vol. printed in purification and properties of penicillin...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Jan., 1967, p. 302-308
Copyright 1967 American Society for Microbiology
Purification and Properties of Penicillin Amidasefrom Bacillus megaterium
C. CHIANG AND R. E. BENNETTSquibb Institute for Medical Research, New Brunswick, New Jersey
Received for publication 18 July 1966
ABSTRACT
A penicillin amidase, obtained from the exogenous medium of a Bacillus mega-
terium culture, was purified approximately 96-fold by means of two cycles of adsorp-tion on, and elution from, Celite, followed by a further fractionation on carboxy-methylcellulose. On the basis of sedimentation centrifugation analysis, the finalpreparation was deemed to be homogeneous with an apparent molecularweight of approximately 120, 000. The enzyme is specific for benzylpeni-cillin and has a pH optimum between 8 and 9. Complete hydrolysis of benzylpeni-cillin was obtained at low substrate concentrations. At higher substrate con-
centrations, the hydrolysis of benzylpenicillin was incomplete, apparently due toenzyme inhibition by phenylacetic acid and 6-aminopenicillanic acid, which were
formed during the hydrolysis. Under the assay conditions, phenylacetic acid was a
competitive inhibitor of penicillin amidase with an inhibitor constant (Ki) of 0.45 M,
whereas 6-aminopenicillanic acid was noncompetitive in nature with a Ki of 2.6 X10-2 M. The Michaelis constant of this enzyme was found to be 4.5 X 10-3 M whenbenzylpenicillin was used as substrate.
An enzyme which hydrolyzes penicillins to givecarboxylic acid and 6-aminopenicillanic acid hasbeen widely demonstrated in bacteria, yeast, andfilamentous fungi (3, 8, 10, 14, 16, 17; R. C.Erickson and R. E. Bennett, Bacteriol. Proc., p.
65, 1961), as well as in plants (H. E. Alburn,N. H. Grant, and D. E. Clark, U.S. Patent3,032,473, 1962) and in animals (G. A. Weit-nauer, U.S. Patent 3,070,511, 1962). The enzymehas been given different names by various investi-gators, depending upon the particular experi-mental conditions and the results obtained (4, 9,12). According to the source from which it isisolated, the penicillin amidase may be classifiedinto two types. In one type, the enzyme, obtainedfrom fungi and Streptomyces species, hydrolyzesphenoxymethylpenicillin much more readily thanit does benzylpenicillin. In the second type, theenzyme, isolated from bacterial species, hydro-lyzes benzylpenicillin much more readily thanphenoxymethylpenicillin. Most of the publishedstudies are concerned either with a crude cell-freepreparation or with intact cells. This paper is a
report on the purification and some characteris-tics of an extracellular penicillin amidase pro-duced by a strain of Bacillus megaterium ATCC14945.
MATERIALS AND METHODS
Substrates. The potassium benzylpenicillin used inthese experiments was commerical grade material pro-duced by E. R. Squibb & Sons, New Brunswick, N.J.,with a potency of 1,500 units per mg. The ammoniumsalts of A2-pentenyl- and n-heptyl penicillins, phenoxy-methylpenicillin (free acid), allylmercaptomethyl-penicillin (penicillin 0), ethylthiomethylpenicillin,phenoxyethylpenicillin, 2(phenylthio)-ethylpenicillin,dimethoxyphenylpenicillin, and 5-methyl-3-phenyl-4-isoxazolylpenicillin were obtained from the collectionofthe Squibb Institute.
