active genes walid mahrez , minerva susana trejo arellano · 24 in eukaryotic cells, ... 74 and has...
TRANSCRIPT

1
H3K36ac is an evolutionary conserved plant histone modification that marks 1
active genes 2
Walid Mahrez12 Minerva Susana Trejo Arellano1 Jordi Moreno-Romero1 Miyuki 3
Nakamura1 Huan Shu2 Paolo Nanni3 Claudia Koumlhler1 Wilhelm Gruissem2 and Lars 4
Hennig1 5
6
1Swedish University of Agricultural Sciences Department of Plant Biology and 7
Linnean Center for Plant Biology PO-Box 7080 SE-75007 Uppsala Sweden 8
2Department of Biology and Zurich-Basel Plant Science Center ETH Zurich CH-8092 9
Zurich Switzerland 10
3Functional Genomics Center Zurich University of ZurichETH Zurich CH-8057 11
Zurich Switzerland 12
13
Author for correspondence 14
Lars Hennig 15
Department of Plant Biology Uppsala BioCenter Swedish University of Agricultural 16
Sciences and Linnean Center for Plant Biology 17
SE-75007 Uppsala Sweden 18
LarsHennigsluse 19
20
Running title H3K36ac is an active histone mark in plants 21
22
Plant Physiology Preview Published on January 13 2016 as DOI101104pp1501744
Copyright 2016 by the American Society of Plant Biologists
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
2
Abstract 23
In eukaryotic cells histones are subject to a large number of post-translational 24
modifications whose sequential or combinatorial action affects chromatin structure and 25
genome function We have identified acetylation at lysine 36 in histone H3 (H3K36ac) 26
as a new chromatin modification in plants The H3K36ac modification is evolutionary 27
conserved in seed plants including the gymnosperm Norway spruce and the 28
angiosperms rice tobacco and Arabidopsis thaliana In Arabidopsis H3K36ac is highly 29
enriched in euchromatin but not in heterochromatin Genome-wide ChIP-seq 30
experiments revealed that H3K36ac peaks at the 5rsquo end of genes mainly on the two 31
nucleosomes immediately distal to the transcription start site independently of gene 32
length H3K36ac overlaps with H3K4me3 and the H2AZ histone variant The histone 33
acetyl transferase GCN5 and the histone deacetylase HDA19 are required for H3K36ac 34
homeostasis H3K36ac and H3K36me3 show negative crosstalk which is mediated by 35
GCN5 and the histone methyl transferase SDG8 Although H3K36ac is associated with 36
gene activity we did not find a linear relationship between H3K36ac and transcript 37
levels suggesting that H3K36ac is a binary indicator of transcription 38
39
40
One sentence summary 41
Profiling of histone H3 modifications by MSMS identified H3K36ac which is 42
introduced at the first 500 bp of active Arabidopsis genes by GCN5 and is conserved in 43
gymnosperms and angiosperms 44
45
Introduction 46
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
3
47
Nuclear DNA of eukaryotes is packed in chromatin the basic unit of which is the 48
nucleosome consisting of 147 bp of DNA wrapped around a protein octamer formed by 49
two copies each of four histones (H2A H2B H3 H4) Chromatin structure and 50
composition are dynamically regulated both temporally and spatially within the nucleus 51
Covalent modifications of histones play an important role in the regulation of gene 52
expression by changing chromatin structure (Strahl et al 2000 Jenuwein et al 2001) 53
Many histone modifications have been identified including acetylation methylation 54
phosphorylation and ubiquitinylation (Cannon 1955 Zhang et al 2007) It has been 55
proposed that histone modifications act in a combinatorial manner to establish specific 56
chromatin states that regulate gene expression (Strahl et al 2000 Jenuwein et al 57
2001) It has been argued however that chromatin-dependent gene activity is only a 58
consequence of the cumulative effect of histone modifications rather than the 59
interpretation of an alphabet (Henikoff 2005) Using high-throughput methods with 60
improved sensitivity specificity and resolution such as mass spectrometry and 61
chromatin immunoprecipitation (ChIP) coupled to sequencing (ChIP-seq) the genomic 62
localization combinatorial patterns and enrichment of histone modifications can be 63
resolved at the level of individual nucleosomes (Liu et al 2005 Kharchenko et al 64
2011) 65
Histone acetylation is a dynamic and reversible modification in which an acetyl 66
group is transferred from acetyl-CoA to histone lysine residues Lysine acetylation 67
neutralizes positive charges of the histone tails thus decreasing their affinity for 68
negatively charged DNA (Hong et al 1993) Acetylated lysines can be recognized by 69
histone code readers such as Bromo domain proteins (Sanchez et al 2014) which then 70
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
4
may promote the recruitment of additional chromatin remodeler or modifier complexes 71
to relax chromatin and facilitate the binding or activation of RNA polymerases 72
(Bannister et al 2011) Histone lysine acetylation is often associated with transcription 73
and has important roles in numerous developmental and biological processes including 74
the regulation of cell cycle flowering time response to environmental conditions and 75
hormone signals (Chen et al 2007 Luo et al 2012) 76
Several amino-terminal lysine residues of histone H3 (K9 K14 K18 K23 and 77
K27) and H4 (K5 K8 K12 K16 and K20) are known to be acetylated in Arabidopsis 78
thaliana (Berr et al 2011) Histone acetylation is dynamically regulated with a usual 79
half-life of 2-3 minutes in human cell lines that rarely exceeds 30-40 minutes (for 80
review see (Iwamoto et al 2015) This rapid turn-over strongly suggests that instead of 81
influencing long-term epigenetic memory acetylation participates in short-term 82
alterations of chromatin states (Barth et al 2010) Homeostasis of histone acetylation is 83
maintained by histone acetyl transferases (HATs) which catalyze the histone 84
acetylation and histone deacetylases (HDACs) which remove the acetyl group (Pandey 85
et al 2002) 86
Histone lysine methylation is another well-studied epigenetic modification with 87
both activating and repressing roles in gene expression Histone lysine residues can be 88
mono- di- or tri-methylated Each distinct methylation state confers different functional 89
outcomes depending on the methylated histone residue the degree of methylation and 90
the chromatin context (Liu et al 2010) Methylated lysines constitute landmarks for 91
binding of chromo MBT Tudor W40 domain or PHD fingers containing proteins 92
which subsequently recruit additional protein complexes resulting in the compaction or 93
relaxation of chromatin structure (Greer et al 2012) 94
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
5
Compared to yeast and animals much less is known about the repertoire of 95
histone modification sites in plants Pioneering work in Arabidopsis includes genome-96
wide chromatin indexing by ChIP (Roudier et al 2011 Sequeira-Mendes et al 2014) 97
and characterization of histone modifications by mass spectrometry (Johnson et al 98
2004 Bergmuumlller et al 2007 Zhang et al 2007) In order to expand this knowledge 99
we implemented high-performance liquid chromatography (HPLC) histone separation 100
in combination with mass spectrometry to identify uncharacterized modifications on the 101
amino-terminal tail of histone H3 in Arabidopsis and its close relative Brassica 102
oleracea (cauliflower) We found H3K36 acetylation (H3K36ac) as a novel histone 103
modification in plants GCN5 a HAT and HDA19 an HDAC contribute to H3K36ac 104
homeostasis H3K36ac is enriched in euchromatic regions and excluded from 105
heterochromatin In addition H3K36ac is generally found at the 5rsquo end of 106
transcriptionally active genes where it overlaps with H3K4me3 and the H2AZ histone 107
variant As H3K36 can also be mono- di- or tri-methylated we tested if the 108
mechanisms of tri-methylation or acetylation of H3K36 are interdependent H3K36ac 109
was increased in a mutant with reduced H3K36me3 and H3K36me3 was increased in 110
gcn5 mutants with reduced H3K36ac Thus acetylation and methylation at H3K36 are 111
in a competitive equilibrium that defines two alternative states 112
113
Results 114
The histone H3 modification profile is conserved in Brassicaceae 115
A histone extract from cauliflower (Brassica oleracea) heads was subjected to reverse-116
phase HPLC and fractions corresponding to H3 based on immunoblot detection were 117
collected and analyzed by liquid chromatography-tandem mass spectrometry (LC-118
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
6
MSMS) using an LTQ-Orbitrap XL instrument For the analysis of long peptides both 119
CID (Collision-Induced Dissociation) and ETD (Electron Transfer Dissociation) 120
fragmentation were performed (Supplemental Fig S1) The high accuracy achieved in 121
the MS measurements allowed to distinguish between tri-methylated and acetylated 122
peptides (Δm = 00364 Da) Data were submitted to the Mascot search engine for 123
identification of proteins and post-translational modifications with a focus on 124
acetylation and methylation This produced a map of H3 acetylation and methylation in 125
cauliflower inflorescences (Supplemental Tab S1) Most of the identified H3 126
modifications and their combinations are consistent with those in Arabidopsis (Johnson 127
et al 2004 Zhang et al 2007) suggesting a high conservation of the histone 128
modifications 129
130
H3K36ac is a new modification in Arabidopsis 131
We identified H3K36ac as a novel H3 modification in plants (Supplemental Tab S1) 132
H3K36ac was found in two independent experiments using different enzymes 133
(Chymotrypsin and ArgC) The confidence for the localization of the modification site 134
was higher than 90 The MSMS spectrum of the ArgC-digested histone H3 shown in 135
Figure 1A details the c and z fragmentation ions for the parent peptide with an observed 136
mass of mz 15048468 The accurate mass of the recorded parent ion was consistent 137
with peptide acetylation (ΔM = 420106 Da) and not tri-methylation 138
(ΔM = 420470 Da) Importantly we detected H3K36 mono- di- and tri-methylation 139
on other H3 peptides confirming that this amino acid can be either methylated or 140
acetylated (Fig 1A Supplemental Fig S2 S3) 141
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
7
To validate and extend the mass spectrometry results a histone extract from 142
Arabidopsis inflorescences was separated by polyacrylamide gel electrophoresis and 143
probed with an anti-H3K36ac antibody The antibody was previously tested for 144
specificity in dot blots and proteinimmuno blots using modified and unmodified 145
peptides and validated for ChIP-seq (Morris et al 2007 Egelhofer et al 2011) The 146
antibody showed no cross-reactivity with eleven different tested acetylated histone 147
peptides (H2AK5ac H3K14ac H3K18ac H3K27ac H3K4ac H3K9ac H4K12ac 148
H4K16ac H4K581216ac H4K5ac H4K8ac) and many unmodified or methylated 149
histone peptides The positive signals for Arabidopsis histone extracts (Fig 2A) 150
confirmed the mass spectrometric identification of the H3K36ac modification in plants 151
Together proteomic and molecular methods support the identification of H3K36ac as a 152
novel histone modification in Brassicacea 153
154
H3K36ac is present in euchromatin 155
To establish the distribution of H3K36ac in the nucleus paraformaldehyde-fixed 156
Arabidopsis root-tip nuclei were immuno-stained using the anti-H3K36ac antibody 157
H3K36ac fluorescence signals were absent from nucleoli and DAPI-dense 158
chromocenters which are mostly comprised of centromeric and pericentromeric repeats 159
In contrast H3K36ac staining was observed in euchromatic regions (Fig 2B-E) 160
indicating that most H3K36ac is associated with less condensed gene-rich regions and 161
possibly active genes To test this prediction formaldehyde-crosslinked ChIP (X-ChIP) 162
was performed using anti-H3K36ac and anti-H3 antibodies Indeed H3K36ac levels 163
were high at the transcribed euchromatic ACTIN7 gene but low at the silent 164
heterochromatic Ta2 transposon (Fig 2F) The increased H3 signal at Ta2 is consistent 165
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

