abkürzungsliste - tu darmstadt publication servicetuprints.ulb.tu-darmstadt.de/491/2/anhang.pdf ·...
TRANSCRIPT

Anhang
A-1
Abkürzungsliste
ANOVA Varianzanalyse
AO Acridin-Orange (Fluoreszenz-Farbstoff, AppliChem, Darmstadt)
BSA Bovine Serum Albumin (Serva, Heidelberg)
C Kohlenstoff
CLPP „Community Level Physiological Profile“, metabolisches Potenzial einer Mikroorganismen-Gesellschaft
dd doppelt destilliert
DFA „Discriminant Function Analysis“, Diskriminanzanalyse
DTT Dithiotheitol (Biomol, Ilvesheim)
EtOH Ethanol
FG Frischgewicht (Material mit Papiertuch abgetupft)
GLM „Generalized Linear Modell“, Varianzanalyse für ungleich replizierte Behandlungen
GUS β-Glucoronidase (Reportergen)
MANOVA Multivariate Varianzanalyse
MPN “Most Problable Number”
MU Methyl-Umbelliferon (Fluka, Buchs, Schweiz)
MUG 4-Methylumbelliferyl-b-D-glucuronid, GUS-Substrat (Fluka, Buchs, CH)
N Stickstoff
NB “Nutrient Broth”, siehe Rezepturen
NMAS “Neff´s Modified Amoebae Salinae”, siehe Rezepturen
OD Optische Dichte bei 590 nm
P Phosphor
PGPR “Plant Growth Promoting Rhizobacteria”, Wuchsfördernde Bakterien
PhAR “Photosynthetic active radiation”, Photosynthetisch wirksame Strahlung im Bereich von 400- 700 nm Wellenlänge (µmol/m2*s)
PMSF Polymethylsulfonylfluorid (AppliChem, Darmstadt)
PVP Polyvinylpyrrolidin (Merck, Darmstadt)
SD Standardabweichung
SDS Natriumdodecylsulfat (AppliChem, Darmstadt)
TG Trockengewicht (gemessen nach 3 d bei 70°C)
X-Gluc 5-Bromo-4-chloro-3-indolyl-β-D-glucuronid, GUS-Substrat (Molecular Probes, Leiden, Niederlande)

Anhang
A-2
Rezepturen Gelistet nach der Reihenfolge im Text. M = Makronährstoffe; m = Mikronährstoffe
NB (Bakterien & Amöben) “Nutrient Broth” (Sigma-Aldrich, München);
verwendete Konzentration 1/10NB = 0,8 g/l
NMAS (Bakterien & Amöben) “Neff´s Modified Amoebae Salinae” (nach Page 1976)
4,8 mg NaCl
0,16 mg MgSO4*7H2O
0,24 mg CaCl2*6H2O
5,68 mg Na2HPO4
5,44 mg KH2PO4
ad. 1000 ml H2Odest
Hoagland´s No. 2 Basal Salt
Mixture (Aeroponisches
System, Kap. 3)
nach Hoagland & Arnon (1938)
120,38 mg MgSO4
303,3 mg KNO3
328,2 mg Ca(NO3)2
57,515 mg (NH4)H2PO4
2,66 mg NaFeEDTA
0,905 mg MnCl2*4H2O
0,11 mg ZnSO4*7H2O
0,93 mg H3BO3
0,04 mg CuSO4*5H2O
ad. 1000 ml H2Odest
Murashige & Skoog-
Nährmedium (Agarsysteme,
Kap. 4)
nach Murashige & Skoog (1962)
332,16 mg CaCl2
0,025 mg CoCl2*6H2O
0,025 mg CuSO4*5H2O
27,8 mg FeSO4. 7H2O
6,2 mg H3BO3
170 mg KH2PO4
0,83 mg KI
1.900 mg KNO3

