znt-1 enhances the activity and surface expression of t ... · znt-1 enhances the activity and...
TRANSCRIPT

ZnT-1 enhances the activity and surface expression of T-type calciumchannels through activation of Ras-ERK signaling
Merav Mor,1,5* Ofer Beharier,1,5* Shiri Levy,1 Joy Kahn,1 Shani Dror,1,5 Daniel Blumenthal,2
Levi A. Gheber,2 Asher Peretz,3 Amos Katz,4,5 Arie Moran,1* and Yoram Etzion5*1Department of Physiology, Faculty of Health Sciences, Ben-Gurion University of the Negev, Beer-Sheva, Israel; 2Departmentof Biotechnology Engineering, Ben-Gurion University of the Negev, Beer-Sheva, Israel; 3Department of Physiology andPharmacology, Sackler Medical School, Tel Aviv University, Tel Aviv, Israel; 4Department of Cardiology, Barzilai MedicalCenter, Ashkelon, Israel; and 5Cardiac Arrhythmia Research Laboratory, Soroka University Medical Center and Ben-GurionUniversity of the Negev, Beer-Sheva, Israel
Submitted 2 December 2011; accepted in final form 7 May 2012
Mor M, Beharier O, Levy S, Kahn J, Dror S, Blumenthal D,Gheber LA, Peretz A, Katz A, Moran A, Etzion Y. ZnT-1enhances the activity and surface expression of T-type calciumchannels through activation of Ras-ERK signaling. Am J PhysiolCell Physiol 303: C192–C203, 2012. First published May 9, 2012;doi:10.1152/ajpcell.00427.2011.—Zinc transporter-1 (ZnT-1) is aputative zinc transporter that confers cellular resistance from zinctoxicity. In addition, ZnT-1 has important regulatory functions, in-cluding inhibition of L-type calcium channels and activation of Raf-1kinase. Here we studied the effects of ZnT-1 on the expression andfunction of T-type calcium channels. In Xenopus oocytes expressingvoltage-gated calcium channel (CaV) 3.1 or CaV3.2, ZnT-1 enhancedthe low-threshold calcium currents (IcaT) to 182 � 15 and 167.95 �9.27% of control, respectively (P � 0.005 for both channels). Asexpected, ZnT-1 also enhanced ERK phosphorylation. Coexpressionof ZnT-1 and nonactive Raf-1 blocked the ZnT-1-mediated ERKphosphorylation and abolished the ZnT-1-induced augmentation ofIcaT. In mammalian cells (Chinese hamster ovary), coexpression ofCaV3.1 and ZnT-1 increased the IcaT to 166.37 � 6.37% comparedwith cells expressing CaV3.1 alone (P � 0.01). Interestingly, surfaceexpression measurements using biotinylation or total internal reflec-tion fluorescence microscopy indicated marked ZnT-1-induced en-hancement of CaV3.1 surface expression. The MEK inhibitor PD-98059 abolished the ZnT-1-induced augmentation of surface expres-sion of CaV3.1. In cultured murine cardiomyocytes (HL-1 cells),transient exposure to zinc, leading to enhanced ZnT-1 expression, alsoenhanced the surface expression of endogenous CaV3.1 channels.Consistently, in these cells, endothelin-1, a potent activator of Ras-ERK signaling, enhanced the surface expression of CaV3.1 channelsin a PD-98059-sensitive manner. Our findings indicate that ZnT-1enhances the activity of CaV3.1 and CaV3.2 through activation ofRas-ERK signaling. The augmentation of CaV3.1 currents by Ras-ERK activation is associated with enhanced trafficking of the channelto the plasma membrane.
extracellular signal-regulated kinase signaling; trafficking
LOW-VOLTAGE-ACTIVATED T-TYPE Ca2� channels (TTCC) are linkedto the pathophysiology of various disease states and are recog-nized as universal players in a plethora of cellular functions,including differentiation and proliferation, neuronal excitability,hormone secretion, muscle contraction, cardiac pacemaker activ-
ity, and transmission of pain signals (6, 24, 32, 42). TTCC aredistinguished among voltage-gated Ca2� channels (VGCC) bytheir low-voltage thresholds for activation and inactivation, fastinactivation, and smaller conductance of Ba2� (49). Molecularcloning demonstrated the existence of protein family consisting ofthree isoforms: voltage-gated calcium channels (CaV) 3.1,CaV3.2, and CaV3.3.
The putative structure of the TTCC �-subunit has the samegeneral topology as the �-subunit of high-voltage-activatedCa2� channels. However, most evidence indicates that thebiophysical properties of TTCC are not affected by auxiliarysubunits (42, 49). In addition, while the modulatory pathwaysthat affect high-voltage-activated Ca2� channels have beenextensively studied, the modulation of TTCC has only latelybeen substantiated (10). Nevertheless, accumulating data indi-cate that TTCC are highly regulated at various levels. Analysisof the density of T currents (IcaT) indicates major expressionchanges during development, as well as in various experimen-tal models of pathological conditions (32). In addition, theTTCC are directly affected by endogenous ligands, such asarachidonic acid and polyunsaturated fatty acids (9, 50), aswell as by the activation of various G protein-coupled receptors(10, 20, 41). TTCC are also modulated by several secondmessengers and protein kinase pathways, including CamKII,PKA, PKC, and Rho kinase (20).
Since TTCC lacks an �-interaction domain, they do notinteract with the regulatory �-subunit that normally controlstrafficking to the plasma membrane of other VGCC (1). Nev-ertheless, changes in the surface expression of T channels havebeen recently demonstrated (2, 47). While CaV3.2 channels aremarkedly retained in intracellular compartments, mutations inthe intracellular loop I-II of CaV3.2 have been shown toprominently enhance the surface expression of this channel andmay have important pathophysiological implications in ab-sence epilepsy (54). CaV3 channels are not necessarily uni-form in terms of regulation. For instance, when CaV3.1 andCaV3.2 are compared, various reports indicate that distincteffects are observed on the two channel types (44, 49). Inaddition, the interdomain loops, which seem to have importantregulatory functions, are not conserved between these twoisoforms.
The transmembrane protein zinc transporter-1 (ZnT-1) is aubiquitously expressed protein that was reported to act as anendogenous inhibitor of L-type calcium channels (LTCC) (4,37, 46). Recently, our group demonstrated that ZnT-1 interactswith the calcium channel �-subunit, leading to a reduction in
* M. Mor and O. Beharier contributed equally to this work. A. Moran andY. Etzion contributed equally to this work.
Address for reprint requests and other correspondence: Y. Etzion, CardiacArrhythmia Research Laboratory, Faculty of Health sciences, Ben-GurionUniv. & Soroka Univ. Medical Center, P.O. Box 151, Beer-Sheva 84101,Israel (e-mail: [email protected]).
Am J Physiol Cell Physiol 303: C192–C203, 2012.First published May 9, 2012; doi:10.1152/ajpcell.00427.2011.
0363-6143/12 Copyright © 2012 the American Physiological Society http://www.ajpcell.orgC192
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from
the surface expression of the LTCC �-subunit (30). ZnT-1 hasa well-documented role in conferring resistance against zinctoxicity (14, 39, 40, 53). In addition, ZnT-1 binds to theamino-terminal regulatory portion of Raf-1 kinase and pro-motes its enzymatic activity, thereby activating the Ras-ERKsignaling cascade (7, 23). Thus, as already outlined before,ZnT-1 may have important and broad regulatory roles, onlyparts of which are directly related to zinc homeostasis (3, 13).
LTCC and TTCC are coexpressed in many tissues andappear to have different functions in a variety of cells, such ascardiomyocytes, pancreatic �-cells, and neurons (17, 38, 45).The goal of the present study was to determine the effects ofZnT-1 on CaV3.1 and CaV3.2 compared with its recentlydescribed effect on LTCC function. Our results indicate that, incontrast to the inhibitory effect of ZnT-1 on LTCC, ZnT-1markedly augments the activity of CaV3.1 and CaV3.2 chan-
nels. We further demonstrate that this functional augmentationrelates to ZnT-1-induced activation of Ras-ERK signaling andis associated with enhanced surface expression of the channels.These findings document a novel regulatory function of ZnT-1and a new mode of regulation of TTCC channels by theRas-ERK signaling cascade.
EXPERIMENTAL PROCEDURES
Expression of CaV3.1, CaV3.2, and ZnT-1 in Xenopus oocytes. Oocytesof mature female Xenopus laevis frogs were excised and prepared aspreviously described (4, 30). Complementary cRNAs were synthe-sized by in vitro transcription with T7 or an SP6 Amplicap High-YieldMessage Maker Kit (Epicentre Technologies, Madison, WI). Therecombinant plasmids used in these reactions were pGEM-HE-�1Hand pGEM-HE-�1G (42, 43), pSPCA1 [rabbit skeletal muscle �2�(35)], and pXen1-ZnT1 [rat SLC30A (23), a kind gift from Dr.
