a genetic diversity study of antifungal lactobacillus
TRANSCRIPT

Accepted Manuscript
A genetic diversity study of antifungal Lactobacillus plantarum isolates
An-Ran Dong, Van Thi Thuy Ho, Raquel Lo, Nidhi Bansal, Mark S. Turner
PII: S0956-7135(16)30389-9
DOI: 10.1016/j.foodcont.2016.07.026
Reference: JFCO 5152
To appear in: Food Control
Received Date: 10 May 2016
Revised Date: 15 July 2016
Accepted Date: 20 July 2016
Please cite this article as: Dong A.-R., Thuy Ho V.T., Lo R., Bansal N. & Turner M.S., A geneticdiversity study of antifungal Lactobacillus plantarum isolates, Food Control (2016), doi: 10.1016/j.foodcont.2016.07.026.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.
brought to you by COREView metadata, citation and similar papers at core.ac.uk
provided by University of Queensland eSpace

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
A genetic diversity study of antifungal Lactobacillus plantarum isolates. 1
2
An-Ran Donga, Van Thi Thuy Hoa, Raquel Loa, Nidhi Bansala and Mark S. Turnera, b* 3
4
aSchool of Agriculture and Food Sciences, University of Queensland, Brisbane, Australia; 5
bQueensland Alliance for Agriculture and Food Innovation, University of Queensland, 6
Brisbane, Australia. 7
8
9
*Corresponding author: 10
Mark S. Turner, School of Agriculture and Food Sciences, University of Queensland, 11
Brisbane, Australia Tel: +61-7 33651171, Fax: +61-7 33651177, E-mail. 12
14
Running head: DNA typing of antifungal Lactobacillus plantarum. 15
Key words: Lactobacillus plantarum, typing, antifungal, Penicillium commune, MLVA 16
17
18
19

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT1
Abstract 20
Lactobacillus plantarum is a lactic acid bacterium commonly found on fruits and vegetables 21
and also used in a variety of food fermentations. Strains from this species are also regularly 22
reported as having antifungal or probiotic activity. Genotyping methods can be used to 23
differentiate strains of the same species thus determining if strains are related or not. 24
However for L. plantarum, the currently used methods have limitations including DNA band 25
profile interpretation difficulty and cost. In this study, a new genotyping method based on 26
multi-locus variable number tandem repeat analysis (MLVA) analysis was developed and 27
compared to a previously reported randomly amplified polymorphic DNA-PCR (RAPD-28
PCR) method for L. plantarum. With a selection of 13 antifungal strains of L. plantarum 29
isolated from heterogeneous sources (cheese, silage, sauerkraut, vegetables and a probiotic 30
product), RAPD-PCR revealed 9 different profiles resulting in a Hunter-Gaston 31
discrimination index (D-value) of 0.94. The new MLVA method which compares the lengths 32
of 4 repetitive regions within LPXTG motif-containing surface protein genes differentiated 33
the 13 L. plantarum strains into 10 different subtypes leading to a D-value of 0.95. 34
Interestingly 11 additional L. plantarum isolates obtained in a previous study during a screen 35
for antifungal activity against the common cheese spoilage mould Penicillium commune all 36
possessed the same RAPD-PCR and MLVA profile as each other and the commercial 37
probiotic strain L. plantarum 299v. This study demonstrates that the new MLVA method can 38
be used to simply and inexpensively differentiate L. plantarum strains and provide 39
information regarding strain relatedness and thus potential insight into strain properties. 40
41
42
43

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT2
1. Introduction 44
Lactobacillus spp. are members of the lactic acid bacteria (LAB) group, are commonly found 45
in the environment and raw foods and have generally recognized as safe (GRAS) status. 46
They are key components in a variety of fermentations whereby their production of organic 47
acids and other antimicrobial compounds enhance the quality and safety of foods (Reis, 48
Paula, Casarotti, & Penna, 2012). Lactobacillus plantarum is one species of particular 49
interest as it is used in dairy, vegetable and meat fermentations, in the conversion of grass to 50
silage and some strains are marketed as a commercial probiotics with health promoting 51
properties (de Vries, Vaughan, Kleerebezem, & de Vos, 2006). In studies where large LAB 52
collections were screened for antifungal activity, L. plantarum was frequently isolated 53
(Cheong et al., 2014; Crowley, Mahony, & van Sinderen, 2013a; Magnusson, Strom, Roos, 54
Sjogren, & Schnurer, 2003). Antifungal activity of L. plantarum strains has been shown to 55
be due to organic acids, phenyllactic acid, cyclic dipeptides and fatty acids (Crowley, 56
Mahony, & van Sinderen, 2013b), however the genetic basis for the production of these 57
compounds has not been characterised at present. 58
59
Genotyping or subtyping methods can provide information about the relatedness of strains 60
within a species. For L. plantarum, the most commonly used subtyping method has been 61
random amplification of polymorphic DNA (RAPD) PCR (Di Cagno et al., 2010; Johansson, 62
Quednau, Molin, & Ahrne, 1995; Rossetti & Giraffa, 2005). Other subtyping methods such 63
as amplified fragment length polymorphism (AFLP) and restriction fragment length 64
polymorphism (RFLP) have been used to characterise L. plantarum and to differentiate it 65
from closely related species (Johansson, Molin, Pettersson, Uhlen, & Ahrne, 1995; Torriani 66
et al., 2001). Two multilocus sequence typing (MLST) schemes for L. plantarum have been 67
developed (de Las Rivas, Marcobal, & Munoz, 2006; Xu et al., 2015). However MLST can 68

