a dynamic, mitotic-like mechanism for bacterial …genesdev.cshlp.org/content/20/23/3269.full.pdfa...
TRANSCRIPT

A dynamic, mitotic-like mechanismfor bacterial chromosome segregationMichael A. Fogel and Matthew K. Waldor1
Genetics Program, Tufts University School of Medicine, and Howard Hughes Medical Institute,Boston, Massachusetts 02111, USA
The mechanisms that mediate chromosome segregation in bacteria are poorly understood. Despite evidence ofdynamic movement of chromosome regions, to date, mitotic-like mechanisms that act on the bacterialchromosome have not been demonstrated. Here we provide evidence that the Vibrio cholerae ParAI and ParBIproteins are components of an apparatus that pulls the origin region of the large V. cholerae chromosome tothe cell pole and anchors it there. ParBI interacts with a conserved origin-proximal, centromere-like site(parSI) that, following chromosome replication, segregates asymmetrically from one pole to the other. Whilesegregating, parSI stretches far away from neighboring chromosomal loci. ParAI forms a dynamic band thatextends from the pole to the segregating ParBI/parSI complex. Movement of ParBI/parSI across the cell occursin concert with ParAI retraction. Deletion of parAI disrupts proper origin localization and segregationdynamics, and parSI no longer separates from nearby regions. These data suggest that ParAI forms a dynamicstructure that pulls the ParBI-bound chromosome to the pole in a process analogous to anaphase of eukaryoticmitosis.
[Keywords: Vibrio cholerae; chromosome segregation; ParA; ParB]
Supplemental material is available at http://www.genesdev.org.
Received September 25, 2006; revised version accepted October 18, 2006.
All dividing cells must have mechanisms to ensure thattheir genomes are faithfully segregated to daughter cells.While the conserved mitotic apparatus used by eukary-otes to direct chromosome segregation is well defined,relatively little is certain regarding the mechanisms thatmediate chromosome segregation in prokaryotes. In re-cent years, it has become clear that bacterial chromo-somes are both highly organized and dynamically segre-gated. Particular chromosomal loci occupy specific sub-cellular locations (Gordon et al. 1997; Webb et al. 1997),and the relative spatial arrangement of most chromo-somal loci correlates with their linear map positions(Teleman et al. 1998; Niki et al. 2000; Viollier et al.2004). Specific chromosomal regions, particularly thoseclose to the origin of replication, are actively and rapidlypositioned in the cell (Glaser et al. 1997; Webb et al.1998; Jensen and Shapiro 1999). The observed move-ments are more rapid than could be accounted for byprevious models of passive segregation mediated bychromosome attachment to the growing cell membrane(Jacob et al. 1963).
Several mechanisms may contribute to the dynamicmovements of bacterial chromosomes (for reviews, seeErrington et al. 2005; Leonard et al. 2005). The observa-
tion in Bacillus subtilis that the DNA replication ma-chinery appears to be localized as a stationary “factory”at the cell center (Lemon and Grossman 1998) led to theproposal that DNA polymerase could provide the bidi-rectional force for segregation (Lemon and Grossman2000). Similarly, RNA polymerase, acting on direction-ally biased genes near the origin, has been hypothesizedto impart both motive force and directionality to segre-gation (Dworkin and Losick 2002; Kruse et al. 2006).While both of these models could potentially explain thesymmetric and bidirectional segregation of the Esche-richia coli and B. subtilis chromosomes from the middleof the cell, it is difficult to directly apply them to theasymmetric segregation process that occurs in Caulo-bacter crescentus (Mohl and Gober 1997; Viollier et al.2004) and in Vibrio cholerae (Fogel and Waldor 2005). Inboth of these organisms, the origin region is located closeto one pole (the “old” pole) early in the cell cycle, andafter replication, one copy remains at that pole while theother traverses the entire length of the cell to the oppo-site (“new”) pole. Consistent with an asymmetric segre-gation pattern from the pole, the replication machineryof C. crescentus localizes to the old pole, and then afterreplication initiation, migrates to the cell center (Jensenet al. 2001). The presence of a moving replisome sug-gested a modification of previous models in which theorigin regions are positioned rapidly by an origin-specificmechanism, and then the bulk of the chromosome is
1Corresponding author.E-MAIL [email protected]; FAX (617) 636-2723.Article is online at http://www.genesdev.org/cgi/doi/10.1101/gad.1496506.
GENES & DEVELOPMENT 20:3269–3282 © 2006 by Cold Spring Harbor Laboratory Press ISSN 0890-9369/06; www.genesdev.org 3269
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

segregated by a replication factory in directions estab-lished by the positioning of the origins at the poles(Jensen et al. 2001).
Most bacterial chromosomes encode orthologs of plas-mid partitioning (Par) proteins near their origins (Gerdeset al. 2000). In plasmids, par loci consist of three com-ponents: a DNA-binding protein (often termed ParB), anATPase (ParA), and a centromere-like site (parS). ParBbinds parS and spreads along the DNA, forming a largenucleoprotein complex. Formation of this complex andits interaction with ParA are required for efficient plas-mid segregation (Ebersbach and Gerdes 2005; Leonard etal. 2005). The Par-family ATPases fall into two distinctphylogenetic groups; type I ParAs contain the conservedWalker-box ATP-binding motif, whereas type II ParAsare structurally related to eukaryotic actin (Gerdes et al.2000). Types I and II ParAs are found in different plasmidfamilies, but only type I par loci have been identified onbacterial chromosomes (Gerdes et al. 2000). Both type Iand type II ParAs form ATP-dependent filamentous poly-mers in vitro (Møller-Jensen et al. 2002; Barillà et al.2005; Lim et al. 2005). Type II ParAs appear to mediateplasmid segregation by polymerizing between plasmidpairs and “pushing” them apart toward the poles(Møller-Jensen et al. 2003). The mechanism by whichtype I plasmid ParAs function is less clear. Type I ParAsfrom some plasmids appear to oscillate back and forth inthe cell (Ebersbach and Gerdes 2001; Lim et al. 2005;Adachi et al. 2006), but it is unknown how oscillationpositions plasmids. Recently, a plasmid ParA was shownto polymerize into radial filaments on ParB-bound DNAin vitro, and a model was proposed in which plasmids arepositioned by ParA pushing in all directions in the cell(Lim et al. 2005).
While the essential role of par loci in plasmid parti-tioning has been long appreciated, their functions in bac-terial chromosome biology is less clear. The B. subtilisPar proteins Soj (ParA) and Spo0J (ParB) are nonessentialbut have effects on chromosome segregation (Ireton et al.1994; Sharpe and Errington 1996; Lee et al. 2003; Wu andErrington 2003; Lee and Grossman 2006). Spo0J binds toat least 8 sites in a large region around the origin (Lin andGrossman 1998), and deletion of spo0J results in an in-creased frequency of anucleate cells (Ireton et al. 1994).Together, Spo0J and Soj appear to facilitate efficientseparation of newly duplicated origins (Lee and Gross-man 2006), but a mechanistic understanding of their rolein chromosome segregation remains to be defined. Incontrast to B. subtilis, the C. crescentus ParA and ParBare essential, and their overexpression or depletion re-sults in defects in cell growth, division, and chromosomesegregation (Mohl and Gober 1997; Mohl et al. 2001).ParB of C. crescentus binds to sites near the origin of thechromosome (Mohl et al. 2001) and localizes as foci atthe extreme poles (Mohl and Gober 1997). While C. cres-centus ParA and ParB affect both cell division and chro-mosome segregation, recent evidence suggests that thecell division defects are due to the effects of chromosomesegregation on the localization of MipZ, an inhibitor ofFtsZ ring formation (Thanbichler and Shapiro 2006).
Thus, while there is strong evidence in several bacteriafor active movement of chromosomal DNA during seg-regation, specific molecular mechanisms that mediatesuch movements have not been clearly elucidated.
V. cholerae is a curved, Gram-negative rod that causesthe severe diarrheal disease cholera. It contains two cir-cular chromosomes, 2.96 and 1.07 Mb (chrI and chrII,respectively). Despite having coordinated replication ini-tiation (Egan et al. 2004), the segregation of the two chro-mosomes is likely governed by distinct mechanisms asthe localization and segregation patterns of their originregions are different (Fogel and Waldor 2005; Fiebig et al.2006). The origin region of chrI is localized near the oldpole, where an asymmetric segregation process is initi-ated in which the duplicated origin segregates across thecell to the new pole. In contrast, the origin of chrII islocalized at the cell center, where after replication, andat a later point in the cell cycle, the duplicated originsmove symmetrically to the quarter positions of the cell,which become the new cell centers.
The distinct localization and segregation dynamics ofthe origin regions of the two chromosomes suggests theexistence of mechanisms capable of discriminating be-tween and interacting with the two origin regions to me-diate their cell cycle-dependent segregation dynamics.Both chromosomes contain unique origin-proximalparAB loci. Here we explored the role of the Par proteinsin the segregation of chrI. Our findings suggest that thechrI Par proteins participate in a mitotic-like pullingmechanism to mediate the asymmetric polar segregationof the origin region of chrI in V. cholerae.
