5 critical factors in plankton abundance
TRANSCRIPT

٤٩
Critical Factors in Plankton Abundance
Plankton are extremely variable in abundance, both spatially and temporally. This is especially true of the phytoplankton, upon which zooplankton depend. Although many factors contribute to variation in the abundance of phytoplankton, water motion is the dominant factor. It regulates two factors that are vital for the growth of phytoplankton populations: (1) the exchange of nutrients with deeper waters and (2) the mixing of waters downward and therefore away from exposure to sunlight. Water motion is important for phytoplankton growth in all of the ocean, but its influence is best introduced in a discussion of the spring diatom increase, which occurs in nearly all areas where there is a seasonal change in day length. Of course, light and nutrients are crucial determinants of phytoplankton, which after all are photosynthetic and require these for growth. Finally, animal grazing affects phytoplankton abundance. It is the purpose of this chapter to discuss the importance and interaction of these various factors on plankton dynamics.
The Seasonal Pattern of Plankton Abundance
In midlatitudes, phytoplankton increase in the spring, decline in summer, and may increase to a lesser extent in fall. There is a predictable seasonal pattern to plankton abundance in temperate and boreal waters of depths of an approximate range of 10-100 m. Figure 1 traces the seasonal changes in phytoplankton, zooplankton, light, and nutrients during the year in a temperate-boreal coastal zone.
Fig. 1 Idealized diagram tracing changes in phytoplankton, zooplankton, light, and nutrients during the year in a temperate-boreal inshore body of water.

٥٠
Usually in the early spring, phytoplankton increase dramatically and are dominated by a few diatom species. This is known as the spring diatom increase. The exact time of phytoplankton increase varies with latitude and year. In Palestinian waters, the peak of phytoplankton abundance (as determined by chlorophyll concentrations) in South Eastern Mediterranean near Palestine occurs in late winter and early spring. Prior to construction of the dam in 1965, the annual Nile floods peaked between August and October was responsible for the major influx of nutrients utilized by the primary producers and consequently triggered a major phytoplankton bloom off the Egyptian coast and along the coast of Palestine. Since the operation of the Aswan Dam in 1965, the water flow, particulate loading and dissolved nutrient input of the Nile into the southeastern Mediterranean has greatly diminished and the seasonality of the phytoplankton blooms have changed from summer/autumn phytoplankton bloom to the winter–spring period.
Generally, the phytoplankton begin to decline as summer approaches. In some locations, the phytoplankton increase again in the fall, followed by a decline to very low abundance again in the winter.
Zooplankton start to increase as the phytoplankton bloom reaches its peak, attaining a maximum in the late spring or early summer. Zooplankton reach their yearly maximum after the spring phytoplankton increase begins to decline. Calanoid copepods dominate this burst of abundance and are the principal grazers of the diatoms. Planktonic larvae of benthic invertebrates are also common in the late spring and in early summer. Following this burst, zooplanktivorous fish and invertebrates become abundant. Comb jellies and jellyfish become especially abundant and are effective predators on copepods and planktonic larvae. These gelatinous creatures may clog plankton nets in summer, when arrow worms and tunicates may also become abundant.
The temperate-boreal pattern is not universal. The strength of the spring phytoplankton bloom varies with latitude. In the Arctic, a single summer peak of phytoplankton abundance is followed by a zooplankton maximum (Figure a). The phytoplankton production lasts as little as 2-4 weeks. In temperate-boreal waters, as we have discussed, a spring phytoplankton increase is followed by a decrease, coinciding with a zooplankton increase (Figure b). In late spring and summer, the zooplankton decline and a peak of phytoplankton may follow in the fall. In the tropics there is no clear alternating pattern of phytoplankton and zooplankton abundance (Figure c). This correlates with the relative lack of seasonality in tropical waters.

٥١
The Spring Diatom Increase and Water Column Parameters
Water Column Stability and Light
Because light irradiance decreases exponentially with increasing depth, there is a compensation depth below which respiration for a given phytoplankton cell exceeds photosynthetic output. Light irradiance decreases exponentially with increasing depth (Figure 3) and becomes a limiting factor to photosynthesis. The compensation depth is that depth at which the amount of oxygen produced by a phytoplankton cell in photosynthesis equals the oxygen consumed in respiration. We can estimate the compensation depth by placing phytoplankton cells in a clear bottle. At depths shallower than the compensation depth, there is a net increase of oxygen over time, whereas at depths greater than the compensation depth, there is a net decrease of oxygen over time. The compensation depth is thus an indicator of the potential of a photosynthesizing cell to be a net producer. The light intensity corresponding to the compensation depth is the compensation light intensity.
Fig. 3 Decrease in light irradiance with increasing depth in coastal water and clear ocean water. Note that the light intensity is plotted on a logarithmic scale; light is decreasing exponentially with depth. The compensation depth is controlled by season, latitude, and transparency of the water column. As the temperate-boreal spring progresses, the increasing photoperiod tends to increase the compensation depth to an eventual maximum. The Arctic winter photoperiod is zero and therefore the compensation depth is at the surface. Suspended matter in coastal waters reduces the compensation depth relative to the open sea. Similarly, as a phytoplankton bloom develops and as suspended matter (seston) becomes trapped in the water column, the compensation depth decreases due to light absorption and shading by particles.

