zhang thesis - ruhr-universit¤t bochum
TRANSCRIPT

1
INTERNATIONAL GRADUATE SCHOOL OF BIOSCIENCES (IGB)
RUHR UNIVERSITÄT BOCHUM
FUNCTIONAL INVESTIGATION OF ODORANT RECEPTOR IN NON -
OLFACTORY TISSUES
FUNKTIONALE UNTERSUCHUNGEN VON RIECHREZEPTOREN IN NICHT-OLFAKTORISCHEN GEWEBEN
Doctoral Dissertation
Weiyi Zhang
Department of Cell Physiology
Thesis advisor: Prof. Dr. Dr. Dr. Hanns Hatt
Bochum, Germany (07.05.07)

2
1. Introduction .......................................................................................................................... 7
1.1 The mammalian olfactory system..................................................................................... 7
1.2 Signal transduction of odorant receptor in the OSN...................................................... 10
1.3 Odorant receptors are not only expressed in olfactory tissues...................................... 11
1.4 Project aims.................................................................................................................... 13
2. Functional investigation of prostate specific G-protein coupled olfactory receptors (PSGR) in prostate cells. ........................................................................................................ 14
2.1 Research background..................................................................................................... 14
2.2 Materials and Methods................................................................................................... 17 2.2.1 Cell culture and transfection. .................................................................................. 17 2.2.2 Antibodies. .............................................................................................................. 18 2.2.3 Western Blotting. .................................................................................................... 18 2.2.4 Single Cell Ca2+ Imaging. ....................................................................................... 19 2.2.5 DNA and siRNA constructs. ................................................................................... 19 2.2.6 Cell Proliferation. .................................................................................................... 20 2.2.7 Apoptosis assay. ...................................................................................................... 20 2.2.8 RT-PCR and primer pairs........................................................................................ 21
2.3 Results. ........................................................................................................................... 21 2.3.1 PSGR responds to β-ionone and to steroid ligands. ................................................ 21 2.3.2 PSGR activation in LNCaP cells............................................................................. 24 2.3.3 PSGR activation elicits phospholipase C mediated cell signaling in LNCaP cells. 28 2.3.4 Primary prostate epithelial cells respond to β-ionone ............................................. 29 2.3.5 Effect of β-ionone on apoptosis and proliferation of LNCaP and primary prostate epithelial cells................................................................................................................... 31
2.4 Discussion ...................................................................................................................... 33
3. Functional investigation of OR51E2 (PSGR) in mammalian spermatozoa.................. 38
3.1 Research background..................................................................................................... 38
3.2 Materials and methods ................................................................................................... 46 3.2.1 Odorants and reagents ............................................................................................. 46 3.2.2 RT-PCR analysis and sequencing ........................................................................... 46 3.2.3 Immunohistochemistry............................................................................................ 46 3.2.4 Sperm preparation and imaging of Ca2+ levels in spermatozoa .............................. 47 3.2.5 Capacitation............................................................................................................. 48 3.2.6 Detection of phosphorylated protein in human spermatozoa.................................. 48
3.3 Results ............................................................................................................................ 49 3.3.1 OR51E2 is functionally expressed in human spermatozoa. .................................... 49 3.3.2 The activation of OR51E2 elicits phospholipase C mediated cellular signaling in human spermatozoa.......................................................................................................... 52 3.3.3 Agonist elicited Ca2+ increase originates from an internal side in sperm cells. ...... 53 3.3.4 Activation of OR51E2 enhances sperm capacitation via PLC activation. .............. 55 3.3.5 Homologs of OR51E2 are present in the testes of various mammalian species. .... 57 3.3.6 OR51E2 and its homologs may have conserved function in spermatozoa from different mammalian species............................................................................................ 59
3.4 Discussion ...................................................................................................................... 61

3
4. Functional investigation of MOR283-2 in mouse spermatogenic cells.......................... 66
4.1 Research background..................................................................................................... 66
4.2 Material and Methods .................................................................................................... 72 4.2.1 Cloning and vector construction. ............................................................................ 72 4.2.2 Cell Culture. ............................................................................................................ 73 4.2.3 RT-PCR................................................................................................................... 74 4.2.4 Real-time quantitative PCR..................................................................................... 75 4.2.5 In-situ hybridization ................................................................................................ 75 4.2.6 Ca2+-imaging. .......................................................................................................... 76 4.2.7 Immunohistochemistry............................................................................................ 77 4.2.8 BrdU proliferation assay ......................................................................................... 77 4.2.9 In situ 3’-end labeling (ISEL) ................................................................................. 77
4.3 Results ............................................................................................................................ 78 4.3.1 Characterization of MOR283-2 expression pattern in testis ................................... 78 4.3.2 Identification of molecular receptive field for MOR283-2..................................... 79 4.3.3 Amylbutyrate induced transient Ca2+ influx in germ cells...................................... 82 4.3.4 Spermatogonia-Sertoli cell coculture. ..................................................................... 83 4.3.5 The effect of amylbutyrate on in vitro spermatogenesis. ........................................ 84 4.3.6 The effect of amylbutyrate on spermatogenic cell proliferation. ............................ 86 4.3.7 Amylbutyrate prevented germ cell apoptosis.......................................................... 87
4.4 Discussion ...................................................................................................................... 88
Summary ................................................................................................................................. 94
Zusammenfassung.................................................................................................................. 95
Reference list........................................................................................................................... 96

4
ACKNOWLEDGEMENTS This thesis work was conducted at the Department of Cell physiology in Ruhr-University Bochum, directed by Prof. Hanns Hatt. During the course of my study, I was a member of International Max-Planck Research School for Chemical Biology (IMPRS-CB). I am grateful to IMPRS-CB for financing me throughout my study, and a special thanks gives to Dr. Jutta Roetter for all the help during the last years. Foremost, I would like to express my gratitude to both of my supervisors Dr. Eva Neuhaus and Prof. Hanns Hatt. During my study, Eva made me progress forward by her impressively theoretical, methodological, and empirical knowledge on numerous aspects of scientific research. I am further grateful for her tremendous support in all the other aspects during my staying in Germany. I would like to thank Prof. Hatt for the mental and instrumental support and encouragement throughout. I would like to further thank Mr. Harry Bartel and Ms. Jasmin Gerkruth for their excellent technical support. I would also like to express my gratitude to all the members in Department of Cell Physiology. I profited a lot from such a friendly network and I would really have nice memory of my time here. Particularly, I would like to thank Jon Barbour, Ying Deng, Ruth Dooley, Julia Doerner, Lian Gelis and Sophie Herling for their scientific help and friendship. Finally, I thank my family for their unconditional support.

5
List of figures Figure 1.1.1 Schematic diagram of olfactory epithelium. Figure 1.2.1 Signal transduction pathway in the OSN. Figure 2.3.1 Heterologous expressed PSGR reponds to β-ionone and special steroid ligands. Figure 2.3.2.1 Endogenous PSGR in LNCaP cells is activated by the same ligands. Figure 2.3.2.2 ADT and β-ionone induced Ca2+ signals in LNCaP cells depend on PSGR expression. Figure 2.3.3. Signal transduction mechanism of endogenous PSGR in LNCaP cells. Figure 2.3.4. Activation of PSGR in primary prostate epithelial cells. Figure 2.3.5. Activation of PSGR in LNCaP and primary prostate epithelial cells reduces cell proliferation involving MAPK signaling. Figure 3.1.1 Schematic representation of a mammalian sperm and the ultrastructure of the flagellum. Figure 3.3.1.1 OR51E2 is functionally expressed in human spermatozoa. Figure 3.3.1.2 Spatiotemporal analysis of Ca2+ signals in single sperm cell. Figure 3.3.2 OR51E2 mediated signaling is via the activation of PLC. Figure 3.3.3 Ca2+ increase induced by OR51E2 is originates from internal sources. Figure 3.3.4 The effect of β-ionone and the correlated signaling components on the sperm capacitation. Figure 3.3.5.1 Degenerate primer design for OR51E2 homogolous sequences. Figure 3.3.5.2 OR51E2 and its homologs are highly conserved and expressed in the testes of different species. Figure 3.3.6 Homologs of OR51E2 show conserved function in sperm cell. Figure 3.4.1 The predicted binding site residues in two-dimensional Snake-plot of human OR51E2. Figure 4.1.1 Spermatogenesis follows an orchestrated differentiation process Figure 4.1.2 Representative differentiation stages and corresponding γH2Ax staining patterns Figure 4.3.1 Expression and localization of MOR283-2 encoding transcripts in adult mouse testis. Figure 4.3.2. Characterization of olfactory receptive field of MOR283-2. Figure 4.3.3. Effect of amylbutyrate on Ca2+ mobilization in spermatogenic cells. Figure 4.3.4. Sertoli-spermatogonia co-cultures. Figure 4.3.5 The effect of amylbutyrate on the in vitro spermatogenesis. Figure 4.3.6 The effect of amylbutyrate on the proliferation of spermatogenic cells. Figure 4.3.7 The effect of amylbutyrate on spermatogenic cell apoptosis.

6
Abbreviations AC Adenylyl cyclase ADT 1,4,6-Androstatriene-3,17-dione AR Androgen receptor CNG channel Cyclic-nucleotide-gated channel DHT 6-Dihydrotestosterone GPCR G-protein coupled receptor OE Olfactory epithelium OR Olfactory receptor OSN Olfactory sensory neuron PLC Phospholipase C PSA Prostate serum antigen PSGR Prostate specific G-protein coupled receptor SERCA Sarco/endoplasmic reticulum Ca2+-ATPase ZP Zona pellucida

7
1. Introduction
1.1 The mammalian olfactory system. The mammalian olfactory system has great ability to detect a tremendous assemblage of
structurally diverse volatile chemical stimuli, namely odorants. The sense of smell (olfaction)
is one of the most ancient senses and allows animals to find food, mates and predators. For
both animals and human beings, olfaction is a very important way to collect environmental
information from the surroundings.
The main olfactory epithelium (MOE) is situated in the roof of the two nasal cavities of the
nose and is specialized to detect fluctuations in the concentration of a large diversity of
airborne molecules and to transduce this information into a stream of neuronal activity which
is conveyed to the brain. The olfactory region of each of the two nasal passages in humans is a
small area of about 2.5 square centimeters containing three principle cell types: sensory
neurons (OSN), sustentacular cells and basal cells (Moulton and Beidler, 1967). Adjacent
sustentacular cells provide trophic, metabolic, and mechanical support for olfactory receptor
neurons. Basal cells are known as progenitor (stem) cells which are capable of processing
mitotic cell division to form olfactory receptor neurons when functionally mature. In fact
OSNs are continually regenerated throughout the lifespan (Beites et al., 2005) and the
olfactory receptor neurons turnover approximately every 40 days (Graziadei and Monti
Graziadei, 1985). The epithelium is kept moist by the secretions of olfactory glands which
also include odorant binding proteins. These bind to hydrophobic odorants and ‘present’ them
to the olfactory receptors.
The olfactory sensory neuron is bipolar. A dendritic process extends to the mucosal surface
where it gives rise to a number of specialized cilia that provide an extensive, receptive surface
for the interaction of odors with the cell. The OSN also gives rise to an axon which projects to
the olfactory bulb of the brain, the first relay in the olfactory system. The axons of the

8
olfactory bulb neurons, in turn, project to subcortical and cortical regions where higher-level
processing of olfactory information allows the discrimination of odorants by the brain (Buck,
1992)
Figure 1.1 Schematic diagram of olfactory epithelium (Mombaerts, 2004). Olfactory epithelium consists of 3
different cell types: olfactory sensory neurons, supporting sustentacular cells, and basal stem cells. The OSN is a
bipolar cell which give rise to specialized cilia, where olfactory receptors are expressed. It also gives rise to an
axon which conveys the information to the brain.
The discrimination between thousands of volatile chemical stimuli is based on a large family
of G-protein coupled odorant receptors (Buck, 1992). It was estimated that there are more
than 1000 odorant receptor genes in rodents and human by whole genome sequence analysis
(Niimura and Nei, 2003; Zhang and Firestein, 2002). In human, in total 906 OR genes and
pseudogenes were identified, among which ~350 OR genes are functional representing 1% of
human genome (Malnic et al., 2004; Niimura and Nei, 2005; Mombaerts, 1999). In mouse,
OR gene family is composed of 1209 genes, 913 of which are likely to encode functional
receptors (Niimura and Nei, 2005). The intact OR genes can be grouped into families. Class I
genes resemble the OR genes of fish but class II genes are unique to terrestrial vertebrates.
The amino acid similarity is 37% on average across the OR repertoire, but it can be as low as
25% between two ORs (Mombaerts, 2004).

9
Each OSN expresses only one OR type (Chess et al., 1994; Malnic et al., 1999) that is derived
from only one of the two alleles present in the genome (Chess et al., 1994; Lomvardas et al.,
2006). However, a particular OR can respond to a spectrum of multiple odorants, which has
been shown in recordings from the OSNs and heterologous expression system (Araneda et al.,
2000; Hatt et al., 1999; Kajiya et al., 2001; Spehr et al., 2003; Touhara et al., 1999; Wetzel et
al., 1999), while a particular ligand can elicit responses from multiple ORs, leading to unique
combinations of ORs for each odorant (Krautwurst et al., 1998; Malnic et al., 1999). ORs are
members of the rhodopsin-like class of G protein coupled receptors (GPCR), which also
includes catecholamine receptors, and their three-dimensional structure contains the seven
helical transmembrane (TM) motifs characteristic of GPCRs (Mombaerts et al., 1996).
Rhodopsin-like GPCRs exist in one of the two conformations: an inactive conformation and
an active conformation that interacts with an intracellular heterotrimeric G protein (Bargmann,
2006). The transition between the two conformational changes occurs through the movement
of various membrane spanning domains around the so-called agonists or antagonists (Gether
and Kobilka, 1998; Petronczki et al., 2003). Agonists stabilize the active form of the receptor,
whereas antagonists can block agonist binding and stabilize the inactive form (Bargmann,
2006). The properties of odorant receptors are likely to follow these rules. The first hints of
putative odorant binding sites were deduced from the sequencing of first OR proteins, which
revealed that transmembrane domains 3 to 6 are more variable between paralogs (Buck, 1992).
Since then, more studies attempted to reveal ligand binding sites of odorant receptor based on
the sequence analysis and point mutations (Hatt, 2004; Singer et al., 1995; Singer et al., 1996).
From these studies, an interesting finding was based on the two human odorant receptors,
OR17-40 and OR17-44, whose sequences are mostly identical but only differ in 12
transmembrane residues localized from TM3 to TM6. Consequently, these residues lead to the
different ligand specificity of the receptors (Hatt, 2004). Additional studies predicted ligand
binding sites by computer based docking of odorants to structural models of the receptors

10
and/or site-directed mutagenesis (Afshar et al., 1998; Doszczak et al., 2007; Floriano et al.,
2000; Floriano et al., 2004; Singer et al., 1996). These studies together with a report using
sequence pair predicted that ligand binding positions of ORs localized in TM3 to TM7 with
contributions from extracellular loops 2 and 3 (Man et al., 2004). In particular, a few ORs
were particularly modeled and the critical amino acids guarding the ligand specificity were
predicted: for the I7 receptor, 6 amino acids of TM4 were predicted to be involved in
recognition of octanal and other aldehydes, and moreover, mutation of Lys164 has the great
affinity to octanal by forming an electrostatic interaction with octanal (Hall et al., 2004); the
odorant binding sites of mOR-EG are situated in a hydrophobic pocket formed by 9 amino
acids on TM3, TM5 and TM6 (Katada et al., 2004); a network of 8 residues in TM3, TM5 and
TM6 was identified as the critical part of the ligand binding pocket of MOR42-3 (Abaffy et
al., 2007); the 23 amino acids of TM2 to TM7 and extracellular loop 2 form the ligand-
binding pocket of the hOR17-4 receptor (Doszczak et al., 2007).
1.2 Signal transduction of odorant receptor in the OSN.
The initial step in activating OSNs is the binding of the odorant to a corresponding receptor in
the membrane of cilia. Odorant receptors are probably subjected to conformational
rearrangements upon ligand binding, similar to the light-induced structural changes of
rhodopsin (Grobner et al., 2000), thereby transmitting the signal from the extracellular to the
intracellular side of the membrane (Paysan and Breer, 2001). Most odorant activated ORs are
linked to the stimulation of adenylyl cyclase III (AC3) (Bakalyar and Reed, 1990) via a Gαs
homologous alpha subunit of trimeric G-protein, namely Golf (Jones and Reed, 1989).
Activation of AC3 elicits the increase of cyclic adenosine-3’, 5’-monophosphate (cAMP)
level in the cilia, which then leads to the opening of cyclic-nucleotide gated (CNG) cation
channels (Zufall et al., 1994). The CNG channels are very permeable to Ca2+ (Dzeja et al.,
1999). The influx of cations through CNG channels depolarizes the cilia membrane, elevates

11
the intracellular Ca2+ concentration, and then triggers a Ca2+-activated chloride conductance
that significantly amplifies the electrical signal.
Figure 1.2 Signal transduction pathway in the OSN. However, in addition to the above general consensus, it was found that some potent odorants
elicit a substantial increase of inositol 1, 4, 5-trisphosphate (IP3) concentration with a rapid
and transient kinetics (Bakalyar and Reed, 1990; Breer et al., 1990; Huque and Bruch, 1986;
Ko and Park, 2006; Ronnett et al., 1993). This process is mediated by isoforms of
phospholipase C (PLC), which was elucidated by the application of PLC inhibitor U73122
(Noe and Breer, 1998). Evidences also indicated that receptor activation linked to PLC is
mediated by G proteins other than Golf (Schandar et al., 1998) and IP3 gated ion channels have
been considered as possible downstream component of this cascade (Lischka et al., 1999).
1.3 Odorant receptors are not only expressed in olfactory tissues.
In recent years, a number of studies showed that odorant receptors are found not only
restricted to be expressed in olfactory epithelium but also in a large variety of non-olfactory
tissues: RT-PCR and in situ hybridization analyses showed that a rat odorant receptor OL1 is
expressed in developing heart (Drutel et al., 1995); a human odorant receptor is expressed at

12
RNA level in erythroid cells (Feingold et al., 1999); in a transgenic mouse line, by staining
the coexpressed marker, an olfactory receptor subtype is expressed in autonomic nervous
system (Weber et al., 2002); degenerate PCR suggested that a few mouse odorant receptors
are expressed in pyramidal neurons in the cerebral cortex (Otaki et al., 2004); RT-PCR
analysis revealed that the rat odorant receptor OL2 is expressed in spleen and insulin-
secretion cell line (Blache et al., 1998); with a transgenic approach it was found that a rat
odorant receptor is expressed in brainstem (Conzelmann et al., 2000; Raming et al., 1998);
during development, some olfactory receptor subtypes have also been found in the cribriform
mesenchyme between the prospective olfactory epithelium and the developing telencephalon
(Schwarzenbacher et al., 2004); Northern blots and RT-PCR analyses revealed that PSGR, a
G protein coupled odorant receptor was found to be expressed in prostate epithelial cells and
overexpressed in prostate cancer epithelial cells (Xu et al., 2000). Additionally, microarray
analysis of mouse odorant receptor gene expression elucidated that some receptors displayed
expression in testis, liver, heart, cerebellum and muscle (Zhang et al., 2004a). Nevertheless,
for none of these receptors a related physiological function has been shown in these non-
olfactory tissues.
However, a recent study put forward an opinion that the ectopically expressed ORs might be
not functional in the corresponding tissues, as the OR superfamily shows widespread, locus-
dependent and heterogeneous expression, in agreement with a neutral or near neutral
evolutionary model for transcription control, whereby functionality is rendered less likely
(Feldmesser et al., 2006). Nevertheless, accumulating functional studies denied the above
opinion, and in particular, the approach of proteomics analysis of human sperm provided a
direct proof that ORs are present at protein level at least in mature spermatozoa (Barbour,
2006; Spehr et al., 2004b), which is confirmed by the immunostaining and western blot
detection of a human odorant receptor in mature spermatozoa (Neuhaus et al., 2006a).

13
A few ORs have been detected in male germ cells of mammals, including human, dog, rat and
mouse (Asai et al., 1996; Parmentier et al., 1992; Vanderhaeghen et al., 1993; Vanderhaeghen
et al., 1997b) which may imply an important role of ORs in chemoreception during sperm-egg
communication. This inference was strengthened by the study of functional characterization
of hOR17-4 on human spermatozoa (Spehr et al., 2003). This study showed that activation of
hOR17-4 by the specific ligand bourgeonal induces a transient increase of intracellular Ca2+
concentration and also induces strong chemotaxis in sperm behaviour studies. A further study
also showed that the membrane bounded AC3 is involved in the signal transduction of
hOR17-4 (Spehr et al., 2004b). Another study of murine testicular olfactory receptor MOR23
further confirmed that activation of MOR23 elicits Ca2+ influx in spermatozoa and may play a
role in chemoreception during sperm-egg communication and thereby regulate fertilization
(Fukuda et al., 2004).
In addition to the studies of testicular ORs in mature spermatozoa, a recent study using in situ
hybridization and reverse transcriptase polymerase chain reaction (RT-PCR) showed that
several OR transcripts in testis are expressed in three developmental stages: late pachytene
spermatocytes, early round spermatids and late round spermatids (Fukuda and Touhara, 2006).
These findings elucidated the developmental expression profile of testicular OR subset during
spermatogenesis. However, the physiological function of these ORs in spermatogenesis
remains to be clarified.
1.4 Project aims
The project aim was a functional characterization of novel odorant receptors and analysis of
the potential physiological role in non-olfactory tissues. This goal was addressed in three
different aspects:
(1) Functional investigation of prostate specific G-protein coupled olfactory receptor
(PSGR) in prostate cells. Refer to chapter 2.

14
(2) Functional investigation of OR51E2 in mammalian spermatozoa. Refer to chapter 3.
(3) Functional investigation of MOR283-2 in mouse spermatogenic cells. Refer to chapter 4.
2. Functional investigation of prostate specific G-protein coupled olfactory receptors
(PSGR) in prostate cells.
2.1 Research background
Prostate cancer is the most-diagnosed malignant growth in men and is the second-leading
cause of male cancer deaths in the majority of western country. Patients with prostate cancer
have several treatment options, including watchful waiting, surgery and radiation. However,
because the growth of prostate cancer is controlled by steroid androgens, mainly
dihydrotestosterone (DHT), the treatment of locally advanced or metastatic tumor reckons on
hormonal therapies targeting androgen receptor (AR). These therapies include: androgen
ablation by physical or chemical castration of the patient to reduce levels of circulating
androgens; treatment with AR antagonist to disrupt receptor activation; or a combination of
both (Chodak et al., 2002; So et al., 2003; Sternberg et al., 2003). A major limitation of
hormonal therapy is that it only offers temporary relief. Biologically prostate cancer
progresses from an androgen-dependent to an androgen-independent state, characterized by
aggressive growth and invasion of distal organs, predominantly the bone (Feldman and
Feldman, 2001).
Several receptors of growth factors, including epidermal growth factor, insulin-growth factor,
fibroblast growth factor, platelet derived growth factor and transforming growth factor-α,
have been implicated in the development and progression of prostate cancer to androgen
independence (Raj et al., 2002). Recently evidences are accumulating to demonstrate the
involvement of GPCRs in neoplastic transformation of prostate(Daaka, 2004). Firstly, the
cancerous prostate contains elevated levels of enzymes that control the expression of GPCR

15
ligands. Secondly, prostate cancer cells produce increased amounts of GPCR ligands,
including follicle-stimulating hormone (Porter et al., 2001), endothelin-1 (Nelson et al., 2003;
Nelson et al., 1996), and lysophosphatidic acid (Xie et al., 2002; Mills and Moolenaar, 2003).
Thirdly, malignant prostate specimens express higher level of GPCRs, including prostate
specific G-protein coupled olfactory receptor (Xu et al., 2000), bradykinin 1 receptor (Taub et
al., 2003), and endothelin 1A receptor (Gohji et al., 2001; Nelson et al., 1996), compared to
benign prostate tissue. The GPCRs transduce their signals primarily via activation of
heterotrimeric G proteins to produce Gα-GTP and Gβγ subunits. The G proteins are divided
into four groups; Gs, Gi, Gq and G12. Gs and Gi regulate mainly adenylyl cyclases that produce
cAMP, Gq regulates phospholipases that control intracellular Ca2+ levels, and G12 regulates
low molecular weight GTPase Rho and other effectors.
The outward symptoms of prostate cancer are not always apparent; therefore, if left untreated,
prostate cancer may metastasise to other part of body or vital organs. When this happens, the
patients have fewer treatment options than they would if the disease had been discovered
earlier. Although PSA test has been found to be very successful in the early detection of
prostate cancer, it is reported that PSA is elevated not only in men with prostate tumors, but
also in men with benign prostate hyperplasia, prostatitis and other non-malignant disorders
(Pannek and Partin, 1997). It has resulted in enthusiasm to discover new prostate-specific
genes that are not only prostate specific but also overexpressed in prostate cancer.
In several attempts to identify novel prostate-specific tumor biomarkers, a prostate-specific G
protein coupled receptor (PSGR) was identified. The PSGR gene encodes a seven
transmembrane-spanning GPCR which belongs to the superfamily of odorant receptors (ORs)
and maps to chromosome 11p15 (Pannek and Partin, 1997; Vanti et al., 2003; Weng et al.,
2005a; Xu et al., 2000; Xu et al., 2006). Northern blot analysis indicates that the expression of
PSGR is almost exclusive to human prostate gland. PCR and Matched Normal/Tumor Tissue
Array study show significant overexpression of PSGR mRNA in prostate tumor tissues (Xia

16
et al., 2001; Xu et al., 2000). In situ RNA hybridization and quantitative real-time PCR
analysis of more than 140 human prostate tissues (normal and tumors) demonstrate that the
mean increase PSGR mRNA expression in prostate intraepithelial neoplasia (PIN) and
malignant prostate cancers was 25 fold higher over normal human prostate tissues and benign
prostatic hyperplasia (BPH) tissues, suggesting PSGR could be a very sensitive and specific
biomarker to distinguish BPH from prostate carcinoma (Pannek and Partin, 1997; Weng et al.,
2005b). Furthermore, it was found out that the PSGR overexpression associated with higher
percentage of pathologic stage, pT3 (non-organ confined), and a higher level of preoperative
serum PSA (Pannek and Partin, 1997; Xu et al., 2006).
The classical model of steroid action involves binding to specific intracellular steroid
receptors, translocation to the nucleus, DNA binding, and activation of specific genes (Beato,
1989). However, in recent years a number of reports indicated additional steroid actions,
including the rapid activation of kinase signaling cascades, modifications of the cytoskeleton,
and modulation of cyclic nucleotide and intracellular calcium levels (Cato et al., 2002; Erkkila
et al., 2002; Heinlein and Chang, 2002; Herve, 2002; Levin, 2001; Losel et al., 2002). A
number of studies introduced the concept of nongenomic steroid hormone actions to explain
observations related to rapid steroid effects. Nongenomic effects were proposed as membrane-
initiated steroid signaling, which is insensitive to inhibitors of transcription and translation
and is, in most cases, insensitive to steroid antagonists (Falkenstein et al., 2000). Nongenomic
steroid actions have been reported for most prominent steroids (Heinlein and Chang, 2002;
Losel et al., 2002), and binding of androgens via androgen-specific membrane receptors have
been described in human prostate tumors and in the LNCaP human prostate cancer cell line
(Kampa et al., 2002; Papakonstanti et al., 2003; Stathopoulos et al., 2003). The activation of
testosterone membrane receptors results in a strong and persistent regression of prostate
cancer cells (Papakonstanti et al., 2003). Although the nature of these membrane steroid sites
was elusive until recently, the identification of a membrane progesterone (Falkenstein et al.,

17
2000) and estrogen receptor (Revankar et al., 2005), and the isolation of a membrane
glucocorticoid-binding protein with homologies with opioid receptors (Evans et al., 2000)
showed that at least some of these proteins belong to the seven-transmembrane G protein
coupled receptors (GPCRs).
In this work we aimed to explore the functional relevance of the expression of an OR in
prostate tissue. We identified the ligands of heterologously expressed PSGR and showed that
in addition to the classical fragrance β-ionone, and the receptor is activated by steroid
hormones. The identified odorant as well as the steroid hormones elicited rapid Ca2+
responses in the LNCaP prostate cancer cell line and in primary prostate epithelial cells.
Activated PSGR moreover causes phosphorylation of p38 and SAPK/JNK MAPKs, resulting
in reduced proliferation rates and an induction of apoptosis.
2.2 Materials and Methods
2.2.1 Cell culture and transfection.
Reagents for cell culture use were purchased from Invitrogen, unless stated otherwise.
HEK293 cells were maintained under standard conditions in MEM supplemented with 10%
FBS, 100 units/ml penicillin and streptomycin, and 2 mM L-glutamine. LNCaP cells were
maintained in RPMI 1640 medium supplemented with 10% FBS and 100 units/ml penicillin
and streptomycin. HEK293 cell transfections with the PSGR containing plasmid were
performed using a standard calcium phosphate precipitation technique; for siRNA
experiments LNCaP cells were transiently transfected with either targeted or scrambled
siRNAs using Exgene500 (Fermentas). Two days after transfection the growth medium was
removed and replaced with standard Ringer solution (140 mM NaCl, 5 mM KCl, 2 mM CaCl2,
2 mM MgCl2, 10 mM Hepes, 10 mM glucose).
Prostate epithelial cells (PECs) were isolated from freshly collected prostate tissue, which was
minced to 1 mm3 pieces and digested for 30 min at 37°C in Ringer solution containing 0.1%

18
trypsin-EDTA. The tissue was dissociated by trituration, washed and a single cell suspension
of PECs was prepared by centrifugation of the remaining tissue pieces. PECs were seeded in
the serum-free Keratinocyte-SFM medium (supplemented with 50 ng/ml human recombinant
epidermal growth factor and 50 µg/ml bovine pituitary extract) in 50-ml flasks and kept at
37°C in a humidified incubator with 5% CO2. When they reached 70–80% confluence, the
cells were trypsinized and subcultured either in Petri dishes for Ca2+-imaging experiments or
in 96 well plates for cell proliferation and apoptosis assays. Cell morphology was checked
with Zeiss Axioskop2 microscope and viewed with 20 X magnification.
2.2.2 Antibodies.
The following primary antibodies were used: (a) rabbit polyclonal antibodies against p44/42
MAPK and against phosphorylated p44/42 MAPK (New England Biolabs); (b) rabbit
polyclonal antibodies against p38 MAPK and phosphorylated p38 MAPK (New England
Biolabs); (c) rabbit polyclonal antibodies against SAPK/JNK and phosphorylated SAPK/JNK
(New England Biolabs). Secondary goat anti-rabbit antibodies conjugated to HRP (Biorad)
were used.
2.2.3 Western Blotting.
LNCaP cells were treated with 500 µM β-ionone for the indicated times, harvested, pelleted
and homogenized in lysis buffer (50 mM Tris-Cl, pH7.4, 150 mM NaCl, 1 mM EDTA, 1%
Triton X-100) with protease inhibitors (Roche Complete® protease inhibitor mixture).
Sample aliquots of the cells were mixed with Laemmli buffer (30% glycerol, 3% SDS, 125
mM Tris/Cl, pH 6.8), resolved by 10% SDS-PAGE and transferred to nitrocellulose
membrane (Protran; Schleicher & Schuell). The nitrocellulose membranes were stained with
Ponceau S (Sigma), blocked with TBST (150 mM NaCl, 50 mM Tris-Cl, 0.1% Triton, pH
7.4), containing 5% nonfat dried milk (Biorad) and incubated with primary antibodies diluted
in 3% dry milk in TBST. After washing and incubation with HRP coupled secondary

19
antibodies, detection was performed with ECL plus (Amersham) on Hyperfilm ECL
(Amersham).
2.2.4 Single Cell Ca2+ Imaging.
For measuring the cytosolic Ca2+ concentration, cells were incubated (30 min/37°C) in Ringer
solution containing 3 µM Fura-2-AM (Molecular Probes). After removal of extracellular
Fura-2, cells were treated with water-soluble adenylate cyclase inhibitor MDL (Calbiochem)
50 µM or phospholipaseC inhibitor U73122 (Calbiochem) 10 µM for 30 min. The
intracellular calcium store depleting reagent thapsigargin (Sigma) was used at 1 µM.
Ratiofluometric Ca2+-imaging was performed as described (Spehr et al., 2003) using a Zeiss
inverted microscope equipped for ratiometric imaging. Images were acquired from randomly
selected fields of view, and integrated fluorescence ratios (f340/f380 ratio) were measured.
Exposure to odorants was accomplished using a specialized microcapillary application system.
Odorants used were a gift of Dr. T. Gerke, Henkel KGaA, Düsseldorf, Germany. Steroid
hormones were purchased from Steraloids (Newport, U.S.A.). Odorants and steroids assayed
for potential activation of PSGR were tested in at least three transfection experiments in
HEK293 cells and tested for activation of LNCaP cells afterwards. All compounds regarded
as ligands led to clear Ca2+ responses in several different experiments (n > 7), whereas they
did not elicit any Ca2+ signals in untransfected dishes. 200 µM ATP was applied as the
positive control at the end of each experiment in HEK293 cells.
2.2.5 DNA and siRNA constructs.
Human PSGR (NM-030774) was amplified from human genomic DNA by PCR using
specific primers which amplify the complete open reading frame and contain EcoRI
restriction sites for further subcloning into pcDNA3 (Invitrogen); the generated plasmid was
verified by sequencing.