2
Abstract 23
In eukaryotic cells histones are subject to a large number of post-translational 24
modifications whose sequential or combinatorial action affects chromatin structure and 25
genome function We have identified acetylation at lysine 36 in histone H3 (H3K36ac) 26
as a new chromatin modification in plants The H3K36ac modification is evolutionary 27
conserved in seed plants including the gymnosperm Norway spruce and the 28
angiosperms rice tobacco and Arabidopsis thaliana In Arabidopsis H3K36ac is highly 29
enriched in euchromatin but not in heterochromatin Genome-wide ChIP-seq 30
experiments revealed that H3K36ac peaks at the 5rsquo end of genes mainly on the two 31
nucleosomes immediately distal to the transcription start site independently of gene 32
length H3K36ac overlaps with H3K4me3 and the H2AZ histone variant The histone 33
acetyl transferase GCN5 and the histone deacetylase HDA19 are required for H3K36ac 34
homeostasis H3K36ac and H3K36me3 show negative crosstalk which is mediated by 35
GCN5 and the histone methyl transferase SDG8 Although H3K36ac is associated with 36
gene activity we did not find a linear relationship between H3K36ac and transcript 37
levels suggesting that H3K36ac is a binary indicator of transcription 38
39
40
One sentence summary 41
Profiling of histone H3 modifications by MSMS identified H3K36ac which is 42
introduced at the first 500 bp of active Arabidopsis genes by GCN5 and is conserved in 43
gymnosperms and angiosperms 44
45
Introduction 46
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
3
47
Nuclear DNA of eukaryotes is packed in chromatin the basic unit of which is the 48
nucleosome consisting of 147 bp of DNA wrapped around a protein octamer formed by 49
two copies each of four histones (H2A H2B H3 H4) Chromatin structure and 50
composition are dynamically regulated both temporally and spatially within the nucleus 51
Covalent modifications of histones play an important role in the regulation of gene 52
expression by changing chromatin structure (Strahl et al 2000 Jenuwein et al 2001) 53
Many histone modifications have been identified including acetylation methylation 54
phosphorylation and ubiquitinylation (Cannon 1955 Zhang et al 2007) It has been 55
proposed that histone modifications act in a combinatorial manner to establish specific 56
chromatin states that regulate gene expression (Strahl et al 2000 Jenuwein et al 57
2001) It has been argued however that chromatin-dependent gene activity is only a 58
consequence of the cumulative effect of histone modifications rather than the 59
interpretation of an alphabet (Henikoff 2005) Using high-throughput methods with 60
improved sensitivity specificity and resolution such as mass spectrometry and 61
chromatin immunoprecipitation (ChIP) coupled to sequencing (ChIP-seq) the genomic 62
localization combinatorial patterns and enrichment of histone modifications can be 63
resolved at the level of individual nucleosomes (Liu et al 2005 Kharchenko et al 64
2011) 65
Histone acetylation is a dynamic and reversible modification in which an acetyl 66
group is transferred from acetyl-CoA to histone lysine residues Lysine acetylation 67
neutralizes positive charges of the histone tails thus decreasing their affinity for 68
negatively charged DNA (Hong et al 1993) Acetylated lysines can be recognized by 69
histone code readers such as Bromo domain proteins (Sanchez et al 2014) which then 70
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
4
may promote the recruitment of additional chromatin remodeler or modifier complexes 71
to relax chromatin and facilitate the binding or activation of RNA polymerases 72
(Bannister et al 2011) Histone lysine acetylation is often associated with transcription 73
and has important roles in numerous developmental and biological processes including 74
the regulation of cell cycle flowering time response to environmental conditions and 75
hormone signals (Chen et al 2007 Luo et al 2012) 76
Several amino-terminal lysine residues of histone H3 (K9 K14 K18 K23 and 77
K27) and H4 (K5 K8 K12 K16 and K20) are known to be acetylated in Arabidopsis 78
thaliana (Berr et al 2011) Histone acetylation is dynamically regulated with a usual 79
half-life of 2-3 minutes in human cell lines that rarely exceeds 30-40 minutes (for 80
review see (Iwamoto et al 2015) This rapid turn-over strongly suggests that instead of 81
influencing long-term epigenetic memory acetylation participates in short-term 82
alterations of chromatin states (Barth et al 2010) Homeostasis of histone acetylation is 83
maintained by histone acetyl transferases (HATs) which catalyze the histone 84
acetylation and histone deacetylases (HDACs) which remove the acetyl group (Pandey 85
et al 2002) 86
Histone lysine methylation is another well-studied epigenetic modification with 87
both activating and repressing roles in gene expression Histone lysine residues can be 88
mono- di- or tri-methylated Each distinct methylation state confers different functional 89
outcomes depending on the methylated histone residue the degree of methylation and 90
the chromatin context (Liu et al 2010) Methylated lysines constitute landmarks for 91
binding of chromo MBT Tudor W40 domain or PHD fingers containing proteins 92
which subsequently recruit additional protein complexes resulting in the compaction or 93
relaxation of chromatin structure (Greer et al 2012) 94
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
5
Compared to yeast and animals much less is known about the repertoire of 95
histone modification sites in plants Pioneering work in Arabidopsis includes genome-96
wide chromatin indexing by ChIP (Roudier et al 2011 Sequeira-Mendes et al 2014) 97
and characterization of histone modifications by mass spectrometry (Johnson et al 98
2004 Bergmuumlller et al 2007 Zhang et al 2007) In order to expand this knowledge 99
we implemented high-performance liquid chromatography (HPLC) histone separation 100
in combination with mass spectrometry to identify uncharacterized modifications on the 101
amino-terminal tail of histone H3 in Arabidopsis and its close relative Brassica 102
oleracea (cauliflower) We found H3K36 acetylation (H3K36ac) as a novel histone 103
modification in plants GCN5 a HAT and HDA19 an HDAC contribute to H3K36ac 104
homeostasis H3K36ac is enriched in euchromatic regions and excluded from 105
heterochromatin In addition H3K36ac is generally found at the 5rsquo end of 106
transcriptionally active genes where it overlaps with H3K4me3 and the H2AZ histone 107
variant As H3K36 can also be mono- di- or tri-methylated we tested if the 108
mechanisms of tri-methylation or acetylation of H3K36 are interdependent H3K36ac 109
was increased in a mutant with reduced H3K36me3 and H3K36me3 was increased in 110
gcn5 mutants with reduced H3K36ac Thus acetylation and methylation at H3K36 are 111
in a competitive equilibrium that defines two alternative states 112
113
Results 114
The histone H3 modification profile is conserved in Brassicaceae 115
A histone extract from cauliflower (Brassica oleracea) heads was subjected to reverse-116
phase HPLC and fractions corresponding to H3 based on immunoblot detection were 117
collected and analyzed by liquid chromatography-tandem mass spectrometry (LC-118
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
6
MSMS) using an LTQ-Orbitrap XL instrument For the analysis of long peptides both 119
CID (Collision-Induced Dissociation) and ETD (Electron Transfer Dissociation) 120
fragmentation were performed (Supplemental Fig S1) The high accuracy achieved in 121
the MS measurements allowed to distinguish between tri-methylated and acetylated 122
peptides (Δm = 00364 Da) Data were submitted to the Mascot search engine for 123
identification of proteins and post-translational modifications with a focus on 124
acetylation and methylation This produced a map of H3 acetylation and methylation in 125
cauliflower inflorescences (Supplemental Tab S1) Most of the identified H3 126
modifications and their combinations are consistent with those in Arabidopsis (Johnson 127
et al 2004 Zhang et al 2007) suggesting a high conservation of the histone 128
modifications 129
130
H3K36ac is a new modification in Arabidopsis 131
We identified H3K36ac as a novel H3 modification in plants (Supplemental Tab S1) 132
H3K36ac was found in two independent experiments using different enzymes 133
(Chymotrypsin and ArgC) The confidence for the localization of the modification site 134
was higher than 90 The MSMS spectrum of the ArgC-digested histone H3 shown in 135
Figure 1A details the c and z fragmentation ions for the parent peptide with an observed 136
mass of mz 15048468 The accurate mass of the recorded parent ion was consistent 137
with peptide acetylation (ΔM = 420106 Da) and not tri-methylation 138
(ΔM = 420470 Da) Importantly we detected H3K36 mono- di- and tri-methylation 139
on other H3 peptides confirming that this amino acid can be either methylated or 140
acetylated (Fig 1A Supplemental Fig S2 S3) 141
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
7
To validate and extend the mass spectrometry results a histone extract from 142
Arabidopsis inflorescences was separated by polyacrylamide gel electrophoresis and 143
probed with an anti-H3K36ac antibody The antibody was previously tested for 144
specificity in dot blots and proteinimmuno blots using modified and unmodified 145
peptides and validated for ChIP-seq (Morris et al 2007 Egelhofer et al 2011) The 146
antibody showed no cross-reactivity with eleven different tested acetylated histone 147
peptides (H2AK5ac H3K14ac H3K18ac H3K27ac H3K4ac H3K9ac H4K12ac 148
H4K16ac H4K581216ac H4K5ac H4K8ac) and many unmodified or methylated 149
histone peptides The positive signals for Arabidopsis histone extracts (Fig 2A) 150
confirmed the mass spectrometric identification of the H3K36ac modification in plants 151
Together proteomic and molecular methods support the identification of H3K36ac as a 152
novel histone modification in Brassicacea 153
154
H3K36ac is present in euchromatin 155
To establish the distribution of H3K36ac in the nucleus paraformaldehyde-fixed 156
Arabidopsis root-tip nuclei were immuno-stained using the anti-H3K36ac antibody 157
H3K36ac fluorescence signals were absent from nucleoli and DAPI-dense 158
chromocenters which are mostly comprised of centromeric and pericentromeric repeats 159
In contrast H3K36ac staining was observed in euchromatic regions (Fig 2B-E) 160
indicating that most H3K36ac is associated with less condensed gene-rich regions and 161
possibly active genes To test this prediction formaldehyde-crosslinked ChIP (X-ChIP) 162
was performed using anti-H3K36ac and anti-H3 antibodies Indeed H3K36ac levels 163
were high at the transcribed euchromatic ACTIN7 gene but low at the silent 164
heterochromatic Ta2 transposon (Fig 2F) The increased H3 signal at Ta2 is consistent 165
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

3
47
Nuclear DNA of eukaryotes is packed in chromatin the basic unit of which is the 48
nucleosome consisting of 147 bp of DNA wrapped around a protein octamer formed by 49
two copies each of four histones (H2A H2B H3 H4) Chromatin structure and 50
composition are dynamically regulated both temporally and spatially within the nucleus 51
Covalent modifications of histones play an important role in the regulation of gene 52
expression by changing chromatin structure (Strahl et al 2000 Jenuwein et al 2001) 53
Many histone modifications have been identified including acetylation methylation 54
phosphorylation and ubiquitinylation (Cannon 1955 Zhang et al 2007) It has been 55
proposed that histone modifications act in a combinatorial manner to establish specific 56
chromatin states that regulate gene expression (Strahl et al 2000 Jenuwein et al 57
2001) It has been argued however that chromatin-dependent gene activity is only a 58
consequence of the cumulative effect of histone modifications rather than the 59
interpretation of an alphabet (Henikoff 2005) Using high-throughput methods with 60
improved sensitivity specificity and resolution such as mass spectrometry and 61
chromatin immunoprecipitation (ChIP) coupled to sequencing (ChIP-seq) the genomic 62
localization combinatorial patterns and enrichment of histone modifications can be 63
resolved at the level of individual nucleosomes (Liu et al 2005 Kharchenko et al 64
2011) 65
Histone acetylation is a dynamic and reversible modification in which an acetyl 66
group is transferred from acetyl-CoA to histone lysine residues Lysine acetylation 67
neutralizes positive charges of the histone tails thus decreasing their affinity for 68
negatively charged DNA (Hong et al 1993) Acetylated lysines can be recognized by 69
histone code readers such as Bromo domain proteins (Sanchez et al 2014) which then 70
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
4
may promote the recruitment of additional chromatin remodeler or modifier complexes 71
to relax chromatin and facilitate the binding or activation of RNA polymerases 72
(Bannister et al 2011) Histone lysine acetylation is often associated with transcription 73
and has important roles in numerous developmental and biological processes including 74
the regulation of cell cycle flowering time response to environmental conditions and 75
hormone signals (Chen et al 2007 Luo et al 2012) 76
Several amino-terminal lysine residues of histone H3 (K9 K14 K18 K23 and 77
K27) and H4 (K5 K8 K12 K16 and K20) are known to be acetylated in Arabidopsis 78
thaliana (Berr et al 2011) Histone acetylation is dynamically regulated with a usual 79
half-life of 2-3 minutes in human cell lines that rarely exceeds 30-40 minutes (for 80
review see (Iwamoto et al 2015) This rapid turn-over strongly suggests that instead of 81
influencing long-term epigenetic memory acetylation participates in short-term 82
alterations of chromatin states (Barth et al 2010) Homeostasis of histone acetylation is 83
maintained by histone acetyl transferases (HATs) which catalyze the histone 84
acetylation and histone deacetylases (HDACs) which remove the acetyl group (Pandey 85
et al 2002) 86
Histone lysine methylation is another well-studied epigenetic modification with 87
both activating and repressing roles in gene expression Histone lysine residues can be 88
mono- di- or tri-methylated Each distinct methylation state confers different functional 89
outcomes depending on the methylated histone residue the degree of methylation and 90
the chromatin context (Liu et al 2010) Methylated lysines constitute landmarks for 91
binding of chromo MBT Tudor W40 domain or PHD fingers containing proteins 92
which subsequently recruit additional protein complexes resulting in the compaction or 93
relaxation of chromatin structure (Greer et al 2012) 94
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
5
Compared to yeast and animals much less is known about the repertoire of 95
histone modification sites in plants Pioneering work in Arabidopsis includes genome-96
wide chromatin indexing by ChIP (Roudier et al 2011 Sequeira-Mendes et al 2014) 97
and characterization of histone modifications by mass spectrometry (Johnson et al 98
2004 Bergmuumlller et al 2007 Zhang et al 2007) In order to expand this knowledge 99
we implemented high-performance liquid chromatography (HPLC) histone separation 100
in combination with mass spectrometry to identify uncharacterized modifications on the 101
amino-terminal tail of histone H3 in Arabidopsis and its close relative Brassica 102
oleracea (cauliflower) We found H3K36 acetylation (H3K36ac) as a novel histone 103
modification in plants GCN5 a HAT and HDA19 an HDAC contribute to H3K36ac 104
homeostasis H3K36ac is enriched in euchromatic regions and excluded from 105
heterochromatin In addition H3K36ac is generally found at the 5rsquo end of 106
transcriptionally active genes where it overlaps with H3K4me3 and the H2AZ histone 107
variant As H3K36 can also be mono- di- or tri-methylated we tested if the 108
mechanisms of tri-methylation or acetylation of H3K36 are interdependent H3K36ac 109
was increased in a mutant with reduced H3K36me3 and H3K36me3 was increased in 110
gcn5 mutants with reduced H3K36ac Thus acetylation and methylation at H3K36 are 111
in a competitive equilibrium that defines two alternative states 112
113
Results 114
The histone H3 modification profile is conserved in Brassicaceae 115
A histone extract from cauliflower (Brassica oleracea) heads was subjected to reverse-116
phase HPLC and fractions corresponding to H3 based on immunoblot detection were 117
collected and analyzed by liquid chromatography-tandem mass spectrometry (LC-118
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
6
MSMS) using an LTQ-Orbitrap XL instrument For the analysis of long peptides both 119
CID (Collision-Induced Dissociation) and ETD (Electron Transfer Dissociation) 120
fragmentation were performed (Supplemental Fig S1) The high accuracy achieved in 121
the MS measurements allowed to distinguish between tri-methylated and acetylated 122
peptides (Δm = 00364 Da) Data were submitted to the Mascot search engine for 123
identification of proteins and post-translational modifications with a focus on 124
acetylation and methylation This produced a map of H3 acetylation and methylation in 125
cauliflower inflorescences (Supplemental Tab S1) Most of the identified H3 126
modifications and their combinations are consistent with those in Arabidopsis (Johnson 127
et al 2004 Zhang et al 2007) suggesting a high conservation of the histone 128
modifications 129
130
H3K36ac is a new modification in Arabidopsis 131
We identified H3K36ac as a novel H3 modification in plants (Supplemental Tab S1) 132
H3K36ac was found in two independent experiments using different enzymes 133
(Chymotrypsin and ArgC) The confidence for the localization of the modification site 134
was higher than 90 The MSMS spectrum of the ArgC-digested histone H3 shown in 135
Figure 1A details the c and z fragmentation ions for the parent peptide with an observed 136
mass of mz 15048468 The accurate mass of the recorded parent ion was consistent 137
with peptide acetylation (ΔM = 420106 Da) and not tri-methylation 138
(ΔM = 420470 Da) Importantly we detected H3K36 mono- di- and tri-methylation 139
on other H3 peptides confirming that this amino acid can be either methylated or 140
acetylated (Fig 1A Supplemental Fig S2 S3) 141
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
7
To validate and extend the mass spectrometry results a histone extract from 142
Arabidopsis inflorescences was separated by polyacrylamide gel electrophoresis and 143
probed with an anti-H3K36ac antibody The antibody was previously tested for 144
specificity in dot blots and proteinimmuno blots using modified and unmodified 145
peptides and validated for ChIP-seq (Morris et al 2007 Egelhofer et al 2011) The 146
antibody showed no cross-reactivity with eleven different tested acetylated histone 147
peptides (H2AK5ac H3K14ac H3K18ac H3K27ac H3K4ac H3K9ac H4K12ac 148
H4K16ac H4K581216ac H4K5ac H4K8ac) and many unmodified or methylated 149
histone peptides The positive signals for Arabidopsis histone extracts (Fig 2A) 150
confirmed the mass spectrometric identification of the H3K36ac modification in plants 151
Together proteomic and molecular methods support the identification of H3K36ac as a 152
novel histone modification in Brassicacea 153
154
H3K36ac is present in euchromatin 155
To establish the distribution of H3K36ac in the nucleus paraformaldehyde-fixed 156
Arabidopsis root-tip nuclei were immuno-stained using the anti-H3K36ac antibody 157
H3K36ac fluorescence signals were absent from nucleoli and DAPI-dense 158
chromocenters which are mostly comprised of centromeric and pericentromeric repeats 159
In contrast H3K36ac staining was observed in euchromatic regions (Fig 2B-E) 160
indicating that most H3K36ac is associated with less condensed gene-rich regions and 161
possibly active genes To test this prediction formaldehyde-crosslinked ChIP (X-ChIP) 162
was performed using anti-H3K36ac and anti-H3 antibodies Indeed H3K36ac levels 163
were high at the transcribed euchromatic ACTIN7 gene but low at the silent 164
heterochromatic Ta2 transposon (Fig 2F) The increased H3 signal at Ta2 is consistent 165
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