Anhang
A-3
370 mg MgSO4*7H2O
16,9 mg MnSO4*H2O
37,3 mg Na2*EDTA
0,25 mg Na2MoO4.2H2O
1.650 mg NH4NO3
8,6 mg ZnSO4*7H2O
100 mg i-Inositol
0,4 mg Thiamine*HCl
ad. 1000 ml H2Odest
Hybridisations-Puffer (FISH,
Kap. 4)
180 µl NaCl 5M
20 µl Tris (HCl) pH 8 1M
1 µl SDS 10%
200 µl para-Formaldehyd 4%
600 µl ddH2O
Wasch-Puffer (FISH, Kap. 4) 2150 µl NaCl 5M
1000 µl Tris (HCl) pH 8 1M
50 µl SDS 10%
500 µl EDTA 0,5M
ad. 50 ml ddH2O
modifiziertes Strullu-Romand-
Medium (VA-Mykorrhiza,
Kap. 5)
nach Declerck et al. (1998)
739 mg MgSO4*7H2O
76 mg KNO3
65 mg KCl
4,1 mg KH2PO4
359 mg Ca(NO3)2*4H2O
8 mg NaFeEDTA
2,45 mg MnSO4*4H2O
0,29 mg ZnSO4*7H2O
1,86 mg H3BO3
0,24 mg CuSO4*5H2O
0,035 mg (NH4)Mo7O24*4H2O
0,0024 mg Na2MoO4*2H2O

Anhang
A-4
0,9 mg Ca-Panthotenat
0,0009 mg Biotin
0,9 mg Pyridoxin
1 mg Thiamin
0,4 mg Cyanocobalamin
1 mg Nicotinsäure
10000 mg Sucrose
3000 mg Gel Gro (ICN, Cost Mesa, USA)
ad. 1000 ml H2Odest
pH 5,5
Inkubationslösung
(histochemischer GUS-
Nachweis, Kap. 6)
Stammlösungen:
X-Gluc: 20mM in Dimethylformalid gelöst (10,44
mg/ml)
Phosphat-Puffer: 1,31 g Na2HPO4 x 2H2O
1,73 g NaH2PO4 x 2H2O
ad. 25 ml H2Obidest, pH 7
K-Ferrocyanid (II) in H2Obidest (21,12 mg/10 ml), pH 7
K-Ferricyanid (III) in H2Obidest (16,46 mg/10 ml), pH 7
EDTA in H2Obidest (93,16 mg/10 ml), pH 7
Inkubationslösung:
610 µl H2Obidest
100 µl Phosphat-Puffer
100 µl K-Ferrocyanid (II)
100 µl K-Ferricyanid (III)
40 µl EDTA
50 µl X-Gluc
Assay-Puffer (fluorometrischer
GUS-Nachweis, Kap. 6)
Stammlösungen:
DTT in H2Obidest (1,54 g/10 ml)
SDS in H2Obidest (1g/ 10 ml)
PMSF in Isopropanol (17,42 mg/1ml)
Phosphat-Puffer: 1,31 g Na2HPO4 x 2H2O
1,73 g NaH2PO4 x 2H2O
ad. 25 ml H2Obidest, pH 7

Anhang
A-5
EDTA in H2Obidest (93,16 mg/10 ml), pH 7
Extraktionspuffer:
86,5 µl H2Obidest
10,0 µl Phosphat-Puffer
0,4 µl EDTA
1,0 µl DTT
1,0 µl SDS
1,0 µl TritonX-100 (Sigma-Aldrich, München)
0,1 µl PMSF
1,5 g PVP
Assay-Puffer:
MUG 8,13 mg/ 20 ml Extraktionspuffer
Eichgerade für
fluorometrischen GUS-
Nachweis (Kap. 6)
Stammlösung 1mM MU
Nullwert: Stopp-Puffer (siehe unten)
Konzentrationen von 20-8000 µmol in Stopp-Puffer
Gesamtproteingehalt,
Bestimmung (Kap. 6)
nach Popov (1975)
BSA-Stammlösung:
BSA in H2Obidest (0,5 mg/ml)
Färbelösung:
Amidoschwarz 10 B (Sigma-Aldrich, München) (0,13
g in 1 ml 100% Essigsäure und 9 ml 100% Methanol),
gefiltert
Waschlösungen:
Acid I 10 ml 100% Essigsäure ad. 100 ml 100%
Methanol
Acid II 10 ml 100% Essigsäure ad. 100 ml 99,8%
Ethanol
Gebrauchslösung:
0,5 ml Färbelösung + 24,4 ml Acid I
Stop-Puffer (fluorometrischer
GUS-Nachweis, Kap. 6)
Na2CO3 (2,12 g/100 ml)