Determination ofenzymatic activity. The enzymaticactivity of the penicillin amidase preparations was de-termined by measuring the amount of 6-aminopenicil-lanic acid or phenylacetic acid produced from a re-
action mixture containing 10 mg of benzylpenicillinin 1 ml of 0.1 M borate buffer, pH 8.7, and the enzymeafter 20 min of incubation at 37 C. The hydroxyl-amine method of Batchelor et al. (1) was used fordetermination of the 6-aminopenicillanic acid, andthe amount of phenylacetic acid was determined by thegas chromatographic method of Niedermayer (15). Aunit of enzymatic activity is defined as the amount ofenzyme required to produce 1 ,umole of 6-amino-penicillanic acid (or phenylacetic acid) per hr underthese test conditions. The rate of hydrolysis was linear
between 0 and 10 units of enzyme per ml during the
302
Vol. 93, No. 1Printed In US.A.
on July 17, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

PENICILLIN AMIDASE FROM BACILLUS MEGATERIUM
first 30 min of incubation. Specific activity is definedas units of enzyme activity per milligram of protein.
Determination of protein and of ammonia. Proteinwas determined on enzyme preparations by use of themethod ofLowry et al. (13), or spectrophotometricallyby the method of Warburg and Christian (19) withbovine serum albumin as standard. The proteincontent in crude broth supernatant fluid was de-termined after the protein had been precipitated with10% trichloroacetic acid and taken into 0.1 N NaOHsolution. In those experiments in which the effect ofthe penicillin amidase on various amides was studied,the amount of ammonia released from the amides wasdetermined by the method of Conway (6).
Growth of B. megaterium. The culture of B. mega-terium ATCC 14945, obtained from S. Murao(University of Tottori, Tottori City, Japan), wasmaintained on agar slants and as lyophilized cellsuspensions. For large-scale experiments, the culturewas grown in 30-liter fermentors according to themethod described in Example 3 by Murao et al. (U.S.Patent 3,144,395, 1964), in which phenylacetic acid isadded to induce maximal production of penicillinamidase. After an incubation period of 70 to 72 hr, thefermentation broth was harvested and passed througha Westphalia centrifuge to remove the solids. Thecell-free supernatant liquid was retained for recoveryof the penicillin amidase.
Purification of enzyme. A batch of 34 liters ofsupernatant liquid, containing 20 enzyme units per ml,was acidified with dilute acetic acid to pH 6.4, andwas mixed with acid-washed Celite (no. 545, Johns-Manville Co., New York, N.Y.) at a ratio of 15 g perliter. The suspension was stirred for 2 hr, during whichperiod sufficient acetic acid was added to maintain thepH at 6.4. The Celite was collected by filtration andwas washed once with 3.4 liters of distilled water. Ap-proximately 95% of the original enzyme activity wasrecovered on the Celite, and less than 1% was lost bythe water wash. The Celite cake was slurried in 24%ammonium sulfate (w/v) in 0.1 M tris(hydroxymethyl)-aminomethane (Tris) buffer (adjusted to pH 8.4 withNH40H) and was transferred to a glass column ofsuitable size (Celite column I). Elution of the enzymefrom the Celite column I was carried out by furtheraddition of the ammonium sulfate solution, and com-plete elution was obtained when approximately 2 litersof eluate had been collected. The eluate was concen-trated in vacuo by maintaining it at 40 C in a waterbath. The protein gradually precipitated from thesolution, and the evaporation was halted at the firstappearance of ammonium sulfate crystals. The pre-cipitated protein was separated by filtration and wasresuspended in a minimal amount of 0.05 M phosphatebuffer, pH 7.0. The suspension was dialyzed against a50-fold volume of 0.025 M phosphate buffer (pH 6.4),with three changes of buffer in a 24-hr period. The finalsolution was clarified by centrifugation, with a totalyield of 58 ml.The enzyme was further purified by fractionation on
a carboxymethylcellulose column at 4 C. The columnwas prepared by applying pressure at 15 psi and by