Results
A YFP-ParBI fusion protein forms discrete polar focithat segregate asymmetrically
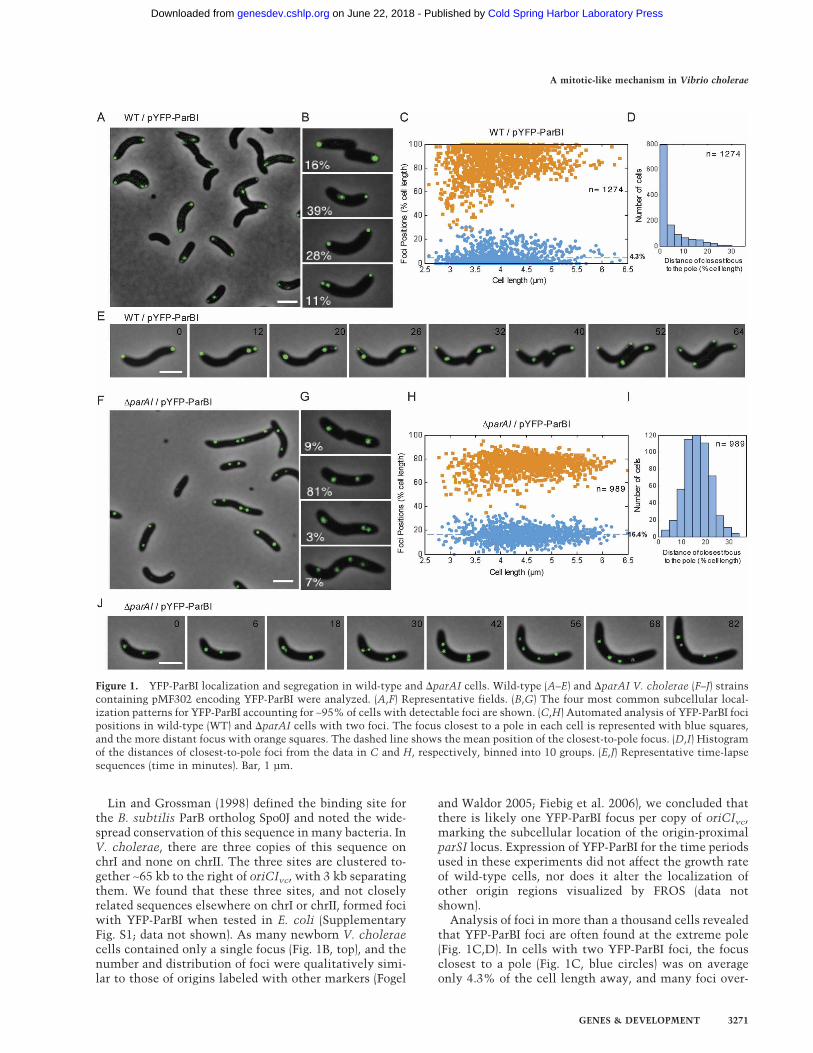
Fusions of GFP to ParB proteins in several plasmid andchromosomal systems form punctate fluorescent foci,marking the location of the ParB–parS nucleoproteincomplex in the cell (Glaser et al. 1997; Lin et al. 1997; Liand Austin 2002). We constructed a fusion of YFP to theN terminus of the ParB protein encoded on chrI (ParBI) inorder to investigate if its subcellular localization wassuggestive of a role in chromosome segregation. YFP-ParBI formed well-defined fluorescent foci in wild-typeV. cholerae cells (Fig. 1A). In contrast, expression of thisfusion protein in E. coli resulted in only diffuse fluores-cence (Supplementary Fig. S1), suggesting that there areV. cholerae specific binding sites for ParBI. Almost all V.cholerae cells with YFP-ParBI foci contained at least onefocus at the extreme pole of the cell (94%). The majorityof cells (67%) contained two foci, one at the extreme poleand the second either at the opposite pole or at an inter-mediate position between the poles (Fig. 1B). The num-ber of foci per cell and their localization was very similarto our observations of the origin of chrI (oriCIvc) detectedwith the lacO/tetO fluorescent repressor–operator sys-tems (FROS) (Fogel and Waldor 2005), suggesting thatParBI likely binds near the origin of chrI.
Fogel and Waldor
3270 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from
Lin and Grossman (1998) defined the binding site forthe B. subtilis ParB ortholog Spo0J and noted the wide-spread conservation of this sequence in many bacteria. InV. cholerae, there are three copies of this sequence onchrI and none on chrII. The three sites are clustered to-gether ∼65 kb to the right of oriCIvc, with 3 kb separatingthem. We found that these three sites, and not closelyrelated sequences elsewhere on chrI or chrII, formed fociwith YFP-ParBI when tested in E. coli (SupplementaryFig. S1; data not shown). As many newborn V. choleraecells contained only a single focus (Fig. 1B, top), and thenumber and distribution of foci were qualitatively simi-lar to those of origins labeled with other markers (Fogel
and Waldor 2005; Fiebig et al. 2006), we concluded thatthere is likely one YFP-ParBI focus per copy of oriCIvc,marking the subcellular location of the origin-proximalparSI locus. Expression of YFP-ParBI for the time periodsused in these experiments did not affect the growth rateof wild-type cells, nor does it alter the localization ofother origin regions visualized by FROS (data notshown).
Analysis of foci in more than a thousand cells revealedthat YFP-ParBI foci are often found at the extreme pole(Fig. 1C,D). In cells with two YFP-ParBI foci, the focusclosest to a pole (Fig. 1C, blue circles) was on averageonly 4.3% of the cell length away, and many foci over-
Figure 1. YFP-ParBI localization and segregation in wild-type and �parAI cells. Wild-type (A–E) and �parAI V. cholerae (F–J) strainscontaining pMF302 encoding YFP-ParBI were analyzed. (A,F) Representative fields. (B,G) The four most common subcellular local-ization patterns for YFP-ParBI accounting for ∼95% of cells with detectable foci are shown. (C,H) Automated analysis of YFP-ParBI focipositions in wild-type (WT) and �parAI cells with two foci. The focus closest to a pole in each cell is represented with blue squares,and the more distant focus with orange squares. The dashed line shows the mean position of the closest-to-pole focus. (D,I) Histogramof the distances of closest-to-pole foci from the data in C and H, respectively, binned into 10 groups. (E,J) Representative time-lapsesequences (time in minutes). Bar, 1 µm.
A mitotic-like mechanism in Vibrio cholerae
GENES & DEVELOPMENT 3271
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

lapped the edge of the cell. The other focus (Fig. 1C,orange squares) was found close to the opposite pole or atintermediate positions between the poles, presumablyrepresenting an actively segregating origin. As expected,time-lapse microscopy (Fig. 1E) revealed that the YFP-ParBI foci segregate with asymmetric dynamics that re-capitulate the segregation pattern of the FROS-labeledorigins (Fogel and Waldor 2005). Late predivisional cellscontain YFP-ParBI foci at both poles; these poles becomethe old poles of each daughter cell. Around the time ofdivision, the polar foci duplicate, and one segregatesacross the cell to the new pole; the other remains at theold pole. The most significant difference between YFP-ParBI foci and the origin-proximal loci studied previ-ously with FROS is the extreme polarity of YFP-ParBI;the FROS foci were rarely at the edge of the cell. Theextreme polarity of the YFP-ParBI foci suggested the pos-sibility that the ParBI–parSI nucleoprotein complex an-chors the chromosome to the pole, as has been proposedfor C. crescentus (Mohl and Gober 1997). The origin, 65kb away, would be expected to be near the pole, but notas close as the anchored site.
ParAI is required for polar positioning and asymmetricsegregation of ParBI
We constructed a V. cholerae strain with a deletion ofparAI. The �parAI strain had an increased frequency offilamentous cells, 1.8% versus 0.4% for wild type, sug-gesting a mild defect in cell division, but otherwise thestrain appeared to grow normally. We examined the lo-calization of YFP-ParBI in the �parAI mutant and foundthat the foci were dramatically mislocalized (Fig. 1F). In�parAI cells, the YFP-ParBI foci were dissociated fromthe cell poles and instead were generally found near thecell center in cells with a single focus or close to the quar-ter positions in cells with two foci (Fig. 1G). In �parAIcells with two foci, the mean position of the closest-to-pole ParBI focus (Fig. 1H, blue circles) was 16.4% of thecell length (vs. 4.3% for wild type), and >98.5% of thefoci were farther away from the pole than the mean valuefor wild type. Not only was the mean position of theclosest-to-pole focus changed, but the distribution of thedistances was significantly wider for the �parAI strain(Fig. 1, cf. D and I). This suggests that YFP-ParBI foci inthe �parAI strain have a greater degree of freedom intheir localization, a behavior that is consistent with lossof an attachment between the ParBI-bound chromosomeand the pole. Recently, mislocalization of an origin-proximal locus in a V. cholerae parAI deletion mutantwas shown by FISH (Saint-Dic et al. 2006). Togetherthese observations suggest that ParAI affects the local-ization of a relatively large origin-proximal domain ofchromosome I.
Remarkably, the asymmetric pattern that character-izes the segregation of oriCIvc was absent in the �parAImutant. Time-lapse analysis of �parAI cells showed gen-erally bidirectional movement of YFP-ParBI foci (Fig. 1J).The presence of separated and symmetrically distributed
YFP-ParBI foci in the �parAI strain (Fig. 1G,J) suggeststhat ParAI-independent mechanisms can still segregateduplicated origins, even though wild-type polar localiza-tion and asymmetric segregation are lost.
