1 mechanisms of resistance x efflux inactivation decreased permeability altered target antibiotics...
TRANSCRIPT

1
Mechanisms of Resistance
XEfflux
Inactivation
Decreasedpermeability
Altered target
Antibiotics exert selective pressure that favors emergence of resistant organisms
Bacteria employ several biochemical strategies to become resistant

2
Genetic Basis of Resistance Spontaneous mutations in endogenous genes
– Structural genes: expanded spectrum of enzymatic activity, target site modification, transport defect
– Regulatory genes: increased expression
Acquisition of exogenous sequences– Usually genes that encode inactivating enzymes or
modified targets, regulatory genes– Mechanisms of DNA transfer: conjugation (cell-cell
contact); transformation (uptake of DNA in solution); transduction (transfer of DNA in bacteriophages)
Expression of resistance genes– Reversible induction/repression systems can affect
resistance phenotypes

3
Spread of Resistance Genes
R
S R
R S
R
Conjugation
Transformation
R

4
Major Classes of Antibiotics
Mechanism of action Major resistance mechanisms
Beta-lactams Inactivate PBPs (peptidoglycan synthesis)
•Beta-lactamases•Low affinity PBPs•Decreased transport
Glycopeptides Bind to precursor of peptidoglycan
•Modification of precursor
Aminoglycosides Inhibit protein synthesis (bind to 30S subunit)
•Modifying enzymes (add adenyl or PO4)
Macrolides Inhibit protein synthesis (bind to 50S subunit)
•Methylation of rRNA•Efflux pumps
Quinolones Inhibit topoisomerases (DNA synthesis)
•Altered target enzyme•Efflux pumps

5
Antibiotic Susceptibility Tests
Minimal inhibitory concentration (MIC)– Reference method. Add standard inoculum to dilutions of
antibiotic. Incubate overnight. MIC is lowest concentration that inhibits growth (can also be performed by agar dilution).
– Interpretation (S or R) is based on achievable drug levels
104 cfu
4 2 1 0.5 0.25 0.12 0g/ml

6
Antibiotic Susceptibility Tests
Kirby-Bauer agar disk diffusion– Paper disk containing antibiotic
is placed on lawn of bacteria, then incubated overnight. Diameter of zone of inhibition is inversely related to MIC (used to establish interpretive breakpoints).
– Standardized for commonly isolated, rapidly growing organisms.

7
Antibiotic Susceptibility Tests
E-test– Strips containing a gradient of
antibiotic are placed on lawn of bacteria and incubated overnight. MIC is determined at point where zone of inhibition intersects scale on strip.
– Combines ease of KB with an MIC method. Particularly useful for S. pneumoniae.

8
-lactam Antibiotics
Monobactams
– Substrate analogs of D-Ala-D-Ala– Covalently bind to PBPs, inhibit final step of
peptidoglycan synthesis
Penicillins
NO
S
Carbapenems
NO
NO
NO
S Cephalosporins
1st gen: GPC, some GNR
2nd gen: some GNR +anaerobes
3rd gen: many GNR, GPC

9
Structure of Peptidoglycan
|L-Ala |D-Glu |L-diA |D-Ala |D-Ala
NAG-NAM-NAG-NAM
-(AA)n-NH2
|L-Ala |D-Glu |L-diA |D-Ala |D-Ala
NAG-NAM-NAG-NAM
-(AA)n-NH2
cytoplasm
Transpeptidation reaction

10
Penicillin-Binding Proteins (PBPs)
Membrane bound enzymes– Catalyze final steps of peptidoglycan synthesis
(transglycosylation and transpeptidation)– Multiple essential PBPs (4-5) - involved in cell
elongation, determination of cell shape, and cell division; essential for cell viability
-lactams acylate active site serine of PBPs, inhibit transpeptidation– Activity determined by affinity for PBPs, stability
against -lactamases, and permeability– Autolysins contribute to bactericidal activity

11
Penicillin-Resistant S. pneumoniae (PRSP)
S. pneumoniae interpretative breakpoints– penicillin susceptible (MIC 0.0625 µg/ml),
intermediate (0.125 -1.0), resistant ( 2.0).
High-level penicillin resistance has risen rapidly in US (0.01% in 1987 to 3% in 1994)– 20-30% of isolates may be non-susceptible (I or R).– High-level PRSP may exhibit cross-resistance to
3rd generation cephalosporins– Serious problem when infection occurs at body
sites where antibiotic availability is limited.
PRSP may be multi-resistant (macrolides, TMP/SXT); strains can spread widely