Anhang
A-6
Tabellenverzeichnis Tab. 1: Ergebnisse einer einfaktoriellen ANOVA zum Einfluss von Amöben auf Parameter
des Wurzelsystems von L. perenne im ersten aeroponischen Versuch 39
Tab. 2: Ergebnisse einer zweifaktoriellen ANOVA zum Einfluss der blockweisen Aufstellung und von Amöben auf die Wurzellänge, die Anzahl der Wurzelspitzen und die Wurzelverzweigung von O. sativa und L. perenne im zweiten aeroponischen Versuch
40
Tab. 3: Ergebnisse der DFA zum Einfluss der Pflanzenart und von Amöben auf das mikrobielle Muster der Substratnutzung von 31 Kohlenstoffquellen
41
Tab. 4: Ergebnisse der DFA zum Einfluss der Pflanzenart (O. sativa/ L. perenne) und von Amöben auf das mikrobielle Muster der Substratnutzung der Kohlenstoffquellen Äpfelsäure, D-Xylose, L-Asparagin, 4-Hydroxy-benzoesäure und L-Threonin
42
Tab. 5: Ergebnisse von zweifaktoriellen Varianzanalysen zum Einfluss von Pflanzenart und Amöben auf die Substratnutzung verschiedener Kohlenstoffquellen auf Basis der maximalen Steigungswerte oder der Zeit zum Erreichen der maximalen Steigung
43
Tab. 6: Ausgewählte Vertreter der Proteobakterien nach Subgruppen und Funktion in der
Rhizosphäre. 48
Tab. 7: Ergebnisse einer einfaktoriellen ANOVA für den Einfluss von Amöben auf die Wurzellänge, Anzahl der Wurzelspitzen und Wurzelverzweigung von L. perenne in Agarsystemen
54
Tab. 8: Ergebnisse einer einfaktoriellen ANOVA für den Einfluss von Amöben auf die Wurzellänge, Anzahl der Wurzelspitzen und Wurzelverzweigung von O. sativa in Agarsystemen
54
Tab. 9: Ergebnisse einer dreifaktoriellen ANOVA zum Einfluss der Pflanzenart, von Mykorrhiza und Amöben auf Gesamt-, Spross- und Wurzelbiomasse bzw. auf das Spross/Wurzel-Verhältnis von P. lanceolata und L. perenne in Monokulturen
67
Tab. 10: Ergebnisse einer zweifaktoriellen ANOVA zum Einfluss von Mykorrhiza und Amöben auf Gesamt-, Spross- und Wurzelbiomasse von P. lanceolata und L. perenne im Konkurrenzversuch und auf das Spross/Wurzel-Verhältnis
68
Tab. 11: Ergebnisse einer dreifaktoriellen ANOVA zum Einfluss der Pflanzenart, von Mykorrhiza und Amöben auf die Wurzellänge, Anzahl der Wurzelspitzen und die Verzweigung von P. lanceolata und L. perenne in Monokulturen
70
Tab. 12: Ergebnisse einer vierfaktoriellen ANOVA für den Einfluss von Amöben und Mykorrhiza bzw. der zusätzlichen Zugabe von Stickstoff und Phosphor auf den maximalen Anstieg der mikrobiellen Respiration in der assimilativen Phase nach C-Substratinduktion im Konkurrenzversuch
74
Tab. 13: Ergebnisse einer zweifaktoriellen ANOVA zum Einfluss von Amöben und Transformante bzw. Ökotyp auf Wurzellänge, Anzahl der Wurzelspitzen und Verzweigung von A. thaliana im ersten Versuch
87