٥٢
In winter, the water density is about the same throughout the water column, and phytoplankton cells are stirred on average to depths that prevent average photosynthesis from permitting the phytoplankton population to increase. Consider the state of the plankton and of the water column in the winter before the spring diatom bloom occurs. At this time the water column is isothermal, with little or no density variation with depth. Near the shore, temperate-boreal winters are times of high wind stress, resulting in extensive overturn of the water column. Because there are no density differences, the water column is unstable, and winds cause extensive vertical mixing.
The mixing depth is the depth above which all water is thoroughly mixed under the wind's influence. Because the winter mixing depth is great due to storms, phytoplankton cells can easily be swept down to great depths, where there is not enough light for photosynthesis. A phytoplankton bloom cannot be initiated because any potential profit in photosynthesis would be lost through mixing to greater depths. So even though the photoperiod may increase as winter progresses, the instability of the water column may preclude the development of a phytoplankton bloom. Along with the increase of photoperiod, water column stability is thus an essential part of the development of the phytoplankton bloom. Water is usually well mixed in winter, and plankton are uniformly distributed. Hence, respiration is about the same at all depths. Because a population increase of phytoplankton requires that total production exceed total respiration, a phytoplankton bloom can occur only when the volume of water in which photosynthesis occurs has a net excess of production over consumption (respiration, in this case).
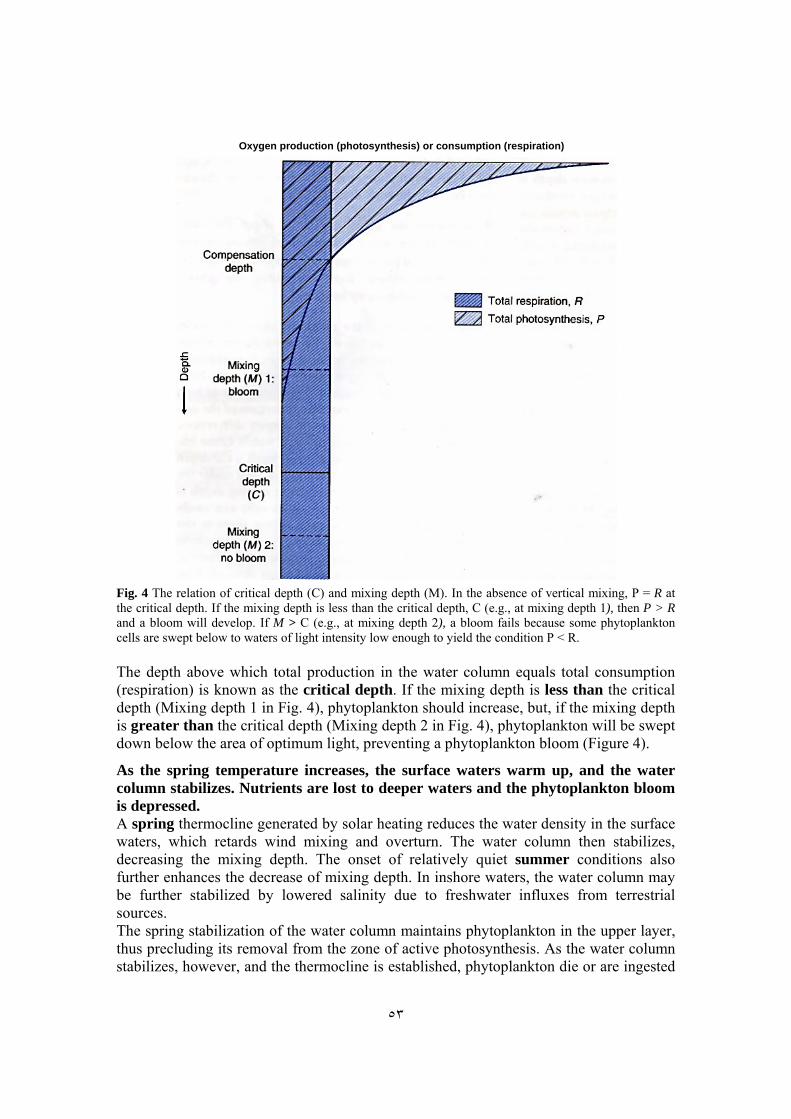
٥٣
Oxygen production (photosynthesis) or consumption (respiration)
Fig. 4 The relation of critical depth (C) and mixing depth (M). In the absence of vertical mixing, P = R at the critical depth. If the mixing depth is less than the critical depth, C (e.g., at mixing depth 1), then P > R and a bloom will develop. If M > C (e.g., at mixing depth 2), a bloom fails because some phytoplankton cells are swept below to waters of light intensity low enough to yield the condition P < R. The depth above which total production in the water column equals total consumption (respiration) is known as the critical depth. If the mixing depth is less than the critical depth (Mixing depth 1 in Fig. 4), phytoplankton should increase, but, if the mixing depth is greater than the critical depth (Mixing depth 2 in Fig. 4), phytoplankton will be swept down below the area of optimum light, preventing a phytoplankton bloom (Figure 4).
As the spring temperature increases, the surface waters warm up, and the water column stabilizes. Nutrients are lost to deeper waters and the phytoplankton bloom is depressed. A spring thermocline generated by solar heating reduces the water density in the surface waters, which retards wind mixing and overturn. The water column then stabilizes, decreasing the mixing depth. The onset of relatively quiet summer conditions also further enhances the decrease of mixing depth. In inshore waters, the water column may be further stabilized by lowered salinity due to freshwater influxes from terrestrial sources. The spring stabilization of the water column maintains phytoplankton in the upper layer, thus precluding its removal from the zone of active photosynthesis. As the water column stabilizes, however, and the thermocline is established, phytoplankton die or are ingested