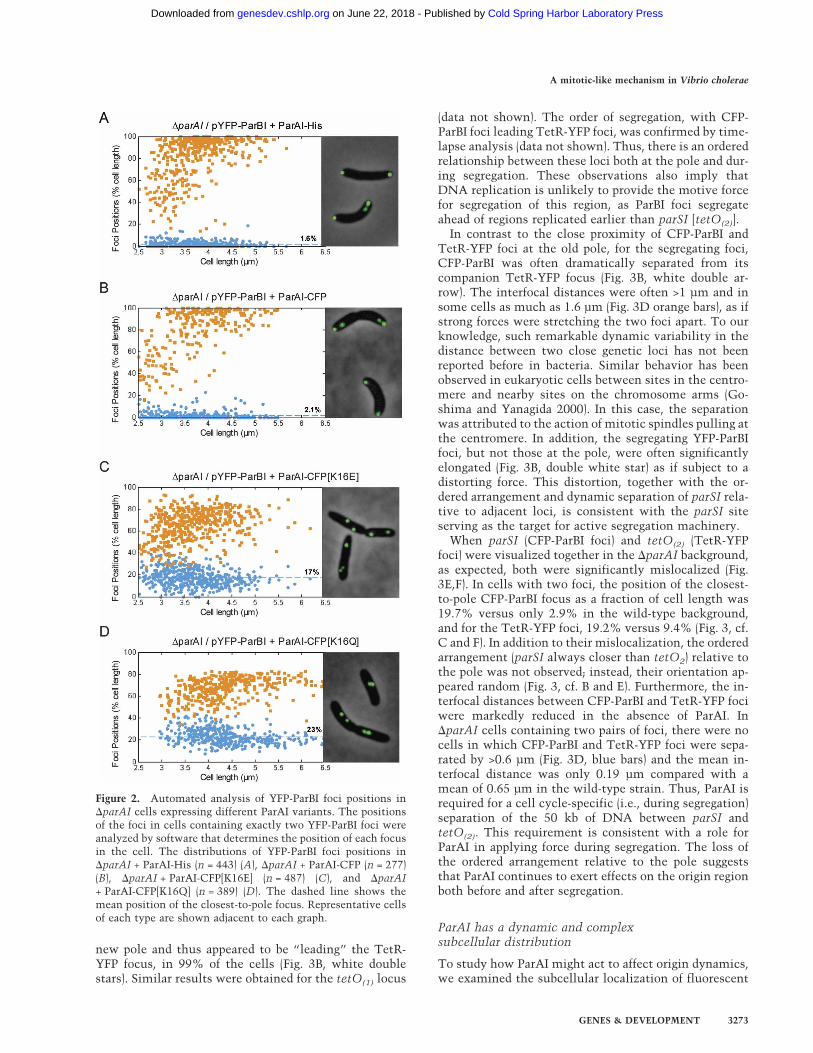
The defects in the �parAI cells are attributable to thedeletion of parAI, as expression of either a His-taggedParAI (ParAI-His) or a fluorescent fusion to ParAI (ParAI-CFP) restored the polar localization of YFP-ParBI foci(Fig. 2A,B, respectively). Moreover, the ability of ParAI tolocalize ParBI to the pole likely requires ATP, as substi-tutions in the conserved ATP-binding motif of ParAIabolished the ability of ParAI-CFP to complement theparAI deletion (Fig. 2C,D). In addition to the parSI site,other origin-proximal loci on both sides of oriCIvc werealso mislocalized in the �parAI mutant (see below).These observations suggest that ParAI is required to po-sition the ParBI–parSI complex at the extreme pole,thereby affecting the localization of a relatively large ori-gin-proximal region.
ParAI promotes separation of the parSI regionfrom neighboring loci during segregation
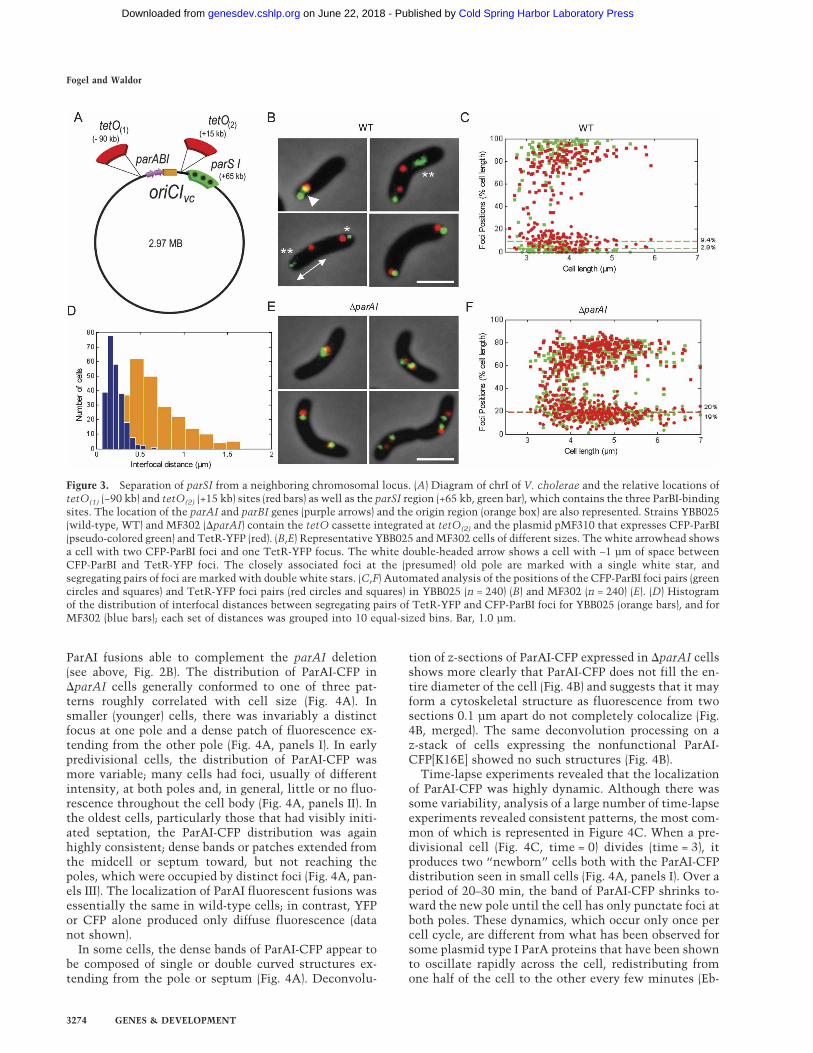
In order to study the spatial relationship between parSIand another origin-proximal locus, we expressed CFP-ParBI and TetR-YFP in a strain containing a tetO arrayintegrated 15 kb to the right of oriCIvc [Fig. 3A, tetO(2)].Under the growth conditions used, the majority (∼85%)of these dual-labeled cells had replicated the origin re-gion of chrI and therefore had four foci, one CFP-ParBIand one TetR-YFP for each copy of the origin region (Fig.3B). As expected from their localization in single fluo-rescence experiments (Fig. 1; Fogel and Waldor 2005),most cells contained one CFP-ParBI and one TetR-YFPfocus at the extrema of one pole, and a second pair eitherat the opposite pole or at intermediate positions (Fig. 3B).In most cells, for the pair of foci closest to a pole (the oldpole), the CFP-ParBI and TetR-YFP foci were adjacent oroverlapping, as might be expected for two loci separatedby only 50 kb (Fig. 3B, single white star). Despite thisclose association, their orientation with respect to thepole was not random. In fact, for 95.2% of cells (n = 849),the CFP-ParBI foci were found to be closer to the polethan the TetR-YFP foci (Fig. 3B, single white star; Fig.3C). Analysis of parSI and a tetO-tagged locus on theother side of the origin [Fig. 3A, tetO(1)] similarly showedthat parSI was consistently closest to the pole (data notshown). The ordered arrangement of these foci both rela-tive to each other and to a cellular structure (the pole)reveals an extremely fine level of spatial organization forthis region of the chromosome.
From the time-lapse experiments with YFP-ParBIalone (Fig. 1D), we know that for cells containing both apolar and an intermediate ParBI focus, the intermediatefocus is in the process of segregating across the cell to theopposite pole. In cells with a segregating pair of CFP-ParBI and TetR-YFP foci, we analyzed the spatial rela-tionship of the two loci during segregation. For thesesegregating foci, the CFP-ParBI focus was closer to the
Fogel and Waldor
3272 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from
new pole and thus appeared to be “leading” the TetR-YFP focus, in 99% of the cells (Fig. 3B, white doublestars). Similar results were obtained for the tetO(1) locus
(data not shown). The order of segregation, with CFP-ParBI foci leading TetR-YFP foci, was confirmed by time-lapse analysis (data not shown). Thus, there is an orderedrelationship between these loci both at the pole and dur-ing segregation. These observations also imply thatDNA replication is unlikely to provide the motive forcefor segregation of this region, as ParBI foci segregateahead of regions replicated earlier than parSI [tetO(2)].