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT3
be costly, involving sequencing of 6-8 housekeeping gene loci for each strain. These MLST 69
methods have identified up to 73 subgroups within L. plantarum suggesting the species is 70
genetically heterogeneous and may have variation in phenotypes such as antifungal activity. 71
72
Although the above genotyping methods have provided valuable information on the L. 73
plantarum species, they have limitations including discriminatory power, difficulties in band 74
profile interpretation or cost. Multilocus variable number tandem repeat analysis (MLVA) is 75
a subtyping method which involves analysis of repetitive regions which can be identified 76
from whole genome sequences using online programs such as Tandem Repeat Finder (TRF) 77
(Benson, 1999). Repeat regions undergo rapid evolution due to slip-strand mispairing 78
leading to good discriminatory power (Keim et al., 2004). DNA fragments are amplified and 79
their lengths can be compared using capillary electrophoresis, high resolution melt analysis 80
(HRMA) or more simply with agarose gel electrophoresis. With respect to foodborne 81
bacteria, MLVA has been successfully used to differentiate strains within Lactobacillus casei 82
(Diancourt et al., 2007), Bifidobacterium longum (Matamoros, Savard, & Roy, 2011), 83
Clostridium tyrobutyricum (Nishihara et al., 2014), Geobacillus spp. (Seale et al., 2012) 84
Krilaviciute & Kuisiene, 2013), Bacillus licheniformis (Dhakal et al., 2013), Staphylococcus 85
aureus (Schouls et al., 2009), Listeria monocytogenes (Chen, Li, Saleh-Lakha, Allen, & 86
Odumeru, 2011) and Salmonella enterica (Kruy, van Cuyck, & Koeck, 2011). 87
88
In this report, a new MLVA method was developed and compared to an established RAPD-89
PCR method for genotyping L. plantarum strains from a variety of sources. Both gave 90
congruent results and showed that antifungal L. plantarum strains are genetically 91
heterogeneous. In addition, the new MLVA method proved to be more discriminatory and 92
easier to interpret than the RAPD-PCR method. 93

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT4
2. Materials and Methods 94
2.1. Bacteria, mould and their growth conditions. 95
Eleven L. plantarum isolates from a variety of sources, one commercial probiotic strain L. 96
plantarum 299v, 12 previously identified antifungal strains and 2 non-antifungal control 97
isolates Weissella soli and Lactococcus lactis (Table 1) were collected from previous research 98
(Cheong et al., 2014). All bacterial isolates were cultured using de Man Rogosa Sharpe 99
(MRS; Oxoid) media containing 1.5% agar and were incubated at 30°C for 24 h under 100
anaerobic conditions (AnaeroGen system; Oxoid). Strains were grown in MRS broth and 101
incubated at 30°C overnight under non-shaking conditions. Bacteria and mould were frozen 102
in 40% glycerol at -80°C for long-term storage. P. commune FRR 4117 was plated out on 103
malt extract agar (MEA; Difco) from a frozen stock, and incubated at room temperature 104
(~23°C) for 7 days. 105
2.2. Antifungal activity testing. 106
All L. plantarum strains were tested against Penicillium commune FRR 4117 on MEA plates 107
using a method similar to that described previously (Cheong et al., 2014). P. commune 108
spores were harvested from MEA plates using 0.2 % (w/v) peptone water (Oxoid), and the 109
concentration was adjusted to an OD600 of 0.5 (~1 × 106 mould spores/ml). The P. commune 110
spore suspension (130 µl) was added to 8 ml of soft MEA (0.7% agar) and was poured onto 111
an MRS agar base (15 ml). L. plantarum strains (5 µl) from pure frozen stocks were spotted 112
onto the fully dried P. commune-containing plates. Plates were first incubated anaerobically 113
at 30°C for 24 h to allow efficient growth of L. plantarum and then incubated aerobically at 114
room temperature (~23°C) for 3 days. Inhibition of P. commune growth was seen when the 115
mould did not grow to the edge of the bacterial colony. Non-antifungal isolates W. soli 33 and 116
L. lactis 38 (Cheong et al., 2014) were included as negative controls in the test and P. 117
commune was able to grow over their colonies. 118

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT5
2.3. RAPD-PCR amplification and gel electrophoresis. 119
The RAPD-PCR method used was adapted from that described previously (Di Cagno et al., 120
2010) using the M13 primer (5'-GAGGGTGGCGGTTCT-3') for amplification. The 121
GenEluteTM Bacterial Genomic DNA Kit (Sigma) was used to extract bacterial DNA from 122
1.5 ml of overnight broth culture using the manufacturer’s instructions, with 200 µl elution 123
solution used. A 25 µl total reaction volume for each sample contained 12.5 µl of GoTaq® 124
Green 2x Master Mix (Promega), 2 µl of primer M13 (final concentration 6 µM), 9.5 µl water 125
and 1 µl of undiluted bacterial DNA template. Negative controls where water replaced the 126
template DNA were included in all runs. Thirty amplification cycles: 94°C / 60 sec, 42°C / 127
20 sec and 72°C / 2 min were carried out after an initial 94°C / 2 min denaturation. A final 128
extension step of 72 °C / 10 min was performed. The PCR products were resolved using a 129
1.5% agarose gel in TAE buffer and stained using SYBR® Safe (Invitrogen). A DNA marker 130
was included in each gel (1 kb plus ladder, Invitrogen). Gels were run at 100 V for 2 h 131
(Mini-300 unit; Major Science) and the banding profiles were captured under UV light 132
(Smart View Pro 1100 Imager System; Major Science). Groupings were identified and the 133
Hunter-Gaston discriminatory index (D-value) was determined (Hunter & Gaston, 1988). 134
2.4. MLVA development. 135
The TRF program (Benson, 1999) was used to identify the DNA repeat regions in L. 136
plantarum WCFS1 (GenBank accession: NC_004567.2). Eighty five repeat regions were 137
located and 23 loci were chosen based on having a repeat sizes ≥ 20bp and a repeat copy 138
number of ≥ 2 based on that done previously (Seale et al., 2012). Primer-BLAST 139
(http://www.ncbi.nlm.nih.gov/tools/primer-blast/) was used to design primers flanking the 140
repeat regions (Ye et al., 2012). Two hundred bp of flanking DNA and the repeat region 141
were analysed by Primer-BLAST and 46 primers for the 23 repeat loci were obtained. Four 142
isolates (278, 291, 1299 and 299v; Table 1) were used to test the 23 sets of primers. The 143

