- of perishable foods
TRANSCRIPT

305.
M I C R O B I A L GROWTH A N D M E A T Q U A L I T Y M I C R O B I A L A N T A G O N I ' S M I N C O N T R O L L I N G GROWTH*
P A T R I C I A A K R A B A W I A N D H . P . R I E M A N N U N I V E R S I T Y O F C A L I F O R N I A , D A V I S
I. Introduction
A. Microbiological c h a r a c t e r i s t i c s of perishable foods - Most mildly preserved, moist foods contain i n i t i a l l y saprophytic
organisms i n numbers t h a t exceed the number of food poisoning organisms which may be present. The saprophytes w i l l i n general a l s o outnumber t h e hazardous microorganisms i n stored foods unless t h e storage conditions or t h e composition of the food i s highly se lec t ive f o r food poisoning species. A s examples can be mentioned t h e rap id and of ten se lec t ive growth of C . perfringens i n cooked food t h a t i s kept at a temperature i n t h e upper-range of t h e growth i n t e r v a l f o r t h i s organism. Staph. aureus may grow p a r t i a l l y s e l e c t i v e l y when foods with moderate concentrations of salt and l o w i n acid a re kept a t room temperature or w a r m e r . t h a t one or several groups of saprophytic organisms present w i l l grow as fast or f a s t e r than the food poisoning species and t h i s can have one of several consequences.
However, it w i l l general ly be found
1. The saprophytes have no or l i t t l e e f f e c t on t h e mul t ip l ica t ion of food poisoning strains--but may produce s igns of spoilage and thus warn the consumer. This seems t o be a common s i t u a t i o n with many moist, un- preserved foods ( r a w meat, f i s h ) and some mildly preserved moist foods.
2. The saprophytes may enhance the growth of food poisoning organisms. A f e w examples are known and they w i l l be discussed b r i e f l y below.
3. The saprophytes i n h i b i t or slow down t h e growth of food poisoning organisms. This general ly require a c e r t a i n composition of t h e food (presence of fermentable carbohydrates, s m a l l amounts of N a C 1 as a preservat ive) , t h e presence of c e r t a i n groups of a c t i v e l y competing saprophytes and storage of t h e food within c e r t a i n tenperature ranges. These conditions preva i l i n t h e preservation of foods (sauerkraut, fermented sausages, e tc . ) by fermentation. However, the safe ty of many f oods--including perishable processed mests--which are not in ten t iona l ly fermented r e s t s on exac t ly the same pr inc ip les . This w i l l be discussed i n some d e t a i l after a review of the l i t e r a t u r e dealing with microbial competition and synergism.
B. Microbial competition i n foods - L i t e r a t u r e -- Effec t of saprophytic organisms on various food poisoning bac ter ia .
* Acknowledgement This study w a s supported by the Agricul tural Resemch Service, U. S. Department of Agriculture, under Grant No. 12-14-100-9184( 73) drnin- i s t e r e d by t h e Eastern U t i l i z a t i o n Research and Development Division, 600 E a s t Mermaid Lane, Philadelphia, Pennsylvania 19118 and by a grant from t h e American Meat I n s t i t u t e Foundation.

306.
1. C. botulinum - A synerg is t ic e f f e c t of l a c t i c acid b a c t e r i a has been observed (Benjamin e t al. 1956) and may be due t o a reduction of the oxidation- reduction p o t e n t i a l or production of growth f a c t o r s . A s i m i l a r e f f e c t has been observed with s t r a i n s such as C . sporosphaeroides and - B. p o l p y x a which faci l i ta ted C. b o t u l i & m type C tox in production i n laboratory media (Bulatova e t :.-It i s also known t h a t the n a t u r a l f l o r a , e .g. Pseudomonas species (Valenzuela e t a. 1967), enables - C . botulinum type E t o grow on t h e surface of f i s h (exposed t o atmos- pheric oxygen). - C . botulinwn i n f r u i t ju ices apparently by oxidizing organic acids with a resulting decrease i n a c i d i t y t o around pH 5 (Meyer and Gunnison, 1929, Tanner e t al. 1940).
--
-- --
Fungi (yeast , mold) have been found t o induce growth of
-- Examples of growth i n h i b i t i o n of C . botulinum are more numerous.
Saprophytes i n r a w , f rozen vegetables can increase the a c i d i t y during defrost ing t o pH5 or less. A similar s i t u a t i o n i s found i n some pre- cooked frozen foods such as chicken a l a king. acid b a c t e r i a resu l ted i n an increase i n a c i d i t y t o pH l e v e l s of 3.5-4.4 during defrost ing and no botulinum t o x i n w a s formed i n inoculated packs ( S d e h and Ordal, 1955a and b) .
