allelic variation at a hypervariable compound microsatellite locus in the ascomycete ascochyta...
TRANSCRIPT

ORIGINAL PAPER
J. Geistlinger áK. Weising áW. J. Kaiser áG. Kahl
Allelic variation at a hypervariable compound microsatellite locusin the ascomycete Ascochyta rabiei
Received: 11 March 1997 /Accepted: 21 June 1997
Abstract The genome of the fungal chickpea pathogenAscochyta rabiei was screened for polymorphisms bymicrosatellite-primed PCR. While ethidium-bromidestaining of electrophoretically separated ampli®cationproducts showed only limited polymorphism among 24Tunisian A. rabiei isolates, Southern hybridization ofpuri®ed PCR fragments to restriction digests of fungalDNA revealed polymorphic DNA ®ngerprints. Oneparticular probe that gave rise to a hypervariable single-locus hybridization signal was cloned from the Syrianisolate AA6 and sequenced. It contained a large com-pound microsatellite harbouring the penta- and decam-eric repeat units (CATTT)n, (CATTA)n, (CATATC-ATTT)n and (TATTT)n. We call this locus ArMS1(Ascochyta rabiei microsatellite 1). Unique ¯anking se-quences were used to design primer pairs for locus-speci®c microsatellite ampli®cation and direct sequenc-ing of additional ArMS1 alleles from Tunisian and Pa-kistani isolates. A high level of sequence variation wasobserved, suggesting that multiple mutational mecha-nisms have contributed to polymorphism. Hybridizationand PCR analyses were performed on the parents and 62monoascosporic F1 progeny derived from a cross be-tween two di�erent mating types of the fungus. Progenyalleles could be traced back to the parents, with onenotable exception, where a longer than expected frag-ment was observed. Direct sequencing of this new lengthallele revealed an alteration in the copy number of theTATTT repeat [(TATTT)53 to (TATTT)65], while theremainder of the sequence was unchanged.
Key words Locus-speci®c microsatellite analysis áMicrosatellite-primed PCR á RFLP analysis áMutagenesis á Phytopathogenic fungi
Introduction
Microsatellites are tandemly reiterated, short DNAsequence motifs (usually 2±5 bp long) that occur atmultiple sites (up to 105) in eukaryotic genomes(Beckmann and Weber 1992; Wang et al. 1994; Fieldand Wills 1996). A key feature of this class of repet-itive DNA is an extraordinarily high level of variationamong taxa, mainly expressed as variation in the copynumber of tandem repeats at a particular locus. Sev-eral techniques have been developed to exploit thisvariability for DNA pro®ling and molecular markergeneration. Firstly, synthetic oligonucleotides comple-mentary to microsatellite motifs can be used as hy-bridization probes for DNA ®ngerprinting, revealingrestriction fragment length polymorphisms (RFLPs) atmultiple loci simultaneously (reviewed by Weising etal. 1995a). Secondly, length variation of individualmicrosatellites can be detected by PCR using a¯anking pair of primers, electrophoretic separation ofthe ampli®cation products, and visualization by auto-radiography or staining (reviewed by Powell et al.1996). Molecular markers of this type have beentermed sequence-tagged microsatellite site (STMS)markers by Beckmann and Soller (1990). Thirdly,mini- or microsatellite-speci®c oligonucleotides can beused as single PCR primers to amplify inter-repeatregions in genomic DNA (Heath et al. 1993; Meyer etal. 1993; Gupta et al. 1994; Zietkiewicz et al. 1994).Electrophoretic separation of such microsatellite-primed PCR (MP-PCR) products commonly results inDNA pro®les that resemble RAPD patterns (Williamset al. 1990; Weising et al. 1995b).