12
Mosaic PBP Genes in PRSP Resistance is due to alterations in endogenous
PBPs– Resistant PBP genes exhibit 20-30% divergence from
sensitive isolates (Science 1994;264:388-393)
– DNA from related streptococci taken up and incorporated into S. pneumoniae genes
Czechoslovakia (1987)
USA (1983)
South Africa (1978)
S SXN
= pen-sensitive S. pneumoniae = Streptococcus ?
PBP 2B

13
International Spread of PRSP
Multiresistant PRSP in Iceland (JID 1993;168:158-63)
– First isolate in 12/88; 17% PRSP in 1992. • Almost 70% of PRSP were serotype 6B; resistant to tet,
chloram, erythro, and TMP/SXT; similar or identical molecular markers.
• Icelandic PRSP identical to multiresistant 6B clone endemic in Spain (popular vacation site).
Possible factors responsible for rapid spread– -lactam use in Iceland low, but high use of
TMP/SXT, tet, etc may have selected for multiresistant clone.
– 57% of population lives in Reykjavik/suburbs, almost 80% of children age 2-6 attend day-care centers.

14
-lactam resistance in Staph. aureus
>90% of strains produce -lactamase– plasmid encoded, confers resistance to penicillin,
ampicillin– these strains are susceptible to penicillinase-
resistant penicillins (e.g. methicillin), 1st generation cephalosporins, and -lactam/-lactamase inhibitor combinations
At many large medical centers, approx 30% of S. aureus are resistant to methicillin and other -lactams

15
Methicillin-resistant S. aureus (MRSA)
MRSA contain novel PBP2a, substitutes for native PBPs; low affinity for all -lactams– MRSA chromosome contains ~ 50kb mec region
not present in MSSA. Acquired from coag-neg Staph spp.
– PBP2a is encoded by mecA gene; expression controlled by mecI, mecR1 and other factors.
Most MRSA are also resistant to macrolides and fluoroquinolones; remain susceptible to vancomycin.
Major nosocomial pathogen; primarily spread on hands of healthcare workers.

16
Enterococci and -lactams
Intrinsically less susceptible to -lactams– PenG/Amp MICs 10-fold higher than other
streptococci, not bactericidal• PenG/Amp + gent (bactericidal) for endocarditis
• Ampicillin alone effective for UTI
– Cephalosporins not active; increase risk of enterococcal infection
Acquired resistance is a new problem– High-level Amp resistance (altered PBPs in E.
faecium); [-lactamase still rare]

17
Vancomycin-resistant Enterococci
Since 1989, a rapid increase in the incidence of infection and colonization with vancomycin-resistant enterococci (VRE) has been reported by U.S. hospitals (MMWR Vol. 44 / No. RR-12)
This poses important problems, including:– Lack of available antimicrobial therapy for VRE
infections because most VRE are also resistant to drugs previously used to treat such infections (e.g. aminoglycosides and ampicillin).
– Possibility that vancomycin-resistance genes present in VRE can be transferred to other gram-positive bacteria (e.g. Staph. aureus )

18
Vancomycin
Member of glycopeptide family– Binds to D-Ala-D-Ala in peptidoglycan precursors– Prevents transglycosylation and transpeptidation– Resistance to -lactams does not confer cross-
resistance to vancomycin
Only active against Gram-positives– Cannot cross outer membrane of Gram-negatives– Primarily used for MRSA, MRSE infections; pts with
penicillin allergy; severe C. difficile disease.

19
MGMG MG MG
Vancomycin
Vancomycin binds to D-Ala-D-Ala; prevents transglycosylation and transpeptidation
Mechanism of Action of Vancomycin