Addition of l a c t i c
C. botulinum grow slowly at pH 5.0 and not a t a l l a t pH 4.5 and it seems t h a t a decrease i n pH i s t h e main f a c t o r i n "natural" inh ib i t ion . Bacteria which s p o i l c r e a m s t y l e corn without a decrease i n pH d id not prevent t o x i n production by type A or B ( M d i n and Greenberg, 1964) and we have similar experiences with formation of type A and E t o x i n i n spoi l ing m e a t . However, it has been reported t h a t Streptococcus lactis and Lactobaci l lus vir idescans i n h i b i t or delay t o x i n production by type E i n r a w f i s h where only very l imi ted fermentation can be expected (Valenzuela e t al. 1967). since it i s known t h a t c e r t a i n s t r a i n s of S t rep t . l a c t i s produce n i s i n . Brevibacterium l i n e n s which grow i n c e r t a i n types of cheese can produce mt ib io t icum active against C . botulinum (Greez, e t al. 1959a,b; 1961, Greez, 1964).
The i n h i b i t i o n may be due t o a n t i b i o t i c s --
L- -
Some C . botulinum s t r a i n s produce a n t i b a c t e r i a l agents, bacter iocins , ac t ive against other C . botulinum s t r a i n s . duced by C. sporogenes and there aze cross reac t ions between t h i s species and C . botulinum with regard t o bac ter ioc in s e n s i t i v i t y (Beerens and Tahon, 19677.
Bacteriocins are a l s o pro-
A bac ter ioc in which i s very ac t ive against C. botulinum E i s produced by some non-toxic botulinum E l i k e orgaks-er, 1964). The importance of bacter iocins i n food i s unknown but it has been found t h a t i s o l a t i o n of C. botulinum from natural. sources can be d i f f i c u l t when bac ter ioc in producing organisms are present; t h e d i f f i c u l t y can be over- come by t r y p s i n addi t ion (Harmon and Kautter, 1967).
Various b a c t e r i d proteases can reduce or destroy t h e t o x i c i t y of butulinum toxin.

2. - C . perfringens
This species seem to be as sens i t ive t o low pH as i s C . botulinum. It has, however, t h e advantzge of being able to l rul t iply very rap id ly a t a high temperature (46OC) and nay therefore under conditions that would i n h i b i t C . botulinum be able t o outgrow acid producing organisms. Also, C . perfr ingens produce bacter iocins (Welchicins) some of which are ac t ive agzinst a broad spectrum of spore formers (Sss,wrnan m d Antohi, 1967).
-
3. Staphylococcus aureus and Salmonella
Staph. aureus and Salmonella are strongly repressed i n foods t h a t ferment e a s i l y such as chicken, turkey and beef p ies . Complete repres- s ion w a s found even when t h e pathogens i n i t i a l l y outnumbered the sapro- phytes hundredfold (Dack and Lippitz, 1962). and-cheese dinner, no repression w a s observed (Petersen, e t al. 1962a.) However Staph. aureus was outgrown by saprophytes i n r z w crab neat (Stabyj -- e t al. -and many d i f f e r e n t organisms compete successful ly with S t a h. aweus d though moderate amounts of salt tend t o faxror the l a t t e r Peterson e t al. 1962b, 1964; Graves znd Fraz ie r , 1963). The mechanism of inhibition--when drop i n pH i s not involved--is unknown.
I n another food, macaroni- --
le --
Staph. aureus i s of ten s t rongly repressed by l a c t i c acid b a c t e r i a i n mildly cured meats such as bacon and bologna sausage (Eddy and Ingram, 1962; Ingram, 1960, 1962; Tonge e t al. 1964; Christiansen and Foster, 1965). confined to t h e surface (Casman e t ail. 1963).
-- When enterotoxin i s produced i n meat the production seems mainly
-- 11. Experimental Studies
A. I n h i b i t i o n of - C . botulinum by curing f a c t o r s alone.
The br ine concentration and pH required f o r i n h i b i t i o n of C . botulinum i n mildly processed meats are wel l known. The results we havz obtained with inoculated packs containing very few saprophytes -e s ta ted i n Table 1 and they agree f a i r l y wel l with the results published by others .
B. I n h i b i t i o n of - C. botulinum i n products with subinhibitory l e v e l s of salt.
It i s known t h a t a l a r g e proportion of t h e mildly processed meat products which are on the market today do not contain l e v e l s of brine concentration and pH s u f f i c i e n t f o r a complete i n h i b i t i o n of C . botulinum (Riemann, 1968). One could therefore expect t h z t t o x i n production would o c c u frequent ly when perishable cured market meats are inoculated with botulinum spores and incubated at elevated temperatures. This, however, i s co t t h e case. Out of 1 2 1 f r e s h l y vacuum packed symples of m e a t products picked up i n super- markets and inoculated with botulinum spores only 11 became toxic .

308.
Table 1. Levels of curing f a c t o r s required -ko prevent formation of botulinum tox in i n meat contain- i n g low numbers of saprophytes.
Brine Type Temperature concentration PH - - E 10' - 30' C 3.3 6.5
A 25' - 30' C 6.7 6.5
Table 2. Composition of per ishable m e a t products which became tox ic after inoculat ion with C . botulinum type A spores and incubation a t 20-3@C! f o r 2-4 days.