٥٤
and egested by zooplankton, sinking below the compensation depth. Because of the stabilization of the water column, these materials and other nutrients are not returned to the surface from greater depths and from the bottom. Toward the end of summer, with the advent of fall storms, the thermocline may be dis-rupted, bringing nutrients toward the surface from the bottom in shallow water. This may result in a fall increase of phytoplankton.
As the water column stabilizes in late spring and summer, phytoplankton organisms denser than seawater, such as diatoms, start to sink from the water column. Such considerations do not hold, however, for phytoplankton (e.g., dinoflagellates) whose swimming abilities circumvent this tendency to sink. If these ideas are correct, then we may conclude that the hydrographic conditions tied to seasonal variation play the primary role in the birth, development, and demise of the spring phytoplankton increase. The stabilization of the water column in spring initially permits the development of the spring increase. The stability of the water column, however, prevents nutrients lost from the surface waters from returning to the surface where light is available for photosynthesis. Furthermore, dense phytoplankton organisms sink out of the water column when spring-summer stability sets in. The poor nutrient situation prevails until the following fall or winter storms overturn the density homogenized water column.

٥٥
Water Column Exchange in Very Shallow Waters and in Estuaries
The importance of water column stability mentioned above may change with the depth of the basin and with season. The spring phytoplankton bloom ends when the water column is strongly stratified, because nutrients are not regenerated from the bottom to the surface. In shallower waters, however, the nutrient exchanges between bottom and surface waters called benthic-planktonic coupling are much stronger and are not shut off by stratification. Many bays and estuaries are very shallow, and even modest wind mixing mixes the water column completely. In late spring and summer, decomposition of organic matter in shallow bottoms, combined with vertical mixing, brings nutrients back to the surface, and a summer phytoplankton bloom occurs that can exhibit higher productivity than the spring event. Beach blooms: Benthic-planktonic coupling figures importantly in a nearshore phenomenon known as beach blooms. These are phytoplankton blooms of a few species of very large diatoms. Even though the coast appears to be very energetic with strong waves, an offshore submerged bar confines the vertically recirculating water to the nearshore region (Figure 5). Dense populations of benthic animals excrete into the water column nutrients that are thoroughly mixed and fuel enormous phytoplankton growth, enough to color the water a deep brown. The diatoms sink to the bottom and are eaten by the invertebrates, although some planktonic consumption occurs as well. Input from waters offshore and from the exposed beaches adds nutrients to the system.
Fig. 5 A benthic-planktonic coupling system. Bivalve excretion provides nutrients for phytoplankton uptake, as do inputs from the beach. Bivalves in the bottom feed on the phytoplankton. A large confined circulation landward of an offshore bar recirculates the water. Water exchange with offshore waters is relatively restricted. In deeper water, especially seaward of the shelfslope break, benthic-planktonic coupling is indirect at best. In some parts of the ocean a surprising amount of material sinks from the surface waters to the bottom, but the return route to the surface is very indirect and depends upon bottom currents. Such currents could not restore organic material to the surface waters for many years.
Spring freshet in estuaries: The extraordinarily high primary production of estuaries owes itself to large inputs of nutrients from the tributaries of the watershed, with a special seasonal peak during the spring freshet (the flood of a river from heavy rain or melted snow). Nutrient input to estuaries occurs as a pulse during the spring freshet, but some estuaries have rather short residence times for water, which is rapidly lost to the sea. In most estuaries, primary productivity increases as the nutrient input of the freshet increases, and decreases as the degree of flow from the estuary increases in late spring and summer.