In fungi, the ubiquitous presence of microsatelliteswas ®rst demonstrated by DNA ®ngerprinting usingmicrosatellite-speci®c oligonucleotides as hybridization
Mol Gen Genet (1997) 256: 298±305 Ó Springer-Verlag 1997
Communicated by E. Cerda -Olmedo
J. Geistlinger á K. Weising á G. Kahl (&)Plant Molecular Biology, Department of Biology,Biocentre N200, University of Frankfurt/Main,Marie-Curie-Str. 9, D-60439 Frankfurt, GermanyFax: +49-69-7982-9268; e-mail: [email protected]
W. J. KaiserUnited States Department of Agriculture,Agricultural Research Service, Washington State University,Pullman, WA 99164-6402, USA

probes (reviewed by Rosewich and McDonald 1994;Weising et al. 1995a). Both yeast-like and ®lamentousfungi have also been investigated by microsatellite-primed PCR (Meyer et al. 1993; Meyer and Mitchell1995; Buscot et al. 1996; Longato and Bonfante 1997).However, only a few recent studies have been devoted tothe cloning of fungal mini- and microsatellites and/orthe generation of STMS markers (Groppe et al. 1995;Osiewacz et al. 1996; Andersen and Nilsson-Tillgren1997).
Ascochyta rabiei (Pass.) Labrousse [teleomorph:Didymella rabiei (Kovachevski) v. Arx] is the most im-portant fungal pathogen of chickpea (Cicer arietinumL.), a pulse crop plant grown in dry areas of Asia, Africaand Southern Europe. This ascomycete is a suitablemodel system for the study of microsatellites in fungi,since at least 25 di�erent microsatellite motifs have beendetected in its genome by extensive oligonucleotide ®n-gerprint analyses (Weising et al. 1991; Morjane et al.1994; Geistlinger et al. 1997). Target sequences recog-nized by the probes (CA)8, (CAA)5, (GACA)4, (GGAT)4and (GTTTGG)3 were found to be particularly abun-dant, and provided highly informative DNA ®nger-prints. However, the oligonucleotide ®ngerprinttechnique involves a lengthy experimental protocol, andis therefore not suitable for large-scale monitoring anddiagnostics of fungal populations. We therefore at-tempted to exploit microsatellite variability amongA. rabiei strains using PCR-based procedures. In thepresent communication we describe the use of micro-satellite-primed PCR, RFLP and locus-speci®c micro-satellite analysis for genotyping A. rabiei isolates. Wealso present sequence and inheritance data on a newtype of fungal microsatellite mainly consisting ofpentanucleotide repeats.
Materials and methods
Fungal material
In 1992, a total of 300 A. rabiei isolates were hierarchically sampled(McDonald and Martinez 1990) from di�erent chickpea-growingregions of Tunisia. For the present study, 24 isolates were selectedthat represented 24 di�erent haplotypes as determined by oligo-nucleotide ®ngerprinting with the four microsatellite probes (CA)8,(CAA)5, (CAT)5 and (GATA)4 (Morjane et al. 1994; H. Morjaneand J. Geistlinger, unpublished results). Single conidia from frozenglycerol stocks were germinated and grown on chickpea seed-mealagar as described (Morjane et al. 1994). After the ®rst vegetativereproduction cycle, a conidial suspension was transferred into liq-uid medium and incubated on a rotary shaker at 20°C. After 5±7days, mycelia were harvested by ®ltration through cheesecloth,lyophilized and stored at )80°C.
To generate a segregating population, the Syrian A. rabiei iso-late AA6 (ICARDA, Aleppo) was crossed with a North Americanisolate of a compatible mating type (MAT-1±1; deposited as ac-cession ATCC 76501 in the American Type Culture Collection;Rockville, Md., USA; Wilson and Kaiser 1995). A total of 62monoascosporic cultures were initiated as previously described(Geistlinger et al. 1997), mycelia were then harvested, lyophilizedand stored at )80°C.