4
may promote the recruitment of additional chromatin remodeler or modifier complexes 71
to relax chromatin and facilitate the binding or activation of RNA polymerases 72
(Bannister et al 2011) Histone lysine acetylation is often associated with transcription 73
and has important roles in numerous developmental and biological processes including 74
the regulation of cell cycle flowering time response to environmental conditions and 75
hormone signals (Chen et al 2007 Luo et al 2012) 76
Several amino-terminal lysine residues of histone H3 (K9 K14 K18 K23 and 77
K27) and H4 (K5 K8 K12 K16 and K20) are known to be acetylated in Arabidopsis 78
thaliana (Berr et al 2011) Histone acetylation is dynamically regulated with a usual 79
half-life of 2-3 minutes in human cell lines that rarely exceeds 30-40 minutes (for 80
review see (Iwamoto et al 2015) This rapid turn-over strongly suggests that instead of 81
influencing long-term epigenetic memory acetylation participates in short-term 82
alterations of chromatin states (Barth et al 2010) Homeostasis of histone acetylation is 83
maintained by histone acetyl transferases (HATs) which catalyze the histone 84
acetylation and histone deacetylases (HDACs) which remove the acetyl group (Pandey 85
et al 2002) 86
Histone lysine methylation is another well-studied epigenetic modification with 87
both activating and repressing roles in gene expression Histone lysine residues can be 88
mono- di- or tri-methylated Each distinct methylation state confers different functional 89
outcomes depending on the methylated histone residue the degree of methylation and 90
the chromatin context (Liu et al 2010) Methylated lysines constitute landmarks for 91
binding of chromo MBT Tudor W40 domain or PHD fingers containing proteins 92
which subsequently recruit additional protein complexes resulting in the compaction or 93
relaxation of chromatin structure (Greer et al 2012) 94
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
5
Compared to yeast and animals much less is known about the repertoire of 95
histone modification sites in plants Pioneering work in Arabidopsis includes genome-96
wide chromatin indexing by ChIP (Roudier et al 2011 Sequeira-Mendes et al 2014) 97
and characterization of histone modifications by mass spectrometry (Johnson et al 98
2004 Bergmuumlller et al 2007 Zhang et al 2007) In order to expand this knowledge 99
we implemented high-performance liquid chromatography (HPLC) histone separation 100
in combination with mass spectrometry to identify uncharacterized modifications on the 101
amino-terminal tail of histone H3 in Arabidopsis and its close relative Brassica 102
oleracea (cauliflower) We found H3K36 acetylation (H3K36ac) as a novel histone 103
modification in plants GCN5 a HAT and HDA19 an HDAC contribute to H3K36ac 104
homeostasis H3K36ac is enriched in euchromatic regions and excluded from 105
heterochromatin In addition H3K36ac is generally found at the 5rsquo end of 106
transcriptionally active genes where it overlaps with H3K4me3 and the H2AZ histone 107
variant As H3K36 can also be mono- di- or tri-methylated we tested if the 108
mechanisms of tri-methylation or acetylation of H3K36 are interdependent H3K36ac 109
was increased in a mutant with reduced H3K36me3 and H3K36me3 was increased in 110
gcn5 mutants with reduced H3K36ac Thus acetylation and methylation at H3K36 are 111
in a competitive equilibrium that defines two alternative states 112
113
Results 114
The histone H3 modification profile is conserved in Brassicaceae 115
A histone extract from cauliflower (Brassica oleracea) heads was subjected to reverse-116
phase HPLC and fractions corresponding to H3 based on immunoblot detection were 117
collected and analyzed by liquid chromatography-tandem mass spectrometry (LC-118
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
6
MSMS) using an LTQ-Orbitrap XL instrument For the analysis of long peptides both 119
CID (Collision-Induced Dissociation) and ETD (Electron Transfer Dissociation) 120
fragmentation were performed (Supplemental Fig S1) The high accuracy achieved in 121
the MS measurements allowed to distinguish between tri-methylated and acetylated 122
peptides (Δm = 00364 Da) Data were submitted to the Mascot search engine for 123
identification of proteins and post-translational modifications with a focus on 124
acetylation and methylation This produced a map of H3 acetylation and methylation in 125
cauliflower inflorescences (Supplemental Tab S1) Most of the identified H3 126
modifications and their combinations are consistent with those in Arabidopsis (Johnson 127
et al 2004 Zhang et al 2007) suggesting a high conservation of the histone 128
modifications 129
130
H3K36ac is a new modification in Arabidopsis 131
We identified H3K36ac as a novel H3 modification in plants (Supplemental Tab S1) 132
H3K36ac was found in two independent experiments using different enzymes 133
(Chymotrypsin and ArgC) The confidence for the localization of the modification site 134
was higher than 90 The MSMS spectrum of the ArgC-digested histone H3 shown in 135
Figure 1A details the c and z fragmentation ions for the parent peptide with an observed 136
mass of mz 15048468 The accurate mass of the recorded parent ion was consistent 137
with peptide acetylation (ΔM = 420106 Da) and not tri-methylation 138
(ΔM = 420470 Da) Importantly we detected H3K36 mono- di- and tri-methylation 139
on other H3 peptides confirming that this amino acid can be either methylated or 140
acetylated (Fig 1A Supplemental Fig S2 S3) 141
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
7
To validate and extend the mass spectrometry results a histone extract from 142
Arabidopsis inflorescences was separated by polyacrylamide gel electrophoresis and 143
probed with an anti-H3K36ac antibody The antibody was previously tested for 144
specificity in dot blots and proteinimmuno blots using modified and unmodified 145
peptides and validated for ChIP-seq (Morris et al 2007 Egelhofer et al 2011) The 146
antibody showed no cross-reactivity with eleven different tested acetylated histone 147
peptides (H2AK5ac H3K14ac H3K18ac H3K27ac H3K4ac H3K9ac H4K12ac 148
H4K16ac H4K581216ac H4K5ac H4K8ac) and many unmodified or methylated 149
histone peptides The positive signals for Arabidopsis histone extracts (Fig 2A) 150
confirmed the mass spectrometric identification of the H3K36ac modification in plants 151
Together proteomic and molecular methods support the identification of H3K36ac as a 152
novel histone modification in Brassicacea 153
154
H3K36ac is present in euchromatin 155
To establish the distribution of H3K36ac in the nucleus paraformaldehyde-fixed 156
Arabidopsis root-tip nuclei were immuno-stained using the anti-H3K36ac antibody 157
H3K36ac fluorescence signals were absent from nucleoli and DAPI-dense 158
chromocenters which are mostly comprised of centromeric and pericentromeric repeats 159
In contrast H3K36ac staining was observed in euchromatic regions (Fig 2B-E) 160
indicating that most H3K36ac is associated with less condensed gene-rich regions and 161
possibly active genes To test this prediction formaldehyde-crosslinked ChIP (X-ChIP) 162
was performed using anti-H3K36ac and anti-H3 antibodies Indeed H3K36ac levels 163
were high at the transcribed euchromatic ACTIN7 gene but low at the silent 164
heterochromatic Ta2 transposon (Fig 2F) The increased H3 signal at Ta2 is consistent 165
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

5
Compared to yeast and animals much less is known about the repertoire of 95
histone modification sites in plants Pioneering work in Arabidopsis includes genome-96
wide chromatin indexing by ChIP (Roudier et al 2011 Sequeira-Mendes et al 2014) 97
and characterization of histone modifications by mass spectrometry (Johnson et al 98
2004 Bergmuumlller et al 2007 Zhang et al 2007) In order to expand this knowledge 99
we implemented high-performance liquid chromatography (HPLC) histone separation 100
in combination with mass spectrometry to identify uncharacterized modifications on the 101
amino-terminal tail of histone H3 in Arabidopsis and its close relative Brassica 102
oleracea (cauliflower) We found H3K36 acetylation (H3K36ac) as a novel histone 103
modification in plants GCN5 a HAT and HDA19 an HDAC contribute to H3K36ac 104
homeostasis H3K36ac is enriched in euchromatic regions and excluded from 105
heterochromatin In addition H3K36ac is generally found at the 5rsquo end of 106
transcriptionally active genes where it overlaps with H3K4me3 and the H2AZ histone 107
variant As H3K36 can also be mono- di- or tri-methylated we tested if the 108
mechanisms of tri-methylation or acetylation of H3K36 are interdependent H3K36ac 109
was increased in a mutant with reduced H3K36me3 and H3K36me3 was increased in 110
gcn5 mutants with reduced H3K36ac Thus acetylation and methylation at H3K36 are 111
in a competitive equilibrium that defines two alternative states 112
113
Results 114
The histone H3 modification profile is conserved in Brassicaceae 115
A histone extract from cauliflower (Brassica oleracea) heads was subjected to reverse-116
phase HPLC and fractions corresponding to H3 based on immunoblot detection were 117
collected and analyzed by liquid chromatography-tandem mass spectrometry (LC-118
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
6
MSMS) using an LTQ-Orbitrap XL instrument For the analysis of long peptides both 119
CID (Collision-Induced Dissociation) and ETD (Electron Transfer Dissociation) 120
fragmentation were performed (Supplemental Fig S1) The high accuracy achieved in 121
the MS measurements allowed to distinguish between tri-methylated and acetylated 122
peptides (Δm = 00364 Da) Data were submitted to the Mascot search engine for 123
identification of proteins and post-translational modifications with a focus on 124
acetylation and methylation This produced a map of H3 acetylation and methylation in 125
cauliflower inflorescences (Supplemental Tab S1) Most of the identified H3 126
modifications and their combinations are consistent with those in Arabidopsis (Johnson 127
et al 2004 Zhang et al 2007) suggesting a high conservation of the histone 128
modifications 129
130
H3K36ac is a new modification in Arabidopsis 131
We identified H3K36ac as a novel H3 modification in plants (Supplemental Tab S1) 132
H3K36ac was found in two independent experiments using different enzymes 133
(Chymotrypsin and ArgC) The confidence for the localization of the modification site 134
was higher than 90 The MSMS spectrum of the ArgC-digested histone H3 shown in 135
Figure 1A details the c and z fragmentation ions for the parent peptide with an observed 136
mass of mz 15048468 The accurate mass of the recorded parent ion was consistent 137
with peptide acetylation (ΔM = 420106 Da) and not tri-methylation 138
(ΔM = 420470 Da) Importantly we detected H3K36 mono- di- and tri-methylation 139
on other H3 peptides confirming that this amino acid can be either methylated or 140
acetylated (Fig 1A Supplemental Fig S2 S3) 141
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
7
To validate and extend the mass spectrometry results a histone extract from 142
Arabidopsis inflorescences was separated by polyacrylamide gel electrophoresis and 143
probed with an anti-H3K36ac antibody The antibody was previously tested for 144
specificity in dot blots and proteinimmuno blots using modified and unmodified 145
peptides and validated for ChIP-seq (Morris et al 2007 Egelhofer et al 2011) The 146
antibody showed no cross-reactivity with eleven different tested acetylated histone 147
peptides (H2AK5ac H3K14ac H3K18ac H3K27ac H3K4ac H3K9ac H4K12ac 148
H4K16ac H4K581216ac H4K5ac H4K8ac) and many unmodified or methylated 149
histone peptides The positive signals for Arabidopsis histone extracts (Fig 2A) 150
confirmed the mass spectrometric identification of the H3K36ac modification in plants 151
Together proteomic and molecular methods support the identification of H3K36ac as a 152
novel histone modification in Brassicacea 153
154
H3K36ac is present in euchromatin 155
To establish the distribution of H3K36ac in the nucleus paraformaldehyde-fixed 156
Arabidopsis root-tip nuclei were immuno-stained using the anti-H3K36ac antibody 157
H3K36ac fluorescence signals were absent from nucleoli and DAPI-dense 158
chromocenters which are mostly comprised of centromeric and pericentromeric repeats 159
In contrast H3K36ac staining was observed in euchromatic regions (Fig 2B-E) 160
indicating that most H3K36ac is associated with less condensed gene-rich regions and 161
possibly active genes To test this prediction formaldehyde-crosslinked ChIP (X-ChIP) 162
was performed using anti-H3K36ac and anti-H3 antibodies Indeed H3K36ac levels 163
were high at the transcribed euchromatic ACTIN7 gene but low at the silent 164
heterochromatic Ta2 transposon (Fig 2F) The increased H3 signal at Ta2 is consistent 165
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

