document
TRANSCRIPT

Zbl. Bakt. Hyg., 1. Abt, Orig. C 2, 339-348 (1981)
Institute of Fisheries, University of Tromse Norway
Biochemical Ecology of Psychrotrophic Strains ofVibrioanguillarum Isolated from Outbreaks of Vibriosis at Low
Temperature
JAN A. OLAFSEN, MARIANNE CHRISTIE!, and JAN RAA
Received May 5, 1981
Summary
Four strains of the fish pathogen Vibrio anguillarum have been isolated from rainbowtrout (Sa/rna trutta}, salmon (Sa/rna salar}, coalfish (Gadus virens) and eel (Anguillaanguilla) kept captive at water temperatures of 1-4 DC for aquaculture or other purposesin the vicinity of Tromso Norway.
Biochemically, they all closely resemble a reference strain, V. anguillarurn NCMB 6.The only important exception was that our strains have lower cardinal temperatures forgrowth, namely 0 DC (minimum), 15-18 DC (optimum) and below 30 DC (maximum),as opposed to an optimum of 30 DC and a maximum of 30-40 DC, respectively, for thereference strain.
The 4 isolates grew optimally in media containing 2 % NaCI, with outer limits forgrowth at 0.5 Ofo and 5 Ofo NaCI. The viability was quickly lost in nutrient media withoutinorganic salt, in membrane-filtered seawater and by freezing or freeze-drying. Theouter pH limits for growth were 5 and 9, respectively. Our strains showed enhancedanaerobic growth yields in the presence of trimethylamine oxide (TMAO).
When examined with immunochemical methods all the strains, including the referencestrain, showed serological cross-reaction, but they clearly differed in antigenic structure.
Key words: Vibrio anguillarum - Vibriosis - Fish pathogen - Psychrotrophic Isolation - Characterization - Biochemical ecology
Introduction
Vibriosis, originally called the "Red Pest Disease" is one of the most frequentlyreported fish diseases. The causative bacterium Vibrio anguillarum has a worldwide distribution and according to Bullock and McLaughlin (1970), outbreaks ofvibriosis have been reported from at least 14 countries, affecting 48 species of fish.The fish species most commonly involved are trout, salmon, plaice, cod, coalfish,
1 Present address: Gades Institute, Department of Microbiology, University of Bergen,Bergen, Norway.
Abbreviations: TMAO = trimethylamine oxide

340 J. A.Olafsen, M. Christie, and J. Raa
herring and eel (Anderson and Conroy, 1970; Evelyn, 1971; Hdstein and Holt,1972; Wolke, 1975).
Vibriosis is primarily a salt water disease, the bacterium being dependent onNaCI for growth, but outbreaks in fresh water (Hacking and Budd, 1971; Rosset aI., 1968) have been reported. In the latter case, however, the isolated bacteriumwas found to require NaCI for growth.
Outbreaks of vibriosis are often of epizootic proportions, and are of economicimportance when they affect fish used for human consumption. There have beenreports of mass mortality of free-living fish due to vibriosis. Foerster (1941) andTester (1942) described epizootics of sardines and herring, respectively; and inNorwegian fjords mass mortalities among yearlings of coalfish due to vibriosishave been reported by Egidius and Andersen (1975). Accordingly, vibriosis is amajor threat to marine fish farming.
The biochemical characteristics of different strains of V. anguillarum have beendescribed by Hendrie et al. (1971) and Evelyn (1971). The latter author proposedan emerging archetype, and a widely accepted scheme for identification was proposed by Levin et al. (1972).
Different biotypes (Smith, 1961) and serotypes (Kiehn and Pacha, 1969) of V.anguillarum have been described. However, it is not known whether these differences affect the manifestation of pathogenicity with respect to different speciesof fish.
Vibriosis is a systemic disease which affects most organs, with symptoms asdescribed by Wolke (1975). Death due to vibriosis may, however, occur withoutany visible disease symptoms. Moreover, V.anguillarum is facultatively pathogenic,and can be isolated from fish without any signs of disease (Mattheis, 1964; Rodscether et aI., 1977).