20
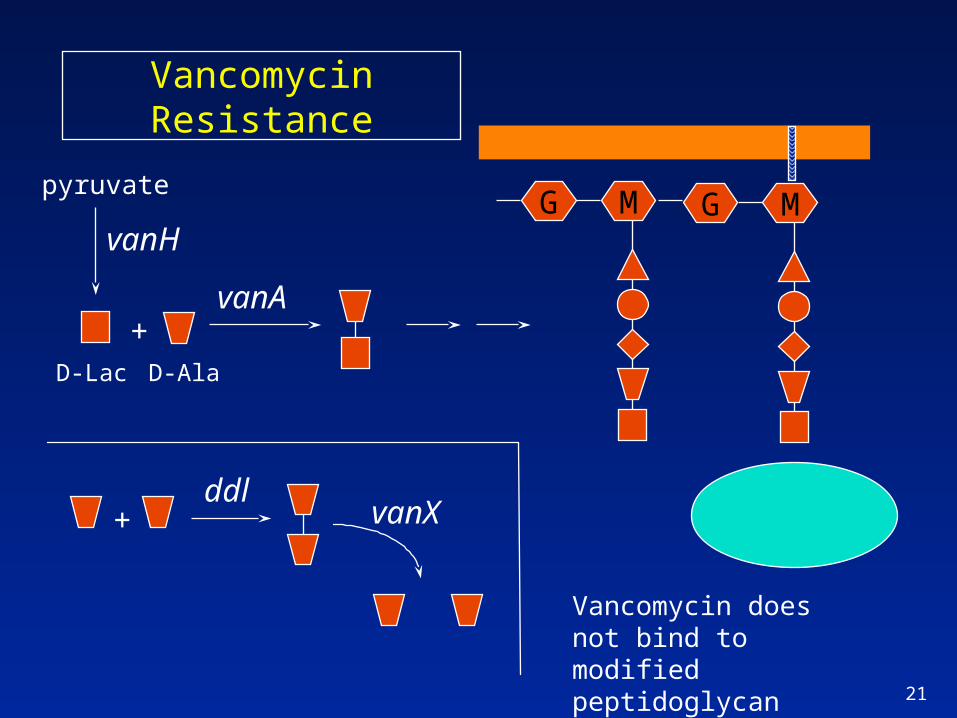
Mechanism of VRE
Acquired high-level resistance– E. faecium and E. faecalis containing vanA or vanB
gene clusters produce modified peptidoglycan containing D-Ala-D-lactate; does not bind vancomycin (MIC = 32 - >256
– Resistance genes are on mobile elements, have spread widely since 1st reports in late 80’s; major focus of infection control
– Multiresistant E. faecium (vancomycin, high-level ampicillin, high-level aminoglycoside) poses therapeutic challenge
Other enterococci contain vanC; low-level, non-transferable resistance; strains have low pathogenicity
21
Vancomycin Resistance
MGMG
+
+
vanA
ddlvanX
D-AlaD-Lac
Vancomycin does not bind to modified peptidoglycan
pyruvate
vanH

22
Epidemiology of VRE
Risk factors for colonization/infection in USA– Severe underlying disease (malignancy, ICU, long
hosp); antibiotics (vancomycin, 3rd gen cephs)
Reservoirs, routes of dissemination not fully understood
– VRE strains can be distinguished by molecular typing (PFGE)
– Multiple patterns are seen in some institutions (endogenous infection from intestinal source?)
– Clonal outbreaks are seen in others (transmission by HCWs?, fomites?)

23
VRSA - An emerging Problem
Several reports of S. aureus with reduced susceptibility to vancomycin since 1997– Japan and U.S. (Michigan, NJ, NY, Illinois)– Vancomycin MIC = 8 g/ml– Isolates obtained from patients with chronic MRSA
infection
No evidence of vanA or vanB Decreased susceptibility due to increased
levels of peptidoglycan and precursors

24
por
in
PG OMPM
-lactam resistance in Gram-negative rods
Factors that increase the MIC (resistance)– Increased enzymatic
inactivation
• High VMAX and/or low KM
• Increased enzyme concentration
– Decreased intracellular concentration
• Decreased influx• Increased efflux)
– Multiple mechanisms may function in the same strain

25
Gram-negative TEM-1 -lactamases
20-30% of E. coli are ampicillin-resistant– Most contain a plasmid-encoded class 2b -
lactamase (TEM-1). – Active against penicillins but not 3rd generation
cephalosporins. Inhibited by clavulanate.
All K. pneumoniae are ampicillin resistant– Contain chromosomal SHV-1 (related to TEM-1)
Most E. coli and K. pneumoniae are susceptible to 1st gen cephs (e.g. cefazolin).
Until recently, all were susceptible to 3rd gen cephalosporins (e.g. ceftriaxone, ceftazadime).