20
PSGR targeted and scrambled hairpin siRNA designs were carried out with siRNA Target
Designer-Version 1.51 (Promega); oligos were synthesized by Invitrogen and ligated into the
pGeneClipTM hMGFP vector (Promega) according to the manufacturers instruction. The best
working siRNA sequence of PSGR was GCTGCCTCCTGTCATCAAT; the oligonucleotide
sequences to generate 5'-target-loop-reverse-complement-3' hairpins were 5’-
TCTCGCTGCCTCTGTCATCAATAAGTTCTCTATTGATGACAGGAGGCAGCCT-3’,
5’-CTGCAGGCTGCCTCCTGTCATCAATAGAGAACTTATTGATGACAGGAGGCAGC-
3’. The following scrambled versions of the siRNA sequence was used as control 5’-
TCTCGTACACTGACCCCCTTTGTAAGTTCTCTACAAAGGGGGTCAGTGTACCT-3’,
5’-CTGCAGGTACACTGACCCCCTTTGTAGAGAACTTACAAAGGGGGTCAGTGTAC-
3’.
2.2.6 Cell Proliferation.
Growing LNCaP cells and primary human prostate cancer epithelial cells were plated in 96
well plates at a density of 5×103 cells/well. After 24 hours at 37°C with 5% CO2, cells were
treated with different concentrations of β-ionone (50 nM to 250 µM), Dihydrotestosterone
(DHT, 10 nM), or with a mixture of β-ionone (250 µM and 100 µM) and DHT (10 nM).
Alternatively, cells were simultaneously stimulated with β-ionone (250 µM) and varying
concentrations of inhibitors for p38 ((RS)-{4-[5-(4-Fluorophenyl)-2-methylsulfanyl-3H-
imidazol-4-yl] pyridin-2-yl}-(1-phenylethyl) amine]), Calbiochem) and JNK (Anthra[1,9-
cd]pyrazol-6(2H)-one1,9-pyrazoloanthrone, Calbiochem). Cell proliferation was investigated
after 3 and 6 days using CyQUANT cell proliferation assay kit (Invitrogen).
2.2.7 Apoptosis assay.
LNCaP cells and primary prostate cancer epithelium cells were treated with 5 µM and 250
µM β-ionone for 3 days observed by phase contrast microscopy. Induction of apoptosis was
monitored using apoptotic DNA ladder kit (Roche).

21
2.2.8 RT-PCR and primer pairs.
RNA of LNCaP and human primary prostate cells was isolated with Trizol reagent
(Invitrogen), digested with DNAseI (Fermentas) and purified again with Trizol before
isolation of polyA+ mRNA with oligo-dT-coated paramagnetic particles (Dynal). cDNA was
synthesized by using MMLV reverse transcriptase (Invitrogen) and oligo(dT18) primer. PCR
was performed with 100 ng template cDNA and specific primer pairs for PSGR, PSA,
androgen receptor, cytokeratin 8, and cytokeratin 18, respectively. The amplifications were
done for 35 cycles (1 min 94°C, 1 min 58°C, 45sec 72°C).
The following primer pairs were used in RT-PCR analysis:
PSGR across intron: forward 5’-CCTCAGCCTTCTGAGTCAGC-3’
reverse 5’-GAGACTGTGACAAGCCCTGG-3’
Androgen receptor(AR): forward 5’-GCCTGTTGAACTCTTCTGAGC-3’
reverse 5’-GCTGTGAAGGTTGCTGTTCCTC-3’
Prostate serum antigen (PSA) forward 5’-TACCCACTGCATCAGGAACA-3’
reverse 5’-CCTTGAAGCACACCATTACA-3’
Cytokeratin 18 (CK18) forward 5’-TGAGACGACGCTCACAGAGCTGA-3’
reverse 5’-TATCCGGCGGGTGGTGGTCTTTT-3’
Cytokeratin 8 (CK8) forward 5’-CTGGAGGCCGCCATTGCAGAT-3’
reverse 5’-CAGACACCAGCTTCCCATCACG-3’
2.3 Results.
2.3.1 PSGR responds to β-ionone and to steroid ligands.
The recently identified human prostate-specific G-protein coupled receptor (PSGR) is
specifically expressed in human prostate tissues and its expression increases significantly in
human prostate intraepithelial neoplasia (PIN) and prostate tumors (approximately 10-fold),
suggesting that PSGR may play an important role in early prostate cancer development and
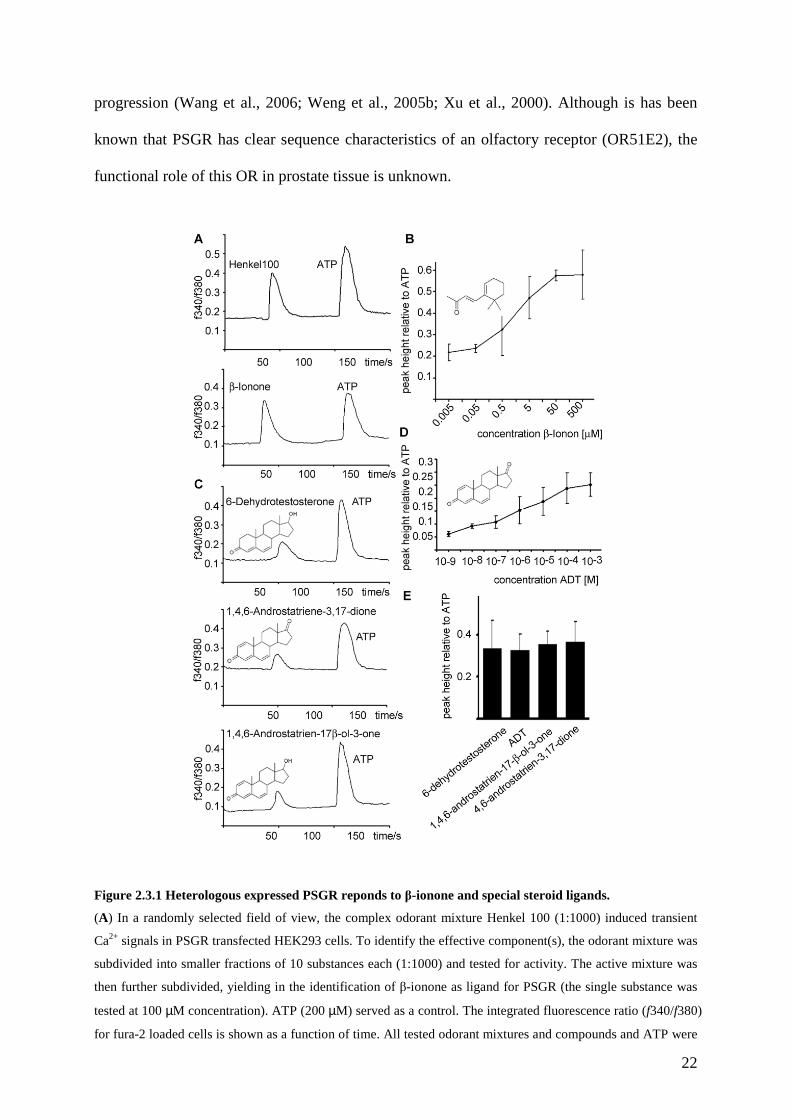
22
progression (Wang et al., 2006; Weng et al., 2005b; Xu et al., 2000). Although is has been
known that PSGR has clear sequence characteristics of an olfactory receptor (OR51E2), the
functional role of this OR in prostate tissue is unknown.
Figure 2.3.1 Heterologous expressed PSGR reponds to β-ionone and special steroid ligands.
(A) In a randomly selected field of view, the complex odorant mixture Henkel 100 (1:1000) induced transient
Ca2+ signals in PSGR transfected HEK293 cells. To identify the effective component(s), the odorant mixture was
subdivided into smaller fractions of 10 substances each (1:1000) and tested for activity. The active mixture was
then further subdivided, yielding in the identification of β-ionone as ligand for PSGR (the single substance was
tested at 100 µM concentration). ATP (200 µM) served as a control. The integrated fluorescence ratio (f340/f380)
for fura-2 loaded cells is shown as a function of time. All tested odorant mixtures and compounds and ATP were

23
applied for 5 s. (B) β-ionone was tested at different concentrations for the activation of recombinant PSGR, the
peak height of the Ca2+ signal relative to the ATP induced Ca2+ signal is displayed as function of the applied
concentration. (C) Testing a steroid library composed of 100 structurally diverse steroids for the activation of
recombinant PSGR resulted in the identification of the active substances 6-dehydrotestosterone, ADT and 1,4,6-
androstatrien-17β-ol-3-one, fluorescence ratios (f340/f380) are shown for fura-2 loaded cells as a function of
time. Steroids (first peak) and ATP (second peak) were applied for 5 s each. (D) ADT was tested at different
concentrations for the activation of recombinant PSGR, the peak height of the Ca2+ signal relative to the ATP
induced Ca2+ signal is displayed as function of the applied concentration. (E) Different steroids containing
double bonds at position 6 and 4 and a keton-group at position 3 were tested for activation of recombinant PSGR,
the peak heights relative to ATP are given (concentrations are 10 µM each).
We cloned PSGR and functionally expressed it in HEK293 heterologous expression system.
We then transiently expressed PSGR in HEK293 cells and determined its ligand specificity by
measuring the cell responses to a mixture of chemical stimuli using ratiofluorometric Ca2+
imaging (Neuhaus et al., 2006b; Wetzel et al., 1999). Our complex odorant mixture (Henkel
100) for the initial ligand screening includes 100 compounds, mainly aromatic and short-chain
aliphatic hydrocarbons, which was used previously to find stimuli for “orphan” ORs (Spehr et
al., 2003; Wetzel et al., 1999). Henkel 100, diluted 1:1000 in Ringer solution, induced
transient Ca2+ responses in 1 to 2% of all cells tested, which is typical for transient OR
transfections (Spehr et al., 2003; Wetzel et al., 1999). By subdividing the mixture into 2
mixtures composed of 50 substances, followed by 5 mixtures composed of 10 substances each,
we identified one submixture and subsequently β-ionone as the only active ligand in the
Henkel 100 mixture (Figure2.3.1A). β-ionone elicited receptor induced Ca2+-responses
already at concentrations of approximately 100 nM (Figure 2.3.1B). Other components of the
same Henkel sub-mixtures were tested as single substances (Cumarine, Traseolide, Fixolide,
Terpineol, Heliotropin) did not result in Ca2+ signals in PSGR expressing HEK293 cells. In
untransfected HEK293 cells, β-ionone induced Ca2+-signals were not observed.
It is a common feature of ORs that they are activated by multiple ligands with different
receptor affinities (Malnic et al., 1999), therefore we continued to test the prostate specific OR
with other substances. As it is discussed in several recent publications that steroid hormones

24
can mediate rapid nongenomic signaling events involving GPCRs, we tested a steroid library
composed of 100 structurally diverse steroids for their ability to activate heterologously
expressed PSGR. By subdividing this mixture, we found that steroids which share some
structural homologies with β-ionone induced transient Ca2+ responses in PSGR expressing
HEK293 cells (Figure 2.3.1C). The presence of an aldehyde group at position 3, together with
at least two double bonds at positions 4 and 6 which are present in 6-dehydrotestosterone,
1,4,6-androstatrien-3,17-dione, and 1,4,6-androstatrien-17β-ol-3-one were key determinants
for effective PSGR ligands (Figure 2.3.1C). 1,4,6-Androstatriene-3,17-dione (ADT) was then
tested at different concentrations for activation of PSGR, and found to function as a ligand
already in the lower nanomolar range (Figure 2.3.1D). We next specifically tested other
steroids with the same structural motif, and found that these substances could also activate
PSGR to a similar extent (Figure 2.3.1E). Some modifications of the group at position 17
were tolerated, whereas the absence of the double bonds at position 6 (e.g. in testosterone) or
4 (e.g. in dihydrotestosterone) abolishes the ability of a ligand to activate the receptor.
Additional single substances, which were tested and found to be inactive were
Androstanedione, 4,16-Androstadiene-3-one, 5-Androstan-3,16-diol, Etiocholanolone, 4-
Androstene-3,6,17-trione, Fernhotz-Acid, and 4-Estren-3-17-diol (data not shown). All
steroids were tested at the same concentrations for unspecific responses in untransfected
HEK293 cells.
2.3.2 PSGR activation in LNCaP cells
We next investigated whether stimulation of PSGR with the identified ligands in the LNCaP
prostate cancer cell line also affected intracellular Ca2+ homeostasis (Figure 2.3.2.1).
Applying the PSGR agonist β-ionone to LNCaP cells elicited a slow, gradual increase in the
intracellular Ca2+ concentration within 5-8 min (Figure 2.3.2.1A). Also the identified steroid
ligands induced a slow rise in the Ca2+ concentration, while androgens without double bonds
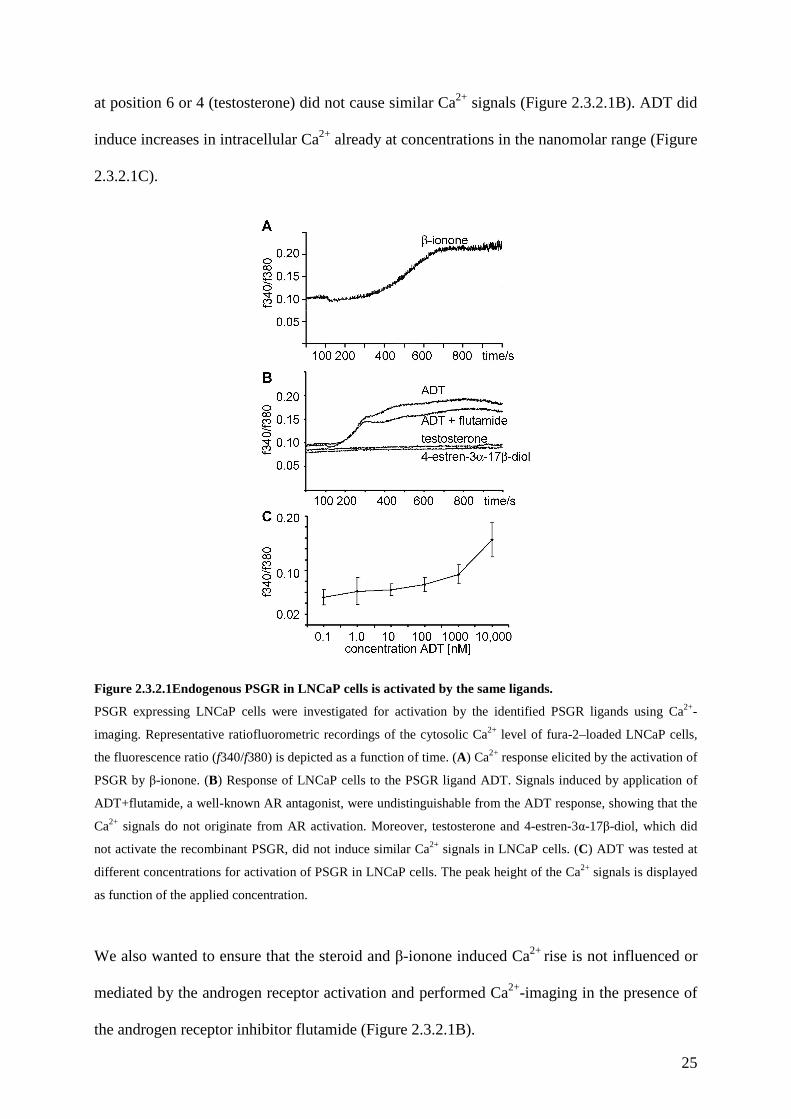
25
at position 6 or 4 (testosterone) did not cause similar Ca2+ signals (Figure 2.3.2.1B). ADT did
induce increases in intracellular Ca2+ already at concentrations in the nanomolar range (Figure
2.3.2.1C).
Figure 2.3.2.1Endogenous PSGR in LNCaP cells is activated by the same ligands.
PSGR expressing LNCaP cells were investigated for activation by the identified PSGR ligands using Ca2+-
imaging. Representative ratiofluorometric recordings of the cytosolic Ca2+ level of fura-2–loaded LNCaP cells,
the fluorescence ratio (f340/f380) is depicted as a function of time. (A) Ca2+ response elicited by the activation of
PSGR by β-ionone. (B) Response of LNCaP cells to the PSGR ligand ADT. Signals induced by application of
ADT+flutamide, a well-known AR antagonist, were undistinguishable from the ADT response, showing that the
Ca2+ signals do not originate from AR activation. Moreover, testosterone and 4-estren-3α-17β-diol, which did
not activate the recombinant PSGR, did not induce similar Ca2+ signals in LNCaP cells. (C) ADT was tested at
different concentrations for activation of PSGR in LNCaP cells. Τhe peak height of the Ca2+ signals is displayed
as function of the applied concentration.
We also wanted to ensure that the steroid and β-ionone induced Ca2+ rise is not influenced or
mediated by the androgen receptor activation and performed Ca2+-imaging in the presence of
the androgen receptor inhibitor flutamide (Figure 2.3.2.1B).

26
Flutamide treatment did not change the amplitude or the kinetics of the ADT induced signal
(Figure 2.3.2.1B). Moreover, steroids which are well known to effectively activate androgen
receptor as e.g. 4-estren-3α,17β-diol and DHT, but do not activate the heterologously
expressed PSGR, elicited no significant Ca2+ increases in LNCaP cells (Figure 2.3.2.1B). The
results demonstrated an identical molecular receptive field of native PSGR to the recombinant
receptor.
To prove that the β-ionone-induced Ca2+ increase in LNCaP cells was due to PSGR activation,
we performed RNAi experiments to reduce the PSGR expression levels. We cloned a PSGR
targeting sequence into a plasmid designed for in vivo expression of short interfering RNAs
(siRNAs). This vector contains GFP as internal fluorescent marker, which enables the
determination of the transfection efficiency and facilitates recording of the Ca2+ responses
specifically in the siRNA expressing cells (Figure 2.3.2.2A). β-ionone induced Ca2+ signals in
siRNA expressing LNCaP cells (orange and turquoise curves in Figure 2.3.2.2B) were then
compared to the Ca2+-increase in neighbouring cells not expressing GFP (blue and pink
curves in Figure 2.3.2.2B). Quantification of the Ca2+ signals revealed that siRNA expression
strongly (~80%) reduced the β-ionone mediated Ca2+-increase (Figure 2.3.2.2B), indicating
that this increase is mediated by activation of PSGR. Expression of GFP alone did not alter
the� β-ionone induced Ca2+-increase (data not shown).
In addition, we investigated whether the identified steroid ligands also induce PSGR
dependent Ca2+-increase in LNCaP cells (Figure 2.3.2.2C, D). Ratiofluorimetric single cell
Ca2+ measurements showed, that ADT induced increases in the Ca2+ concentration were
strongly reduced in siRNA expressing cells, which were identified by GFP expression (Figure
2.3.2.2C, D). All steroids that have been identified as PSGR ligands in HEK293 cells were
able to induce an increase in the intracellular Ca2+-concentration of LNCaP cells.
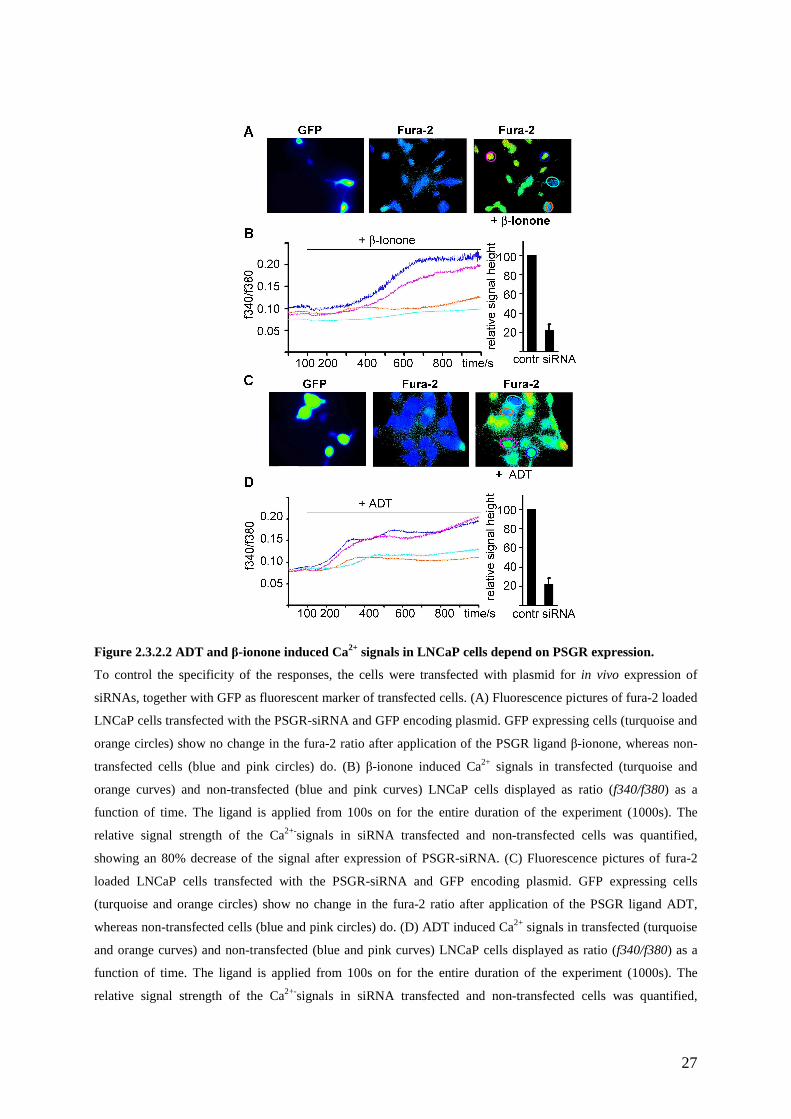
27
Figure 2.3.2.2 ADT and β-ionone induced Ca2+ signals in LNCaP cells depend on PSGR expression.
To control the specificity of the responses, the cells were transfected with plasmid for in vivo expression of
siRNAs, together with GFP as fluorescent marker of transfected cells. (A) Fluorescence pictures of fura-2 loaded
LNCaP cells transfected with the PSGR-siRNA and GFP encoding plasmid. GFP expressing cells (turquoise and
orange circles) show no change in the fura-2 ratio after application of the PSGR ligand β-ionone, whereas non-
transfected cells (blue and pink circles) do. (B) β-ionone induced Ca2+ signals in transfected (turquoise and
orange curves) and non-transfected (blue and pink curves) LNCaP cells displayed as ratio (f340/f380) as a
function of time. The ligand is applied from 100s on for the entire duration of the experiment (1000s). The
relative signal strength of the Ca2+-signals in siRNA transfected and non-transfected cells was quantified,
showing an 80% decrease of the signal after expression of PSGR-siRNA. (C) Fluorescence pictures of fura-2
loaded LNCaP cells transfected with the PSGR-siRNA and GFP encoding plasmid. GFP expressing cells
(turquoise and orange circles) show no change in the fura-2 ratio after application of the PSGR ligand ADT,
whereas non-transfected cells (blue and pink circles) do. (D) ADT induced Ca2+ signals in transfected (turquoise
and orange curves) and non-transfected (blue and pink curves) LNCaP cells displayed as ratio (f340/f380) as a
function of time. The ligand is applied from 100s on for the entire duration of the experiment (1000s). The
relative signal strength of the Ca2+-signals in siRNA transfected and non-transfected cells was quantified,

28
showing an 80% decrease of the signal after expression of PSGR-siRNA. Data are the mean from 5 independent
experiments. Error bars represent SEM.
2.3.3 PSGR activation elicits phospholipase C mediated cell signaling in LNCaP cells.
To understand the origin of the β-ionone induced Ca2+ rise we performed a series of
experiments to further elucidate the transduction mechanism that is used by PSGR. Most
GPCRs are coupled to either adenylate cyclase (AC) or phospholipase C (PLC) via
heterotrimeric G-proteins. ORs in the olfactory epithelium couple to adenylate cyclase via the
activation of the specific G-protein Gαolf which in turn activates ACIII to convert ATP to
cAMP, ultimately opening cAMP gated ion channels (Jones and Reed, 1989; Zufall et al.,
1994). Phospholipase C is activated by Gαq proteins and converts phosphatidylinositol (PIP2)
to inositol triphosphate (IP3) and diacylglycerol (DAG). IP3 then opens membrane channels to
release calcium from the endoplasmic reticulum, DAG increases the activity of intracellular
enzymes, as e.g. protein kinase C.
To determine the origin of the agonist-evoked Ca2+ rise in LNCaP cells, we used extracellular
media with varying Ca2+ content. The β-ionone-evoked Ca2+ increase still occurs (with the
similar signal amplitude) after removal of the extracellular Ca2+, suggesting that most of the
Ca2+ entered the cell from an intracellular source (Figure 2.3.3A). This is further confirmed by
the fact, that depletion of the Ca2+ stores with thapsigargin, an inhibitor of intracellular
calcium (SERCA) pumps, before application of the ligand completely abolished the β-ionone
induced cytosolic Ca2+ increase (Figure 2.3.3B). To find out which signaling pathway is
involved in the β-ionone induced Ca2+ influx we treated LNCaP cells with inhibitors of key
enzymes. Pre-incubation with the adenylate cyclase inhibitor MDL caused no changes in the
β-ionone induced Ca2+ increase, whereas pretreatment with the phospholipaseC inhibitor
U73122 completely abolished the Ca2+ signal (Figure 2.3.3C). Τhe inactive analogue U73343
did not change the Ca2+ signal induced by β-ionone in LNCaP cells.
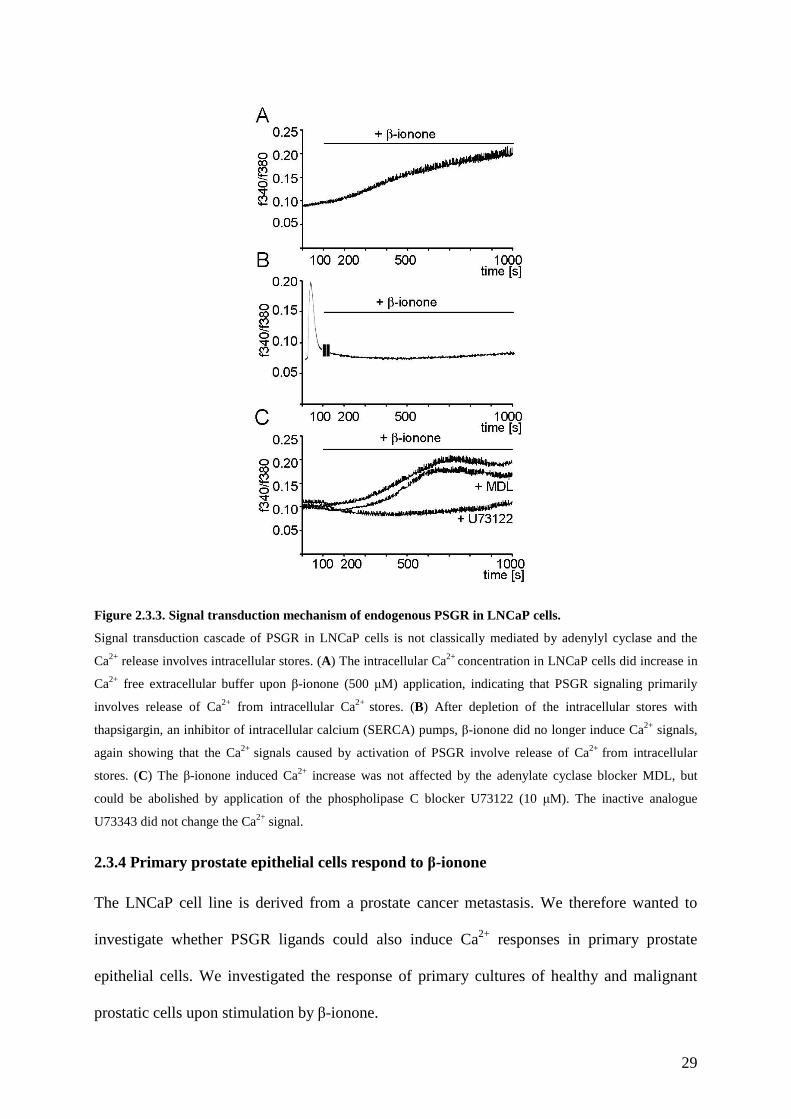
29
Figure 2.3.3. Signal transduction mechanism of endogenous PSGR in LNCaP cells.
Signal transduction cascade of PSGR in LNCaP cells is not classically mediated by adenylyl cyclase and the
Ca2+ release involves intracellular stores. (A) The intracellular Ca2+ concentration in LNCaP cells did increase in
Ca2+ free extracellular buffer upon β-ionone (500 µΜ) application, indicating that PSGR signaling primarily
involves release of Ca2+ from intracellular Ca2+ stores. (B) After depletion of the intracellular stores with
thapsigargin, an inhibitor of intracellular calcium (SERCA) pumps, β-ionone did no longer induce Ca2+ signals,
again showing that the Ca2+ signals caused by activation of PSGR involve release of Ca2+ from intracellular
stores. (C) The β-ionone induced Ca2+ increase was not affected by the adenylate cyclase blocker MDL, but
could be abolished by application of the phospholipase C blocker U73122 (10 µΜ). The inactive analogue
U73343 did not change the Ca2+ signal.
2.3.4 Primary prostate epithelial cells respond to β-ionone
The LNCaP cell line is derived from a prostate cancer metastasis. We therefore wanted to
investigate whether PSGR ligands could also induce Ca2+ responses in primary prostate
epithelial cells. We investigated the response of primary cultures of healthy and malignant
prostatic cells upon stimulation by β-ionone.

30
Figure 2.3.4. Activation of PSGR in primary prostate epithelial cells.
To investigate PSGR signaling in primary prostate epithelial cells, epithelial cell culture was performed from
prostatic tissue that was obtained from resections arising from CaP (prostate cancer epithelial, PCE) or healthy
prostatic tissue (prostate epithelial, PE). (A) To investigate whether the cultured cells resemble differentiated
secretory epithelial cells, we checked for expression of marker RNAs by RT-PCR. The cultured primary cells, as
well as LNCaP cells, express AR, PSA, PSGR and the cytokeratins 8 and 18, which are hallmarks of prostatic
secretory cells. (B) Phase-contrast picture of the epithelial cell culture from healthy prostate tissue. Primary
prostate epithelial cells were investigated for activation by the identified PSGR ligands using Ca2+-imaging. β-
ionone and ADT induced Ca2+ signals are displayed as ratio (f340/f380) as a function of time. The ligands are
applied from 100s on for the entire duration of the experiment (1000s). The β-ionone induced Ca2+ increase
could be abolished by application of the phopsholipaseC blocker U73122, similar as in LNCaP cells, indicating
that the signaling mechanism is similar in both cell types. (C) Phase-contrast picture and Ca2+-imaging of the
epithelial cell culture from prostate cancer, performed as described in (B).
The characteristics of our primary cultures resembled that of human prostatic epithelial cells,
with expression of marker RNAs for epithelial prostate cancer cells (cytokeratin 8, cytokeratin
18, prostate specific antigen, and androgen receptor, shown in Figure 2.3.4A) as well as a
typical morphology (Figure 2.3.4B, C). Moreover, RT-PCR analysis also revealed that the
cells express PSGR (Figure 2.3.4A).

31
We then investigated whether the primary cells can also be activated by the PSGR ligands β-
ionone (500 µΜ) and the steroid ADT (50 µΜ) (Figure 2.3.4D, E).
Applying both agonists elicited a slow, gradual increase in the intracellular Ca2+ concentration
within approximately 10 min (Figure 2.3.4D, E). The Ca2+ signal was overall similar to the
signal in LNCaP cells. In addition, we could block the PSGR induced Ca2+ rise in the primary
cells by pre-incubation of the cells with the phospholipase C inhibitor U73122 (Figure 2.3.4D,
E). Together these results show that PSGR is expressed in primary prostate epithelial cells and
can be activated in a similar manner as in LNCaP cells.
2.3.5 Effect of β-ionone on apoptosis and proliferation of LNCaP and primary prostate
epithelial cells
In order to examine the effect of PSGR signaling in prostate cancer cells, we treated cells with
250 µm β-ionone for 72-h and 144-h, which resulted in a significant reduction of LNCaP and
primary prostate cell (normal prostate epithelial cells and prostate cancer epithelial cells)
proliferation (Figure 2.3.5A, C, E) with a maximal inhibition at 144-h treatment (~50%) for
the LNCaP cell line.
To modulate cell proliferation, we tested DHT in all three cell types and found the expected
induction of the proliferation rates. When either LNCaP or primary cells were treated with a
mixture of DHT and β-ionone, the proliferative effect of DHT was suppressed (Figure 2.3.5A,
C, E).
We then examined whether the reduced cell proliferation rates observed after β-ionone
treatment were caused by increased apoptosis rates. Treatment of LNCaP (Figure 2.3.5B) and
primary (Figure 2.3.5D, F) cells with either 5 or 250 µm β-ionone caused an increase in
apoptotic cells as detected by DNA fragmentation.