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT6
thermal cycling conditions were: 95°C / 2 min, followed by 30 cycles of 95°C / 15 sec, 63°C 144
/ 30 sec, 72°C / 2 min followed by a final extension step of 72°C / 2 min. The PCR products 145
were resolved using a 2.5% agarose gel with conditions as described above. Four primers 146
which generated PCR products with the greatest variation in length using the 4 tested strains 147
were named variable number tandem repeat 1 (VNTR1), VNTR2, VNTR3 and VNTR4 148
(Table 2) and were used to amplify DNA from all strains. Based on different sizes of the 149
PCR products, bands were scored from 1 to 6, with 1 being the largest PCR fragment. 150
151
3. Results 152
3.1. RAPD-PCR analysis of antifungal L. plantarum isolates from a variety of sources. 153
In previous work, antifungal L. plantarum strains were identified in a screen of isolates from 154
fruits and vegetables (Cheong et al., 2014). Testing of additional food and probiotic L. 155
plantarum strains listed in Table 1 for antifungal activity against P. commune was carried out. 156
It was found that all L. plantarum isolates inhibited P. commune growth similar to that shown 157
for strains 299v, ASCC 3005 and 885 (Figure 1). To explore the genetic relatedness of 158
antifungal L. plantarum strains, a previously described RAPD-PCR genotyping method was 159
adopted (Di Cagno et al., 2010). Thirteen selected L. plantarum isolates from a variety of 160
sources including cheese, silage, sauerkraut, green beans and a commercial probiotic (Table 161
1) were analysed by RAPD-PCR with primer M13. Based on product profiles, the sizes of 162
amplified products ranged from approximately 200bp to 2,500bp, and nine main clusters (A 163
to I) were identified from 13 isolates (Figure 2). Two PCR fragments from isolates ASCC 164
276 and ASCC 281 (group A) were observed and the sizes were around 1,800bp and 350bp. 165
A similar pattern was shown in strain ASCC 2973, however the 350 bp PCR product was not 166
present, therefore it was differentiated into group B. Highly similar banding patterns were 167

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT7
observed from isolates ASCC 278, ASCC 291 and ASCC 3005 (group C). Unique banding 168
patterns divided isolates ASCC 289, ASCC 368, ASCC 1239, ASCC 1299 and ASCC 1398 169
into 5 different groups (D, E, F, G and H). The probiotic strain 299v produced the same 170
banding pattern as strain 897 (group I) obtained previously from the antifungal screening 171
study (Cheong et al., 2014). The 9 groups identified from the 13 isolates resulted in a 172
Hunter-Gaston discrimination index (D-value) of 0.94 (Table 3). 173
3.2. MLVA development and analysis of L. plantarum isolates from a variety of sources. 174
To simplify the interpretation of band profiles and to determine if a more discriminatory 175
method could be devised, a MLVA method was investigated. Eighty-five repetitive regions 176
were identified in the L. plantarum WCFS1 genome (NC_004567.2) using the TRF program. 177
PCR primers were designed from repetitive regions which were ≥20bp in length and 178
consisted of ≥2 repeats. These 23 primer sets were used to amplify DNA from four strains 179
which produced a mix of the same (ASCC 278, ASCC 291) and different (ASCC 1299 and 180
299v) RAPD-PCR subtypes. Four of the 23 primer sets generated PCR products on all tested 181
isolates and showed variation in PCR product lengths. Interestingly all 4 genes within which 182
the VNTRs were located were found to encode repetitive LPXTG motif anchored cell surface 183
proteins. The binding sites for each set of primers along with the repeat numbers for L. 184
plantarum WCFS1 are shown in Figure 3. These four primer sets were used to amplify all 13 185
L. plantarum strains analysed using RAPD-PCR described above (Figure 4). Based on the 186
size differences of the amplified PCR products from the 4 VNTR loci, the 13 isolates could 187
be classified into 10 groups (Table 3). L. plantarum strains ASCC 276 and ASCC 281, 188
which had the same RAPD-PCR profiles, could be differentiated into two groups using 189
MLVA with the length of VNTR4 being clearly different between the two isolates (Figure 190
4D). Like RAPD-PCR, MLVA clustered group C isolates (ASCC 278, ASCC 291 and 191