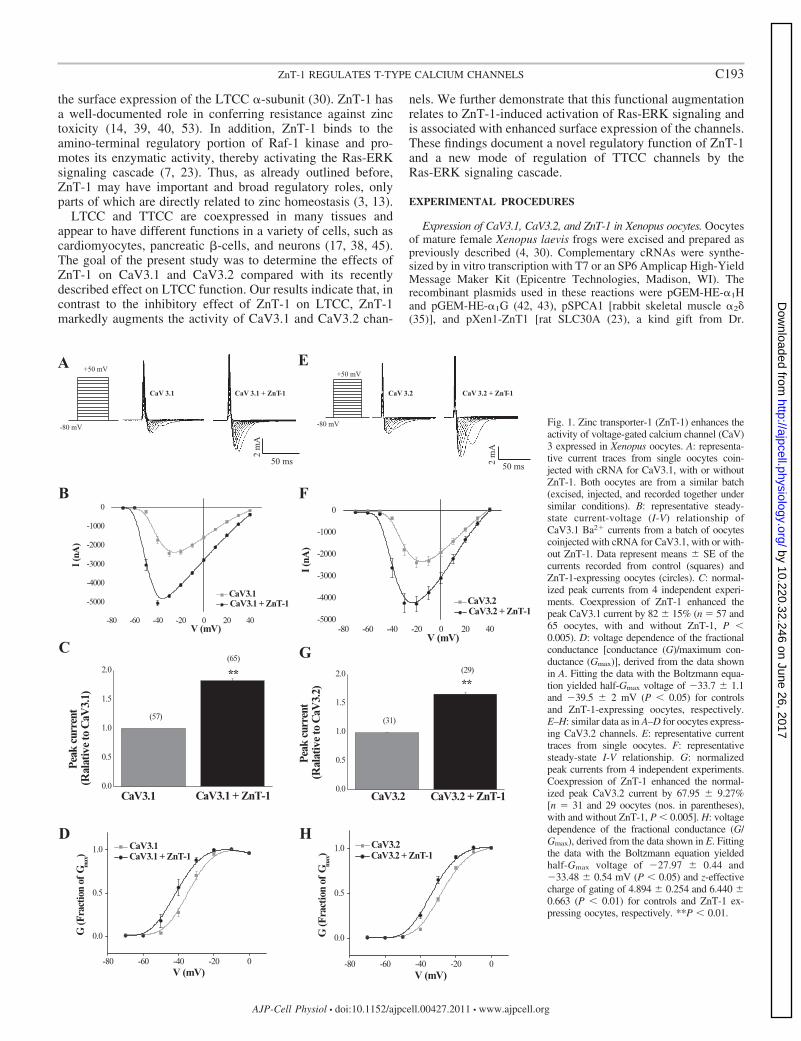
Fig. 1. Zinc transporter-1 (ZnT-1) enhances theactivity of voltage-gated calcium channel (CaV)3 expressed in Xenopus oocytes. A: representa-tive current traces from single oocytes coin-jected with cRNA for CaV3.1, with or withoutZnT-1. Both oocytes are from a similar batch(excised, injected, and recorded together undersimilar conditions). B: representative steady-state current-voltage (I-V) relationship ofCaV3.1 Ba2� currents from a batch of oocytescoinjected with cRNA for CaV3.1, with or with-out ZnT-1. Data represent means � SE of thecurrents recorded from control (squares) andZnT-1-expressing oocytes (circles). C: normal-ized peak currents from 4 independent experi-ments. Coexpression of ZnT-1 enhanced thepeak CaV3.1 current by 82 � 15% (n � 57 and65 oocytes, with and without ZnT-1, P �0.005). D: voltage dependence of the fractionalconductance [conductance (G)/maximum con-ductance (Gmax)], derived from the data shownin A. Fitting the data with the Boltzmann equa-tion yielded half-Gmax voltage of �33.7 � 1.1and �39.5 � 2 mV (P � 0.05) for controlsand ZnT-1-expressing oocytes, respectively.E–H: similar data as in A–D for oocytes express-ing CaV3.2 channels. E: representative currenttraces from single oocytes. F: representativesteady-state I-V relationship. G: normalizedpeak currents from 4 independent experiments.Coexpression of ZnT-1 enhanced the normal-ized peak CaV3.2 current by 67.95 � 9.27%[n � 31 and 29 oocytes (nos. in parentheses),with and without ZnT-1, P � 0.005]. H: voltagedependence of the fractional conductance (G/Gmax), derived from the data shown in E. Fittingthe data with the Boltzmann equation yieldedhalf-Gmax voltage of �27.97 � 0.44 and�33.48 � 0.54 mV (P � 0.05) and z-effectivecharge of gating of 4.894 � 0.254 and 6.440 �0.663 (P � 0.01) for controls and ZnT-1 ex-pressing oocytes, respectively. **P � 0.01.
C193ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from
Anthony Muslin]. Oocytes were injected with equal quantities (2.5ng) of cRNA of either CaV3.1 or CaV3.2, with or without cRNA ofZnT-1 (2.5 ng). In addition, �2� was injected into all oocytes. Injectedoocytes were stored at 18°C for 4 days in NDE96 solution before theiruse for experiments.
Barium current recordings in Xenopus oocytes. Current was mon-itored by a two-electrode voltage clamp (TEVC) using Gene Clamp500 amplifier (Molecular Devices, Sunnyvale, CA), as previouslydescribed (4, 30). The bath solution contained the following (in mM):40 Ba(OH)2, 50 NaOH, 2 KOH, and 5 HEPES (titrated to pH 7.5 withmethanesulfonic acid). Currents were measured in response to 200-ms-long voltage-clamp pulses generated every 2 s from a holdingpotential of �80 mV to test potentials between �80 and �50 mV.Barium currents in water-injected oocytes or cells injected with thecRNA of ZnT-1 alone were negligible.
Cell culture and transfection. Human embryonic kidney cells (HEKT-293) and Chinese hamster ovary (CHO) cells were maintained inhigh-glucose Dulbecco’s modified Eagle’s medium supplementedwith (vol/vol) 1% penicillin-streptomycin, 1% L-glutamine, and 10%fetal bovine serum at 37°C in a humidified 5% CO2 incubator.HEKT-293 cells were subcultured into 60-mm dishes 1 day beforetransfection and seeded to reach 50–70% confluence. Transient trans-fection of HEKT-293 cells was carried out utilizing calcium phos-phate transfection protocol. CHO cells were transfected utilizing
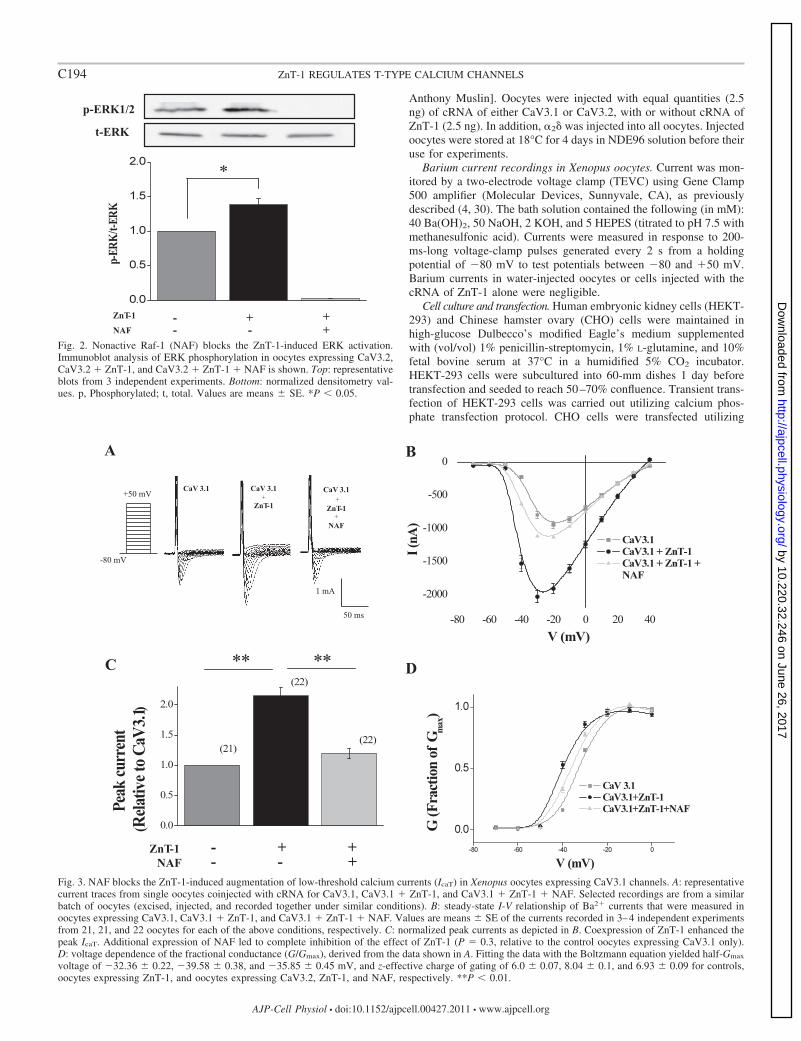
Fig. 2. Nonactive Raf-1 (NAF) blocks the ZnT-1-induced ERK activation.Immunoblot analysis of ERK phosphorylation in oocytes expressing CaV3.2,CaV3.2 � ZnT-1, and CaV3.2 � ZnT-1 � NAF is shown. Top: representativeblots from 3 independent experiments. Bottom: normalized densitometry val-ues. p, Phosphorylated; t, total. Values are means � SE. *P � 0.05.
Fig. 3. NAF blocks the ZnT-1-induced augmentation of low-threshold calcium currents (IcaT) in Xenopus oocytes expressing CaV3.1 channels. A: representativecurrent traces from single oocytes coinjected with cRNA for CaV3.1, CaV3.1 � ZnT-1, and CaV3.1 � ZnT-1 � NAF. Selected recordings are from a similarbatch of oocytes (excised, injected, and recorded together under similar conditions). B: steady-state I-V relationship of Ba2� currents that were measured inoocytes expressing CaV3.1, CaV3.1 � ZnT-1, and CaV3.1 � ZnT-1 � NAF. Values are means � SE of the currents recorded in 3–4 independent experimentsfrom 21, 21, and 22 oocytes for each of the above conditions, respectively. C: normalized peak currents as depicted in B. Coexpression of ZnT-1 enhanced thepeak IcaT. Additional expression of NAF led to complete inhibition of the effect of ZnT-1 (P � 0.3, relative to the control oocytes expressing CaV3.1 only).D: voltage dependence of the fractional conductance (G/Gmax), derived from the data shown in A. Fitting the data with the Boltzmann equation yielded half-Gmax
voltage of �32.36 � 0.22, �39.58 � 0.38, and �35.85 � 0.45 mV, and z-effective charge of gating of 6.0 � 0.07, 8.04 � 0.1, and 6.93 � 0.09 for controls,oocytes expressing ZnT-1, and oocytes expressing CaV3.2, ZnT-1, and NAF, respectively. **P � 0.01.