extensive washing with 0.025 M phosphate buffer, pH6.4. A 45-ml amount of a solution containing 6 mg ofprotein per ml, which had been obtained from the pre-vious step, was placed on a cellulose column (4.7 by47 cm). The column was continuously eluted at a flowrate of 100 ml/hr by the addition of 0.025 M phosphatebuffer, pH 6.4. About 750 ml of eluate was collected,and most of the impurities and pigmented materialswere contained in this eluate. At this point, the elutingsolution was changed to 0.1 M phosphate buffer, pH6.5. Then, another 460 ml of eluate was collected be-fore any appreciable activity was eluted. A subsequent565-ml fraction, which contained most of the enzymeactivity, was obtained. The pooled active eluate wasmixed with acid-washed Celite at a ratio of 75 g ofCelite per liter and was stirred for 1 hr. The Celite,upon which the enzyme was adsorbed, was collected,slurried with 24% ammonium sulfate in 0.1 M Trisbuffer (pH 8.4), and transferred to a glass column(Celite column II). Additional ammonium sulfatesolution was added to the column until 4 ml of eluateper g of Celite had been collected. This latter eluatewas concentrated in vacuo, and the precipitated pro-tein was recovered as described above for Celite col-umn I. The protein was suspended in a minimalamount of 0.05 M phosphate buffer, pH 7.0, and wasdialyzed against a 200-fold volume of the same bufferwith three changes over a 16-hr period. The final vol-ume of this solution was 10 ml. This preparation wasused in most of the studies.
RESULTS
Purification. The data obtained in a typicalenzyme purification experiment are presented inTable 1. The first cycle of Celite adsorption wasevidently the most efficient step, since 95%c of theenzyme was recovered from the fermentationbroth in this fraction. During the process of con-centrating the ammonium sulfate solution, it wasfrequently observed that a considerable amountof protein was coagulated on the liquid surfaceas insoluble material. This denatured protein nodoubt accounts for the significant loss in enzymeactivity in this step. In subsequent small-scaleexperiments, the percentage of recovery wasusually higher than that obtained on the largecolunm described above.
Ultracentrifuge studies. The final enzyme prep-arations were found to be homogeneous in molec-ular species when samples were examined in aSpinco model E ultracentrifuge equipped with aSchlieren diaphragm at an angle of 70°. In atypical experiment, in which the protein concen-tration was 0.8% in 0.05 M phosphate buffer (pH7.0) after reaching a maximal speed of 44,700rev/min for a period of 64 min at a temperatureof 24 C, a single component was found in thesedimentation pattern. Figure 1 illustrates thesedimentation patterns taken at various time
303VOL. 93, 1967
on July 17, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

TABLE 1. Purification of penicillin amidase
SpecificFraction Vol ~~~~Total Total activity Puri- RFraction Vol ; activity( protein (units per fication Recovery
ml units mg %Crude broth supernatant fluid............ 34,000 4.56 23,000 19.8 1 100First-cycle Celite adsorption and ammonium sulfate
precipitation................................... 58 3.15 340 930 47 69Carboxymethylcellulose column fractionation . ... 565 2.36 160 1,470 75 52Second-cycle Celite adsorption and ammonium sul-
fate precipitation............................... 10 1.53 81 1,900 96 33
a Unit of enzyme activity is defined as the amount of enzyme required to produce 1 ,umole of 6-amino-penicillanic acid (or phenylacetic acid) per hr in 0.1 M borate buffer (pH 8.7) at 37 C.
~CrFIG.. Sedimntationpatternof thepurifedpenicilin amidse. Picures tkenat.6,32,.nd.64mn.afte
......n44,00. ev/in.t.2.C
periods. The sedimentation constant of this en-zyme protein was calculated from the data to be5.5 Svedberg units. The apparent molecularweight was estimated to be about 120,000 in aseparated sedimentation-equilibrium experimentconducted at 9,341 rev/min at 20 C for 47 hr,according to the method employed by Van Holdeand Baldwin (18). (Thanks are due to James B.Ifft for his assistance in this experiment.) Thepartial specific volume of this protein was as-sumed to be 0.75 ml/g (7).