In contrast to the close proximity of CFP-ParBI andTetR-YFP foci at the old pole, for the segregating foci,CFP-ParBI was often dramatically separated from itscompanion TetR-YFP focus (Fig. 3B, white double ar-row). The interfocal distances were often >1 µm and insome cells as much as 1.6 µm (Fig. 3D orange bars), as ifstrong forces were stretching the two foci apart. To ourknowledge, such remarkable dynamic variability in thedistance between two close genetic loci has not beenreported before in bacteria. Similar behavior has beenobserved in eukaryotic cells between sites in the centro-mere and nearby sites on the chromosome arms (Go-shima and Yanagida 2000). In this case, the separationwas attributed to the action of mitotic spindles pulling atthe centromere. In addition, the segregating YFP-ParBIfoci, but not those at the pole, were often significantlyelongated (Fig. 3B, double white star) as if subject to adistorting force. This distortion, together with the or-dered arrangement and dynamic separation of parSI rela-tive to adjacent loci, is consistent with the parSI siteserving as the target for active segregation machinery.
When parSI (CFP-ParBI foci) and tetO(2) (TetR-YFPfoci) were visualized together in the �parAI background,as expected, both were significantly mislocalized (Fig.3E,F). In cells with two foci, the position of the closest-to-pole CFP-ParBI focus as a fraction of cell length was19.7% versus only 2.9% in the wild-type background,and for the TetR-YFP foci, 19.2% versus 9.4% (Fig. 3, cf.C and F). In addition to their mislocalization, the orderedarrangement (parSI always closer than tetO2) relative tothe pole was not observed; instead, their orientation ap-peared random (Fig. 3, cf. B and E). Furthermore, the in-terfocal distances between CFP-ParBI and TetR-YFP fociwere markedly reduced in the absence of ParAI. In�parAI cells containing two pairs of foci, there were nocells in which CFP-ParBI and TetR-YFP foci were sepa-rated by >0.6 µm (Fig. 3D, blue bars) and the mean in-terfocal distance was only 0.19 µm compared with amean of 0.65 µm in the wild-type strain. Thus, ParAI isrequired for a cell cycle-specific (i.e., during segregation)separation of the 50 kb of DNA between parSI andtetO(2). This requirement is consistent with a role forParAI in applying force during segregation. The loss ofthe ordered arrangement relative to the pole suggeststhat ParAI continues to exert effects on the origin regionboth before and after segregation.
ParAI has a dynamic and complexsubcellular distribution
To study how ParAI might act to affect origin dynamics,we examined the subcellular localization of fluorescent
Figure 2. Automated analysis of YFP-ParBI foci positions in�parAI cells expressing different ParAI variants. The positionsof the foci in cells containing exactly two YFP-ParBI foci wereanalyzed by software that determines the position of each focusin the cell. The distributions of YFP-ParBI foci positions in�parAI + ParAI-His (n = 443) (A), �parAI + ParAI-CFP (n = 277)(B), �parAI + ParAI-CFP[K16E] (n = 487) (C), and �parAI+ ParAI-CFP[K16Q] (n = 389) (D). The dashed line shows themean position of the closest-to-pole focus. Representative cellsof each type are shown adjacent to each graph.
A mitotic-like mechanism in Vibrio cholerae
GENES & DEVELOPMENT 3273
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from
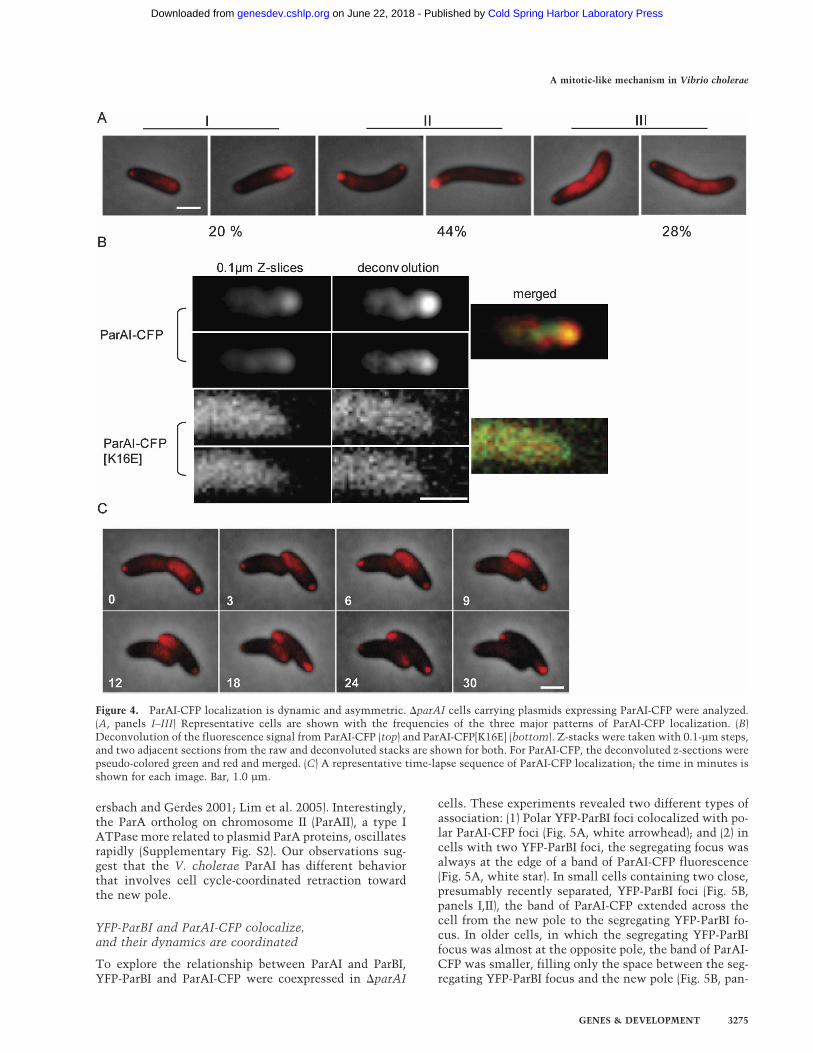
ParAI fusions able to complement the parAI deletion(see above, Fig. 2B). The distribution of ParAI-CFP in�parAI cells generally conformed to one of three pat-terns roughly correlated with cell size (Fig. 4A). Insmaller (younger) cells, there was invariably a distinctfocus at one pole and a dense patch of fluorescence ex-tending from the other pole (Fig. 4A, panels I). In earlypredivisional cells, the distribution of ParAI-CFP wasmore variable; many cells had foci, usually of differentintensity, at both poles and, in general, little or no fluo-rescence throughout the cell body (Fig. 4A, panels II). Inthe oldest cells, particularly those that had visibly initi-ated septation, the ParAI-CFP distribution was againhighly consistent; dense bands or patches extended fromthe midcell or septum toward, but not reaching thepoles, which were occupied by distinct foci (Fig. 4A, pan-els III). The localization of ParAI fluorescent fusions wasessentially the same in wild-type cells; in contrast, YFPor CFP alone produced only diffuse fluorescence (datanot shown).
In some cells, the dense bands of ParAI-CFP appear tobe composed of single or double curved structures ex-tending from the pole or septum (Fig. 4A). Deconvolu-
tion of z-sections of ParAI-CFP expressed in �parAI cellsshows more clearly that ParAI-CFP does not fill the en-tire diameter of the cell (Fig. 4B) and suggests that it mayform a cytoskeletal structure as fluorescence from twosections 0.1 µm apart do not completely colocalize (Fig.4B, merged). The same deconvolution processing on az-stack of cells expressing the nonfunctional ParAI-CFP[K16E] showed no such structures (Fig. 4B).
Time-lapse experiments revealed that the localizationof ParAI-CFP was highly dynamic. Although there wassome variability, analysis of a large number of time-lapseexperiments revealed consistent patterns, the most com-mon of which is represented in Figure 4C. When a pre-divisional cell (Fig. 4C, time = 0) divides (time = 3), itproduces two “newborn” cells both with the ParAI-CFPdistribution seen in small cells (Fig. 4A, panels I). Over aperiod of 20–30 min, the band of ParAI-CFP shrinks to-ward the new pole until the cell has only punctate foci atboth poles. These dynamics, which occur only once percell cycle, are different from what has been observed forsome plasmid type I ParA proteins that have been shownto oscillate rapidly across the cell, redistributing fromone half of the cell to the other every few minutes (Eb-
Figure 3. Separation of parSI from a neighboring chromosomal locus. (A) Diagram of chrI of V. cholerae and the relative locations oftetO(1) (−90 kb) and tetO(2) (+15 kb) sites (red bars) as well as the parSI region (+65 kb, green bar), which contains the three ParBI-bindingsites. The location of the parAI and parBI genes (purple arrows) and the origin region (orange box) are also represented. Strains YBB025(wild-type, WT) and MF302 (�parAI) contain the tetO cassette integrated at tetO(2) and the plasmid pMF310 that expresses CFP-ParBI(pseudo-colored green) and TetR-YFP (red). (B,E) Representative YBB025 and MF302 cells of different sizes. The white arrowhead showsa cell with two CFP-ParBI foci and one TetR-YFP focus. The white double-headed arrow shows a cell with ∼1 µm of space betweenCFP-ParBI and TetR-YFP foci. The closely associated foci at the (presumed) old pole are marked with a single white star, andsegregating pairs of foci are marked with double white stars. (C,F) Automated analysis of the positions of the CFP-ParBI foci pairs (greencircles and squares) and TetR-YFP foci pairs (red circles and squares) in YBB025 (n = 240) (B) and MF302 (n = 240) (E). (D) Histogramof the distribution of interfocal distances between segregating pairs of TetR-YFP and CFP-ParBI foci for YBB025 (orange bars), and forMF302 (blue bars); each set of distances was grouped into 10 equal-sized bins. Bar, 1.0 µm.
