4 somac!adaptaons! !and!selec’on!
TRANSCRIPT

4
Soma'c adapta'ons and selec'on

Increased capacity of recogni'on without overloading the genome with duplicates: Combinatorial associa'on
of polypep'de chains

Combinatorial associa'on of pep'des, TLRs in human
Cell Death and Differen.a.on (2006) 13, 816–825

The estimated numbers of mouse gene segments are: heavy chain κ light chain λ light chain V 300-1000 300 2 D 13 0 0 J 4 4 3 Combinations of gene segments: Heavy chain : 300 x 13 x 4 = 1.6 x 104
Light chain κ : 300 x 4 = 1.2 x 103
Light chain λ : 2 x 3 = 6 Combinatorial association of heavy and light chains :1.6 x 10exp4 x (1.2 x 103 + 6) = 1.9 x 10exp7 possible antibodies. (In fact the natural number of combinations is much larger due to the mechanisms of P- and N-recombination and somatic mutations.)
Combination of gene segments and of polypeptides contribute to antibody diversity

Alterna've splicing Mutually exclusive alterna've splicing
RNA level, not inheritable
Soma'c adapta'ons
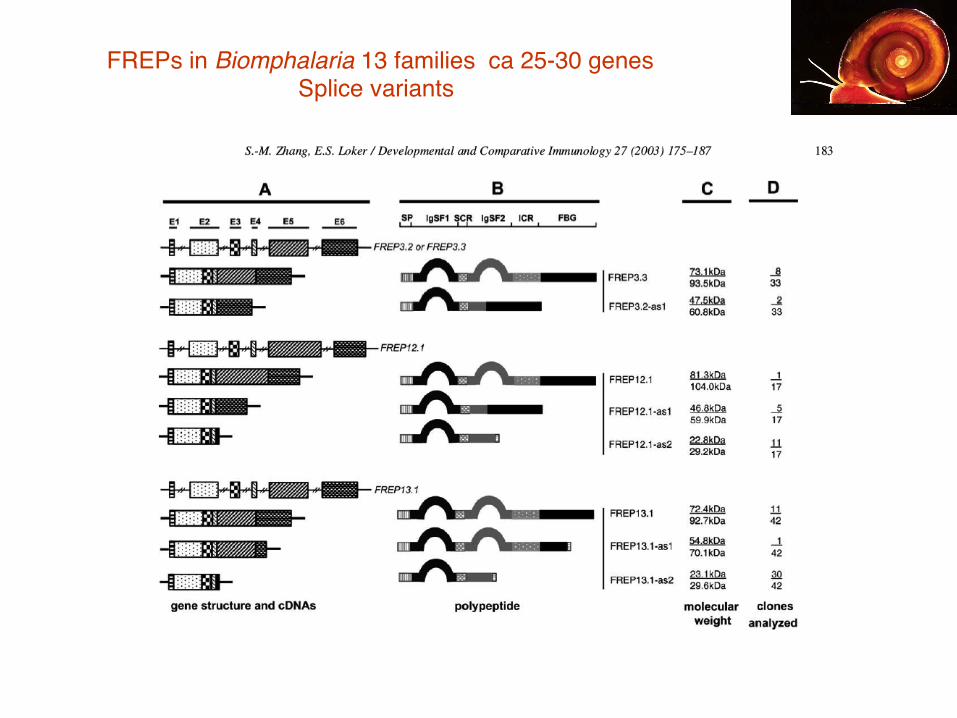
FREPs in Biomphalaria 13 families ca 25-30 genes!! ! ! !Splice variants

Wernert T et al 2003 J Biol.Chem 278 26319-‐26322
Pep'doglycan recogni'on proteins (PGRPs) are innate immunity molecules that contain a conserved pep'doglycan-‐binding type 2 amidase domain. PGRPs are ubiquitous in most animals. Insects have mul'ple PGRP genes that are classified into short (S) and long (L) transcripts and are oXen alterna'vely spliced into up to 19 different proteins. PGRPs have also been iden'fied in mollusks, echinoderms, and vertebrates), but plants and some metazoa, including nematodes such as Caenorhabdi.s elegans and crustaceans such as Daphnia, do not have PGRPs. PGRP genes usually form clusters that suggest their origin by gene duplica'on.