32
Figure 2.3.5. Activation of PSGR in LNCaP and primary prostate epithelial cells reduces cell proliferation
involving MAPK signaling.
PSGR activation resulted in reduced cell proliferation and an induction of apoptosis in LNCaP cells and in
primary prostate epithelial cells. (A) Proliferation on LNCaP cells after application of DHT, a well-known
inducer of prostate cell proliferation, β-ionone, and a mixture of DHT and β-ionone for 3 and 6 days, relative to
control conditions without any treatment. β-ionone slowed cell proliferation, and could even reduce the DHT
stimulated increased proliferation rates. (B) LNCaP cells were treated with different concentrations of β-ionone
for 72 hours, induction of apoptosis was investigated by DNA fragmention, which can be seen by the increased
intensities of low molecular weight bands. The same experiments were performed with the primary prostate
cancer epithelial cells (C, D) and healthy primary prostate epithelial cells (E, F), which were treated with DHT
and β-ionone for either 3 or 6 days, showing that the PSGR ligand β-ionone could also reduce cell proliferation
in these cells. (D, F) β-ionone also induced apoptosis in primary prostate epithelial cells. (G) PSGR stimulation
by β-ionone results in phosphorylation of SAP/JNK and p38 MAPK in LNCaP cells. Western blot analysis of
total cell lysate of untreated LNCaP cells (0) and cells that were treated with β-ionone for 5, 10, 20 and 30 min.
Lysates were probed for the presence of phosphorylated SAP/JNK MAPK and phosphorylated p38 MAPK using
antibodies that specifically recognize the phosphorylated form of the proteins. Detection of the total amounts of
SAP/JNK and p38 are shown as controls. (H) Inhibition of p38 and JNK MAPK abolished the effect of�β-
ionone on LNCaP cell proliferation. The inhibitors were applied together with β-ionone for 6d (I+p38; I+JNK)

33
and cell proliferation rates were measured, control cells (C) and β-ionone (I) exposed cells were treated with
equal concentrations of DMSO as used for the inhibitors. Treatment of LNCaP cells with only the inhibitors did
not influence cell proliferation (data not shown).
2.4 Discussion
In the present study, we describe the functional characterization of PSGR, an “orphan”
prostate specific G-protein coupled receptor that is overexpressed in prostate cancer cells.
PSGR belongs to the superfamily of ORs (OR51E2), which are expressed in the neurons of
the olfactory epithelium where they detect volatile molecules. In addition, members of the OR
family have been found in various ectopic tissues, including sperm, where they were
implicated in reproductive functions. In both tissues, ORs are responsible for the recognition
of diverse chemical signals. In an attempt to determine the significance of the OR PSGR in
prostate tissue, we were successful in the de-orphanization of the receptor and identified β-
ionone and steroid hormones with a similar structural motif as PSGR ligands. PSGR
stimulation affects Ca2+ homoestasis in LNCaP prostate cancer cell lines and primary prostate
epithelial cells and seems to play an important role in prostate cancer cell proliferation.
The PSGR induced Ca2+ increase can be abolished by blocking phospholipase C. It still takes
place in Ca2+ free extracellular solution, but not after depletion of the intracellular Ca2+ stores
with thapsigargin. These are the properties of Gαq-coupled receptors. PSGR is therefore the
second example of a mammalian OR that can mobilize Ca2+ elevation via an endogenous PLC
signaling pathway and does not couple to adenylyl cyclase using cAMP as second messenger.
Previous biochemical measurements have shown that some odorants increase inositol-1,4,5-
trisphosphate (IP3) in mammalian olfactory neurons (Boekhoff et al., 1990; Breer et al., 1990;
Breer, 1993; Ronnett et al., 1993). However, the corresponding ORs were not identified.
Odorant-activated PLC signaling also occurs in microvillar cells of the olfactory epithelium
that contain PLCβ2 and IP3 receptor type 3 (IP3R-3), as well as a plasma membrane transient
receptor potential (TRP) channel TRPC-6 (Elsaesser et al., 2005). Recently it was shown, that

34
ORs can directly activate the PLC pathway to stimulate Ca2+ elevation, as OR U131 was
shown to mediate odor stimulation of the PLC/IP3 pathway in Odora cells (Liu et al., 2006).
Previous studies demonstrated that there is excessive signaling by some GPCRs such as
endothelin A receptor (Godara et al., 2005), bradykinin 1 receptor (Taub et al., 2003), follicle
stimulating hormone receptor (Ben-Josef et al., 1999) and thrombin receptor (Chay et al.,
2002; Cooper et al., 2003) in prostate cancers due to strong overexpression of the respective
receptors. The neuropeptides neurotensin and bombesin, acting through Gq-coupled receptors,
were shown to activate AR and enhance the androgen-independent growth of prostate cancer
cells (Dai et al., 2002; Lee et al., 2001). AR activation by bombesin was shown to be
dependent on ERK1/2 and non-receptor tyrosine kinase Src followed by EGF receptor
transactivation (Xiao et al., 2003). Reduction of RGS2, which can inhibit several Gq-coupled
GPCRs, is implicated in solid prostate tumor development (Silva et al., 2003) and was
recently associated with the acquisition of androgen-independence by prostate cancer cells
(Cao et al., 2006). Regulation of the Gq-coupled signaling pathway by RGS2 may therefore
be physiologically important in the regulation of prostate cancer cell growth. Also the orphan
prostate-specific olfactory receptor PSGR has been shown to be overexpressed in prostate
cancer (Weng et al., 2005b; Weng et al., 2005a; Xu et al., 2000; Xu et al., 2006). PSGR also
affects prostate cancer cell growth, but differs from the above described examples of Gq
coupled GPCRs in prostate cells due to the observed reduction in the cell proliferation rates
upon receptor stimulation.
We here show, that PSGR can be activated by the odorant β-ionone, as well as by steroid
hormones, and is therefore a novel membrane receptor being responsible for mediating rapid,
non-genomic effects of steroids on intracellular signaling cacades. Evidences for non-genomic
steroid effects and distinct receptors involved exist for all steroid groups including vitamin D
(3) and thyroid hormones (Losel and Wehling, 2003). According to the Mannheim Criteria
(Falkenstein et al., 2000), non-genomic responses to steroids must be observed within min, at

35
low steroid concentrations, should not be affected by inhibitors of transcription or translation,
and should be present in the presence of antagonists for the classic receptors. All of these
criteria hold true for the activation of PSGR in LNCaP cells by the testosterone derivatives
that we identified as PSGR ligands. Most importantly, we proved the transmission of the rapid
steroid effect being PSGR mediated by showing that a reduction of the amount of PSGR in
RNAi experiments abolished the steroid induced Ca2+ influx. Membrane androgen sites have
already been shown on the cell surface of LNCaP (Kampa et al., 2002) and in the AR negative
prostate cancer cell line DU-145 (Hatzoglou et al., 2005). Similar to PSGR, these binding
sites are preferentially expressed in prostate cancer cells as compared to normal cells
(Dambaki et al., 2005; Stathopoulos et al., 2003). 5α-dihydrotestosterone (DHT) was shown
to produce a concentration-dependent fast and transient increase in the intracellular Ca2+
concentration in LNCaP cells, which was abolished by removal of extracellular Ca2+ or L-
type Ca2+ channel inhibitors (Steinsapir et al., 1991; Sun et al., 2006). The DHT response was
shown to be linked to a GPCR, but was insensitive to the inhibition of phospholipase C (Sun
et al., 2006). Similar to other non-classical membrane steroid receptors (Losel et al., 2002;
Mendiberri et al., 2006; Picotto et al., 1999), PSGR signaling involves phospholipase C
mediated release of calcium from intracellular stores. Due to the fact that we did not observe
activation of the recombinant PSGR by DHT, there might be very likely additional steroid
binding sites on the surface of LNCaP cells. We performed our studies additionally on tissue
isolated from prostate cancer patients and found that these cells also respond to PSGR ligand
with a rapid, phospholipase C dependent Ca2+ influx.
One of the steroid ligands we identified is 6-dehydrotestosterone, which can be made
endogenously by conversion of testosterone. Testosterone, which is produced by Leydig cells
of the testes, is the major androgen in most mammalian species, but androgen metabolites can
make up a significant fraction of circulating steroids. Testosterone conversion to
dihydrotestosterone (DHT) is known to amplify the testosterone action on androgen receptors

36
(ARs). Testosterone was shown to be converted to 6-dehydrotestosterone, together with 6β-
hydroxytestosterone, 2β-hydroxytestosterone, and 15β-hydroxytestosterone, by a cytochrome
P450 belonging to the CYP3A family (Halvorson et al., 1990; Nagata et al., 1986). The
cytochrome P-450 (CYP450) monooxygenase system consists of enzymes that, among a
multitude of other functions, are responsible for metabolism of steroid hormones. The organ
that expresses the highest levels of CYP450 is the liver, which plays the dominant role in
steroid hormone metabolism. 6-dehydrotestosterone binds the steroid binding domain of rat
androgen-binding protein (rABP) (Danzo et al., 1991), indicating that it might have
physiological relevance. The other steroid structures that we identified, solely based on
similarity to� β-ionone and 6-dehydrotestosterone, are not known to occur in the human body
as major metabolites. ATD (1,4,6-androstatriene-3,17-dione) is known to function as
aromatase (CYP19) inhibitor preventing the aromatization of androgens to estrogens (Brodie
et al., 1983), and is used in pharmaceutical treatments of cancer, including cancer of the
prostate. Interestingly, this component also has a negative proliferatory effect on prostate
epithelial cells, which express PSGR.
Testosterone membrane binding in LNCaP cells results in a rapid modification of the actin
cytoskeleton and an increased PSA secretion (Kampa et al., 2002; Papakonstanti et al., 2003).
In addition, the activation of membrane androgen receptors is known to induce cell apoptosis
and to reduce cancer cell migration and adhesion (Hatzoglou et al., 2005; Hendriksen et al.,
2006; Kampa et al., 2002). We show here, that PSGR signaling also influences proliferation
and apoptosis of LNCaP, as well as primary prostate cancer cells. It has been suggested that
the membrane androgen receptor induced Ca2+ elevation reduces AR expression and promotes
apoptosis (Hatzoglou et al., 2005; Kampa et al., 2002), similar antiproliferative effects via the
induction of store dependent Ca2+ signaling have been reported for the G-protein coupled
P2Y-purinergic receptor (Thebault et al., 2005). In general, the Ca2+ content in the
endoplasmic reticulum seems to play a crucial role in the regulation of prostate cancer cell

37
apoptosis (McConkey et al., 1996; Skryma et al., 2000; Vanden et al., 2002). The thapsigargin
induced apoptosis has been correlated to the activation of c-jun NH2 terminal kinase (JNK), a
member of the MAPK family (Engedal et al., 2002). MAPK family members which are
involved in the transduction of signals from the cell surface to the nucleus, have also more
generally been implicated in the regulation of apoptosis. The three major members of the
MAPK family are the extracellular stress regulated kinase (ERK), which contributes to cell
differentiation, proliferation and survival, and JNK and p38, which can promote apoptosis.
We investigated the involvement of MAPK family members in the PSGR signaling cascade
and found, that p38 and JNK, but not ERK1/2 were phosphorylated upon PSGR activation.
Moreover, we could show that inhibition of p38 and JNK outweighed the effect of β-ionone
on the cell proliferation rate of LNCaP cells. The inhibitor of p38 MAPK already had strong
effects at concentrations near the IC50 of the inhibitor, while JNK inhibitor only worked at
much higher concentration, where also p38 might already be at least partially inhibited. The
involvement of p38 MAPK in the PSGR signaling cascade therefore seems relatively clear,
while the participation of JNK is not. p38 generally plays an important role in induction of
apoptosis in prostate cancer cells (Tanaka et al., 2003) and may therefore contribute to cancer
development, it was shown to be necessary for human prostate cancer cell apoptosis induced
by 2-methoxyestradiol, an endogenous metabolite of estradiol-17β and by phorbol 12-
myristate 13-acetate/PKC (Davoodpour and Landstrom, 2005; Shimada et al., 2003; Shimada
et al., 2004). Also the reported P2Y receptor mediated invasiveness of prostate cancer cells is
affected by p38 (Chen et al., 2004). JNK can also control cell life or death, but its impact of
JNK on prostate cancer is more controversial (Shimada et al., 2006). Non activated p38 is
expressed in non-neoplastic prostate epithelial or basal cells, and is strongly activated in
hyperplastic or cancer cells from well and moderately differentiated cancers, but is absent in
poorly differentiated cancers or metastatic lesions (Uzgare et al., 2003).

38
Rapid, non-genomic, steroid actions have already been identified in cells bearing or not
intracellular receptors, but with the exception of progesterone and estradiol, for which seven-
loop GPCRs have been identified, solely evidences existed for membrane-related steroid
actions. In the present study, we showed the existence of another specific G-protein coupled
membrane steroid receptor, PSGR, which belongs to the superfamily of odorant receptors. In
addition, we found that major intracellular signaling cascades involved in cell survival and/or
apoptosis are activated by PSGR. In future, PSGR may be used as a membrane steroid
receptor in cancer control strategies as major or adjuvant chemotherapeutic agent, providing
new possible targets for cancer chemotherapy.
3. Functional investigation of OR51E2 (PSGR) in mammalian spermatozoa.
3.1 Research background
In mammals, the mature spermatozoon is a fairly small, highly polarized and
hydrodynamically shaped motile cell that consists of a head and a tail. The head contains the
condensed nucleus with all genetic traits from the father, whereas the tail provides the motility
necessary to transport the sperm to the site of fertilization and to assure the appropriate
orientation to penetrate the coatings of the ovum.
The nucleus forms the greater part of the head and it is covered on its anterior two-thirds by
the acrosome that is a cap-like membrane limited structure, derived from Golgi. The sperm
acrosome plays an important role at the site of sperm-zona (egg) binding during the
fertilization process. Immediately after sperm-zona binding, the outer membrane fuses with
the overlying plasma membrane, releasing enzymatic contents of the acrosomal at the site of
binding. These enzymes include a variety of acid glycohydrolases, proteinases, phophatases,
esterases and aryl sulfatases (bou-Haila and Tulsiani, 2000).
The tail (flagellum) is structurally divided into four major parts: the connecting piece, the
midpiece, the principle piece, and the end piece [Figure 3.1.1 (Turner, 2003)]. The connecting

39
piece attaches to the implantation fossa of nucleus in the sperm head. From the remnant of the
centriole at this point, the axoneme extends throughout the length of flagellum. Not only the
axoneme but also the midpiece begins at the connecting piece and is characterized by the
presence of 9 outer dense fibers that lie outside each of the 9 outer axonemal microtubule
doublets and by a mitochondrial sheath that encloses the outer dense fibers and axoneme. The
midpiece terminates about one-fourth of the way down the sperm flagellum at the annulus,
which marks the beginning of the principle piece. At this point, the mitochondrial sheath ends
and the outer dense fibers associated with outer axonemal doublets 3 and 8 are replaced by the
2 longitudinal columns of the fibrous sheath. The presence of fibrous sheath and of only 7
(rather than 9) outer dense fibers surrounding the axoneme defines the principal piece. The
end piece contains only axoneme surrounded by plasma membrane. The significant difference
between sperm and ciliated/flagellated eukaryotic cells is that only sperm contains
mitochondrial sheath, outer dense fibers, and fibrous sheath besides axoneme [Figure 3.1.1
(Turner, 2003)]. The axoneme acts as flagellar motor. Activation of axonemal dynein ATPase
causes the sliding of adjacent outer doublet microtubules, which results in flagellar bending
(Tash and Means, 1982). The central pair and radial spokes regulate the size and shape of
axonemal bending in a calcium-dependent fashion (Wargo and Smith, 2003). The
mitochondrial sheath is a source of ATP for sperm. Sperm mitochondria are located only in
mitochondrial sheath of the midpiece and produce ATP for the cell. Sperm mitochondria have
been associated with several unique proteins that are not found in the mitochondria of somatic
cell. These proteins include sperm specific isoforms of lactate dehydrogenase and hexokinase
(Burgos et al., 1995; Travis et al., 1998). The outer dense fibers provide structural support. It
has been suggested that the role of the fibers provide passive elasticity to the motile flagellum
and structural support to the flagellum. However, the functions in sperm motility remain
speculative (Turner, 2003). The fibrous sheath also provides structural and a rigid support for
the flagellum and determines its planar beat (Lindemann et al., 1992). Moreover, multiple

40
members of both motility and metabolism related pathways localize to the fibrous sheath. It is
likely that it serves as a scaffold and organizing center for multiple signaling and metabolic
cascades that are critical for normal flagellar function (Brown et al., 2003; Eddy et al., 2003;
Miki et al., 2002; Turner et al., 1999).
Figure 3.1.1 Schematic representation of a mammalian sperm and the ultrastructure of the flagellum
(Turner, 2003). (A) Mammalian sperm flagella are structurally divided into 4 areas: the connecting piece,
midpiece, principal piece, and end piece. The end of the midpiece and start of the principal piece are demarcated
by the annulus. (B) Schematic cross-section through a representative segment of the midpiece showing the
plasma membrane (PM) and mitochondrial sheath (MS) surrounding the 9 outer dense fibers (ODFs). Within the
ODFs are the components of the axoneme: the 9 outer microtubule doublets of the axoneme (OMDA) with
associated dynein arms (DA) and radial spokes (RS) and the central pair of microtubule doublets (CP). (C)
Schematic cross-section through a representative segment of the principal piece showing the PM surrounding 7
ODFs. ODFs 3 and 8 have been replaced by the two longitudinal columns of the fibrous sheath (LC). The two
columns are connected by transverse ribs (TR). (D) Schematic cross-section through a representative segment of
the end piece. The ODFs and FS tapered at the termination of the principal piece and are no longer present in the
end piece, thus leaving only the PM to surround the axoneme.

41
While immotile during testicular maturation and epididymal transit, ejaculated spermatozoa
exhibit distinct tail beating patterns and swimming behaviors that are governed by
intracellular Ca2+ waves. Spermatozoa undergo a ripening process inside the female genital
tracts, known as capacitation. In the capacitation process, sperm cells undergo a series of
biochemical and functional changes, including an increase in membrane fluidity, cholesterol
efflux, increase of intracellular Ca2+ concentration, increased protein tyrosine phosphorylation
and induction of hyperactivation.
Ca2+ has a very important role in capacitation (Breitbart, 2002). It has been reported that there
is an increase in intracellular Ca2+ concentration in sperm cell during capacitation in several
mammalian species (Naz and Rajesh, 2004). Extracellular Ca2+ appears to be necessary for
the completion of capacitation but an increase of Ca2+ from intracellular stores is also required
for capacitation (Marquez et al., 2007). Influx of extracellular Ca2+ involves in various types
of channels localized on the plasma membrane of the sperm. These channels include voltage-
and agonist- regulated Ca2+ permeable channels (Darszon et al., 2005; Darszon et al., 2006;
Jimenez-Gonzalez et al., 2006). Expulsion of excess Ca2+ through the sperm plasma
membrane relies on ATP-driven Ca2+ pumps and Na+/Ca2+ exchangers (Wennemuth et al.,
2003). Although sperm cells are lack of endoplasmic reticulum, at least two intracellular
stores are present in the tiny cells within 5 µm of each other (Publicover et al., 2007). An
intracellular store localized on the outer acrosomal membrane is formed by an isoform of
sarcoplamic/endoplasmic reticulum Ca2+-ATPases (SERCAs) and IP3 receptors (Kuroda et al.,
1999; Lawson et al., 2007; Minelli et al., 2000; Walensky and Snyder, 1995). A second Ca2+
store is present in the connecting piece of mammalian sperm as the redundant nuclear
envolope (a remnant of excess membrane after nuclear condensation collects behind the
nuclear) and membrane vesicles (opposite to the plasmalemma in the cytoplasmic droplet of
sperm) contain calreticulin (a Ca2+ store protein), IP3 receptors and ryanodine receptor
(Harper et al., 2004; Ho and Suarez, 2003; Naaby-Hansen et al., 2001). Accumulation and

42
mobilization of Ca2+ from one or both stores are believed to initiate and/or control
hyperactivation in sperm of various mammalian species (Ho and Suarez, 2003; Marquez et al.,
2007), and to regulate the flagellum-beating pattern in human sperm (Harper et al., 2004).
Tyrosine phosphorylation of protein represents another very important aspects of capacitation.
It has also been shown to be associated with hyperactivated motility, sperm-zona binding and
acrosome reaction. The degree of tyrosine phosphorylation is correlated with capacitative
state of spermatozoa (Naz and Rajesh, 2004). The flagellum seems to be the major component
of sperm cell that undergoes tyrosine phosphorylation in most species (Naz and Rajesh, 2004).
In human sperm, at least 7 proteins are phosphorylated and 14 proteins are
autophosphorylated during capacitation. Some of them have been identified, including
fertilization antigen 1 (FA-1 antigen), protein A kinase anchoring proteins (AKAPs), calcium
binding and tyrosine phosphorylation-regulated protein (CABYR) (Naz and Rajesh, 2004).
FA-1 antigen has been shown to play an important role in sperm capacitation and sperm-zona
binding (Kaplan and Naz, 1992; Naz and Ahmad, 1994; Naz and Zhu, 2002). AKAP82, pro-
AKAP-82 and FSP 95 are the most prominent tyrosine phosphorylated proteins during
capacitation (Carrera et al., 1996; Mandal et al., 1999). Disruption of AKAP4 leads to defects
in sperm flagellum and motility (Miki et al., 2002). CABYR is localized on the principle
piece of sperm and it has been suggested to be involved in crosstalk between tyrosine
phosphorylation and Ca2+ mobilization in signal transduction pathway (Naaby-Hansen et al.,
2002).
After capacitation, the next step required for fertilization is called the acrosome reaction. The
acrosome reaction allows sperm to penetrate zona pellucida (ZP) and fuse with the egg. This
process is associated with the release of digestive enzymes and the fusion of the acrosome and
plasma membrane in the sperm head (bou-Haila and Tulsiani, 2000). The human ZP is
composed of four proteins, ZP1, ZP2, ZP3 and ZP4 (Caballero-Campo et al., 2006; Moos et
al., 1995). During the acrosome reaction, there is an increase in intracellular Ca2+

43
concentration, which involves at least three separate and sequentially linked Ca2+ events.
Initially, intracellular Ca2+ is elevated to micromolar levels within 50 ms and relaxes back to
the resting value within 200 ms. This process is associated with the opening of T-type voltage
gated ion channels (Arnoult et al., 1999). The second intracellular Ca2+ rise is slower,
sustained, and lasting several min and is necessary for AR completion. ZP binding causes an
IP3 increase which in turn releases Ca2+ from intracellular Ca2+ stores (De et al., 2002). The
generation of IP3 and the emptying of intracellular stores lead to further increase of
intracellular Ca2+ concentration due to the opening of store operated channels at the plasma
membrane (Darszon et al., 2006; Fukami et al., 2003). Progesterone is also known to induce a
Ca2+ response similar to ZP and may help to penetrate through the egg coat by inducing
hyperactivated flagellar movements (Harper et al., 2004).
During acrosome reaction, there is also an increase in tyrosine phosphorylation of sperm
proteins. In human sperm, ZP3/progesterone induced acrosome reaction is associated with an
increase of tyrosine phosphorylation of 95±5 kDa protein only in capacitated cells (Brewis
and Moore, 1997; Luconi et al., 1995). Additional to 95 kDa protein, other phosphorylated
proteins during acrosome reaction in human spermatozoa have also been characterized
including galactosyltransferase and FA-1 antigen (Baldi et al., 1996).
Two active mechanisms of sperm guidance have been shown in mammals, namely
thermotaxis and chemotaxis (Eisenbach and Giojalas, 2006). Thermotaxis is termed as the
directed movement of cells in a temperature gradient (from the cooler to the warmer).
Chemotaxis is termed as the movement of cells in the direction of chemoattractant that
attracts specific cells in a concentration gradient. The available information about the
molecular mechanisms of sperm chemotaxis in mammals is limited and mainly includes the
identity of some chemotaxis receptors and chemoattractant-induced intracellular Ca2+ changes
(Eisenbach and Giojalas, 2006).

44
The identification of odorant receptors in mammalian sperm and their localization to the
midpiece of the mature spermatozoa raise the possibility that some of these receptors might be
chemotaxis receptors (Eisenbach and Giojalas, 2006; Spehr et al., 2003). The first study
carried out in 1992 has demonstrated transcription of ~20 mamalian OR genes in male germ
cells, particularly in late stages of spermatogenesis (Parmentier et al., 1992). By employing
both RT-PCR and RNase protection assays, another 20 OR clones from each tested species
including mouse, rat, dog and human were found in testis cDNA and a maximal total number
of 50 OR genes was estimated (Vanderhaeghen et al., 1997b; Vanderhaeghen et al., 1997a). A
similar number was confirmed by using microarray assays to detect 66 OR genes in mouse
testis (Zhang et al., 2004b). Thus a following major challenge is to decipher the potential
physiological role of these receptors in sperm cells.
Employing polyclonal antibodies against specific ORs revealed that these ORs are localized
in the mid-piece or the base of the flagellum of spermatozoa from dog and human (Neuhaus et
al., 2006a; Vanderhaeghen et al., 1993; Walensky et al., 1995). Identification and
characterization of human OR17-4 (Spehr et al., 2003) and mouse OR MOR23 (Fukuda et al.,
2004) provided a further understanding of the potent role of ORs in mammalian sperm cells.
The activation of hOR17-4 by its specific ligand bourgeonal couples the signaling pathway to
the membrane bound isoform of adenylyl cyclase (Spehr et al., 2004b) and leads to the
activation of PKA (Neuhaus et al., 2006a). Most interestingly, a combination of
microcapillary accumulation assays and video motion analysis of swimming sperm revealed
that the ligand bourgeonal has strong and dose–dependent effects on sperm chemotaxis and
chemokinesis as well as hyperactive flagellar beating frequency (Spehr et al., 2004a; Spehr et
al., 2004b). These effects were completely abolished in the presence of the specific antagonist
of hOR17-4 undecanal (Spehr et al., 2004a; Spehr et al., 2004b). Therefore, OR17-4 and/or
other ORs might be involved in chemical guided sperm navigation and have an effect in
sperm-egg interaction.

45
In the study of MOR23, the transcripts were found in mouse testis and in the olfactory
epithelium (Asai et al., 1996) and in situ hybridization showed that the MOR23 mRNA is
present in late pachytene spermatogenic cells and round spermatids (Fukuda et al., 2004). The
MOR23 ligand lyral elicits transient intracellular calcium increase in dissociated
spermatogenic cells and mature spermatozoa (Fukuda et al., 2004). In a sperm accumulation
assay, spermatozoa migrated towards an increasing gradient of lyral. Tracking and sperm
flagellar analysis suggested that the Ca2+ increase caused by MOR23 activation leads to
modulation of flagellar configuration, resulting in chemotaxis (Fukuda et al., 2004).
OR51E2 (PSGR) was initially found to be almost exclusively expressed in prostate tissue (Xia
et al., 2001; Xu et al., 2000), additionally it has also been found in brain, colon and liver(Yuan
et al., 2001). Moreover, it has been demonstrated that OR51E2 displayed a very high
evolutionarily conservation: the mouse OR51E2 homolog diverges only at four positions from
the rat homolog RA1 protein, while both the rat and mouse homologs share 93% amino acid
identity to the human OR51E2 (Yuan et al., 2001). This suggested a possible conserved
physiological function of this receptor besides chemoreception in the epithelium and in the
prostate cancer.
In the present study, we found that OR51E2 is functionally expressed in human testis.
Activation of OR51E2 triggers the activation of PLC mediated pathway, the Ca2+ release of
intracellular stores and enhancement of capacitation. Sequencing analysis of OR51E2
homologs in various species revealed that they share more than 90% amino acid identity.
After conducting Ca2+ imaging on the mature spermatozoa from four species, we showed that
the cells responded to the same ligand, β-ionone.

46
3.2 Materials and methods
3.2.1 Odorants and reagents
Odorants used in the study were kindly provided by Dr. T. Gerke (Henkel KGaA, Düsseldorf,
Germany). Odorant solutions were directly suspended by sonication in the buffer used in each
assay at indicated concentration. The adenylyl cyclase III inhibitor (Calbiochem) MDL 13320
was used at 50 µM; SQ22536 was used at 10 mM; PLC inhibitor U733122 and the inactive
analogue U73343 were used at 10 µM; thapsigargin was used at 1 µM.
3.2.2 RT-PCR analysis and sequencing
RNA was extracted from testes (~100 mg) of various species by Trizol reagent and cDNA
was synthesized as described in the previous chapter. After amplification, the PCR products
(~100 ng) were used for sequence analysis.
The following primer pairs were used in the study.
OR51E2 degenerate primer (also used in the sequencing analysis):
Forward 3’-ATGAGTTCCTGCAACTTCAC-5’
Reverse 3’-CACAGCTGATCTTGAACATAGC-5’
3.2.3 Immunohistochemistry
Purified spermatozoa were incubated with standard Ringer’s solution and placed on poly-L-
lysine-coated coverslips and fixed in 4% PFA. Cells were permeablilized with 0.1% Triton X-
100 in PBS containing 1% cold-water fish skin gelatin (Sigma) and incubated with anti-PSGR
antibody (1:100) in PBS/gelatin/Triton X-100. After washing, coverslips were incubated with
fluorescently labeled secondary anti-rabbit IgG (1:1000) and mounted in ProLong Antifade
(Molecular Probes). All fluorescence images were obtained with a confocal microscope and
further processed with Photoshop.

47
3.2.4 Sperm preparation and imaging of Ca2+ levels in spermatozoa
For the imaging of Ca2+ levels in mouse sperm, spermatozoa were isolated from cauda
epididymis and incubated for 15 min at 37°C in Ham’s F10 medium (Invitrogen) containing
15 mM NaHCO3 (Sigma) and 5 mg/ml globulin free bovine serum albumin (BSA) (Sigma) to
disperse the spermatozoa. The sperm suspension was then collected in a 1.5 ml Eppendorf
tube, and F10 medium was added to a final volume of 1 ml. The cells were incubated for
10min at 37°C in an atmosphere of 5% CO2/95% air. The upper 50 µl layer of the sperm
suspension containing motile spermatozoa was transferred in to a new 1.5 ml Eppendorf tube
and loaded with fura-2 supplemented with 0.02% Cremophor/EL in Ringer’s solution. The
mixture was then incubated for 30 min at 37°C, and a 100 µl sample of loaded sperm
suspension was placed on a Concanavalin A (ConA) coated glass-bottom dish (WPI).
Sperm from other species were freshly collected from the epididymis of different animals
obtained from veterinary surgery and then inbubated in swim up medium (F-10 medium
containing 15 mM NaHCO3 supplemented with 0.3% BSA) for 30 min. The upper layer of the
suspension was collected and gently spun (500 g, 5 min) to collect the motile spermatozoa.
The cells were then loaded with 7.5µM fura-2 in Ringer for 30min and placed on ConA
coated glass-bottom dishes.
Human sperm were freshly obtained from young healthy donors and a PercollTM density
gradient centrifugation was performed after liquefaction (30 min at 35.5°C) to isolate mature
and motile sperm. In brief, liquefied semen was overlaid on a two-layer Percoll (Amersham
Biosciences) density gradient consisting of 5 ml 80% and 5 ml 55% isotonic PercollTM in
Ham’s F-10 medium (Invitrogen). After 40-min centrifugation at 500 g at room temperature
the pellet was collected, washed in standard Ringer’s solution (140 mM NaCl, 5 mM KCl, 2
mM CaCl2, 2 mM MgCl2, 10 mM Hepes, 10 mM glucose), and again centrifuged for 15 min.
Intracellular Ca2+ levels were then measured as described in the previous chapter.