Stability. Samples of purified penicillin amidasein 0.05 M phosphate buffer, pH 7.0, were stablewhen stored at 4 C for a 3-month period. Nochange in activity was noted when enzyme sam-ples were diluted with 0.1 M borate buffer, pH8.4, to a protein concentration of 1 mg/ml andwere stored for 8 hr at -20, 5, 24, and 37 C.With the same protein concentration, 70% of theenzyme activity was lost at 45 C, and the enzymewas completely denatured at 60 C. A study ofpenicillin amidase stability in solutions of differ-ent pH is summarized in Table 2. Maximal en-
zyme stability was observed at 24 C for 9 hr overthepH range of 6.5 to 10.05.
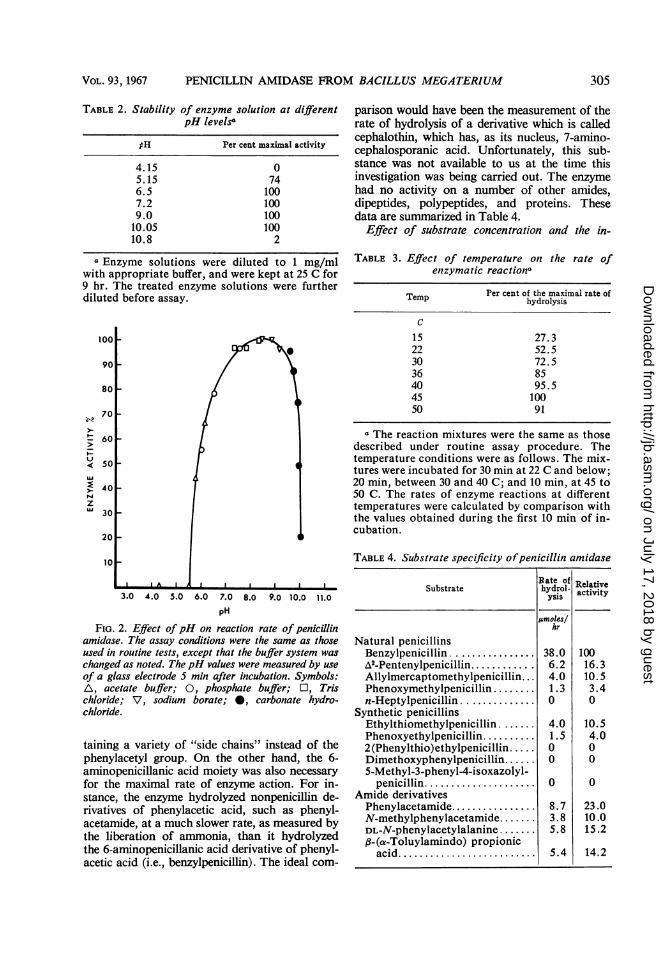
Optimal pH and temperature. The enzyme hasan optimal pH range between 8 and 9. A rapiddecrease in activity was observed when the pHwas raised above 9.5 or lowered below 7.5 (seeFig. 2). The decrease in activity was not the re-sult of chemical inactivation of substrate (benzyl-penicillin) or of product (6-aminopenicillanicacid), since both compounds were found to beunchanged after incubation at 37 C for 20 min insolutions more alkaline than pH 10 and moreacidic thanpH 5.The effect of temperature on the initial rate of
enzyme reaction is summarized in Table 3. Al-though a temperature of 45 C was optimal forenzymatic activity, it was not suitable for routineuse, because the enzyme protein was considerablyinactivated at this temperature.
Substrate specificity. The penicillin amidase ofB. megaterium was found to be rather specific forbenzylpenicillin. Under the same assay conditions,it was much less active on other penicillins con-
304 CHIANG AND BENNETI' J. BACrERIOL.
on July 17, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
VOL. 93, 1967 PENICILLIN AMIDASE FROM BACILLUS MEGATERIUM
TABLE 2. Stability of enzyme solution at differentpH levels"
pH Per cent maximal activity
4.15 05.15 746.5 1007.2 1009.0 10010.05 10010.8 2
Enzyme solutions were diluted to 1 mg/mlwith appropriate buffer, and were kept at 25 C for9 hr. The treated enzyme solutions were furtherdiluted before assay.