٥٦
Light Required by Phytoplankton Energy from solar sources is expressed in terms of energy units (such as langleys per minute, g cal cm-2 min-1). The angle of the sun at different times of day, the latitude, and other factors contribute to the spectral distribution of light that strikes the sea surface and the amount of back-scattering. The spectral distribution of light striking the sea surface includes a large part of the ultraviolet-infrared spectrum; however, only visible parts of the spectrum penetrate to great depths. At temperate latitudes in clear weather during the summer, the maximum energy striking the sea surface is about 1.4 langleys per minute. About one-half the total radiant energy is in the infrared region of the spectrum and so is not available to marine photosynthetic organisms. Light attenuation Light is attenuated in the water column through absorption and scattering. Scattering can be accomplished by (1) water molecules, (2) dissolved organic matter, (3) particulate organic and (4) inorganic material, and (5) living plankton themselves.
Figure 7 shows attenuation values for different wavelengths of light. In the clear open ocean, the attenuation spectrum of light transmission maximizes transmission at about 480 nanometers (nm). In turbid inshore waters, however, a maximum transmission occurs at longer wavelengths, approximately 500-550 nm.
Fig. 7 Attenuation of different wavelengths of light with increasing depth (each curve is labeled by depth in meter) below the sea surface. Wavelengths is in angstroms (1 Å × 0.1 = 1 nm)

٥٧
Ultraviolet light Because ultraviolet light has detrimental effects on DNA, its penetration is of great interest. In moderately turbid coastal waters, incident light with a wavelength of 380 nm or less is almost attenuated at a depth of 1-2 m, but in very clear parts of the ocean, 20 m may be required to remove 90 percent of the radiation entering the surface.
light intensity and depth variations Incident light near the surface is intense enough to inhibit photosynthesis through bleaching of photosynthetic pigments, such as chlorophyll-a, or the arresting of pigment production. With increasing depth, light energy is absorbed to the extent that the inhibitory effect disappears. Photosynthetic pigments Photosynthetic phytoplankton use chlorophyll-a, chlorophyll-c, and a variety of "accessory" pigments to utilize light energy from most of the visible spectrum. Within the usable wavelengths of 400-700 nm, the light absorbed by phytoplankton pigments can be divided into (a) light of greater than 600 nm, which is mainly absorbed by chlorophyll, and (b) light of less than 600 nm, which is mainly absorbed by accessory pigments. The combined absorption of chlorophyll and accessory pigments allows the yield of photosynthesis to be constant over a large portion of the visible light spectrum. Photosynthesis and Light intensity Figure 8 illustrates a theoretical photosynthesis light curve showing photosynthetic rate as a function of light intensity. Photosynthesis increases with increasing light intensity and then reaches a plateau at a maximal value Pmax, then it is inhibited by high light intensity.
Fig. 8 The relationship between photosynthesis and light intensity.

٥٨
At the compensation light intensity, the photosynthetic rate (measured in terms of oxygen evolution) equals the amount of oxygen consumed in respiration. Because the amount of light reaching a phytoplankton cell varies over a day, the compensation light intensity is usually expressed on a 24-hour basis. It is assumed here, for simplicity, that respiration occurs at the same rate in the light and the dark (a shaky assumption because of light-accelerated respiration, known as photorespiration). An average 24-hour compensation light intensity is in the range of 3-13 langleys per day in temperate seas. The physiological adjustment to surrounding light conditions involves changes of some of the following morphological and biochemical factors: • total photosynthetic pigment content, • pigment proportions, • morphology of the chloroplast, • chloroplast arrangement, • and availability of Calvin cycle enzymes.
For example, under strong light conditions, diatom chloroplasts shrink and aggregate. Adaptations to changes in light intensity usually occur within 1 day. Deep-water phytoplankton can respond partially to low light intensities by increasing the total pigment content.
Nutrients Required by Phytoplankton
Nutrients occur in dissolved and particulate form • dissolved − dissolved inorganic form (e.g., ammonium, nitrate) or in − dissolved organic form (e.g., amino acids, vitamins). • Particulate organic forms
In oceans, dissolved organics, particulate organics, and living biomass occur, on average, in the approximate proportions 100:10:2. Amounts of required nutrient • Major nutrient elements are required in great amounts and include carbon,
nitrogen, phosphorus, oxygen, silicon, magnesium, potassium, and calcium. • Trace nutrients elements are required in far smaller amounts and include iron,
copper, and vanadium. Organic nutrients include vitamins. Type of nutrients uptake • Autotrophic the uptake of inorganic nutrients in association with photosynthesis. • auxotrophic uptake are the uptake organic nutrients in association with
photosynthesis. • Heterotrophic uptake refers to uptake of organic substances for nutrition in the
absence of photosynthesis. Many phytoplankton can absorb peptides and even engulf particles.