48
3.2.5 Capacitation
2 ml Ham-F10 supplemented with 0.3% globulin-fatty acid free BSA (sigma A7030) were
placed in two 15 ml Falcon tubes. 1ml of fresh ejaculation was charged into a Pasteur pipette,
carefully released at the bottom of the tube. Probes were incubated for 1h at 37°C, 5% CO2 in
a 45° rack. 1 ml of the upper phase was carefully recovered and placed in a 15 ml tube, and 9
ml F10 culture medium supplemented with 0.3% BSA was added and centrifuged at 400g for
10min. One of the two pellets was resuspended in 1ml culture medium supplemented with
2.6% BSA, and the other with 0.3% BSA as negative control. Sperm motility and
concentration of the selected populations was determined and concentrations adjusted to
5×106/ml. Up to 2 ml per tube were incubated 180 min at 37°C, 5% CO2 in a 45° rack. 5 µΜ
β-ionone and 10 µΜ U73122 (Calbiochem) were added to the above medium in specific
experiments as described in the text. After capacitation procedure, sperm cells were stained
with 2.7 µM Merocyanine 540 (Sigma) in PBS at 37°C for 10 min. Capacitated cells are
determined by bright red fluorescence around acrosome region, and uncapacitated cells show
no fluorescence.
3.2.6 Detection of phosphorylated protein in human spermatozoa.
After treatment, spermatozoa were pelleted by centrifugation at 500 g for 5 min and washed
two times in PBS. Total cell lysates were extracted in Laemmli buffer and heated for 5 min at
100°C. Equivalent amount of protein was loaded, resolved by 8% SDS-PAGE and transferred
to nitrocellulose membrane. The membrane were subsequentially blocked with TBST
containing 5% blocking reagent from ECL advanced detection kit (Amersham) and incubated
with an anti-phosphotyrosine monoclonal antibody (Biomol) diluted to 1:1000 in 3% blocking
reagent in TBST. After washing and incubation with HRP coupled secondary antibodies,
detection was performed with ECL advance (Amersham) on hyperfilm ECL (Amersham).

49
The signals were quantified by densitometry with a Chemilimager v5.5 (Alpha Innotech
Corporation).
3.3 Results
3.3.1 OR51E2 is functionally expressed in human spermatozoa.
OR51E2 that is also named as prostate-specific G-protein coupled receptor (PSGR) was
shown to be expressed in prostate epithelial cells. According to EST database analysis and a
recent membrane proteomics study on human sperm (Barbour, 2006), PSGR is also supposed
to be expressed in human testis and mature spermatozoa. To investigate whether PSGR
(OR51E2) as an OR is expressed in the OE and whether PSGR is also present in testis, we
performed RT-PCR analysis of cDNA from a biopsy of human testis and olfactory epithelium.
Using a primer pair amplifying the intact open reading frame, we showed that the transcripts
of OR51E2 are present in testis as well as in the olfactory epithelium (Figure 3.3.1.A), and no
amplification was observed from the mRNA control. To ensure that the amplified DNA was
not from the possible trace amount of contaminating genomic DNA, we designed a pair of
intron-spanning primers across an intron between 5’-untranslated region (5-UTR) and ORF.
With this primer pair, the target band from cDNA should show the size of 810 base pairs,
which could be easily distinguished from the band potentially amplified from genomic DNA
(>15,000 base pairs) (Figure 3.3.1 B). Both RT-PCR results showed that the transcripts of
OR51E2 are present in human testicular cells. To further elucidate that the receptor is
expressed in human sperm cells at protein level and figure out the localization of the receptor
in human spermatozoa, we performed immunohistochemistry assays by employing the
specific antibody against the N-terminus of OR51E2. The immunostaining results showed that
OR51E2 is localized to the middle and principle piece of human spermatozoa and vesicular
structures of the sperm head (Figure 3.3.1 C).
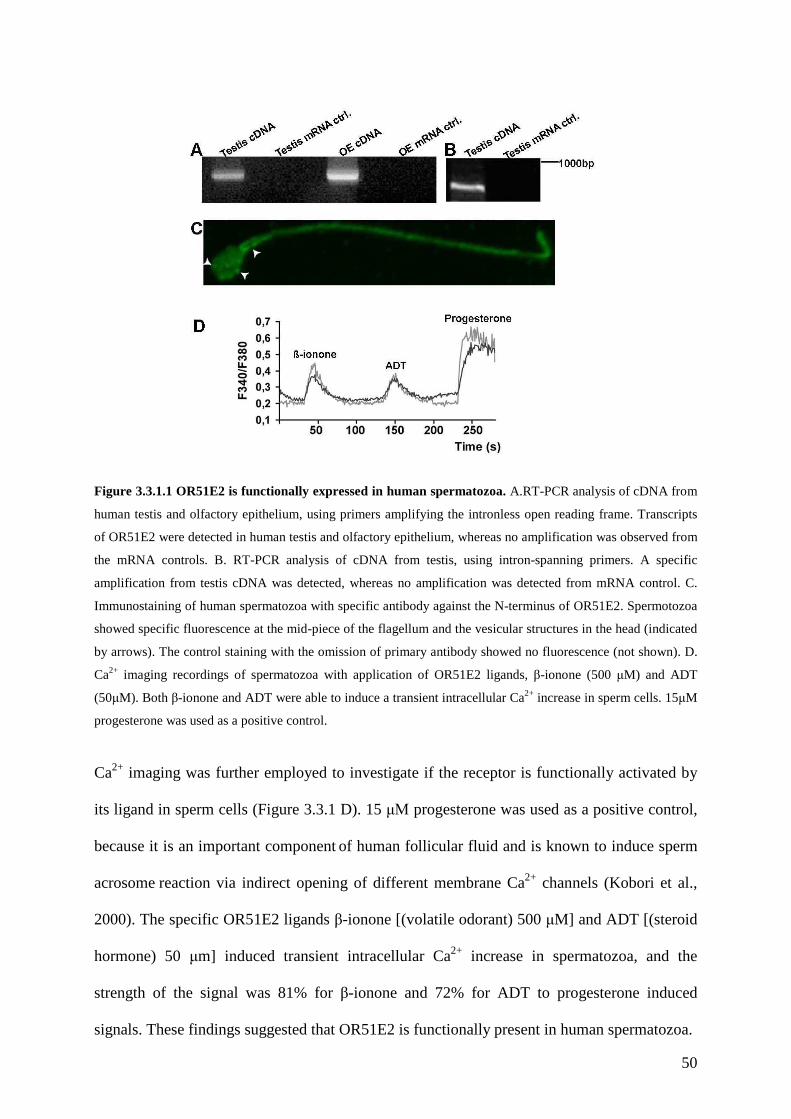
50
Figure 3.3.1.1 OR51E2 is functionally expressed in human spermatozoa. A.RT-PCR analysis of cDNA from
human testis and olfactory epithelium, using primers amplifying the intronless open reading frame. Transcripts
of OR51E2 were detected in human testis and olfactory epithelium, whereas no amplification was observed from
the mRNA controls. B. RT-PCR analysis of cDNA from testis, using intron-spanning primers. A specific
amplification from testis cDNA was detected, whereas no amplification was detected from mRNA control. C.
Immunostaining of human spermatozoa with specific antibody against the N-terminus of OR51E2. Spermotozoa
showed specific fluorescence at the mid-piece of the flagellum and the vesicular structures in the head (indicated
by arrows). The control staining with the omission of primary antibody showed no fluorescence (not shown). D.
Ca2+ imaging recordings of spermatozoa with application of OR51E2 ligands, β-ionone (500 µM) and ADT
(50µM). Both β-ionone and ADT were able to induce a transient intracellular Ca2+ increase in sperm cells. 15µM
progesterone was used as a positive control.
Ca2+ imaging was further employed to investigate if the receptor is functionally activated by
its ligand in sperm cells (Figure 3.3.1 D). 15 µΜ progesterone was used as a positive control,
because it is an important component of human follicular fluid and is known to induce sperm
acrosome reaction via indirect opening of different membrane Ca2+ channels (Kobori et al.,
2000). The specific OR51E2 ligands β-ionone [(volatile odorant) 500 µM] and ADT [(steroid
hormone) 50 µm] induced transient intracellular Ca2+ increase in spermatozoa, and the
strength of the signal was 81% for β-ionone and 72% for ADT to progesterone induced
signals. These findings suggested that OR51E2 is functionally present in human spermatozoa.

51
It has been suggested that the generation of odorant-induced Ca2+ signals in different
compartments of a sperm cell could correspond with the functional localization of the receptor
and its signal transduction machinery (Spehr et al., 2004b).
Figure 3.3.1.2 Spatiotemporal analysis of Ca2+ signals in single sperm cell. A. The Fura-2 loaded sperm cells
showed clear staining on the head, mid-piece and principle piece of flagellum. B. Spatiotemporal Ca2+ signals
induced by 500 µM β-ionone. The slopes of Ca2+ responses in the head, mid-piece and principle piece were
shown in blue, pink, and yellow, respectively. The cytosolic Ca2+ level is depicted as the integrated fluorescence
ratio (f340/f380) and viewed as a function of time. An application of β-ionone at 9s induced an instant intracellular
Ca2+ increase in the principle piece and a delayed response in the mid piece, whereas the response of the head
region showed significant delay. C. The average latency of different compartments of responding sperm cells.
From the recording of 15 sperm cells, the average latency for the 3 recorded compartments is 6.7±0.7s, 3.9±0.9s
and 2.4±0.8s, respectively. D. Spatiotemporal Ca2+ signals induced by 5µM tharpsigargin. The slopes of Ca2+
responses in the head, mid-piece and principle piece were shown in blue, pink, and yellow, respectively. An
application of tharpsigargin at 9s induced a synchronous response in three different subcellular compartments in
the cells (the difference in latency is less than 0.2s.). E. The average latency of different compartment of
thapsigargin responded sperm cell. The maximal difference is less than 0.2 s.
Spatiotemporal properties of β-ionone induced Ca2+ mobilization in individual spermatozoon
was analyzed. The Ca2+ mobilization in different compartments of spermatozoon (head,
flagellar mid piece and flagellar principle piece) was recorded separately by using a high-
resolution imaging (Figure 3.3.1.2 A and B). From the recordings of 15 individual sperm cells,

52
we showed that the Ca2+ mobilization initiated from the principle part of flagellum and
gradually transmitted to the midpiece and became maximized in the head region. By
comparing the average latency after application of the odorant (Figure 3.3.1.2 C), we found
that sperm head, midpiece and principle piece of flagellum gave responses to β-ionone with
the latency of 6.7±0.7s; 3.9±0.9s and 2.4±0.8s, respectively. However, by analyzing the
spatiotemporal response pattern induced by thapsigargin, we showed that the thapsigargin
induced more synchronous responses in different compartments of sperm cells (Figure 3.3.1.2
D and E). The maximal difference in latency of thapsigargin induced Ca2+ signals in three
compartments is less then 0.2s (Figure 3.3.1.2 E).
3.3.2 The activation of OR51E2 elicits phospholipase C mediated cellular signaling in
human spermatozoa.
After showing that OR51E2 is functionally expressed in human spermatozoa, we further
investigated the potential signaling pathway following receptor activation. In the LNCaP
prostate cell line, OR51E2 induced signaling that involves activation of PLC. To investigate if
the PLC signaling is also activated after stimulation of OR51E2 in sperm cells, the PLC
inhibitor U73122 was co-applied with the ligands for 5s in Ca2+ imaging on fura-2 loaded
human spermatozoa (Figure 3.3.2 B and D). We found that the application of 10 µM U73122
almost completely abolished the responses induced by the OR51E2 ligands (β-ionone and
ADT). However, its inactive analogue U73343 did not display the same effect at the same
concentration (data not shown).
A previous report pointed out that odorant receptor induced Ca2+ signaling in sperm leads to
the activation of membrane-bound isoform of AC (Spehr et al., 2004b). In order to further
clarify the role of AC in the OR51E2 signaling cascade, we applied the two AC inhibitors:
MDL-12330A (250 µM) for both soluble and membrane bounded isoforms and SQ22536 (10
mM) selectively for membrane forms. Consistently with our above findings, neither of the AC
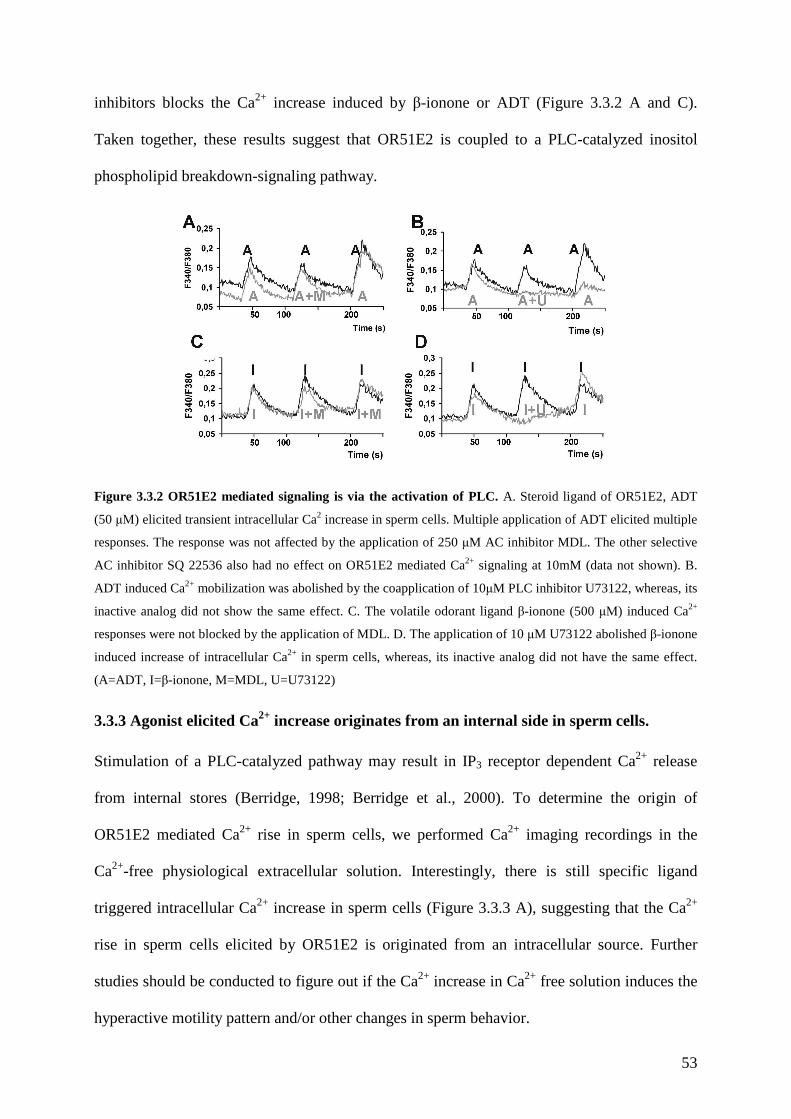
53
inhibitors blocks the Ca2+ increase induced by β-ionone or ADT (Figure 3.3.2 A and C).
Taken together, these results suggest that OR51E2 is coupled to a PLC-catalyzed inositol
phospholipid breakdown-signaling pathway.
Figure 3.3.2 OR51E2 mediated signaling is via the activation of PLC. A. Steroid ligand of OR51E2, ADT
(50 µΜ) elicited transient intracellular Ca2 increase in sperm cells. Multiple application of ADT elicited multiple
responses. The response was not affected by the application of 250 µM AC inhibitor MDL. The other selective
AC inhibitor SQ 22536 also had no effect on OR51E2 mediated Ca2+ signaling at 10mM (data not shown). B.
ADT induced Ca2+ mobilization was abolished by the coapplication of 10µM PLC inhibitor U73122, whereas, its
inactive analog did not show the same effect. C. The volatile odorant ligand β-ionone (500 µΜ) induced Ca2+
responses were not blocked by the application of MDL. D. The application of 10 µM U73122 abolished β-ionone
induced increase of intracellular Ca2+ in sperm cells, whereas, its inactive analog did not have the same effect.
(A=ADT, I=β-ionone, M=MDL, U=U73122)
3.3.3 Agonist elicited Ca2+ increase originates from an internal side in sperm cells.
Stimulation of a PLC-catalyzed pathway may result in IP3 receptor dependent Ca2+ release
from internal stores (Berridge, 1998; Berridge et al., 2000). To determine the origin of
OR51E2 mediated Ca2+ rise in sperm cells, we performed Ca2+ imaging recordings in the
Ca2+-free physiological extracellular solution. Interestingly, there is still specific ligand
triggered intracellular Ca2+ increase in sperm cells (Figure 3.3.3 A), suggesting that the Ca2+
rise in sperm cells elicited by OR51E2 is originated from an intracellular source. Further
studies should be conducted to figure out if the Ca2+ increase in Ca2+ free solution induces the
hyperactive motility pattern and/or other changes in sperm behavior.

54
Figure 3.3.3. Ca2+ increase induced by OR51E2 is originates from internal sources. A.The ligands of
OR51E2 β-ionone (500 µM) and ADT (50 µM) elicited transient Ca2+ mobilization in Ca2+ free extracellular
solution, similar as in Ca2+ containing solution. B. 5 µM thapsigargin led to the depletion of internal “stores” in
sperm cells and abolished the OR51E2 induced intracellular Ca2+ increase.
Although the presence of intracellular Ca2+ stores in sperm is controversial, emerging
evidences showed that some store specific proteins and channels are present in sperm (Harper
et al., 2004; Ho and Suarez, 2003; Kuroda et al., 1999; Lawson et al., 2007; Minelli et al.,
2000; Naaby-Hansen et al., 2001; Walensky and Snyder, 1995). Thapsigargin, an inhibior of
sarco-/endo-plasmic reticulum Ca2+/ATPase pumps (Thomas and Hanley, 1994), triggers
capacitative calcium entry in many cell types, leading to increase of intracellular Ca2+ and
depletion of Ca2+ stores (Putney, Jr. and McKay, 1999). In sperm cells, it has also been shown
that the application of thapsigargin resulted in the mobilization of intracellular Ca2+ and
acrosome reaction (Dorval et al., 2002; Walensky and Snyder, 1995). Here, we used
thapsigargin as a store-depleting reagent to further confirm and characterize the origin of
intracellular Ca2+ increase induced by the activation of OR51E2 in human spermatozoa. We
found that the application of 5 µM thapasigargin for 200s led to the depletion of the internal

55
Ca2+ sources. This directly resulted in the almost complete abolishment of OR51E2 induced
Ca2+ response in Ca2+ free extracellular solution (Figure 3.3.3 B). This provided us with
additional evidence supporting the conclusion that activation of OR51E2 leads to Ca2+
increase via internal Ca2+ sources.
3.3.4 Activation of OR51E2 enhances sperm capacitation via PLC activation.
After showing that OR51E2 induces a Ca2+ increase in human spermatozoa, we wanted to
know whether this Ca2+ increase is able to induce any change in sperm capacitation state.
Capacitation is a very important step of sperm-egg interaction and only capacitated sperm
cells can further process through acrosome reaction. For in vitro capacitation, mammalian
spermatozoa require removal of cholesterol from the sperm surface by BSA. As a
consequence, capacitation leads to a change of membrane fluidity and phosphorylation of
specific proteins at tyrosine sites. The markers of these changes can be therefore used as tools
to monitor capacitation in vitro. Merocyanine 540 (M540) was used as a marker of membrane
scramble to stain capacitated cells. In many species, fully capacitated sperm cells display
strong fluorescent staining of M540 on the membrane, whereas the noncapacitated cells show
no fluorescence (Flesch et al., 2001; Gadella et al., 1999; Rathi et al., 2001).
In the F10 medium supplemented with 2.6% BSA, ejaculated spermatozoa were incubated for
3 hours in the presence or absence of 5 µM β-ionone. The cells were then stained with M540
and the number of stained cells was counted under a fluorescence microscope. By calculating
the ratio of M540 stained cells to the total cell number, we showed that β-ionone significantly
enhanced capacitation and 10 µM U73122 abolished the effect (Figure 3.3.4A). However, the
U73122 alone did not significantly affect capacitation. At the end of the capacitation, tyrosine
phosphorylation status was examined (Figure 3.3.4 C and D). All the treatments led to protein
tyrosine phosphorylation in human sperm by showing two dominant tyrosine phosphorylated
protein bands at 85 kDa and 105 kDa and an additional 68 kDa band. However, incubation

56
with 5 µM β-ionone increased tyrosine phosphorylation by showing stronger signal and a
fourth band at 48 kDa. The upregulation effect was suppressed by 10 µM U73122 and the
U73122 alone did not significantly change the tyrosine phosphorylation.
Figure 3.3.4 The effect of β-ionone and the correlated signaling components on the sperm capacitation. A.
In the presence of 2.6% BSA, the sperm cells show 37% capacitated cells. 5 µM β-ionone enhanced capacitation
significantly, and this effect was blocked by U73122. However, 10 µM U73122 alone did not change the
percentage of capacitated cells significantly. B. In the absence of BSA, 5 µΜ β-ionone significantly increased
capacitation. C. Western blot detection of tyrosine phosphorylated protein in capacitated sperm. The detected
tyrosine phosphorylated protein bands were indicated by the asterisks. D. Tyrosine phosphorylation was
quantified by densitometry with a chemilimager 5.5 (Alpha Innotech). The graph depicts the average from 3
independent experiments. Error bars represent SEM.(I= β-ionone; U=U73122; ctrl.=control)
Furthermore, we also tested the effect of β-ionone on sperm capacitation in F10 medium with
the omission of BSA. In this case, the basal level of capacitation was much smaller
(8.3±2.5%). This is also expected, as without BSA in the medium cholesterol cannot be
efficiently removed from the sperm membrane. Furthermore, in the presence of 5 µM β-
ionone (in F10 medium only), the percentage of capacitated cells is significantly higher than
basal level (Figure 3.3.4 B).

57
3.3.5 Homologs of OR51E2 are present in the testes of various mammalian species.
It has been found that OR51E2 displayed an evolutionary conservation: the mouse OR51E2
homolog diverges only at four positions from the homologous rat protein RA1, while both the
rat and mouse homologs share 93% amino acid identity to the human OR51E2 (Yuan et al.,
2001). Therefore, we would like to test the degree of conservation of OR51E2 in more
mammalian species. Since we have discovered that OR51E2 was able to induce intracellular
Ca2+ increase in human sperm, we were particularly interested in whether the OR51E2
transcripts are present in the testes of other mammalian species. First of all, we aligned the
DNA sequences of OR51E2 and its homologs from mouse, human, chimpanzee and rhesus
monkey (Figure3.3.5.1). Interestingly, we found a highly conserved region with 100%
identity at the N-terminus with 20 base pairs, and another highly conserved region of 22 base
pairs at C-terminus. We designed a primer pair bases on these two conserved regions to
amplify OR51E2 homologs in testis cDNA from cat, dog, ferret, guinea pig, horse, rabbit and
rat. The transcripts of OR51E2 homologs were detectable in the testes out of all these seven
species, (Figure 3.3.5.2 A).
Figure 3.3.5.1 Degenerate primer design for OR51E2 homogolous sequences. The conserved regions at N-
terminus and C-terminus are labelled in red, and these regions were further used as primers to amplify putative
OR51E2 homologs from other mammalian species.
For the testis cDNA from mouse, we used specifically designed intron-spanning primers
across an intron between the 5’-UTR and the ORF. RT-PCR results with these primers
demonstrated that the transcripts of mouse OR51E2 homolog are present in the testis (Figure
3.3.5A).
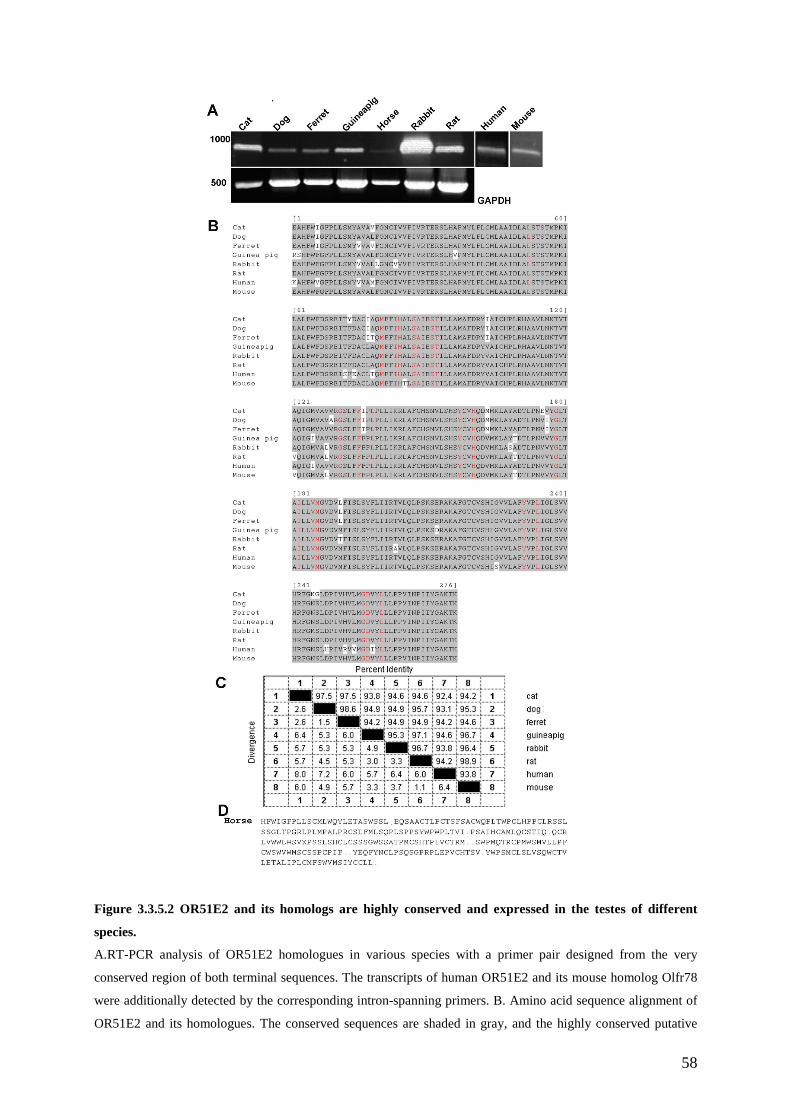
58
Figure 3.3.5.2 OR51E2 and its homologs are highly conserved and expressed in the testes of different
species.
A.RT-PCR analysis of OR51E2 homologues in various species with a primer pair designed from the very
conserved region of both terminal sequences. The transcripts of human OR51E2 and its mouse homolog Olfr78
were additionally detected by the corresponding intron-spanning primers. B. Amino acid sequence alignment of
OR51E2 and its homologues. The conserved sequences are shaded in gray, and the highly conserved putative

59
ligand binding sites were labelled in red. C. The table of sequence distance of OR51E2 homologs. These
homologs share very conserved primary amino acid sequences to human OR51E2. D. The homolog in horse
seems to be a pseudogene, as multiple stop codons (dots in the sequence) appear in the putative coding sequence.
After sequencing the amplified PCR products of putative OR51E2 homologs, we found out
that the horse homolog is a pseudogene by showing multiple stop codons in the open reading
frame (Figure 3.3.5 D). The others turned out to be the intact homologs of OR51E2. The
sequence alignment analysis of OR51E2 homologs showed that the DNA sequences have 86-
91% identity to human OR51E2. The amino acid sequences of functional homologs share
more than 92% identity to human OR51E2 with variations at different positions (Figure
3.3.5C). According to the prediction (Man et al., 2004), 22 sequence positions are predicted as
odorant binding sites. They are disposed on transmembrane helices 2 to 7 and on the second
extracellular loop of odorant receptors. We particularly checked these positions for all the
functional homologs, and intriguingly they share 100% identity to human OR51E2 at these
sites (Figure 3.3.5.B). This suggests that they are potentially able to recognize the similar
ligands as the human receptor.
3.3.6 OR51E2 and its homologs may have conserved function in spermatozoa from
different mammalian species.
Following the identification of the transcripts of OR51E2 in testicular cells from various
species, we tested if these receptors may have conserved function. We collected fresh sperm
cells from cat, guineapig, rabbit, mouse and horse, and stimulated these sperm cells with β-
ionone that was identified as human OR51E2 ligand, followed by performing fura-2 based
Ca2+ imaging on these cells. The viability of the cells was controlled by the reponses to K+8.6.
As we predicted from sequence alignment analysis, they all responded to β-ionone, however,
with different kinetics and different response amplitude (Figure 3.3.6). Cat sperm showed an
instantaneous response, and lasted for about 20s (Figure 3.3.6 A and G). Sperm cells from
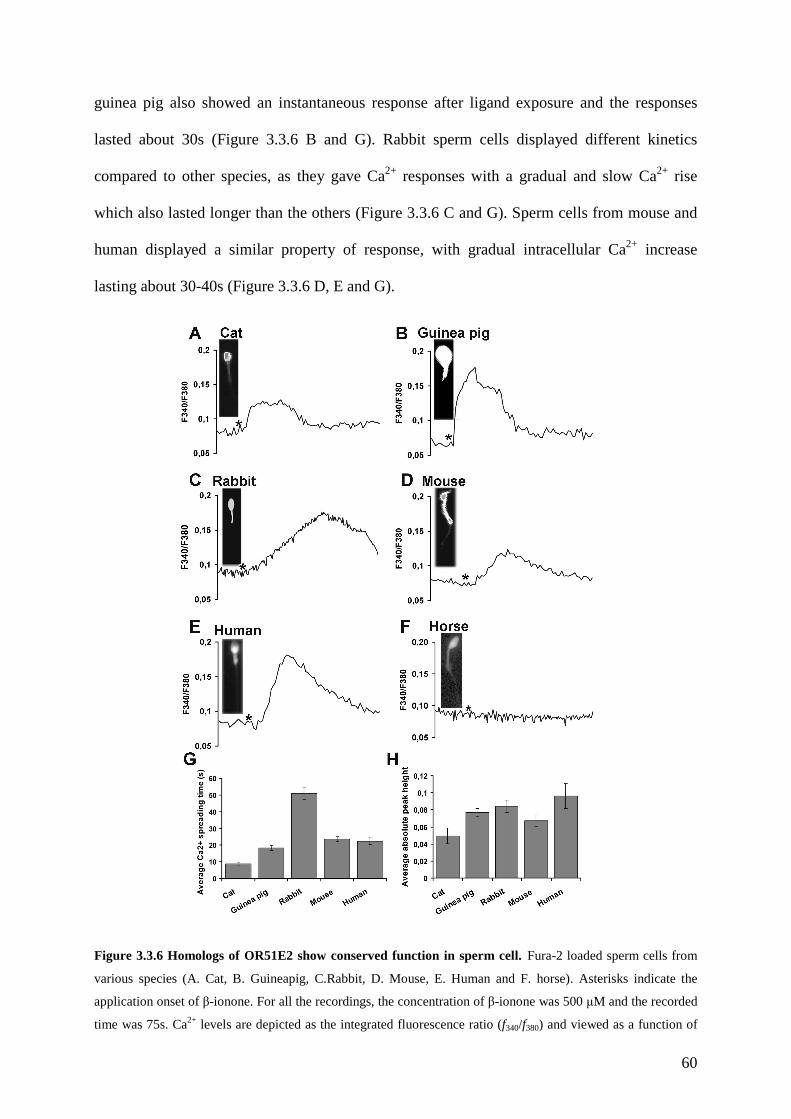
60
guinea pig also showed an instantaneous response after ligand exposure and the responses
lasted about 30s (Figure 3.3.6 B and G). Rabbit sperm cells displayed different kinetics
compared to other species, as they gave Ca2+ responses with a gradual and slow Ca2+ rise
which also lasted longer than the others (Figure 3.3.6 C and G). Sperm cells from mouse and
human displayed a similar property of response, with gradual intracellular Ca2+ increase
lasting about 30-40s (Figure 3.3.6 D, E and G).
Figure 3.3.6 Homologs of OR51E2 show conserved function in sperm cell. Fura-2 loaded sperm cells from
various species (A. Cat, B. Guineapig, C.Rabbit, D. Mouse, E. Human and F. horse). Asterisks indicate the
application onset of β-ionone. For all the recordings, the concentration of β-ionone was 500 µM and the recorded
time was 75s. Ca2+ levels are depicted as the integrated fluorescence ratio (f340/f380) and viewed as a function of

61
time. G. The average Ca2+ spreading speed was interpreted as the average time scale from the onset of response
to the maximal response in β-ionone stimulated sperm cells of each species. H. The average absolute peak height
of β-ionone induced Ca2+ responses in different species. Error bars represent SEM.
As expected, sperm cells collected from horse did not respond to β-ionone, but to the positive
control K+8.6 (Figure 3.3.6 F). This further supported that the OR51E2 homolog is not
functional in horse sperm cells. Among all the tested species, human, guineapig and rabbit
sperm cells showed the highest activity (largest response amplitude) (Figure 3.3.6.H), and
they also showed the closest protein distances to each other. All the above data demonstrate
that OR51E2 homologs in different species share not only the conserved sequences, but also
may have a conserved function as they respond to the same ligand.
3.4 Discussion
Our current study revealed the potential physiological role of a novel testicular receptor,
OR51E2. The signaling cascade of this receptor is coupled to a PLC mediated signaling
pathway. Extracellular Ca2+ is not necessary for the OR51E2 elicited Ca2+ mobilization in
human sperm. Additionally, we found that OR51E2 is evolutionarily conserved, and the
homologs display high conservation in the putative ligand binding positions and responded to
the same ligand. Consequently, OR51E2 homologs may have a highly conserved function in
sperm of different species.
Accumulating evidences show that at least some ORs are localized in the sperm cells together
with the signaling cascade members known to be expressed in the OE. Human OR17-4
(Neuhaus et al., 2006a), dog OR DTMT (Vanderhaeghen et al., 1993) and rat ORs (Walensky
et al., 1998) have been localized to the midpiece of sperm cells. Moreover, β-arrestin
(Neuhaus et al., 2006a; Walensky et al., 1995), adenylyl cyclases (Spehr et al., 2004b), and
Golf (Spehr et al., 2004b) were localized to middle piece and/or principle piece of sperm cells.
In the current study, we found that the OR51E2 is localized to the midpice, principle piece
and vesicular structures in the head region of the sperm by using a specific antibody against