Anhang
A-7
Tab. 14: Ergebnisse einer zweifaktoriellen ANOVA zum Einfluss der Behandlung mit NPA und Amöben auf die Wurzellänge, die Anzahl der Wurzelspitzen und die Verzweigung der Transformanten DR5 im zweiten Versuch
88
Tab. 15: Ergebnisse von zweifaktoriellen Varianzanalysen zum Einfluss der Behandlung mit NPA (NPA) und Amöben (AMO) auf die als Farbintensität und GUS-Aktivität gemessene Expression von DR5::GUS und ARR5::GUS im zweiten Versuch
91
Abbildungsverzeichnis Abb. 1: Modell des “Microbial loop” nach Clarholm (1985) 4
Abb. 2: Modell der „Hormonal interactions“ nach Bonkowski & Brandt (2002) 6
Abb. 3: Aufbau einer primären Wurzel 7
Abb. 4: Gliederung der durchgeführten Versuche nach Untersuchungszielen und Substratkomplexität
12
Abb. 5: Aufbau eines aeroponischen Systems 28
Abb. 6: Einfluss von Amöben auf den täglichen Längenzuwachs der längsten Wurzel in cm bei O. sativa und L. perenne im ersten aeroponischen Versuch
37
Abb. 7: Einfluss von Amöben auf die Spross-Biomasse von O. sativa und L. perenne im zweiten aeroponischen Versuch
38
Abb. 8: Einfluss von Amöben auf die Anzahl und die Gesamtlänge von Haupt- und Seitenwurzeln von L. perenne im ersten aeroponischen Versuch
39
Abb. 9: Einfluss von Amöben auf die Wurzel-Gesamtlänge und die Anzahl an Wurzelspitzen von O. sativa und L. perenne im zweiten aeroponischen Versuch
40
Abb. 10: Einfluss der Pflanzenart und von Amöben auf die Diskriminanzanalyse des Musters der Substratnutzung von 31 C-Quellen von mikrobiellen Rhizoplanen-Gesellschaften
41
Abb. 11: Einfluss der Pflanzenart und von Amöben auf die Diskriminanzanalyse des Musters der Substratnutzung von fünf Substraten von mikrobiellen Rhizoplanen-Gesellschaften
42
Abb. 12: Einfluss von Amöben auf Wurzel-Gesamtlänge und Anzahl an Wurzelspitzen von L. perenne in Agarsystemen
53
Abb. 13: Einfluss von Amöben auf Anzahl und mittlere Länge von Primärwurzeln von O. sativa in Agarsystemen
55
Abb. 14: Einfluss von Amöben auf die Initiation von Primordien und die Streckung von Primordien zu Lateralwurzeln bei O. sativa in Agarsystemen
55
Abb. 15: Einfluss von Amöben auf die Verteilung von Lateralwurzeln von O. sativa nach Längenklassen in Agarsystemen
56

Anhang
A-8
Abb. 16: Einfluss von Amöben auf die Verteilung der Bakterienzellen nach Volumenklassen
57
Abb. 17: Einfluss von Amöben auf die Dichte von Eubakterien und α-Proteobakterien bzw. auf die Dichte von β-, γ-Proteobakterien und der Nitrospira-Gruppe in der Rhizosphäre von O. sativa
57
Abb. 18: Einfluss von Mykorrhiza und Amöben auf die Spross- und Wurzelbiomasse von P. lanceolata und L. perenne in Monokulturen
68
Abb. 19 Einfluss von Amöben und Mykorrhiza auf die Sprossbiomasse von P. lanceolata (linke Balken) und L. perenne (rechte Balken) und deren gemeinsame Wurzelbiomasse im Konkurrenzversuch
69
Abb. 20: Einfluss von Amöben und Mykorrhiza auf den N-Gehalt im Spross von P. lanceolata und L. perenne in Monokulturen
70
Abb. 21: Einfluss von Amöben und Mykorrhiza auf die Gesamtwurzellänge von P. lanceolata und L. perenne in Monokulturen
71
Abb. 22: Einfluss von Amöben und Mykorrhiza auf die Gesamtwurzellänge von P. lanceolata und L. perenne im Konkurrenzversuch
72
Abb. 23: Einfluss von Amöben und Mykorrhiza auf die mikrobielle Basalatmung in der Rhizosphäre von L. perenne in Monokultur und der Mischkultur von P. lanceolata und L. perenne
73
Abb. 24: Einfluss von Amöben auf die Wurzel-Gesamtlänge und die Anzahl an Wurzelspitzen der Transformanten ARR5 und DR5 im ersten Versuch
87
Abb. 25: Einfluss von Amöben und NPA-Behandlung auf die Anzahl an Wurzelspitzen der Transformanten DR5 im zweiten Versuch
88
Abb. 26: Einfluss von Amöben auf die GUS-Expression der Cytokinin-sensitiven Transformante ARR5 und der Auxin-sensitiven Transformante DR5 im ersten Versuch
89
Abb. 27: Einfluss von Amöben und NPA-Behandlung auf die DR5::GUS-Expression in Wurzelspitzen von Keimlingen und Altpflanzen im zweiten Versuch
90
Abb. 28: Einfluss von Amöben und NPA-Behandlung auf die ARR5::GUS-Expression in Wurzelspitzen von Keimlingen und Expression bezüglich des Proteingehalts von Altpflanzen im zweiten Versuch
91
Abb. 29: Schema der Seitenwurzelbildung, Seitenwurzelstreckung und mögliche Zonen der Nährstoffaufnahme (3mm Radius) an einer typischen Hauptwurzel von O. sativa ohne und mit Amöben
97