Type of products Brine concentration pH after incubation
Vacuum packed s l i ced ham
3.2 - 4.8% Higher than 5.6 Sl iced vacuum packed smoked chicken and turkey

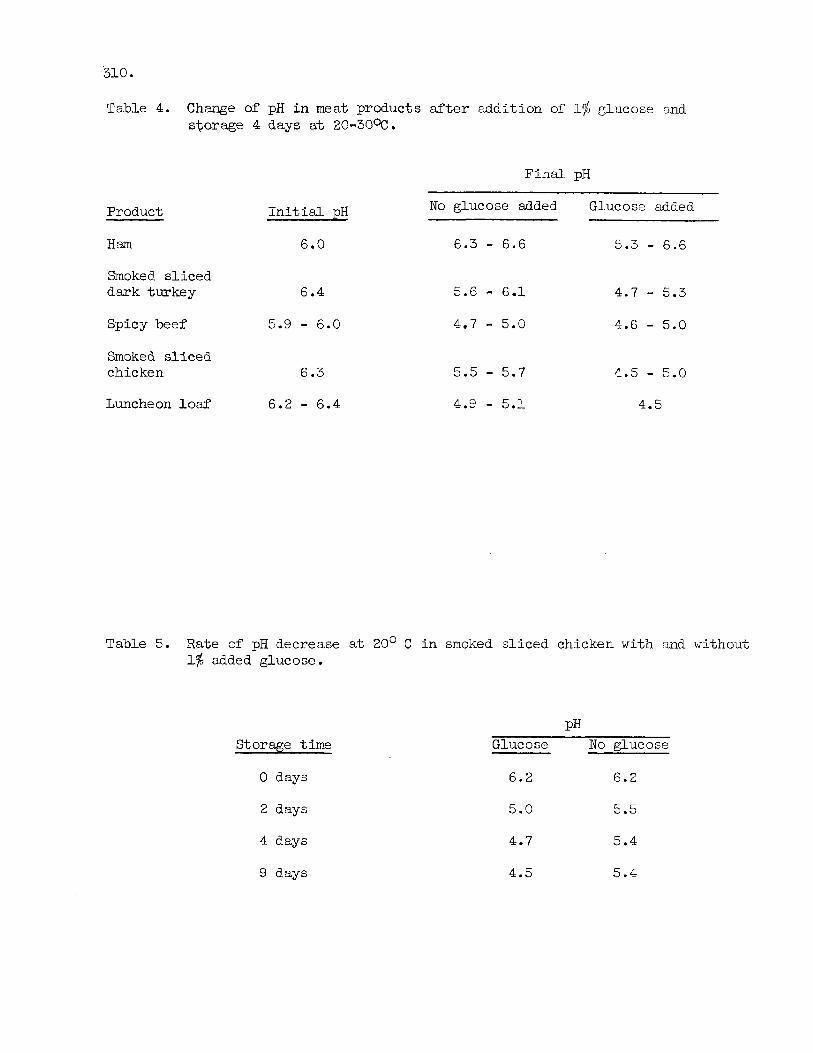
(Table 2) and these 11 were c h a r a c t e r i s t i c i n having pH V'JllL:; : J ~ o v ~ : 5.6 after incubation f o r several days at 2O-3O0C. pH values. Typical examples are shown i n Table 3. The ztudy showr;d t h a t many perishable meat products undergo acid fermentation when exposed t o .an elevated storage temperature. The fermentation must be due t o rdded carbohydrate since meat i tself does not contain enough carbohydrate t o permit fermentation t o proceed t o such low pH values. The e f f e c t of added carbohydrates was confirmed i n experiments where glucose w a s added t o d i f fe ren t m e a t products and pH measured before and after incubation (Table 4 ) . rapid (Table 5). (Table 4). at supermarkets (24-48 hours after packaging) and analyzed f o r compositlon including content of glucose (measured by T e s tape) and t h e r e s u l t s a e presented i n Table 6. The results suggest t h a t addi t ion of glucose i s de te r - mined not only by type of product but also a c h a r a c t e r i s t i c of t h e manufacturer. Some manufacturers apparently add glucose up t o 2% or more t o nost of t h e i r products and t h i s gives t h e i r products an addi t ional b u i l t - i n safe ty f a c t o r .
A l l the non-toxic prodilct:; had lower
The drop i n pH can be qui te However, addi t ion of glucose did not i n v a i a b l y cause pH drop
A number of m e a t products were sampled a t t h e time they arr ived
It i s of i n t e r e s t t o know what kind of organisms are responsible f o r the acid formation i n glucose containing perishable meat products. The following s tudies were car r ied out t o obtain information on t h i s point .
Table 3. Composition and change i n pH i n some perishable meat products d>Jring storage at 2O-3O0C f o r 2-4 days. addi t ion of botulinum spores are marked *.
Samples which became toxic d t e r
Product Percent br ine
* H a
*Sliced dark turkey
Spicy beef
*Smoked s l i c e d chicken
Spiced luncheon loaf
Salami
Pepper loaf
Bologna
Sliced Beef
4.5 - 4.8
2.6 - 4.1 4.6 - 6.5 2.9 - 3.4
3.2 - 3.6 4.5
3.6 - 4.5 3.6 - 5.3
5.2
Before storage
6.0
6.2 - 6.5 5.9 - 6.0
6.3
6.2
4.9
5.4
5.2 - 6.3
5.9
After storage
6.2 - 6.6
5.2 - 6.7 4.8 - 5.0
5.7
4.5 - 5.0 4.6 - 4.7 4.4 - 4.8 4.4 - 4.7 5.1 - 5.4
310.