Fogel and Waldor
3274 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from
ersbach and Gerdes 2001; Lim et al. 2005). Interestingly,the ParA ortholog on chromosome II (ParAII), a type IATPase more related to plasmid ParA proteins, oscillatesrapidly (Supplementary Fig. S2). Our observations sug-gest that the V. cholerae ParAI has different behaviorthat involves cell cycle-coordinated retraction towardthe new pole.
YFP-ParBI and ParAI-CFP colocalize,and their dynamics are coordinated
To explore the relationship between ParAI and ParBI,YFP-ParBI and ParAI-CFP were coexpressed in �parAI
cells. These experiments revealed two different types ofassociation: (1) Polar YFP-ParBI foci colocalized with po-lar ParAI-CFP foci (Fig. 5A, white arrowhead); and (2) incells with two YFP-ParBI foci, the segregating focus wasalways at the edge of a band of ParAI-CFP fluorescence(Fig. 5A, white star). In small cells containing two close,presumably recently separated, YFP-ParBI foci (Fig. 5B,panels I,II), the band of ParAI-CFP extended across thecell from the new pole to the segregating YFP-ParBI fo-cus. In older cells, in which the segregating YFP-ParBIfocus was almost at the opposite pole, the band of ParAI-CFP was smaller, filling only the space between the seg-regating YFP-ParBI focus and the new pole (Fig. 5B, pan-
Figure 4. ParAI-CFP localization is dynamic and asymmetric. �parAI cells carrying plasmids expressing ParAI-CFP were analyzed.(A, panels I–III) Representative cells are shown with the frequencies of the three major patterns of ParAI-CFP localization. (B)Deconvolution of the fluorescence signal from ParAI-CFP (top) and ParAI-CFP[K16E] (bottom). Z-stacks were taken with 0.1-µm steps,and two adjacent sections from the raw and deconvoluted stacks are shown for both. For ParAI-CFP, the deconvoluted z-sections werepseudo-colored green and red and merged. (C) A representative time-lapse sequence of ParAI-CFP localization; the time in minutes isshown for each image. Bar, 1.0 µm.
A mitotic-like mechanism in Vibrio cholerae
GENES & DEVELOPMENT 3275
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

els III,IV). In late predivisional cells (Fig. 5B, panels V,VI),YFP-ParBI foci are at each pole, and ParAI-CFP was al-most completely located at the cell center or septumfrom which it extended outward toward the two old
poles. Thus, before division is complete, both ParAI andParBI are positioned to repeat the cycle.
In order to analyze the relationship between YFP-ParBIand ParAI-CFP fluorescence in a large number of cells,
Figure 5. YFP-ParBI colocalizes with ParAI-CFP at the pole and at the edge of ParAI-CFP bands. �parAI cells contained plasmidsexpressing YFP-ParBI and ParAI-CFP. (A) Phase, YFP, CFP, and merged images of a representative cell showing the two distinct typesof colocalization: foci at the pole (white arrowhead) and YFP-ParBI-focus at the edge of ParAI-CFP bands (white star). (B) Representativecells of different lengths and at different points during chromosome segregation showing the relationship of ParAI-CFP localizationwith YFP-ParBI. (C) Graphical representation of the relationship between YFP-ParBI separation (green circles) and the localization ofParAI-CFP fluorescence along the cell’s length (red surface) in 314 cells. For each cell, width-averaged ParAI-CFP fluorescence (Z-axis)at each point along the midline (X-axis) is graphed versus the distance between the YFP-ParBI foci of that cell. The positions of theYFP-ParBI foci (green) are overlaid on the graph (at an arbitrary Z-axis height of 0.3 for visibility). Dashed arrows point to images ofcells (panels a–d) representative of data at the indicated position of the graph. The width-averaged ParAI-CFP fluorescence values werebackground-subtracted, then normalized as a fraction of total cellular fluorescence. The positions along the midline (X-axis) and thedistance between YFP-ParBI foci (Y-axis) were normalized as a fraction of total cell length. Only cells with above-background signalfor both fusions and wild-type localization of the YFP-ParBI foci were analyzed. (D) Time-lapse analysis of �parAI cells expressingParAI-CFP and YFP-ParBI showing that ParAI-CFP retracts toward the new pole ahead of segregating YFP-ParBI foci. Bar, 1.0 µm.
Fogel and Waldor
3276 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

we designed software to quantify the distribution of totalcellular ParAI-CFP fluorescence along the length of thecell (see Materials and Methods). Figure 5C shows thisanalysis for 314 cells that contained two YFP-ParBI foci.Cells with short distances between YFP-ParBI foci haveParAI-CFP spread over a large fraction of the cell (Fig. 5C,panel a). In cells with greater separation between theirYFP-ParBI foci, the highest intensities of ParAI-CFP fluo-rescence are found closer to the distant pole (Fig. 5C,panels b,c). When the YFP-ParBI foci are completely seg-regated to opposite poles, the dramatic redistribution ofParAI-CFP fluorescence from the poles to a large regioncentered at mid-cell is readily apparent (Fig. 5C, panel d).At all cell sizes, ParAI-CFP was present at the old polewhere the nonsegregating YFP-ParBI focus resides, sug-gesting that ParAI interacts with ParBI at the old polethroughout the cell cycle. Time-lapse analysis of cellscoexpressing YFP-ParBI and ParAI-CFP confirm the rela-tionship between segregating YFP-ParBI foci and ParAI-CFP (Fig. 5D). Segregating YFP-ParBI foci (Fig. 5D, greenfoci) appear to be pulled by retracting ParAI-CFP (Fig. 5D,red fluorescence) to the new pole. At time 0, the cell inFigure 5D is about to divide. From time 0 to 12 min,ParAI-CFP relocalizes such that it extends from the newpole of each daughter cell to the old pole, where theYFP-ParBI foci have duplicated. Between 12 and 16 min,the YFP-ParBI foci segregate across the cell at the edge ofretracting ParAI-CFP. From 16 to 20 min, the YFP-ParBIfocus of the bottom cell continues to follow ParAI-CFPretraction, while the YFP-ParBI focus of the top cell hasalready reached the pole; in this cell, ParAI-CFP has re-distributed to the middle of the cell, where it will be
positioned to repeat the cycle. The in vivo dynamics ofthe ParBI and ParAI fluorescent fusions suggest thatParAI both pulls the ParBI-bound DNA across the cell,and anchors it at the pole.
Mutations in the ATP-binding motif of ParAI abolishits dynamics
As the activity of many Walker-type ParA proteins hasbeen shown to require ATP hydrolysis (Leonard et al.2005), we made two separate substitution mutations,K16E and K16Q, in the predicted ATP-binding pocket ofthe ParAI-CFP fusion protein. In a plasmid ortholog ofParAI, these substitutions have been reported to blockATP binding (K16E) and to allow binding but preventhydrolysis (K16Q), respectively (Fung et al. 2001). As de-scribed above, neither construct was capable of comple-menting the mislocalization of YFP-ParBI foci in the�parAI background (Fig. 2C,D). We examined the sub-cellular distribution of these constructs to investigatethe effects of these mutations on ParAI dynamics. Inboth wild-type and �parAI backgrounds, ParAI-CFP[K16E] showed only homogeneous diffuse fluores-cence (Fig. 6A) and had no detectable dynamics in time-lapse experiments (data not shown). The K16Q mutationsimilarly resulted in diffuse fluorescence and lack ofwild-type localization and dynamics; however, it alsoformed faint foci (Fig. 6A). When YFP-ParBI was coex-pressed with ParA-CFP[K16Q], their foci were always co-localized (Fig. 6B). The loss of ParAI dynamics with bothsubstitutions suggests that ATP binding and hydrolysisare required for the dynamic localization of ParAI. The
Figure 6. Mutations in the conservedATPase domain of ParAI abolish ParAI-CFP localization. (A) Representative im-ages of �parAI cells expressing wild-typeParAI-CFP (top), ParAI-CFP [K16E](middle), and ParAI-CFP [K16Q] (bottom).(B) Phase, YFP, CFP, and merged images ofa representative cell expressing both YFP-ParBI and ParAI-CFP[K16Q] showing theircolocalization. Bar, 1.0 µm.
A mitotic-like mechanism in Vibrio cholerae
GENES & DEVELOPMENT 3277
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

colocalization of the K16Q mutant, but not the K16Emutant, with YFP-ParBI foci suggests that the ATP-bound state, but not the unbound state (presumably thecase for the K16E substitution), may promote the inter-action of ParAI with ParBI.