100
901-
80-
701-
60
<50
t 40
z,,, I
)F
)1_
30 _-
201_
10
3.0 4.0 5.0 6.0 7.0 8.0 9.0 10.0 11.0
pH
FIG. 2. Effect of pH on reaction rate of penicillinamidase. The assay conditions were the same as thoseused in routine tests, except that the buffer system waschanged as noted. ThepH values were measured by useof a glass electrode S min after incubation. Symbols:A\, acetate buffer; 0, phosphate buffer; O, Tris
chloride; V, sodium borate; *, carbonate hydro-chloride.
taining a variety of "side chains" instead of thephenylacetyl group. On the other hand, the 6-aminopenicillanic acid moiety was also necessaryfor the maximal rate of enzyme action. For in-stance, the enzyme hydrolyzed nonpenicillin de-rivatives of phenylacetic acid, such as phenyl-acetamide, at a much slower rate, as measured bythe liberation of ammonia, than it hydrolyzedthe 6-aminopenicillanic acid derivative of phenyl-acetic acid (i.e., benzylpenicillin). The ideal com-
parison would have been the measurement of therate of hydrolysis of a derivative which is calledcephalothin, which has, as its nucleus, 7-amino-cephalosporanic acid. Unfortunately, this sub-stance was not available to us at the time thisinvestigation was being carried out. The enzymehad no activity on a number of other anides,dipeptides, polypeptides, and proteins. Thesedata are summarized in Table 4.
Effect of substrate concentration and the in-
TABLE 3. Effect of temperature on the rate ofenzymatic reactiona
Temnp Per cent of the maximal rate ofhydrolysis
C
15 27.322 52.530 72.536 8540 95.545 10050 91
The reaction mixtures were the same as thosedescribed under routine assay procedure. Thetemperature conditions were as follows. The mix-tures were incubated for 30 min at 22 C and below;20 min, between 30 and 40 C; and 10 min, at 45 to50 C. The rates of enzyme reactions at differenttemperatures were calculated by comparison withthe values obtained during the first 10 min of in-cubation.
TABLE 4. Substrate specificity ofpenicillin amidase
SubstrateRat ofRelativeSubstrate hydrol- atvt
ysis ativity
pmoleslhr
Natural penicillinsBenzylpenicillin. 38.0 100A'-Pentenylpenicillin........... . 6.2 16.3Allylmercaptomethylpenicillin... 4.0 10.5Phenoxymethylpenicillin ........ 1.3 3.4n-Heptylpenicillin .............. 0 0
Synthetic penicillinsEthylthiomethylpenicillin.. 4.0 10.5Phenoxyethylpenicillin .......... 1.5 4.02 (Phenylthio) ethylpenicillin .. 0 0Dimethoxyphenylpenicillin.a. . . 0 05-Methyl-3-phenyl-4-isoxazolyl-