Schmucker et al. 2000 Cell. 2000 Jun 9;101(6):671-84, Science. 2005 Aug 18
> 30000 isoforms of DSCAM molecules
Brites D.et al, Biol Evol. 2008 Jul;25(7):1429-‐39.

Schema?c representa?on of Daphnia Dscam cytoplasmic tails (A) Daphnia magna tail structure and splicing possibili?es result in 4 alterna?ve forms.
Brites D et al. Mol Biol Evol 2008;25:1429-‐1439
© The Author 2008. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolu'on. All rights reserved. For permissions, please e-‐mail: [email protected]

Brites et al, 2013 More than one way to produce protein diversity: duplica?on and limited alterna?ve splicing of an adhesion molecule
gene in basal arthropods. “Evolu'on”, submiged
Pressure for DSCAM diversity and oriign of alterna've splicing mechanisms

DSCAM in Ixodes No hypervariable forms. Many germ-‐line full DSCAM genes
Many genes (>55)
mpst of them without any alterna've splicing capacity of the extracellular part

Botryllus Fester
Fester reconize the polymorphic determinants of the fuhc product on the surface of Botryllus ampullae cells leading to fusion or rejec'on.!
!Sushi -‐repeat: C3b/C4b-‐binding domain, short consensus repeat Scr
(apolipoprotein-‐H, complement system, blood coagula'on system, transglutaminases,adhesionproteins)

“Each individual appears to express both the full-‐length fester gene, plus an individual-‐specific subset of up to 36 alterna'vely spliced variants, encoding proteins with different extracellular and intracellular coding regions, as well as puta've secreted forms.”
Immunity, Volume 25, Issue 1, July 2006, Pages 163-‐173
Botryllus
Fester reconizes the polymorphic determinants of the fuhc product on the surface of Botryllus ampullae cells leading to fusion or rejec'on.!

…But apparently there was s'll room for further improvement of individual responses, for increasing the capacity of adapta'on during a life'me.
Further soma'c diversifca'on at the individual level
Leading to changes inheritable in cell lineages
Soma'c muta'ons Soma'c gene conversion
at the DNA level
Clonal amplifica'on possible
(Implies cell prolifera'on)

Zhang et al. Science. 2004 Jul 9;305(5681):251-4.

Zhang et al. Science. 2004 Jul 9;305(5681):251-4. Soma'c varia'ons in FREP genes of Biomphalaria

Diversity of 185/333 transcripts. Detected after LPS injection, produced by coelomocytes
David P. Terwilliger et al. Physiol. Genomics 2006;26:134-144
©2006 by American Physiological Society



Rearrangement of gene (DNA) segments Inheritable changes
Vertebrates
due to two types of enzymes AID and RAG
the most important ones and TdT
Analogies and homologies in the soma'c genera'on of immune repertoires of Vertebrates: Agnathans and Gnathostomes
“Do-‐it yourself” Kit
Soma'c adapta'ons

Two enzymes involved with the preceding somatic events: AID and RAG
AID: Activation -Induced ( Cytidine) deaminase
• Removes the amino group from the cytidine, replace sit by a uracil that trigger a repair mechanism , hence possible mutations depending on which base is reinserted inplace of
the U
• Involved in three separate somatic diversification processes: somatic mutation, gene conversion and class switch recombination of IgSF members of Gnathostomes
• Could it be involved in LRR somatic modifications in agnathans? • i.e. could a AID homolog be involved in an analogous process? Yes
• Pancer Z et al Nature Imm. 2007 8:647-656
• RAG 1 and 2 (Recombination activating gene)!• The recombina'on ac'va'ng gene 1 component is thought to contain most of the cataly'c
ac'vity, while the N-‐terminal of the recombina'on ac'va'ng gene 2 component is thought to form a binding scaffold for the 'ght associa'on of the complex with DNA.
• • Present in Echinoderms , expressed early during gastrulation, hardly at all in hemocytes
• Rast J et al PNAS 2006 103: 3728-3733