62
the N-terminus of OR51E2. Previous studies demonstrated that Gq/11(Walensky and Snyder,
1995), PLCβ1(Walensky and Snyder, 1995), and a Ca2+ store composed of IP3 receptor and an
isoform of sarcoplamic/endoplasmic reticulum Ca2+-ATPases (SERCAs) were also localized
to the acrosome region of the head and the midpiece (Kuroda et al., 1999; Lawson et al., 2007;
Minelli et al., 2000; Walensky and Snyder, 1995).
It has further been suggested that the localization of receptors and their signaling components
may also be correlated with the pattern of spatiotemporal Ca2+ response in sperm cells (Spehr
et al., 2004b). Using a similar method, we showed that Ca2+ mobilization induced by the
application of β-ionone spread gradually from the principle piece of the tail to the head with
significant latency, which might be due to the distribution of OR51E2 and its signaling
components in the sperm cells. However, the inhibitor of SERCA pump, thapsigargin that
releases Ca2+ from intracellular Ca2+ stores (localized to the acrosome cap), in contrast, was
shown to induce synchronous Ca2+ signals in different sub-cellular compartments in sperm
cells. Although it is not yet known how the spatiotemporal response pattern in the sperm is
generated, the evidences provided in our current study may imply that it is correlated with
different intracellular Ca2+ regulating mechanisms.
We demonstrated that inhibition of PLC by U73122 abolished β-ionone induced Ca2+
signaling in sperm cells, suggesting activation of OR51E2 involves a PLC mediated pathway.
It is known that in olfactory sensory neurons, ORs mediate the responses by depolarizing the
OSN membrane, elevating intracellular Ca2+ concentration and generating action potentials in
the axon. Adenylyl cyclase is generally accepted to be the key player in olfactory signal
transduction in vertebrates (Belluscio et al., 1998; Brunet et al., 1996; Wong et al., 2000).
However, odorants were known to elicit more than one transduction cascade. Studies from
different groups showed that many odorants did not induce Ca2+ responses in transgenic mice
with deficiency of Gαolf and a subunit of CNG channel, which are essential components of the

63
AC/cAMP mediated signaling cascade (Belluscio et al., 1998; Brunet et al., 1996; Jones and
Reed, 1989; Zhao and Reed, 2001). However, these animals were still able to accumulate IP3,
which indicates that PLC may not participate in transducing signals for odor detection and
reorganization like AC but rather participate in some other biological events (Noe and Breer,
1998). Nevertheless, in a native system, the identity of ORs whose activation directly couples
to the PLC/IP3 pathway was not yet elucidated, until we showed that native OR51E2 (PSGR)
coupling to PLC/IP3 mediated Ca2+ signaling in prostate cells (Chapter 2). AC/cAMP pathway
is also believed to play an important role in sperm physiology (Fraser et al., 2005). Activated
AC results in cAMP production which activates the PKA activation and tyrosine
phosphorylation which are essential for the following acrosome reaction (Breitbart, 2002).
ORs have been shown to be able to activate membrane bound isoform of AC, accordingly
leading to sperm chemotaxis and hyperactivity (Spehr et al., 2003; Spehr et al., 2004b).
Nevertheless, AC/cAMP pathway is not the only active pathway in sperm cells triggered by
the activation of GPCRs. So far twelve phospholipase C (PLC) isozymes have been identified,
and they are divided into six classes, β-, γ-, δ-, ε-, ς-, and η-type, on the basis of structure and
activation mechanisms. Some of them have already been shown to be expressed in sperm cells
and to be functionally correlated to male fertility: mutations of PLC β1 leads to reduction of
acrosome reaction, fertilization and embryo development (Choi et al., 2001); cAMP triggers
PLC γ1-IP3 receptor-calcium signaling at the connecting piece of boar spermatozoa
(Harayama et al., 2005); PLC γ4 is required for Ca2+ mobilization and zona pellucida induced
acrosome reaction (Fukami et al., 2001; Fukami et al., 2003). PLC ς as a sperm specific PLC
isoform triggers the development of a fertilized egg into an embryo (Saunders et al., 2002).
Moreover, the OE specific isoform of PLC, PLCβ2 has also been revealed to be present in
human spermatozoa by a proteomics approach (Barbour, 2006).
Furthermore, we also showed that the extracellular Ca2+ is not essential for β-ionone induced
intracellular Ca2+ increase, as sperm cells still showed responses to β-ionone in Ca2+ free

64
extracellular solution. This observation was consistent with the finding that store depletion
with thapsigargin almost completely abolished β-ionone induced Ca2+ response in sperm cells.
The Ca2+ released from intracellular stores of sperm has been suggested to have important
biological function in spermatozoa. The intracellular Ca2+ release, but not an external Ca2+
entry, plays an important role in providing Ca2+ to initiate sperm hyperactivation (Marquez et
al., 2007). Ca2+ release from an intracellular Ca2+ store localized to a compartment of the RNE
directly affects axonemes and is responsible for initiating hyperactivated motility (Ho and
Suarez, 2003).
Capacitation of sperm cells is essential for fertilization virtually in every mammalian species.
Some GPCRs were found to affect capacitation in different aspects. β-adrenergic receptors
and α2-adrenergic receptor are functional in capacitation and acrosome reaction: β-adrenergic
receptor agonists accelerate capacitation in uncapacitated sperm; α2-adrenergic receptor
agonist inhibits acrosome reaction in capacitated sperm (oya-Osiguwa et al., 2006; oya-
Osiguwa and Fraser, 2007). The ligand of adrenergic receptors, adrenaline was shown to
stimulate cAMP production in uncapacitated spermatozoa but to inhibit it in capacitated cells
(oya-Osiguwa et al., 2006). Another study showed that µ-opioid receptor has a function on
induction of capacitation and acrosome reaction (Albrizio et al., 2005). In the current study,
we demonstrated that OR51E2 played a role in regulating sperm capacitation. By employing
M540 staining in order to detect fully capacitated sperm cells, we found there is a significant
PLC-dependent enhancement of human sperm capacitation mediated by β-ionone, indicating
that β-ionone induced intracellular Ca2+ increase is important for regulating capacitation. The
results were further confirmed by the detection of tyrosine phophorylated proteins in
capacitated sperm, which also demonstrated a PLC-dependent increase pattern. Some
evidences showed that in progesterone mediated acrosome reaction, inhibition of G protein,
phospholipase A2 and phospholipase C consequently leads to the decreased acrosome reacted

65
rate in capacitated human sperm (Pietrobon et al., 2005). However, the mechanisms of how
ORs and the PLC are involved in regulation of capacitation needs to be further elucidated.
OR51E2 is an evolutionarily conserved gene. Using degenerate primers, we detected OR51E2
homologs in testes of 9 different mammalian species: cat, dog, ferret (belonging to Carnivora),
guinea pig, rat, mouse (belonging to Rodentia), rabbit (belonging to Lagomorpha), and human
(belonging to Primates), showing conserved testicular expression. Sequencing analysis
revealed that the homologous receptors share more than 89% DNA identity and more than
92% amino acid identity. RT-PCR amplification only showed a very weak band amplified
from horse testis RNA, the following sequencing analysis indicated that the horse homolog is
a pseudogene by showing multiple stop codons in the open reading frame. Actually some OR
pseudogenes may have stable transcripts, but mRNA of others may undergo nonsense-
mediated decay (Feldmesser et al., 2006; Maquat, 2005). The horse OR51E2 homolog may
belong to the first class of pseudogenes, which could explain the weak amplification from this
dysfunctional homolog.
Figure 3.4.1 The predicted binding site residues in two-dimensional Snake-plot of human OR51E2. Helical
net representation of a typical OR. The predicted binding site residues are shaded by grey color. (Man et al.,
2004)

66
For the intact homologs, we further compared the primary protein sequences. Overall they
share more than 92% homology to human OR51E2, but they have variations at different
positions. It has been predicted that 22 amino acids may play an important role in constructing
the odorant binding site of the ORs (Man et al., 2004). They are disposed on transmembrane
helices 2 to 7 and on the second extracellular loop of the receptor and clustered around a
pocket in a structural model of ORs, mostly facing the inner lumen (Man et al., 2004). We
specially compared these putative positions for conservation among different species, these
sites on the homologs are 100% identical to human OR51E2. This explained the fact that the
homologs respond to the same ligand as the human receptor.
Ca2+ imaging results showed us that sperm cells from different species indeed respond to β-
ionone and the overall signals look similar among all the species in our study. Spermatozoa
give transient responses to β-ionone, and the time scale until response decay was similar.
Rabbit sperm displayed a strong and more gradual Ca2+ increase and a strong band in the RT-
PCR analysis indicated a high abundance of the transcript, which might contribute to explain
the phenomenon. However, this question needs further investigation to figure out the quantity
of the transcripts of these homologs in each species. The horse homolog did not show
response to β-ionone but showed responses to the positive K+8.6 control, which is consistent
with the observation in RT-PCR analysis. These data supported the conclusion that the horse
homolog is a dysfunctional pseudogene. The results together may suggest a conserved
physiological role of OR51E2 homologous receptors in mammalian sperm physiology.
4. Functional investigation of MOR283-2 in mouse spermatogenic cells.
4.1 Research background
Spermatogenesis involves a number of unique processes, including meiosis, haploid gene
expression, formation of the acrosome and the flagellum, removal of histones from the

67
chromatin and their replacement with protamines and nuclear condensation. However, it also
processes features characteristic of typical differentiating tissue, including self-renewal from a
stem cell population, a series of cell divisions closely associated with stepwise developmental
processes and progression from morphologically undifferentiated to highly differentiated cells.
Spermatogenesis occurs in successive mitotic, meiotic and post-meiotic phases (Figure 4.1.1).
Figure 4.1.1 Spermatogenesis follows an orchestrated differentiation process (Kimmins et al., 2004).
Spermatogenesis in mice initiates at about postnatal day 4; the mitotic phase lasts ~10 days;
the meiotic phase lasts ~11 days and the post-meiotic phase lasts ~14 days (Bellve et al.,
1977). In total, sexual maturation takes 36~42 days. During the mitotic phase stem cells
divide 6 times to successively form type A intermediate and type B spermatogonia. The final
division produces preleptotene spermatocytes, which begin the meiotic phase and undergo the
last cell cycle S phase of spermatogenesis. During meiotic phase homologous chromosomes
recombine to exchange genetic materials. This is followed by two meiotic divisions that occur

68
in rapid succession with DNA replication to produce spermatids, the post-meiotic cells.
Spermatids are then remodelled into spermatozoa by the process of acrosome formation,
nuclear condensation, flagellar development and loss of the majority of the cytosol (Fawcett,
1993).
Meiosis is a specialized type of cell division that gives rise to haploid gametes. The first round
of cell division has a long prophase that is subdivided into leptotene, zygotene, pachytene,
diplotene, and diakinesis stages. The following second round of cell division is made up of
metaphase, anaphase and telophase. These stages are distinguishably different in form and
relationships of the pairs of homologous chromosomes and their degree of condensation. The
leptotene stage is only slightly different from the early stage of mitosis. The nuclei of meitotic
cells are bigger than other cells. Pairs of homologous chromosomes are like long slender
threads. During the zygotene stage the homologous chromosomes come together, forming the
haploid number of synaptic pairs. In the pachytene stage, the pairs of chromosomes contract
longitudinally, becoming thicker and more conspicuous. The two members of each synaptic
pair also split longitudinally forming tetrads consisting of four parallel chromatids. In the
following diplotene stage, corresponding segments of pared chromosomes are exchanged in a
process unique to meiosis, called crossing-over. The parental genes are mixed and rearranged
in new combinations. The diakinesis is the continuation of the diplotene. It is usually difficult
to demarcate both states. The chromosomes condense and become more compact. The
metaphase. From now on the processes resemble that of mitosis again. The nuclear membrane
is completely dissolved, the mitotic spindle fully developed. The homologous chromosomes
do still stay together. During the anaphase the homologous chromosomes are separated from
each other. It is followed by the telophase, then by interkinesis (this state corresponds to the
so-called quiescence or interphase state). Shortly afterwards the second mitotic division with

69
the usual states of prophase, metaphase, anaphase and telophase occurs. Here the chromatids
are separated from each other (Fawcett, 1993).
Figure 4.1.2 Representative differentiation stages (Fawcett, 1993) and corresponding γH2Ax staining
patterns (Mahadevaiah et al., 2001). The schematic drawings of each representative stages of spermatogenesis
is shown in upper panel. Spermatogonia (A); Leptotene spermatocytes (L); Pachytene spermatocytes (P); Round
spermatid (7). The corresponding γH2Ax expression is shown in lower panel. Spermatogonia, spermatids and
spermatozoa are devoid of γH2Ax, whereas, in early prophase spermatocytes (leptotene to zygotene), γ-H2AX
displays a punctate appearance or has become restricted to a tadpole-shaped structure. When pachytene
processes, γ-H2AX is restricted to the sex body throughout the pachytene stage.
The reduction in chromosome number is achieved by undergoing one round of DNA
duplication followed by two rounds of chromosome segregation. During the first round of
segregation, meiosis I, chromosomes must find their cognate homolog, pair and undergo
reciprocal recombination, leading to the establishment of physical connections between them
(Petronczki et al., 2003). In order to deal with incomplete synapsis of the X-Y pair, mammals
have adopted a strategy that involves seclusion of the sex chromosomes to a subnuclear
compartment known as the sex body. This subnuclear domain distinguishes the sex chromatin
from the rest of the chromatin during pachynema (when the autosomes are completely
synapsed) and diplonema (Handel, 2004). Histone H2AX is a variant of the core histone H2A
and is ubiquitously distributed throughout the genome. Mitotic cells phosphorylate H2AX at
serine 139 in response to DNA damage, inducing the formation of nuclear foci of
phosphorylated H2AX (γH2AX) at the sites of damage. Therefore, γH2AX can be used as a

70
marker for double strand breaks (Fernandez-Capetillo et al., 2004). Analysis of H2AX
phosphorylation during mouse meiosis revealed that γH2AX occurs in all intermediate and B
spermatogonia and in preleptotene to zygotene spermatocytes (Mahadevaiah et al., 2001).
Type A spermatogonia and round spermatids do not exhibit γH2AX foci but show
homogeneous nuclear γH2AX staining, whereas in pachytene spermatocytes γH2AX is only
present in the sex body. Therefore, γH2AX can be used as a specific marker for certain stage
of meiosis, and in particular is used as a marker for pachytene spermatocytes, as the γH2AX
staining is restricted to the sex body (Mahadevaiah et al., 2001).
The post-meiotic stage termed as spermiogenesis is a very intricate and precise
morphogenetic process during which many structural and functional changes are necessary in
order to produce mature spermatozoa. The round spermatid is identified as the first cell type
of beginning spermiogenesis. In the spermatid cytoplasm, microtubules increase in number
and form specialized structures called manchette. Until this stage of spermatid differentiation,
the tail of the future spermatozoon is represented only by the axoneme (a highly-conserved
cytoskeletal structure) enclosed in a flagellar membrane. At the place where the flagellar
membrane is continuous with the membrane of the cell body, dense material adhering to its
cytoplasmic face condenses to form a ring. This ring is the precursor of the annulus that will
later be situated at the junction of the mid piece and principal piece of the sperm tail (Fawcett,
1993). The presence of an axoneme can be confirmed by the linear a-tubulin immunoreactive
pattern along each filamentous projection, ending in a coiled tip (Marh et al., 2003).
The somatic cells in testicular tubule are Sertoli cells which produce factors required by
developing germ cells. The number of Sertoli cells determines testicular size, germ cell
numbers per testis and spermatozoa output (Mruk and Cheng, 2004). Moreover, Sertoli cells
also provide a specialized, protected environment within the seminiferous tubules of the testis
for germ cell development. Adjacent Sertoli cells form tight junctions with each other such

71
that nothing larger than 1000 Daltons can pass from the outside to the inside of the tubule,
forming the so-called blood-testis barrier. At the beginning of meiosis, germ cells located
outside of the barrier pass through the tight junctions. Once beyond the barrier, germ cells are
dependent on Sertoli cells to provide nutrients and growth factors. Sertoli cells provide factors
required to fuel germ cell metabolism, growth regulatory factors, insulin-like growth factors,
fibroblast growth factors, epidermal growth factor and hormones that regulate the
development of the male reproductive structures (Mruk and Cheng, 2004), or feedback to
regulate the hormonal signals affecting Sertoli cells (Ramaswamy et al., 2000). Nevertheless,
as a very complicated progress, spermatogenesis involves a precisely regulated sequence of
differentiation steps, with adult fertility achieved through the integration of a succession of
hormonal and local factors driving both somatic and germ cell maturation. The precise
mechanisms of the regulation of spermatogenesis or the proteins or hormones involved in
sperm cell maturation remains unclear.
Since discovery of the olfactory receptor gene superfamily in 1991(Buck and Axel, 1991),
accumulating evidences showed that ORs are also ectopically expressed in testis, especially in
spermatozoa (Asai et al., 1996; Parmentier et al., 1992; Vanderhaeghen et al., 1993;
Vanderhaeghen et al., 1997b). The functions of some of these ORs in regulating male fertility
have also been discovered in human and mouse (Fukuda et al., 2004; Spehr et al., 2003).
Additionally, a recent study employed degenerate PCR and in situ hybridization showed that a
subset of OR transcripts are localized in three developmental stages of germ cells: late
pachtyene spermatocytes, early round spermatids and late round spermatids (Fukuda and
Touhara, 2006). The signal transduction components of olfaction and GPCR signal
transduction are also found to be expressed in spermatogenic cells. Gαi1, Gαi2 Gαi3 Gαo were
detected in spermatocytes and spermatids (Karnik et al., 1992), ACIII and a type of 4A
phophodiesterase were found to be expressed in mouse round spermatids (Gautier-Court et al.,

72
1998; Naro et al., 1996). Moreover, G-protein coupled receptor kinase 2 (GRK2) and β-
arrestin 2 were found in elongated spermatids (Walensky et al., 1995). Like OSN,
spermatogenic cells probably utilize ORs and GPCRs as signal transducing elements.
Nevertheless, the potential function of this testicular subset of ORs in spermatogenesis was
not yet discovered.
In the present study, we studied a novel mouse OR, MOR283-2, which is expressed in the
olfactory epithelium as well as in testis, particularly in spermatogonia and primary
spermatocytes. We cloned the receptor and functionally express it in heterologous HEK293
cell expression system. By conducting calcium imaging on MOR283-2 expressed HEK cells,
we were able to show the molecular receptive field of MOR283-2. Moreover, the
endogenous ligand is present in the secretion of Sertoli cells. To elucidate the function of the
MOR283-2, we examined the developmental expression pattern of MOR283-2 in mouse
testis. Furthermore, we established a sertoli-spermatogonia coculture system and were able to
monitor a physiological role of MOR283-2 in regulating spermatogenesis.
4.2 Material and Methods
4.2.1 Cloning and vector construction.
MOR283-2 receptor DNA was amplified from mouse genomic DNA by PCR using specific
primers that amplify the complete open reading frame and contain restriction sites for further
subclonings. The receptor construct containing EcoRV and HpaI restriction sites was cloned
into pcDNA3 vector (Invitrogen).
The following primer pairs have been used in this experiment
MM_11.21.01_up: 5’ ACC ATC GAT CGA TAT CGC TGT GCA GGG ATC CCA GAG
A-3’
MM_11.21.01_down: 5’ TCA GTT AAC ATG GAT CCA CCA GCA CGT CAG CAC
CTC-3’

73
4.2.2 Cell Culture.
HEK293 cells were cultivated in MEM medium (Invitrogen) supplemented with 10% FBS
and antibiotics. Semiconfluent HEK cells were transfected in 35 mm dishes with the
MOR283-2 plasmid by using the calcium-phosphate precipitation method. Calcium imaging
measurements were carried out 48–72 hours after transfection.
Mouse type-A spermatogonia and Sertoli cells were prepared and maintained in culture as
previously described (Iwanami et al., 2006). Briefly, 7-day-old mouse testis was dissected,
the tunica albuginea was removed, and the seminiferous tubules were digested in DMEM
containing 1 mg/ml collagenase (Sigma) and 0.2 mg/ml DNAse (Roche) combination at
37°C for 5 min, and subsequently in 0.05% trypsin/0.02% EDTA at 37°C for 5 min with
gentle agitation. The cells were then seeded onto culture dishes with a total medium volume
of 4ml per dish, and maintained at 37°C for 3 days and at 33°C for the subsequent days in
5% CO2. Half of the medium was exchanged every day. The culture medium was comprised
of DMEM medium supplemented with 10% serum, 0.1% non-essential amino acids, 10
ng/ml mouse epidermal growth factor, 10 ng/ml insulin-like growth factor, 1 ng/ml follicle
stimulating hormone, 133 µIU human growth hormone, 5 µg/ml transferrin, 5 µM retinol, 0.1
µM testosterone, 0.01% nucleosides and antibiotics (Iwanami et al., 2006). In odorant treated
cells, 5 µM amylbutyrate and bourgeonal were added to the culture medium, respectively.
Primary culture of Sertoli cells was prepared from 20-day old CD1 mice by the same
sequential enzymatic treatments as described above. The resulting cells were plated at a
density of 106/cm2 on Petri dishes in DMEM medium. Cells were maintained in a humidified
atmosphere of 95% air and 5% O2 at 32°C. To obtain a culture of Sertoli cells that was more
than 95%, cultures were hypotonically shocked with 20 mM Tris (pH7.4) 48 hours after
plating to lyse contaminating germ cells, then washed twice with medium as described
previously (Syed et al., 1999). After cultures were maintained for 72 hours, Ringer solution
was substituted for medium in the culture dish and collected after 24 hours, aliquoted, frozen

74
and passed through a 22 µM nylon filter prior to application onto the cells for Ca2+ imaging
experiments. The viability of the transfected HEK cells incubated with Ringer solution was
checked by 0.4% trypan blue solution and the viability was more than 95% and similar to the
cells maintained in culture medium.
4.2.3 RT-PCR.
The RNA of mice testes was isolated with Trizol Reagent (Invitrogen) and cDNA was
synthesized by using MMLV reverse transcriptase (Invitrogen) and oligo (dT12-18) primer.
Amplification was performed with 1ng cDNA and specific primer pairs for MOR283-2, TP1
(transition protein 1), TP2 (transition protein 2), PRM-2 (promatine 2), respectively. The
amplification was done for 35 cycles (94° 1 min, 58° 1 min, 72° 45 sec).
The following primer pairs have been used in this part of experiment:
mm-P19 fw: 5’-CTC AGC CCT CGG TCA AAA GAA TC-3’ rv: 5’-ACG GGG TCG GGG AGA AAG TC-3’ mm-TH2B fw: 5’- AAG GTT ACT TGA GCC ATG CC -3’ rv: 5’- CGA TGC GCT CGA AGA TGT CTG-3’ mm-TP1 fw: 5’-GCC GCA AGC TAA AGA CTC ATG G-3’ rv: 5’-CTC CTG CCC CGT GTT GTT TG-3’ mm-TP2 fw: 5’-CTC GTG GCC TCA AAG TCA CAC CAG-3’ rv: 5’-GAC GGC CTT TCT CTT GCT CAC TTT CC-3’ mm-16s-RNA: fw: 5’-TGG GCT CAT CAA GGT GAA CGG-3’ rv: 5’- GGT CCG ATC GTA TTG GAT GAG G -3’ PRM-2: fw: 5’-GAG CGC GTA GAG GAC TAT GG-3’ rv: 5’-GCA AGT GAC TTC CTT GGC TC-3’
MM11.21.01(across intron) : Upper: 5’-CAC CCG CTG AAC TAC AT-3’ Lower: 5’- AAA GGA ACT GGG CAA TAC ATA CAT-3’

75
4.2.4 Real-time quantitative PCR.
Real time quantitative PCR was performed in DNA Engine Opticon2 System using SyBr
Green detection and Eurogentec q-PCR master kit, according to manufacturer’s
recommendations. The following amplification protocol was used: 40 cycles of 15s at 95°C,
20s at 60°C and 40s at 72°C. Cycle numbers obtained at the log-linear phase of the reaction
were plotted against a standard curve prepared with serially diluted cDNA samples.
Expression of the target gene was normalized to 16S ribosome RNA levels. The delta Ct
(cycle threshold) method was used to calculate relative expression levels, as previously
described (Livak and Schmittgen, 2001). Results are reported as fold change in gene
expression relative to control conditions. (In Sertoli-spermatogonia culture, control is
untreated cultures. In developmental testis tissue, P1 testis RNA is taken as control.)
4.2.5 In-situ hybridization
Digoxigenin (DIG)-labelled sense and antisense probes were prepared from full length
MOR283-2 subcloned pcDNA3 plasmid. After linearization of the plasmid, the sense and
antisense probes were transcribed using T7 and SP6 RNA polymerase together with DIG
RNA labelling mix (Roche). The testes from 10-15-week old mice were immediately
embedded in tissue freezing medium (Leica) on dry ice and cryostat sections (10 µM) were
cut on Leica cryostat and mounted on superfrost glass slide. Slides were postfixed in 4% PFA
in PBS at 4°C for 20 min), acetylated in 250 ml 0.1 M triethonolaminhydrochloride solution
with 600 µl acetanhydride and then incubated overnight at 55°C, with 200 µL hybridization
buffer (50% Formamid, 5×SSC, 5×Denhardts, 2.5 mM EDTA, 50 µg/ml Heparin, 250 µg/ml
tRNA, 500 µg/ml sperm DNA and 0.1% Tween 20) containing DIG-labelled probe (0.8
µl/200 µl hybridization buffer). The sections were washed at 65°C with 0.2×SSC (30mM
NaCl and 3mM sodium citrate) and then blocked with 10% goat serum in PBS buffer. This
was followed by incubation with 1:1000 alkaline-phosphotase (AP) conjugated anti-DIG Fab

76
fragment (Roche). The sections were then incubated with NBT/BCIP (20 µl/ml) to develop
the signals (Roche).
4.2.6 Ca2+-imaging.
Imaging of HEK293 cells was performed as described previously (Neuhaus et al., 2006b).
Briefly, cells were incubated for 30 min at 37°C in the Ringer solution (140 mM NaCl, 5 mM
KCl, 2 mM CaCl2, 2 mM MgCl2, 10 mM Hepes, pH7.4) containing 3 µM Fura-2 (Molecular
Probes) and. The final concentration of amylbutyrate was 500 µM. Ca2+ imaging experiments
were performed on the stage of a Zeiss inverted microscope equipped for ratiometric imaging.
Odorants used were a gift from Dr. T. Gerke, Henkel KGaA, Düsseldorf, Germany. Odorants
assayed for potential activation of MOR283-2 were tested in at least three transfection
experiments in HEK cells. All compounds regarded as ligands led to clear Ca2+ responses in
several different experiments (n > 7), whereas they did not elicit any Ca2+ signals in
untransfected HEK cells. 200 µM ATP was applied as the positive control at the end of each
experiment in HEK cells.
For imaging of spermatogenic cells, seminiferous tubules were isolated from mouse testis in
PBS buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, pH 7.4), and
cells were dissociated as previously reported (Fukuda et al., 2004). In brief, pieces of
seminiferous tubules were treated for 10 min with 0.05% trypsin and 0.2 mg/ml DNase I
(Roche). The trypsin treated tubules were washed with Ringer buffer supplemented with 15
mM NaHCO3 and then spermatogenic cells were isolated on an uncoated glass-bottomed dish.
Cells were loaded in Ringer buffer with 3 µM Fura2-AM at 37°C for 20 min. The imaging
procedure was conducted as described above. K+8.6 (135 mM KCl, 5 mM NaCl, 2 mM CaCl2,
1 mM MgCl2, 10 mM glucose, 10 mM lactic acid, 1 mM pyruvic acid, 30 mM HEPES, pH8.6)
was used as a positive control at the end of each experiment in spermatogenic cells.

77
4.2.7 Immunohistochemistry
Sertoli-spermatogonia cells were maintained as described above for 10 or 14 days on poly-l-
lysine coated coverslips. The cells were blocked in 10% goat serum, 1% BSA and 1% gelatin
overnight. γ-H2Ax antibody (1:100) and tubulin antibody (1:100), were used, followed by
secondary fluorescent anti-rabbit IgG (1:1000) to determine the temporal appearance of XY
bivalent in primary spermatocytes (pachytene spermatocytes) and axoneme in round
spermatids, respectively. Controls consisted of omission of the primary antibody. Statistics
were done by counting 500 cells and repeated for 3 different cultures for each staining.
Significance was tested by paired T-test.
4.2.8 BrdU proliferation assay
For the BrdU proliferation assay, before seeding onto the culture dish, testicular cells were
incubated with BrdU for 1 hour according to the kit instruction (Amersham). An anti-BrdU
monoclonal antibody (diluted 1:10) (Amersham) and an antimouse fluorescence secondary
antibody (diluted 1:1000) were used to reveal labelled cells in culture.
4.2.9 In situ 3’-end labeling (ISEL)
Cultured tubule segments were fixed on slides using a modified squash preparation as
previously described (Hendriksen et al., 2006; Kotaja et al., 2004). In brief, seminiferous
tubules were dissected in a petri dish containing PBS buffer under microscope and segments
(~1 mm) were transferred to glass microscope slide in 15 µl buffer. They were carefully
squashed under a coverslip to form a cell monolayer. Slides were frozen in liquid nitrogen
and the coverslip was removed with a scalpel. Slides were then fixed in 4% PFA at 4°C for
20min and were incubated in 2×SSC at 80°C for 20 min followed by washing twice with
water and once with Proteinase K buffer (10 µg/ml in 50 mM Tris and 1.5 mM calcium
acetate) for 5 min each. The slides were then treated with proteinase K (10 µg/ml, Roche) at
37°C for 30 min. A 20µl aliquot of 3’-end labelling reaction mixture was applied to one 2-

78
mm-long squashed segment. The slides were kept in a humidified box and incubated at 37°C
for 1 hour and then washed three times with TBST for 10 min each. An anti-Dig-AP
monoclonal antibody (1:500) in 10% goat serum containing PBS was applied and the slides
were incubated at room temperature for 1 hour followed by three 5-minute washes with
TBST. Finally, the labelled cells were visualized by incubating the slides with NBT/BCIP
(Roche) for 0.5-2 min.
4.3 Results
4.3.1 Characterization of MOR283-2 expression pattern in testis
To elucidate the function of MOR283-2, we first examined the expression pattern of
MOR283-2 at the mRNA level. By conducting the RT-PCR, MOR283-2 transcripts were
found to be present in mouse olfactory epithelium, as well as in testis (Figure 4.3.1A). As
GPCRs have intronless open reading frames, we designed an intron-spanning primer pair
across an intron between the 5’-UTR and the open reading frame of MOR283-2 to exclude
that the amplified DNA comes from the trace amount of contaminating genomic DNA. This
prime pair gave a potent ~400 bp amplification fragment from the testicular cells of mouse
different developmental stages (Figure 4.3.1 B). In order to determine the expression level of
MOR283-2 during the testis development, we further utilized this primer pair in real-time
RT-PCR analysis. This analysis revealed that the level of MOR283-2 transcripts was
developmentally regulated during testis maturation. From the quantitative PCR analysis, we
discovered that the MOR283-2 mRNA was most abundant at postnatal day 10 (P10).
Taking the amount of MOR283-2 mRNA of P1 mouse testis as a basal level (as 1 fold), we
can see that at P10, the receptor mRNA level is 1.7±0.3 fold of the basal level. Then the
amount of transcripts gradually declined to 1.2±0.1 fold at P20, and further declined to 0.8
±0.1 fold of basal level at P30. From P30 to P60 (from the end of puberty to adulthood), the
receptor expression level stays steadier by showing only little decrease (Figure 4.3.1C). We

79
next examined the expression pattern of MOR283-2 transcripts in the testis by in situ
hybridization to determine the exact cell types expressing MOR283-2. Hybridization signals
were observed in the cells close to the basal membrane (Figure 4.3.1D). About 20% of
tubules show hybridization stainings. By further analysing the nuclear morphologies of
theses cells, it was determined that MOR283-2 transcripts were mainly localized in
spermatogonia and primary spermatocytes (indicated by arrows).
Figure 4.3.1 Expression and localization of MOR283-2 encoding transcripts in adult mouse testis. (A) RT-
PCR detection of MOR283-2 expression in mouse testis and olfactory epithelium, using the primer pair specific
for the open reading frame (ORF) of MOR283-2. (B) RT-PCR detection of MOR283-2 expression profile
during testis maturation, using the primer pair across the intron between 5’-UTL and ORF. (C) Changes in
expression level of MOR283-2 during mouse testis maturation compared with postnatal day 1.(D) Detection of
MOR283-2 transcripts with an antisense probe. MOR283-2 encoding mRNA was detected in spermatogonia
and early stage of spermatogenic cells, indicated by the arrows.
4.3.2 Identification of molecular receptive field for MOR283-2
ORs were reported to be able to respond to a complex of odorant stimuli (Spehr et al., 2003).
Therefore, Henkel 100, containing 100 odorants, was used for ligand screening. Upon the
application of Henkel 100, we observed a transient Ca2+ influx in HEK293 cells transfected

80
with MOR283-2 (Figure 4.3.2A). Moreover, no intracellular Ca2+ increase was observed in
non-transfected/mock transfected HEK cells. By further dividing Henkel 100 into smaller
fractions, we discovered that one single compound, namely amylbutyrate, was specifically
recognized by MOR283-2 (Figure 4.3.2A). The molecular receptive field of MOR283-2 was
established based on the chemical structure of amylbutyrate. By screening the active and
inactive substances sharing a similar sub-structural motif to amylbutyrate, we concluded that
the ethyl acid ester group is an essential determinant for being a cognate MOR283-2 ligand
(Figure 4.3.2B). Moreover, amylbutyrate (500 µM) turned out to be the most active agonist
for MOR283-2 as the relative Ca2+ increase in transfected HEK cells induced by
amylbutyrate to ATP is 67.1±19.1%, which is much higher than the other active compounds
(20%-40%). (Figure 4.3.2D)
Since MOR283-2 is found to be expressed in immature spermatogenic cells, we questioned
whether an endogenous ligand is present in the secretion of Sertoli cells. It is known that
spermatogenesis is highly dependent upon the somatic Sertoli cells to produce factors
required by spermatogenic cells (Griswold, 1998; Walker and Cheng, 2005). We then
collected the secretion of Sertoli cells from the culture by incubating the cultured Sertoli cells
in Ringer’s solution for 24 hours and applied the supernatant on the HEK 293 cells
transfected with MOR283-2. Remarkably, the secretion of Sertoli cells induced transient
intracellular Ca2+ increase in MOR283-2 transfected cells but not in mock transfected or non-
transfected cells (Figure 4.3.2C). After normalizing the Ca2+ imaging signal from Sertoli cell
secretion stimulated cells against the positive control (stimulated with ATP), we elucidated
that the relative signal is 44.7±11.9% (Figure 4.3.2D), which is smaller than amylbutyrate
induced signals. However, this could be due to the fact that the concentration of applied
amylbutyrate might be much higher than the concentration of endogenous MOR283-2 ligand
in the Sertoli cell supernatant.