penicillin ................... 0 0Amide derivativesPhenylacetamide................ 8.7 23.0N-methylphenylacetamide....... 3.8 10.0DL-N-phenylacetylalanine ....... 5.8 15.2fl-(a-Toluylamindo) propionic
acid...................... 5.4 14.2
305
0
1
1 1 A I I I I
L- on July 17, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
CHIANG AND BENNEIT
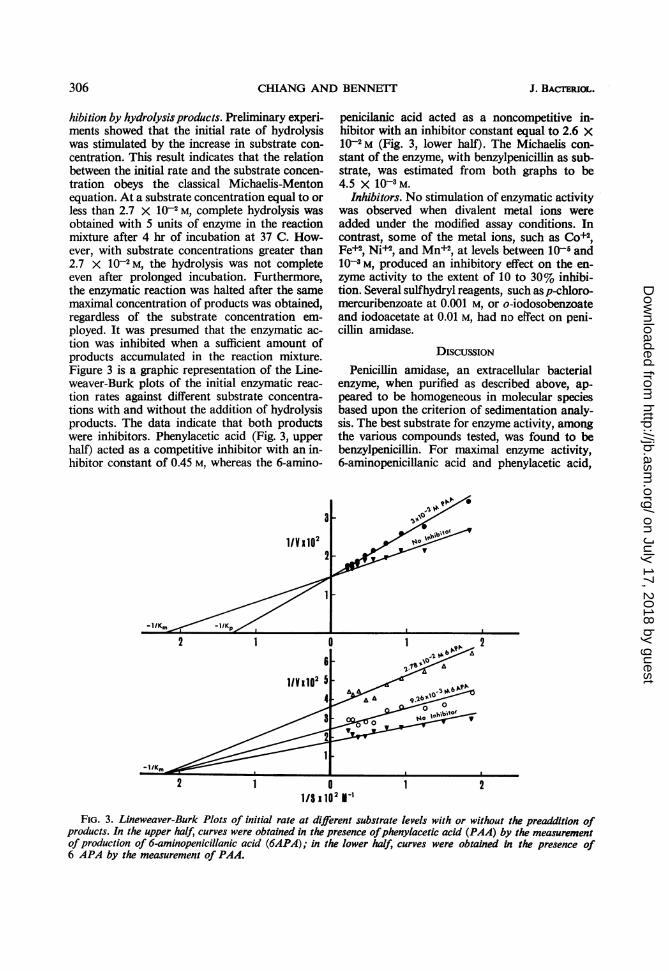
hibition by hydrolysis products. Preliminary experi-ments showed that the initial rate of hydrolysiswas stimulated by the increase in substrate con-centration. This result indicates that the relationbetween the initial rate and the substrate concen-tration obeys the classical Michaelis-Mentonequation. At a substrate concentration equal to orless than 2.7 X 102 M, complete hydrolysis wasobtained with 5 units of enzyme in the reactionmixture after 4 hr of incubation at 37 C. How-ever, with substrate concentrations greater than2.7 X 102 M, the hydrolysis was not completeeven after prolonged incubation. Furthermore,the enzymatic reaction was halted after the samemaximal concentration of products was obtained,regardless of the substrate concentration em-ployed. It was presumed that the enzymatic ac-tion was inhibited when a sufficient amount ofproducts accumulated in the reaction mixture.Figure 3 is a graphic representation of the Line-weaver-Burk plots of the initial enzymatic reac-tion rates against different substrate concentra-tions with and without the addition of hydrolysisproducts. The data indicate that both productswere inhibitors. Phenylacetic acid (Fig. 3, upperhalf) acted as a competitive inhibitor with an in-hibitor constant of 0.45 M, whereas the 6-amino-
penicilanic acid acted as a noncompetitive in-hibitor with an inhibitor constant equal to 2.6 X10-2 M (Fig. 3, lower half). The Michaelis con-stant of the enzyme, with benzylpenicillin as sub-strate, was estimated from both graphs to be4.5 X 10- M.
Inhibitors. No stimulation of enzymatic activitywas observed when divalent metal ions wereadded under the modified assay conditions. Incontrast, some of the metal ions, such as Co+2,Fe+2, Ni+2, and Mn+2, at levels between 10-6 and10-3 M, produced an inhibitory effect on the en-zyme activity to the extent of 10 to 30% inhibi-tion. Several sulfhydryl reagents, such as p-chloro-mercuribenzoate at 0.001 M, or o-iodosobenzoateand iodoacetate at 0.01 M, had no effect on peni-cillin amidase.