DNA isolation
DNA was extracted from lyophilized mycelia as described previ-ously (Weising et al. 1991; Morjane et al. 1994). In short, myceliawere ground to a ®ne powder, dipersed in homogenization bu�er(2% cetyl trimethylammonium bromide, 1.4 M NaCl, 20 mMEDTA, 0.2% b-mercaptoethanol, 100 mM Tris-HCl pH 8.0), andincubated at 60°C for 30±45 min. After extraction with an equalvolume of chloroform/isoamylalcohol (24:1), phases were sepa-rated by centrifugation (5000 ´ g; 30 min). Nucleic acids wereprecipitated from the aqueous phase by adding 0.6 vol. of isopro-panol, and collected by centrifugation (5000 ´ g; 30 min). Pelletswere washed with 70% ethanol, dissolved in 0.5 ml of TE bu�er(10 mM Tris-HCl, 1 mM EDTA pH 8.0), and treated with 10 unitsof DNase-free RNase A for 3±5 h. RNase was removed by anadditional phenol/chloroform extraction step, DNA was precipi-tated with ethanol, centrifuged and dissolved in an appropriatevolume of TE bu�er.
Microsatellite-primed PCR (MP-PCR)
PCR with microsatellite-speci®c primers was performed in 25-llvolumes in a Perkin Elmer/Cetus TC-9600 thermocycler. ThePCR mixtures contained 20 mM Tris-HCl pH 8.4, 50 mM KCl,1.6 mM MgCl2, 0.2 mM dNTPs, 1 pmol/ll of primers[(GTTTGG)3, (GGAT)4, (GATA)4, (GACA)4 and (GAA)5, ei-ther singly or in various combinations], 0.03 U/ll Taq DNApolymerase (GoldStar, Eurogentech) and 1 ng/ll of templateDNA. After initial denaturation (94°C, 1 min), PCR was run for37 cycles consisting of a 94°C denaturing step (30 s), an an-nealing step (60 s) and a 72°C elongation step (60 s). Annealingtemperatures for the di�erent primers were calculated accordingto Thein and Wallace (1986). If two primers were used in asingle reaction, the lower temperature was selected for annealing.At the end of the run, a ®nal extension period was included(72°C; 150 s). Ampli®cation products were separated on 1.5%agarose gels in Tris-borate-EDTA bu�er (Sambrook et al. 1989),stained with ethidium bromide and photographed under UV il-lumination.
Cloning and sequencing of MP-PCR fragments
A subset of MP-PCR products obtained from the Syrian isolateAA6 was further analyzed by cloning and sequencing. Ethidiumbromide-stained bands were excised from agarose gels and passedthrough Whatman paper and glass wool using a syringe. DNA wasprecipitated, washed in 70% ethanol and redissolved in TE bu�er.The recovered PCR products were blunted with T4 DNA poly-merase, dNTPs and Mg2+ (Sambrook et al. 1989), and subse-quently ligated into the EcoRV site of the pBluescript II SK (�)phagemid (Stratagene). Ligation products were transformed intocompetent Escherichia coli SURE cells (Stratagene) using standardprocedures (Sambrook et al. 1989). Bacteria were grown in LuriaBroth supplemented with the appropriate antibiotics at 37°C.Plasmids were isolated using the alkaline lysis method (Sambrooket al. 1989). Plasmid inserts as well as puri®ed PCR products weresequenced with the ABI PRISM Dye Terminator Cycle SequencingReady Reaction Kit (Applied Biosystems), employing AmpliTaqDNA polymerase FS, T7 and T3 primers and the GeneAmp PCRSystem 9600 (Perkin-Elmer/Cetus). Sequence data generated by theABI 373 sequencer (Applied Biosystems) have been deposited inthe EMBL database (Accession No. Y09794). Some MP-PCRproducts were sequenced directly, using nested primers (primersequences correspond to bp 64±84 and bp 353±373 of the EMBLdatabase accession number Y09794). Homology searches in theGenBank and EMBL databases were performed using theBLASTN program (Altschul et al. 1990).
299
Locus-speci®c PCR (STMS analysis)
PCR primers for locus-speci®c microsatellite ampli®cation werededuced from ¯anking regions of the pentanucleotide repeat foundin a cloned MP-PCR product from isolate AA6. Primers weredesigned as 20mers with a GC content of 40%. Genomic DNAs ata concentration of 1 ng/ll were primed with 1 pmole/ll each ofthe oligonucleotides 5¢-GTAGAGTAGTTGTAGCTACT-3¢ and5¢-GGATCAATTACTAGGTTGCT-3¢ and 0.2 mM of eachdNTP. PCR bu�er conditions and polymerase concentration wereas described for MP-PCR. After 30 s of predenaturation at 96°C,37 cycles were run with 20 s denaturation at 94°C, 60 s annealingat 52°C and 60 s primer extension at 72°C, followed by a ®nalextension step (72°C, 150 s).