Table 4. Change of p H i n meat products after addi t ion of' 1% glucose and storage 4 days at 20-30%.
Fina l pH
Product I n i t i a l pH
Ham 6.0
Smoked sliced dark turkey 6.4
Spicy beef 5.9 - 6.0
Smoked s l iced chicken 6.3
Luncheon loaf 6.2 - 6.4
~~~-
No glucose added Glucose added
6.3 - 6.6 5.3 - 6.6
5.6 - 6.1 4.7 - 5.3 4.7 - 5.0 4.6 - 5.0
5.5 - 5.7 4.5 - 5.0 4.9 - 5.1 4.5
Table 5. R a t e of p H decrease a t 20' C i n smoked s l iced chicken with and without 1% added glucose.
Storage t i m e
0 days
2 days
4 days
9 days
PH Glucose No glucose
6.2 6.2
5.0 5.5
4.7 5.4
4.5 5.4

511.
Table 6. Concentration of glucose i n f r e s h l y packed (24-48 hours under r e f r i g e r a t i o n ) perishable meat products. 24- means higher than 2%
Manufacturer Product Percent glue ose
A Hot dog
B
C
F
G
H
Pork sausage Pork sausage with sage
A l l beef bologna Pepper loaf Cooked ham
Smoked turkey Smoked ham Smoked chicken
Chopped pork Beef sdami Bologna Spiced luncheon Liver loaf Smoked beef Cooked ham Beef loaf Chopped ham Franks All beef f ranks
Cooked ham Ham l o a f Picnic ham
103s
Summer sausage Pressed pork
Corned Beef Smoked turkey Smoked dark turkey Smoked chicken Pastrami Sliced beef Sliced ham All beef salami
2+
0.1 - 0.25 2l-
0.1 0 0 - 0.1 0.1 0.1 0.1
2+ 0.25 - 2 2+ 0 - 2 0.1 - 0.25 0 0 - 0.1 0.25 - 0.5 0 2+ 2+
0.1 - 0.25 0.1 - 0.25 0.1
0.5 - 2.0 0 - 0.1 0 - 0.1 0 - 0.1 0 - 0.1 0 - 0.5 0.1 - 0.25 0.1 - 0.25 0.1 0.25

312.
Table 6 - continued
Manufacturer
I
J
K
L
M
Spiced lunch loaf Cooked ham Bonele s s ham Chopped ham Gel l ied cooked beef Beef loaf Beef bologna Smoked beef loa f Liver sausage
Sl iced chicken
Liver sausage Pork sausages Cot to salami Bologna Chopped ham Old fashion loaf Ham salad spread Fresh sausage
Spiced loaf Bologna Cooked ham Braunschwe ige r
Turkey s l i c e s Beef s l i c e s Corned beef s l i c e s Smoked ham Cooked ham Chopped ham Liver sausage Sliced ham Ham slices
Percent glucose
0 .25 - 0.5 0 - 0.1 0 2+ 0 0.1 2+ 0.1 2+
0
2-F 0.5 2+ 2+ 2+ 2-F 24- 2+
0.25 2+ 2+ 2+
0 - 2.0 0.1 - 0.25 0.1 0.1 0 - 2.0 0 - 2.0 2+ 0.1 0 - 0.1

313.
C . Ef fec t of saprophytes and glucose on toxigenesis by C . botulinum. - Samples of vacuum packed bologna and s l i ced dark tilrkcy wcre
inoculated with 1000 spopes of Clostridium botulinum type A."The incuba- t i o n periods var ied from 1 t o 1 6 days at both ZOOC and 3OoC. M t c r incubation, t he m e a t samples were examined f o r t he growth of organisms other than C . botulinum. -
All meat samples where C . botulinum did not grow were streaked on All Purpose Medium (APT) plus-O.~-m c reso l green. made t o d i s t ingu i sh colonies which produced acid, but almost all of t he colonies f e l l i n t o t h i s category. Therefore, several organisms were picked a t random from each meat samples. Seventeen s t r a i n s of acid- producing cocci were i so la ted , several of which were s imi la r .
Attempts were
A spec ia l mixture of blood and most of t h e cons t i tuents of stock APT agar w a s then formulated. Difco A€" agar contains Tween 80 and when mixed with blood t h i s compound prevented t h e c h a r a c t e r i s t i c and d i s t i n c - t i v e hemolysis of C . botulinum. When t h e acid-producing cocci and C . botulinum were both grown on p l a i n APT agar, t he re were no d i s t i n c t i v e f e a t u r e s t o allow t h e i r separation, and several. of t h e cocci s t r a i n s were not able t o grow on p l a i n blood agar. Therefore, APT agar w a s mixed with- out Tween 80 and then 5% cow blood w a s added f o r assays of cocci-botulinum growth .
-
The cocci s t r a i n s were divided i n t o two groups on t h e b a s i s of ca ta lase a c t i v i t y . Seven s t r a i n s of ca t a l a se pos i t ive and t e n s t r a i n s of ca t a l a se negative cocci were t e s t e d .