26
Extended Spectrum Beta-lactamases (ESBLs)
Changes in 1-5 amino acids near active site serine of TEM-1 (or SHV-1) greatly increase activity against 3rd gen cephalosporins and monobactams.– TEMs 3-29, SHVs 2-6; still inhibited by clavulanate – Carbapanems are only reliable -lactams vs ESBL
producers– Mainly seen in E. coli and K. pneumoniae– Located on transferable plasmids that may carry
additional resistance genes

27
Ceftazidime, Imipenem and ESBLs
During early 1990s, ESBL-producing Klebsiella became increasingly common at a hospital in NYC. In 1996 cephalosporin use was sharply curtailed to attempt reduce the ESBL burden (JAMA 1998;280:1233-37)
1995 1996
Median monthly use (grams)
Ceftazidime 383 66
Imipenem 197 474
Ceftazidime-resistant Klebsiella (nososcomial)
150 84
Imipenem-resistant P. aeruginosa (nosocomial)
67 113

28
ampC -lactamases
Several Enterobacteriaceae, including Enterobacter, Citrobacter , and Serratia, contain an inducible, chromosomal gene coding for a -lactamase (ampC)
– Very active in vitro against 1st gen cephs; low activity against 3rd gen cephs; not inhibited by clavulanate
– These organisms are naturally resistant to cefazolin, cefoxitin (strong inducers of ampC)
– Usually sensitive to 3rd gen cephs (poor inducers of ampC)

29
Regulation of ampC
AmpD
+
Peptidoglycan
autolysins
Peptidoglycan
[AmpR]- [AmpR]+
ampC+-lactam-
ase
Recycling of peptidoglycan produces NAM-tripeptide Normally catabolized by AmpD (NAM-tripeptide amidase) and recycled
into new peptidoglycan
NAM-tripeptide is also a positive activator of AmpR
Increases transcription of ampC

30
Resistance due to derepression of ampC
Many strains of Enterobacter and Citrobacter develop resistance to 3rd gen cephs during therapy. Resistant variants contain mutations that inactivate AmpD– NAM-tripeptide accumulates, causes stable
derepression of ampC– Increased levels of AmpC -lactamase inactivates
3rd gen cephalosporins
Resistant strains remain susceptible to imipenem (a carbapenem)– Poorly hydrolyzed, targets low copy PBP

31
-lactam Resistance in P. aeruginosa
Naturally resistant to many antibiotics– Outer membrane lacks high permeability porins
present in Enterobacteriaceae.– Pump mechanism actively exports antibiotics
Acquired resistance is common– Inducible ampC -lactamase
Imipenem resistance due to mutations that inactivate porin D2 (basic AA transporter)– Sole transporter of imipenem– Mutations in D2 decrease imipenem influx; -
lactamase inactivates sufficient drug to confer resistance.

32
Quinolones Inhibit topoisomerases/DNA synthesis
– Trap enzyme-DNA complex after strand breakage– DNA gyrase (topo II) (gyrA/gyrB)
• Primary target in Gram-negatives
– Topoisomerase IV [parC/parE (grlA/grlB in S.aur)] • Primary target in Gram-positives
Acquired resistance– Mutations in DNA gyrase and topo IV subunits
• Mainly gyrA and parC (grlA)
• Stepwise increase in resistance results from sequential mutations in primary and secondary targets
– Efflux pumps• P. aeruginosa, S. aureus, S. pneumoniae

33
Rapid Appearance of Ciprofloxacin Resistance in S. aureus
After the introduction of ciprofloxacin in the late ’80s there was rapid increase in resistance among MRSA– Prior to introduction of cipro at the Atlanta VAMC,
0% of MRSA were cipro-resistant. One year after introduction, 79% of MRSA were cipro-resistant (JID 1991;163:1279-85).
• More than one clone developed resistance
• One-half of pts had been previously treated with cipro (given for other infections)
• One year later 91% were resistant
• 13% of MSSA also became cipro-resistant

34
Quinolone-Resistant Campylobacter jejuni in Minnesota
During 1992-8 resistant isolates increased from 1.3 to 10.2% (NEJM 1999;340:1525-32).
Foreign travel was the major risk factor– Mexico, Caribbean, Asia– Prior antibiotic therapy accounted for 15% of resistance
There was also an increase in domestically acquired resistant isolates that was temporally related to the introduction of quinolones for treatment of poultry in 1995– Quinolone-resistant isolates were cultured from 20% (18/91)
of retail chicken products– 6/7 resistant subtypes (PCR-RFLP) from chicken were also
isolated from humans

35
Optimism and Concern on Many Fronts
NEJM (8/14/97): “In Finland, after nationwide reductions in the use of macrolide antibiotics for outpatient therapy, there was a significant decline in the frequency of erythromycin resistance among group A streptococci isolated from throat swabs and pus samples.”
NEJM (9/4/97): “We report high-level resistance to multiple antibiotics, including all the drugs recommended for plague prophylaxis and therapy, in a clinical isolate of Y. pestis. The resistance genes were carried by a plasmid that could conjugate to other Y. pestis isolates.”