81
Figure 4.3.2. Characterization of olfactory receptive field of MOR283-2. (A) The complex odorant mixture
Henkel 100 induced transient Ca2+ mobilization in transfected HEK293 cells. (B) Representative Ca2+ imaging
recordings of responding MOR283-2 transfected HEK293 cells to 500 µM amylbutyrate. 100µM ATP was used
as a positive control. (C) Structural correlated molecules were individually tested for the capability of activating
MOR283-2, collectively identified by the presence of the group of ethyl acid ester. The left panel of the table
indicates the active compounds (tested at 500 µM) and the right panel indicates the inactive compounds (tested
at 500 µM). (D) HEK293 cells transfected with the recombinant MOR283-2 responded to the Sertoli cell
secretion which was collected by incubating Sertoli cells in Ringer’s solution for 24 hours. 100 µM ATP was
used as a positive control. (E) Relative peak heights compared to ATP. The average Ca2+ signal elicited by
amylbutyrate is normalized to 67.1±19.1% to ATP. The one elicited by the secretion is normalized to
44.7±11.9% to ATP.
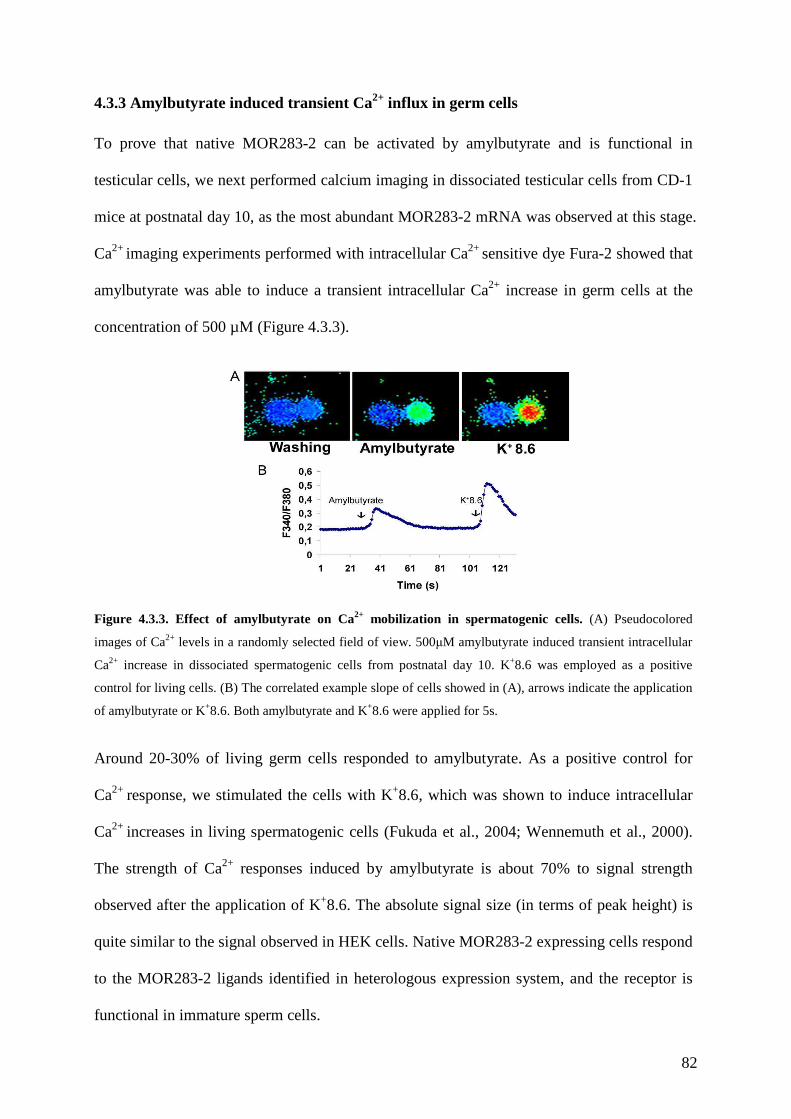
82
4.3.3 Amylbutyrate induced transient Ca2+ influx in germ cells
To prove that native MOR283-2 can be activated by amylbutyrate and is functional in
testicular cells, we next performed calcium imaging in dissociated testicular cells from CD-1
mice at postnatal day 10, as the most abundant MOR283-2 mRNA was observed at this stage.
Ca2+ imaging experiments performed with intracellular Ca2+ sensitive dye Fura-2 showed that
amylbutyrate was able to induce a transient intracellular Ca2+ increase in germ cells at the
concentration of 500 µM (Figure 4.3.3).
Figure 4.3.3. Effect of amylbutyrate on Ca2+ mobilization in spermatogenic cells. (A) Pseudocolored
images of Ca2+ levels in a randomly selected field of view. 500µM amylbutyrate induced transient intracellular
Ca2+ increase in dissociated spermatogenic cells from postnatal day 10. K+8.6 was employed as a positive
control for living cells. (B) The correlated example slope of cells showed in (A), arrows indicate the application
of amylbutyrate or K+8.6. Both amylbutyrate and K+8.6 were applied for 5s.
Around 20-30% of living germ cells responded to amylbutyrate. As a positive control for
Ca2+ response, we stimulated the cells with K+8.6, which was shown to induce intracellular
Ca2+ increases in living spermatogenic cells (Fukuda et al., 2004; Wennemuth et al., 2000).
The strength of Ca2+ responses induced by amylbutyrate is about 70% to signal strength
observed after the application of K+8.6. The absolute signal size (in terms of peak height) is
quite similar to the signal observed in HEK cells. Native MOR283-2 expressing cells respond
to the MOR283-2 ligands identified in heterologous expression system, and the receptor is
functional in immature sperm cells.

83
4.3.4 Spermatogonia-Sertoli cell coculture.
To investigate the physiological role of MOR283-2 in germ cells, testicular cells from
postnatal day 7 mice were isolated and cultured in the presence or absence of 5 µM
amylbutyrate by the Sertoli-spermatogonia co-culture method. 5 µM amylbutyrate was able
to elicits intracellular Ca2+ increase in MOR283-2 expressed HEK cells and in spermatogenic
cell. Bourgeonal was employed in the culture as a control odorant, as it was shown to be
incapable of inducing Ca2+ increase in immature mouse sperm cells due to a lack of
corresponding receptors (Fukuda et al., 2004). The phase-contrast photo of cultured cells
(Figure 4.3.4 A) showed that spermatogonial cells in co-culture system were able to
differentiate into flagellated round spermatids (with arrow indication).
Figure 4.3.4.Sertoli-spermatogonia co-cultures. (A) Phase contrast photo of the cells in Sertoli-
spermatogonia co-culture for 14 days. The arrow indicates flagellated round spermatids differentiated from
spermatogonia. (B) RT-PCR detection of specific molecular markers for haploidy round spermatids. TP1,
transition protein 1; TP2, transition protein 2; PRM2, promatine-2.
To validate the successful differentiation at molecular level, we performed RT-PCR on the
cells in culture for 14 days (first spermatids emerged in the culture) to detect the specific
round spermatid markers TP1, TP2, and PRM2. These markers are exclusively transcribed in
round spermatids (Bunick et al., 1990; Hecht, 1990; Marret et al., 1998). The RT-PCR results
(Figure 4.3.4B) elucidated that post-culture cells in all different treatments contained the

84
transcripts of these markers as well as the transcripts of MOR283-2, indicating the co-
cultured spermatogonia were able to differentiate in vitro.
4.3.5 The effect of amylbutyrate on in vitro spermatogenesis.
To generate haploid spermatids, diploid spermatogonia need to enter the meiotic phase and
differentiate into tetraploid meiotic cells (pachytene spermatocytes). One significant feature
for meiotic cells is sex chromatin breaking, which can be monitored by the distribution of the
phospho-isoform of histone protein H2Ax (γH2Ax). During spermatogenesis, H2Ax is
phosphorylated throughout the chromatin in leptotene spermatocytes, and γH2Ax gradually
decreases during zygotene stage. By pachytene stage, γH2Ax is undetectable on autosomal
chromatin and restricted to the sex body (at a subnuclear location) during pachytene and
diplotene stages (meiotic progress) (Mahadevaiah et al., 2001). To evaluate the effects of
amylbutyrate on the meiotic steps of the spermatogenic process, we examined the
immunoreactive patterns of γH2Ax in cells cultured for 10 days (Figure 4.3.4 D), as we
found that most pachytene spermatocytes appeared at day 10 in the culture. The number of
spermatogenic cells with the subnuclear dot localized phospho-H2Ax staining was counted,
and the ratio to the total germinal cells was calculated (Figure 4.3.4 E). The results showed a
significant increase of the percentage of meiotic cells when co-culture was conducted with
addition of amylbutyrate (5 µM) compared to the cultures maintained under the control
condition or to cultures treated with bourgeonal (5 µM). In control and bourgeonal treated
cultures, 35.2%±10.6% and 27.4±7.4% cells showed the sex-body restricted γH2Ax
staining pattern. In amylbutyrate treated cells, the percentage was 60.3±4.5%, which is
significantly higher than in the control cultures (Figure 4.3.4 E). We further performed
quantitative PCR to analyse the expression amount of pachytene spermatocyte marker p19
which is temporally transcribed in tetraploid meiotic cells (Weiss et al., 1997). By taking the
expression level of p19 in non-treated control cells as basal level (normalized to 1), the
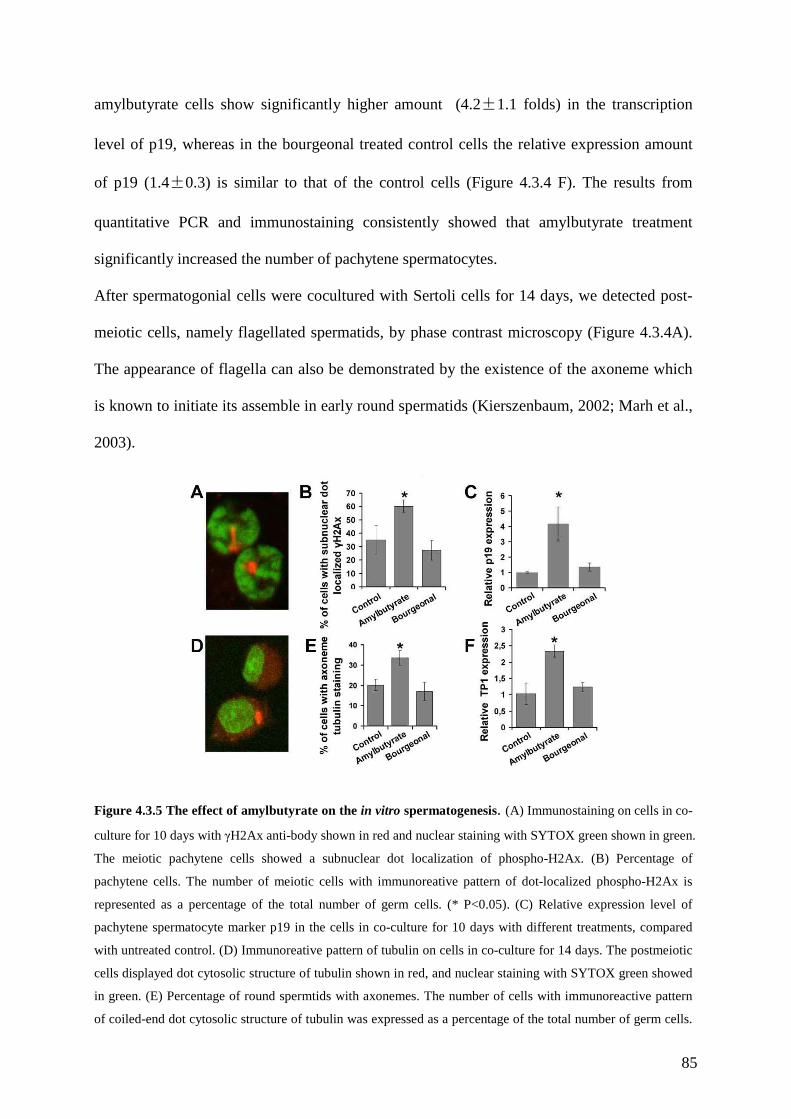
85
amylbutyrate cells show significantly higher amount (4.2±1.1 folds) in the transcription
level of p19, whereas in the bourgeonal treated control cells the relative expression amount
of p19 (1.4±0.3) is similar to that of the control cells (Figure 4.3.4 F). The results from
quantitative PCR and immunostaining consistently showed that amylbutyrate treatment
significantly increased the number of pachytene spermatocytes.
After spermatogonial cells were cocultured with Sertoli cells for 14 days, we detected post-
meiotic cells, namely flagellated spermatids, by phase contrast microscopy (Figure 4.3.4A).
The appearance of flagella can also be demonstrated by the existence of the axoneme which
is known to initiate its assemble in early round spermatids (Kierszenbaum, 2002; Marh et al.,
2003).
Figure 4.3.5 The effect of amylbutyrate on the in vitro spermatogenesis. (A) Immunostaining on cells in co-
culture for 10 days with γH2Ax anti-body shown in red and nuclear staining with SYTOX green shown in green.
The meiotic pachytene cells showed a subnuclear dot localization of phospho-H2Ax. (B) Percentage of
pachytene cells. The number of meiotic cells with immunoreative pattern of dot-localized phospho-H2Ax is
represented as a percentage of the total number of germ cells. (* P<0.05). (C) Relative expression level of
pachytene spermatocyte marker p19 in the cells in co-culture for 10 days with different treatments, compared
with untreated control. (D) Immunoreative pattern of tubulin on cells in co-culture for 14 days. The postmeiotic
cells displayed dot cytosolic structure of tubulin shown in red, and nuclear staining with SYTOX green showed
in green. (E) Percentage of round spermtids with axonemes. The number of cells with immunoreactive pattern
of coiled-end dot cytosolic structure of tubulin was expressed as a percentage of the total number of germ cells.

86
(* P<0.05) (F) Relative expression level of haploid cell marker TP1 in the cells in co-culture for 14 days with
different treatments, compared with untreated control.
The existence of axoneme was demonstrated by a dot-like tubulin immunoreactive pattern in
the experiments shown here (Figure 4.3.4 G). The ratio of the number of round spermatids
containing axoneme to the total germinal cell number was determined (Figure 4.3.4 H) and
the statistic results demonstrated that the number of haploid flagellated spermatids (33.5±
3.5%) is significantly larger in amybutyrate treated culture than under control conditions
(20.1±2.8% for non-treated control cells, 17.0±4.3% for bourgeonal treated cells) (Figure
4.3.4 H). The quantitative PCR analysis of TP1, the haploid marker also showed that
amylbutyrate treatment significantly increased the transcription level of TP1 (2.3 fold),
whereas bourgeonal treatment did not show a similar effect. (Figure 4.3.4 I). We infer that
introducing amylbutyrate into culture medium significantly increased the percentage of post-
meiotic cells.
4.3.6 The effect of amylbutyrate on spermatogenic cell proliferation.
To elucidate whether the effect on spermatogenesis is partially or fully due to the change of
germ cell proliferation, we performed a BrdU proliferation assay on the culture germ cells
and examined the proliferation of the cells cultured for 10 days and 14 days, respectively. By
comparing the percentage of positive BrdU immunoreactive cells to total germ cells, we
found no difference among different treatments either in the cells from 10-day culture or 14-
day culture (Figure 4.3.4 C). In cells cultured for 10 days, all different treated conditions
displayed the similar effect on proliferation by displaying about 70% of proliferating cells. In
cells cultured for 14 days, different treatments still did not exhibit any notable difference.
This suggested that activation of MOR283-2 enhanced spermatogonia differentiation but not
through an increase of germ cell proliferation.

87
Figure 4.3.6 The effect of amylbutyrate on the proliferation of spermatogenic cells. The number of BrdU
possitively immunoreacted germ cells is represented as a percentage of the total number of germ cells. Error
bars represent standard deviation.
4.3.7 Amylbutyrate prevented germ cell apoptosis
To investigate the effect of amylbutyrate on spermatogenic cell apoptosis, we kept
spermatogenic cells in seminiferous tubules to maintain the conditions as close to the
physiological environment as possible. Apoptosis of germ cells was induced by incubating
the fragments of seminiferous tubules in serum- and hormone-free medium to exclude
necessary survival factors. To identify the apoptotic cells we performed ISEL analysis of
squash preparation from mouse seminiferous tubules taken immediately after culture in
serum free conditions for 4 hours in the presence or absence of 5 µM amylbutyrate. In the
samples cultured for 4 hours in the presence of amylbutyrate almost no apoptotic cells were
observed, whereas in the samples cultured in the absence of amylbutyrate or in the presence
of the control odorant bourgeonal there was a larger amount of apoptotic cells (Figure 4.3.7),
indicating that amylbutyrate potentially prevents immature germ cells from apoptosis.

88
Figure 4.3.7 The effect of amylbutyrate on spermatogenic cell apoptosis. ISEL analysis of the amylbutyrate
mediated inhibition of in vitro induced apoptosis of sperm cells. The apoptotic cells displayed DNA
fragmentation indicated by the positive staining of 3’-end labelled dig-UTP transferred by terminal transferase.
Amylbutyrate treated cells showed a decrease of apoptotic cells in comparison to control cells.
4.4 Discussion
The transcripts of a subset of ORs have recently been shown to be present in spermatogenic
cells in mouse testis. However, the physiological role of ORs during spermatogenesis
remains enigmatic. In the present study we have shown that MOR283-2 is expressed in
mouse germ cells and the transcription of MOR283-2 is developmentally regulated.
Amylbutyrate is the most active ligand for MOR283-2 and the endogenous ligand is present
in the secretion of Sertoli cells. A proportion of germ cells responded to amylbutyrate, which
was indicated by the transient increase of intracellular Ca2+ concentration upon amylbutyrate
application. Treating spermatogonial cells with 5 µM amylbutyrate in spermatogonia-Sertoli
coculture enhances of spermatogenesis without concurrently affecting germ cell proliferation.
The activation of MOR283-2 was shown to prevent immature germinal cells from apoptosis.
Native MOR283-2 is present in mouse testis throughout spermatogenesis shown by RT-PCR.
Quantitative RT-PCR revealed that the transcription of MOR283-2 is regulated during testis
maturation. The highest level of MOR283-2 transcripts appeared at postnatal day 10,
declined gradually during maturation and stayed steady after puberty, suggesting that there is
a temporal regulation of MOR283-2 transcription during spermatogenesis. Moreover, it is
known that mouse spermatogenesis initiates at postnatal day 7 and the first pachytene
spermatocytes appear at postnatal day 10 (Olesen et al., 2004). The coincidence that maximal
transcription of MOR283-2 appeared almost simultaneously with the emergence of the first
pachytene spermatocytes may demonstrate that the physiological role of MOR283-2 in
spermatogenic cells could be directly correlated with the initiation of spermatogenesis and
meiosis. Therefore, it may serve as an important factor for sperm maturation at the initial
steps. In situ hybridization discovered that MOR283-2 transcripts are predominantly detected

89
in spermatogonia and primary spermatogenic cells. The localization is different from other
ORs which were identified in testis, as these ORs are localized to more mature stage of
sperm development (Fukuda et al., 2004; Fukuda and Touhara, 2006; Walensky et al., 1998).
ORs have been found to be expressed in the developmental stages from pachytene
spermatocytes to late round spermatids (Fukuda and Touhara, 2006). MOR23 is expressed in
elongated spermatids (Fukuda et al., 2004), and two rat ORs were localized to the round
spermatids (Walensky et al., 1998). The different OR localizations may correlate with
different functions during development. Interestingly, not only ORs are found in developing
sperm cells, but also the components of olfactory signaling cascade are found to be expressed
in these cells. Type 4A phophodiesterase, which is mainly expressed in odorant receptor
neurons of the olfactory epithelium (Cherry and Davis, 1999) is also expressed at a high level
in round spermatids (Naro et al., 1996). Transcripts of ACIII predominantly expressed in the
olfactory system (Bakalyar and Reed, 1990) is expressed in developing germ cells,
particularly on the acrosomal membrane of developing spermatids (Gautier-Court et al.,
1998). Moreover, ACIII null mice display a greatly reduced fertilizing ability, which could
be partially caused by a loss of fertility during early spermatid differentiation (Livera et al.,
2005).
Restoration of spermatogenesis in a culture system is very important to understand the
biological event related to germ cell proliferation and differentiation. Spermatogonia-Sertoli
cell co-culture was shown to be a powerful tool to monitor spermatogenesis in vitro
(Lacham-Kaplan, 2004) and the cultured cells successfully progress through meiosis and
differentiate into round sperm cells (step VII) with axonemes and waveform flagella
(Hofmann et al., 1994; Iwanami et al., 2006; Marh et al., 2003). Moreover, it has been
reported that round spermatids developed in vitro by similar culture methods can produce
normal offspring (Marh et al., 2003). Therefore, we employed this system in our study, as the
coculture method allowed to address some aspects of the meiotic process (Godet et al., 2004;

90
Weiss et al., 1997). In our study, cells under different treatment conditions differentiated into
round spermatids with axonemes and flagella, which was confirmed by RT-PCR
amplification of transcripts of three temporal molecular markers of round spermatids: TP1,
TP2 and PRM2. The differentiation in culture is not disrupted by the application of odorants,
as each type of differentiated cells did not synchronously appear or run into the next
differentiation step. The differentiation is more gradual, e.g. pachytene spermatocytes first
appeared at 4-day culture but the largest amount of them appeared until six days later,
whereas in the 14-day culture, they are still present with other cells types. These findings
suggests that despite the limited number of cells that undergo meiosis and become round
spermatids, amylbutyrate and the control odorant had no negative influence for completion of
the meiotic process of spermatogenic cells and showed no disturbance of germ cell
maturation in terms of morphology or at the molecular level.
Another in vitro culture model has been employed to study the germ cells apoptosis. In this
model, testicular tubule fragments were cultured in serum- and hormone- free medium. It has
been shown previously that this treatment is effective to induce germ cell apoptosis (Erkkila
et al., 1997). Intriguingly, the germ cells in this culture were effectively prevented from
undergoing apoptosis when amylbutyrate was additionally added to the medium, whereas the
control odorant bourgeonal did not show the same effect. As this culture contained only
seminiferous tubules and very few interstitial Leydig cells, we can assume that the effect of
amylbutyrate should act directly on spermatogenic cells. Furthermore, since no MOR283-2
transcripts were detected either in Leydig cells or Sertoli cells, we could deduce that
amylbutyrate is directly involved in the regulation of spermatogenic cell survival. Sertoli
cells are known to secrete survival factors and thereby inhibit germ cell apoptosis. Lactate is
one of these factors, it is provided by Sertoli cells and is required for germ cell metabolism
and has been shown to reduce the number of apoptotic cells in seminiferous tubule (Erkkila
et al., 1997). Follicle-stimulating hormones (FSH) and testosterone (Vigier et al., 2004)

91
whose receptors are possessed by Sertoli cells act as hormonal signals that regulate
spermatogenesis and display the ability to reduce germinal cell apoptosis. Therefore, these
factors play roles in regulating spermatogenesis as well as spermatogenic cell survival. We
have shown that an endogenous MOR283-2 ligand is present in the secretion of Sertoli cells.
Therefore, we can propose that the endogenous ligand could partially serve as a survival
factor for spermatogenic cells. Further investigations will be conducted to figure out the
mechanism by which MOR283-2 suppresses the stress-induced apoptosis in spermatogenic
cells. Odorant receptors have been shown to be involved in regulation of cell survival and
selectively rescue a population of OSNs from apoptosis. Odorant stimuli rescue OSNs in a
cAMP-dependent manner by activating the MAPK/CREB-dependent pathway, which
activates corresponding transcriptional pathways (Watt et al., 2004). This activity-dependent
survival could have particular significance in the environment, in that the persistence of
odorant-detecting cells would be dictated by the odorants encountered by the animal (Watt et
al., 2004). Although it is completely unknown if the odorant stimulates the survival of
spermatogenic cells by the similar mechanism as in OSNs, the effect is mediated via a
specific odorant receptor in spermatogenic cells.
The most remarkable finding of the current study is that amylbutyrate treatment significantly
enhanced the number of the meiotic cells (pachytene spermatocytes) and postmeiotic cells
(round spermatids), indicating that activation of MOR283-2 by the ligand has a positive
effect on driving spermatogonia to enter meitotic phase and moreover plays an important role
in the completion of meiosis. The assumption that MOR283-2 is important for initiation of
meiosis was supported by the observation that the number of pachytene spermatocytes is
increased and that the transcription level of p19 mRNA is higher [in pachytene
spermatocytes from stages III-IV onward (Marret et al., 1998)]. It has been reported that at
the beginning of meiosis germ cells located outside of the testis-blood barrier have to pass
through the tight junctions formed by Sertoli cells. Additionally Sertoli cells are directly

92
needed in the movement of developing germ cells (Mruk and Cheng, 2004). Many hormones
and growth factors provided by Sertoli cells may be involved in this process (Mruk and
Cheng, 2004). It has been shown that a subset of ORs are expressed in primordial germ cells
just prior to the meiotic phase in human fetal testis and it has been suggested that specific
members of the OR family may have a function in germ cell movements at the initial phase
of meiosis (Goto et al., 1999; Goto et al., 2001). However, further investigations are required
to elucidate the mechanism of MOR283-2 involved regulating the initial steps of
spermatogenesis. The effect of MOR283-2 in early steps of spermiogenesis is substantiated
by the enhanced appearance of round spermatids and the higher content of TP1 mRNA [in
spermatids from steps 6 to 15-16 of spermiogenesis (Marret et al., 1998)] in the amylbutyrate
treated culture. Some of the hormones and factors secreted by Sertoli cells have been shown
to be involved in male germ cell differentiation. β-nerve growth factor and members of TGF
family have been discover to regulate the second meiotic division of rat spermatocytes
(Damestoy et al., 2005; Perrard et al., 2007). Noteworthy, it has also been discovered that
some odorant stimuli have influences on increasing sperm density and accelerating male
maturation (Koyama, 2004), but the mechanism is not yet known. Other testicular ORs have
been demonstrated to display distinct stage-specific expression pattern, suggesting that ORs
might recognize hormones and growth factors secreted by Sertoli cells at each different
development stage (Fukuda and Touhara, 2006). Taken all together, it may suggest that via
activating MOR283-2, amylbutyrate and the non-identified endogenous ligand act as
important factors to mediate germ cell maturation at different developmental stages.
However, the molecular mechanism underlying transient Ca2+ influx into germ cells and
signaling events which trigger the regulation of spermatogenic cell differentiation through
activated MOR283-2 remains to be further clarified.
In conclusion, our study demonstrated that MOR283-2 is expressed and functions as an
active chemosensory receptor in immature spermatogenic cells. The ligand of MOR283-2,

93
amylbutyrate, prevented male germ cells from undergoing apoptosis. Moreover, the
activation of MOR283-2 by amylbutyrate leads to the promotion of male germ cell
differentiation without affecting germ cell proliferation. Therefore, MOR283-2 plays an
important role in the initiation of spermatogenesis and serves as a survival factor for germ
cells. Furthermore, the endogenous ligand of MOR283-2 is likely present in the secretion of
Sertoli cells. These findings highlight the novel function of testicular ORs in sperm cell
maturation.

94
Summary
Olfaction is one of the most ancient senses to get information from the environment. The molecular mechanism underlying the detection and discrimination of thousands of odorants is based on the largest gene family in the mammalian genome, the odorant receptors (ORs). Besides their expression in the cilia of olfactory sensory neurons, ORs are also expressed in non-olfactory tissues. Ectopic expression of ORs in spermatozoa was correlated with a function in the chemical communication between sperm and the egg. However, in other non-olfactory tissues, OR function remains enigmatic. In the present study, we discovered novel functions of olfactory receptors in non-olfactory tissues, which are presented in three sections. 1). An uncharacterized human OR named “prostate specific G-protein coupled receptor (PSGR)” due to its expression in prostate epithelial cells was cloned and functionally characterized in a heterologous expression system. The identified ligands induce a Ca2+ influx in a prostate cancer epithelial cell line (LNCaP). Ligand treatment additionally resulted in a time dependent activation of members of the MAPK family. The ligand was also found to be a potent inhibitor of cell proliferation and an inducer of apoptosis. Similar results were obtained using primary prostate epithelial cells from resection specimens, from cancer as well as from healthy tissue. Due to the fact that PSGR was found to be overexpressed in prostate cancer, ligands could be developed as novel therapeutic agents for the treatment of prostate cancer. 2). We found another OR, OR51E2, to be functionally expressed in human testis. Using specific intron-spanning primer pairs, we showed that the transcripts of OR51E2 are present in testicular cells. The receptor was localized to the mid-piece and vesicles in the head of human spermatozoa by immunohistochemistry. Furthermore, we demonstrated that the activation of OR51E2 elicits phospholipase C mediated cellular signaling, and that extracellular Ca2+ is not necessary for the OR51E2 elicited Ca2+ mobilization in human sperm. Moreover, activation of OR51E2 enhances human sperm capacitation in a PLC-dependent manner. Additionally, we found that OR51E2 has an evolutionarily conserved testicular expression pattern, and its homologs in various mammalian species display identical putative ligand binding positions and responded to the same ligands. Consequently, OR51E2 homologs may have a highly conserved function in sperm of various mammalian species. 3). We further found that MOR283-2 is expressed in and functions as an active chemosensory receptor in immature spermatogenic cells. The ligand of MOR283-2 (amylbutyrate) was identified using the recombinantly expressed receptor. Application of this ligand to freshly dissociated testicular tubules prevented male germ cells from undergoing apoptosis. Moreover, the activation of MOR283-2 leads to enhanced differentiation of in vitro cultivated male germ cells, without affecting germ cell proliferation. Therefore, MOR283-2 plays an important role in the initiation of spermatogenesis and serves as a survival factor for germ cells. Furthermore, the endogenous ligand of MOR283-2 is likely present in the secretion of Sertoli cells. These findings highlight the novel function of testicular ORs in sperm cell maturation.