DISCUSSIONPenicillin amidase, an extracellular bacterial
enzyme, when purified as described above, ap-peared to be homogeneous in molecular speciesbased upon the criterion of sedimentation analy-sis. The best substrate for enzyme activity, amongthe various compounds tested, was found to bebenzylpenicillin. For maximal enzyme activity,6-aminopenicillanic acid and phenylacetic acid,
0
1/8 1 102 1-1
FIG. 3. Lineweaver-Burk Plots of initial rate at different substrate levels with or without the preaddition ofproducts. In the upper half, curves were obtained in the presence ofphenylacetic acid (PAA) by the measurementofproduction of 6-aminopenicillanic acid (6APA); in the lower half, curves were obtained in the presence of6 APA by the measurement of PAA.
306 J. BACTERIOL.
on July 17, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 93, 1967 PENICILLIN AMIDASE FROM BACILLUS MEGATERIUM
which are moieties of benzylpenicillin, were re-
quired. Although this penicillin amidase couldutilize as substrate various penicillins other thanbenzylpenicillin, or certain derivatives of phenyl-acetic acid, the reaction rates in these experi-ments were much slower. Therefore, in view ofthe substrate specificity required for penicillinamidase activity, it appears unlikely that this en-zyme can be classified as either a deacylase, as
suggested by Cole (5), or an acyl transferase, assuggested by Kaufmann and Bauer (11). For thesame reasons, the more correct nomenclature forthis enzyme would appear to be penicillin amidase,according to Sakaguchi and Murao (17), ratherthan benzylpenicillin acylase, as suggested byHuang et al. (9).Although no attempt was made to measure the
residual enzyme bound on the cells, most of theenzyme appears to be excreted into the exogenousmedium. The maximal growth of the bacterialcells occurred after an incubation period of 20 to24 hr. At this time, the extracellular enzyme con-centration began gradually to increase, and con-tinued during the observation period of 96 hr.There was no evidence of lysis in the bacteriathroughout the incubation period.The role played by penicillin amidase in the
metabolism of B. megaterium is still obscure. Al-though the cell releases this penicillin-inactivatingenzyme into the medium, the growth of the or-
ganism remains sensitive to benzylpenicillin. Theinhibition of bacterial growth by benzylpenicillinwas observed by use of several media containingantibiotic concentrations as low as 0.1 ,g/ml(Basch, personal communication).
Batchelor et al. (2) observed that, although6-aminopenicillanic acid inhibited the enzymatichydrolysis of phenoxymethylpenicillin by Strepto-myces lavendulae BRL. 198, phenoxyacetic acidwas without effect. However, as noted above,both 6-aminopenicillanic acid and phenylaceticacid were shown to inhibit penicillin amidase ob-tained from B. megaterium. Since penicillin ami-dase of B. megaterium and the enzyme obtainedfrom S. lavendulae are both penicillin-hydrolyzingenzymes, these results appear to be inconsistent.This seeming inconsistency may be resolved if oneconsiders that the inhibition of the enzyme ob-tained from S. lavendulae by 6-aminopenicillanicacid could be easily demonstrated because of thenoncompetitive character of the compound, withan inhibitor constant close to the value of theMichaelis constant of the enzyme. Conversely,the inhibitory effect of phenylacetic acid may nothave been detected, because it would appear tobe a competitive inhibitor of the enzyme with a
much greater inhibitor constant.
ACKNOWLEDGMENTS
We are grateful to W. R. Frazier for his assistancein growing the large batches of bacterial culture, to S.Murao for the culture of B. megaterium, and to W. W.Wainio of Rutgers University for the use of the Spincomodel E ultracentrifuge. Thanks are also due to D.Perlman and P. Arnow for reading the manuscript.
LITERATURE CITED
1. BATCHELOR, F. R., E. B. CHAIN, T. L. HARDY, K.R. L. MANSFORD, AND G. N. ROLINSON. 1961.6-Aminopenicillanic acid. III. Isolation andpurification. Proc. Roy. Soc. (London) Ser.B 154:498-508.