C194 ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from

Lipofectamine 2000 (Invitrogen, Carlsbad, CA), according to themanufacturer’s instructions. The recombinant plasmids used for ex-pressing CaV3.1 and ZnT-1 were as follows: pEGFP-N1 [murineCaV3.1-EGFP (enhanced green fluorescent protein), a kind gift fromDr. Norbert Klugbauer], SLC30A-pJJ19 [rat ZnT-1 tagged with myc(rZnT-Myc), a kind gift from Dr. Anthony Muslin]. Cells transfectedwith pEGFP-N1 were cotransfected with either SLC30A-pJJ19 or anempty pcDNA3 vector. The total amounts of plasmids and transfec-tion reagent were similar in each treatment group. Measurements wereperformed 36–48 h after transfection. In some experiments, ERKsignaling was inhibited, by incubating the cells with the MEK-1inhibitor PD-98059 (25 M). The compound was dissolved in DMSOdiluted 1:500 and added to the culture 2 h before the experiment.DMSO alone was diluted 1:500 and added to all other wells of theseexperiments, at the same time that PD-98059 was applied.
Electrical recordings of T currents in CHO cells. Patch-clamp re-cordings were performed using whole cell configuration. Signals wereamplified using an Axopatch 200B patch-clamp amplifier (MolecularDevices, Sunnyvale, CA) sampled at 2 kHz and filtered at 800 Hz viaa four-pole Bessel low-pass filter. Data were acquired using pClamp9.2 software (Molecular Devices) and an IBM-compatible Pentium IVcomputer in conjunction with a DigiData 1322A interface (MolecularDevices). The patch pipettes were pulled from borosilicate glass(Warner Instrument) with a resistance of 3–6 M. For Ba2� currentrecordings in CHO cells, the intracellular pipette solution contained
the following (in mM): 130 KCl, 5 MgATP, 5 EGTA, 10 HEPES,adjusted with KOH to pH 7.4 (300 mosmol). The extracellularsolution contained the following (in mM): 140 NaCl, 4 KCl, 2 BaCl2,1.2 MgCl2, 11 glucose, and 5.5 HEPES, adjusted with NaOH to pH7.4 (315 mosmol). Series resistances (5–13 M) were compensated(75–90%) and periodically monitored. Barium currents in nontrans-fected CHO cells were negligible (not shown).
HL-1 cells. Cultured murine cardiac cells (HL-1 cells) were kindlyprovided by Dr. W. Claycomb (New Orleans, LA) and were main-tained as described previously (3). Briefly, cells were plated ingelatin/fibronectin-coated culture vessels and maintained in Claycombmedium (JRH Biosciences) supplemented with 10% fetal bovineserum, 0.1 mM norepinephrine, 2 mM glutamine, 100 units/ml pen-icillin-streptomycin. For induction of endogenous expression ofZnT-1 (3), HL-1 cells were incubated with 100 M Zn2� for 6 h andthen washed and kept in regular nominally zinc-free medium until theexperiment 36 h thereafter. To evaluate the effect of endothelin-1(ET-1) on surface expression of TTCC, 30 nM ET-1 were applied for2 h to HL-1 cells. Thereafter, surface biotinylation was performed asdescribed below. To verify the effect of ET-1 on ERK phosphoryla-tion, cells were exposed to ET-1 (30 nM) for 10 min, Thereafter, cellswere homogenized, and Western blot analysis was performed.
Western blot analysis. To harvest cellular proteins, cultured cellswere washed three times in phosphate-buffered saline (PBS), scrapedwith a rubber-policeman, and homogenized by sonication in 150 l
Fig. 4. NAF blocks the ZnT-1 induced augmentation of IcaT in Xenopus oocytes expressing CaV3.2 channels. A: representative current traces from single oocytescoinjected with cRNA for CaV3.2, CaV3.2 � ZnT-1, and CaV3.2 � ZnT-1 � NAF. Selected recordings are from a similar batch of oocytes (excised, injected,and recorded together under similar conditions). B: steady-state I-V relationship of Ba2� currents that were measured in oocytes expressing CaV3.2, CaV3.2 �ZnT-1, and CaV3.2 � ZnT-1 � NAF. Values are means � SE of the currents recorded in 3–4 independent experiments from 28, 25, and 24 oocytes for eachof the above conditions, respectively. C: normalized peak currents as depicted in B. Coexpression of ZnT-1 enhanced the peak IcaT in a similar manner as in Fig.1. The expression of NAF led to complete inhibition of the effect of ZnT-1 (P � 0.37, relative to the control oocytes expressing CaV3.2 only). D: voltagedependence of the fractional conductance (G/Gmax), derived from the data shown in A. Fitting the data with the Boltzmann equation yielded half-Gmax voltageof �27.48 � 0.61, �30.85 � 0.51, and �24.09 � 0.28 mV, and z-effective charge of gating of 6.197 � 0.341, 6.563 � 0.401, and 5.829 � 0.277 for controls,oocytes expressing ZnT-1, and oocytes expressing CaV3.2, ZnT-1, and NAF, respectively. **P � 0.01.
C195ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from

Choi homogenizing buffer containing the following (in mM): 20Tris·HCl, 320 sucrose, 0.2 EDTA, 0.5 EGTA, and a protease inhibitormixture (Boehringer Complete Protease Inhibitor Mixture, RocheMolecular Biochemicals). The homogenate was cleared at 14,000 gfor 15 min at 4°C, and the supernatant frozen and stored at �80°C forfuture use. Xenopus oocytes were lysed with Nonidet P-40 lysis buffer(0.5% Nonidet P-40; Igepal), containing the following (in mM): 137NaCl, 50 NaF, 5 EDTA, 10 Tris (pH 7.5), 1 NaVO3, and a proteaseinhibitor mixture as above. Lysates were cleared by low-speedcentrifugation (820 g for 15 min at 4°C), and the supernatantfrozen and stored at �80°C for future use. Protein concentrationswere determined utilizing Bradford assay (Bio-Rad). Immunoblotanalysis was performed as previously described (4, 30). Phospho-ERK-1/2 was detected by anti-phospho-ERK-1/2 (Sigma-Aldrich).Total-ERK was detected by anti-total-ERK-1/2 (Sigma-Aldrich).ZnT-1 myc expression was detected using anti-sera against ZnT-1,as previously described (4, 30).
Surface biotinylation. The surface expression of CaV3 channelswas assessed as previously described for CaV1.2 channels (30).Briefly, HEKT-293 cells were subcultured in 60-mm culture dishesachieving 50% confluence. The following day, HEKT-293 cells werecotransfected with a plasmid containing GFP-tagged CaV3.1 (pEGFP-
N1) and either ZnT-1 (SLC30A-pJJ19) or an empty vector (pcDNA3).The experiments were carried out 48 h following the transfection. Allprocedures were done on ice at 0°C. Cell monolayers were washedthree times with ice-cold PBS, incubated for 30 min at 4°C, andshaken gently, in the presence of 0.5 mg/ml of the membrane-impermeable reagent Sulfo-NHS-SS-biotin in PBS. The cells werethen washed three times with PBS supplemented with 10% BSA(Sigma) and incubated in the same solution, but without the biotin, foran additional 5 min to remove any unbound NHS-SS-biotin. Follow-ing three additional cycles of PBS washing, the cells were scraped into150-l harvest buffer [20 mM Tris pH 8.0, 137 mM NaCl, 5 mMEDTA, 5 mM EGTA, 10% glycerol, 0.5% Triton X-100, 50 mM NaF,20 mM benzamide (Sigma), and protease inhibitor cocktail (Roche)].The scraped cells were then collected into Eppendorf tubes prechilledon ice, sonicated for 10 s, and centrifuged at 5,000 g for 10 min at4°C. The supernatant was collected, and protein content was deter-mined using the Bio-Rad assay kit with bovine serum albumin as astandard. All samples were adjusted to similar protein concentration.From each sample, 15 l were retained and marked as “total protein”.Thirty microliters of Immobilized NeutrAvidin resin (Thermo Scien-tific) were added to the remaining samples, and the reaction mixturewas gently rotated end over end at 4°C for 10 h. Thereafter, resin was
Fig. 5. ZnT-1 enhances the activity of CaV3.1channels expressed in Chinese hamster ovary(CHO) cells. A: superimposed traces of ICaT inCHO cells expressing Cav3.1 without (middle)or with (right) ZnT-1. Cells were depolarizedfrom a holding potential of �90 mV. Left: thevoltage-clamp protocol. B: steady-state I-V rela-tionship of Ca2� currents. Values are means �SE of the currents recorded from 6 cells in eachgroup. C: normalized peak currents as depictedin A. Coexpression of ZnT-1 enhanced the rela-tive normalized peak current by 66.37 � 6.37%(n � 6 for both, P � 0.005). D: calculated meanGmax values for controls and ZnT-1-expressingcells (12.06 � 0.37 and 16.71 � 1.4 S, respec-tively, P � 0.01). E: voltage dependence of thefractional conductance (G/Gmax), derived fromthe data shown in A. Fitting the data with theBoltzmann equation yielded half-Gmax voltageof �37.9 � 0.46 and �42.3 � 0.45 mV (P �0.01) and z-effective charge of gating of 4.63 �0.31 and 4.38 � 0.13 (P � 0.21) for controls andZnT-1-expressing cells, respectively. Of note,error bars are included, but are too small to beseen. **P � 0.01.
C196 ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from
spun down (6,000 g, 5 min) and washed three times with ice-coldharvest buffer. The released proteins were retained, marked as “mem-brane fraction”, and stored at �80°C for further analysis. Western blotanalysis of the total protein and membrane fraction samples was doneusing anti-GFP antibody (Santa Cruz Biotechnology) for detectingGFP-tagged CaV3.1. Endogenous CaV3.1 channels in HL-1 cellswere detected using anti-CaV3.1 and antibodies (Alomone Laborato-ries). Ponceau staining was used to confirm equal protein loading.