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT8
ASCC 3005) and group I isolates (299v and 897) together with all the other isolates being 192
different to each other, demonstrating that the two typing methods are highly consistent. The 193
discrimination of the two isolates (ASCC 276 and ASCC 281) from RAPD-PCR group A 194
gave MLVA a slightly improved D-value of 0.95 (Table 3). 195
Eleven other antifungal L. plantarum strains (Table 1) which were isolated from the previous 196
Cheong et al (2014) study were analysed by both RAPD-PCR and MLVA. They all 197
produced identical RAPD-PCR and MLVA profiles as each other and strains 897 and 299v 198
(data not shown). 199
200
4. Discussion 201
In this study a new MLVA method was developed and validated using antifungal L. 202
plantarum strains from a variety of sources. This method was found to have greater 203
discriminatory power than the previously reported RAPD-PCR method and also allow for 204
simpler band profile interpretation. It was observed that genetically distinct strains contained 205
antifungal activity against P. commune which agrees with the knowledge that different 206
antifungal inhibitory compounds (phenyllactic acid, fatty acids and cyclic dipeptides) are 207
produced by different L. plantarum strains. The antifungal mechanism(s) of the strains in the 208
current study are not yet known, however by using genotyping techniques, strains of the same 209
subtype are predicted to produce the same antifungal compound(s). Genotyping methods are 210
also useful for distinguishing unrelated strains which may have different phenotypes leading 211
to different performance abilities in food processing situations. Therefore, the new MLVA 212
method developed here is useful in the early stages of L. plantarum strain characterisation 213
and development for subsequent food technology applications. 214
215

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT9
To date, molecular characterisation and comparison between antifungal isolates of L. 216
plantarum has been limited. However, the complete genome sequence of antifungal L. 217
plantarum 16 has been published (Crowley, Bottacini, Mahony, & van Sinderen, 2013) and 218
genome shuffling of L. plantarum IMAU10014 has been shown to improve antifungal 219
activity (Wang et al., 2013). The two genotyping methods used here were found to provide 220
good discriminatory power and congruent results with a selection of L. plantarum strains. 221
The first method involving RAPD-PCR with an M13 primer generated a high number of 222
bands and generated consistent profiles between runs and also for different DNA templates 223
from the same strain. Other RAPD-PCR primers that have been used for L. plantarum 224
including P4 and P7 (Di Cagno et al., 2010) were also tested, but only a few bands were 225
generated (data not shown). An inherent issue with RAPD-PCR is that slight modifications 226
in DNA extraction methods (Cocconcelli, Porro, Galandini, & Senini, 1995) or PCR cycling 227
conditions can lead to variations in banding profiles and it can be difficult to interpret 228
profiles, in particular where faint bands are present in one isolate and not another. Therefore, 229
a new method was developed based on DNA repeat regions in the L. plantarum genome 230
which also relies on agarose gel electrophoresis but interpretation is made simpler as only one 231
band is produced for each VNTR PCR and the size differences are clear. Gold standard 232
methods including MLST have been developed for L. plantarum (de Las Rivas et al., 2006; 233
Xu et al., 2015), but the cost of Sanger sequencing of 6-8 housekeeping gene loci per isolate 234
prohibits labs from doing this analysis routinely. With the costs of next generation 235
sequencing falling rapidly, it is likely that draft genome sequence-based typing will become 236
commonplace. 237
238
Interestingly it was found that all 4 VNTR loci were from large repetitive LPXTG-family 239
surface proteins. This was not deliberately intended, as the TRF program was used to 240

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT10
identify repeat regions in the L. plantarum genome. In similar studies in B. licheniformis and 241
Geobacillus spp., TRF did not identify any VNTR regions from within surface protein genes 242
(Dhakal et al., 2013; Seale et al., 2012). The number of LPXTG anchoring domain 243
containing proteins in L. plantarum is at the high end when compared to other Firmicutes 244
(Boekhorst, de Been, Kleerebezem, & Siezen, 2005). L. plantarum WCFS1 contains 27 such 245
proteins and these types of proteins are involved in adhesion to bacterial and host cells, 246
surfaces and mucus. Proteins and domains which are repeated can provided greater binding 247
affinity or allow greater extension from the cell surface (Foster & Hook, 1998). Since these 248
proteins are extracellularly exposed, they can undergo fast evolutionary changes in response 249
to changes in the environment in which the cell exists, such as that seen in the case of S. 250
aureus protein A (Shopsin et al., 1999). One LPXTG-containing surface protein that is well 251
characterised in L. plantarum is the mannose specific adhesin Msa (Pretzer et al., 2005) and a 252
repeat region in the encoding gene was identified by the TRF program where 23 loci were 253
initially identified. Primers were designed against the repeat region in the msa gene and PCR 254
was carried out on several of our isolates. Variations in repeat length were seen, a result in 255
agreement with variations in repeat number previously identified (Gross, Snel, Boekhorst, 256
Smits, & Kleerebezem, 2010). However only half of the isolates generated a PCR product, 257
making msa unsuitable for use as a genotyping target. These results are in agreement with 258
Pretzer et al (2005) where the absence of msa in several L. plantarum allowed for the 259
discovery of its role in mannose specific cell agglutination. 260
261
Interestingly all the 12 L. plantarum strains collected in our previous work, in which a 262
collection of 897 LAB were screened for antifungal activity (Cheong et al., 2014), contained 263
identical RAPD-PCR and MLVA genotypes. These isolates were from a variety of sources 264
but the majority were isolated from vegetables from the same retail outlet. Strains shown in 265