6
MSMS) using an LTQ-Orbitrap XL instrument For the analysis of long peptides both 119
CID (Collision-Induced Dissociation) and ETD (Electron Transfer Dissociation) 120
fragmentation were performed (Supplemental Fig S1) The high accuracy achieved in 121
the MS measurements allowed to distinguish between tri-methylated and acetylated 122
peptides (Δm = 00364 Da) Data were submitted to the Mascot search engine for 123
identification of proteins and post-translational modifications with a focus on 124
acetylation and methylation This produced a map of H3 acetylation and methylation in 125
cauliflower inflorescences (Supplemental Tab S1) Most of the identified H3 126
modifications and their combinations are consistent with those in Arabidopsis (Johnson 127
et al 2004 Zhang et al 2007) suggesting a high conservation of the histone 128
modifications 129
130
H3K36ac is a new modification in Arabidopsis 131
We identified H3K36ac as a novel H3 modification in plants (Supplemental Tab S1) 132
H3K36ac was found in two independent experiments using different enzymes 133
(Chymotrypsin and ArgC) The confidence for the localization of the modification site 134
was higher than 90 The MSMS spectrum of the ArgC-digested histone H3 shown in 135
Figure 1A details the c and z fragmentation ions for the parent peptide with an observed 136
mass of mz 15048468 The accurate mass of the recorded parent ion was consistent 137
with peptide acetylation (ΔM = 420106 Da) and not tri-methylation 138
(ΔM = 420470 Da) Importantly we detected H3K36 mono- di- and tri-methylation 139
on other H3 peptides confirming that this amino acid can be either methylated or 140
acetylated (Fig 1A Supplemental Fig S2 S3) 141
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
7
To validate and extend the mass spectrometry results a histone extract from 142
Arabidopsis inflorescences was separated by polyacrylamide gel electrophoresis and 143
probed with an anti-H3K36ac antibody The antibody was previously tested for 144
specificity in dot blots and proteinimmuno blots using modified and unmodified 145
peptides and validated for ChIP-seq (Morris et al 2007 Egelhofer et al 2011) The 146
antibody showed no cross-reactivity with eleven different tested acetylated histone 147
peptides (H2AK5ac H3K14ac H3K18ac H3K27ac H3K4ac H3K9ac H4K12ac 148
H4K16ac H4K581216ac H4K5ac H4K8ac) and many unmodified or methylated 149
histone peptides The positive signals for Arabidopsis histone extracts (Fig 2A) 150
confirmed the mass spectrometric identification of the H3K36ac modification in plants 151
Together proteomic and molecular methods support the identification of H3K36ac as a 152
novel histone modification in Brassicacea 153
154
H3K36ac is present in euchromatin 155
To establish the distribution of H3K36ac in the nucleus paraformaldehyde-fixed 156
Arabidopsis root-tip nuclei were immuno-stained using the anti-H3K36ac antibody 157
H3K36ac fluorescence signals were absent from nucleoli and DAPI-dense 158
chromocenters which are mostly comprised of centromeric and pericentromeric repeats 159
In contrast H3K36ac staining was observed in euchromatic regions (Fig 2B-E) 160
indicating that most H3K36ac is associated with less condensed gene-rich regions and 161
possibly active genes To test this prediction formaldehyde-crosslinked ChIP (X-ChIP) 162
was performed using anti-H3K36ac and anti-H3 antibodies Indeed H3K36ac levels 163
were high at the transcribed euchromatic ACTIN7 gene but low at the silent 164
heterochromatic Ta2 transposon (Fig 2F) The increased H3 signal at Ta2 is consistent 165
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

7
To validate and extend the mass spectrometry results a histone extract from 142
Arabidopsis inflorescences was separated by polyacrylamide gel electrophoresis and 143
probed with an anti-H3K36ac antibody The antibody was previously tested for 144
specificity in dot blots and proteinimmuno blots using modified and unmodified 145
peptides and validated for ChIP-seq (Morris et al 2007 Egelhofer et al 2011) The 146
antibody showed no cross-reactivity with eleven different tested acetylated histone 147
peptides (H2AK5ac H3K14ac H3K18ac H3K27ac H3K4ac H3K9ac H4K12ac 148
H4K16ac H4K581216ac H4K5ac H4K8ac) and many unmodified or methylated 149
histone peptides The positive signals for Arabidopsis histone extracts (Fig 2A) 150
confirmed the mass spectrometric identification of the H3K36ac modification in plants 151
Together proteomic and molecular methods support the identification of H3K36ac as a 152
novel histone modification in Brassicacea 153
154
H3K36ac is present in euchromatin 155
To establish the distribution of H3K36ac in the nucleus paraformaldehyde-fixed 156
Arabidopsis root-tip nuclei were immuno-stained using the anti-H3K36ac antibody 157
H3K36ac fluorescence signals were absent from nucleoli and DAPI-dense 158
chromocenters which are mostly comprised of centromeric and pericentromeric repeats 159
In contrast H3K36ac staining was observed in euchromatic regions (Fig 2B-E) 160
indicating that most H3K36ac is associated with less condensed gene-rich regions and 161
possibly active genes To test this prediction formaldehyde-crosslinked ChIP (X-ChIP) 162
was performed using anti-H3K36ac and anti-H3 antibodies Indeed H3K36ac levels 163
were high at the transcribed euchromatic ACTIN7 gene but low at the silent 164
heterochromatic Ta2 transposon (Fig 2F) The increased H3 signal at Ta2 is consistent 165
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

8
with earlier reports and might be related to the more compact chromatin structure in 166
heterochromatin (Rehrauer et al 2010 Shu et al 2012) Together these results suggest 167
that H3K36ac is present on active genes in euchromatin but not on heterochromatic loci 168
169
H3K36ac is associated with the first two nucleosomes of transcriptionally active 170
genes 171
We performed a native ChIP-seq (N-ChIP-seq) experiment to determine the genome-172
wide localization of H3K36ac at mono-nucleosome resolution (Supplemental Tab S2) 173
To control for nucleosome density H3K36ac reads were normalized to the anti-H3 read 174
counts After mapping to the five Arabidopsis chromosomes the highest enrichment 175
was found along chromosome arms In contrast pericentromeric heterochromatic 176
regions were considerably less enriched (Supplemental Fig S4) Furthermore the 177
H3K36ac coverage profile was similar to the gene density distribution consistent with 178
an enrichment of H3K36ac at or near genes These results support the H3K36ac 179
immunolocalisation data To explore the distribution of H3K36ac along gene bodies in 180
more detail averaged coverage profiles were plotted around transcriptional start sites 181
(TSS) and transcriptional termination sites (TTS) for all annotated gene models (Fig 182
3A) The results show a strong enrichment of H3K36ac between 35 bp and 516 bp 183
downstream of the TSS (positions with 50 of maximal peak height) In contrast 184
H3K36ac was not enriched in promoter regions in gene bodies beyond the first 500 bp 185
distal to the TSS or around transcriptional termination sites (TTS) 186
Next we tested whether the width of the H3K36ac peak depended on gene 187
length by plotting averaged H3K36ac profiles separately for bins of increasing gene 188
length (Fig 3B) This analysis revealed that position shape and width of the H3K36ac 189
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

9
enrichment peak are independent of gene length and that H3K36ac covered only about 190
480 bp downstream of the TSS This invariant pattern of the H3K36ac modification 191
allowed the calculation of a gene-specific H3K36ac score as the mean H3K36ac signal 192
within a TSS-distal 35 bp to 516 bp window The H3K36ac score has a bimodal 193
distribution (Fig 3C) that allowed to separate 16431 genes with low H3K36ac (score 194
lt=27) from 10920 genes with high H3K36ac (score gt27) To avoid confounding effects 195
of alternative transcripts only genes with a single annotated transcript were included in 196
this analysis Since H3K36ac is found at genes and the differential enrichment of this 197
modification allows the clear distinction of two gene classes we asked if its presence is 198
a binary signal for transcription Globally genes with higher H3K36ac scores had also 199
higher mRNA levels (Fig 3D) Only active but not inactive genes had the pronounced 200
H3K36ac peak near the TSS (Fig 3E) However the relationship between H3K36ac 201
and transcript levels is not linear but strongly sigmoidal (red trend line in Supplemental 202
Fig S5) indicating that H3K36ac levels are only weakly proportional to transcription 203
rate and instead are a qualitative indicator of transcriptional activity This interpretation 204
is consistent with the bimodal distribution of H3K36ac scores (Fig 3C) 205
Since H3K36ac is associated with gene activity we analyzed its relationship to 206
other chromatin marks associated with transcription namely H3K4me3 H3K36me2 207
H3K36me3 and the histone variant H2AZ Similar to H3K36ac position shape and 208
width of the H3K4me3 and H2AZ enrichment peaks are independent of gene length 209
(Supplemental Fig S6) In contrast H3K36me3 and H3K36me2 profiles depend 210
strongly on gene length Next averaged H3K4me3 H3K36me2 H3K36me3 and 211
H2AZ profiles were plotted separately for five bins of increasing H3K36ac scores All 212
tested chromatin marks showed a positive correlation with H3K36ac but the correlation 213
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

10
with H3K4me3 and H3K36me3 was most pronounced (Supplemental Fig S7) 214
H3K4me3 H3K36me3 and H2AZ are restricted to the 5rsquo-end of genes while 215
H3K36me2 increases towards the 3rsquo-end To better compare peak positions averaged 216
profiles for the analyzed marks were plotted together (Fig 4) This analysis revealed 217
that H3K36ac and H2AZ are localized closest to the TSS and likely associated with 218
transcription initiation and the transition to transcription elongation (H3K36ac peaks at 219
199 bp and H2AZ at 252 bp distal to the TSS) Downstream of the H2AZ peak 220
H3K4me3 and H3K36me3 (peaks at 368 bp and 629 bp respectively) become more 221
dominating until H3K36me2 which is associated with elongating RNA Polymerase II 222
(Pol II) (Oh et al 2008) increases towards the 3rsquo-end of genes 223
Profiling of H3K36ac specifically along active genes showed the highest 224
enrichment around the TSS irrespective of feature length (Fig 5A B) consistent with 225
the distribution obtained when all genes were used (Fig 3A) Enrichment of 226
H3K36me3 on the other hand also reaches a peak right after the TSS but then extends 227
further towards the 3rsquo-end (Fig 5C D) Interestingly we observed that the enrichment 228
of H3K36me3 along the whole gene body increased with gene length (Fig 5D) This is 229
consistent with the model that H3K36me3 is associated with Pol II processivity (Oh et 230
al 2008) 231
Together H3K36ac is most enriched on +1 and +2 nucleosomes of the TSS of 232
transcriptionally active genes and correlates with H3K4me3 H3K36me2 H3K36me3 233
and the histone variant H2AZ 234
235
H3K36ac depends on GCN5 in Arabidopsis 236
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

11
Histone acetylation is established by HATs Arabidopsis has 12 different HAT genes 237
that are grouped into four classes (Pandey et al 2002) of which GCN5 (GENERAL 238
CONTROL NON DEREPRESSIBLE PROTEIN 5) is highly expressed in most 239
sporophytic tissues (Zimmermann et al 2004) In yeast GCN5 acetylates H3K14 (Roth 240
et al 2001) Because GCN5 is conserved in plants and because the H3 sequences 241
around K14 and K36 are similar (STGGK14AP versus STGGVK36KP) we tested if 242
Arabidopsis GCN5 acetylates H3K36 using the gcn5-1 mutant for X-ChIP assays The 243
TSS regions of two genes with high H3K36ac levels were analyzed using qPCR Their 244
H3K36ac levels were reduced more than five-fold in gcn5-1 (Fig 6A) indicating that 245
GCN5 is required for full H3K36 acetylation in vivo but that other HATs can also 246
acetylate H3K36 explaining the residual H3K36ac in gcn5-1 247
To test whether GCN5 can directly acetylate H3K36 in vitro HAT assays were 248
performed using amino-terminal H3 fragments and purified recombinant GST and GST-249
GCN5 Acetylated histone fragments were probed with anti-H3K36ac antibody A 250
strong signal was obtained when using GCN5 enzyme while no signal was obtained 251
with control GST or H3 alone (Fig 6B) The in vivo and in vitro assays therefore 252
establish Arabidopsis GCN5 as an H3K36 acetyl transferase and strongly suggest that it 253
is the main enzyme depositing H3K36ac in vivo 254
255
Competing modifications targeting H3K36 256
H3K36 can also be mono- di- or tri- methylated at its ɛ-amino group The Arabidopsis 257
histone lysine methyltransferase SET DOMAIN GROUP 8 (SDG 8) deposits di- and tri-258
methylation on H3K36 (Zhao et al 2005 Xu et al 2008) Considering that GNC5 and 259
SDG8 both modify H3K36 we tested the mechanistic dependency of acetylation and 260
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