Total internal reflection fluorescence microscopy. Analysis of totalinternal reflection fluorescence microscopy (TIRFM) was done aspreviously described (30). CHO cells grown in a glass-bottom dishwere cotransfected with CaV3.1/GFP, with or without ZnT-1. Twenty-four hours following transfection, the cells were fixed for 20 min with4% paraformaldehyde/PBS and then imaged in standard PBS solutionat room temperature. Imaging was performed on an in-house builtprism-based TIRFM system, using the 488-nm line of an argon ionlaser (Melles Griot) and imaged utilizing a SPOT charge-coupleddevice camera (Diagnostic Instruments, Sterling Heights, MI),mounted on a Zeiss Axioplan2 upright microscope with a �40 1.3numerical aperture objective (Zeiss, Jena, Germany). To quantifychanges in TIRFM of EGFP-labeled proteins, fluorescent intensitieswere normalized according to the following equation: relative fluo-rescence intensity � F(t)/F(e), where F(t) is the intensity of TIRFMmeasurements, and F(e) is the intensity of epifluorescence measure-ments. The relative fluorescence intensity was determined using theImage J software package.
Statistical analysis. Values were expressed as means � SE. Stu-dent’s t-test or one-way ANOVA were used as required to determinestatistical significance of differences between means. For results thatwere distributed in a non-Gaussian manner, the Mann-Whitney non-parametric test was used, as indicated. Statistical significance was set atP � 0.05. In Figs. 1–10, * signifies P � 0.05, and ** signifies P � 0.01.
RESULTS
ZnT-1 enhances TTCC function. Regulation of TTCC by ZnT-1was first assessed by measuring Ba2� currents in Xenopus oocytesexpressing either CaV3.1 or CaV3.2 channels (Fig. 1). Overex-
pression of ZnT-1 augmented the peak IcaT of CaV3.1 and CaV3.2expressing oocytes to 182 � 15 and 167.95 � 9.27% of control,respectively (Fig. 1, C and G), and enhanced the calculatedmaximum conductance (Gmax) of CaV3.1 and CaV3.2 expressingoocytes to 165.9 � 8.4 and 152.9 � 3.8% of control, respectively(not shown). In addition, ZnT-1 induced a small but significantshift of the half-Gmax voltage of CaV3.1 from �33.7 � 1.1 to�39.5 � 2 mV (Fig. 1D) and a similar shift of the half-Gmax
voltage of CaV3.2 from �27.97 � 0.44 to �33.48 � 0.54 mV(Fig. 1H).
ZnT-1 enhances TTCC function through activation of Ras-ERKsignaling. ZnT-1 has previously been shown to bind and activateRaf-1 kinase, leading to activation of Ras-ERK signaling (7, 23).Therefore, we investigated whether the regulatory effect of ZnT-1on CaV3 channels is mediated by the activation of this signalingpathway. For this purpose, ZnT-1 and nonactive Raf-1 (NAF)were coexpressed in Xenopus oocytes. As expected, coexpressionof NAF with ZnT-1 inhibited the ability of ZnT-1 to enhanceERK phosphorylation (Fig. 2). In addition, NAF completelyinhibited the ZnT-1-induced enhancement of IcaT in oocytesexpressing either CaV3.1 (Fig. 3) or CaV3.2 (Fig. 4). Expressionof NAF in the absence of ZnT-1 seemed to have a mild inhibitoryeffect on IcaT by itself (not shown). Taken together, the resultssupport the notion that ZnT-1 augments the activity of CaV3channels through activation of Ras-ERK signaling. It should benoted that, in a different set of experiments, we also tested theeffect of the MEK inhibitor PD-98059 on the ZnT-1-inducedenhancement of IcaT in Xenopus oocytes. Although in someexperiments the drug indeed blocked the effect of ZnT-1, unex-plained variability did not allow definite conclusion using thisdrug in Xenopus oocytes.
ZnT-1 enhances TTCC function in mammalian cells. The effectof ZnT-1 on TTCC function was further assessed in mamma-lian cells. Similar to the findings in oocytes, IcaT increased to
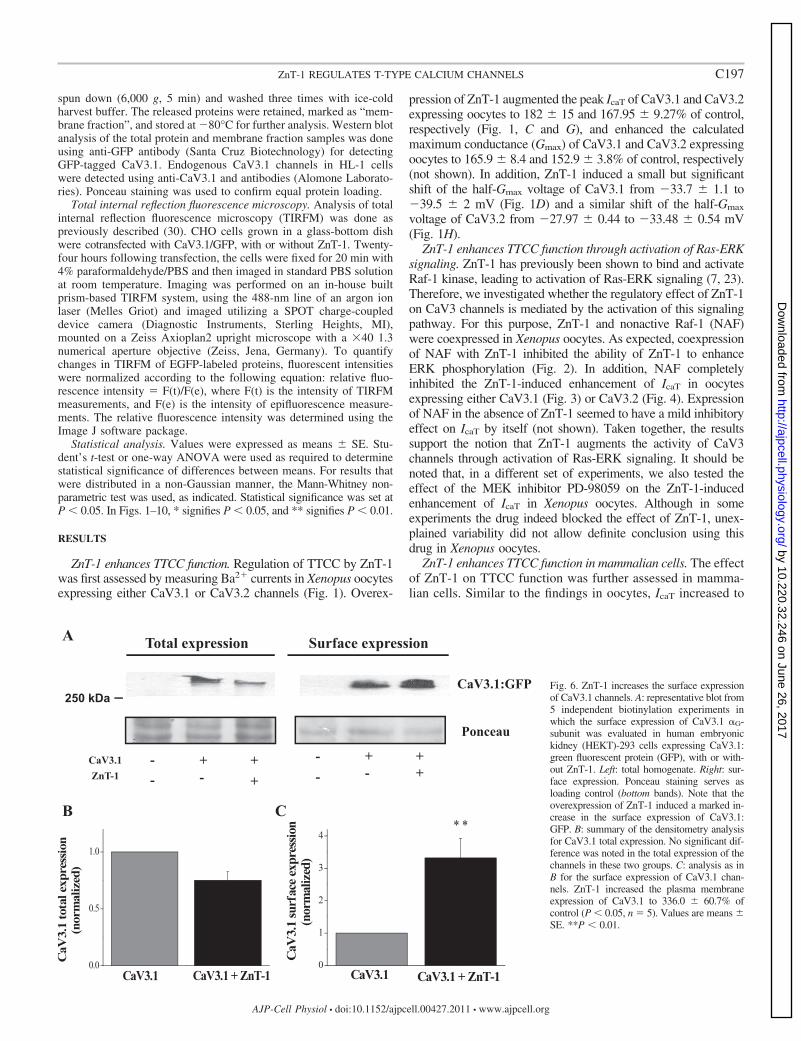
Fig. 6. ZnT-1 increases the surface expressionof CaV3.1 channels. A: representative blot from5 independent biotinylation experiments inwhich the surface expression of CaV3.1 �G-subunit was evaluated in human embryonickidney (HEKT)-293 cells expressing CaV3.1:green fluorescent protein (GFP), with or with-out ZnT-1. Left: total homogenate. Right: sur-face expression. Ponceau staining serves asloading control (bottom bands). Note that theoverexpression of ZnT-1 induced a marked in-crease in the surface expression of CaV3.1:GFP. B: summary of the densitometry analysisfor CaV3.1 total expression. No significant dif-ference was noted in the total expression of thechannels in these two groups. C: analysis as inB for the surface expression of CaV3.1 chan-nels. ZnT-1 increased the plasma membraneexpression of CaV3.1 to 336.0 � 60.7% ofcontrol (P � 0.05, n � 5). Values are means �SE. **P � 0.01.
C197ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from
166.37 � 6.37% in CHO cells coexpressing CaV3.1 andZnT-1, compared with cells expressing CaV3.1 alone (Fig. 5,A–C). ZnT-1 also enhanced the calculated Gmax to 138.55 �1.4% of control (Fig. 5D) and induced a small, but significant,shift in the half-Gmax voltage from �37.90 � 0.46 mV incontrol cells to �42.30 � 0.45 mV in cells coexpressingCaV3.1 channels and ZnT-1 (Fig. 5E).