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT11
Table 1 with numbers in the 800s were isolated from vegetables purchased from the same 266
store, while strains 170 and 377 were from different stores. It is possible that these 267
vegetables were all harvested from the same farm or farming region and then distributed to 268
different retailers, thereby explaining why they have identical genotypes. It is therefore 269
likely that they are clonal and share the same antifungal mechanism of action. Interestingly 270
they have the same genotype with the commercial probiotic strain 299v. L. plantarum 299v 271
is a well-researched probiotic strain and has shown potential in the treatment of irritable 272
bowel syndrome (Ducrotte, Sawant, & Jayanthi, 2012; Niedzielin, Kordecki, & Birkenfeld, 273
2001) and cardiovascular disease (Naruszewicz, Johansson, Zapolska-Downar, & Bukowska, 274
2002). It was shown that 299v has antifungal activity and therefore may find application in 275
food both as a probiotic and biopreservative. It is also possible that the strains identified 276
from the antifungal LAB screen (Cheong et al., 2014) have probiotic properties similar to 277
299v, but more work is needed to test this hypothesis. 278
279
In conclusion, is was demonstrated that a newly developed MLVA genotyping method can be 280
used to differentiate antifungal L. plantarum strains. Future work understanding the 281
antifungal mechanism of action and the genetic basis of this activity of different strain 282
subtype groups will be of interest. Genotyping methods thus can provide valuable 283
information to underpin the development and application of biopreservative strains to reduce 284
mould spoilage in food. 285
286
287
Acknowledgements 288
We thank Whitney Beddoes for her work in the initial stages of the project and Dairy 289
Innovation Australia Limited for the provision of strains. 290

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT12
291
292
References 293
294
Benson, G. (1999). Tandem repeats finder: a program to analyze DNA sequences. Nucleic 295
Acids Res, 27(2), 573-580. 296
Boekhorst, J., de Been, M. W., Kleerebezem, M., & Siezen, R. J. (2005). Genome-wide 297
detection and analysis of cell wall-bound proteins with LPxTG-like sorting motifs. Journal of 298
Bacteriology, 187(14), 4928-4934. 299
Chen, S., Li, J., Saleh-Lakha, S., Allen, V., & Odumeru, J. (2011). Multiple-locus variable 300
number of tandem repeat analysis (MLVA) of Listeria monocytogenes directly in food 301
samples. International Journal of Food Microbiology, 148(1), 8-14. 302
Cheong, E. Y. L., Sandhu, A., Jayabalan, J., Kieu Le, T. T., Nhiep, N. T., My Ho, H. T., et al. 303
(2014). Isolation of lactic acid bacteria with antifungal activity against the common cheese 304
spoilage mould Penicillium commune and their potential as biopreservatives in cheese. Food 305
Control, 46, 91-97. 306
Cocconcelli, P. S., Porro, D., Galandini, S., & Senini, L. (1995). Development of RAPD 307
protocol for typing of strains of lactic acid bacteria and Enterococci. Letters in Applied 308
Microbiology, 21(6), 376-379. 309
Crowley, S., Bottacini, F., Mahony, J., & van Sinderen, D. (2013). Complete genome 310
sequence of Lactobacillus plantarum strain 16, a broad-spectrum antifungal-producing lactic 311
acid bacterium. Genome Announc, 1(4), e00533. 312
Crowley, S., Mahony, J., & van Sinderen, D. (2013a). Broad-spectrum antifungal producing 313
lactic acid bacteria and their application in fruit models. Folia Microbiologica, 58(4), 291-314
299. 315

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT13
Crowley, S., Mahony, J., & van Sinderen, D. (2013b). Current perspectives on antifungal 316
lactic acid bacteria as natural bio-preservatives. Trends in Food Science & Technology, 33(2), 317
93-109. 318
de Las Rivas, B., Marcobal, A., & Munoz, R. (2006). Development of a multilocus sequence 319
typing method for analysis of Lactobacillus plantarum strains. Microbiology, 152(1), 85-93. 320
de Vries, M. C., Vaughan, E. E., Kleerebezem, M., & de Vos, W. M. (2006). Lactobacillus 321
plantarum: survival, functional and potential probiotic properties in the human intestinal 322
tract. International Dairy Journal, 16(9), 1018-1028. 323
Dhakal, R., Chauhan, K., Seale, R. B., Deeth, H. C., Pillidge, C. J., Powell, I. B., et al. 324
(2013). Genotyping of dairy Bacillus licheniformis isolates by high resolution melt analysis 325
of multiple variable number tandem repeat loci. Food Microbiology, 34(2), 344-351. 326
Di Cagno, R., Minervini, G., Sgarbi, E., Lazzi, C., Bernini, V., Neviani, E., et al. (2010). 327
Comparison of phenotypic (Biolog System) and genotypic (random amplified polymorphic 328
DNA-polymerase chain reaction, RAPD-PCR, and amplified fragment length polymorphism, 329
AFLP) methods for typing Lactobacillus plantarum isolates from raw vegetables and fruits. 330
International Journal of Food Microbiology, 143(3), 246-253. 331
Diancourt, L., Passet, V., Chervaux, C., Garault, P., Smokvina, T., & Brisse, S. (2007). 332
Multilocus sequence typing of Lactobacillus casei reveals a clonal population structure with 333
low levels of homologous recombination. Appl Environ Microbiol, 73(20), 6601-6611. 334
Ducrotte, P., Sawant, P., & Jayanthi, V. (2012). Clinical trial: Lactobacillus plantarum 299v 335
(DSM 9843) improves symptoms of irritable bowel syndrome. World J Gastroenterol, 336
18(30), 4012-4018. 337
Foster, T. J., & Hook, M. (1998). Surface protein adhesins of Staphylococcus aureus. Trends 338
Microbiol, 6(12), 484-488. 339
Gross, G., Snel, J., Boekhorst, J., Smits, M. A., & Kleerebezem, M. (2010). Biodiversity of 340
mannose-specific adhesion in Lactobacillus plantarum revisited: strain-specific domain 341
composition of the mannose-adhesin. Benef Microbes, 1(1), 61-66. 342