A f a c t o r i a l expe r imnt w a s then conducted t o determine t h e inh ib i to ry e f f e c t , i f any, which these cocci had on the growth of - C . botulinum type A. The following f a c t o r s were examined: 1% and 3% NaC1, pH 6 and 7, 0 and 100 ppm n i t r i t e , 0.1% and 1.0% glucose, incubation temperatures of 25OC and 35%, and two d i l u t i o n s of cocci. varying amounts of salt, n i t r i t e , and glucose, and pH w a s lowered t o 6 by adding 0.75% glucono d e l t a lactone. they w e r e t o be used and then s tored overnight at 10°C.
APT aga.r plus 5$ cow blood was mixed with
The p l a t e s were poured one day before
One mi l l ion spores of C. botulinum type A were evenly spread on the APT-BA p l a t e s . 10-1 and p l a t e s which had been seeded with spores. The p l a t e s were immediately placed i n anaerobic jars and dupl ica tes were incubated at 25OC and 35OC f o r t h ree days.
S & S f i l t e r p a p e r d i s c s 6 mm i n diameter were dipped i n d i l u t i o n s of t h e var ious s t r a i n s of cocci and placed on the
After incubation, t h e p l a t e s were examined and zones where C . botulinum w a s unable t o grow were measured around t h e f i l t e r paper d l s c s . The s i ze of t h e zones of i nh ib i t i on var ied depending on t h e s t r a i n used and t h e f a c t o r s involved. glucose and n i t r i t e increased t h e inh ib i to ry e f f e c t s of t he cocci . Certain s t r a i n s of both ca t a l a se negative Streptococcus and cz t a l a se pos i t ive Staphylococcus were able t o take advantage of these favorable high concen- t r a t i o n s to i n h i b i t t he growth of C. botulinum. The temperature of incLtbn.- t i o n and the d i l u t i o n of t h e inh ib i to ry organisms had no s ign i f i can t
Low i n i t i d pH and high concentrations of NaC1,

314.
e f f e c t s on botulinum growth. f i n a l l y iden t i f i ed as Strep. faecium seemed t o be s l i g h t l y more inh ib i tory than the o thers .
One s t r a i n of ca t a l a se negative cocci
It was la te r used i n inh ib i t i on s tud ie s i n ground beef.
Fresh ground round w a s divided i n t o t e n gram port ions. H a l f of t h e samples were "cooked" by autoclaving f o r 15 minutes before they were placed i n s m a l l p l a s t i c p e t r i d i shes and the other ha l f of t h e meat samples were used r a w . 100, 10,000, and 1,000,000 spores of C . botulinum type A and loo, 10-2, and d i l u t i o n s of 16 hours ATP cu l tu re s of t h e most inh ib i tory s t r a i n of Strep. faecium were used t o inoculate t h e meat.
The e f f e c t s of t he following f a c t o r s have been explored: Glucose, NaC1, incubation temperature. The s tud ie s are being continued using complete f a c t o r i a l designs.
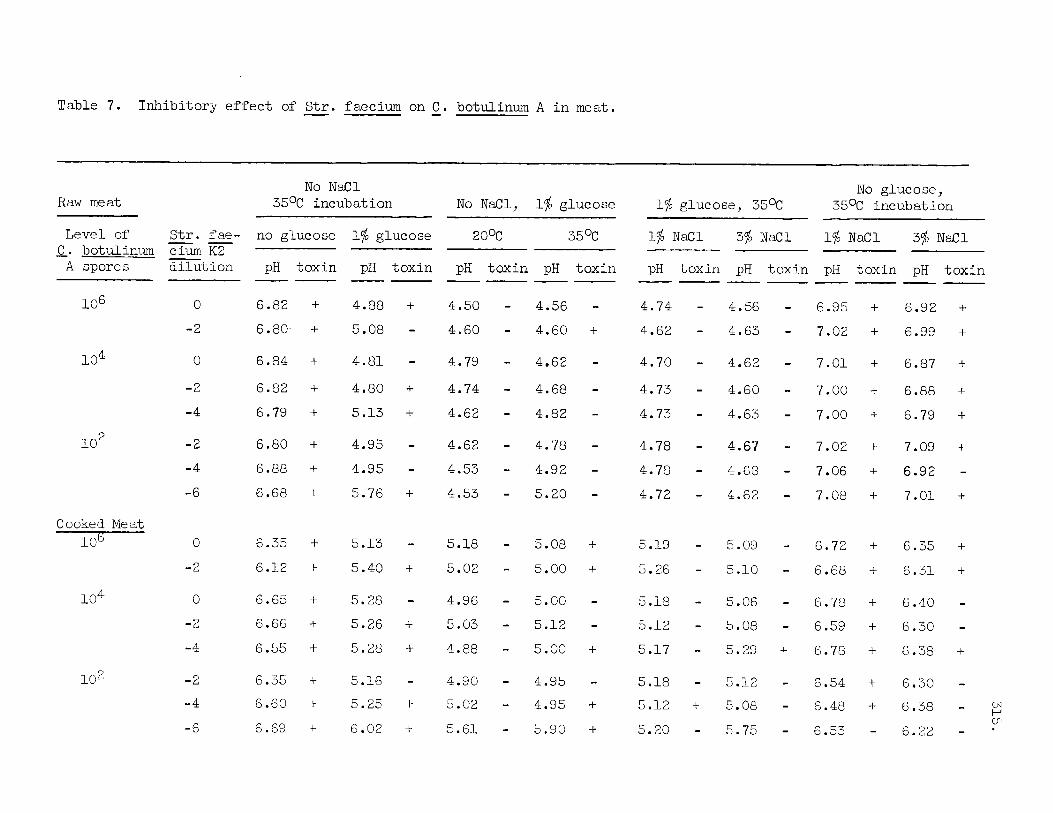
Table 7 i l l u s t r a t e s t h e inh ib i tory e f f e c t s of Strep. faecium on C . botulinum 62A. added t o the meat except f o r a minor inh ib i t i on i n r a w m e a t when 3% NaCl i s added. f e s t e spec ia l ly when 1 og 3% N a C l i s a l so added. The inh ib i tory e f f e c t i s more pronounced i n r a w than i n cooked meat; t h e l a t t e r having a s l i g h t l y higher pH value. The inh ib i tory e f f e c t seems a l s o more pronounced a t ZO°C compared t o 35OC.