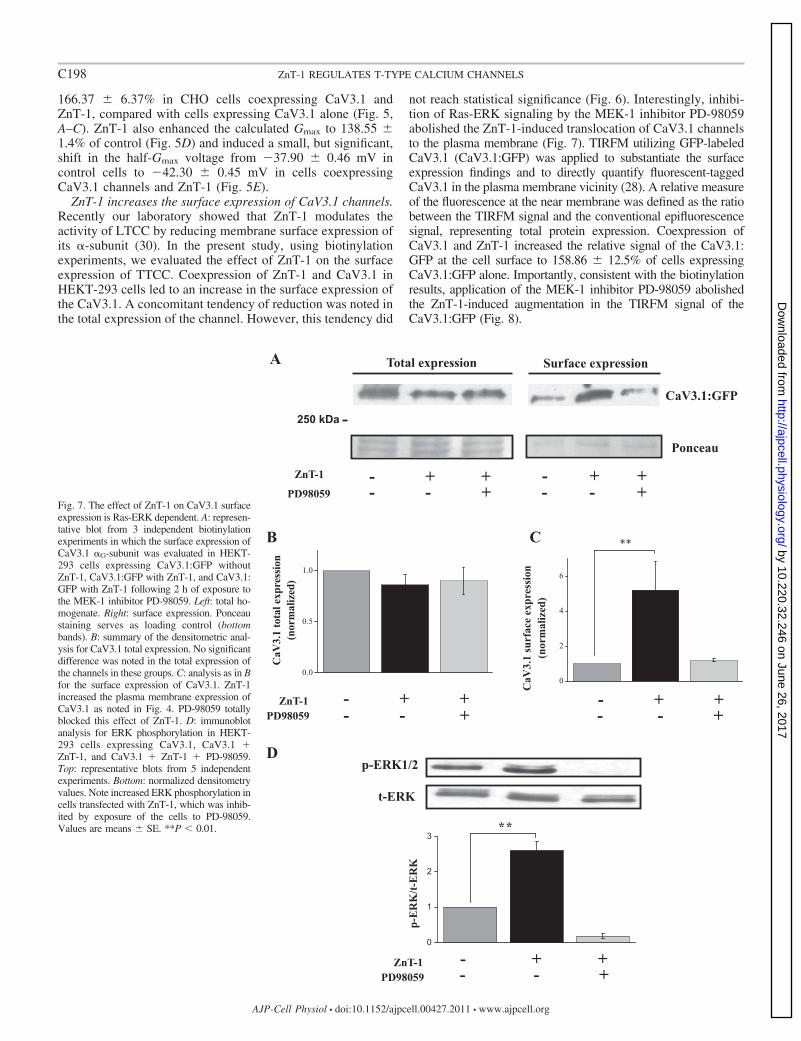
ZnT-1 increases the surface expression of CaV3.1 channels.Recently our laboratory showed that ZnT-1 modulates theactivity of LTCC by reducing membrane surface expression ofits �-subunit (30). In the present study, using biotinylationexperiments, we evaluated the effect of ZnT-1 on the surfaceexpression of TTCC. Coexpression of ZnT-1 and CaV3.1 inHEKT-293 cells led to an increase in the surface expression ofthe CaV3.1. A concomitant tendency of reduction was noted inthe total expression of the channel. However, this tendency did
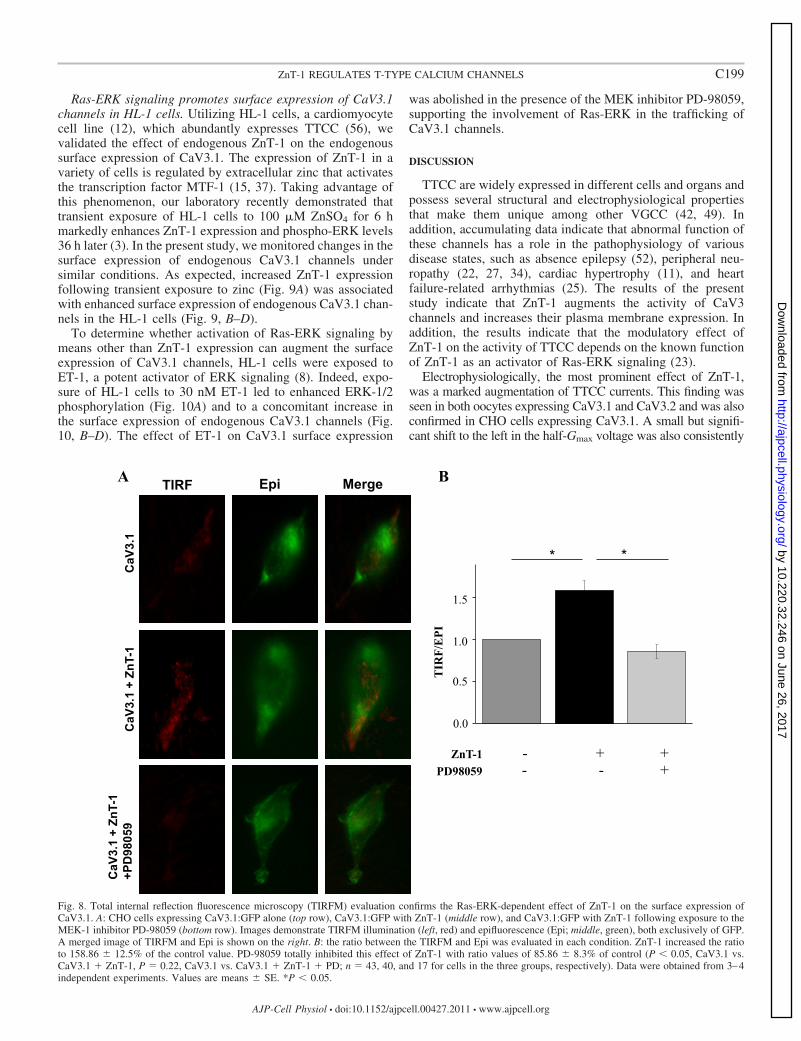
not reach statistical significance (Fig. 6). Interestingly, inhibi-tion of Ras-ERK signaling by the MEK-1 inhibitor PD-98059abolished the ZnT-1-induced translocation of CaV3.1 channelsto the plasma membrane (Fig. 7). TIRFM utilizing GFP-labeledCaV3.1 (CaV3.1:GFP) was applied to substantiate the surfaceexpression findings and to directly quantify fluorescent-taggedCaV3.1 in the plasma membrane vicinity (28). A relative measureof the fluorescence at the near membrane was defined as the ratiobetween the TIRFM signal and the conventional epifluorescencesignal, representing total protein expression. Coexpression ofCaV3.1 and ZnT-1 increased the relative signal of the CaV3.1:GFP at the cell surface to 158.86 � 12.5% of cells expressingCaV3.1:GFP alone. Importantly, consistent with the biotinylationresults, application of the MEK-1 inhibitor PD-98059 abolishedthe ZnT-1-induced augmentation in the TIRFM signal of theCaV3.1:GFP (Fig. 8).
Fig. 7. The effect of ZnT-1 on CaV3.1 surfaceexpression is Ras-ERK dependent. A: represen-tative blot from 3 independent biotinylationexperiments in which the surface expression ofCaV3.1 �G-subunit was evaluated in HEKT-293 cells expressing CaV3.1:GFP withoutZnT-1, CaV3.1:GFP with ZnT-1, and CaV3.1:GFP with ZnT-1 following 2 h of exposure tothe MEK-1 inhibitor PD-98059. Left: total ho-mogenate. Right: surface expression. Ponceaustaining serves as loading control (bottombands). B: summary of the densitometric anal-ysis for CaV3.1 total expression. No significantdifference was noted in the total expression ofthe channels in these groups. C: analysis as in Bfor the surface expression of CaV3.1. ZnT-1increased the plasma membrane expression ofCaV3.1 as noted in Fig. 4. PD-98059 totallyblocked this effect of ZnT-1. D: immunoblotanalysis for ERK phosphorylation in HEKT-293 cells expressing CaV3.1, CaV3.1 �ZnT-1, and CaV3.1 � ZnT-1 � PD-98059.Top: representative blots from 5 independentexperiments. Bottom: normalized densitometryvalues. Note increased ERK phosphorylation incells transfected with ZnT-1, which was inhib-ited by exposure of the cells to PD-98059.Values are means � SE. **P � 0.01.
C198 ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from
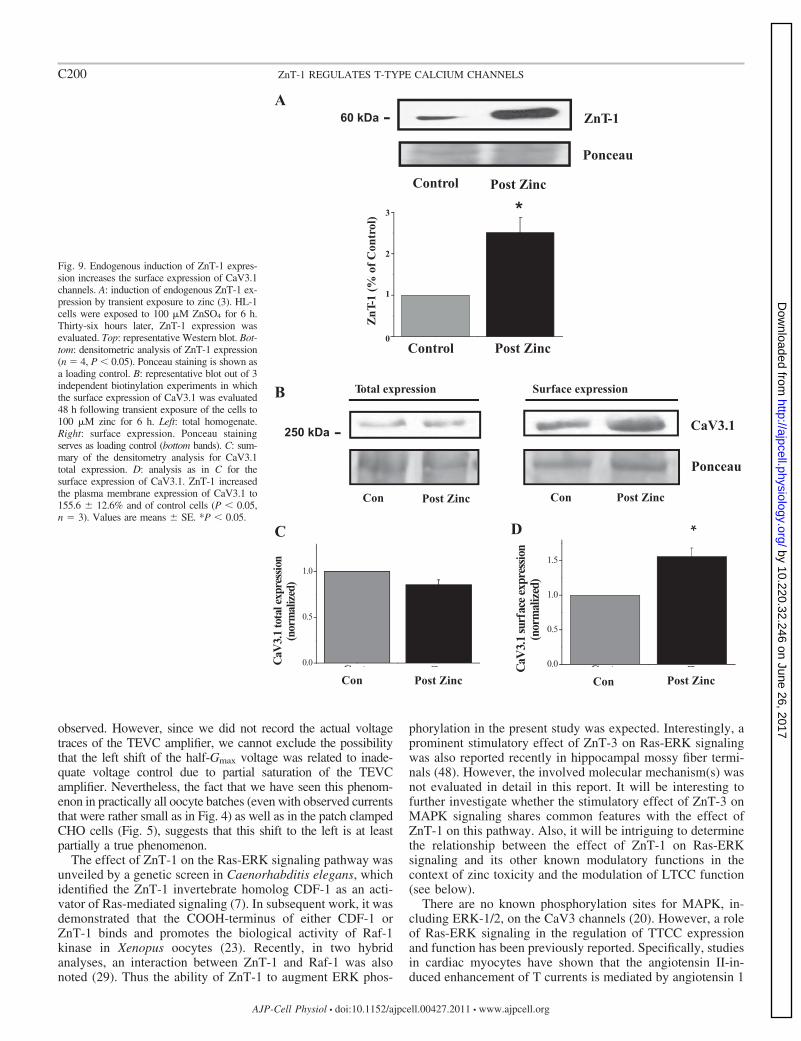
Ras-ERK signaling promotes surface expression of CaV3.1channels in HL-1 cells. Utilizing HL-1 cells, a cardiomyocytecell line (12), which abundantly expresses TTCC (56), wevalidated the effect of endogenous ZnT-1 on the endogenoussurface expression of CaV3.1. The expression of ZnT-1 in avariety of cells is regulated by extracellular zinc that activatesthe transcription factor MTF-1 (15, 37). Taking advantage ofthis phenomenon, our laboratory recently demonstrated thattransient exposure of HL-1 cells to 100 M ZnSO4 for 6 hmarkedly enhances ZnT-1 expression and phospho-ERK levels36 h later (3). In the present study, we monitored changes in thesurface expression of endogenous CaV3.1 channels undersimilar conditions. As expected, increased ZnT-1 expressionfollowing transient exposure to zinc (Fig. 9A) was associatedwith enhanced surface expression of endogenous CaV3.1 chan-nels in the HL-1 cells (Fig. 9, B–D).