Southern analysis
Aliquots of fungal genomic DNA samples were digested withHinfI, electrophoresed in 1.0% agarose gels in TBE bu�er, dena-tured, neutralized and transferred onto a nylon membrane (Sam-brook et al. 1989). Cloned MP-PCR fragments were labeled by nicktranslation with [a32P]dCTP (Amersham), and hybridized to theblots containing the HinfI-digested fungal DNA samples understandard conditions (Sambrook et al. 1989). Blots were washed2 ´ 30 min with 0.1 ´ SSC at room temperature, followed by a10-min stringent wash at 63°C. Autoradiographs were developedon Kodak X-Omat-AR ®lms using intensifying screens.
Results
MP-PCR and RFLP analysis of A. rabiei isolates
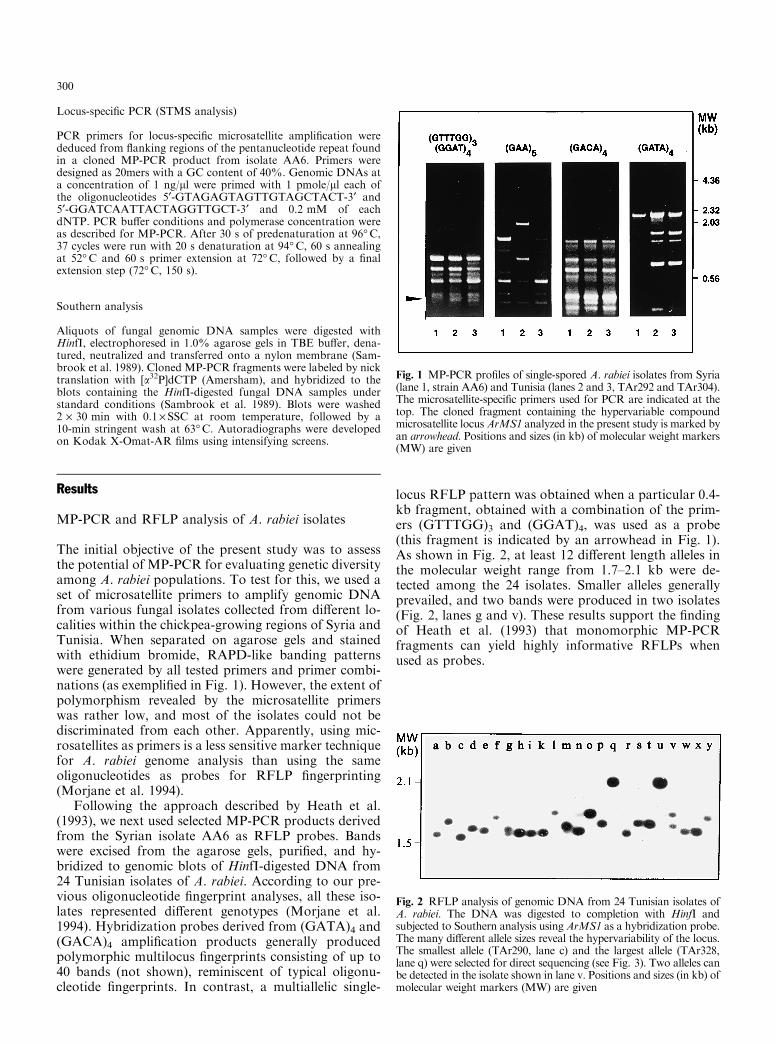
The initial objective of the present study was to assessthe potential of MP-PCR for evaluating genetic diversityamong A. rabiei populations. To test for this, we used aset of microsatellite primers to amplify genomic DNAfrom various fungal isolates collected from di�erent lo-calities within the chickpea-growing regions of Syria andTunisia. When separated on agarose gels and stainedwith ethidium bromide, RAPD-like banding patternswere generated by all tested primers and primer combi-nations (as exempli®ed in Fig. 1). However, the extent ofpolymorphism revealed by the microsatellite primerswas rather low, and most of the isolates could not bediscriminated from each other. Apparently, using mic-rosatellites as primers is a less sensitive marker techniquefor A. rabiei genome analysis than using the sameoligonucleotides as probes for RFLP ®ngerprinting(Morjane et al. 1994).