V. anguillarum is referred to as an opportunistic pathogen (Bullock and McLaughlin, 1970) and there are indications that disease outbreaks are usually initiated by alteration in the host-pathogen relationship, frequently caused by environmental changes (Wedemeyer, 1970). It is our experience from the Tromseregion in the northern part of Norway that vibriosis outbreaks are more frequentin the winter period. The fish may under these conditions be sufficiently weakenedto allow local strains of V. anguillarum, which are able to grow at low temperature, to attack successfully. The present paper reports outbreaks of vibriosis atlow temperature, and describes some biochemical and serological properties relatedto the growth, survival and ecology of the strains involved.
Materials and Methods
Isolation of the bacterium
Strains of V. anguillarum from rainbow trout, salmon, coalfish and eels were isolatedfrom blood samples withdrawn from the heart of living fish with clear disease symptoms.
The strains from rainbow trout and salmon (V. anguillarum UTC 100-1 and UTC100-3) were isolated from a marine fish farm at Hakeya near Trornse. The fish had beenkept in free-floating (120 m2 X 4 m) nets in the sea, and samples were taken duringJanuary and February after outbreaks of vibriosis. The seawater temperature was then1-2°C. The fish affected by the disease were yearlings of rainbow trout and salmonthat had been transferred to seawater during the summer and autumn, and the weightwas 100-400 g. Outbreaks of vibriosis occurred among the salmon and rainbow trout

Biochemical Ecology of Psychrotrophic Strains of Vibrio anguillarum 341
in this fish farm during the winter in three subsequent years (1973-75), whereas no summer-outbreaks were recorded.
V. anguillarum UTC 100-4 was isolated from coalfish held in floating nets at Sornmarey near Trornse for commercial purposes. These fish had been kept alive in densepopulations from late summer until winter, when they got vibriosis. The water temperature at that time was 3-4 0c.
The strain V. anguillarum UTC 100-2 was isolated from eels caught in brackish water(Rossfjordvarn near Trornse) and kept in tanks supplied with running fresh water at2-4 °c , as described by Redscetheret al. (1977).
Characterization and growth conditions of the different strains
Blood samples were plated on Bacto Tryptose Blood Agar Base (Difco) containmg5 Ofo (v/v) human erythrocytes and 30 g NaClIl in order to examine the haemolytic property. The identification of the bacterium was carried out according to the scheme ofLevin et al. (1972), as described by Redscether et al. (1977).
The growth medium (Vibrio-medium A) contained, in g/I: yeast extract (0.5), glucose(0.1) and trypton (2.0) in a mineral medium containing NaCI (20), NH.CI (1.0), (NH.)2S0.(1.0), MgSO.· 7H20 (0.1), KH2P0 4 (1.5), NaH2P0 4 (0.5) and Na2HP04 (1.6) in tapwater. pH was adjusted to 7, unless otherwise stated. Growth rate and growth yield(maximum turbidity) was measured in a nephelometer.
lmmunochemical methods
Antiserum was prepared by injection of killed V. anguillarum cells in a rabbit. Lyophilized , washed V. anguillarum cells were suspended in sodium phosphate buffer (0.01molll, pH 7.3) containing 9 g NaClII, mixed with an equal volume of Freunds completeadjuvant (Behring Werke A.G., Marburg Lahn, W. Germany) . This was injected subcutaneously (1 mg/ml) followed by 6 weekly intramuscular injections (2, 4, 6, 8, 10, and20 mg/ml) and a booster dose (20 mg/ml) 3 weeks later. The rabbit was bled one weekafter the 7th injection, and the serum stored at -20 °C.