Anhang
A-9
Bilder (in der Reihenfolge der beschriebenen Versuche)
Aeroponisches System
Bild 1: Aufbau mit Zentralbehältern und Wurzelkammern im ersten Versuch.
Bild 2: Reis-Keimlinge in präparierter Pipettenspitze mit Nährmedium (links) und bei der Inokulation (rechts).
Anhang
A-10
Aeroponisches System
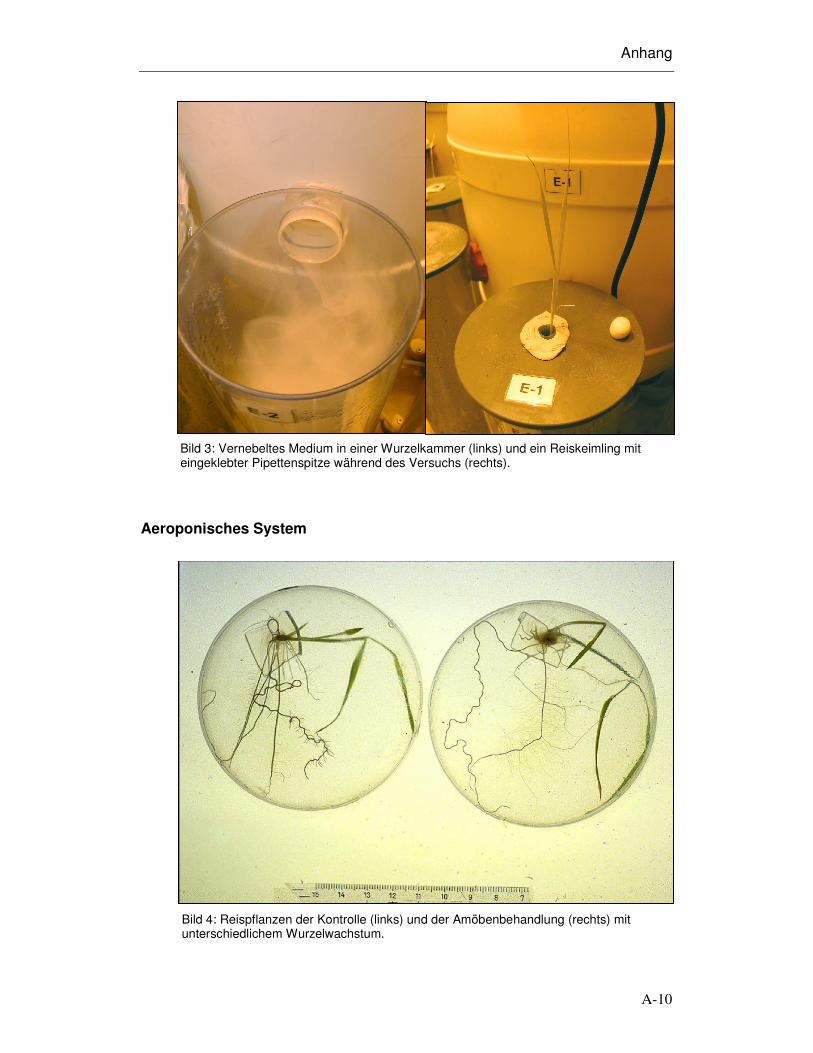
Bild 3: Vernebeltes Medium in einer Wurzelkammer (links) und ein Reiskeimling mit eingeklebter Pipettenspitze während des Versuchs (rechts).
Bild 4: Reispflanzen der Kontrolle (links) und der Amöbenbehandlung (rechts) mit unterschiedlichem Wurzelwachstum.