APOBEC family phylogeny (AID enzyme involved in soma'c adapta'ons) AID a rela'vely recent member from an old family
Although the family forms part of a larger superfamily of deaminases distributed throughout the biological world, the AID/APOBEC family itself is restricted to vertebrates with homologs of AID (a DNA deaminase that triggers an'body gene diversifica'on) and of APOBEC2 (unknown func'on) iden'fiable in sequence databases from bony fish, birds,
amphibians, and mammals.

A homolog of RAG1-‐RAG2 in sea urchins not detected in coelomocytes but during embryonic development. Func'on???

S Fugmann Semin Immunol 2009 22: 10-‐16

ASSEMBLY(AND(DIVERSIFICATION(OF(GENES(ENCODING(ANTIGEN(RECEPTORS
JAWLESS(VERTEBRATES
LRRNT LRRCTLRR&modules52 3 41
LRR&modulesn7 8 96
LRR&modules52 3 41
LRR&modulesn7 8 96
LRRNT LRRCTNT CTUUU
U U
U
AID
Homology(searchExtension(by(DNA(polymerase
LRR&modules52 3 41
LRR&modulesn7 8 96
LRRNT LRRCT2 5 7 9NT CT
Copied(LRR(modules
VLR(gene(assembly
JAWED(VERTEBRATES
V D J C
V D J C
V D J C
AID
V D J C
U U
ErrorSprone(repair
Point(mutation
Somatic(hypermutation
VψV D J C
AID
VψV D J C
U U
Gene(conversion(trace
Copy(from(ψV
Gene(conversion
DNA(deletion
ClassSswitch(recombination
V D J Cγ
Replaced(exon
S
(
AID
V D J Cµ Cγ(
Sµ Sγ
UUU U
RAG1SRAG2(V(D)J(recombinationTDT

The Immunoglobulin loci in Vertebrates
• Cluster organiza'on in chondrichthyes(VJC Or VDJC mul'ple copies)
• Translocon organiza'on VVVVn JJJn C pr VVVVn JJJn C from Teleots(cluster conserved in some cases)
• Limited rearranging capacity in birds gene conversion


Soma'c gene conversion
A hyperconversion mechanism generates the chicken light chain preimmune repertoire. Reynaud CA, Anquez V, Grimal H, Weill JC.Cell. 1987 Feb 13;48(3):379-‐88
Combinatorial rearrangement
EMBO J. 1988 Aug;7(8):2409-‐15. Organiza'on and rearrangement of immunoglobulin M genes in the amphibian Xenopus.
Schwager J, Grossberger D, Du Pasquier L.

Many different processes selected during evolution can make the number of receptors much larger than the number of genes that encode them
Population level • Polymorphism (receptors and effectors: lectins, C’ related, AMPs, srcr, Ig, TCR, MHC
etc)
Individual level • Peptides Combinatorial association of polypeptide chains: Ig H.L, TCR αβ γδ,
TLRs, PGRPs,… • • Nucleic acids • RNA • Alternative splicing: e.g. SRCRs,PGRPs, FREPs, DSCAM, • (Arthropods, Mollusks, Echinoderms, Urochordates, Vertebrates) • Post transcriptional level: 185/333 (Echinoderms) • DNA
Somatic rearrangement: Ig, TCR,VLR (combinatorial joining) (Vertebrates) Somatic gene conversion: Ig Vertebrates.LRR Agnathans
Somatic mutations: Ig from sharks on. Mollusks? Heavy chain class switch: From shark to mammals