٥٩
Marine snow Toward the end of rich phytoplankton blooms, some species become sticky and exhibit negative buoyancy. As they sink, they aggregate and form particles. This resulting fragile organic aggregate material, known as marine snow (Figure 9), is often found to be enriched by a variety of planktonic organisms and detrital products of plankton. Marine snow persists in relatively quiet water and can be a major mechanism of transport of material to deeper waters. Marine snow aggregates are rich in organic matter, derived from various sources including decaying phytoplankton cells, mucus from various gelatinous zooplankton, and marine bacteria. In the eastern Pacific, the gelatinous houses of larvacean tunicates are a major part of the larger marine snow particle aggregates. Sometimes the aggregates are rich enough in organic matter and bacterial activity to cause the material to become anoxic.
Fig. 9 Marine snow derives from collisions of macromolecules and from the degradation of dead plankton. Fragments of gelatinous zooplankton have been colonized by bacteria, and other fragments have adhered to the original larger particle.
Nitrogen
Importance: Nitrogen (N) is required for the synthesis of proteins Occurrence forms: occurs in • three inorganic dissolved forms: ammonium (NH4+), nitrate (NO3
-), and nitrite (NO2
-). • dissolved organic forms, such as urea, amino acids, and peptides.
Ammonium is usually the preferred form of nitrogen from a nutritional perspective because no chemical reduction is required to be used in protein synthesis. Dissolved ammonium can inhibit the uptake of nitrate. Ammonium can also be taken up more ef-ficiently at low light levels than is the case for nitrate. Nitrate and nitrite must be reduced by the enzymes nitrate reductase, making their uptake a chemically slower process. The highest concentration of dissolved nitrogen that occurs in the ocean is that of nitrate (roughly 100 micromolar: 100 µM), and nitrate is often the most abundant form of nitrogen in eutrophic coastal waters. Upwelling and storm-induced turbulence carry nitrate to the euphotic zone. Under certain circumstances, however, ammonium can surpass nitrate in abundance (usually when the nitrate is used up).

٦٠
Nitrite is generally the rarest of the three nitrogen bearing nutrients and behaves similarly to nitrate in phytoplankton nutrient uptake, it must be reduced by the enzymes nitrite reductase. The dissolved concentrations of all three forms of nitrogen increase in the temperate-boreal winter and decrease in the spring and summer when phytoplankton populations build up.
Sources of Nitrogen New Sources Attributed to nitrogen supplied from nutrient-rich pool of deeper waters by upwelling, or through river/land, or atmospheric input (Some Cyanobacteria can fix free nitrogen (N2). As long as phytoplankton cells are not mixed downward too far, this supply, combined with light, allows phytoplankton to grow.
Regeneration Sources Attributed to nitrogen supplied from a recycling process that occurs within the euphotic zone such as the excretion processes by zooplankton and bacteria, which excrete nitrogen, usually in the form of ammonia. This is used by phytoplankton.
New production versus regenerated production The amount of primary production attributable to nutrient supply from deeper waters is known as new production. By contrast, The amount of primary production attributable to nutrient supply from regeneration sources is known as regeneration production. Because regeneration production comes from a recycling process that occurs within the system, the new production is often of greater interest in regional estimates of primary productivity because it represents the outcome of nutrient supply from outside the system.
Nitrogen cycling in the sea is intimately involved with microbial transformations. Nitrogen cycling involves a large pool of gaseous nitrogen in the atmosphere. Nitrogen may be incorporated into marine food chains through the process of nitrogen fixation (ac-complished by some bacteria, cyanobacteria, and yeasts). Gaseous nitrogen is converted eventually to nitrogen in proteins. Several distinct groups of bacteria transform nitrogen from one form to another. Some nitrifying bacteria convert ammonium ion to nitrite, whereas other species of nitrifying bacteria oxidize nitrite to nitrate. Both processes require the presence of oxygen. Under anaerobic conditions, denitrifying bacteria reduce nitrate to ammonium ion. Nitrate-reducing bacteria can return nitrate to the atmosphere, in the form of nitrogen gas.
Nitrogen fixation and denitrification In the global ocean, nitrogen is gained by nitrogen fixation and lost by the process of denitrification. Nitrogen fixation is an anaerobic process. In some seas, nitrogen fixation is responsible for as much as 20 percent of the input of nitrogen into phytoplankton, but elsewhere the role of nitrogen fixation is trivial. The process of denitrification is most common in anaerobic environments such as inshore water columns and sediments. Denitrification and nitrogen fixation are not coupled processes, so there is a possibility that one will dominate the other. Current evidence suggests that there is a slight excess of denitrification, which implies that nitrogen is a more limiting nutrient element than phosphorus, at least as a world average.