Anhang
A-11
Bild 5: Fluoreszenz-in-situ-Hybridisation (FISH) von Eubakterien (blau) und α-Proteobakterien (rot/rosa) in der Rhizosphäre von Reis der Kontrollbehandlung (links) und der Amöbenbehandlung (rechts).
Bild 6: Fluoreszenz-in-situ-Hybridisation (FISH) von β-Proteobakterien (blau), γ-Proteobakterien (grün) und Bakterien der Nitrospira-Gruppe (rot) in der Rhizosphäre von Reis der Kontrollbehandlung (links) und der Amöbenbehandlung (rechts).

Anhang
A-12
Bodensysteme
Bild 8: Arabidopsis thaliana in Kulturflaschen während des ersten Versuchs zur Untersuchung des Einflusses von Amöben auf die Auxin- und Cytokinin-Aktivität in Wurzelspitzen. Bei der hier gezeigten Transformante ARR5 ist die Rosette schwach ausgeprägt.
Bild 7: Plantago lanceolata in Kulturflaschen am Ende des Versuches zur Untersuchung der Wechselwirkungen zwischen Protozoen und VA-Mykorrhiza. Die Aluminiumfolie verhindert die Algenbildung in den Gefäßen im Bereich des Bodens.

Anhang
A-13
GUS-Expression in den Wurzelspitzen
Bild 9: Transparente Boxen mit gaspermeabler Membran, wie sie im zweiten Versuch mit Arabidopsis thaliana (im Bild zu einem frühen Versuchszeitpunkt) und im Konkurrenzversuch zwischen Lolium perenne und Plantago lanceolata eingesetzt wurden.
Bild 10: GUS-Expression in den Wurzelspitzen der Auxin-sensitiven Transformante DR5 (links) und der Cytokinin-sensitiven Transformante ARR5. Die Rahmen stellen den Messbereich dar, in dem die Farbintensität der Wurzelspitze und des Hintergrundes gemessen wurde. Für DR5-Wurzelspitzen wurden quadratische Messbereiche und für ARR5-Wurzelspitzen rechteckige Messbereiche mit einem Kantenverhältnis von 2:1 ausgewählt, da sich die Bereiche der GUS-Aktivitäten räumlich unterschieden.

Publikationsliste
Teile der vorliegenden Arbeit sowie weiterer Projekte während meiner
Promotionszeit wurden wie folgt publiziert bzw. auf Tagungen präsentiert:
BONKOWSKI M, KREUZER K, SCHEU S (2001) Biodiversity of protozoa in the
rhizosphere of plants: effects on activity, performance and diversity of
microorganisms and on plant growth. Poster, Tagung “Biodiversity and Global
Change” (BIOLOG) des BMBF, Bonn.
KREUZER K, SCHEU S, BONKOWSKI M (2002) Interaktionen von
Mikroorganismen, Protozoen und Pflanzen: Wirkungen auf Wurzelarchitektur
und das Wachstum von Weidelgras (Lolium perenne) und Reis (Oryza sativa).
Vortrag, Tagung der DGB, AG Bodenbiologie, München-Neuherberg.
BONKOWSKI M, KREUZER K, SCHEU S (2003) Biodiversity of protozoa in the
rhizosphere of plants: effects on activity, performance and diversity of
microorganisms and on plant growth. Poster, Symposium “Sustainable use and
conservation of biological diversity” des BMBF, Berlin.
KREUZER K, KERN CH, HERDLER S, SCHEU S, BONKOWSKI M (2004) Protozoan
effects on root architecture of rice (Oryza sativa). Poster, Tagung der
Deutschen Gesellschaft für Protozoologie, Innsbruck.
HENKES, G, KREUZER K, SCHEU S, BONKOWSKI M (2004) Combined effects of
protozoa and VA-mycorrhiza on growth of wheat (Triticum aestivum L.).
Poster, Tagung der Deutschen Gesellschaft für Protozoologie, Innsbruck.