The double diffusion precipitation test (Ouchterlony, 1949) was carried out in agar(12 gil) containing 8.5 g NaClIl. The wells had a diameter of 4 mm, and precipitationlines were recorded after 48 h at 20 °C. The slide agglutination test was used as describedby Conroy and Withnell (1974) with monovalent antisera. The indirect haemagglutination test with sheep erythrocytes treated with a solution of 6.25 mg tannic acid/I (E Merck ,Darmstadt, W. Germany, was performed as described by Tender and Redscether (1974).The treated erythrocytes were mixed with 10 mg of lyophilized bacteria/m!.
Infe ction studies
Infection studies were carried out with groups of five rainbow trout, each weighing100-400 g in two 500 I concrete tanks with aerated sea-water at 4-6 °C. The bacteriawere washed once by centrifugation at 3,000 X g, and resuspended in 20 g NaClIl. Theexperimental fish were injected with 1 ml of 107 bacteria in 20 g NaClII, whereas thecontrol group received 1 ml of 20 g NaCI /1.
Results
Four strains of V. anguillarum were obtained in pure culture by the samplingmethods described. Electron microscopy of the isolates revealed comma-shaped,polarly flagellated cells. The isolated strains were characterized biochemicallyaccording to Levin et aI. (1972), and compared with the reference strain NCMB 6(Hendrie et aI., 1971) from cod (Gadus callarias] (Table 1). The four isolated

342 ].A.Olafsen, M.Christie, and ].Raa
Table 1. Biochemical properties of Vibrio anguillarum isolated from rainbow trout,salmon, eel and coalfish; and of the reference strain NCMB 6. Tests were carried out asdescribed by Redstetber et aI. (1977)
Our isolates Referencestrain
UTC 100 NCMB 6Strain 1-4
Gram strainMotilityGrowth at 0 DCGrowth at 30 DCGrowth in 00/0 NaClGrowth in 8 % NaClOxidase (Kovacs)AmylaseGelatinaseArginine dihydrolaseHaemolysisIndole formation from tryptophanNitrate reductaseVoges-ProskauerGlucose fermentationMannitol fermentationSucrose fermentation "-Galactose fermentationSorbitol fermentationSensitivity to novobiocinSensitivity to 0/129 (2.4 diamino-6,7-diisopropylptcridin)Sensitivity to neomycin
++
+++++++
++
+++-1-
+
+
+
++++++++++++++++
Strain UTC 100-1 isolated from rainbow trout (Salmo gairdneri)Strain UTC 100-2 isolated from eel (Anguilla anguilla)Strain UTC 100-3 isolated from salmon (Salmo salar)Strain UTC 100-4 isolated from coalfish (Gadus virens)Reference strain NCMB 6 isolated from cod (Gadus callarias),,- Strain UTC 100-1 and UTC 100-3 fermented sucrose, whereas strain UTC 100-2 and
UTC 100-4 did not.
strains had the same physiological and biochemical properties, except for theability to ferment sucrose. Unlike the reference strain, our cold water isolates grewat 0 DC, but not at 30 DC. They also gave negative Voges-Proskauer reaction, aproperty which is variable between V. anguillarum strains (Evelyn, 1971).
Immune serum against V. anguillarum UTC 100-1 did not agglutinate any ofthe other isolates, using the slide agglutination test. The double diffusion precipitation test revealed differences in antigenic structure between different isolates ofV. anguillarum (Fig. 1). However, all the strains except that from coalfish haverelated or partly related antigenic determinants, the reference strain V. anguillarumNCMB 6 being most like the strain UTC 100-1, to which the antiserum was made.

Biochemical Ecology of Psychrotrophic Strains of Vibrio anguilla rum 343
®®
Fig. 1. Precipitation lines of different V. anguillarum strains tested in double diffusionin agar against rabbit immune serum to V. anguillarum UTC 100-1 (Is).1: V. anguillarum UTC 100-1 (from rainbow trout),2: V. anguillarum UTC 100-2 (from eel),3: V. anguillarum UTC 100-3 (from salmon),4: V. anguillarum UTC 100-4 (from coalfish),6: V. anguilla rum NCMB 6 (from cod),
10: V. anguilla rum HI 10 (from rainbow trout in the south of Norway).