95
Zusammenfassung
Der Geruchssinn, welcher phylogenetisch zu den ältesten Sinnessystemen zählt, dient zur Aufnahme von Duftinformation aus der Umwelt und ermöglicht die Diskriminierung von Tausenden von Düften. Der Prozess der olfaktorischen Wahrnehmung basiert auf spezialisierten Rezeptorproteinen, die zur Genfamilie der olfaktorischen Rezeptoren (OR) gehören. Olfaktorische Rezeptoren befinden sich normalerweise in der Zilienmembran der Riechsinneszellen der Nase, sind aber auch in nicht-olfaktorischem Gewebe exprimiert (ektopische Expression). In Spermien vermitteln ektopisch exprimierte OR chemotaktische Prozesse beim Finden der Eizelle, für andere Gewebe ist die physiologische Relevanz ektopischer OR noch unklar. In der vorliegenden Arbeit konnte die Funktionen verschiedener ektopisch exprimierter Geruchsrezeptoren in nicht-olfaktorischen Geweben aufgeklärt werden, wobei folgende drei Aspekte fokussiert wurden:
1) Der humane “prostate specific G-protein coupled receptor (PSGR)”, der zur OR-Familie gehört, konnte kloniert und funktional in Bezug auf das molekulare rezeptive Feld charakterisiert werden. Die identifizierten Liganden wurden hinsichtlich ihrer physiologischen Wirkung in einer epithelialen Prostatakrebs Zelllinie (LNCaP) untersucht. Die Stimulation von PSGR in LnCAP Zellen führt zu einem Anstieg der cytosolischen Calcium-Konzentration und zur Aktivierung von MAP-Kinasen. PSGR-Liganden haben einen proliferationshemmenden Effekt, der möglicherweise aus der Aktivierung apoptotischer Signalwege resultiert. Entsprechende Untersuchungen an primären Epithelzellen von dissoziierten Gewebeproben, sowohl von gesunder Prostata als auch von Prostata-Tumoren, zeigten dieselben Ergebnisse. Die Befunde legen nahe, dass PSGR-Liganden gegebenenfalls neue therapeutische Wirkstoffe zur Behandlung von Prostatakrebs darstellen könnten. 2) Durch RNA Nachweis im menschlichen Hoden und durch immunzytochemische Antikörper-Färbungen in Spermien konnte die Expression eines weiteren OR (OR51E2) in humanen Spermien gezeigt werden. Die Lokalisation im Mittelstück steht im Einklang mit dem räumlich-zeitlichen Antwortmuster der OR51E2-induzierten Calcium-Signale. Der durch den spezifischen Liganden β-Ionon ausgelöste Calcium-Anstieg wird durch den PLC-Inhibitor U73122 geblockt. Weitere Analysen zeigten, dass das Calcium aus intrazellulären Speichern stammt, ein OR-PLC-Signalweg scheint demnach in Spermien vorhanden zu sein. Die Aktivierung von OR51E2 reguliert PLC abhängig die Spermien Kapazitierung. Die Sequenzierung von OR51E2-Homologen aus Hoden-RNA verschiedener Säugetierspezies zeigte eine mehr als 90%ige Übereinstimmung in der übersetzten Aminosäuresequenzen. Die Spermien verschiedener Spezies antworteten alle auf den gleichen Liganden wie der humane OR51E2 mit Ca2+-Anstiegen. OR51E2 könnte also als evolutionär hoch konservierter Rezeptor eine wichtige, möglicherweise auch konservierte Funktion in der Regulierung der Spermien-Funktion in verschiedenen Spezies übernehmen. 3) Darüber hinaus konnte die Expression eines murinen OR, MOR283-2, in unreifen Spermatogonien nachgewiesen werden. Durch Klonierung und rekombinante Expression konnte Amylbutyrat als Ligand von MOR283-2 identifiziert werden. Amylbutyrat fördert die Differenzierung s kultivierter männlicher Keimzellen und verhindert deren Apoptose, ohne einen Einfluss auf die Proliferation dieser Zellen zu nehmen. MOR283-2 spielt daher möglicherweise eine wichtige Rolle in der Initiierung der Spermatogenese und dient als ein Überlebensfaktor für Keimzellen. Ca2+-Imaging Untersuchungen mit dem rekombinanten Rezeptor zeigten, dass der endogene Ligand für MOR283-2 von Sertoli-Zellen sekretiert werden könnte. Diese Befunde weisen auf eine neue Funktion testikulärer ORs in der Spermienreifung hin.

96
Reference list
Reference List
Abaffy,T., Malhotra,A., and Luetje,C.W. (2007). The molecular basis for ligand specificity in a mouse olfactory receptor: a network of functionally important residues. J. Biol. Chem. 282, 1216-1224.
Afshar,M., Hubbard,R.E., and Demaille,J. (1998). Towards structural models of molecular recognition in olfactory receptors. Biochimie 80, 129-135.
Albrizio,M., Guaricci,A.C., Maritato,F., Sciorsci,R.L., Mari,G., Calamita,G., Lacalandra,G.M., Aiudi,G.G., Minoia,R., Dell'Aquila,M.E., and Minoia,P. (2005). Expression and subcellular localization of the mu-opioid receptor in equine spermatozoa: evidence for its functional role. Reproduction. 129, 39-49.
Araneda,R.C., Kini,A.D., and Firestein,S. (2000). The molecular receptive range of an odorant receptor. Nat. Neurosci. 3, 1248-1255.
Arnoult,C., Kazam,I.G., Visconti,P.E., Kopf,G.S., Villaz,M., and Florman,H.M. (1999). Control of the low voltage-activated calcium channel of mouse sperm by egg ZP3 and by membrane hyperpolarization during capacitation. Proc. Natl. Acad. Sci. U. S. A 96, 6757-6762.
Asai,H., Kasai,H., Matsuda,Y., Yamazaki,N., Nagawa,F., Sakano,H., and Tsuboi,A. (1996). Genomic structure and transcription of a murine odorant receptor gene: differential initiation of transcription in the olfactory and testicular cells. Biochem. Biophys. Res. Commun. 221, 240-247.
Bakalyar,H.A. and Reed,R.R. (1990). Identification of a specialized adenylyl cyclase that may mediate odorant detection. Science 250, 1403-1406.
Baldi,E., Luconi,M., Bonaccorsi,L., Krausz,C., and Forti,G. (1996). Human sperm activation during capacitation and acrosome reaction: role of calcium, protein phosphorylation and lipid remodelling pathways. Front Biosci. 1, d189-d205.
Barbour, J. THE OLFACTORY RECEPTOR ASSOCIATED PROTEOME. 2006. Ref Type: Thesis/Dissertation
Bargmann,C.I. (2006). Comparative chemosensation from receptors to ecology. Nature 444, 295-301.
Beato,M. (1989). Gene regulation by steroid hormones. Cell 56, 335-344.
Beites,C.L., Kawauchi,S., Crocker,C.E., and Calof,A.L. (2005). Identification and molecular regulation of neural stem cells in the olfactory epithelium. Exp. Cell Res. 306, 309-316.
Belluscio,L., Gold,G.H., Nemes,A., and Axel,R. (1998). Mice deficient in G(olf) are anosmic. Neuron 20, 69-81.
Bellve,A.R., Cavicchia,J.C., Millette,C.F., O'Brien,D.A., Bhatnagar,Y.M., and Dym,M. (1977). Spermatogenic cells of the prepuberal mouse. Isolation and morphological characterization. J. Cell Biol. 74, 68-85.
Ben-Josef,E., Yang,S.Y., Ji,T.H., Bidart,J.M., Garde,S.V., Chopra,D.P., Porter,A.T., and Tang,D.G. (1999). Hormone-refractory prostate cancer cells express functional follicle-stimulating hormone receptor (FSHR). J. Urol. 161, 970-976.
Berridge,M.J. (1998). Neuronal calcium signaling. Neuron 21, 13-26.
Berridge,M.J., Lipp,P., and Bootman,M.D. (2000). Signal transduction. The calcium entry pas de deux. Science 287, 1604-1605.
Blache,P., Gros,L., Salazar,G., and Bataille,D. (1998). Cloning and tissue distribution of a new rat olfactory receptor-like (OL2). Biochem. Biophys. Res. Commun. 242, 669-672.

97
Boekhoff,I., Tareilus,E., Strotmann,J., and Breer,H. (1990). Rapid activation of alternative second messenger pathways in olfactory cilia from rats by different odorants. EMBO J. 9, 2453-2458.
bou-Haila,A. and Tulsiani,D.R. (2000). Mammalian sperm acrosome: formation, contents, and function. Arch. Biochem. Biophys. 379, 173-182.
Breer,H. (1993). Second messenger signalling in olfaction. Ciba Found. Symp. 179, 97-109.
Breer,H., Boekhoff,I., and Tareilus,E. (1990). Rapid kinetics of second messenger formation in olfactory transduction. Nature 345, 65-68.
Breitbart,H. (2002). Intracellular calcium regulation in sperm capacitation and acrosomal reaction. Mol. Cell Endocrinol. 187, 139-144.
Brewis,I.A. and Moore,H.D. (1997). Molecular mechanisms of gamete recognition and fusion at fertilization. Hum. Reprod. 12, 156-165.
Brodie,A.M., Garrett,W.M., Hendrickson,J.R., Tsai-Morris,C.H., and Williams,J.G. (1983). 1. Estrogen antagonists. Aromatase inhibitors, their pharmacology and application. J. Steroid Biochem. 19, 53-58.
Brown,P.R., Miki,K., Harper,D.B., and Eddy,E.M. (2003). A-kinase anchoring protein 4 binding proteins in the fibrous sheath of the sperm flagellum. Biol. Reprod. 68, 2241-2248.
Brunet,L.J., Gold,G.H., and Ngai,J. (1996). General anosmia caused by a targeted disruption of the mouse olfactory cyclic nucleotide-gated cation channel. Neuron 17, 681-693.
Buck,L. and Axel,R. (1991). A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell 65, 175-187.
Buck,L.B. (1992). A novel multigene family may encode odorant receptors. Soc. Gen. Physiol Ser. 47, 39-51.
Bunick,D., Johnson,P.A., Johnson,T.R., and Hecht,N.B. (1990). Transcription of the testis-specific mouse protamine 2 gene in a homologous in vitro transcription system. Proc. Natl. Acad. Sci. U. S. A 87, 891-895.
Burgos,C., Maldonado,C., Gerez de Burgos,N.M., Aoki,A., and Blanco,A. (1995). Intracellular localization of the testicular and sperm-specific lactate dehydrogenase isozyme C4 in mice. Biol. Reprod. 53, 84-92.
Caballero-Campo,P., Chirinos,M., Fan,X.J., Gonzalez-Gonzalez,M.E., Galicia-Chavarria,M., Larrea,F., and Gerton,G.L. (2006). Biological effects of recombinant human zona pellucida proteins on sperm function. Biol. Reprod. 74, 760-768.
Cao,X., Qin,J., Xie,Y., Khan,O., Dowd,F., Scofield,M., Lin,M.F., and Tu,Y. (2006). Regulator of G-protein signaling 2 (RGS2) inhibits androgen-independent activation of androgen receptor in prostate cancer cells. Oncogene 25, 3719-3734.
Carrera,A., Moos,J., Ning,X.P., Gerton,G.L., Tesarik,J., Kopf,G.S., and Moss,S.B. (1996). Regulation of protein tyrosine phosphorylation in human sperm by a calcium/calmodulin-dependent mechanism: identification of A kinase anchor proteins as major substrates for tyrosine phosphorylation. Dev. Biol. 180, 284-296.
Cato,A.C., Nestl,A., and Mink,S. (2002). Rapid actions of steroid receptors in cellular signaling pathways. Sci. STKE. 2002, RE9.
Chay,C.H., Cooper,C.R., Gendernalik,J.D., Dhanasekaran,S.M., Chinnaiyan,A.M., Rubin,M.A., Schmaier,A.H., and Pienta,K.J. (2002). A functional thrombin receptor (PAR1) is expressed on bone-derived prostate cancer cell lines. Urology 60, 760-765.
Chen,L., He,H.Y., Li,H.M., Zheng,J., Heng,W.J., You,J.F., and Fang,W.G. (2004). ERK1/2 and p38 pathways are required for P2Y receptor-mediated prostate cancer invasion. Cancer Lett. 215, 239-247.
Cherry,J.A. and Davis,R.L. (1999). Cyclic AMP phosphodiesterases are localized in regions of the mouse brain associated with reinforcement, movement, and affect. J. Comp Neurol. 407, 287-301.

98
Chess,A., Simon,I., Cedar,H., and Axel,R. (1994). Allelic inactivation regulates olfactory receptor gene expression. Cell 78, 823-834.
Chodak,G.W., Keane,T., and Klotz,L. (2002). Critical evaluation of hormonal therapy for carcinoma of the prostate. Urology 60, 201-208.
Choi,D., Lee,E., Hwang,S., Jun,K., Kim,D., Yoon,B.K., Shin,H.S., and Lee,J.H. (2001). The biological significance of phospholipase C beta 1 gene mutation in mouse sperm in the acrosome reaction, fertilization, and embryo development. J. Assist. Reprod. Genet. 18, 305-310.
Conzelmann,S., Levai,O., Bode,B., Eisel,U., Raming,K., Breer,H., and Strotmann,J. (2000). A novel brain receptor is expressed in a distinct population of olfactory sensory neurons. Eur. J. Neurosci. 12, 3926-3934.
Cooper,C.R., Chay,C.H., Gendernalik,J.D., Lee,H.L., Bhatia,J., Taichman,R.S., McCauley,L.K., Keller,E.T., and Pienta,K.J. (2003). Stromal factors involved in prostate carcinoma metastasis to bone. Cancer 97, 739-747.
Daaka,Y. (2004). G proteins in cancer: the prostate cancer paradigm. Sci. STKE. 2004, re2.
Dai,J., Shen,R., Sumitomo,M., Stahl,R., Navarro,D., Gershengorn,M.C., and Nanus,D.M. (2002). Synergistic activation of the androgen receptor by bombesin and low-dose androgen. Clin. Cancer Res. 8, 2399-2405.
Dambaki,C., Kogia,C., Kampa,M., Darivianaki,K., Nomikos,M., Anezinis,P., Theodoropoulos,P.A., Castanas,E., and Stathopoulos,E.N. (2005). Membrane testosterone binding sites in prostate carcinoma as a potential new marker and therapeutic target: study in paraffin tissue sections. BMC. Cancer 5, 148.
Damestoy,A., Perrard,M.H., Vigier,M., Sabido,O., and Durand,P. (2005). Transforming growth factor beta-1 decreases the yield of the second meiotic division of rat pachytene spermatocytes in vitro. Reprod. Biol. Endocrinol. 3, 22.
Danzo,B.J., Parrott,J.A., and Skinner,M.K. (1991). Analysis of the steroid binding domain of rat androgen-binding protein. Endocrinology 129, 690-696.
Darszon,A., Acevedo,J.J., Galindo,B.E., Hernandez-Gonzalez,E.O., Nishigaki,T., Trevino,C.L., Wood,C., and Beltran,C. (2006). Sperm channel diversity and functional multiplicity. Reproduction. 131, 977-988.
Darszon,A., Nishigaki,T., Wood,C., Trevino,C.L., Felix,R., and Beltran,C. (2005). Calcium channels and Ca2+ fluctuations in sperm physiology. Int. Rev. Cytol. 243, 79-172.
Davoodpour,P. and Landstrom,M. (2005). 2-Methoxyestradiol-induced apoptosis in prostate cancer cells requires Smad7. J. Biol. Chem. 280, 14773-14779.
De,B.G., Michaut,M., Trevino,C.L., Tomes,C.N., Yunes,R., Darszon,A., and Mayorga,L.S. (2002). The intraacrosomal calcium pool plays a direct role in acrosomal exocytosis. J. Biol. Chem. 277, 49326-49331.
Dorval,V., Dufour,M., and Leclerc,P. (2002). Regulation of the phosphotyrosine content of human sperm proteins by intracellular Ca2+: role of Ca2+-adenosine triphosphatases. Biol. Reprod. 67, 1538-1545.
Doszczak,L., Kraft,P., Weber,H.P., Bertermann,R., Triller,A., Hatt,H., and Tacke,R. (2007). Prediction of Perception: Probing the hOR17-4 Olfactory Receptor Model with Silicon Analogues of Bourgeonal and Lilial. Angew. Chem. Int. Ed Engl.
Drutel,G., Arrang,J.M., Diaz,J., Wisnewsky,C., Schwartz,K., and Schwartz,J.C. (1995). Cloning of OL1, a putative olfactory receptor and its expression in the developing rat heart. Receptors. Channels 3, 33-40.
Dzeja,C., Hagen,V., Kaupp,U.B., and Frings,S. (1999). Ca2+ permeation in cyclic nucleotide-gated channels. EMBO J. 18, 131-144.
Eddy,E.M., Toshimori,K., and O'Brien,D.A. (2003). Fibrous sheath of mammalian spermatozoa. Microsc. Res. Tech. 61, 103-115.

99
Eisenbach,M. and Giojalas,L.C. (2006). Sperm guidance in mammals - an unpaved road to the egg. Nat. Rev. Mol. Cell Biol. 7, 276-285.
Elsaesser,R., Montani,G., Tirindelli,R., and Paysan,J. (2005). Phosphatidyl-inositide signalling proteins in a novel class of sensory cells in the mammalian olfactory epithelium. Eur. J. Neurosci. 21, 2692-2700.
Engedal,N., Korkmaz,C.G., and Saatcioglu,F. (2002). C-Jun N-terminal kinase is required for phorbol ester- and thapsigargin-induced apoptosis in the androgen responsive prostate cancer cell line LNCaP. Oncogene 21, 1017-1027.
Erkkila,K., Aito,H., Aalto,K., Pentikainen,V., and Dunkel,L. (2002). Lactate inhibits germ cell apoptosis in the human testis. Mol. Hum. Reprod. 8, 109-117.
Erkkila,K., Henriksen,K., Hirvonen,V., Rannikko,S., Salo,J., Parvinen,M., and Dunkel,L. (1997). Testosterone regulates apoptosis in adult human seminiferous tubules in vitro. J. Clin. Endocrinol. Metab 82, 2314-2321.
Evans,S.J., Searcy,B.T., and Moore,F.L. (2000). A subset of kappa opioid ligands bind to the membrane glucocorticoid receptor in an amphibian brain. Endocrinology 141, 2294-2300.
Falkenstein,E., Tillmann,H.C., Christ,M., Feuring,M., and Wehling,M. (2000). Multiple actions of steroid hormones--a focus on rapid, nongenomic effects. Pharmacol. Rev. 52, 513-556.
Fawcett,D.W. (1993). Male reproductive system. In A text book of histology, (New York: Chapman&Hall), pp. 816-858.
Feingold,E.A., Penny,L.A., Nienhuis,A.W., and Forget,B.G. (1999). An olfactory receptor gene is located in the extended human beta-globin gene cluster and is expressed in erythroid cells. Genomics 61, 15-23.
Feldman,B.J. and Feldman,D. (2001). The development of androgen-independent prostate cancer. Nat. Rev. Cancer 1, 34-45.
Feldmesser,E., Olender,T., Khen,M., Yanai,I., Ophir,R., and Lancet,D. (2006). Widespread ectopic expression of olfactory receptor genes. BMC. Genomics 7, 121.
Fernandez-Capetillo,O., Lee,A., Nussenzweig,M., and Nussenzweig,A. (2004). H2AX: the histone guardian of the genome. DNA Repair (Amst) 3, 959-967.
Flesch,F.M., Brouwers,J.F., Nievelstein,P.F., Verkleij,A.J., van Golde,L.M., Colenbrander,B., and Gadella,B.M. (2001). Bicarbonate stimulated phospholipid scrambling induces cholesterol redistribution and enables cholesterol depletion in the sperm plasma membrane. J. Cell Sci. 114, 3543-3555.
Floriano,W.B., Vaidehi,N., and Goddard,W.A., III (2004). Making sense of olfaction through predictions of the 3-D structure and function of olfactory receptors. Chem. Senses 29, 269-290.
Floriano,W.B., Vaidehi,N., Goddard,W.A., III, Singer,M.S., and Shepherd,G.M. (2000). Molecular mechanisms underlying differential odor responses of a mouse olfactory receptor. Proc. Natl. Acad. Sci. U. S. A 97, 10712-10716.
Fraser,L.R., oya-Osiguwa,S., Baxendale,R.W., Mededovic,S., and Osiguwa,O.O. (2005). First messenger regulation of mammalian sperm function via adenylyl cyclase/cAMP. J. Reprod. Dev. 51, 37-46.
Fukami,K., Nakao,K., Inoue,T., Kataoka,Y., Kurokawa,M., Fissore,R.A., Nakamura,K., Katsuki,M., Mikoshiba,K., Yoshida,N., and Takenawa,T. (2001). Requirement of phospholipase Cdelta4 for the zona pellucida-induced acrosome reaction. Science 292, 920-923.
Fukami,K., Yoshida,M., Inoue,T., Kurokawa,M., Fissore,R.A., Yoshida,N., Mikoshiba,K., and Takenawa,T. (2003). Phospholipase Cdelta4 is required for Ca2+ mobilization essential for acrosome reaction in sperm. J. Cell Biol. 161, 79-88.
Fukuda,N. and Touhara,K. (2006). Developmental expression patterns of testicular olfactory receptor genes during mouse spermatogenesis. Genes Cells 11, 71-81.

100
Fukuda,N., Yomogida,K., Okabe,M., and Touhara,K. (2004). Functional characterization of a mouse testicular olfactory receptor and its role in chemosensing and in regulation of sperm motility. J. Cell Sci. 117, 5835-5845.
Gadella,B.M., Miller,N.G., Colenbrander,B., van Golde,L.M., and Harrison,R.A. (1999). Flow cytometric detection of transbilayer movement of fluorescent phospholipid analogues across the boar sperm plasma membrane: elimination of labeling artifacts. Mol. Reprod. Dev. 53, 108-125.
Gautier-Court, Salanova,M., and Conti,M. (1998). The olfactory adenylyl cyclase III is expressed in rat germ cells during spermiogenesis. Endocrinology 139, 2588-2599.
Gether,U. and Kobilka,B.K. (1998). G protein-coupled receptors. II. Mechanism of agonist activation. J. Biol. Chem. 273, 17979-17982.
Godara,G., Cannon,G.W., Cannon,G.M., Jr., Bies,R.R., Nelson,J.B., and Pflug,B.R. (2005). Role of endothelin axis in progression to aggressive phenotype of prostate adenocarcinoma. Prostate 65, 27-34.
Godet,M., Damestoy,A., Mouradian,S., Rudkin,B.B., and Durand,P. (2004). Key role for cyclin-dependent kinases in the first and second meiotic divisions of rat spermatocytes. Biol. Reprod. 70, 1147-1152.
Gohji,K., Kitazawa,S., Tamada,H., Katsuoka,Y., and Nakajima,M. (2001). Expression of endothelin receptor a associated with prostate cancer progression. J. Urol. 165, 1033-1036.
Goto,T., Adjaye,J., Rodeck,C.H., and Monk,M. (1999). Identification of genes expressed in human primordial germ cells at the time of entry of the female germ line into meiosis. Mol. Hum. Reprod. 5, 851-860.
Goto,T., Salpekar,A., and Monk,M. (2001). Expression of a testis-specific member of the olfactory receptor gene family in human primordial germ cells. Mol. Hum. Reprod. 7, 553-558.
Graziadei,P.P. and Monti Graziadei,G.A. (1985). Neurogenesis and plasticity of the olfactory sensory neurons. Ann. N. Y. Acad. Sci. 457, 127-142.
Griswold,M.D. (1998). The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 9, 411-416.
Grobner,G., Burnett,I.J., Glaubitz,C., Choi,G., Mason,A.J., and Watts,A. (2000). Observations of light-induced structural changes of retinal within rhodopsin. Nature 405, 810-813.
Hall,S.E., Floriano,W.B., Vaidehi,N., and Goddard,W.A., III (2004). Predicted 3-D structures for mouse I7 and rat I7 olfactory receptors and comparison of predicted odor recognition profiles with experiment. Chem. Senses 29, 595-616.
Halvorson,M., Greenway,D., Eberhart,D., Fitzgerald,K., and Parkinson,A. (1990). Reconstitution of testosterone oxidation by purified rat cytochrome P450p (IIIA1). Arch. Biochem. Biophys. 277, 166-180.
Handel,M.A. (2004). The XY body: a specialized meiotic chromatin domain. Exp. Cell Res. 296, 57-63.
Harayama,H., Murase,T., and Miyake,M. (2005). A cyclic adenosine 3',5'-monophosphate stimulates phospholipase Cgamma1-calcium signaling via the activation of tyrosine kinase in boar spermatozoa. J. Androl 26, 732-740.
Harper,C.V., Barratt,C.L., and Publicover,S.J. (2004). Stimulation of human spermatozoa with progesterone gradients to simulate approach to the oocyte. Induction of [Ca(2+)](i) oscillations and cyclical transitions in flagellar beating. J. Biol. Chem. 279, 46315-46325.
Hatt,H. (2004). Molecular and cellular basis of human olfaction. Chem. Biodivers. 1, 1857-1869.
Hatt,H., Gisselmann,G., and Wetzel,C.H. (1999). Cloning, functional expression and characterization of a human olfactory receptor. Cell Mol. Biol. (Noisy. -le-grand) 45, 285-291.
Hatzoglou,A., Kampa,M., Kogia,C., Charalampopoulos,I., Theodoropoulos,P.A., Anezinis,P., Dambaki,C., Papakonstanti,E.A., Stathopoulos,E.N., Stournaras,C., Gravanis,A., and Castanas,E. (2005). Membrane androgen

101
receptor activation induces apoptotic regression of human prostate cancer cells in vitro and in vivo. J. Clin. Endocrinol. Metab 90, 893-903.
Hecht,N.B. (1990). Regulation of 'haploid expressed genes' in male germ cells. J. Reprod. Fertil. 88, 679-693.
Heinlein,C.A. and Chang,C. (2002). The roles of androgen receptors and androgen-binding proteins in nongenomic androgen actions. Mol. Endocrinol. 16, 2181-2187.
Hendriksen,P.J., Dits,N.F., Kokame,K., Veldhoven,A., van Weerden,W.M., Bangma,C.H., Trapman,J., and Jenster,G. (2006). Evolution of the androgen receptor pathway during progression of prostate cancer. Cancer Res. 66, 5012-5020.
Herve,J.C. (2002). Non-genomic effects of steroid hormones on membrane channels. Mini. Rev. Med. Chem. 2, 411-417.
Ho,H.C. and Suarez,S.S. (2003). Characterization of the intracellular calcium store at the base of the sperm flagellum that regulates hyperactivated motility. Biol. Reprod. 68, 1590-1596.
Hofmann,M.C., Hess,R.A., Goldberg,E., and Millan,J.L. (1994). Immortalized germ cells undergo meiosis in vitro. Proc. Natl. Acad. Sci. U. S. A 91, 5533-5537.
Huque,T. and Bruch,R.C. (1986). Odorant- and guanine nucleotide-stimulated phosphoinositide turnover in olfactory cilia. Biochem. Biophys. Res. Commun. 137, 36-42.
Iwanami,Y., Kobayashi,T., Kato,M., Hirabayashi,M., and Hochi,S. (2006). Characteristics of rat round spermatids differentiated from spermatogonial cells during co-culture with Sertoli cells, assessed by flow cytometry, microinsemination and RT-PCR. Theriogenology 65, 288-298.
Jimenez-Gonzalez,C., Michelangeli,F., Harper,C.V., Barratt,C.L., and Publicover,S.J. (2006). Calcium signalling in human spermatozoa: a specialized 'toolkit' of channels, transporters and stores. Hum. Reprod. Update. 12, 253-267.
Jones,D.T. and Reed,R.R. (1989). Golf: an olfactory neuron specific-G protein involved in odorant signal transduction. Science 244, 790-795.
Kajiya,K., Inaki,K., Tanaka,M., Haga,T., Kataoka,H., and Touhara,K. (2001). Molecular bases of odor discrimination: Reconstitution of olfactory receptors that recognize overlapping sets of odorants. J. Neurosci. 21, 6018-6025.
Kampa,M., Papakonstanti,E.A., Hatzoglou,A., Stathopoulos,E.N., Stournaras,C., and Castanas,E. (2002). The human prostate cancer cell line LNCaP bears functional membrane testosterone receptors that increase PSA secretion and modify actin cytoskeleton. FASEB J. 16, 1429-1431.
Kaplan,P. and Naz,R.K. (1992). The fertilization antigen-1 does not have proteolytic/acrosin activity, but its monoclonal antibody inhibits sperm capacitation and acrosome reaction. Fertil. Steril. 58, 396-402.
Karnik,N.S., Newman,S., Kopf,G.S., and Gerton,G.L. (1992). Developmental expression of G protein alpha subunits in mouse spermatogenic cells: evidence that G alpha i is associated with the developing acrosome. Dev. Biol. 152, 393-402.
Katada,S., Tanaka,M., and Touhara,K. (2004). Structural determinants for membrane trafficking and G protein selectivity of a mouse olfactory receptor. J. Neurochem. 90, 1453-1463.
Kierszenbaum,A.L. (2002). Sperm axoneme: a tale of tubulin posttranslation diversity. Mol. Reprod. Dev. 62, 1-3.
Kimmins,S., Kotaja,N., Davidson,I., and Sassone-Corsi,P. (2004). Testis-specific transcription mechanisms promoting male germ-cell differentiation. Reproduction. 128, 5-12.
Ko,H.J. and Park,T.H. (2006). Dual signal transduction mediated by a single type of olfactory receptor expressed in a heterologous system. Biol. Chem. 387, 59-68.

102
Kobori,H., Miyazaki,S., and Kuwabara,Y. (2000). Characterization of intracellular Ca(2+) increase in response to progesterone and cyclic nucleotides in mouse spermatozoa. Biol. Reprod. 63, 113-120.
Kotaja,N., Kimmins,S., Brancorsini,S., Hentsch,D., Vonesch,J.L., Davidson,I., Parvinen,M., and Sassone-Corsi,P. (2004). Preparation, isolation and characterization of stage-specific spermatogenic cells for cellular and molecular analysis. Nat. Methods 1, 249-254.
Koyama,S. (2004). Primer effects by conspecific odors in house mice: a new perspective in the study of primer effects on reproductive activities. Horm. Behav. 46, 303-310.
Krautwurst,D., Yau,K.W., and Reed,R.R. (1998). Identification of ligands for olfactory receptors by functional expression of a receptor library. Cell 95, 917-926.
Kuroda,Y., Kaneko,S., Yoshimura,Y., Nozawa,S., and Mikoshiba,K. (1999). Are there inositol 1,4,5-triphosphate (IP3) receptors in human sperm? Life Sci. 65, 135-143.
Lacham-Kaplan,O. (2004). In vivo and in vitro differentiation of male germ cells in the mouse. Reproduction. 128, 147-152.
Lawson,C., Dorval,V., Goupil,S., and Leclerc,P. (2007). Identification and localisation of SERCA 2 isoforms in mammalian sperm. Mol. Hum. Reprod.
Lee,D.K., Duan,H.O., and Chang,C. (2001). Androgen receptor interacts with the positive elongation factor P-TEFb and enhances the efficiency of transcriptional elongation. J. Biol. Chem. 276, 9978-9984.
Levin,E.R. (2001). Cell localization, physiology, and nongenomic actions of estrogen receptors. J. Appl. Physiol 91, 1860-1867.
Lindemann,C.B., Orlando,A., and Kanous,K.S. (1992). The flagellar beat of rat sperm is organized by the interaction of two functionally distinct populations of dynein bridges with a stable central axonemal partition. J. Cell Sci. 102 ( Pt 2), 249-260.
Lischka,F.W., Zviman,M.M., Teeter,J.H., and Restrepo,D. (1999). Characterization of inositol-1,4,5-trisphosphate-gated channels in the plasma membrane of rat olfactory neurons. Biophys. J. 76, 1410-1422.
Liu,G., Badeau,R.M., Tanimura,A., and Talamo,B.R. (2006). Odorant receptors directly activate phospholipase C/inositol-1,4,5-trisphosphate coupled to calcium influx in Odora cells. J. Neurochem. 96, 1591-1605.
Livak,K.J. and Schmittgen,T.D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402-408.
Livera,G., Xie,F., Garcia,M.A., Jaiswal,B., Chen,J., Law,E., Storm,D.R., and Conti,M. (2005). Inactivation of the mouse adenylyl cyclase 3 gene disrupts male fertility and spermatozoon function. Mol. Endocrinol. 19, 1277-1290.
Lomvardas,S., Barnea,G., Pisapia,D.J., Mendelsohn,M., Kirkland,J., and Axel,R. (2006). Interchromosomal interactions and olfactory receptor choice. Cell 126, 403-413.
Losel,R., Feuring,M., and Wehling,M. (2002). Non-genomic aldosterone action: from the cell membrane to human physiology. J. Steroid Biochem. Mol. Biol. 83, 167-171.
Losel,R. and Wehling,M. (2003). Nongenomic actions of steroid hormones. Nat. Rev. Mol. Cell Biol. 4, 46-56.
Luconi,M., Bonaccorsi,L., Krausz,C., Gervasi,G., Forti,G., and Baldi,E. (1995). Stimulation of protein tyrosine phosphorylation by platelet-activating factor and progesterone in human spermatozoa. Mol. Cell Endocrinol. 108, 35-42.
Mahadevaiah,S.K., Turner,J.M., Baudat,F., Rogakou,E.P., de,B.P., Blanco-Rodriguez,J., Jasin,M., Keeney,S., Bonner,W.M., and Burgoyne,P.S. (2001). Recombinational DNA double-strand breaks in mice precede synapsis. Nat. Genet. 27, 271-276.