2. BATCHELOR, F. R., E. B. CHAIN, M. RICHARDS,AND G. N. ROLINSON. 1961. 6-Aminopenicil-lanic acid. VI. Formation of 6-aminopenicil-lanic acid from penicillin by enzymic hydrolysis.Proc. Roy. Soc. (London) Ser. B 154:522-531.
3. CLARIDGE, C. A., A. GOUREVITCH, AND J. LEIN.1960. Bacterial penicillin amidase. Nature 187:237-238.
4. CLARIDGE, C. A., J. R. LuTTINGER, AND J. LEIN.1963. Specificity of penicillin amidases. Proc.Soc. Exptl. Biol. Med. 113:1008-1012.
5. COLE, M. 1964. Properties of penicillin deacylaseenzyme of Escherichia coli. Nature 203:519-520.
6. CONWAY, E. J. 1947. Microdiffusion analysis andvolumetric error. Cosby Lockwood and Sons,Ltd., London, England.
7. EDSALL, J. T. 1953. The size, shape and hydrationof protein molecules, p. 549. In H. Neurath andK. Bailey [ed.], The proteins, vol. I, part B.Academic Press, Inc., New York.
8. HUANG, H. T., A. R. ENGLISH, T. A. SETO, G. M.SHULL, AND B. A. SOBIN. 1960. Enzymatic hy-drolysis of the side chain of penicillins. J. Am.Chem. Soc. 82:3790-3791.
9. HUANG, H. T., T. A. SETO, AND G. M. SHULL.1963. Distribution and substrate specificity ofbenzylpenicillin acylase. Appl. Microbiol. 11:1-6.
10. KAMEDA, Y., Y. KIMURA, E. TOYOURA, AND T.OMARI. 1961. A method for isolating bacteriacapable of producing 6-aminopenicillanic acidfrom benzylpenicillin. Nature 191:1122-1123.
11. KAUFMANN, W., AND K. BAUER. 1964. Variety ofsubstrates for a bacterial benzylpenicillin-splitting enzyme. Nature 203:520.
12. KAUFMANN, W., K. BAUER, AND H. OFFE. 1960.Enzymatic cleavage and resynthesis of penicil-lins. Antimicrobial Agents Ann., p. 1-5.
13. LOWRY, 0. H., N. J. ROSEBROUGH, A. L. FARR,AND R. J. RANDALL. 1951. Protein measure-ment with the Folin phenol reagent. J. Biol.Chem. 193:265-275.
14. MURAO, S., AND Y. KISHIDA. 1961. Studies onpenicillin amidase. Part V. Studies on the peni-cillin-amidase producing microorganisms. J.Agr. Chem. Soc. Japan 35:610-615.
15. NIEDERMAYER, A. 0. 1964. Determination of
307
on July 17, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CHIANG AND BENNETT
phenylacetic acid in penicillin fermentationmedia by means of gas chromatography. Anal.Chem. 193:938-939.
16. ROLiNsON, G. N., F. R. BATCHELOR, D. BUTrER-WORTH, J. CAMERON-WOOD, M. COLE, G. C.EUSTACE, M. V. HART, M. RIcHARDs, AND E.B. CHAIN. 1960. Formation of 6-aminopenicil-lanic acid from penicillin by enzymatic hy-drolysis. Nature 187:236-237.
17. SAKAGUCHI, K., AND S. MURAO. 1950. A pre-liminary report on a new enzyme, "penicillin-amidase." J. Agr. Chem. Soc. Japan 23:411.
18. VAN HOLDE, K. E., AND R. L. BALDWIN. 1958.Rapid attainment of sedimentation equilibrium.J. Phys. Chem. 62:734.
19. WARBURG, 0., AND W. CHRISTIAN. 1941. Iso-lierung und Kristallisation des garungsfermentsenolase. Biochem. Z. 310:384-421.
308 J. BACrtRIOL.
on July 17, 2018 by guesthttp://jb.asm
.org/D
ownloaded from