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT14
Hunter, P. R., & Gaston, M. A. (1988). Numerical index of the discriminatory ability of 343
typing systems: an application of Simpson's index of diversity. J Clin Microbiol, 26(11), 344
2465-2466. 345
Johansson, M. L., Molin, G., Pettersson, B., Uhlen, M., & Ahrne, S. (1995). Characterization 346
and species recognition of Lactobacillus plantarum strains by restriction-fragment-length-347
polymorphism (RFLP) of the 16s ribosomal RNA gene. Journal of Applied Bacteriology, 348
79(5), 536-541. 349
Johansson, M. L., Quednau, M., Molin, G., & Ahrne, S. (1995). Randomly amplified 350
polymorphic DNA (RAPD) for rapid typing of Lactobacillus plantarum strains. Letters in 351
Applied Microbiology, 21(3), 155-159. 352
Keim, P., Van Ert, M. N., Pearson, T., Vogler, A. J., Huynh, L. Y., & Wagner, D. M. (2004). 353
Anthrax molecular epidemiology and forensics: using the appropriate marker for different 354
evolutionary scales. Infection Genetics and Evolution, 4(3), 205-213. 355
Kruy, S. L., van Cuyck, H., & Koeck, J. L. (2011). Multilocus variable number tandem repeat 356
analysis for Salmonella enterica subspecies. European Journal of Clinical Microbiology and 357
Infectious Diseases, 30(4), 465-473. 358
Magnusson, J., Strom, K., Roos, S., Sjogren, J., & Schnurer, J. (2003). Broad and complex 359
antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol 360
Lett, 219(1), 129-135. 361
Matamoros, S., Savard, P., & Roy, D. (2011). Genotyping of Bifidobacterium longum subsp. 362
longum strains by multilocus variable number of tandem repeat analysis. Journal of 363
Microbiological Methods, 87(3), 378-380. 364
Naruszewicz, M., Johansson, M. L., Zapolska-Downar, D., & Bukowska, H. (2002). Effect of 365
Lactobacillus plantarum 299v on cardiovascular disease risk factors in smokers. American 366
Journal of Clinical Nutrition, 76(6), 1249-1255. 367
Niedzielin, K., Kordecki, H., & Birkenfeld, B. (2001). A controlled, double-blind, 368
randomized study on the efficacy of Lactobacillus plantarum 299v in patients with irritable 369

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT15
bowel syndrome. European Journal of Gastroenterology and Hepatology, 13(10), 1143-370
1147. 371
Nishihara, M., Takahashi, H., Sudo, T., Kyoi, D., Kawahara, T., Ikeuchi, Y., et al. (2014). 372
Multilocus variable-number of tandem repeat analysis (MLVA) for Clostridium 373
tyrobutyricum strains isolated from cheese production environment. International Journal of 374
Food Microbiology, 190, 61-65. 375
Pretzer, G., Snel, J., Molenaar, D., Wiersma, A., Bron, P. A., Lambert, J., et al. (2005). 376
Biodiversity-based identification and functional characterization of the mannose-specific 377
adhesin of Lactobacillus plantarum. Journal of Bacteriology, 187(17), 6128-6136. 378
Reis, J. A., Paula, A. T., Casarotti, S. N., & Penna, A. L. B. (2012). Lactic acid bacteria 379
antimicrobial compounds: Characteristics and applications. Food Engineering Reviews, 4(2), 380
124-140. 381
Rossetti, L., & Giraffa, G. (2005). Rapid identification of dairy lactic acid bacteria by M13-382
generated, RAPD-PCR fingerprint databases. Journal of Microbiological Methods, 63(2), 383
135-144. 384
Schouls, L. M., Spalburg, E. C., van Luit, M., Huijsdens, X. W., Pluister, G. N., van Santen-385
Verheuvel, M. G., et al. (2009). Multiple-locus variable number tandem repeat analysis of 386
Staphylococcus aureus: comparison with pulsed-field gel electrophoresis and spa-typing. 387
PLoS One, 4(4), e5082. 388
Seale, R. B., Dhakal, R., Chauhan, K., Craven, H. M., Deeth, H. C., Pillidge, C. J., et al. 389
(2012). Genotyping of present-day and historical Geobacillus species isolates from milk 390
powders by high-resolution melt analysis of multiple variable-number tandem-repeat loci. 391
Appl Environ Microbiol, 78(19), 7090-7097. 392
Shopsin, B., Gomez, M., Montgomery, S. O., Smith, D. H., Waddington, M., Dodge, D. E., et 393
al. (1999). Evaluation of protein A gene polymorphic region DNA sequencing for typing of 394
Staphylococcus aureus strains. J Clin Microbiol, 37(11), 3556-3563. 395