٦١
Phosphorus
Role The biochemical role of phosphorus (P) is different from that of nitrogen because phosphorus is used primarily in the energy cycle of the cell. Adenosine triphosphate (ATP) is a crucial energy source in all enzymatic reactions.
Occurrence forms Phosphorus occurs in the ocean as
• inorganic phosphate (PO4), • dissolved organic phosphorus (P), and • particulate phosphorus.
Phosphate is the form preferred by phytoplankton and exchanges rapidly between phyto-plankton and seawater. Phosphate is taken up very rapidly by phytoplankton, and the con-centration in surface waters is usually quite low.
• At the usual alkaline pH of seawater, organic phosphates are easily hydrolyzed to inorganic phosphate
• Alkaline phosphatase: enzyme of algae and bacteria helps converting organic phosphate into inorganic phosphate, which is subsequently taken up; phosphatase can occur intra- or extracellularly.
Phosphorus is recycled rapidly between the water and phytoplankton and is often a rate-limiting step in primary productivity as a result. Grazing and excretion by the zooplankton allow rapid regeneration in the plankton. As phytoplankton detritus settles from the water column, the sediments accumulate phosphorus. Benthic decomposition results in the diffusion of phosphorus from the bottom. The remixing of the water column in fall and winter returns phosphorus to the surface waters. The limiting nutrient? A limiting nutrient is a chemical necessary for algal growth — but is available in smaller quantities than needed for algae to increase their abundance. Once the limiting nutrient in a water body is exhausted, the population of phytoplankton stops expanding. If more of the limiting nutrient is added, larger algal populations will result until their growth is again limited by nutrients or by other limiting environmental factors. A simpler method to determine what is limiting algal growth is one based on knowledge of the uptake of nutrients by algae. Surface water samples (0.5- to 1-m depth) are collected during critical periods of maximum algal biomass and analyzed chemically for available nitrogen (ammonia and nitrate) and available phosphorus (soluble orthophosphate). This will provide an assessment of the amounts of nutrients that are still available in excess amounts after the peak biomass has occurred; those present in excess amounts relative to the amounts needed for growth are not limiting growth, i.e., the amount of algal available nutrients present in the water is sufficient to allow the algae to grow at a rate independent of the concentration of the nutrient. Under some conditions, both nutrients may be present in growth-limiting amounts, and it may be a combination of these nutrients which is limiting maximum biomass development. In some water bodies, both (N and P) may be present in sufficient amounts and some factor other than

٦٢
nitrogen or phosphorus concentrations, such as light or a toxic substance, would be limiting maximum algal production. In ocean, nitrogen is believed to be the main element limiting phytoplankton growth, Rather than phosphorus. Usually a shortage of nitrogen is believed to be responsible for stopping the growth of phytoplankton populations. A shortage of phosphorus in a cell must be extreme before it affects energy-related processes, such as enzymatic reactions, and photochemical processes in photosynthesis.
Nitrogen/Phosphorus Ratio in the Sea The growth of phytoplankton results in the simultaneous depletion of both nitrogen and phosphorus. Those elements are available in ocean water (N:P = 15:1) in very nearly the proportions usually required by phytoplankton (N:P = 16:1). Phytoplankton particles are more enriched in nitrogen (N:P = 16:1), which suggests that in as much as N and P are taken up in photosynthesis, nitrogen is a limiting factor in primary production. These proportions are known as Redfield ratios, after their discoverer. As we mentioned in the discussion of nitrogen, there appears to be a slight excess of denitrification, leading to the apparent difference in N:P ratios in phytoplankton and in water. Despite the probable correctness of this hypothesis, numerous examples exist of phytoplanktonic species that deviate from the "typical" N:P uptake ratio.
Silicon Silicic acid, which contains the nutrient element silicon, is a constituent of seawater and is essential for the skeletons of diatoms. Depletion of silicon inhibits cell division and eventually suppresses the metabolic activity of the cell. In natural waters depletion of sil-icon can limit diatom populations and may direct the plankton succession (see below) toward phytoplankton lacking a siliceous test. As diatoms sink from surface waters, they remove silicon, which may partially cause the demise of the spring diatom increase found in so many temperate and higher-latitude waters.
Trace Substances and the Iron Hypothesis
Inorganic trace metals, such as iron, manganese, and zinc, have important functions in oxidase systems (iron is the cofactor in the oxygen evolution step of photosynthesis) and serve as cofactors for enzymes essential for plant growth (e.g., molybdenum, zinc, cobalt, copper, and vanadium play such a role). Iron limits the photosynthetic potential of phytoplankton. Experimental studies found that phytoplankton growth was strongly increased in water in bottles to which iron had been added. Chelators may be important in affecting the utilization of trace metals. Phytoplankton may synthesize and release chelating substances into the water to alter the availability of trace metals such as iron. During glacial maxima in the Pleistocene epoch, iron was eroded from the land and supplied at a much higher rate to the ocean. Organic trace nutrient substances, particularly vitamins, may also be of great significance in the sea. Almost all marine phytoplankton species are auxotrophic and require cobalamine, thiamine, or biotin. In mixed cultures, vitamin production and release by one species may stimulate the growth of another species, although most vitamin production is probably by bacteria.