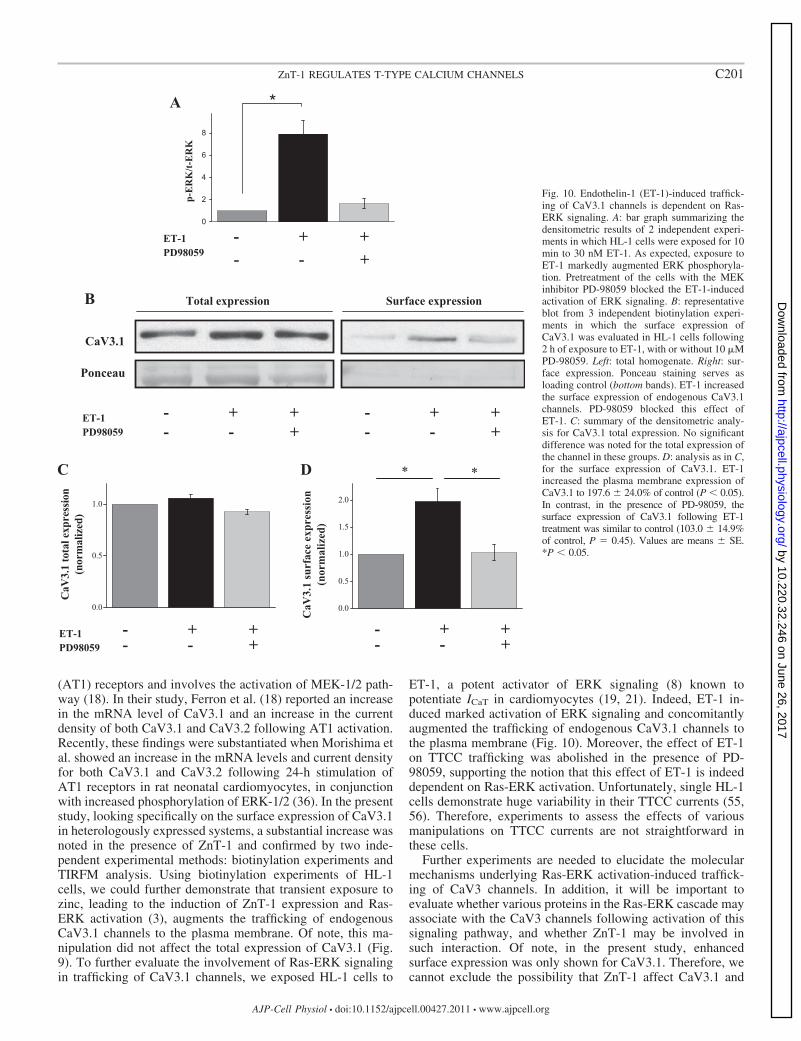
To determine whether activation of Ras-ERK signaling bymeans other than ZnT-1 expression can augment the surfaceexpression of CaV3.1 channels, HL-1 cells were exposed toET-1, a potent activator of ERK signaling (8). Indeed, expo-sure of HL-1 cells to 30 nM ET-1 led to enhanced ERK-1/2phosphorylation (Fig. 10A) and to a concomitant increase inthe surface expression of endogenous CaV3.1 channels (Fig.10, B–D). The effect of ET-1 on CaV3.1 surface expression
was abolished in the presence of the MEK inhibitor PD-98059,supporting the involvement of Ras-ERK in the trafficking ofCaV3.1 channels.
DISCUSSION
TTCC are widely expressed in different cells and organs andpossess several structural and electrophysiological propertiesthat make them unique among other VGCC (42, 49). Inaddition, accumulating data indicate that abnormal function ofthese channels has a role in the pathophysiology of variousdisease states, such as absence epilepsy (52), peripheral neu-ropathy (22, 27, 34), cardiac hypertrophy (11), and heartfailure-related arrhythmias (25). The results of the presentstudy indicate that ZnT-1 augments the activity of CaV3channels and increases their plasma membrane expression. Inaddition, the results indicate that the modulatory effect ofZnT-1 on the activity of TTCC depends on the known functionof ZnT-1 as an activator of Ras-ERK signaling (23).
Electrophysiologically, the most prominent effect of ZnT-1,was a marked augmentation of TTCC currents. This finding wasseen in both oocytes expressing CaV3.1 and CaV3.2 and was alsoconfirmed in CHO cells expressing CaV3.1. A small but signifi-cant shift to the left in the half-Gmax voltage was also consistently
Fig. 8. Total internal reflection fluorescence microscopy (TIRFM) evaluation confirms the Ras-ERK-dependent effect of ZnT-1 on the surface expression ofCaV3.1. A: CHO cells expressing CaV3.1:GFP alone (top row), CaV3.1:GFP with ZnT-1 (middle row), and CaV3.1:GFP with ZnT-1 following exposure to theMEK-1 inhibitor PD-98059 (bottom row). Images demonstrate TIRFM illumination (left, red) and epifluorescence (Epi; middle, green), both exclusively of GFP.A merged image of TIRFM and Epi is shown on the right. B: the ratio between the TIRFM and Epi was evaluated in each condition. ZnT-1 increased the ratioto 158.86 � 12.5% of the control value. PD-98059 totally inhibited this effect of ZnT-1 with ratio values of 85.86 � 8.3% of control (P � 0.05, CaV3.1 vs.CaV3.1 � ZnT-1, P � 0.22, CaV3.1 vs. CaV3.1 � ZnT-1 � PD; n � 43, 40, and 17 for cells in the three groups, respectively). Data were obtained from 3–4independent experiments. Values are means � SE. *P � 0.05.
C199ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from
observed. However, since we did not record the actual voltagetraces of the TEVC amplifier, we cannot exclude the possibilitythat the left shift of the half-Gmax voltage was related to inade-quate voltage control due to partial saturation of the TEVCamplifier. Nevertheless, the fact that we have seen this phenom-enon in practically all oocyte batches (even with observed currentsthat were rather small as in Fig. 4) as well as in the patch clampedCHO cells (Fig. 5), suggests that this shift to the left is at leastpartially a true phenomenon.
The effect of ZnT-1 on the Ras-ERK signaling pathway wasunveiled by a genetic screen in Caenorhabditis elegans, whichidentified the ZnT-1 invertebrate homolog CDF-1 as an acti-vator of Ras-mediated signaling (7). In subsequent work, it wasdemonstrated that the COOH-terminus of either CDF-1 orZnT-1 binds and promotes the biological activity of Raf-1kinase in Xenopus oocytes (23). Recently, in two hybridanalyses, an interaction between ZnT-1 and Raf-1 was alsonoted (29). Thus the ability of ZnT-1 to augment ERK phos-
phorylation in the present study was expected. Interestingly, aprominent stimulatory effect of ZnT-3 on Ras-ERK signalingwas also reported recently in hippocampal mossy fiber termi-nals (48). However, the involved molecular mechanism(s) wasnot evaluated in detail in this report. It will be interesting tofurther investigate whether the stimulatory effect of ZnT-3 onMAPK signaling shares common features with the effect ofZnT-1 on this pathway. Also, it will be intriguing to determinethe relationship between the effect of ZnT-1 on Ras-ERKsignaling and its other known modulatory functions in thecontext of zinc toxicity and the modulation of LTCC function(see below).
There are no known phosphorylation sites for MAPK, in-cluding ERK-1/2, on the CaV3 channels (20). However, a roleof Ras-ERK signaling in the regulation of TTCC expressionand function has been previously reported. Specifically, studiesin cardiac myocytes have shown that the angiotensin II-in-duced enhancement of T currents is mediated by angiotensin 1
Fig. 9. Endogenous induction of ZnT-1 expres-sion increases the surface expression of CaV3.1channels. A: induction of endogenous ZnT-1 ex-pression by transient exposure to zinc (3). HL-1cells were exposed to 100 M ZnSO4 for 6 h.Thirty-six hours later, ZnT-1 expression wasevaluated. Top: representative Western blot. Bot-tom: densitometric analysis of ZnT-1 expression(n � 4, P � 0.05). Ponceau staining is shown asa loading control. B: representative blot out of 3independent biotinylation experiments in whichthe surface expression of CaV3.1 was evaluated48 h following transient exposure of the cells to100 M zinc for 6 h. Left: total homogenate.Right: surface expression. Ponceau stainingserves as loading control (bottom bands). C: sum-mary of the densitometry analysis for CaV3.1total expression. D: analysis as in C for thesurface expression of CaV3.1. ZnT-1 increasedthe plasma membrane expression of CaV3.1 to155.6 � 12.6% and of control cells (P � 0.05,n � 3). Values are means � SE. *P � 0.05.
C200 ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from
(AT1) receptors and involves the activation of MEK-1/2 path-way (18). In their study, Ferron et al. (18) reported an increasein the mRNA level of CaV3.1 and an increase in the currentdensity of both CaV3.1 and CaV3.2 following AT1 activation.Recently, these findings were substantiated when Morishima etal. showed an increase in the mRNA levels and current densityfor both CaV3.1 and CaV3.2 following 24-h stimulation ofAT1 receptors in rat neonatal cardiomyocytes, in conjunctionwith increased phosphorylation of ERK-1/2 (36). In the presentstudy, looking specifically on the surface expression of CaV3.1in heterologously expressed systems, a substantial increase wasnoted in the presence of ZnT-1 and confirmed by two inde-pendent experimental methods: biotinylation experiments andTIRFM analysis. Using biotinylation experiments of HL-1cells, we could further demonstrate that transient exposure tozinc, leading to the induction of ZnT-1 expression and Ras-ERK activation (3), augments the trafficking of endogenousCaV3.1 channels to the plasma membrane. Of note, this ma-nipulation did not affect the total expression of CaV3.1 (Fig.9). To further evaluate the involvement of Ras-ERK signalingin trafficking of CaV3.1 channels, we exposed HL-1 cells to
ET-1, a potent activator of ERK signaling (8) known topotentiate ICaT in cardiomyocytes (19, 21). Indeed, ET-1 in-duced marked activation of ERK signaling and concomitantlyaugmented the trafficking of endogenous CaV3.1 channels tothe plasma membrane (Fig. 10). Moreover, the effect of ET-1on TTCC trafficking was abolished in the presence of PD-98059, supporting the notion that this effect of ET-1 is indeeddependent on Ras-ERK activation. Unfortunately, single HL-1cells demonstrate huge variability in their TTCC currents (55,56). Therefore, experiments to assess the effects of variousmanipulations on TTCC currents are not straightforward inthese cells.