12
methylation at H3K36 We performed X-ChIP-seq using the anti-H3K36ac antibody in 261
sdg8-2 and an anti-H3K36me3 antibody in gcn5-1 because in our experience X-ChIP is 262
more robust than N-ChIP when comparing genotypes Crosstalk between GCN5 and 263
SDG8 could result in reciprocal effects on H3K36ac and H3K36me3 enrichments 264
respectively To avoid confounding effects from inactive genes we restricted our 265
analysis to active protein-coding genes (see Methods) Globally H3K36ac levels in 266
genes increased in sdg8-2 in particular close to the TSS (Fig 7A) In addition 267
H3K36ac increased close to the TTS of genes in which its levels are normally low 268
Similarly enrichment of H3K36me3 increased in gcn5-1 and was highest in the center 269
of genes (Fig 7B) This increase in enrichment of acetylation and methylation in sdg8-2 270
and gcn5-1 respectively indicates that both modifications affect each other Because 271
the differences were found in distinct regions along gene bodies this crosstalk probably 272
does not reflect a direct competition for the H3K36 but an indirect functional repressive 273
interaction between the acetylation and methylation enzymes 274
275
HDA19 but not HDA6 affects H3K36ac levels 276
Eighteen different HDACs have been identified in Arabidopsis and grouped into three 277
different classes (Pandey et al 2002) To identify HDACs responsible for H3K36 278
deacetylation we tested the possible involvement of HDA6 and HDA19 which are two 279
major Arabidopsis HDACs HDA6 removes acetyl groups from H3K9ac H3K14ac 280
H3K18ac H3K23ac H3K27ac H4K5ac H4K8ac and H4K12ac (reviewed in 281
(Hollender et al 2008)) To test if HDA6 deacetylates H3K36ac we performed X-ChIP 282
assays with wild-type hda6-1 and 35SHDA6-FLAG plants using two genes with high 283
H3K36ac levels in wild type No significant differences of H3K36ac levels were found 284
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

13
in HDA6 loss-of-function or overexpressing plants (Supplemental Fig S8) suggesting 285
that HDA6 does not deacetylate H3K36 in vivo at least at these two loci In contrast a 286
considerable increase of H3K36ac levels was found for the same genes in an HDA19 287
RNAi line (Fig 8) indicating that HDA19 is one of the Arabidopsis HDACs that 288
deacetylase H3K36ac 289
290
H3K36ac is a conserved histone modification in plants 291
Histones are among the most highly conserved eukaryotic proteins and K36 is found in 292
H3 of mouse yeast and plants such as Arabidopsis tobacco wheat rice and Norway 293
spruce (Fig 9A) To test if H3K36ac exists only in Brassicaceae or also in other plant 294
species histones were extracted from tobacco rice wheat and spruce as representatives 295
of monocotyledonous and dicotyledonous angiosperms as well as gymnosperms The 296
presence of H3K36ac was analyzed using protein immunoblotting with anti-H3K36ac 297
antibody A distinct H3K36ac band was visible in all histone preparations (Fig 9B) 298
indicating that H3K36ac is a highly conserved modification in seed plants and likely has 299
a conserved biological function 300
To test whether the H3K36ac enrichment on active genes is conserved in other 301
plant species X-ChIP was performed using Norway spruce cell culture Two loci with 302
low expression (MA_16054g0010 and the EnSpm transposon-like locus MA_10270349) 303
and two loci with high expression (MA_491379g0010 and MA_94077g0010) were 304
selected for ChIP analysis (Supplemental Fig S9) using two primer sets for each gene ndash 305
one set probing a region upstream of the transcriptional unit and the other set probing a 306
region downstream of the TSS in the gene body near the start codon Similar to the 307
results in Arabidopsis strong H3K36ac ChIP signals were present at the active genes 308
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

14
MA_491379g0010 and MA_94077g0010 but no or only weak signals were present at 309
the silent or lowly expressed genes (Fig 9C) In addition H3K36ac was much higher 310
enriched distal of the TSS than in the promoter region Together these results suggest 311
that H3K36ac is present in the 5-region of active genes in the angiosperm Arabidopsis 312
and the gymnosperm Norway spruce revealing a wide conservation of this modification 313
among seed plants 314
315
Discussion 316
Applying the sensitive and specific CIDETD LC-MSMS method (Syka et al 2004) to 317
histone extracts from cauliflower inflorescences we have identified a novel H3 318
acetylation on K36 in plants CIDETD LC-MSMS allows sequencing of long highly 319
charged peptides (gt20 aa) and even of intact proteins and has been successfully used 320
already to determine histone PTMs (Taverna et al 2007 Xiong et al 2010 Jufvas et 321
al 2011) Our analysis confirmed most of the known Arabidopsis lysine acetylation 322
and methylation modifications We did not detect phosphorylated H3 peptides which 323
require phospho-peptide enrichment during histone extraction (Zhang et al 2007) 324
However our approach identified previously unreported plant histone modifications 325
including H3K36ac that we analyzed in more detail A complete list of the identified H3 326
post-translational modification is reported in Supplemental Table S1 327
H3K36ac was previously found in yeast Tetrahymena and mammals (Morris et 328
al 2007) Using a high-specificity antibody we confirmed the MSMS identification of 329
H3K36ac in plants In addition to cauliflower H3K36ac was also found in several other 330
seed plants The presence of H3K36ac in yeast ciliates animals and plants suggests an 331
evolutionary conserved function 332
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

15
In Arabidopsis H3K36ac is associated with euchromatin and excluded from 333
heterochromatic chromocenters It is enriched within the first 500 bp and particularly at 334
+1 and +2 nucleosomes distal to the TSS of transcriptionally active genes irrespective 335
of their lengths This presence of H3K36ac distal to the TSS of Arabidopsis genes is in 336
contrast to yeast in which H3K36ac is highest at promoters but mostly absent from 337
transcribed regions (Morris et al 2007) The restriction of H3K36ac to the 5rsquo-end of 338
genes and the positive correlation with gene expression suggests that it is associated 339
with the early phase of transcription but not with elongation Although H3K36ac marks 340
active genes its levels are not significantly correlated with transcription rates H3K36ac 341
rather marks transcribed genes in a qualitative manner with full acetylation reached 342
even at moderate transcription rates 343
The peak of H3K36ac enrichment in Arabidopsis coincided well with the H2AZ 344
peak at the 5rsquo-end of gene bodies It is not clear whether the different width of the 345
H2AZ and H3K36ac peaks is genuine or the result of the different ChIP methods in our 346
study and the report for H2AZ (Zilberman et al 2008) The coincidence of H2AZ and 347
H3K36ac suggests that nucleosomes in which H3K36 is acetylated often contain the 348
histone variant H2AZ It remains to be established however whether incorporation or 349
presence of H2AZ favors acetylation of H3K36 andor H3K36ac favors H2AZ 350
incorporation Similar to H3K36ac and H2AZ H3K4me3 and H3K36me3 peak at 5rsquo-351
ends of transcriptionally active genes In the case of H3K4me3 the peak width is 352
independent of gene length but the peak summit is shifted to the TTS The H3K36me3 353
peak is shifted even further towards the 3rsquo-end than the H3K4me3 peak The extension 354
of the H3K36me3 peak strongly depends on gene length H3K36me2 increases 355
downstream of the H3K36me3 peak and reaches its maximum only at the 3rsquo-end The 356
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

16
gradient of H3K36ac H2AZ H3K4me3 H3K36me2 and H3K36me3 along transcribed 357
genes provides a spatial map of histone modifications along transcription units 358
Histone acetylation is regulated by antagonistic HAT and HDAC activities The 359
Arabidopsis HAT GCN5 acetylates H3K36 in vitro and is necessary to achieve full 360
H3K36ac levels in vivo These results are consistent with the reported activity of GCN5 361
in yeast (Morris et al 2007) supporting a conserved mechanism for H3K36 acetylation 362
in eukaryotes although as described above the H3K36ac patterns differ between yeast 363
and plants (Morris et al 2007) 364
The Arabidopsis genome encodes 18 HDAC proteins that are grouped into three 365
main subfamilies (RPD3 HD2 and SIR2 family) (Pandey et al 2002) We found that 366
H3K36ac was increased in HDA19 RNAi lines suggesting that HDA19 is required for 367
H3K36ac homeostasis on target genes in vivo It remains to be established whether 368
other HDACs also modulate H3K36ac levels 369
Despite the evolutionary conservation of H3K36 modifications animals and 370
yeast seem to use them in functionally different mechanisms than plants In animals and 371
yeast H3K36me3 is associated with transcriptional elongation peaking towards the 3rsquo-372
end and H3K36ac in yeast is restricted to promoters In plants H3K36me3 peaks in the 373
5rsquo-half of the gene and H3K36ac is restricted to an approximately 480 bp distal to the 374
TSS In Arabidopsis H3K36ac and H3K36me3 are found in overlapping regions 375
downstream of the TSS Our genome-wide analysis revealed a negative crosstalk 376
between the HAT GCN5 and the HKMT SDG8 This resulted in increased H3K36ac 377
levels when H3K36me3 is reduced and vice versa the distinct spatial patterns of 378
H3K36ac and H3K36me3 were however not affected Further work will be required to 379
understand the mechanistic basis for this crosstalk It is tempting to speculate that 380
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

17
acetylation and methylation at H3K36 form a molecular switch in which one of the two 381
dominates However since nucleosomes have two H3 molecules concomitant 382
acetylation and methylation at H3K36 in the same nucleosome are possible It remains 383
to be shown whether such nucleosomes with asymmetrical K36 modifications do indeed 384
exist 385
386
Experimental procedures 387
Plant material 388
The following Arabidopsis thaliana lines were previously described HDA19-RNAi1 389
(Zhou et al 2005) gcn5-1 (Vlachonasios et al 2003) 35SFlag-HDA6 (Earley et al 390
2006) hda6-1 (Aufsatz et al 2002) and sdg8-2 (Xu et al 2008) The HDA19-RNAi1 391
was shown to have no off-target effects on the close HDA19 homolog HDA6 (Zhou et 392
al 2005 Mehdi et al 2015) Seeds were sown on 05times basal salts Murashige and 393
Skoog (MS) medium (Duchefa Haarlem The Netherlands) stratified at 4degC for one 394
day and then transferred to growth chambers for germination at 20degC under long-day 395
(LD 16 h light) photoperiods for ten days The seedlings were transferred to pots 396
containing soil and cultivated in growth chambers under LD conditions Tobacco 397
(Nicotiana tabacum) wheat (Triticum aestivum) and rice (Oryza sativa) were grown in 398
a greenhouse at 22degC in LD Cauliflower (Brassica oleracea) was obtained from a local 399
supermarket Norway spruce (Picea abies) needles were collected from an 400
approximately 15-years-old wild-grown tree in a forest close to Uppsala (Sweden) 401
located at 5981ordm N 1766ordm E For ChIP and RT-qPCR in Norway spruce embryogenic 402
calli (line 111811) derived from a zygotic embryo were used (von Arnold et al 403
2008) 404
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

18
405
Histone extraction and purification from plants 406
To extract histones from inflorescences of cauliflower and Arabidopsis or from fully 407
developed leaves of tobacco wheat rice and spruce approximately 3 g of material were 408
ground to a fine powder and homogenized for 15 min in histone extraction buffer [025 409
M sucrose 1 mM CaCl2 15 mM NaCl 60 mM KCl 5 mM MgCl2 15 mM PIPES pH 410
70 05 Triton X-100 including protease inhibitors cocktail (Roche) and 10 mM 411
sodium butyrate] After centrifugation for 20 min at 4degC and 4500timesg pellets were 412
dissolved in 01 M H2SO4 After further centrifugation for 10 min at 17000timesg total 413
histones were precipitated from the supernatant with concentrated Trichloroacetic acid 414
to a final concentration of 33 The histone pellet was washed twice with acetone-01 415
HCl air dried dissolved either in 1times Laemmli buffer and subjected to SDS-PAGE 416
followed by protein immunoblotting using anti-H3 (cat no 07-690 UpstateMillipore) 417
and anti-H3K36ac (cat no 39379 ActiveMotif) antibodies or fractionated by RP-418
HPLC 419
420
Histone fractionation by RP-HPLC and PTM identification by tandem mass 421
spectrometry 422
Bulk histones from cauliflower were prepared as described above The pellet was 423
dissolved in 100 μl of water sonicated for 1 min and centrifuged for 1 min at 10000timesg 424
The supernatant was gently aspirated by pipetting and loaded on an ECLIPS XDB-C8 425
(46times150 mm) (Agilent) connected to an Agilent HP1100 binary HPLC system Histone 426
variants were separated and eluted according to (Tweedie-Cullen et al 2009) The 427
fractions containing H3 were combined dried and dissolved in 50 mM ammonium 428
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

19
bicarbonate pH 80 After reduction with dithiothreitol (DTT 10 mM) and alkylation 429
with iodoacetamide (IAA 40 mM) histones H3 were digested for 2 h at 37degC with 430
Chymotrypsin (enzymesubstrate ratio 150 (ww)) (Promega Switzerland) or with 431
Arg_C (150 (ww)) (Roche Switzerland) Samples were desalted with C18 ZipTips 432
(Millipore Switzerland) following the manufacturerrsquos recommendations and dried 433
Peptides were resolubilized in 10 μl of 5 acetonitrile 01 trifluoracetic acid and 434
were analyzed by LC-MSMS on an LTQ-Orbitrap XL-ETD mass spectrometer 435
(Thermo Fischer Scientific Bremen Germany) coupled to an Eksigent-Nano-HPLC 436
system (Eksigent Technologies Dublin CA USA) Solvent composition at the two 437
channels was 02 formic acid 1 acetonitrile for channel A and 02 formic acid 438
80 acetonitrile for channel B Peptides were resolubilized in 10μl of 5 acetonitrile 439
and 01 formic acid and loaded on a self-made tip column (75 microm times 70 mm) packed 440
with reverse-phase C18 material (AQ 3 μm 200 Aring Bischoff GmbH Leonberg 441
Germany) Elution was performed with a flow rate of 200 nl min by a gradient from 3 442
to 10 of B in 6 min 35 B in 38 min 97 B in 45 min One scan cycle comprised of 443
a full-scan MS survey spectrum followed by up to 6 sequential CID and ETD MSMS 444
scans on the 3 most intense signals above a threshold of 500 Full-scan MS spectra 445
(300minus2000 mz) were acquired in the FT-Orbitrap with a resolution of 60 000 at 400 mz 446
after accumulation to a target value of 200000 while CID and ETD MSMS spectra 447
were recorded in the linear ion trap (target value of 1e4 for ion trap MSn scans) The 448
ETD anion target value was set at 3e5 and activation time at 50 ms Supplementary 449
activation was employed to enhance the fragmentation efficiency for 2+ precursors and 450
charge state dependent ETD time enabled The ETD reaction time was 120 ms and 451
isolation width was 3 mz Charge state screening was enabled and singly charge states 452
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