103
Malnic,B., Godfrey,P.A., and Buck,L.B. (2004). The human olfactory receptor gene family. Proc. Natl. Acad. Sci. U. S. A 101, 2584-2589.
Malnic,B., Hirono,J., Sato,T., and Buck,L.B. (1999). Combinatorial receptor codes for odors. Cell 96, 713-723.
Man,O., Gilad,Y., and Lancet,D. (2004). Prediction of the odorant binding site of olfactory receptor proteins by human-mouse comparisons. Protein Sci. 13, 240-254.
Mandal,A., Naaby-Hansen,S., Wolkowicz,M.J., Klotz,K., Shetty,J., Retief,J.D., Coonrod,S.A., Kinter,M., Sherman,N., Cesar,F., Flickinger,C.J., and Herr,J.C. (1999). FSP95, a testis-specific 95-kilodalton fibrous sheath antigen that undergoes tyrosine phosphorylation in capacitated human spermatozoa. Biol. Reprod. 61, 1184-1197.
Maquat,L.E. (2005). Nonsense-mediated mRNA decay in mammals. J. Cell Sci. 118, 1773-1776.
Marh,J., Tres,L.L., Yamazaki,Y., Yanagimachi,R., and Kierszenbaum,A.L. (2003). Mouse round spermatids developed in vitro from preexisting spermatocytes can produce normal offspring by nuclear injection into in vivo-developed mature oocytes. Biol. Reprod. 69, 169-176.
Marquez,B., Ignotz,G., and Suarez,S.S. (2007). Contributions of extracellular and intracellular Ca2+ to regulation of sperm motility: Release of intracellular stores can hyperactivate CatSper1 and CatSper2 null sperm. Dev. Biol. 303, 214-221.
Marret,C., Avallet,O., Perrard-Sapori,M.H., and Durand,P. (1998). Localization and quantitative expression of mRNAs encoding the testis-specific histone TH2B, the phosphoprotein p19, the transition proteins 1 and 2 during pubertal development and throughout the spermatogenic cycle of the rat. Mol. Reprod. Dev. 51, 22-35.
McConkey,D.J., Greene,G., and Pettaway,C.A. (1996). Apoptosis resistance increases with metastatic potential in cells of the human LNCaP prostate carcinoma line. Cancer Res. 56, 5594-5599.
Mendiberri,J., Rauschemberger,M.B., Selles,J., and Massheimer,V. (2006). Involvement of phosphoinositide-3-kinase and phospholipase C transduction systems in the non-genomic action of progesterone in vascular tissue. Int. J. Biochem. Cell Biol. 38, 288-296.
Miki,K., Willis,W.D., Brown,P.R., Goulding,E.H., Fulcher,K.D., and Eddy,E.M. (2002). Targeted disruption of the Akap4 gene causes defects in sperm flagellum and motility. Dev. Biol. 248, 331-342.
Mills,G.B. and Moolenaar,W.H. (2003). The emerging role of lysophosphatidic acid in cancer. Nat. Rev. Cancer 3, 582-591.
Minelli,A., Allegrucci,C., Piomboni,P., Mannucci,R., Lluis,C., and Franco,R. (2000). Immunolocalization of A1 adenosine receptors in mammalian spermatozoa. J. Histochem. Cytochem. 48, 1163-1171.
Mombaerts,P. (1999). Molecular biology of odorant receptors in vertebrates. Annu. Rev. Neurosci. 22, 487-509.
Mombaerts,P. (2004). Genes and ligands for odorant, vomeronasal and taste receptors. Nat. Rev. Neurosci. 5, 263-278.
Mombaerts,P., Wang,F., Dulac,C., Chao,S.K., Nemes,A., Mendelsohn,M., Edmondson,J., and Axel,R. (1996). Visualizing an olfactory sensory map. Cell 87, 675-686.
Moos,J., Faundes,D., Kopf,G.S., and Schultz,R.M. (1995). Composition of the human zona pellucida and modifications following fertilization. Hum. Reprod. 10, 2467-2471.
Moulton,D.G. and Beidler,L.M. (1967). Structure and function in the peripheral olfactory system. Physiol Rev. 47, 1-52.
Mruk,D.D. and Cheng,C.Y. (2004). Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis. Endocr. Rev. 25, 747-806.

104
Naaby-Hansen,S., Mandal,A., Wolkowicz,M.J., Sen,B., Westbrook,V.A., Shetty,J., Coonrod,S.A., Klotz,K.L., Kim,Y.H., Bush,L.A., Flickinger,C.J., and Herr,J.C. (2002). CABYR, a novel calcium-binding tyrosine phosphorylation-regulated fibrous sheath protein involved in capacitation. Dev. Biol. 242, 236-254.
Naaby-Hansen,S., Wolkowicz,M.J., Klotz,K., Bush,L.A., Westbrook,V.A., Shibahara,H., Shetty,J., Coonrod,S.A., Reddi,P.P., Shannon,J., Kinter,M., Sherman,N.E., Fox,J., Flickinger,C.J., and Herr,J.C. (2001). Co-localization of the inositol 1,4,5-trisphosphate receptor and calreticulin in the equatorial segment and in membrane bounded vesicles in the cytoplasmic droplet of human spermatozoa. Mol. Hum. Reprod. 7, 923-933.
Nagata,K., Liberato,D.J., Gillette,J.R., and Sasame,H.A. (1986). An unusual metabolite of testosterone. 17 beta-Hydroxy-4,6-androstadiene-3-one. Drug Metab Dispos. 14, 559-565.
Naro,F., Zhang,R., and Conti,M. (1996). Developmental regulation of unique adenosine 3',5'-monophosphate-specific phosphodiesterase variants during rat spermatogenesis. Endocrinology 137, 2464-2472.
Naz,R.K. and Ahmad,K. (1994). Molecular identities of human sperm proteins that bind human zona pellucida: nature of sperm-zona interaction, tyrosine kinase activity, and involvement of FA-1. Mol. Reprod. Dev. 39, 397-408.
Naz,R.K. and Rajesh,P.B. (2004). Role of tyrosine phosphorylation in sperm capacitation / acrosome reaction. Reprod. Biol. Endocrinol. 2, 75.
Naz,R.K. and Zhu,X. (2002). Molecular cloning and sequencing of cDNA encoding for human FA-1 antigen. Mol. Reprod. Dev. 63, 256-268.
Nelson,J., Bagnato,A., Battistini,B., and Nisen,P. (2003). The endothelin axis: emerging role in cancer. Nat. Rev. Cancer 3, 110-116.
Nelson,J.B., Chan-Tack,K., Hedican,S.P., Magnuson,S.R., Opgenorth,T.J., Bova,G.S., and Simons,J.W. (1996). Endothelin-1 production and decreased endothelin B receptor expression in advanced prostate cancer. Cancer Res. 56, 663-668.
Neuhaus,E.M., Mashukova,A., Barbour,J., Wolters,D., and Hatt,H. (2006a). Novel function of beta-arrestin2 in the nucleus of mature spermatozoa. J. Cell Sci. 119, 3047-3056.
Neuhaus,E.M., Mashukova,A., Zhang,W., Barbour,J., and Hatt,H. (2006b). A specific heat shock protein enhances the expression of mammalian olfactory receptor proteins. Chem. Senses 31, 445-452.
Niimura,Y. and Nei,M. (2003). Evolution of olfactory receptor genes in the human genome. Proc. Natl. Acad. Sci. U. S. A 100, 12235-12240.
Niimura,Y. and Nei,M. (2005). Evolutionary changes of the number of olfactory receptor genes in the human and mouse lineages. Gene 346, 23-28.
Noe,J. and Breer,H. (1998). Functional and molecular characterization of individual olfactory neurons. J. Neurochem. 71, 2286-2293.
Olesen,C., Moller,M., and Byskov,A.G. (2004). Tesmin transcription is regulated differently during male and female meiosis. Mol. Reprod. Dev. 67, 116-126.
Otaki,J.M., Yamamoto,H., and Firestein,S. (2004). Odorant receptor expression in the mouse cerebral cortex. J. Neurobiol. 58, 315-327.
oya-Osiguwa,S.A. and Fraser,L.R. (2007). Cathine, an amphetamine-related compound, acts on mammalian spermatozoa via beta1- and alpha2A-adrenergic receptors in a capacitation state-dependent manner. Hum. Reprod. 22, 756-765.
oya-Osiguwa,S.A., Gibbons,R., and Fraser,L.R. (2006). Identification of functional alpha2- and beta-adrenergic receptors in mammalian spermatozoa. Hum. Reprod. 21, 1555-1563.

105
Pannek,J. and Partin,A.W. (1997). Prostate-specific antigen: what's new in 1997. Oncology (Williston. Park) 11, 1273-1278.
Papakonstanti,E.A., Kampa,M., Castanas,E., and Stournaras,C. (2003). A rapid, nongenomic, signaling pathway regulates the actin reorganization induced by activation of membrane testosterone receptors. Mol. Endocrinol. 17, 870-881.
Parmentier,M., Libert,F., Schurmans,S., Schiffmann,S., Lefort,A., Eggerickx,D., Ledent,C., Mollereau,C., Gerard,C., Perret,J., and . (1992). Expression of members of the putative olfactory receptor gene family in mammalian germ cells. Nature 355, 453-455.
Paysan,J. and Breer,H. (2001). Molecular physiology of odor detection: current views. Pflugers Arch. 441, 579-586.
Perrard,M.H., Vigier,M., Damestoy,A., Chapat,C., Silandre,D., Rudkin,B.B., and Durand,P. (2007). beta-Nerve growth factor participates in an auto/paracrine pathway of regulation of the meiotic differentiation of rat spermatocytes. J. Cell Physiol 210, 51-62.
Petronczki,M., Siomos,M.F., and Nasmyth,K. (2003). Un menage a quatre: the molecular biology of chromosome segregation in meiosis. Cell 112, 423-440.
Picotto,G., Vazquez,G., and Boland,R. (1999). 17beta-oestradiol increases intracellular Ca2+ concentration in rat enterocytes. Potential role of phospholipase C-dependent store-operated Ca2+ influx. Biochem. J. 339 ( Pt 1), 71-77.
Pietrobon,E.O., Soria,M., Dominguez,L.A., Monclus,M.L., and Fornes,M.W. (2005). Simultaneous activation of PLA2 and PLC are required to promote acrosomal reaction stimulated by progesterone via G-proteins. Mol. Reprod. Dev. 70, 58-63.
Porter,A.T., F A C R O, and Ben-Josef,E. (2001). Humoral mechanisms in prostate cancer: A role for FSH. Urol. Oncol. 6, 131-138.
Publicover,S., Harper,C.V., and Barratt,C. (2007). [Ca2+]i signalling in sperm--making the most of what you've got. Nat. Cell Biol. 9, 235-242.
Putney,J.W., Jr. and McKay,R.R. (1999). Capacitative calcium entry channels. Bioessays 21, 38-46.
Raj,G.V., Barki-Harrington,L., Kue,P.F., and Daaka,Y. (2002). Guanosine phosphate binding protein coupled receptors in prostate cancer: a review. J. Urol. 167, 1458-1463.
Ramaswamy,S., Marshall,G.R., McNeilly,A.S., and Plant,T.M. (2000). Dynamics of the follicle-stimulating hormone (FSH)-inhibin B feedback loop and its role in regulating spermatogenesis in the adult male rhesus monkey (Macaca mulatta) as revealed by unilateral orchidectomy. Endocrinology 141, 18-27.
Raming,K., Konzelmann,S., and Breer,H. (1998). Identification of a novel G-protein coupled receptor expressed in distinct brain regions and a defined olfactory zone. Receptors. Channels 6, 141-151.
Rathi,R., Colenbrander,B., Bevers,M.M., and Gadella,B.M. (2001). Evaluation of in vitro capacitation of stallion spermatozoa. Biol. Reprod. 65, 462-470.
Revankar,C.M., Cimino,D.F., Sklar,L.A., Arterburn,J.B., and Prossnitz,E.R. (2005). A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science 307, 1625-1630.
Ronnett,G.V., Cho,H., Hester,L.D., Wood,S.F., and Snyder,S.H. (1993). Odorants differentially enhance phosphoinositide turnover and adenylyl cyclase in olfactory receptor neuronal cultures. J. Neurosci. 13, 1751-1758.
Saunders,C.M., Larman,M.G., Parrington,J., Cox,L.J., Royse,J., Blayney,L.M., Swann,K., and Lai,F.A. (2002). PLC zeta: a sperm-specific trigger of Ca(2+) oscillations in eggs and embryo development. Development 129, 3533-3544.

106
Schandar,M., Laugwitz,K.L., Boekhoff,I., Kroner,C., Gudermann,T., Schultz,G., and Breer,H. (1998). Odorants selectively activate distinct G protein subtypes in olfactory cilia. J. Biol. Chem. 273, 16669-16677.
Schwarzenbacher,K., Fleischer,J., Breer,H., and Conzelmann,S. (2004). Expression of olfactory receptors in the cribriform mesenchyme during prenatal development. Gene Expr. Patterns. 4, 543-552.
Shimada,K., Nakamura,M., Ishida,E., Kishi,M., and Konishi,N. (2003). Roles of p38- and c-jun NH2-terminal kinase-mediated pathways in 2-methoxyestradiol-induced p53 induction and apoptosis. Carcinogenesis 24, 1067-1075.
Shimada,K., Nakamura,M., Ishida,E., Kishi,M., Matsuyoshi,S., and Konishi,N. (2004). The molecular mechanism of sensitization to Fas-mediated apoptosis by 2-methoxyestradiol in PC3 prostate cancer cells. Mol. Carcinog. 39, 1-9.
Shimada,K., Nakamura,M., Ishida,E., and Konishi,N. (2006). Molecular roles of MAP kinases and FADD phosphorylation in prostate cancer. Histol. Histopathol. 21, 415-422.
Silva,A.P., Salim,A.C., Bulgarelli,A., de Souza,J.E., Osorio,E., Caballero,O.L., Iseli,C., Stevenson,B.J., Jongeneel,C.V., de Souza,S.J., Simpson,A.J., and Camargo,A.A. (2003). Identification of 9 novel transcripts and two RGSL genes within the hereditary prostate cancer region (HPC1) at 1q25. Gene 310, 49-57.
Singer,M.S., Oliveira,L., Vriend,G., and Shepherd,G.M. (1995). Potential ligand-binding residues in rat olfactory receptors identified by correlated mutation analysis. Receptors. Channels 3, 89-95.
Singer,M.S., Weisinger-Lewin,Y., Lancet,D., and Shepherd,G.M. (1996). Positive selection moments identify potential functional residues in human olfactory receptors. Receptors. Channels 4, 141-147.
Skryma,R., Mariot,P., Bourhis,X.L., Coppenolle,F.V., Shuba,Y., Vanden,A.F., Legrand,G., Humez,S., Boilly,B., and Prevarskaya,N. (2000). Store depletion and store-operated Ca2+ current in human prostate cancer LNCaP cells: involvement in apoptosis. J. Physiol 527 Pt 1, 71-83.
So,A.I., Hurtado-Coll,A., and Gleave,M.E. (2003). Androgens and prostate cancer. World J. Urol. 21, 325-337.
Spehr,M., Gisselmann,G., Poplawski,A., Riffell,J.A., Wetzel,C.H., Zimmer,R.K., and Hatt,H. (2003). Identification of a testicular odorant receptor mediating human sperm chemotaxis. Science 299, 2054-2058.
Spehr,M., Schwane,K., Heilmann,S., Gisselmann,G., Hummel,T., and Hatt,H. (2004a). Dual capacity of a human olfactory receptor. Curr. Biol. 14, R832-R833.
Spehr,M., Schwane,K., Riffell,J.A., Barbour,J., Zimmer,R.K., Neuhaus,E.M., and Hatt,H. (2004b). Particulate adenylate cyclase plays a key role in human sperm olfactory receptor-mediated chemotaxis. J. Biol. Chem. 279, 40194-40203.
Stathopoulos,E.N., Dambaki,C., Kampa,M., Theodoropoulos,P.A., Anezinis,P., Delakas,D., Delides,G.S., and Castanas,E. (2003). Membrane androgen binding sites are preferentially expressed in human prostate carcinoma cells. BMC. Clin. Pathol. 3, 1.
Steinsapir,J., Socci,R., and Reinach,P. (1991). Effects of androgen on intracellular calcium of LNCaP cells. Biochem. Biophys. Res. Commun. 179, 90-96.
Sternberg,C.N., Pansadoro,V., Calabro,F., Schnetzer,S., Giannarelli,D., Emiliozzi,P., De,P.F., Scarpone,P., De,C.P., Pizzo,M., Platania,A., and Amini,M. (2003). Can patient selection for bladder preservation be based on response to chemotherapy? Cancer 97, 1644-1652.
Sun,Y.H., Gao,X., Tang,Y.J., Xu,C.L., and Wang,L.H. (2006). Androgens induce increases in intracellular calcium via a G protein-coupled receptor in LNCaP prostate cancer cells. J. Androl 27, 671-678.
Syed,V., Gomez,E., and Hecht,N.B. (1999). Messenger ribonucleic acids encoding a serotonin receptor and a novel gene are induced in Sertoli cells by a secreted factor(s) from male rat meiotic germ cells. Endocrinology 140, 5754-5760.

107
Tanaka,Y., Gavrielides,M.V., Mitsuuchi,Y., Fujii,T., and Kazanietz,M.G. (2003). Protein kinase C promotes apoptosis in LNCaP prostate cancer cells through activation of p38 MAPK and inhibition of the Akt survival pathway. J. Biol. Chem. 278, 33753-33762.
Tash,J.S. and Means,A.R. (1982). Regulation of protein phosphorylation and motility of sperm by cyclic adenosine monophosphate and calcium. Biol. Reprod. 26, 745-763.
Taub,J.S., Guo,R., Leeb-Lundberg,L.M., Madden,J.F., and Daaka,Y. (2003). Bradykinin receptor subtype 1 expression and function in prostate cancer. Cancer Res. 63, 2037-2041.
Thebault,S., Lemonnier,L., Bidaux,G., Flourakis,M., Bavencoffe,A., Gordienko,D., Roudbaraki,M., Delcourt,P., Panchin,Y., Shuba,Y., Skryma,R., and Prevarskaya,N. (2005). Novel role of cold/menthol-sensitive transient receptor potential melastatine family member 8 (TRPM8) in the activation of store-operated channels in LNCaP human prostate cancer epithelial cells. J. Biol. Chem. 280, 39423-39435.
Thomas,D. and Hanley,M.R. (1994). Pharmacological tools for perturbing intracellular calcium storage. Methods Cell Biol. 40, 65-89.
Touhara,K., Sengoku,S., Inaki,K., Tsuboi,A., Hirono,J., Sato,T., Sakano,H., and Haga,T. (1999). Functional identification and reconstitution of an odorant receptor in single olfactory neurons. Proc. Natl. Acad. Sci. U. S. A 96, 4040-4045.
Travis,A.J., Foster,J.A., Rosenbaum,N.A., Visconti,P.E., Gerton,G.L., Kopf,G.S., and Moss,S.B. (1998). Targeting of a germ cell-specific type 1 hexokinase lacking a porin-binding domain to the mitochondria as well as to the head and fibrous sheath of murine spermatozoa. Mol. Biol. Cell 9, 263-276.
Turner,R.M. (2003). Tales from the tail: what do we really know about sperm motility? J. Androl 24, 790-803.
Turner,R.M., Eriksson,R.L., Gerton,G.L., and Moss,S.B. (1999). Relationship between sperm motility and the processing and tyrosine phosphorylation of two human sperm fibrous sheath proteins, pro-hAKAP82 and hAKAP82. Mol. Hum. Reprod. 5, 816-824.
Uzgare,A.R., Kaplan,P.J., and Greenberg,N.M. (2003). Differential expression and/or activation of P38MAPK, erk1/2, and jnk during the initiation and progression of prostate cancer. Prostate 55, 128-139.
Vanden,A.F., Skryma,R., Shuba,Y., Van,C.F., Slomianny,C., Roudbaraki,M., Mauroy,B., Wuytack,F., and Prevarskaya,N. (2002). Bcl-2-dependent modulation of Ca(2+) homeostasis and store-operated channels in prostate cancer cells. Cancer Cell 1, 169-179.
Vanderhaeghen,P., Schurmans,S., Vassart,G., and Parmentier,M. (1997a). Molecular cloning and chromosomal mapping of olfactory receptor genes expressed in the male germ line: evidence for their wide distribution in the human genome. Biochem. Biophys. Res. Commun. 237, 283-287.
Vanderhaeghen,P., Schurmans,S., Vassart,G., and Parmentier,M. (1997b). Specific repertoire of olfactory receptor genes in the male germ cells of several mammalian species. Genomics 39, 239-246.
Vanderhaeghen,P., Schurmans,S., Vassart,G., and Parmentier,M. (1993). Olfactory receptors are displayed on dog mature sperm cells. J. Cell Biol. 123, 1441-1452.
Vanti,W.B., Nguyen,T., Cheng,R., Lynch,K.R., George,S.R., and O'Dowd,B.F. (2003). Novel human G-protein-coupled receptors. Biochem. Biophys. Res. Commun. 305, 67-71.
Vigier,M., Weiss,M., Perrard,M.H., Godet,M., and Durand,P. (2004). The effects of FSH and of testosterone on the completion of meiosis and the very early steps of spermiogenesis of the rat: an in vitro study. J. Mol. Endocrinol. 33, 729-742.
Walensky,L.D., Roskams,A.J., Lefkowitz,R.J., Snyder,S.H., and Ronnett,G.V. (1995). Odorant receptors and desensitization proteins colocalize in mammalian sperm. Mol. Med. 1, 130-141.
Walensky,L.D., Ruat,M., Bakin,R.E., Blackshaw,S., Ronnett,G.V., and Snyder,S.H. (1998). Two novel odorant receptor families expressed in spermatids undergo 5'-splicing. J. Biol. Chem. 273, 9378-9387.

108
Walensky,L.D. and Snyder,S.H. (1995). Inositol 1,4,5-trisphosphate receptors selectively localized to the acrosomes of mammalian sperm. J. Cell Biol. 130, 857-869.
Walker,W.H. and Cheng,J. (2005). FSH and testosterone signaling in Sertoli cells. Reproduction. 130, 15-28.
Wang,J., Weng,J., Cai,Y., Penland,R., Liu,M., and Ittmann,M. (2006). The prostate-specific G-protein coupled receptors PSGR and PSGR2 are prostate cancer biomarkers that are complementary to alpha-methylacyl-CoA racemase. Prostate 66, 847-857.
Wargo,M.J. and Smith,E.F. (2003). Asymmetry of the central apparatus defines the location of active microtubule sliding in Chlamydomonas flagella. Proc. Natl. Acad. Sci. U. S. A 100, 137-142.
Watt,W.C., Sakano,H., Lee,Z.Y., Reusch,J.E., Trinh,K., and Storm,D.R. (2004). Odorant stimulation enhances survival of olfactory sensory neurons via MAPK and CREB. Neuron 41, 955-967.
Weber,M., Pehl,U., Breer,H., and Strotmann,J. (2002). Olfactory receptor expressed in ganglia of the autonomic nervous system. J. Neurosci. Res. 68, 176-184.
Weiss,M., Vigier,M., Hue,D., Perrard-Sapori,M.H., Marret,C., Avallet,O., and Durand,P. (1997). Pre- and postmeiotic expression of male germ cell-specific genes throughout 2-week cocultures of rat germinal and Sertoli cells. Biol. Reprod. 57, 68-76.
Weng,J., Ma,W., Mitchell,D., Zhang,J., and Liu,M. (2005a). Regulation of human prostate-specific G-protein coupled receptor, PSGR, by two distinct promoters and growth factors. J. Cell Biochem. 96, 1034-1048.
Weng,J., Wang,J., Cai,Y., Stafford,L.J., Mitchell,D., Ittmann,M., and Liu,M. (2005b). Increased expression of prostate-specific G-protein-coupled receptor in human prostate intraepithelial neoplasia and prostate cancers. Int. J. Cancer 113, 811-818.
Wennemuth,G., Babcock,D.F., and Hille,B. (2003). Calcium clearance mechanisms of mouse sperm. J. Gen. Physiol 122, 115-128.
Wennemuth,G., Westenbroek,R.E., Xu,T., Hille,B., and Babcock,D.F. (2000). CaV2.2 and CaV2.3 (N- and R-type) Ca2+ channels in depolarization-evoked entry of Ca2+ into mouse sperm. J. Biol. Chem. 275, 21210-21217.
Wetzel,C.H., Oles,M., Wellerdieck,C., Kuczkowiak,M., Gisselmann,G., and Hatt,H. (1999). Specificity and sensitivity of a human olfactory receptor functionally expressed in human embryonic kidney 293 cells and Xenopus Laevis oocytes. J. Neurosci. 19, 7426-7433.
Wong,S.T., Trinh,K., Hacker,B., Chan,G.C., Lowe,G., Gaggar,A., Xia,Z., Gold,G.H., and Storm,D.R. (2000). Disruption of the type III adenylyl cyclase gene leads to peripheral and behavioral anosmia in transgenic mice. Neuron 27, 487-497.
Xia,C., Ma,W., Wang,F., Hua,S., and Liu,M. (2001). Identification of a prostate-specific G-protein coupled receptor in prostate cancer. Oncogene 20, 5903-5907.
Xiao,D., Qu,X., and Weber,H.C. (2003). Activation of extracellular signal-regulated kinase mediates bombesin-induced mitogenic responses in prostate cancer cells. Cell Signal. 15, 945-953.
Xie,Y., Gibbs,T.C., Mukhin,Y.V., and Meier,K.E. (2002). Role for 18:1 lysophosphatidic acid as an autocrine mediator in prostate cancer cells. J. Biol. Chem. 277, 32516-32526.
Xu,L.L., Stackhouse,B.G., Florence,K., Zhang,W., Shanmugam,N., Sesterhenn,I.A., Zou,Z., Srikantan,V., Augustus,M., Roschke,V., Carter,K., McLeod,D.G., Moul,J.W., Soppett,D., and Srivastava,S. (2000). PSGR, a novel prostate-specific gene with homology to a G protein-coupled receptor, is overexpressed in prostate cancer. Cancer Res. 60, 6568-6572.
Xu,L.L., Sun,C., Petrovics,G., Makarem,M., Furusato,B., Zhang,W., Sesterhenn,I.A., McLeod,D.G., Sun,L., Moul,J.W., and Srivastava,S. (2006). Quantitative expression profile of PSGR in prostate cancer. Prostate Cancer Prostatic. Dis. 9, 56-61.

109
Yuan,T.T., Toy,P., McClary,J.A., Lin,R.J., Miyamoto,N.G., and Kretschmer,P.J. (2001). Cloning and genetic characterization of an evolutionarily conserved human olfactory receptor that is differentially expressed across species. Gene 278, 41-51.
Zhang,X. and Firestein,S. (2002). The olfactory receptor gene superfamily of the mouse. Nat. Neurosci. 5, 124-133.
Zhang,X., Rogers,M., Tian,H., Zhang,X., Zou,D.J., Liu,J., Ma,M., Shepherd,G.M., and Firestein,S.J. (2004a). High-throughput microarray detection of olfactory receptor gene expression in the mouse. Proc. Natl. Acad. Sci. U. S. A 101, 14168-14173.
Zhang,X., Rogers,M., Tian,H., Zhang,X., Zou,D.J., Liu,J., Ma,M., Shepherd,G.M., and Firestein,S.J. (2004b). High-throughput microarray detection of olfactory receptor gene expression in the mouse. Proc. Natl. Acad. Sci. U. S. A 101, 14168-14173.
Zhao,H. and Reed,R.R. (2001). X inactivation of the OCNC1 channel gene reveals a role for activity-dependent competition in the olfactory system. Cell 104, 651-660.
Zufall,F., Firestein,S., and Shepherd,G.M. (1994). Cyclic nucleotide-gated ion channels and sensory transduction in olfactory receptor neurons. Annu. Rev. Biophys. Biomol. Struct. 23, 577-607.

110
Weiyi Zhang Overbergstrasse. 8
D-44801 Bochum, Germany Phone: +49-234-3228104,
Email: [email protected]; [email protected]
EDUCATION
• PhD Department of Cellphysiology, Ruhr-Universität Bochum, Bochum, Germany 2004-present • Pre-PhD Department of Cellphysiology, Ruhr-Universität Bochum, Bochum, Germany 2003-2004 • Master Department of Biology, Xiamen University, Xiamen, China 2001-2003 • Bachelor Department of Biology, Xiamen University, Xiamen, China 1997-2001
RESEARCH EXPERIENCE International Max-Planck Chemical Biology Program, Ruhr-Universität Bochum, Germany 2003-present Doctoral thesis research conducted with Dr. Eva Neuhaus and Prof. Dr.Dr. Hanns Hatt Thesis: Investigation of functions of olfactory receptors in non-olfactory tissues. Molecular biology and Biochemistry Program, Xiamen University, Xiamen, China 2001-2003 Master study conducted with Dr. Shengcai Lin Thesis: Determinants that control specific docking of TAB1 to p38alpha Department of Biology, Xiamen University, Xiamen, China 1997-2001 Bachelor degree thesis study Thesis: Genetic investigation of p62 gene in liver cancer.
POSTERS AND TALKS 1.Weiyi Zhang, Hanns Hatt, Eva Neuhaus. Investigation into the dimerization and ligand identification of human olfactory receptors. FEBS Lecture Course on Cellular Signaling and 4th Dubrovnik Signaling conference. May 21-27, 2004. Dubrovnik Croatia. (Poster) 2.Weiyi Zhang, Anastasia Moshukova, Hanns Hatt, Eva Neuhaus. A specific heat shock protein as an enhancer for the expression of mammalian olfactory receptor proteins. 6th Meeting of the German Neuroscience Society and 30th Göttingen Neurobiology Conference. Feb 17-20, 2005. (Poster) 3.Weiyi Zhang, Hanns Hatt, Eva Neuhaus. Functional investigation of prostate specific G-protein coupled odorant receptor. The 5th conference of Asian-pacific organization of cell biology. Oct.28-31,2006. Beijing China (Talk) 4. Weiyi Zhang, Hanns Hatt, Eva Neuhaus. Functional investigation of prostate specific G-protein coupled odorant receptor. The 46th annual meeting of American Society of Cell Biology. Dec 9-13, 2006. San Diego USA (Poster) 5. Max-Planck Research School Annual Retreat, Nov 2004, Rom, Italy. (Talk) 6.Max-Planck Research School Annual Retreat, Sep 2005, Jena, Germany. (Talk and poster)

111
REFERENCES Dr. Eva Neuhaus Ruhr-Universität Bochum Department of Cell physiology Universitätstr. 150 Gebaude ND4 D-44780 Bochum Germany Phone: +49-234-3224315 Email: [email protected] Prof. Dr.Dr.Hanns Hatt Ruhr-Universität Bochum Department of Cell physiology Universitätstr. 150 Gebaude ND4 D-44780 Bochum Germany Phone: +49-234-3224586 Email: hanns.hatt@ ruhr-uni-bochum.de Prof. Dr. Martin Engelhard Max Planck Institute of Molecular Physiology Dept. of Physical Biochemistry Otto-Hahn-Strasse 11 44227 Dortmund Germany Phone: +49- 231-133-2302 Email: [email protected]
PUBLICATIONS
1. Eva Neuhaus*, Weiyi Zhang*, Joachim Noldus, Hanns Hatt. Functional investigation of prostate specific G-protein coupled olfactory receptor (PSGR). (in preparation) 2. Weiyi Zhang, Hanns Hatt, Eva Neuhaus. Promotion of mouse spermatogenesis by activating odorant receptor MOR283-2. (in preparation) 3. Buchwald B, Zhang G, Vogt-Eisele A, Zhang W, Ahangari R, Griffin JW, Hatt H, Toyka KV, Sheikh KA. Different monoclonal anti-ganglioside Abs show distinct effects on presynaptic motor nerve terminals: implications for immune neuropathies. (submitted). 4. Neuhaus EM, Mashukova A, Zhang W, Barbour J, Hatt H. A specific heat shock protein enhances the expression of mammalian olfactory receptor proteins. Chem Senses. 2006 Jun; 31(5): 445-52. 5. Neuhaus EM, Gisselmann G, Zhang W, Dooley R, Stortkuhl K, Hatt H. Odorant receptor heterodimerization in the olfactory system of Drosophila melanogaster. Nat Neurosci. 2005 Jan;8(1): 15-7 6. Peng X, Zhang J, Wang S, Lin Z, Zhang W. Immuno-capture PCR for detection of Aeromonas hydrophila. J Microbiol Methods. 2002 May; 49(3): 335-8.