The indirect haemagglutination assay with tannin-treated sheep erythrocytes alsoshowed some cross reactions among all strains.
Blood serum from fish with vibriosis as well as healthy fish caught in the coldenvironment near Tromse and tested in double diffusion in agar, gave a precipitation line with all the strains of V. anguillarum we have isolated from this environment, but not with the reference strain NCMB 6, nor a strain HI 10 (Instituteof Marine Research Culture Collection, strain HI 10, Bergen, Norway) isolatedfrom rainbow trout in the south of Norway. This precipitation factor in the fishserum may have been naturally induced by exposure to the local strains of thepathogen.
Fig. 2 shows growth curves and maximum growth of V. anguillarum UTC100-1 at different NaCI concentrations. The shortest lag-period of growth and thehighest growth rate resulted at about 2 % NaCl, with outer limits for growth atabout 0.5 and 5010 NaCl, but survival between 0.2 and 9010. In distilled water,or in nutrient media with no inorganic salts added, it lost its viability very quickly;a suspension of 109 bacteria per ml distilled water was sterile after 30 min at20 °C. The bacterium had an appreciable viability in heat-sterilized sea water,but not if the water was filtered through a 0.45 f-l membrane filter (Fig. 5).
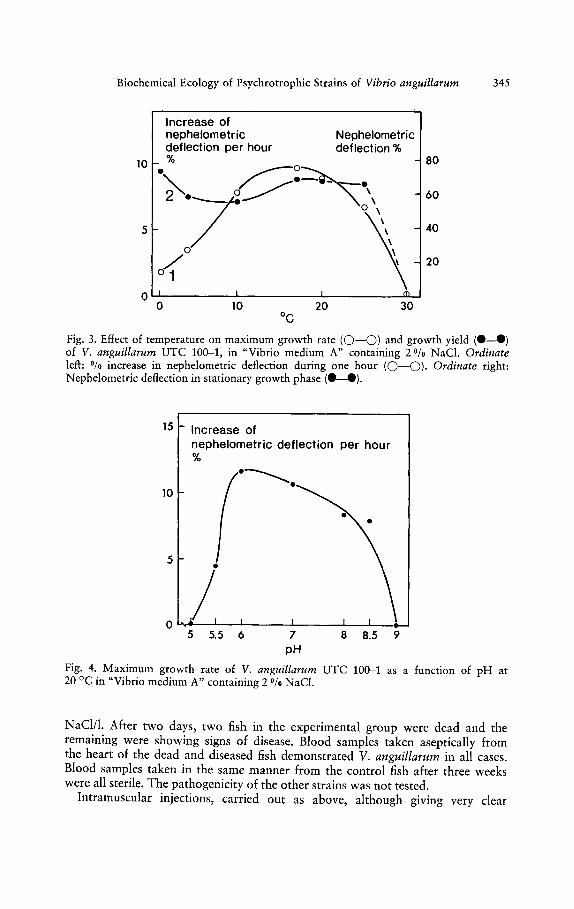
The maximum temperature for growth was slightly below 30 °C, optimum atabout 15 °C and minimum at O°C (Fig. 3). The growth yield (as turbidity) wasalmost the same at all the temperatures permitting growth.
Growth was maximal at neutral and slightly acidic conditions (Fig. 4). Theouter pH limits for growth were 5 and 9. Outside these limits the bacterium lostits viability very quickly.
Freezing and freeze-drying drastically reduced the viability of V. anguilla rum.At - 20 °C in the growth medium there was a 50010 reduction in the viable countafter 15 h, and the viability was gradually lost during subsequent storage at thistemperature.

~44 ].A.Olafsen, M.Christie, and ].Raa
0.6'10
10 15Time (hours)
Nephelometric ._----0-0-0 2 '10
deflection ~
% /I
~ /0 ••"3.5'10\CJ .......I / ..0.0 O.S'!
o ! /0/ / 0/
./ /,o'r/20
80
40
60
15
10
Increase ofnephelom.deflectionper hour%
®
."."•
5
•J
I•,i•oL..-__....l-__---l. .L-__--"---'
o 2 3 4% NaCI
Fig. 2. Effect of NaCl on growth of V. anguillarum UTC 100-1 in "Vibrio medium A"at 20°C. Fig. (A) shows growth curves, and fig. (B) maximum growth rates at differentNaCl concentrations.