Further experiments are needed to elucidate the molecularmechanisms underlying Ras-ERK activation-induced traffick-ing of CaV3 channels. In addition, it will be important toevaluate whether various proteins in the Ras-ERK cascade mayassociate with the CaV3 channels following activation of thissignaling pathway, and whether ZnT-1 may be involved insuch interaction. Of note, in the present study, enhancedsurface expression was only shown for CaV3.1. Therefore, wecannot exclude the possibility that ZnT-1 affect CaV3.1 and
Fig. 10. Endothelin-1 (ET-1)-induced traffick-ing of CaV3.1 channels is dependent on Ras-ERK signaling. A: bar graph summarizing thedensitometric results of 2 independent experi-ments in which HL-1 cells were exposed for 10min to 30 nM ET-1. As expected, exposure toET-1 markedly augmented ERK phosphoryla-tion. Pretreatment of the cells with the MEKinhibitor PD-98059 blocked the ET-1-inducedactivation of ERK signaling. B: representativeblot from 3 independent biotinylation experi-ments in which the surface expression ofCaV3.1 was evaluated in HL-1 cells following2 h of exposure to ET-1, with or without 10 MPD-98059. Left: total homogenate. Right: sur-face expression. Ponceau staining serves asloading control (bottom bands). ET-1 increasedthe surface expression of endogenous CaV3.1channels. PD-98059 blocked this effect ofET-1. C: summary of the densitometric analy-sis for CaV3.1 total expression. No significantdifference was noted for the total expression ofthe channel in these groups. D: analysis as in C,for the surface expression of CaV3.1. ET-1increased the plasma membrane expression ofCaV3.1 to 197.6 � 24.0% of control (P � 0.05).In contrast, in the presence of PD-98059, thesurface expression of CaV3.1 following ET-1treatment was similar to control (103.0 � 14.9%of control, P � 0.45). Values are means � SE.*P � 0.05.
C201ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from

CaV3.2 through different mechanisms, depending on the chan-nel type. Interestingly, a recent study by Dey et al. (16) foundthat the stimulatory effect of leukemia inhibitory factor onTTCC function in HEKT-293 cells was associated with in-creased membrane expression of GFP-tagged CaV3.2 channelsand was blocked by the MEK-1/2 inhibitor U-0126.
LTCC and TTCC are coexpressed in a variety of cells andorgans and appear to have differential roles in functional terms(17, 38, 45). Our laboratory’s recent reports indicate thatZnT-1 inhibits the LTCC by interaction with its �-subunit,which leads to a reduction in the surface expression of theLTCC �-subunit (30). Therefore, based on our present andprevious findings, it appears that ZnT-1 has an importantfunctional role in regulation of the delicate balance between thesurface expression and function of LTCC and TTCC. Whilethe actual domain of interaction between ZnT-1 and the �-sub-unit has not yet been identified, it will be intriguing to evaluatewhether there is a relationship between the ability of ZnT-1 tobind Raf-1 and thereby augment ICaT, and its ability to interactwith the LTCC �-subunit, leading to LTCC inhibition.
ZnT-1 is a member of a 10-gene protein family and has aputative role in zinc homeostasis. Among the ZnT proteins,ZnT-1 is the most ubiquitously expressed and is the onlyfamily member that has a well-documented ability to reducethe extracellular levels of Zn2� and thereby confer resistanceagainst zinc toxicity (13, 37, 39, 40). The expression of ZnT-1is regulated through the metal-responsive transcription factorMTF-1 (26) and is enhanced by exposure of cells to increasedlevels of Zn2� (13, 31, 33, 37). Considering the well-knownsensitivity of CaV3 channels to extracellular levels of Zn2�
(20, 51), our present data may suggest a regulatory feedbackloop in which Zn2�, which is released from presynaptic vesi-cles of glutamatergic neurons in the brain (5), inhibits the localactivity of CaV3 channels. On the other hand, the releasedZn2� also increases the local expression of ZnT-1, whichrestores the activity of CaV3 channels by activation of Ras-ERK signaling.
In summary, the present study elucidates a modulatory effectof ZnT-1 on the activity of CaV3.1 and CaV3.2 channels. Thestimulatory effect of ZnT-1 on TTCC function is in contrast tothe previously described role of ZnT-1 as an inhibitor of LTCCfunction. We have shown that the effect of ZnT-1 on TTCCresults from its known biological activity as an activator ofRas-ERK signaling and involves enhanced trafficking ofCaV3.1 channels to the plasma membrane. The above findingsare likely to bear important functional implications due to theinvolvement of TTCC in a variety of biological processes.
ACKNOWLEDGMENTS
The authors thank Dr. Nathan Dascal for technical assistance and forimportant scientific comments.
GRANTS
This work was supported by Israel Science Foundation Grant 992/07 (A.Moran and Y. Etzion) and Deutsche Forschungsgemeinschaf Grant MO1932/1-1 (A. Moran).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
M.M., O.B., A.K., A.M., and Y.E. conception and design of research;M.M., O.B., S.L., J.K., S.D., D.B., and A.P. performed experiments; M.M.,
O.B., S.L., J.K., S.D., D.B., L.A.G., and Y.E. analyzed data; M.M., O.B., J.K.,A.M., and Y.E. interpreted results of experiments; M.M., S.D., A.P., and Y.E.prepared figures; M.M., O.B., A.K., A.M., and Y.E. edited and revisedmanuscript; M.M., O.B., S.L., J.K., S.D., D.B., L.A.G., A.P., A.K., A.M., andY.E. approved final version of manuscript; O.B., L.A.G., A.M., and Y.E.drafted manuscript.
REFERENCES
1. Arias JM, Murbartian J, Vitko I, Lee JH, Perez-Reyes E. Transfer ofbeta subunit regulation from high to low voltage-gated Ca2� channels.FEBS Lett 579: 3907–3912, 2005.
2. Baumgart JP, Vitko I, Bidaud I, Kondratskyi A, Lory P, Perez-ReyesE. I-II loop structural determinants in the gating and surface expression oflow voltage-activated calcium channels. PLos One 3: e2976, 2008.
3. Beharier O, Dror S, Levy S, Kahn J, Mor M, Etzion S, Gitler D, KatzA, Muslin AJ, Moran A, Etzion Y. ZnT-1 protects HL-1 cells fromsimulated ischemia-reperfusion through activation of Ras-ERK signaling.J Mol Med (Berl) 90: 127–138, 2012.
4. Beharier O, Etzion Y, Katz A, Friedman H, Tenbosh N, Zacharish S,Bereza S, Goshen U, Moran A. Crosstalk between L-type calciumchannels and ZnT-1, a new player in rate-dependent cardiac electricalremodeling. Cell Calcium 42: 71–82, 2007.
5. Bitanihirwe BK, Cunningham MG. Zinc: the brain’s dark horse. Syn-apse 63: 1029–1049, 2009.
6. Bourinet E, Alloui A, Monteil A, Barrere C, Couette B, Poirot O,Pages A, McRory J, Snutch TP, Eschalier A, Nargeot J. Silencing ofthe Cav3.2 T-type calcium channel gene in sensory neurons demonstratesits major role in nociception. EMBO J 24: 315–324, 2005.
7. Bruinsma JJ, Jirakulaporn T, Muslin AJ, Kornfeld K. Zinc ions andcation diffusion facilitator proteins regulate Ras-mediated signaling. DevCell 2: 567–578, 2002.
8. Cadwallader K, Beltman J, McCormick F, Cook S. Differential regu-lation of extracellular signal-regulated protein kinase 1 and Jun N-terminalkinase 1 by Ca2� and protein kinase C in endothelin-stimulated Rat-1cells. Biochem J 321: 795–804, 1997.
9. Chemin J, Nargeot J, Lory P. Chemical determinants involved inanandamide-induced inhibition of T-type calcium channels. J Biol Chem282: 2314–2323, 2007.
10. Chemin J, Traboulsie A, Lory P. Molecular pathways underlying themodulation of T-type calcium channels by neurotransmitters and hor-mones. Cell Calcium 40: 121–134, 2006.
11. Chiang CS, Huang CH, Chieng H, Chang YT, Chang D, Chen JJ,Chen YC, Chen YH, Shin HS, Campbell KP, Chen CC. The Ca(v)3.2T-type Ca(2�) channel is required for pressure overload-induced cardiachypertrophy in mice. Circ Res 104: 522–530, 2009.
12. Claycomb WC, Lanson NA Jr, Stallworth BS, Egeland DB, DelcarpioJB, Bahinski A, Izzo NJ Jr. HL-1 cells: a cardiac muscle cell line thatcontracts and retains phenotypic characteristics of the adult cardiomyo-cyte. Proc Natl Acad Sci U S A 95: 2979–2984, 1998.
13. Cousins RJ, Liuzzi JP, Lichten LA. Mammalian zinc transport, traffick-ing, and signals. J Biol Chem 281: 24085–24089, 2006.
14. Cousins RJ, McMahon RJ. Integrative aspects of zinc transporters. JNutr 130: 1384S–1387S, 2000.
15. Devergnas S, Chimienti F, Naud N, Pennequin A, Coquerel Y,Chantegrel J, Favier A, Seve M. Differential regulation of zinc effluxtransporters ZnT-1, ZnT-5 and ZnT-7 gene expression by zinc levels: areal-time RT-PCR study. Biochem Pharmacol 68: 699–709, 2004.