Following the approach described by Heath et al.(1993), we next used selected MP-PCR products derivedfrom the Syrian isolate AA6 as RFLP probes. Bandswere excised from the agarose gels, puri®ed, and hy-bridized to genomic blots of HinfI-digested DNA from24 Tunisian isolates of A. rabiei. According to our pre-vious oligonucleotide ®ngerprint analyses, all these iso-lates represented di�erent genotypes (Morjane et al.1994). Hybridization probes derived from (GATA)4 and(GACA)4 ampli®cation products generally producedpolymorphic multilocus ®ngerprints consisting of up to40 bands (not shown), reminiscent of typical oligonu-cleotide ®ngerprints. In contrast, a multiallelic single-
locus RFLP pattern was obtained when a particular 0.4-kb fragment, obtained with a combination of the prim-ers (GTTTGG)3 and (GGAT)4, was used as a probe(this fragment is indicated by an arrowhead in Fig. 1).As shown in Fig. 2, at least 12 di�erent length alleles inthe molecular weight range from 1.7±2.1 kb were de-tected among the 24 isolates. Smaller alleles generallyprevailed, and two bands were produced in two isolates(Fig. 2, lanes g and v). These results support the ®ndingof Heath et al. (1993) that monomorphic MP-PCRfragments can yield highly informative RFLPs whenused as probes.
Fig. 1 MP-PCR pro®les of single-spored A. rabiei isolates from Syria(lane 1, strain AA6) and Tunisia (lanes 2 and 3, TAr292 and TAr304).The microsatellite-speci®c primers used for PCR are indicated at thetop. The cloned fragment containing the hypervariable compoundmicrosatellite locus ArMS1 analyzed in the present study is marked byan arrowhead. Positions and sizes (in kb) of molecular weight markers(MW) are given
Fig. 2 RFLP analysis of genomic DNA from 24 Tunisian isolates ofA. rabiei. The DNA was digested to completion with HinfI andsubjected to Southern analysis usingArMS1 as a hybridization probe.The many di�erent allele sizes reveal the hypervariability of the locus.The smallest allele (TAr290, lane c) and the largest allele (TAr328,lane q) were selected for direct sequencing (see Fig. 3). Two alleles canbe detected in the isolate shown in lane v. Positions and sizes (in kb) ofmolecular weight markers (MW) are given
300

Characterization of a cloned MP-PCR productcontaining a compound microsatellite
The 0.4-kb MP-PCR fragment that revealed the hyper-variable single-locus RFLP was cloned and sequenced.As shown in Fig. 3 (uppermost line), more than half ofthe fragment consisted of a compound microsatellite of235 bp, ¯anked by unique sequences of 74 and 59 bp,respectively. As expected, the 5¢ and 3¢ ends of thefragment contained the primer sequences used for MP-PCR, i.e. (GTTTGG)3, and (CCTA)4, the inverse com-plement of (GGAT)4. The compound microsatellite inthe interior of the fragment exclusively consisted of AT-rich penta- and decanucleotide repeats with a consensussequence of 5¢-[(C/T)AT(A/T)(A/T)]n-3¢. We called thismicrosatellite locus ArMS1 (Ascochyta rabiei microsat-ellite 1).