There i s no e f f e c t of Strep. faecium when no glucose i:
I n the presence of 1% glucose the inh ib i tory e f f e c t becomes mani-
D. I d e n t i f i c a t i o n of i nh ib i to ry saprophytes.
Streptococci .
The ca t a l a se negative cocci i so l a t ed occurred mainly i n p a i r s but were also found i n s m a l l chains. They showed no pigmentation and, accord- i n g t o Bergey's Manual, they are more l i k e Streptococcus f a e c a l i s than any other group.
Fur ther t e s t s of Sharpe and Fryer (1966) suggested t h a t t h e organ- Growth a t 5OoC i s a f ea tu re i s m s are va r i an t s of Streptococcus faecium.
used t o d i f f e r e n t i a t e between S. faecium and t h e other enterococci. However, t h i s c h a r a c t e r i s t i c may be l o s t on repeated subculturing (Sharpe and Fryer, 1966).
Strep. f a e c a l i s can reduce te t razol ium t o the insoluble red formazan so t h a t t h e colonies have dark red center . Strep. faecium i s not able t o reduce te t razol ium, (Barnes, 1956), and t h e s t r a i n s i so l a t ed from bologna were white on the te t razol ium glucose agar.
Other cha rac t e r i s t i c s , such as t h e a b i l i t y t o ferment melibiose and mannitol, d i f f e r from t h e proposed r e s u l t s . Suff ice it t o say t h a t the cocc i i so l a t ed are more l i k e Strep. faecium than they are l i k e S t rep , f a e c a l i s . Thei r c h a r a c t e r i s t i c s are shown i n Table 8.
315.
++
cu
m
Wa
?
?
++
Lncu
ab
?
?
II
Wn
L
oa
dd
.
.
II
*c
u
PW
dd
.
.
I+
a0
m
a
dd
.
.
II
00
-+
*
??
+I
coco
QO
*m
.
.
++
NO
C
OC
O
WW
.
.
ON
I
a 0
rl
+I
+
+ P
a3
W
+ rl 0
F
I
cu a
* I
0
F
d
I
cu W
d
I
0,
F
d I
ri
03
d
+ d
00
W
0
* 0 4
++
a
m
co
b
am
.
.
++
00
00
bP
.
.
II
OM
W
a
d-
+
..
I
I
mm
F
b
**
.
.
II
QC
U
ma
d
*
..
I
I
*N
b
W
*d
.
.
++
O
M
*L
O
?t
++
c
um
c
op
W
W
..
cu
d
II
++
ln
ri
am
? "!
++
c
um
F
W
WW
.
.
II
F;
a
rJ
II
ma
mL
o
e?
++
c
oo
00
LO
m
..
I
I
QC
U
rl
o
mu
)
..
I+
MO
r
ld
m
m
..
++
mc
u
Wa
'?e
ON
I
Il
l
mm
ri
o
m0
.
..
b
aP
++
+
++
+ +
+ I
Na
m
000
..
.
am
a,
P
LO
F
..
.
aW
a .
..
a
ac
o
PF
P
Il
l
Il
l
r-
co
cu
a
Wa
a
a,
m
00
cu
l
nm
m
..
. cu
a, U
I r
io
r-
LoLoLo
..
.
..
.
*d
d
Il
l
Il
l
co
a,
N
FF
b
dd
d
..
.
Il
l
I+
+
LD
mm
Il
l
11
1
Il
l
WM
03
m
oa
,
0N
r-
l
mo
a
..
.
dl
nd
.
..
*
U)
m
I+
+
..
.
*d
m
21 k
0
++
+
++
+ +
++
0
03
03
c
oc
oa
.
..
L
oo
m
ma
a
4
u
a, 4-1 4-1 a,
..
.
Wa
a
WW
W
Nd
W
Il
l
c\! 0
rl cu 0
rl

316.