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT16
Torriani, S., Clementi, F., Vancanneyt, M., Hoste, B., Dellaglio, F., & Kersters, K. (2001). 396
Differentiation of Lactobacillus plantarum, L. pentosus and L. paraplantarum species by 397
RAPD-PCR and AFLP. Systematic and Applied Microbiology, 24(4), 554-560. 398
Wang, H. K., Sun, Y., Chen, C., Sun, Z., Zhou, Y. C., Shen, F. D., et al. (2013). Genome 399
shuffling of Lactobacillus plantarum for improving antifungal activity. Food Control, 32(2), 400
341-347. 401
Xu, H., Liu, W., Zhang, W., Yu, J., Song, Y., Menhe, B., et al. (2015). Use of multilocus 402
sequence typing to infer genetic diversity and population structure of Lactobacillus 403
plantarum isolates from different sources. Bmc Microbiology, 15(1), 241. 404
Ye, J., Coulouris, G., Zaretskaya, I., Cutcutache, I., Rozen, S., & Madden, T. (2012). Primer-405
BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC 406
Bioinformatics, 13, 134. 407
408
409
410
Figure legends. 411
Figure 1. Antifungal activity assay of L. plantarum strains 299v, ASCC 3005 and 885 and 412
negative control LAB W. soli 33 and L. lactis 38 against P. commune. 413
414
Figure 2. RAPD-PCR analysis of selected L. plantarum strains using the M13 primer. The 415
groups are indicated above the strain name with a single capital letter. M is the DNA marker 416
and NC is the no template negative control PCR. 417
418

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT17
Figure 3. Selected VNTR regions from four LPXTG motif-containing surface protein 419
encoding genes from L. plantarum WCFS1. Primer binding sites are shown as arrow heads 420
and repeat numbers from strain WCFS1 are shown. The numbered ruler refers to nucleotides. 421
422
Figure 4. MLVA of selected L. plantarum strains. Agarose gels show VNTR1 (A), VNTR2 423
(B), VNTR3 (C) and VNTR4 (D) PCR products. M is the DNA marker and NC is the no 424
template negative control PCR. Scores for each band are based on migration, with the lowest 425
number corresponding to the largest sized PCR product. Equivalent sized PCR products are 426
assigned the same number. 427
428
429
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT18
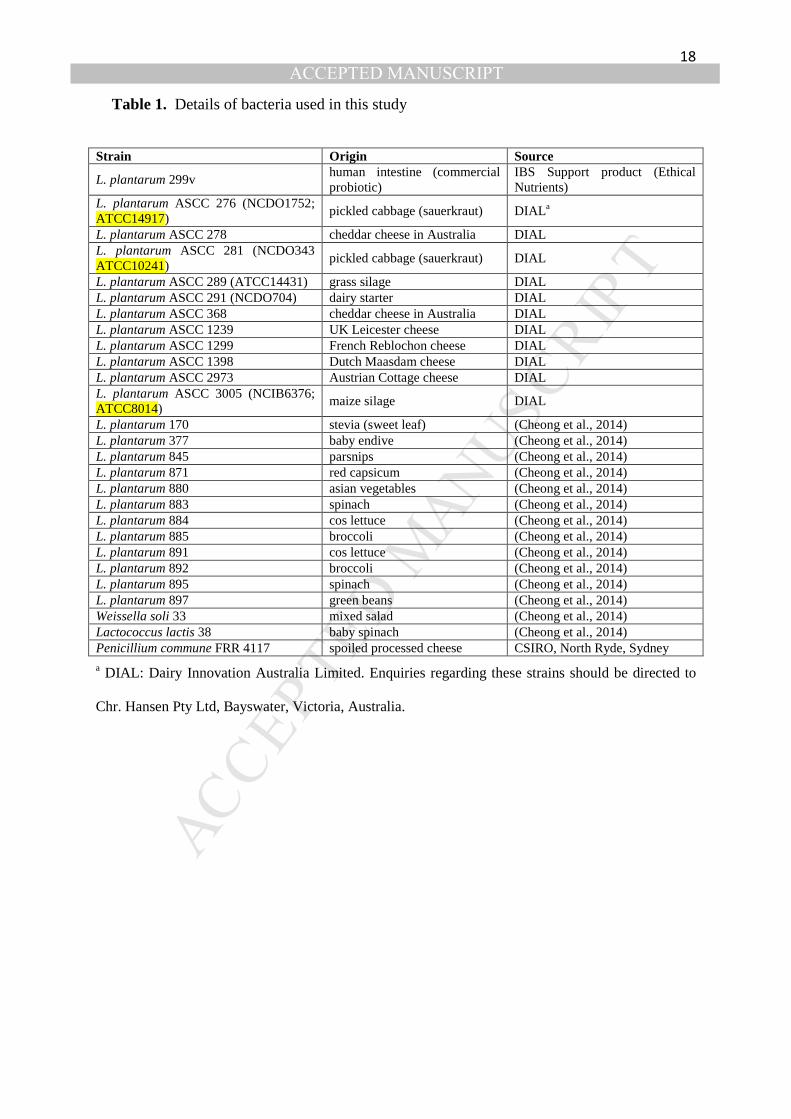
Table 1. Details of bacteria used in this study
Strain Origin Source
L. plantarum 299v human intestine (commercial probiotic)
IBS Support product (Ethical Nutrients)
L. plantarum ASCC 276 (NCDO1752; ATCC14917)
pickled cabbage (sauerkraut) DIALa
L. plantarum ASCC 278 cheddar cheese in Australia DIAL L. plantarum ASCC 281 (NCDO343 ATCC10241)
pickled cabbage (sauerkraut) DIAL
L. plantarum ASCC 289 (ATCC14431) grass silage DIAL L. plantarum ASCC 291 (NCDO704) dairy starter DIAL L. plantarum ASCC 368 cheddar cheese in Australia DIAL L. plantarum ASCC 1239 UK Leicester cheese DIAL L. plantarum ASCC 1299 French Reblochon cheese DIAL L. plantarum ASCC 1398 Dutch Maasdam cheese DIAL L. plantarum ASCC 2973 Austrian Cottage cheese DIAL L. plantarum ASCC 3005 (NCIB6376; ATCC8014)
maize silage DIAL
L. plantarum 170 stevia (sweet leaf) (Cheong et al., 2014) L. plantarum 377 baby endive (Cheong et al., 2014) L. plantarum 845 parsnips (Cheong et al., 2014) L. plantarum 871 red capsicum (Cheong et al., 2014) L. plantarum 880 asian vegetables (Cheong et al., 2014) L. plantarum 883 spinach (Cheong et al., 2014) L. plantarum 884 cos lettuce (Cheong et al., 2014) L. plantarum 885 broccoli (Cheong et al., 2014) L. plantarum 891 cos lettuce (Cheong et al., 2014) L. plantarum 892 broccoli (Cheong et al., 2014) L. plantarum 895 spinach (Cheong et al., 2014) L. plantarum 897 green beans (Cheong et al., 2014) Weissella soli 33 mixed salad (Cheong et al., 2014) Lactococcus lactis 38 baby spinach (Cheong et al., 2014) Penicillium commune FRR 4117 spoiled processed cheese CSIRO, North Ryde, Sydney
a DIAL: Dairy Innovation Australia Limited. Enquiries regarding these strains should be directed to
Chr. Hansen Pty Ltd, Bayswater, Victoria, Australia.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
19
Table 2. Details of VNTR regions analysed in L. plantarum
VNTR locus
Locus tag of gene in WCFS1
Repeating sequences Copy number in WCFS1
Gene product Primers D-value
VNTR 1 lp_2588 AACCAGGAGTTACTGAACCGGAAAAACCTGGGACTACCGAACCCGAGA
8.5 LPXTG anchor cell wall protein with collagen binding domain (Pfam05737)
5’-CCAGACGTTCCCGAAAATCC-3’ 5’-GCTGTTCTGGTGAGACGGTA-3’
0.79
VNTR 2 lp_2958 TGGTCAGAACTAGAGTCCACAGCGCTTGAACTTTCCGAACTAGAG
7.3 WapA cell surface protein with LPXTG anchor and collagen binding domain (Pfam05737)
5’-CTCGAGCTTTCAGAACCGGA-3’ 5’-AAGTCGTTAACGGTGGGAGC-3’
0.59
VNTR 3 lp_3001 GGTTTAGTCGTGTTGCTACCATTTTCATCC
3.2 LPXTG anchor cell wall protein 5’-CCGTGGTGCTAGTTTCATGC-3’ 5’-TAAACCCGACGAAAACGGCA-3’
0.73
VNTR 4 lp_3059 TTGACCCGGTTCTTCCGGTTGACTCGGTTGCTCGGG
5.6 AapA cell surface protein with LPXTG anchor and mucus binding domain (Pfam06458)
5’-GCAGTTCCCCTGGTTTTTCG-3’ 5’-GCCGATCAGGTTGTAGTGGT-3’
0.79
430