Discussion
Our findings show that ParAI, ParBI, and parSI (ParABSI)are components of a mechanism that mediates the polarlocalization and asymmetric segregation of the origin re-gion of chrI. ParBI interacts with parSI sites near oriCIvc,apparently forming a complex that localizes and asym-metrically segregates with the origin (Figs. 1B,D,F, 2A).ParAI is required for the extreme polar localization of theParBI–parSI complex and its pattern of segregation (Fig.1C,E,G). This type I ATPase is also required for dramaticcell cycle-dependent separation of parSI from neighbor-ing loci, a phenomenon consistent with active DNAmovement (Fig. 3). Moreover, our observations suggest afunctional relationship between segregating ParBI fociand ParAI dynamics, as ParAI-CFP retraction toward thenew pole coincided with YFP-ParBI movement. Takingour data together, we propose the following model forParABI-mediated segregation of the origin region of chrIvia a pulling mechanism (Fig. 7): In late predivisionalcells containing two fully replicated and segregated chro-
mosomes, the origin of each chromosome is attached tothe pole by an interaction between ParAI, anchored to anas-of-yet unknown polar protein/structure, and ParBI,bound to the chromosome at the origin-proximal parSIsite (Fig. 7A). At some point, perhaps related to the as-sembly of the cell division machinery, ParAI nucleates atthe forming septum and polymerizes outward as bandsor networks of polymers toward both poles. The nextround of DNA replication yields sister copies of the ori-gin region, each containing a ParBI–parSI complex. Onecomplex is captured by the ParAI already present at theold pole; the other ParBI–parSI complex is captured bythe ParAI extending from the closing septum that willbecome the new pole (Fig. 7B). The completion of cyto-kinesis produces two daughter cells in which ParBI-bound DNA is pulled across the cell by the retractingParAI polymers (Fig. 7C,D).
Our findings provide the first in vivo evidence for mi-totic-like pulling forces mediating DNA movement inbacteria. Given the localization of ParAI ahead of segre-gating YFP-ParBI foci, ParAI pulling is strongly suggestedby its requirement for both the distortion of YFP-ParBIfoci (Fig. 3B) and their separation from nearby regionsduring segregation (Fig. 3B,D,E). A pulling mechanismsuch as this provides an uncomplicated explanation forthe asymmetric segregation pattern of the origin regionof chrI in V. cholerae. While the dynamics of chromo-some segregation have been described in only a few bac-teria, given the widespread presence of chromosome-en-coded Par proteins related to V. cholerae ParAI and ParBIin diverse bacteria (Gerdes et al. 2000), it is likely thatthe Par-directed pulling segregation mechanism de-scribed here is conserved in many other systems.
ParBI localizes with parSI sites near the origin
The formation of visible foci by fluorescent ParBI fusionssuggests that, like other ParB proteins, ParBI forms alarge nucleoprotein complex on chrI DNA. In compari-son to the distribution of the eight or more Spo0J-bindingsites over a 800-kb region that spans the origin of the B.subtilis chromosome (Lin and Grossman 1998), the threeparSI sites on V. cholerae chrI are much closer together.These conserved ParB-binding sites, originally identifiedin the V. cholerae genome by Lin and Grossman (1998),were recently shown to be functional for ParABI-medi-ated plasmid stabilization (Saint-Dic et al. 2006). Thegreater number and distribution of parS sites in B. sub-tilis may reflect an adaptation of the Par origin-localizingsystem to facilitate large-scale compaction and localiza-tion of the chromosome during sporulation. As in V.cholerae, the four to five parS sites in C. crescentus areclustered together within an ∼10-kb region, close to theorigin (Mohl et al. 2001). The similarity of the distribu-tions of the V. cholerae and C. crescentus parS sitesalong with the polar localization of their respective ori-gins suggest that the C. crescentus and V. cholerae chrIpar systems likely carry out the same function for originlocalization, although they may have other roles thatdiffer.
Figure 7. Model of ParAI-mediated segregation of the ParBI-bound origin of chrI. (A) Older, predivisional, cells contain twofully replicated and segregated chromosomes. The origin of eachchromosome is attached to the pole by an interaction betweenParAI, anchored to an as-of-yet unknown polar protein/struc-ture, and ParBI, bound to the chromosome at the origin-proxi-mal parSI site. (B) At some point, perhaps related to the assem-bly of the cell division machinery, ParAI nucleates at the form-ing septum and polymerizes outward as bands or networks ofpolymers toward both poles. The next round of DNA replicationyields sister copies of the origin region, each containing a ParBI–parSI complex. One complex is captured by the ParAI alreadypresent at the old pole; the other ParB–parSI complex is cap-tured by the ParAI extending from the closing septum that willbecome the new pole. (C,D) The completion of cytokinesis pro-duces two daughter cells in which ParBI-bound DNA is beingpulled across the cell by the retracting ParAI polymers.
Fogel and Waldor
3278 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

ParAI does not influence bulk nucleoid segregation
The ParABSI system primarily functions to position andsegregate the origin region of chrI. Deletion of parAI didnot dramatically disrupt partitioning of chrI to daughtercells, supporting the emerging view that bacterial chro-mosome segregation is mediated by multiple, likelyoverlapping or redundant mechanisms (Errington et al.2005). Similarly, in C. crescentus, it was shown thatMreB is required for proper positioning of the origin butnot for other regions of the chromosome (Gitai et al.2005). As C. crescentus origin localization is so similarto that of V. cholerae, it is possible that MreB plays simi-lar roles in both bacteria, either as a separate mechanismor perhaps as a scaffolding for Par-mediated segregation,as proposed by Gitai et al. (2005). It seems likely thatbulk chromosome segregation may be directed by con-served processes such as replication, transcription, andcondensation; perhaps additional, species-specific,mechanisms might mediate the localization of specificchromosome regions such as the origin and terminus.
The parAI mutant has only a subtle growth defect de-spite the dramatic mislocalization of origin proximal re-gions in this background. The function of the extremepolar localization of chromosomal origins in bacteriasuch as V. cholerae and C. crescentus is unclear. Perhapsthere are particular environmental conditions in whichthere is a strong advantage to having a more elongatedand/or organized genome. If the chrI par genes do influ-ence the segregation of origin-distal regions of the chro-mosome, then redundant mechanisms must be able tocompensate in their absence. It will be interesting toexamine if mutations in other genes implicated in chro-mosome segregation, such as mreB and mukB, have syn-thetic effects with parAI, as has been recently reportedfor soj in B. subtilis (Lee and Grossman 2006).
It is possible that there is cross-talk between the chrIand chrII par systems; the absence of a significant growthdefect in the �parAI mutant may be attributable to theactivity of ParAII, encoded in the par locus on chrII.While ParAII is phylogenetically grouped with plasmidParA proteins, both are type I ATPases, and they are 45%similar. Consistent with this idea, the localization andsegregation of the ParB–parSI complex in the �parAIbackground resembles the dynamics of oriCIIvc. Whilethe mechanism to properly localize the parSI site at thepole is clearly dependent on ParAI, it is possible thatParAII might substitute for ParAI in other importantsteps during segregation. One interesting possibility isthat the ParABI–parSI system may have a function spe-cifically in origin separation, as is the case for Soj in B.subtilis (Lee and Grossman 2006). If so, ParAII mightpromote the separation of duplicated ParBI–parSI com-plexes, thus partially suppressing the parAI deletion.
Pulling or pushing DNA
Polymerizing ParA proteins could localize DNA by pro-viding either “pushing” or “pulling” motive force(Møller-Jensen et al. 2002; Barillà et al. 2005). To date,
the only well-established mechanism for Par-mediatedDNA segregation is the ParM–ParR system of plasmidR1. ParM (a type II ParA ATPase) polymerizes bidirec-tionally between ParR-bound (ParB-like) plasmids. Inthis system, the ParM–ParR interaction stabilizes theParM filament, facilitating its continued polymeriza-tion, thereby pushing the plasmid clusters in oppositedirections in the cell (Møller-Jensen et al. 2002, 2003;Garner et al. 2004). Recently, another elegant mecha-nism was proposed for SopA/SopB-mediated segregationof the F plasmid that also invokes a pushing force. In thiscase, radial asters of SopA filaments pushing in all direc-tions were hypothesized to mediate F segregation (Lim etal. 2005). In both models, the polymerizing ParA-likeprotein is found between segregating ParB–DNA com-plexes. Therefore, the region of ParA fluorescence in-creases with the distance between ParB complexes. Thisis not the case with the V. cholerae ParAI and ParBIproteins; rather, ParAI is found between the segregatingParBI focus and the new pole; fluorescent ParAI“shrinks” toward the new pole with increased ParBI seg-regation, consistent with a pulling mechanism.