Ques'ons Is all this diversity useful? (concentra'on of specific products) Advantages Individualiza'on, economy Problems Regula'on of expression, autommunity

Selec'on? Understood fro Gnathostomes only.
Te MHC, its origin.
Large repertoires of TCR and An'bodies are useful…. but also dangerous

Germ-‐line diversity can be selected in the phylogeny. Large diversity generated randomly by soma'c processes has to be selected soma'cally.
• Autoimmunity control: rela'vely well understood in vertebrates Central dele'on (thymus, AIRE)(Bone marrow)
Peripheral inhibi'on (regulatory T cells)
• Sites and means of selec'on (MHC, stroma of lymphoid organs).
• Lymphocyte (+Lymphoid organs) = good adapta'on to the necessity of selec'on.
• One cell/one “an'gen” receptor. If one cell is “autoimmune” there is just this one cell to eliminate.
• Once selected the lymphocyte is an efficient effector, prolifera'ng fast, filling par'ally the genera'on gap with pathogens.
• Darwinian varia'on selec'on inside the body!
ORIGIN OF LYMPHOCYTES? Build up of the vertebrate adap've system plan

Class I and Class II pathways of presenta'on Class I internal, self, viruses via LMP TAP ClassII external via lyzosomes
MHC

The two basic pathways of an'gen presenta'on: ClassI and ClassII MHC mediated

Agnathan ? Boehm’s group report AAI mee'ng, 2010: FoxN1, VLRA in situ
Courtesy John Horton

Bajoghli et al. 2010 Nature, in press

Selec'ng and maintaining a diverse T-‐cell repertoire Ananda W. Goldrath and Michael J. Bevan 402, 6-13

MHC Class I molecule
Peptide binding region
Ig sf C1 domain
Ig sf C1 domain

A model for the origin of MHC class I and class II
Flajnik et al
Other models suggest that Class II came first
Kaufman et al

Fundamental Immunology 2008
MHC architectures across Vertebrates

ABC? PROTEASOME
C’3 Bf TNFSF X Y Z
12p13 6p23.1 19p13.1-‐.3 1q21-‐25 9q33-‐34
RXRB -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ COL11A2
RING3(BRD2) LMP2 LMP7 MECL1* TAP
NOTCH4 PBX2 TNX CYP21 C4
MHCclassI RAGE TNFSF-‐3 -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ TAPBP -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐
-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ CACNA1A4 COL5A3 BRD4 -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ NOTCH3 -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ CYP2 C3
FcRN+
Basigin TNFSF-‐7,9 VAV1 -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ C5aR+
RXRG CACNA1A6 COL11A1 BRDT -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ NOTCH2 PBX1 TNR
-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ CD1 PO
TNFSF-‐4,6,18 VAV3 -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐
RXRA CACNA1B COL5A1 BRD3 -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ LMPZ ABC2? NOTCH1 PBX3 HXB
-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ C5
-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐
TNFSF8,15 VAV2
TAPBP-‐LIKE Toll 4
-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ α2M -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐
CD4, LAG3 -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐ TAPBP-‐R C3aR
NK comp
Duplica'on, Pre-‐jawless fish >520 mya
Duplica'on, Pre-‐jawed fish >440 mya
Igsf C1
Flajnik et al Fundamental Iimmunology, 5th ed in press!
Amphioxus data suggest that human chromosome 9q34 paralogue has retained many ancestral MHC linked genes(Abi-Rached et al, Nature Genetics 2002)!!

The Proto-‐MHC of Placozoans, a Region Specialized in Cellular Stress and Ubiqui?na?on/Proteasome Pathways Jaanus Suurväli, Luc Jouneau, Dominique Thépot, Simona Grusea, Pierre Pontaro|, Louis Du Pasquier, Sirje Rüütel Boudinot and Pierre Boudinot
J Immunol 2014; 193:2891-‐2901

TCR gene organiza'on in fish and human