis, and Vascular Biology Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on May 18, 2018
http://atvb.ahajournals.org/D
ownloaded from