Table 8. Charac t e r i s t i c s of s t reptococci i so l a t ed from meats.
S t r a i n s
m H1 - - K 2 - Cat a lase
Hemolysis
Aerobic acid production from:
Arabinose
Glyc e r ol
Gluc o se
Lactose
Mannitol
M e 1 i b i o se
R a f f inose
Salacin
Sucrose
Sorbi t ol
Trehalose
Glycerol, anaerobic acid product ion
L i t m u s milk
Gelat i n hydrolysis
Starch hydrolysis
Growth a t lo0 C
Growth at 45O C
Growth at 50° C
Growth a t pH 9 .6
none (gamma) none none
+
- no change
-
- no change
-
- no change
- -
-k
+ + +

317.
Table 8 - continued
Growth i n 0.1% methylene blue milk
Growth on 40% b i l e i n blood agax
Growth i n 12% NaCl
Growth i n 15% N a C l
Te t r a z oliwn reduct ion
Source
+ 1- +
+ + +
bologna, bologna bologna,
1 day a t 3 O°C 3OoC 3OoC
incubated 1 day, 4 day,
Staphylococci.
U s i n g the c l a s s i f i c a t i o n of Baird-Parker (1963, 1966), t h e ca ta lase pos i t ive cocci i so la ted belong t o t h e Staphylococcus subgroups I1 and V which correspond t o Staph. epidermidis i n Bergey' s Manual (Baird-Parker, 1962) . These cocci are found mainly i n p a i r s but also occur singly, i n short chains or i n groups.
The cocci a re c l a s s i f i e d Staphylococci as opposed t o Micrococci because of t h e i r a b i l i t y t o produce acid from glucose under anaerobic conditions (Baird-Parker, 1965) . Their c h a r a c t e r i s t i c s are shown i n Table 9 .
111. Discussion.
It seems Clem- t h a t perishable meat products with t h e i r content of saprophytic microorganisms will have a b u i l t - i n s a f e t y f a c t o r against C . botulinum and other food poisoning b a c t e r i a if glucose (1% or more) is- added. Glucose i s fermented t o acid by several saprophytes and the r e s u l t i n g low pH i n h i b i t s t o x i n formation by C. botulinum. t r a t i o n s of NaC1, even 1% seem t o support the-inhibitory e f f e c t . observations explain why a v a r i e t y of market meats with subinhibitory l e v e l s of N a C l and pH are not botulinogenic. Fecal streptococci, espec ia l ly S t r . fecium, and staphylococci a re of importance i n i n h i b i t i n g C. botulinum i n glucose containing meats and most if not d l of t h e i r inh ix i tory e f f e c t i s i s due t o acid formation.
Small concen- These
-

318.
Table 9 . Cha rac t e r i s t i c s of staphylococci i so l a t ed from meats.
S t r a i n s
Pigment Coagulase production Catalase production Phosphatase production VP t e s t , ace to in production Litmus milk
Aerobic acid production from:
Arab i no se Dextrin G a lac t o se G 1 yc e r ol Glucose Lactose Maltose Mannitol
Glucose, anaerobic acid
Hippurate hydrolysis G e l at i n hydroly s i s Urea u t i l i z a t i o n Nitrate reduct ion Growth i n 15% N a C l Growth i n 12% N a C l Growth at 10% Growth at 45% Growth at pH 9.6 Growth i n 0.1% methylene
Growth on 40% b i l e i n
product ion
blue milk
blood agar
Baird-Parker c l a s s i f i c a t i o n
Source
yellow - + + + acid, c oagul at i on
+ + - + + + + + + + + + ' Subgroup I1
turkey, incubated 1 day a t 30%
c 1 white
-
+ - + acid, coagulation, p a r t i a l d iges t ion
+ + - + + i + + + + + +
Subgroup V
turkey,
30oC 8 day,
N 2
white
-
- + - + acid
+ Subgroup V
bologna,
30% 1 day,

319.
It i s obvious t h a t these organisms must multiply t o form la rge populations i n order t o produce acid enough t o i n h i b i t C . botulinum and although t h e i r presence i s t o be preferred t o the presence-of botulinum toxin many people w i l l probably question the d e s i r a b i l i t y of l a r g e populations of - S t r . fecium and Staphylococci i n foods. It would therefore seem appropriate t o search for other acid producing less objectionable microorganisms t h a t could be added t o perishable meat products and other perishable foods as a safeguard against - C . botulinum and other food poisoning bac ter ia .
References
1. Baird-Parker, A. C . (1962) J. Appl. Bact. 25(3) 352. - - - 2. Baird-Parker, A. C . (1963) J. Gen. Microbiol. - 30, 409.
3. Baird-Parker, A. C . (1965) - - J. Gen. Microbiol. - 38, 363.
4. Baird-Parker, A. C . (1966) &I "Ident i f ica t ion Methods f o r Microbiologists,
- -
Par t A" (B. M. Gibbs and F , A. Skinner, eds.) pp. 59-63. Academic Press.
4a. Barnes, E . M. (1956) J. B a c t . 19(2) , 193.