Lebenslauf
Persönliche Daten Knut Kreuzer
Geboren am 06.05.1971 in Darmstadt Verheiratet seit 07.06.2002 Ein Sohn (1), eine Tochter (0)
Studium
1991-1994 Grundstudium der Biologie an der TU Darmstadt
1994-2001 Hauptstudium der Biologie an der TU Darmstadt
Fächerkombination der Diplomprüfungen: Botanik, Ökologie, Phytomedizin, Landschaftsarchitektur. Thema der Diplomarbeit: „Einfluss von Lumbriciden, Collembolen und der Verteilung von Streu auf die Konkurrenz zwischen Lolium perenne (Poaceae) und Trifolium repens (Fabaceae)“. Gesamtnote: „sehr gut“
2001-2004 Promotion im Fachbereich Biologie der TU Darmstadt, Institut für Zoologie. Thema: „Einfluss von Bodenamöben (Acanthamoeba castellanii Neff) auf das Wachstum von Pflanzen und die bakterielle Rhizosphärengesellschaft
Tätigkeiten
1999-2000 Studentische Hilfskraft bei der IBACON GmbH, Roßdorf: Entwicklung und Betreuung von Testsystemen für Nutzarthopoden
1994-1999 Studentische Hilfskraft bei der GSI, Darmstadt-Wixhausen: Betreuung sensibler Messeinrichtungen
Betreuungstätigkeiten an der TU Darmstadt
1997 Limnisch-ökologisches Praktikum 2001 Marine-ökologisches Praktikum 2002-2003 Studienarbeit „Einfluss von Protozoen (Acanthamoeba
castellanii) und Mykorrhiza (Glomus intraradices) auf Wurzelanatomie, Wachstum und Nährstoffaufnahme von Reis (Oryza sativa)“
2002-2004 Forschungspraktikum und Diplomarbeit „Der Einfluss von Protozoen (Acanthamoeba castellanii) auf die Wurzelarchitektur und das Wachstum von Tabak (Nicotiana attenuata)“
2002-2004 Forschungspraktikum und Diplomarbeit: „Einfluss von Protozoen und Mykorrhiza auf das Wachstum von Weizen in einem Splitroot-Experiment“

Lebenslauf
98
Grundwehrdienst 1990-1991 Panzergrenadier-Lehrbataillon 353, Hammelburg Schulausbildung 1990 1982-1990
Abitur (Gesamtnote 2,0) Ludwig-Georgs-Gymnasium, Darmstadt
1978-1982 Rehbergschule, Roßdorf

Eidesstattliche Erklärung
Eidesstattliche Erklärung
Hiermit erkläre ich an Eides statt, dass ich die vorliegende Dissertation
selbständig und nur mit den angegebenen Hilfsmitteln angefertigt habe.
Ich habe noch keinen weiteren Promotionsversuch unternommen.

Die vorliegende Arbeit wurde unter Leitung von Prof. Dr. Stefan Scheu am
Institut für Zoologie des Fachbereichs Biologie der Technischen Universität
Darmstadt durchgeführt und von dem Bundesministerium für Bildung und
Forschung (BMBF) im Rahmen des Diversitätprogramms “Biodiversity and
Global Change” (BIOLOG) gefördert.