To determine the molecular basis for the lengthvariation observed by RFLP analysis, two Tunisianisolates representing the smallest (Fig. 2, lane c) and thelargest RFLP fragment (Fig. 2, lane u), as well as oneisolate from Pakistan, were selected for direct sequenc-ing. Sequencing primers were derived from the uniqueDNA regions ¯anking the ArMS1 locus (Fig. 3). As isshown in the lower half of Fig. 3, the basic compositionof the ArMS1 locus was the same in all three alleles.However, the numbers of repetitions of particular motifswere drastically di�erent, and some base exchanges werealso observed. Thus, the Tunisian isolate representingthe largest allele in RFLP analysis (i.e. TAr328) wascharacterized by (in 5¢)>3¢ direction) a (CATTTT-ATTT)2 dimer, followed by 69 CATTT units and a(CATATCATTT)10 repeat. The TAr290 isolate, on theother hand, produced the smallest allele in RFLP anal-ysis and instead contained a (TATTT)21 followed by asingle CATTT motif and a (CATATCATTT)5 repeat.
The two outermost (CATTT)5 motifs were identical inboth isolates, as were the ¯anking unique sequences. Thearchitecture of the TAr290 allele closely resembled thatof the Pakistani isolate PK6 and only di�ered from thelatter by the number of CATTT- and TATTT-repeatunits.
In order to con®rm the results from RFLP analysisshown in Fig. 2, we also analyzed the collection of Tu-nisian isolates by locus-speci®c PCR (i. e. STMS anal-ysis) using primers that ¯ank the ArMS1 microsatellite.PCR products were separated on agarose gels andstained with ethidium bromide (not shown). While thePCR data generally con®rmed the RFLP results, somediscrepancies were also observed. For example, the twoisolates that displayed the largest RFLP fragments of2.1 kb (Fig. 2, lanes q and u) yielded ampli®cationproducts of 290 bp and 600 bp, respectively. This resultdemonstrates that similar-sized RFLP alleles are notnecessarily identical, and suggests that the actual allelenumber may be much higher than implied by RFLPdata. In two out of 24 A. rabiei genotypes, two loci weredetected by Southern hybridization (this is most obviousin isolate TAr335; Figure 2, lane v). These isolates also
Fig. 3 Allelic variation at the ArMS1 locus. The uppermost sequence(AA6) represents the MP-PCR fragment originally cloned, andincludes the microsatellite and its ¯anking regions (see EMBLAccession No. Y09794 for complete sequence data). All othersequences were identi®ed by direct sequencing of puri®ed PCRproducts, using nested primers that bind within the unique 59-bp and74-bp ¯anking regions, respectively. The upper three sequences arederived from the parents and one F1 mycelium of a cross between theSyrian isolate AA6 (P1) and the North American isolate ATCC 76501(P2). The triangle indicates an insertion of 10 bases (TATATTATTA)that is speci®c for the P2 allele. The lower three sequences are derivedfrom three randomly chosen A. rabiei isolates from Tunisia andPakistan. Binding sites for MP-PCR, STMS and nested sequencingprimers are indicated at the top
301
showed two ampli®cation products with the STMSprimers. Locus-speci®c PCR of one isolate each of A. pisiLIB. and A. fabae Speg. yielded single-locus ampli®ca-tion products of 260 and 280 bp, respectively, demon-strating the conservation of the ArMS1 ¯anking regionswithin the genus Ascochyta. Whether these products arealso polymorphic and contain a microsatellite in heter-ologous species, remains to be examined.