5. Beerens, H., and Tahon, M. M. (1967) I n "Botulism 1966" (M. Ingram and
- - -
T. A. Roberts, eds.) , pp. 424-428. Chazan and H a l l , London.
6. Benjamin, M. J. W., Wheather, D. M., and Shepherd, P. A. (1956) J. Appl. Bac ter io l . 19, 159. - -
7. Bergeys Manual, 7th Ed.
8. Bulatova, T. I., Matveev, K. I., and Samsonova, V . S. (1967) I n "Botulism 1966" (M. Ingram and T. A. Roberts, eds . ) , pp. 3 9 1 - 3 9 r and H a , London.
Chapman
9. Casman, E. P., McCoy, D. W., and Brandley, P. J. (1963) - Appl. Microbiol. ll. 498.
10. Christiansen, L. M., and Foster, E. M. (1965). Appl. Microbiol. _. 13, 1023.
ll. Dack, G . M., and Lippi tz , G. (1962) Appl. Microbiol. - 10, 472.
1 2 . Eddy, B. P., and Ingram, M. (1962) -- J. Appl. Bacter iol . - 25, 237 .
13. Graves, R . R., and Fraz ier , W. C. (1963). Appl. Microbiol. - 11, 513.
14. Grecz, N. (1964) I n "Microbial Inh ib i tors i n Foods" (P?. Molin, e d . ) , pp. 307-320. Almqzst and Wiksell, Stockholm.
15. Grecz, N., Wagenaar, R. O., and Dack, G. M. (1959a). - Appl. Microbiol. 1, 33.

320.
1 6 . Grecz, N., Wagenaar, R. O., and Dack, G. M. (1959b). Appl. Microbiol. 7, 228. -
1 7 . Grecz, N., Dack, G. M., and Hedrick, L. R. (1961) J. Food Sei. 26, 72. - _L- - 18. Grecz, N., Upadhyay, T., and Tang, T. C . (1967) - - Can. J. Microbiol.
13, 287.
19. Harmon, S. M., and Kautter, D. A. (1967) Bacter io l . - Proc. p. 5.
20. Ingram, M. (1960) - J. Appl. Bacter iol . - 23, 206.
21. Ingram, M. (1962) - J. Appl. Bac ter io l . - 25, 259.
22. Kautter, D. A. (1964) - J. Food Sci. - 29, 843.
23. Malin, B., and Greenberg, R . A. (1964) I n "Microbial Inh ib i to r s i n Food" (N. Molin, ed.) , Almqvist and Wikseii, Stockholm.
24. Meyer, K. F., and Gunnison, J. B. (1929), - J. In fec t . Diseases, - 45, 135.
25. Peterson, A. C., Black, J. J., and Gunderson, M. F. (1962a) Appl. Microbiol. - 10, 16.
26. Peterson, A. C., Black, J. J., and Gunderson, M. F. (1962b) Appl. Microbiol. - 10, 23.
27. Peterson, A. C. , Black, J. J., and Gunderson, M. F. (1964) Appl. Microbiol. 1 2 , 70.
L
28. Pivnick, H., and Barnett , H. (1965) Food Technol. - 19, 1164.
29, Riemann, H. (1968) Western Meat Packer
30. Saleh, M. A., and Ordal, Z. J. (1955a) __.- Food Res. - 20, 332.
31. Saleh, M. A., and Ordal, Z. J. (195513) Food -- R e s . - 20, 340,
32. Sasasman, A., and Antohi, M. (1967) Proceedings, Workshop on Anaerobic Bacteria. Montreal, Canada.
I n s t i t u t de Microbiologie e t d'Hygiene de L 'UniversitC de
33. Sharpe, M. El isabeth, and Fryer, T. F. (1966) - I n " Iden t i f i ca t ion Methods f o r Microbiologists, P u t A" (B. M. Gibbs and F. A. Skinner, eds . ) , pp. 65-68. Academic Press.
34. Stabyj, B. M., Dollar, A. M., and Liston, J. (1965). J. Food -- Sei . - 30, 344.
35. Tanner, F. W., Beamer, P. R., and Rickher, C . J. (1940) -- Food Res. - 5, 323.
36. Tonge, R. J. Baird-Parker, A. C . and Covett, J. J. (1964) - - J. Appl. Bacterial. - 27, 252.
37. Valenzuela, S., Niekerson, J. T. R., Campbell, C. , and Goldblith, S . A. (1967) Chapman and H d l , London.
I n "Botulism 1966" (M. Ingram and T . A. Roberts, eds.) , pp. 224-235.

321.
RIEMANN: Paper In .
BERNHOLM': Thank you very much D r . Rierzann. Movlng on to osu" next top ic , our speaker w i l l be Vincent DelGiudice, Manager, Sani ta t ion, Microbiology and Chemical Department, Qua l i ty Assurance Division, Amour and Company, Chicago. M r . DelGiudice has a M.S. from Northwestern Universizy i n Bacteriology and has been with Armour f o r some 27 years, spending most of h i s time i n applied san i t a t ion for at least t h e last 25 years. He w i l i t s l k t o us today about "Losses t o the Meat Industry Due t o Microbial Growth.ff
# # # i f # # # # # # #