Several recent observations regarding the biochemis-try of ParA proteins and of ParA–ParB interactions sug-gest how ParAI may pull the ParBI–parSI complex to-ward the new pole. In vitro, in the presence of ATP, ParAmonomers can polymerize and form filamentous struc-tures (Møller-Jensen et al. 2002, 2003; Barillà et al. 2005).The nature of the structure of ParAI in V. cholerae re-quires future investigation; many of our images raise thepossibility that this structure is composed of many ParAIpolymers, possibly even a network or lattice of smallParAI polymers. In some plasmid systems, ParA bindingto ParB stimulates ParA hydrolysis of ATP, promotingits disassociation from ParB and depolymerization. As-suming that similar processes occur in vivo with the V.cholerae ParAI and ParBI proteins, then the interactionof ParAI filaments with the ParBI–parSI complex wouldresult in their depolymerization, thereby pulling the ori-gin region of chrI toward the new pole via a “Brownianratchet” mechanism (Raj and Peskin 2006). In this typeof mechanism, ParBI, present as part of a large ParBI/parSI complex, would interact with ATP-bound ParAImolecules at the ends of ParAI polymers (within a largerfilamentous structure) and in so doing, stimulate theirATPase activity. ATP hydrolysis by the terminal subunitwould result in its dissociation as a monomer ofParAI • ADP (i.e., depolymerization). If the ParBI–parSIcomplex is only free to diffuse toward the polymer (itwould be prevented from diffusing away by simulta-neous contact with multiple ParAI polymers), it willcome into contact with the new leading edge of theParAI polymer, and again experience a positive interac-tion that can be propagated along the DNA, shifting theaverage position of nearby regions closer to the polymeredge. The observed retraction of ParAI-CFP as YFP-ParBIsegregates (Fig. 6) is consistent with the idea that ParBIstimulates depolymerization of ParAI polymers. AsParAI-CFP fluorescence is consistently associated withthe nonsegregating YFP-ParBI foci at the old pole, it is
A mitotic-like mechanism in Vibrio cholerae
GENES & DEVELOPMENT 3279
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

possible that ParAI is undergoing a constant cycle of po-lymerization and depolymerization to anchor ParBI–parSI at the pole. This type of equilibrium might involvean unknown polar protein that stimulates ParAI poly-merization, whereas ParBI–parSI antagonizes it. Whilethe biochemistry of this interaction is still highly specu-lative, it is consistent with our observations regardingsubstitutions in ParAI, as a mutation likely to preventParAI ATP binding abolished ParAI–ParBI colocaliza-tion, whereas a different substitution predicted to allowATP binding, but not ATP hydrolysis, colocalized withParBI. Further studies of the biochemistry of these pro-teins will be important to understand the details of ParAIdynamics and how ParAI–ParBI interactions could gen-erate force for chromosome movement.
Materials and methods
Strains and plasmids
A list of the strains and plasmids used in this workis availablein Supplementary Table S1. All V. cholerae strains used in thisstudy were derived from the sequenced clinical isolate N16961(Heidelberg et al. 2000). The �parAI mutation was made byallele exchange, as described (Fogel and Waldor 2005), usingpMF158, a derivative of pCVD442 (Donnenberg and Kaper 1991)containing homology with regions flanking parAI. The deletionwas confirmed by PCR. yfp-parBI was constructed by “splicingby overlap extension” PCR (Horton et al. 1989) and then in-serted into pBAD33 and pBAD18 (Guzman et al. 1995) to makeplasmids pMF302 and pMF341, respectively. parAI-yfp andparAI-cfp fusions were constructed by inserting parAI upstreamof the yfp or cfp gene of plasmids p4414 and p4416, containingmonomeric versions of yfp and cfp (generously provided by S.Bunnell, Tufts Medical School, Boston, MA). After determiningthat the sequences of the resulting fusions were correct, theywere subcloned into pBAD33, yielding pMF320 and pMF321.ParAI K16E (pMF322) and K16Q (pMF323) substitutions wereintroduced into these plasmids by PCR mutagenesis using theQuickChange kit protocol (Stratagene) and verified by sequenc-ing. pMF310 was constructed by inserting tetR-yfp, derivedfrom pLAU53 (Lau et al. 2003) downstream from cfp-parBI inplasmid pMF303 (pBAD33-cfp-parBI). Strains YBB025 andMF302, which harbor tet operators inserted in vc0018, wereconstructed using allele exchange vector pYBA016, a pCVD442derivative with the tetO cassette from pLAU44 (Lau et al. 2003)ligated into the XbaI site of a fragment of vc0018.
Microscopy
Cells were routinely prepared for microscopy by inoculatingfresh single colonies taken from LB plates into M63 minimalmedia containing 0.1% casamino acids and 0.2% glucose andgrown at 37°C to a density of 0.3–0.5 OD600 units. Expression ofthe fluorescent fusions from pBAD plasmids was induced byaddition of 0.08% arabinose for 20–30 min. For experimentswith strains containing the tetO cassette, 80 nM Anhydrotet-racycline was added to the culture media at the same time asarabinose, to reduce TetR-YFP binding as described previously(Lau et al. 2003). Ten microliters to 20 µL of the broth culturewas adsorbed onto thin agarose pads on microscope slides andallowed to settle for 3–4 min; then, the remaining media wasaspirated and a coverslip were placed on top. Slides were sealedwith nail polish. Images were acquired with a Zeiss Axioplane
2 microscope equipped with a 100× �-plan lens, filter sets forYFP and CFP fluorescence, and a cooled CCD Hamamatsu Orcacamera. Openlab 3.0 software was used for image acquisitionand processing.
Image analysis and measurements
Using MatLab software (MathWorks), we developed automatedimage analysis programs to facilitate localization of fluorescentfoci in large numbers of cells as well as for analysis of othertypes of fluorescence signals. Briefly, during data collection, setsof phase-contrast and fluorescence images were collected foreach field of cells. Segmentation of the phase-contrast imagesinto individual cells was done by binary thresholding followedby region detection and shape filters. For each cell body, mor-phological shrinking operations were performed on the binaryimage to create a central skeleton that perfectly represented theindividual curvature of the cell. The poles were calculated asthe two points on each side of the long axis of the cell that weremost distant from the midpoint of the skeleton (similar to anapproach for identifying poles in Viollier et al. 2004). The gapbetween the poles and the skeleton was filled in by linear in-terpolation to create a complete curved midline. For each cellbody, the corresponding region from the fluorescence image wasanalyzed and foci were detected by determining points of localmaxima in fluorescence intensity. The position of each focus inthe cell was determined by finding the closest point on thecurved midline and measuring along the curve to each pole. InFigure 5C, the average ParAI-CFP intensity along the curvedmidline was calculated for each cell. For each pixel along themidline axis, the fluorescence intensity of all of the pixels in thecell lying along the perpendicular line from that point weresummed and then averaged. This information is represented asan average intensity along the long axis of the cell and plottedagainst the position of YFP-ParBI foci in the same cell. Decon-volution of ParAI-CFP fluorescence was performed with Veloc-ity software (Improvision) using a calculated point-spread func-tion on an image-stack of 0.1-µm sections.
Acknowledgments
We thank members of the Waldor laboratory, M. Dorer, and M.Berkmen for helpful discussions and critical reading of themanuscript; S. Bunnell for reagents; P. McGrath for sharing soft-ware; and A. Vaughan for programming advice. We acknowl-edge the support of NIH and HHMI.
References
Adachi, S., Hori, K., and Hiraga, S. 2006. Subcellular positioningof F plasmid mediated by dynamic localization of SopA andSopB. J. Mol. Biol. 356: 850–863.
Barillà, D., Rosenberg, M.F., Nobbmann, U., and Hayes, F. 2005.Bacterial DNA segregation dynamics mediated by the poly-merizing protein ParF. EMBO J. 24: 1453–1464.
Donnenberg, M.S. and Kaper, J.B. 1991. Construction of an eaedeletion mutant of enteropathogenic Escherichia coli by us-ing a positive-selection suicide vector. Infect. Immun.59: 4310–4317.
Dworkin, J. and Losick, R. 2002. Does RNA polymerase helpdrive chromosome segregation in bacteria? Proc. Natl. Acad.Sci. 99: 14089–14094.
Ebersbach, G. and Gerdes, K. 2001. The double par locus ofvirulence factor pB171: DNA segregation is correlated withoscillation of ParA. Proc. Natl. Acad. Sci. 98: 15078–15083.
Ebersbach, G. and Gerdes, K. 2005. Plasmid segregation mecha-nisms. Annu. Rev. Genet. 39: 453–479.
Fogel and Waldor
3280 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

Egan, E.S., Lobner-Olesen, A., and Waldor, M.K. 2004. Synchro-nous replication initiation of the two Vibrio cholerae chro-mosomes. Curr. Biol. 14: R501–R502.
Errington, J., Murray, H., and Wu, L.J. 2005. Diversity and re-dundancy in bacterial chromosome segregation mecha-nisms. Philos. Trans. R. Soc. Lond. B Biol. Sci. 360: 497–505.
Fiebig, A., Keren, K., and Theriot, J.A. 2006. Fine-scale time-lapse analysis of the biphasic, dynamic behaviour of the twoVibrio cholerae chromosomes. Mol. Microbiol. 60: 1164–1178.
Fogel, M.A. and Waldor, M.K. 2005. Distinct segregation dy-namics of the two Vibrio cholerae chromosomes. Mol. Mi-crobiol. 55: 125–136.
Fung, E., Bouet, J.Y., and Funnell, B.E. 2001. Probing the ATP-binding site of P1 ParA: Partition and repression have differ-ent requirements for ATP binding and hydrolysis. EMBO J.20: 4901–4911.
Garner, E.C., Campbell, C.S., and Mullins, R.D. 2004. Dynamicinstability in a DNA-segregating prokaryotic actin homolog.Science 306: 1021–1025.
Gerdes, K., Møller-Jensen, J., and Bugge Jensen, R. 2000. Plasmidand chromosome partitioning: Surprises from phylogeny.Mol. Microbiol. 37: 455–466.