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT20
Table 3. Clusters identified by RAPD-PCR and VNTR with D-values
Isolate RAPD-PCR type VNTR Score and type ASCC 276 A 3-1-3-2 = A ASCC 281 A 3-1-3-4 = A' ASCC 2973 B 5-1-3-4 = B ASCC 278 C 4-3-1-3 = C ASCC 291 C 4-3-1-3 = C ASCC 3005 C 4-3-1-3 = C ASCC 289 D 1-2-2-2 = D ASCC 368 E 3-1-2-4 = E ASCC 1239 F 3-1-4-2 = F ASCC 1299 G 2-2-2-4 = G ASCC 1398 H 6-1-2-3 = H
299v I 4-1-2-1 = I 897 I 4-1-2-1 = I
D-value 0.94 0.95 431
432

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 1.
299v
ASCC 3005 38
33
885

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 2.
ASC
C 2
973
M NC ASC
C 2
76
ASC
C 2
81
ASC
C 2
78
ASC
C 2
91
ASC
C 3
005
ASC
C 2
89
ASC
C 3
68
ASC
C 1
239
ASC
C 1
299
ASC
C 1
398
299v
897
M
A B C D E F G H I
12,000
bp
5,000
2,0001,650
1,000850
650
400
300
200

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 3.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 4.
bpAVNTR1
BVNTR2
CVNTR3
DVNTR4
ASC
C 3
005
ASC
C 2
973
ASC
C 1
398
ASC
C 1
299
ASC
C 1
239
ASC
C 3
68
ASC
C 2
91
ASC
C 2
89A
SCC
281
ASC
C 2
78
ASC
C 2
76
M 299v
897
NCM

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Highlights
• A new MLVA method for typing Lactobacillus plantarum strains was developed
• MLVA targeted repetitive regions within 4 genes encoding LPXTG surface proteins
• MLVA was more discriminatory than a previously reported RAPD-PCR method
• Antifungal L. plantarum strains are genetically heterogeneous