٦٣
Phytoplankton Succession Seasonal changes in dominance are common in the phytoplankton. These changes are known as phytoplankton succession, and the mechanisms are complex.
• Diatoms early in the successional sequence are autotrophic, requiring only inorganic nutrients for their survival, whereas species later in the successional sequence may be auxotrophic, requiring nutrients such as vitamins that they cannot produce themselves. The auxotrophy of later species suggests that later species cannot reach great abundance until the flowering of earlier species.
• Earlier diatom species often have large cell sizes, which allow them to store nutrients. Later species often are smaller in cell size, and thus the surface area (relative to cell volume) that is exposed to nutrient uptake is increased.
• The depletion of limiting nutrient such as silica may result in demise of the diatoms in late spring and favor phytoplankton groups that do not have a siliceous test.
• Groups such as dinoflagellates may depend on exudates and nutrients produced in the excretion and decomposition of species earlier in the successional sequence. Dinoflagellates are known typically to require more nutrients that they cannot manufacture themselves than is the case for diatoms, which perhaps explains the successional sequence in the plankton.
• Allelopathy-the production of toxic compounds by one organism to inhibit another-may play a role in succession. Cyanobacteria may dominate eutrophic lakes and can inhibit the development of diatom populations. Cell-free filtrates of cyanobacteria cultures inhibit the growth of diatoms isolated from the same lake. Cyanobacteria blooms may thus alternate with diatom outbursts in lakes and some polluted estuaries.
During succession, changes in relative abundance of phytoplankton species occur, but it is important to remember that all species are present at all times of the year. Otherwise there would be no seed population from which a population increase of a given species could develop. In some cases, a population of cysts in the bottom sediment may help initiate blooms of some species.
The diversity of phytoplankton also tends to increase as succession progresses. This may reflect an increasing diversity of nutrient sources, especially an increase in organic substances in the water.
Later phytoplankton successional dominants also are often ornate and adorned with spines. This may be a response to a temporal increase in the presence of predators, or to an increase in surface area/volume ratio, which would increase nutrient uptake efficiency. It may also be a mechanism to induce rotation as the phytoplankton cell is sinking, which may break up the boundary layer around the phytoplankton cells and increase nutrient access.
The pattern of seasonal succession seems generally to reflect the geographic distribution of phytoplankton. Phytoplankton species that bloom early in succession are typical of nutrient-rich (eutrophic) coastal waters, whereas those that occur later dominate nutrient-poor (oligotrophic) offshore environments.
Stages of diatom dominance are most pronounced in mid- to high latitudes and are indistinct in tropical locations.

٦٤
Rate of Nutrient Uptake The relationship between nutrient concentration and uptake rate by phytoplankton. Uptake rate of nutrients by phytoplankton may be measured directly (e.g., in terms of ammonium taken up per cell per unit time), or indirectly (in terms of the cell doubling rate). We assume here, for simplicity, that the faster the nutrient uptake, the faster the cell doubling rate.
Nutrient uptake usually follows the general pattern illustrated in Figure 12, which shows the relationship of cell doublings per day, D (which is an estimate of nutrient uptake, as mentioned earlier), as a function of nutrient concentration, C. The cell doubling rate increases with increasing nutrient concentration but then reaches a plateau, at a value of Dmax. Half saturation concentration, or K: The nutrient concentration at which half the maxi-mum cell doubling rate occurs is known as the half saturation concentration, or K. This is a useful measure of nutrient uptake, which we shall use later.
Fig. 12 Growth rate (D = doublings per day) of the diatom Asterionella japonica (circles) as a function of C (which, in this case, is the nitrate concentration in µg atoms L-1). The half-saturation constant K is estimated at 1.5 by finding the value of C on the green curve that corresponds to Dmax/2.
Patterns of nutrient uptake at different habitats (different nutrient concentrations) Open-ocean environments usually have far lower nutrient concentrations than do inner-shelf waters. As a result, we may expect phytoplankton living in these two different habitats to evolve differing patterns of nutrient uptake. Open-ocean phytoplankton should be able to take up nutrients efficiently from low en-vironmental concentrations. They would be expected, however, to have low maximum

٦٥
uptake rates, because they never encounter high nutrient concentrations. By contrast, inshore or inner-shelf phytoplankton live at higher nutrient concentrations should be relatively inefficient at low nutrient concentrations but able to take up nutrients at far higher nutrient concentrations than the open-ocean phytoplankton. Expected curves for both types are shown in Figure 13.
Fig. 13 A theoretical representation of the differences in nutrient uptake expected for a phytoplankton clone (1) adapted to high nutrient levels near-shore versus another clone (2) adapted to offshore low nutrient levels. Dmax 1 is the maximum doubling rate for clone 1, and K1 is the estimated corresponding value of K (same procedure for clone 2).
The difference leads to a prediction, that, the half-saturation concentration K should be greater for inner-shelf phytoplankton. For coastal phytoplankton, K is usually greater than 1 µ mole/L for nitrate uptake, whereas oceanic phytoplankton have values of about 0.1-0.2 µ mole/L. Clones of the same diatom species show high and low K values, depending on whether the clones are isolated from near-shore or oceanic waters, respectively. Oceanic phytoplankton are more efficient at taking up nutrients at low nutrient concentration. Thus, they may be competitively superior to coastal forms in the low nutrient concentrations of the open sea but unable to take up high concentrations. Differences in nutrient limitation among competing species may also contribute to coexistence. For example, species 1 may be superior at phosphate uptake whereas species 2 may be superior at silica uptake. Thus, under conditions of varying nutrient concentration different species may be favored. Such coexistence has been shown for lake phytoplankton.
Can interspecific competition for nutrients determine the success of some phytoplankton species relative to others? Many phytoplankton species seem to coexist with no competitive exclusion and similar nutrient preferences. This coexistence has been characterized by the ecologist G. Evelyn Hutchinson as "the paradox of the plankton."
Conditions in the ocean may be too variable and may shift too rapidly for one species to drive others to extinction by means of greater efficiency of nutrient uptake. Many species may be relatively more efficient under conditions that change periodically.