16. Dey D, Shepherd A, Pachuau J, Martin-Caraballo M. Leukemiainhibitory factor regulates trafficking of T-type Ca2� channels. Am JPhysiol Cell Physiol 300: C576–C587, 2011.
17. Diana MA, Otsu Y, Maton G, Collin T, Chat M, Dieudonne S. T-typeand L-type Ca2� conductances define and encode the bimodal firingpattern of vestibulocerebellar unipolar brush cells. J Neurosci 27: 3823–3838, 2007.
18. Ferron L, Capuano V, Ruchon Y, Deroubaix E, Coulombe A, RenaudJF. Angiotensin II signaling pathways mediate expression of cardiacT-type calcium channels. Circ Res 93: 1241–1248, 2003.
19. Furukawa T, Ito H, Nitta J, Tsujino M, Adachi S, Hiroe M, MarumoF, Sawanobori T, Hiraoka M. Endothelin-1 enhances calcium entrythrough T-type calcium channels in cultured neonatal rat ventricularmyocytes. Circ Res 71: 1242–1253, 1992.
20. Huc S, Monteil A, Bidaud I, Barbara G, Chemin J, Lory P. Regulationof T-type calcium channels: signalling pathways and functional implica-tions. Biochim Biophys Acta 1793: 947–952, 2009.
C202 ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from

21. Izumi T, Kihara Y, Sarai N, Yoneda T, Iwanaga Y, Inagaki K,Onozawa Y, Takenaka H, Kita T, Noma A. Reinduction of T-typecalcium channels by endothelin-1 in failing hearts in vivo and in adult ratventricular myocytes in vitro. Circulation 108: 2530–2535, 2003.
22. Jagodic MM, Pathirathna S, Nelson MT, Mancuso S, Joksovic PM,Rosenberg ER, Bayliss DA, Jevtovic-Todorovic V, Todorovic SM.Cell-specific alterations of T-type calcium current in painful diabeticneuropathy enhance excitability of sensory neurons. J Neurosci 27: 3305–3316, 2007.
23. Jirakulaporn T, Muslin AJ. Cation diffusion facilitator proteins modu-late Raf-1 activity. J Biol Chem 279: 27807–27815, 2004.
24. Kim D, Park D, Choi S, Lee S, Sun M, Kim C, Shin HS. Thalamiccontrol of visceral nociception mediated by T-type Ca2� channels. Science302: 117–119, 2003.
25. Kinoshita H, Kuwahara K, Takano M, Arai Y, Kuwabara Y, YasunoS, Nakagawa Y, Nakanishi M, Harada M, Fujiwara M, Murakami M,Ueshima K, Nakao K. T-type Ca2� channel blockade prevents suddendeath in mice with heart failure. Circulation 120: 743–752, 2009.
26. Langmade SJ, Ravindra R, Daniels PJ, Andrews GK. The transcriptionfactor MTF-1 mediates metal regulation of the mouse ZnT1 gene. J BiolChem 275: 34803–34809, 2000.
27. Latham JR, Pathirathna S, Jagodic MM, Choe WJ, Levin ME, NelsonMT, Lee WY, Krishnan K, Covey DF, Todorovic SM, Jevtovic-Todorovic V. Selective T-type calcium channel blockade alleviates hy-peralgesia in ob/ob mice. Diabetes 58: 2656–2665, 2009.
28. Lavi Y, Edidin MA, Gheber LA. Dynamic patches of membrane pro-teins. Biophys J 93: L35–L37, 2007.
29. Lazarczyk M, Pons C, Mendoza JA, Cassonnet P, Jacob Y, Favre M.Regulation of cellular zinc balance as a potential mechanism of EVER-mediated protection against pathogenesis by cutaneous oncogenic humanpapillomaviruses. J Exp Med 205: 35–42, 2008.
30. Levy S, Beharier O, Etzion Y, Mor M, Buzaglo L, Shaltiel L, GheberLA, Kahn J, Muslin AJ, Katz A, Gitler D, Moran A. Molecular basisfor zinc transporter 1 action as an endogenous inhibitor of L-type calciumchannels. J Biol Chem 284: 32434–32443, 2009.
31. Liuzzi JP, Blanchard RK, Cousins RJ. Differential regulation of zinctransporter 1, 2, and 4 mRNA expression by dietary zinc in rats. J Nutr131: 46–52, 2001.
32. Lory P, Bidaud I, Chemin J. T-type calcium channels in differentiationand proliferation. Cell Calcium 40: 135–146, 2006.
33. McMahon RJ, Cousins RJ. Mammalian zinc transporters. J Nutr 128:667–670, 1998.
34. Messinger RB, Naik AK, Jagodic MM, Nelson MT, Lee WY, ChoeWJ, Orestes P, Latham JR, Todorovic SM, Jevtovic-Todorovic V. Invivo silencing of the Ca(V)32 T-type calcium channels in sensory neuronsalleviates hyperalgesia in rats with streptozocin-induced diabetic neurop-athy. Pain 145: 184–195, 2009.
35. Mikami A, Imoto K, Tanabe T, Niidome T, Mori Y, Takeshima H,Narumiya S, Numa S. Primary structure and functional expression of thecardiac dihydropyridine-sensitive calcium channel. Nature 340: 230–233,1989.
36. Morishima M, Wang Y, Akiyoshi Y, Miyamoto S, Ono K. Telmisartan,an angiotensin II type 1 receptor antagonist, attenuates T-type Ca2�
channel expression in neonatal rat cardiomyocytes. Eur J Pharmacol 609:105–112, 2009.
37. Nolte C, Gore A, Sekler I, Kresse W, Hershfinkel M, Hoffmann A,Kettenmann H, Moran A. ZnT-1 expression in astroglial cells protects
against zinc toxicity and slows the accumulation of intracellular zinc. Glia48: 145–155, 2004.
38. Ono K, Iijima T. Cardiac T-type Ca(2�) channels in the heart. J Mol CellCardiol 48: 65–70, 2010.
39. Palmiter R, Findley S. Cloning and functional characterization of amammalian zinc transporter that confers resistance to zinc. EMBO J 14:639–649, 1995.
40. Palmiter RD, Huang L. Efflux and compartmentalization of zinc bymembers of the SLC30 family of solute carriers. Pflügers Arch 447:744–751, 2004.
41. Perez-Reyes E. G protein-mediated inhibition of Cav3.2 T-type channelsrevisited. Mol Pharmacol 77: 136–138, 2010.
42. Perez-Reyes E. Molecular physiology of low-voltage-activated T-typecalcium channels. Physiol Rev 83: 117–161, 2003.
43. Perez-Reyes E, Lee JH, Cribbs LL. Molecular characterization of twomembers of the T-type calcium channel family. Ann N Y Acad Sci 868:131–143, 1999.
44. Perez-Reyes E, Lory P. Molecular biology of T-type calcium channels.CNS Neurol Disord Drug Targets 5: 605–609, 2006.
45. Satin LS. Localized calcium influx in pancreatic beta-cells: its signifi-cance for Ca2�-dependent insulin secretion from the islets of Langerhans.Endocrine 13: 251–262, 2000.
46. Segal D, Ohana E, Besser L, Hershfinkel M, Moran A, Sekler I. A rolefor ZnT-1 in regulating cellular cation influx. Biochem Biophys ResCommun 323: 1145–1150, 2004.
47. Shcheglovitov A, Vitko I, Bidaud I, Baumgart JP, Navarro-GonzalezMF, Grayson TH, Lory P, Hill CE, Perez-Reyes E. Alternative splicingwithin the I-II loop controls surface expression of T-type Ca(v)31 calciumchannels. FEBS Lett 582: 3765–3770, 2008.
48. Sindreu C, Palmiter RD, Storm DR. Zinc transporter ZnT-3 regulatespresynaptic Erk1/2 signaling and hippocampus-dependent memory. ProcNatl Acad Sci U S A 108: 3366–3370, 2011.
49. Talavera K, Nilius B. Biophysics and structure-function relationship ofT-type Ca2� channels. Cell Calcium 40: 97–114, 2006.
50. Talavera K, Staes M, Janssens A, Droogmans G, Nilius B. Mechanismof arachidonic acid modulation of the T-type Ca2� channel �1G. J GenPhysiol 124: 225–238, 2004.
51. Traboulsie A, Chemin J, Chevalier M, Quignard JF, Nargeot J, LoryP. Subunit-specific modulation of T-type calcium channels by zinc. JPhysiol 578: 159–171, 2007.
52. Tsakiridou E, Bertollini L, de Curtis M, Avanzini G, Pape HC.Selective increase in T-type calcium conductance of reticular thalamicneurons in a rat model of absence epilepsy. J Neurosci 15: 3110–3117,1995.
53. Tsuda M, Imaizumi K, Katayama T, Kitagawa K, Wanaka A, To-hyama M, Takagi T. Expression of zinc transporter gene, ZnT-1, isinduced after transient forebrain ischemia in the gerbil. J Neurosci 17:6678–6684, 1997.
54. Vitko I, Bidaud I, Arias JM, Mezghrani A, Lory P, Perez-Reyes E. TheI-II loop controls plasma membrane expression and gating of Cav32T-type Ca2� channels: a paradigm for childhood absence epilepsy muta-tions. J Neurosci 27: 322–330, 2007.
55. Xia M, Salata JJ, Figueroa DJ, Lawlor AM, Liang HA, Liu Y,Connolly TM. Functional expression of L- and T-type Ca2� channels inmurine HL-1 cells. J Mol Cell Cardiol 36: 111–119, 2004.
56. Yang Z, Shen W, Rottman JN, Wikswo JP, Murray KT. Rapidstimulation causes electrical remodeling in cultured atrial myocytes. J MolCell Cardiol 38: 299–308, 2005.
C203ZnT-1 REGULATES T-TYPE CALCIUM CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00427.2011 • www.ajpcell.org
by 10.220.32.246 on June 26, 2017http://ajpcell.physiology.org/
Dow
nloaded from