V. anguillarum is facultatively anaerobic. The anaerobic growth in the presenceand absence of TMAO was measured as described by Strem et al. (1979). V.anguillarum UTC 100-1 was grown in Vibrio medium A containing 20 mmolglucosell to which was added 60 mmol TMAO/l. The growth yield in the presenceof TMAO was considerably higher than in its absence, namely 82 and 38, respectively (measured as 0/ 0 nephelometric deflection), which is in accordance withthe results reported by Strem et al. (1979) for Proteus sp.
Infection studies were carried out in order to assess the virulence of the strainisolated from rainbow trout. Five rainbow trout were injected intraperitoneaJlywith 107 live bacteria in 20 g NaCIII and 5 control fish were injected with 20 g
Biochemical Ecology of Psychrotrophic Strains of Vibrio anguillarum 345
80
o
Increase ofnephelometric Nephelometricdeflection per hour deflection %
10 I- %
'~ ~~.
S" 2 '7 0\\\ :::o \
/ ~ 2001
I ,;,
10 20 30°c
Fig. 3. Effect of temperature on maximum growth rate (0-0) and growth yield (e-e)of V. anguillarum UTC 100-1, in "Vibrio medium A" containing 2 % NaC!. Ordinateleft: % increase in nephelometric deflection during one hour (0-0). Ordinate right:Nephelometric deflection in stationary growth phase (e ).
15 Increase ofnephelometric deflection per hour%
8 8.5 95 5.5 6 7pH
Fig. 4. Maximum growth rate of V. anguillarum UTC 100-1 as a function of pH at20 °C in "Vibrio medium A" containing 2 Ofo NaCI.
NaCl/I. After two days, two fish in the experimental group were dead and theremaining were showing signs of disease. Blood samples taken aseptically fromthe heart of the dead and diseased fish demonstrated V. anguillarum in all cases.Blood samples taken in the same manner from the control fish after three weekswere all sterile. The pathogenicity of the other strains was not tested.
Intramuscular injections, carried out as above, although giving very clear

346 J.A.Olafsen, M.Christie, and J.Raa
8
6
•
10
•
9
•
832
--:!~,. ------h........... h. -"i.r--::II:i-......,,-......::~.......
"""""",,\
~\
oo
2
cCIa
III
.Do 4>
4 5 6 7Time (weeks)
Fig. 5. Survival of V. anguillarum UTC 100-1 in heat sterilized sea water(e-e) andin heat sterilized sea water filtered through a 0.45 fl membrane filter (6-6) at 20 DC,
disease symptoms in the course of time, did not kill the fish within a week. Thediseased fish were then sacrificed, and V. anguillarum was demonstrated in allthe blood samples.
Discussion
Anderson and Conroy (1970) claimed that vibriosis was more common duringthe warmer months of the year, and according to Roberts (1975) outbreaks ofvibriosis were often closely related to an increase in the environmental temperature. However, our psychrotrophic strains of Vibrio anguillarum have been isolated from fish kept captive in sea water at 1-4 DC, and our strain V. anguillarumUTC 100-1, isolated from rainbow trout, is pathogenic to this fish species wheninjected intraperitoneally at 4-6 DC. In addition to this, it is our experience fromthe arctic region of Norway that fish are more susceptible to vibriosis when thewater temperature is extremely low. The psychrotrophic strains of V. anguillarumare well adapted to these temperatures, whereas the immune system of the fishmay not function optimally tHildeman, 1962).
V. anguillarum is an opportunistic pathogen that can cause disease when thefish is stressed, for example due to high population density, excessive handling(Wedemeyer, 1970) copper- (Rodscether et al., 1977) or crude-oil pollution (Minchew and Yarbrough, 1977). Our results suggest that also low water temperaturesmay render the fish susceptible to vibriosis.