20
were rejected Precursor masses selected twice for MSMS were excluded for further 453
selection for 120 s Samples were acquired using internal lock mass calibration set on 454
mz 4290887 and 4451200 455
MS raw-files were converted into Mascot generic files (mgf) with Mascot 456
Distiller software 2420 (Matrix Science Ltd London UK) Peak lists were searched 457
using Mascot Server 23 against the Arabidopsis TAIR10 protein database Search 458
parameters were requirement for chymotryptic or ArgC ends one missed cleavage 459
allowed peptide tolerance plusmn 6 ppm MSMS tolerance plusmn 05 Da Carbamidomethylation 460
of cysteine was set as fixed modification while oxidation (methionine) acetylation (N-461
term protein lysine) mono- di- and tri-methylation (arginine lysine) were set as 462
variable To assess the location of the post-translational modification sites we used the 463
site localization analysis provided by Mascot The presence of modification sites was 464
further validated by manual inspection of spectra 465
466
Chromatin Immunopreciptation and ChIP-seq 467
Native and cross-linked chromatin immunoprecipitation (N-ChIP and X-ChIP) assays 468
were performed as described (Bernatavichute et al 2008 Exner et al 2009) using 469
gene-specific primers (Supplemental Tab S3) See supplemental information for 470
details For ChIP-seq leaf number six of plants grown for 35 days under short-day 471
conditions (8 h light) was harvested at zeitgeber time ZT=7 and used for nuclei 472
isolation Supplemental Table S2 gives a summary of the ChIP-seq experiments 473
474
Bioinformatics and ChIP-seq data analysis 475
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

21
Illumina reads were mapped to the Arabidopsis reference TAIR10 genome using 476
bowtie2 (version 21) (Langmead et al 2012) SAM file output from bowtie was 477
converted to BAM format using SAMTOOLS (version 14) (Li et al 2009) and 478
imported into R (version 2152) (httpwwwR-projectorg) using functions from the 479
Rsamtools package All subsequent analysis was performed in R Identical reads present 480
more than 25 times were considered as PCR artifacts and filtered out using the 481
filterDuplReads function from package HtSeqTools Replicates were pooled samples 482
normalization to identical sequencing depth and H3K36ac signals were normalized to 483
histone signals to control for variable nucleosome density using functions from package 484
nucleR (Flores et al 2011) H3K4me3 H3K36me2 H3K36me3 and H2AZ data were 485
taken from the literature (Oh et al 2008 Zilberman et al 2008) Gene expression data 486
are from (Shu et al 2012) and match cultivation conditions plant age harvest time and 487
tissue used here for ChIP-seq The expression cutoff to consider a gene as active was a 488
value of 6 in the microarray data from (Shu et al 2012) resulting in a set of 12102 489
active protein coding genes Heat maps were generated using the open source tool 490
deepTools (Ramirez et al 2014) 491
492
Constructs and recombinant protein expression and purification 493
The coding region for the amino-terminal tail of H3 (amino acids 1-50) was amplified 494
by PCR from Arabidopsis cDNA using primers LH1625 (CTATTC-495
TCTTAAAGCAACAGTTCC forward) and LH1626 (CACCATGGCTCGTACCAA 496
reverse) The PCR product was cloned into vector pET100-D-TOPO according to the 497
manufacturerrsquos recommendations (Invitrogen) pGEX4T1 was obtained from 498
(Amersham Life Science Stockholm Sweden) pQE30-GCN5 was previously 499
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

22
described (Benhamed et al 2006) Recombinant amino-terminal 6xHis tagged H31-50 500
was expressed in Escherichia coli BL21pLys Rosetta cells and purified on TALON 501
metal affinity resin (Clontech Stockholm Sweden) according to manufacturerrsquos 502
recommendations Recombinant GST and GST-GCN5 were expressed in BL21pLys 503
Rosetta and purified with Glutathione Sepharose 4B resin according to the 504
manufacturerrsquos recommendations (Amersham Life Science) Purified proteins were 505
stored at -20degC until use in HAT activity assays 506
507
Histone acetyl transferase activity assays 508
HAT assays using recombinant GST-GCN5 expressed in E coli were performed as 509
follows 1 microg of purified His-tagged histone H31-50 was mixed with purified 510
recombinant GST-GCN5 or with GST alone in HAT buffer [025 μgμl Acetyl-CoA 511
(Sigma-Aldrich) 50 mM Tris-HCl (pH 80) 10 glycerol (vv) 1 mM dithiothreitol 512
complete EDTA‐free protease inhibitor (Roche Indianapolis IN USA) and 10 mM 513
sodium butyrate] Acetylation reactions were incubated for 60 min at 30degC with gentle 514
shaking and terminated by addition of 2times SDS Laemmli sample buffer followed by 515
heating for 5 min to 100degC The reaction products were electrophoretically resolved on 516
15 SDS-PAGE gels and subjected to protein immunoblotting using anti-H3K36ac 517
(cat no 39379 Active Motif) and anti-H3 (cat no 07-690 UpstateMillipore) 518
antibodies 519
520
RNA isolation and RT-qPCR 521
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

23
RNA extraction and reverse transcription were performed as described previously 522
(Alexandre et al 2009) using gene-specific primers (Supplemental Tab S3) See 523
supplemental information for details 524
525
Immunostaining 526
Immunostaining was performed as described previously (Jasencakova et al 2000) with 527
some modifications See supplemental information for details 528
529
Accession numbers 530
Sequencing reads are deposited in Gene Expression Omnibus (GSE74841) 531
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

24
Supplemental Material 532
533
Supplemental_Materialpdf contains 534
- Supplemental Methods (RNA isolation and RT-qPCR ChIP and ChIP-seq 535
Immunostaining Sequence alignment of H3 amino-terminal tails) 536
- Supplemental Figures 537
Figure S1 Histone separation by RP-HPLC 538
Figure S2 Identification of H3K36me2 539
Figure S3 Identification of H3K36me2 540
Figure S4 Genome-wide distribution of H3K36ac on the Arabidopsis genome 541
Figure S5 H3K36ac is a qualitative mark for transcriptional activity 542
Figure S6 Dependency of histone marks on gene length 543
Figure S7 H3K36ac positively correlates with H3K4me3 H3K36me3 and 544
H2AZ 545
Figure S8 HDA6 does not affect H3K36ac steady state levels on AT3G04720 or 546
AT3G28060 547
Figure S9 Expression levels of Norway spruce genes used for ChIP 548
- Supplemental Tables 549
Table S1 List of histone H3 modifications that were identified in B oleracea 550
Table S2 Summary of ChIP-seq experiments 551
Table S3 Lists of primers used 552
553
554
Acknowledgments 555
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

25
We thank Cecilia Waumlrdig for technical help We thank Keqiang Wu Craig Pikaard and 556
Dao-Xiu Zhou for kindly providing seeds of HDA19-RNAi1 35SFlag-HDA6 gcn5-1 557
and the pQE30-GCN5 construct respectively We thank the FGCZ for Illumina 558
sequencing This work was supported by grants from the Swiss National Science 559
Foundation the Swedish Science Foundation VR and the Knut-and-Alice-Wallenberg 560
Foundation 561
562
Author contributions 563
WM JM MN and SH performed the experiments WM MT SH PH and LH analyzed 564
data WM CK and LH planned the experiments WM MT CK WG and LH wrote the 565
manuscript 566
567
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

26
Figure legends 568
569
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum 570
of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 571
peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-572
purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale 573
for the y-axis represents the relative abundance of the parent ion Above the spectrum is 574
the peptide sequence in which predicted c-type ions which contain the aminoterminus 575
of the peptide are immediately above the sequence Predicted z-type ions which 576
contain the carboxyterminus are immediately below the sequence ions observed in the 577
spectrum which represent masses associated with the fragmented peptides from the 578
MSMS analyses are underlined 579
580
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from 581
Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by 582
protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization 583
of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were 584
stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac 585
antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a 586
section through the nucleus that was analyzed using ImageJ software (E) Quantitative 587
line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 588
μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac 589
antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable 590
element 591
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

27
592
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac 593
enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment 594
was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is 595
independent of gene length Metagene plots are shown for the first (black) second (red) 596
third (green) and forth (blue) length quartile where the black line represents the 25 597
shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean 598
H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two 599
separate populations of genes with scores of less or more than 27 respectively (D) 600
Relation between H3K36ac score and transcriptional state H3K36ac scores are shown 601
for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) 602
H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and 603
highest (green) mRNA levels respectively Only active genes have the characteristic 604
H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012) 605
606
Figure 4 Metagene plots of active chromatin marks around the TSS Phased 607
enrichment of H3 modifications and the histone variant H2AZ around the TSS The 608
peak of enrichment is indicated for each profile Data are from (Reed et al 1996 609
Zilberman et al 2008) and from this study 610
611
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) 612
H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb 613
upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS 614
(B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments 615
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

28
are similar in level and breadth irrespective of the length and the fraction of the gene 616
being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an 617
absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative 618
scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further 619
towards the 3rsquo-end Longer genes have higher and wider levels of H3K36me3 Genes 620
were ordered by length 621
622
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on 623
AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-624
ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative 625
to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 626
Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B 627
resin blotted and probed with anti-GST antibodies M weight marker Right HAT 628
assays were performed using recombinant His-H3 as substrate The reaction products 629
were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed 630
with anti-H3K36ac antibodies Anti-H3 was used as control 631
632
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of 633
H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in 634
the gcn5-1 background Shaded areas represent 95 confidence intervals 635
636
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and 637
AT3G28060 are affected by HDA19 downregulation Ten-day-old seedlings were used 638
for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac 639
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

29
relative to H3 is shown as mean plusmn SEM 640
641
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the 642
amino-terminal tails of H3 from mouse yeast and several plants shows conservation of 643
lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) 644
wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were 645
resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac 646
antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) 647
H3K36ac is found downstream (downstr) of the TSS on transcribed genes 648
649
Literature Cited 650
Alexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) 651
Arabidopsis MSI1 is required for negative regulation of the response to drought 652
stress Mol Plant 2 675-687 653
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative 654
histone deacetylase needed to enhance DNA methylation induced by double-655
stranded RNA EMBO J 21 6832-6841 656
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications 657
Cell Res 21 381-395 658
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone 659
modifications Trends Biochem Sci 35 618-626 660
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and 661
TAF1HAF2 interact to regulate histone acetylation required for light-responsive 662
gene expression Plant Cell 18 2893-2903 663
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

30
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational 664
modifications of histone H2B-variants isolated from Arabidopsis thaliana J 665
Proteome Res 6 3655-3668 666
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-667
wide association of histone H3 lysine nine methylation with CHG DNA 668
methylation in Arabidopsis thaliana PLoS One 3 e3156 669
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation 670
during plant development Biochim Biophys Acta 1809 567-576 671
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and 672
polyamides Microchimica Acta 43 555-588 673
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant 674
development and polyploidy Biochim Biophys Acta 1769 295-307 675
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M 676
Viegas W Pikaard CS (2006) Erasure of histone acetylation by Arabidopsis 677
HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 678
20 1283-1293 679
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA 680
Cheung MS Day DS Gadel S Gorchakov AA Gu T Kharchenko PV Kuan S 681
Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of 682
histone-modification antibody quality Nat Struct Mol Biol 18 91-93 683
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W 684
Koumlhler C Hennig L (2009) The chromodomain of LIKE 685
HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and 686
function during Arabidopsis development PLoS ONE 4 e5335 687
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

31
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome 688
positioning Bioinformatics 27 2149-2150 689
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and 690
inheritance Nat Rev Genet 13 343-357 691
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative 692
simplicity Proc Natl Acad Sci U S A 102 5308-5309 693
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J 694
Integr Plant Biol 50 875-885 695
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA 696
binding properties of histone H4 amino terminus Thermal denaturation studies 697
reveal that acetylation markedly reduces the binding constant of the H4 tail to 698
DNA J Biol Chem 268 305-314 699
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master 700
switch for nuclear and cell differentiation Comm Integr Biol 8 e1056950 701
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 702
acetylation of euchromatin and heterochromatin is cell cycle dependent and 703
correlated with replication rather than with transcription Plant Cell 12 2087-704
2100 705
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080 706
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE 707
(2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct 708
combinations of post-translational modifications Nucleic Acids Res 32 6511-709
6518 710
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

32
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational 711
modifications in primary human fat cells PLoS One 6 712
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo 713
PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower 714
G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive 715
analysis of the chromatin landscape in Drosophila melanogaster Nature 471 716
480-485 717
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat 718
Methods 9 357-359 719
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G 720
Durbin R (2009) The Sequence AlignmentMap format and SAMtools 721
Bioinformatics 25 2078-2079 722
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant 723
Biol 61 395-420 724
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) 725
Single-nucleosome mapping of histone modifications in S cerevisiae PLoS Biol 726
3 727
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and 728
remodeling in plant abiotic stress responses Biochim Biophys Acta 1819 129-729
136 730
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) 731
MSI1 functions in a HDAC complex to fine-tune ABA signaling Plant Cell 732
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