Gitai, Z., Dye, N.A., Reisenauer, A., Wachi, M., and Shapiro, L.2005. MreB actin-mediated segregation of a specific region ofa bacterial chromosome. Cell 120: 329–341.
Glaser, P., Sharpe, M.E., Raether, B., Perego, M., Ohlsen, K., andErrington, J. 1997. Dynamic, mitotic-like behavior of a bac-terial protein required for accurate chromosome partition-ing. Genes & Dev. 11: 1160–1168.
Gordon, G.S., Sitnikov, D., Webb, C.D., Teleman, A., Straight,A., Losick, R., Murray, A.W., and Wright, A. 1997. Chromo-some and low copy plasmid segregation in E. coli: Visualevidence for distinct mechanisms. Cell 90: 1113–1121.
Goshima, G. and Yanagida, M. 2000. Establishing biorientationoccurs with precocious separation of the sister kinetochores,but not the arms, in the early spindle of budding yeast. Cell100: 619–633.
Guzman, L.M., Belin, D., Carson, M.J., and Beckwith, J. 1995.Tight regulation, modulation, and high-level expression byvectors containing the arabinose PBAD promoter. J. Bacte-riol. 177: 4121–4130.
Heidelberg, J.F., Eisen, J.A., Nelson, W.C., Clayton, R.A.,Gwinn, M.L., Dodson, R.J., Haft, D.H., Hickey, E.K., Peter-son, J.D., Umayam, L., et al. 2000. DNA sequence of bothchromosomes of the cholera pathogen Vibrio cholerae. Na-ture 406: 477–483.
Horton, R.M., Hunt, H.D., Ho, S.N., Pullen, J.K., and Pease, L.R.1989. Engineering hybrid genes without the use of restric-tion enzymes: Gene splicing by overlap extension. Gene 77:61–68.
Ireton, K., Gunther, N.W., and Grossman, A.D. 1994. spo0J isrequired for normal chromosome segregation as well as theinitiation of sporulation in Bacillus subtilis. J. Bacteriol.176: 5320–5329.
Jacob, F., Brenner, S., and Cuzin, F. 1963. On the regulation ofDNA replication in bacteria. Cold Spring Harbor Symp.Quant. Biol. 23: 329–348.
Jensen, R.B. and Shapiro, L. 1999. The Caulobacter crescentussmc gene is required for cell cycle progression and chromo-some segregation. Proc. Natl. Acad. Sci. 96: 10661–10666.
Jensen, R.B., Wang, S.C., and Shapiro, L. 2001. A moving DNAreplication factory in Caulobacter crescentus. EMBO J. 20:4952–4963.
Kruse, T., Blagoev, B., Løbner-Olesen, A., Wachi, M., Sasaki, K.,Iwai, N., Mann, M., and Gerdes, K. 2006. Actin homolog
MreB and RNA polymerase interact and are both required forchromosome segregation in Escherichia coli. Genes & Dev.20: 113–124.
Lau, I.F., Filipe, S.R., Søballe, B., Økstad, O.A., Barre, F.X., andSherratt, D.J. 2003. Spatial and temporal organization of rep-licating Escherichia coli chromosomes. Mol. Microbiol. 49:731–743.
Lee, P.S. and Grossman, A.D. 2006. The chromosome partition-ing proteins Soj (ParA) and Spo0J (ParB) contribute to accu-rate chromosome partitioning, separation of replicated sisterorigins, and regulation of replication initiation in Bacillussubtilis. Mol. Microbiol. 60: 853–869.
Lee, P.S., Lin, D.C., Moriya, S., and Grossman, A.D. 2003. Ef-fects of the chromosome partitioning protein Spo0J (ParB) onoriC positioning and replication initiation in Bacillus subti-lis. J. Bacteriol. 185: 1326–1337.
Lemon, K.P. and Grossman, A.D. 1998. Localization of bacterialDNA polymerase: Evidence for a factory model of replica-tion. Science 282: 1516–1519.
Lemon, K.P. and Grossman, A.D. 2000. Movement of replicat-ing DNA through a stationary replisome. Mol. Cell 6: 1321–1330.
Leonard, T.A., Møller-Jensen, J., and Löwe, J. 2005. Towardsunderstanding the molecular basis of bacterial DNA segre-gation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 360: 523–535.
Li, Y. and Austin, S. 2002. The P1 plasmid in action: Time-lapsephotomicroscopy reveals some unexpected aspects of plas-mid partition. Plasmid 48: 174–178.
Lim, G.E., Derman, A.I., and Pogliano, J. 2005. Bacterial DNAsegregation by dynamic SopA polymers. Proc. Natl. Acad.Sci. 102: 17658–17663.
Lin, D.C. and Grossman, A.D. 1998. Identification and charac-terization of a bacterial chromosome partitioning site. Cell92: 675–685.
Lin, D.C., Levin, P.A., and Grossman, A.D. 1997. Bipolar local-ization of a chromosome partition protein in Bacillus subti-lis. Proc. Natl. Acad. Sci. 94: 4721–4726.
Mohl, D.A. and Gober, J.W. 1997. Cell cycle-dependent polarlocalization of chromosome partitioning proteins in Caulo-bacter crescentus. Cell 88: 675–684.
Mohl, D.A., Easter, J., and Gober, J.W. 2001. The chromosomepartitioning protein, ParB, is required for cytokinesis in Cau-lobacter crescentus. Mol. Microbiol. 42: 741–755.
Møller-Jensen, J., Jensen, R.B., Löwe, J., and Gerdes, K. 2002.Prokaryotic DNA segregation by an actin-like filament.EMBO J. 21: 3119–3127.
Møller-Jensen, J., Borch, J., Dam, M., Jensen, R.B., Roepstorff, P.,and Gerdes, K. 2003. Bacterial mitosis: ParM of plasmid R1moves plasmid DNA by an actin-like insertional polymer-ization mechanism. Mol. Cell 12: 1477–1487.
Niki, H., Yamaichi, Y., and Hiraga, S. 2000. Dynamic organiza-tion of chromosomal DNA in Escherichia coli. Genes &Dev. 14: 212–223.
Raj, A. and Peskin, C.S. 2006. The influence of chromosomeflexibility on chromosome transport during anaphase A.Proc. Natl. Acad. Sci. 103: 5349–5354.
Saint-Dic, D., Frushour, B.P., Kehrl, J.H., and Kahng, L.S. 2006.A parA homolog selectively influences positioning of thelarge chromosome origin in Vibrio cholerae. J. Bacteriol.188: 5626–5631.
Sharpe, M.E. and Errington, J. 1996. The Bacillus subtilis soj-spo0J locus is required for a centromere-like function in-volved in prespore chromosome partitioning. Mol. Micro-biol. 21: 501–509.
Teleman, A.A., Graumann, P.L., Lin, D.C., Grossman, A.D.,and Losick, R. 1998. Chromosome arrangement within a
A mitotic-like mechanism in Vibrio cholerae
GENES & DEVELOPMENT 3281
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

bacterium. Curr. Biol. 8: 1102–1109.Thanbichler, M. and Shapiro, L. 2006. MipZ, a spatial regulator
coordinating chromosome segregation with cell division inCaulobacter. Cell 126: 147–162.
Viollier, P.H., Thanbichler, M., McGrath, P.T., West, L., Mee-wan, M., McAdams, H.H., and Shapiro, L. 2004. Rapid andsequential movement of individual chromosomal loci to spe-cific subcellular locations during bacterial DNA replication.Proc. Natl. Acad. Sci. 101: 9257–9262.
Webb, C.D., Teleman, A., Gordon, S., Straight, A., Belmont, A.,Lin, D.C., Grossman, A.D., Wright, A., and Losick, R. 1997.Bipolar localization of the replication origin regions of chro-mosomes in vegetative and sporulating cells of B. subtilis.Cell 88: 667–674.
Webb, C.D., Graumann, P.L., Kahana, J.A., Teleman, A.A., Sil-ver, P.A., and Losick, R. 1998. Use of time-lapse microscopyto visualize rapid movement of the replication origin regionof the chromosome during the cell cycle in Bacillus subtilis.Mol. Microbiol. 28: 883–892.
Wu, L.J. and Errington, J. 2003. RacA and the Soj-Spo0J systemcombine to effect polar chromosome segregation in sporu-lating Bacillus subtilis. Mol. Microbiol. 49: 1463–1475.
Fogel and Waldor
3282 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from

10.1101/gad.1496506Access the most recent version at doi: 20:2006, Genes Dev.
Michael A. Fogel and Matthew K. Waldor segregationA dynamic, mitotic-like mechanism for bacterial chromosome
Material
Supplemental
http://genesdev.cshlp.org/content/suppl/2006/11/16/20.23.3269.DC1
References
http://genesdev.cshlp.org/content/20/23/3269.full.html#ref-list-1
This article cites 49 articles, 22 of which can be accessed free at:
License
ServiceEmail Alerting
click here.right corner of the article or
Receive free email alerts when new articles cite this article - sign up in the box at the top
Copyright © 2006, Cold Spring Harbor Laboratory Press
Cold Spring Harbor Laboratory Press on June 22, 2018 - Published by genesdev.cshlp.orgDownloaded from