The four isolates have closely related biochemical properties, and show closeresemblance to the reference strain NCMB 6. When examined with immuno-

Biochemical Ecology of Psychrotrophic Strains of Vibrio anguillarum 347
chemical methods our strains, and the reference strain, showed some cross-reaction, but with clearly different antigenic patterns, which is in accordance withAnderson and Conroy (1970). It seems likely, therefore, that immunization offish with V. anguillarum antigens will only be effective against resident strains inthe affected fish stock.
Our strains of V. anguillarum survive better if particles (detritus) are presentin the sea water. This is consistent with the findings of Simudu et aI. (1971), whodemonstrated that marine Vibrio species were most frequently bound to particlesin the sea.
V. anguillarum is facultatively anaerobic, and exhibits enhanced anaerobicgrowth yields in the presence of TMAO. Anaerobic respiration with TMAO aselectron acceptor has been demonstrated among several marine bacteria iStrern,1971; Strem et aI., 1979). Growth of V. anguillarum should accordingly befavoured in the vicinity of marine fish farms, where food waste rich in TMAOand carbohydrate collects into anaerobic sediments. This, as well as the markedability of V. anguillarum to utilize sugar, may explain why this bacterium wasfound to be the predominant organism in sugar-polluted marine sediments (.JuulJensen and Larsen, 1976).
Acknowledgement
We wish to express our gratitude to the late Professor Einar Bruun, University ofTromse, for his help and interest in supplying fish for these studies. The skillful technicalassistance of Miss Edel Olsen is greatly appreciated.
The project has been supported by the Norwegian Council for Fishery Research(NFFR).
References
Anderson, ]. I. W., Conroy, D. A.: Vibrio diseases in marine fishes. In: A symposium ondiseases of fishes and shellfishes (Ed. Snieszko, S. F.), pp. 266-272, American FisheriesSociety, special publication No.5. Washington/D.C. 1970
Bullock, G. L., McLaughlin, ]. ]. A.: Advances in knowledge concerning bacteria pathogenic to fishes (1954-1968). In: A symposium on diseases of fishes and shellfishes (Ed.Snieszko, S. F.), pp. 231-242. American Fisheries Society, Special publication No.5.WashingtonlD.C.1970
Conroy, D. A., Withnell, G. C.: The use of a slide agglutination test as an aid in thediagnosis of Vibrio diseases in fish. Riv. Ital. Piscicolthena de Ittiopatol. Anno IX, nr. 3,69-74 (1974)
Egidius, E., Andersen, K. (1975). Smaseidoden i 1974 - vibriose. (An epizootic of vibriosisin young saithe in 1974). Fiskets Gang 22, 362-365 (1975) (In Norwegian).
Evelyn, T. P. T.: First records of vibriosis in Pacific salmon cultured in Canada, andtaxonomic status of the responsible bacterium Vibrio anguillarum. J. Fish. Res. Bd.Can. 28, 517-525 (1971)
Foerster, R. E.: The mortality of young pilchards 1941. Fish. Res. Bd. Can. Progr. Rep.Pacific Coast Station 48, 3-8 (1941)
Hacking, M. A., Budd, [: Vibrio infection in tropical fish in a freshwater aquarium. J.Wildt. Dis. 7,273-280 (1971)
Hendrie, M. S., Hodgkiss, W., Shewan, ]. M.: Proposal that the species Vibrio anguillarum
24 Zbl. Bakt. Hyg., I. Abt. Orig. C 2

348 J.A. Olafsen, M. Christie, and J.Raa