33
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD 733
Lieb JD Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a 734
highly conserved histone modification J Biol Chem 282 7632-7640 735
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin 736
modification and gene expression in Arabidopsis PLoS Genet 4 e1000077 737
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J 738
Mount DW Jorgensen RA (2002) Analysis of histone acetyltransferase and 739
histone deacetylase families of Arabidopsis thaliana suggests functional 740
diversification of chromatin modification among multicellular eukaryotes 741
Nucleic Acids Res 30 5036-5055 742
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible 743
platform for exploring deep-sequencing data Nucleic Acids Res 42 W187-91 744
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects 745
responsiveness to gibberellins in Arabidopsis Plant Physiology 112 337-342 746
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N 747
Patrignani A Rombauts S Shu H Van de PeY Vuylsteke M Weigel D Zeller 748
G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis 749
transcriptome profiling Plant Physiol 152 487-499 750
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 751
81-120 752
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D 753
Caillieux E Duvernois-Berthet E Al-Shikhley L Giraut L Despres B 754
Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) 755
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

34
Integrative epigenomic mapping defines four main chromatin states in 756
Arabidopsis EMBO J 30 1928-1938 757
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader 758
to druggable target Biochim Biophys Acta 1839 676-685 759
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE 760
Bastolla U Gutierrez C (2014) The functional topography of the Arabidopsis 761
genome is organized in a reduced number of linear motifs of chromatin states 762
Plant Cell 26 2351-2366 763
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of 764
DNA accessibility in plant chromatin Nat Commun 3 1281 765
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 766
403 41-45 767
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein 768
sequence analysis by electron transfer dissociation mass spectrometry Proc Natl 769
Acad Sci U S A 101 9528-9533 770
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT 771
Hunt DF Allis CD (2007) Long-distance combinatorial linkage between 772
methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 773
104 2086-2091 774
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-775
translational modifications on synaptic nuclear and histone proteins in the adult 776
mouse brain J Proteome Res 8 4966-4982 777
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

35
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of 778
ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis 779
growth development and gene expression Plant Cell 15 626-638 780
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47 781
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and 782
acetylation in core histones of Neurospora crassa Biochemistry 49 5236-5243 783
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH 784
(2008) Di- and tri- but not monomethylation on histone H3 lysine 36 marks 785
active transcription of genes involved in flowering time regulation and other 786
processes in Arabidopsis thaliana Mol Cell Biol 28 1348-1360 787
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-788
translational modification patterns in Arabidopsis thaliana PLoS ONE 2 e1210 789
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by 790
expression of FLOWERING LOCUS C requires methylation of histone H3 K36 791
Nat Cell Biol 7 1156-1160 792
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is 793
involved in jasmonic acid and ethylene signaling of pathogen response in 794
Arabidopsis Plant Cell 17 1196-1204 795
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and 796
DNA methylation are mutually antagonistic chromatin marks Nature 456 125-797
129 798
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator 799
Arabidopsis microarray database and analysis toolbox Plant Physiol 136 2621-800
2632 801
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

36
802
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 1 Identification of H3K36ac in cauliflower MSMS fragmentation spectrum of the [M+2H]2+ parent ion at mz 15048474 This peptide was identified as the H3 peptide (histone H3 amino acids 27ndash40) derived from ArgC digested RP-HPLC-purified cauliflower H3 The position of acetylation at Lysine 36 is displayed the scale for the y-axis represents the relative abundance of the parent ion Above the spectrum is the peptide sequence in which predicted c-type ions which contain the aminoterminus of the peptide are immediately above the sequence Predicted z-type ions which contain the carboxyterminus are immediately below the sequence ions observed in the spectrum which represent masses associated with the fragmented peptides from the MSMS analyses are underlined
200 400 600 800 1000 1200 1400mz
0
25
50
75
100
Rel
ativ
eA
bund
ance
37739
67521
49744
746004243875465
6226847605
2612115916 9867571740616865215035311 9123982539 13498423319 30297 127584100664 14922612119811142414616 144880
z+1(
4)2+
z+2(
4)2+
z+1(
5)2+
z+2(
5)2+
z+1(
6)2+
z+2(
6)2+
z+1(
7)2+
z+2(
7)2+
z+1(
8)2+
z+2(
8)2+
z+1(
9)2+
z+2(
9)2+
z+1(
10)2+
z+2(
10)2+
z+2(
11)2+
z+1(
12)2+
z+2(
12)2+
z+1(
7)+
z+2(
7)+
z+2(
6)+
z+2(
4)+
z+1(
1)+
c(2)
+
c(5)
+
c(6)
+
c(7)
+
c(8)
+
c(9)
+ c(10
)+
[M+
4H]4+
[M+
4H]2+bullbull
c(6)
2+
K27 S A P T T G G V K36ac K P H R40
c(2)+ c(5)+ c(6)+ c(7)+ c(8)+ c(9)+ c(10)+c(6)2+
z+1(1)+z+1(4)2+
z+2(4)2+
z+1(5)2+
z+2(5)2+z+2(6)+z+1(6)2+
z+2(6)2+
z+1(7)+z+2(7)+z+1(7)2+
z+2(7)2+
z+1(8)2+
z+2(8)2+z+1(9)2+
z+2(9)2+z+1(10)2+
z+2(10)2+z+2(11)2+z+1(12)2+
z+2(12)2+
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 2 Detection of H3K36ac in Arabidopsis (A) Acid-extracted histones from Arabidopsis inflorescences were resolved on a 15 SDS-PAGE gel followed by protein immunoblotting using anti-H3 and anti-H3K36ac antibodies (B-D) Localization of H3K36ac in root tip nuclei Root tip interphase nuclei of Arabidopsis plants were stained with DAPI (B) and analyzed for immunofluorescence using an anti-H3K36ac antibody (C) (D) The overlay of DAPI and H3K36ac signal The line indicates a section through the nucleus that was analyzed using ImageJ software (E) Quantitative line profiles of DAPI (blue) and H3K36ac (red) florescence intensities Scale bars 5 μm (F) X-ChIP assays were performed using IgG anti-H3 and anti-H3K36ac antibodies with primers against the active ACTIN7 gene and the silent Ta2 transposable element
010203040506070
Inte
nsity
DAPIH3K36ac
A
B C D
E
F
Inpu
t
IgG
H3
H3K
36ac
ACTIN 7
Ta2
α-H3 α-H3K36ac
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-
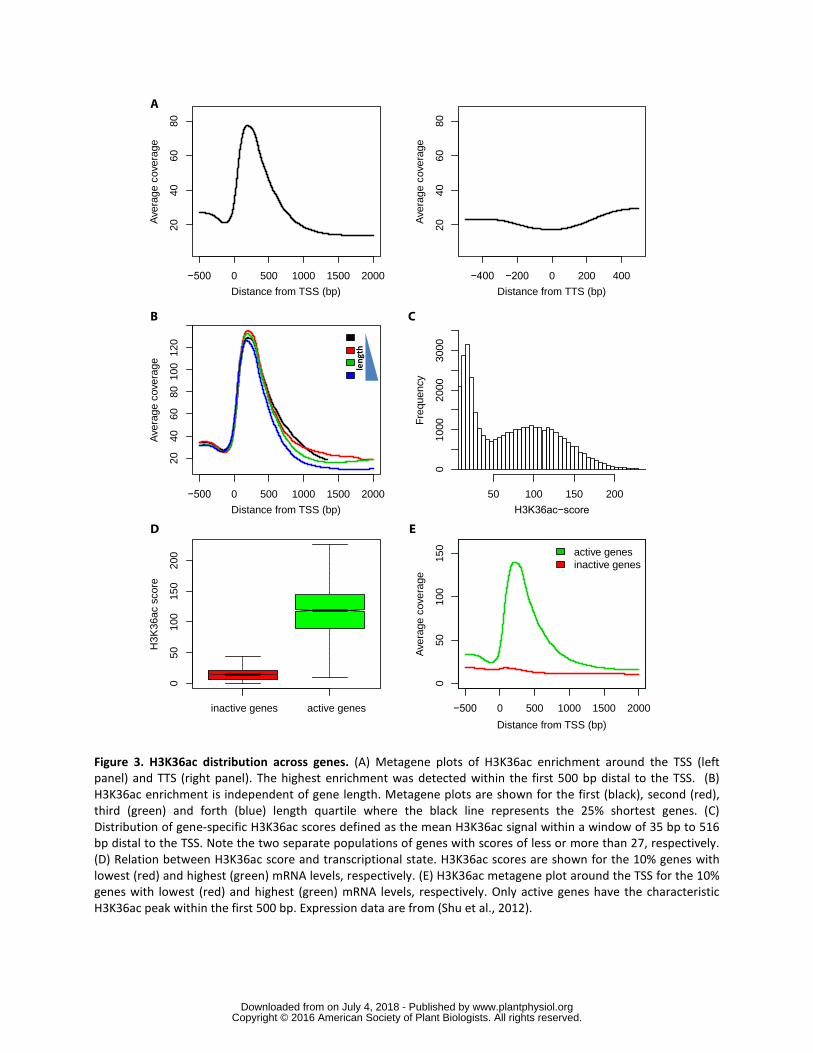
Figure 3 H3K36ac distribution across genes (A) Metagene plots of H3K36ac enrichment around the TSS (left panel) and TTS (right panel) The highest enrichment was detected within the first 500 bp distal to the TSS (B) H3K36ac enrichment is independent of gene length Metagene plots are shown for the first (black) second (red) third (green) and forth (blue) length quartile where the black line represents the 25 shortest genes (C) Distribution of gene-specific H3K36ac scores defined as the mean H3K36ac signal within a window of 35 bp to 516 bp distal to the TSS Note the two separate populations of genes with scores of less or more than 27 respectively (D) Relation between H3K36ac score and transcriptional state H3K36ac scores are shown for the 10 genes with lowest (red) and highest (green) mRNA levels respectively (E) H3K36ac metagene plot around the TSS for the 10 genes with lowest (red) and highest (green) mRNA levels respectively Only active genes have the characteristic H3K36ac peak within the first 500 bp Expression data are from (Shu et al 2012)
minus500 0 500 1000 1500 2000
2040
6080
Distance from TSS (bp)
Aver
age
cove
rage
minus400 minus200 0 200 400
2040
6080
Distance from TTS (bp)
Aver
age
cove
rage
minus500 0 500 1000 1500 2000
2040
6080
100
120
Distance from TSS (bp)
Aver
age
cove
rage
H3K36acminusscore
Freq
uenc
y
50 100 150 200
010
0020
0030
00
inactive genes active genes
050
100
150
200
H3K
36ac
sco
re
minus500 0 500 1000 1500 2000
050
100
150
Distance from TSS (bp)
Aver
age
cove
rage
active genesinactive genes
leng
th
A
B C
D E
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-
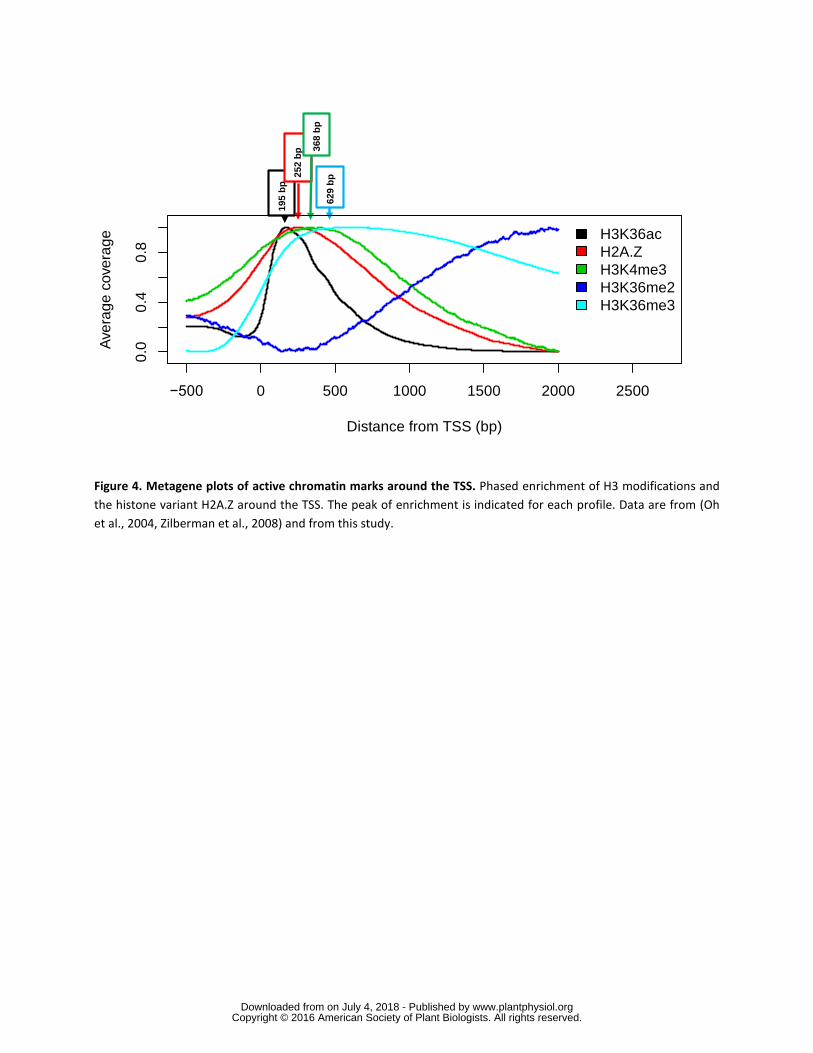
Figure 4 Metagene plots of active chromatin marks around the TSS Phased enrichment of H3 modifications and the histone variant H2AZ around the TSS The peak of enrichment is indicated for each profile Data are from (Oh et al 2004 Zilberman et al 2008) and from this study
195
bp25
2 bp
368
bp62
9 bp
minus500 0 500 1000 1500 2000 2500
00
04
08
Distance from TSS (bp)
Aver
age
cove
rage H3K36ac
H2AZH3K4me3H3K36me2H3K36me3
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 5 H3K36ac and H3K36me3 distribution across transcribed genes (A B) H3K36ac enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (A) and on a relative scale from TSS to TTS (B) H3K36ac was mainly found in the first 500 bp of the gene body The enrichments are similar in level and breadth irrespective of the length and the fraction of the gene being covered (C D) H3K36me3 enrichment along transcribed genes is shown on an absolute scale from 2 kb upstream to 4 kb downstream of the TSS (C) and on a relative scale from TSS to TTS (D) H3K36me3 localizes close to the TSS and expands further towards the 3rsquo end Longer genes have higher and wider levels of the trimethylation Genes were ordered by length
A B C D
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 6 GCN5 is required for H3K36 acetylation (A) H3K36ac levels on AT3G04720 and AT3G28060 depend on GCN5 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM (B) H3K36ac acetyltransferase activity of GCN5 Left Recombinant GST and GST-GCN5 were purified with Glutathione Sepharose 4B resin blotted and probed with anti-GST antibodies M weight marker Right HAT assays were performed using recombinant His-H3 as substrate The reaction products were resolved on a 15 SDS-PAGE gel transferred to PVDF membrane and probed with anti-H3K36ac antibodies Anti-H3 was used as control
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
l
00
02
04
06
08WSgcn5-1
AT3G28060
IgG H3K36ac00
01
02WSgcn5-1
M
GST
-GC
N5
GST
anti H3K36ac
anti H3
H31 GST GST-GCN5
A
B
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 7 Competing modifications of H3K36 by GCN5 and SGD8 (A) Gain of H3K36ac along genes in the sdg8-2 background (D) Gain of H3K36me3 along genes in the gcn5-1 background Shaded areas represent 95 confidence intervals
minus5
0
10
20
30D
iffer
ence
ina v
erag
e co
vera
ge
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36me3 gcn5minuswt
minus2 0 2 4
Distance from TSS
minus2 0 2 4
Distance from TTS (Kb)
H3K36ac sdg8minuswt
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 8 HDA19 affects H3K36ac levels H3K36ac levels on AT3G04720 and AT3G28060 are affected by HDA19 downregulation 10 days old seedlings were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies The enrichment of H3K36ac relative to H3 is shown as mean plusmn SEM
AT3G04720
IgG H3K36ac
Rel
H3K
36ac
leve
ls
00
02
04
06
08 ColHDA19 RNAi1
AT3G28060
IgG H3K36ac00
02
04
06 ColHDA19 RNAi1
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Figure 9 H3K36ac is conserved in seed plants (A) Amino acid alignment of the amino-terminal tails of H3 from mouse yeast and several plants shows conservation of lysine 36 (B) Acid-extracted histones prepared from tobacco (Nicotiana tabacum) wheat (Triticum aestivum) rice (Oryza sativa) and Norway spruce (Picea abies) were resolved on 15 SDS-PAGE gels followed by immunoblotting using anti-H3K36ac antibodies (C) H3K36ac X-ChIP in Norway spruce Shown are mean plusmn SEM (n=3) H3K36ac is found downstream (downstr) of the TSS on transcribed genes
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Parsed CitationsAlexandre C Moumlller-Steinbach Y Schoumlnrock N Gruissem W Hennig L (2009) Arabidopsis MSI1 is required for negative regulationof the response to drought stress Mol Plant 2 675-687
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Aufsatz W Mette MF van der WiJ Matzke M Matzke AJ (2002) HDA6 a putative histone deacetylase needed to enhance DNAmethylation induced by double-stranded RNA EMBO J 21 6832-6841
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bannister AJ Kouzarides T (2011) Regulation of chromatin by histone modifications Cell Res 21 381-395Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Barth TK Imhof A (2010) Fast signals and slow marks the dynamics of histone modifications Trends Biochem Sci 35 618-626Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Benhamed M Bertrand C Servet C Zhou DX (2006) Arabidopsis GCN5 HD1 and TAF1HAF2 interact to regulate histoneacetylation required for light-responsive gene expression Plant Cell 18 2893-2903
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bergmuumlller E Gehrig PM Gruissem W (2007) Characterization of post-translational modifications of histone H2B-variants isolatedfrom Arabidopsis thaliana J Proteome Res 6 3655-3668
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bernatavichute YV Zhang X Cokus S Pellegrini M Jacobsen SE (2008) Genome-wide association of histone H3 lysine ninemethylation with CHG DNA methylation in Arabidopsis thaliana PLoS One 3 e3156
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Berr A Shafiq S Shen WH (2011) Histone modifications in transcriptional activation during plant development Biochim BiophysActa 1809 567-576
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Cannon CG (1955) The interactions and structure of the-CONH-group in amides and polyamides Microchimica Acta 43 555-588Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen ZJ Tian L (2007) Roles of dynamic and reversible histone acetylation in plant development and polyploidy Biochim BiophysActa 1769 295-307
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Earley K Lawrence RJ Pontes O Reuther R Enciso AJ Silva M Neves N Gross M Viegas W Pikaard CS (2006) Erasure ofhistone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance Genes Dev 20 1283-1293
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Egelhofer TA Minoda A Klugman S Lee K Kolasinska-Zwierz P Alekseyenko AA Cheung MS Day DS Gadel S Gorchakov AAGu T Kharchenko PV Kuan S Latorre I Linder-Basso D Luu Y Ngo Q Perry M Re (2011) An assessment of histone-modificationantibody quality Nat Struct Mol Biol 18 91-93
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Exner V Aichinger E Shu H Wildhaber T Alfarano P Caflisch A Gruissem W Koumlhler C Hennig L (2009) The chromodomain ofLIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development PLoSONE 4 e5335
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Flores O Orozco M (2011) nucleR a package for non-parametric nucleosome positioning Bioinformatics 27 2149-2150 wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Greer EL Shi Y (2012) Histone methylation a dynamic mark in health disease and inheritance Nat Rev Genet 13 343-357Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Henikoff S (2005) Histone modifications combinatorial complexity or cumulative simplicity Proc Natl Acad Sci U S A 102 5308-5309Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hollender C Liu Z (2008) Histone deacetylase genes in Arabidopsis development J Integr Plant Biol 50 875-885Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hong L Schroth GP Matthews HR Yau P Bradbury EM (1993) Studies of the DNA binding properties of histone H4 aminoterminus Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 tail to DNA JBiol Chem 268 305-314
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Iwamoto M Hiraoka Y Haraguchi T (2015) The nuclear pore complex acts as a master switch for nuclear and cell differentiationComm Integr Biol 8 e1056950
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jasencakova Z Meister A Walter J Turner BM Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin iscell cycle dependent and correlated with replication rather than with transcription Plant Cell 12 2087-2100
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jenuwein T Allis CD (2001) Translating the histone code Science 293 1074-1080Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson L Mollah S Garcia BA Muratore TL Shabanowitz J Hunt DF Jacobsen SE (2004) Mass spectrometry analysis ofArabidopsis histone H3 reveals distinct combinations of post-translational modifications Nucleic Acids Res 32 6511-6518
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jufvas A Stralfors P Vener AV (2011) Histone variants and their post-translational modifications in primary human fat cells PLoSOne 6
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kharchenko PV Alekseyenko AA Schwartz YB Minoda A Riddle NC Ernst J Sabo PJ Larschan E Gorchakov AA Gu T Linder-Basso D Plachetka A Shanower G Tolstorukov MY Luquette LJ Xi R Jung YL (2011) Comprehensive analysis of the chromatinlandscape in Drosophila melanogaster Nature 471 480-485
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Langmead B Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2 Nat Methods 9 357-359Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Li H Handsaker B Wysoker A Fennell T Ruan J Homer N Marth G Abecasis G Durbin R (2009) The Sequence AlignmentMapformat and SAMtools Bioinformatics 25 2078-2079
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu C Lu F Cui X Cao X (2010) Histone methylation in higher plants Annu Rev Plant Biol 61 395-420Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Liu CL Kaplan T Kim M Buratowski S Schreiber SL Friedman N Rando OJ (2005) Single-nucleosome mapping of histonemodifications in S cerevisiae PLoS Biol 3
Pubmed Author and Title wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

CrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Luo M Liu X Singh P Cui Y Zimmerli L Wu K (2012) Chromatin modifications and remodeling in plant abiotic stress responsesBiochim Biophys Acta 1819 129-136
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Mehdi S Derkacheva M Ramstrom M Kralemann L Bergquist J Hennig L (2015) MSI1 functions in a HDAC complex to fine-tuneABA signaling Plant Cell
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Morris SA Rao B Garcia BA Hake SB Diaz RL Shabanowitz J Hunt DF Allis CD Lieb JD Strahl BD (2007) Identification ofhistone H3 lysine 36 acetylation as a highly conserved histone modification J Biol Chem 282 7632-7640
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oh S Park S van Nocker S (2008) Genic and global functions for Paf1C in chromatin modification and gene expression inArabidopsis PLoS Genet 4 e1000077
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Pandey R Muller A Napoli CA Selinger DA Pikaard CS Richards EJ Bender J Mount DW Jorgensen RA (2002) Analysis ofhistone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatinmodification among multicellular eukaryotes Nucleic Acids Res 30 5036-5055
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ramirez F Dundar F Diehl S Gruning BA Manke T (2014) deepTools a flexible platform for exploring deep-sequencing dataNucleic Acids Res 42 W187-91
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Reed JW Foster KR Morgan PW Chory J (1996) Phytochrome b affects responsiveness to gibberellins in Arabidopsis PlantPhysiology 112 337-342
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Rehrauer H Aquino C Gruissem W Henz SR Hilson P Laubinger S Naouar N Patrignani A Rombauts S Shu H Van de PeYVuylsteke M Weigel D Zeller G Hennig L (2010) AGRONOMICS1 A new resource for Arabidopsis transcriptome profiling PlantPhysiol 152 487-499
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roth SY Denu JM Allis CD (2001) Histone acetyltransferases Annu Rev Biochem 70 81-120Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Roudier F Ahmed I Berard C Sarazin A Mary-Huard T Cortijo S Bouyer D Caillieux E Duvernois-Berthet E Al-Shikhley L GirautL Despres B Drevensek S Barneche F Derozier S Brunaud V Aubourg S Schnittger (2011) Integrative epigenomic mappingdefines four main chromatin states in Arabidopsis EMBO J 30 1928-1938
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sanchez R Meslamani J Zhou MM (2014) The bromodomain from epigenome reader to druggable target Biochim Biophys Acta1839 676-685
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sequeira-Mendes J Araguez I Peiro R Mendez-Giraldez R Zhang X Jacobsen SE Bastolla U Gutierrez C (2014) The functionaltopography of the Arabidopsis genome is organized in a reduced number of linear motifs of chromatin states Plant Cell 26 2351-2366
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Shu H Wildhaber T Siretskiy A Gruissem W Hennig L (2012) Distinct modes of DNA accessibility in plant chromatin Nat Commun3 1281
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-

Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Strahl BD Allis CD (2000) The language of covalent histone modifications Nature 403 41-45Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Syka JE Coon JJ Schroeder MJ Shabanowitz J Hunt DF (2004) Peptide and protein sequence analysis by electron transferdissociation mass spectrometry Proc Natl Acad Sci U S A 101 9528-9533
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Taverna SD Ueberheide BM Liu Y Tackett AJ Diaz RL Shabanowitz J Chait BT Hunt DF Allis CD (2007) Long-distancecombinatorial linkage between methylation and acetylation on histone H3 N termini Proc Natl Acad Sci U S A 104 2086-2091
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tweedie-Cullen RY Reck JM Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic nuclearand histone proteins in the adult mouse brain J Proteome Res 8 4966-4982
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Vlachonasios KE Thomashow MF Triezenberg SJ (2003) Disruption mutations of ADA2b and GCN5 transcriptional adaptor genesdramatically affect Arabidopsis growth development and gene expression Plant Cell 15 626-638
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
von Arnold S Clapham D (2008) Spruce embryogenesis Methods Mol Biol 427 31-47Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xiong L Adhvaryu KK Selker EU Wang Y (2010) Mapping of lysine methylation and acetylation in core histones of Neurosporacrassa Biochemistry 49 5236-5243
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xu L Zhao Z Dong A Soubigou-Taconnat L Renou JP Steinmetz A Shen WH (2008) Di- and tri- but not monomethylation onhistone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsisthaliana Mol Cell Biol 28 1348-1360
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhang K Sridhar VV Zhu J Kapoor A Zhu JK (2007) Distinctive core histone post-translational modification patterns inArabidopsis thaliana PLoS ONE 2 e1210
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhao Z Yu Y Meyer D Wu C Shen WH (2005) Prevention of early flowering by expression of FLOWERING LOCUS C requiresmethylation of histone H3 K36 Nat Cell Biol 7 1156-1160
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou C Zhang L Duan J Miki B Wu K (2005) HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling ofpathogen response in Arabidopsis Plant Cell 17 1196-1204
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zilberman D Coleman-Derr D Ballinger T Henikoff S (2008) Histone H2AZ and DNA methylation are mutually antagonisticchromatin marks Nature 456 125-129
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zimmermann P Hirsch-Hoffmann M Hennig L Gruissem W (2004) Genevestigator Arabidopsis microarray database and analysistoolbox Plant Physiol 136 2621-2632
Pubmed Author and TitleCrossRef Author and TitleGoogle Scholar Author Only Title Only Author and Title
wwwplantphysiolorgon July 4 2018 - Published by Downloaded from Copyright copy 2016 American Society of Plant Biologists All rights reserved
- Parsed Citations
- Article File
- Figure 1
- Figure 2
- Figure 3
- Figure 4
- Figure 5
- Figure 6
- Figure 7
- Figure 8
- Figure 9
- Parsed Citations
-