Danksagung
Danksagung Bei STEFAN SCHEU bedanke ich mich für die Bereitstellung des interessanten
Themas und eines Arbeitsplatzes, der mir den Freiraum zur Umsetzung unserer Ideen gab; für die wissenschaftlich gehaltvollen Diskussionen und den stets vorbildhaften Umgang, den ich als motivierend, sachlich wie herzlich kennen lernen durfte. Dem Ober-Rhizosphärologen MICHAEL BONKOWSKI danke ich für meine
Betreuung, die vielen Anregungen und die enge und intensive Zusammenarbeit trotz seiner limitierten verfügbaren Zeit. Ich bedaure sehr, die gemeinsame und aus meiner Sicht fruchtbare Zusammenarbeit zur Erforschung der Mechanismen in der Rhizosphäre nicht weiterführen zu können. Frau CORNELIA ULLRICH-EBERIUS gilt mein Dank für die interessierte Betreuung
bei den Fragestellungen der Phytohormone, die hierbei intensive Nutzung Ihres Labors und für die engagierte Übernahme des Ko-Referats. Frau FELICITAS PFEIFFER und Herrn HANS-PETER HARRES möchte ich im Voraus
für die Teilnahme an der Prüfungskommission und ihre konstruktive Kritik hierbei danken. Den Neuschwaben MICHAEL HEETHOFF und SVEN MARHAN danke ich herzlichst
für die beste Promotionszeit, die ich – u.a. durch Euch – je hatte; für die intensiven Dialoge wissenschaftlicher und „interdisziplinärer“ Themen sowohl auf dem europäischen Festland als auch auf vorgelagerten Eilanden; und für den gut getimten Exodus gen Schwabenland, um mir für die Diss hinderliche Ablenkungen zu ersparen (aus demselben Grund auch Danke an das SOMMERWETTER 2004 und die DEUTSCHE FUßBALL-NATIONALMANNSCHAFT). Wobei ich bei der AG wäre. Ich habe mich hier sehr wohl gefühlt. Wer auch
immer das sein mag - alle zukünftigen Kolleginnen und Kollegen werden sich an Euch messen lassen müssen. Ich bedanke mich neben dem fachlichen Austausch für die teilweise stoische Bereitwilligkeit, sich auf Feiern an meinen Spielideen zu beteiligen, mich am Ententeich nicht ganz allein mit dem Feuer zu lassen und auch meine ausgeprägte Mittwochs-Neugier zu ertragen. Ich möchte mich darüber hinaus namentlich bei den „Jungs“ - ALEX, JENSI, dem MC, MICHAEL L., OLAF, STEPHAN und SWANTJE - für das ein oder andere nette Gespräch auch mal zwischen zwei Bierchen bedanken, im Speziellen AUNT LILI und MARK für die Hilfe bei der ein oder anderen fachspezifischen Frage und natürlich auch den „Hühnern“ vom Raum 08/15 - KATJA, SILKE und SUSANNE - für das ermöglichte Ausleben meiner Chauvinismen und die Reduzierung frauenspezifischer Gesprächsthemen in meiner Gegenwart. Fachlich bedanke ich mich bei „unseren“ Diplomanden CHRISTIANE, SEBASTIAN und last but not least GUNNAR (welcher nun die Entwicklung von Split-Root-Systemen einer schillernden Karriere als Off-Shore-Speedboat-Piloten vorzieht) für die mehr oder weniger erfolgreiche Zusammenarbeit, um mich gleichzeitig für die fehlende Unterstützung, sei es aus mangelnder fachlicher oder zeitlicher Kompetenz, zu entschuldigen. GABI, DORA und DIANA seien für die stets unterstützende Anwesenheit und dafür gedankt, in unserem Chaos die organisatorische Übersicht behalten zu wollen. „Spasiba” to our russian guys A.U. and A.T. for crucial discussions at any time before sunrise.

Danksagung
CHRISTOPH DICKLER danke ich für die Zusammenarbeit bei den Phytohormon-Versuchen und für das Verständnis meiner durch das enge Zeitbudget bedingte Wechsellaunigkeit und die dadurch belastete Freundschaft. Ich hoffe, dass Deine Promotion ebenfalls ein Happy End finden wird. Ich danke DIETRICH HERTEL (Georg-August-Universität Göttingen) für die
freundliche Einführung und intensive Nutzung von „winrhizo“. Thanks to MORIO IIJIMA (Nagoya University, Japan) for helpful advices according rice roots. JUSTYNA ADAMCYK und MICHAEL WAGNER (vormals TU München, jetzt Uni Wien) danke ich für die Zusammenarbeit bei der Bearbeitung der FISH-Proben. Herzlichen Dank an ANJA MEISTER, dass Sie mich in manch vertrackter
Situation unterstützt und damals den Kontakt zu Stefan hergestellt hat. Bei meinem Freundeskreis (der mich wahrscheinlich weniger vermisst haben
wird als ich ihn) bedanke ich mich für das Verständnis meiner Prioritätensetzung während der angespannten Experimental- und Schreibphase(n). Ich freue mich schon auf das nun bevorstehende Resozialisierungsprogramm. Auch meine Familie hat zuletzt weitgehend auf mich verzichten müssen – ich
danke herzlichst für Ihr Verständnis. Der alles entscheidende Faktor war hierbei die allumfassende Rücksichtnahme und Unterstützung meiner Frau KATHARINA, unseres Sohnes LASSE - und unserer zukünftigen Tochter MERRIT - mich völlig auf die Promotion konzentrieren zu können. Dies ist ein großer Vertrauens- und Liebesbeweis und ich hoffe, Euch meine Dankbarkeit zeigen zu können. IlEggd.