Bergman 1909, Vibrio piscium David 1927 and Vibrio ichthyodermis (Wells and ZoBell)Shewan, Hobbs and Hodgkiss 1960 be combined as a single species, Vibrio anguillarum.Int. J. system. Bact. 21, 64-68 (1971)
Hildeman, W. H.: Immunogenetic studies of poikilothermic animals. Amer. Nat. 46,195-204 (1962)
Hdstein, T., Holt, G.: The occurrence of vibrio disease in wild Norwegian fish. J. FishBioI. 4, 33-37 (1972)
Juul Jensen, N., Larsen, j. L.: Forskningsprosjekt vedrorende marine vibriopopulationersforhold til forurensningskomponenter, isser kullhydratforurensning i marine biopter.Dansk Vet. Tidsskr. 59 (12),521-524 (1976) (in Danish)
Kiehn, E. D., Pacha, R. E.: Characterization and relatedness of marine vibrios pathogenicto fish: deoxyribonucleic acid homology and base composition. J. Bact. 100, 1248-1255(1969)
Levin, M. A., Wolke, R. E., Cabelli, V. j.: Vibrio anguillarum as a cause of disease inwinter flounder (Pseudopleuronectes americanus). Canad. J. Microbiol. 18, 1585-1592(1972)
Mattheis, T.: Das Vorkommen von Vibrio anguillarum in Ostseefischen. Z. Fischer. N. F.XII, 259-263 (1964)
Minchew, C. D., Yarbrough, J. D.: The occurrence of fin rot in mullet (Mugil cephalus)associated with crude oil contamination of an estuarine pond ecosystem. J. Fish. BioI.10,319-323 (1977)
Ouchterlony, 0.: Antigen-antibody reactions in gels. Acta path. micro bioI. scand. 26,507-515 (1949)
Roberts, R. j.: The effects of temperature on diseases and their histopathological manifestations in fish. In: The pathology of fishes. (Eds. Ribelin, W. E., Migaki, G.), pp. 447to 479. Wisconsin, University of Wisconsin Press 1975
Ross, A. J., Martin, j. E., Bressler, V.: Vibrio anguillarum from an epizootic in rainbowtrout (Salmo gairdneri) in U.S.A. Bull. Off. Intern. Epizoot. 69, 1139-1148 (1968)
Redsaither, M. C., Olafsen, j. A., Raa, j., Myhre, K., Steen, j. B.: Copper as an initiatingfactor of vibriosis (Vibrio anguillarum) in eel (Anguilla anguilla). J. Fish BioI. 10,17-21 (1977)
Simudu, U., Ashino, K., Kaneko, E.: Bacterial flora of phyto- and zoo-plankton in theinshore water of Japan. Canad. J. Microbiol. 17, 1157-1160 (1971)
Smith, 1. W.: A disease of finnock due to Vibrio anguillarum. J. gen. Microbiol, 24, 247to 252 (1961)
Strom, A. R.: Bakterielle omsetninger i anaerobt sildeekstrakt. Thesis, Department ofBiochemistry, Norwegian Institute ofTechnology, University of Trondheim, Norway1971 (In Norwegian with English summary)
Strem, A. R., Olafsen, j. A., Larsen, H.: Trimethylamine oxide: A terminal electronacceptor in anaerobic respiration of bacteria. J. gen. Microbiol. 112, 315-320 (1979)
Tester, A. L.: Herring mortality along the south-east coast of Vancouver Island. FishRes. Bd. Can. Progr. Rep. Pacific Coast Station 52, 11-15 (1942)
Tender, 0., Redscetber, M.: Indirect haemagglutination for demonstration of antibodiesto Aspergillus fumigatus. Acta path. microbiol. scand. 82B (6),871-878 (1974)
Wedemeyer, 0.: The role of stress in the disease resistance of fish. In: A symposium ondiseases of fishes and shellfishes (Ed. Snieszko, S. P.), pp. 30-35. American FisheriesSociety, Special publication No.5. WashingtonlD.C. 1970
Wolke, R. E.: Pathology of bacterial and fungal diseases affecting fish. In: The pathologyof fishes (Eds. Ribelin, W. E., Migaki, G.), pp. 33-116. Wisconsin, University of Wisconsin Press 1975
Prof. Jan Raa, Institute of Fisheries, University of Tromse, P. O. Box 488, N-9001Tromse, Norway