٦٦
Zooplankton Grazing
Grazing rate (G) is the volume of water from which cells (prey) are removed per unit time by a single zooplankter. We can say for example that the grazing rate per individual is 20 µl h-1. Synonyms include searching rat، filtering rate, filtration rate, clearance rate.
The impact of zooplankton grazing on phytoplankton abundance is quite variable. - Zooplankton usually increase after the peak of the phytoplankton bloom has passed,
owing to • the time lag in reproduction of the zooplankton; Zooplankton such as copepods have
a much longer generation time (weeks) than phytoplankton (hours to days), • the grazing down of the phytoplankton standing crop as the zooplankton population
increases. A large population of zooplankton could graze a phytoplankton population to near-extinction. This would cause a collapse of the zooplankton until the phytoplankton recovered. Such overgrazing would thus result in strong oscillations in the zooplankton population.
When the phytoplankton organisms reach very low densities, however, the zooplankton may not be able to find them. A low density refuge for phytoplankton allows a subsequent increase when zooplankton decrease in abundance, and the severity of zooplankton oscillations may diminish.
- For a brief time, the advent of zooplankton may even stimulate phytoplankton growth, owing to animal excretion. Eventually, zooplankton may cause significant reductions in the phytoplankton
Spatial alternations of abundance of phytoplankton and zooplankton Spatial alternations of abundance of phytoplankton and zooplankton occur in some regions. This would suggest that an area currently rich in zooplankton but poor in phytoplankton had been rich in phytoplankton until the zooplankton grazed them down. Spatial patterns of alternation between phytoplankton and zooplankton are therefore indications of temporal patterns of phytoplankton growth and subsequent zooplankton population growth and strong grazing. In the North Sea, alternating areas of phytoplankton and zooplankton dominance occur, suggesting cycles of dominance by phytoplankton production and then by grazing (Figure 14).

٦٧
Fig. 14 Distribution of chlorophyll a and copepod carbon on a survey in the North Sea, showing an inverse relationship between phytoplankton and zooplankton standing stock. Phytoplankton are most abundant toward the left, whereas zooplankton are most abundant toward the right.

٦٨
Zooplankton Feeding Feeding rate (f), or ingestion rate, is the number of cells (prey) eaten per unit time per animal. It is usually expressed in cells ingested individual-1 time-1.
Feeding behavior of zooplankton varies with phytoplankton cell concentration (cells per unit volume). Figure 15 shows the relationship between phytoplankton cell concentration and ingestion rate I, which is the cells ingested per animal per hour (cells ingested individual-1 time-1). I increases linearly with cell concentration until a maximum is reached, whereupon no further increases in ingestion occurs. Below the maximum, the feeding response of the copepod is indicative of an animal that searches, encounters, and feeds on particles in direct proportion to their concentration. Then there appears to be a saturation level, and ingestion increases no further. Feeding may cease because the feeding appendages are saturated with food, or because digestion efficiency would decrease if material were moved through the gut any faster. There is also some evidence for a lower threshold, below which the copepod does not expend energy to search for food and to feed.
Fig. 15 Ingestion rate I (cell. copepod-1h-1) for the copepod Callous Pacific's a. a function of diatom cull concentration. (After Frost, 1972.) Feeding behavior of zooplankton varies with phytoplankton cell size and species type. Cell size and species type also influence feeding in copepods and other zooplankton. There is some evidence that copepods can shift the preferred particle size, depending upon the abundance of various food particles in the environment. Copepods feed more rapidly on two-celled chains of diatoms than on single cells, and also feed at higher rates on diatom species with larger cell size, relative to smaller cells. As mentioned earlier, copepod feeding involves movement of feeding appendages. Under viscous conditions of the water it is not possible to strain phytoplankton cells out of the water on a sieve. Instead, feeding appendages actively select different phytoplankton cells. In copepods, selection of particles involves chemical detection, and the animals tend to avoid inert particles, relative to live phytoplankton cells. Many zooplankton avoid toxic phytoplankton, but tintinnids actually feed preferentially on these forms.