Expansion of a (TATTT)n repeat during meiosis
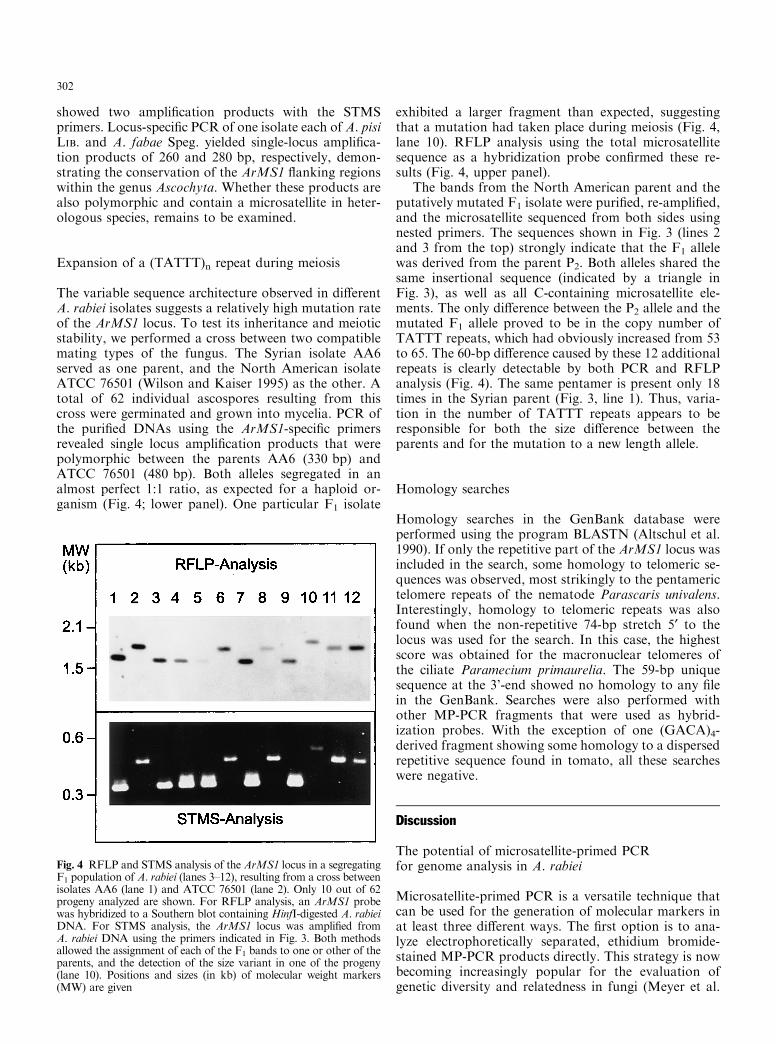
The variable sequence architecture observed in di�erentA. rabiei isolates suggests a relatively high mutation rateof the ArMS1 locus. To test its inheritance and meioticstability, we performed a cross between two compatiblemating types of the fungus. The Syrian isolate AA6served as one parent, and the North American isolateATCC 76501 (Wilson and Kaiser 1995) as the other. Atotal of 62 individual ascospores resulting from thiscross were germinated and grown into mycelia. PCR ofthe puri®ed DNAs using the ArMS1-speci®c primersrevealed single locus ampli®cation products that werepolymorphic between the parents AA6 (330 bp) andATCC 76501 (480 bp). Both alleles segregated in analmost perfect 1:1 ratio, as expected for a haploid or-ganism (Fig. 4; lower panel). One particular F1 isolate
exhibited a larger fragment than expected, suggestingthat a mutation had taken place during meiosis (Fig. 4,lane 10). RFLP analysis using the total microsatellitesequence as a hybridization probe con®rmed these re-sults (Fig. 4, upper panel).
The bands from the North American parent and theputatively mutated F1 isolate were puri®ed, re-ampli®ed,and the microsatellite sequenced from both sides usingnested primers. The sequences shown in Fig. 3 (lines 2and 3 from the top) strongly indicate that the F1 allelewas derived from the parent P2. Both alleles shared thesame insertional sequence (indicated by a triangle inFig. 3), as well as all C-containing microsatellite ele-ments. The only di�erence between the P2 allele and themutated F1 allele proved to be in the copy number ofTATTT repeats, which had obviously increased from 53to 65. The 60-bp di�erence caused by these 12 additionalrepeats is clearly detectable by both PCR and RFLPanalysis (Fig. 4). The same pentamer is present only 18times in the Syrian parent (Fig. 3, line 1). Thus, varia-tion in the number of TATTT repeats appears to beresponsible for both the size di�erence between theparents and for the mutation to a new length allele.
Homology searches
Homology searches in the GenBank database wereperformed using the program BLASTN (Altschul et al.1990). If only the repetitive part of the ArMS1 locus wasincluded in the search, some homology to telomeric se-quences was observed, most strikingly to the pentamerictelomere repeats of the nematode Parascaris univalens.Interestingly, homology to telomeric repeats was alsofound when the non-repetitive 74-bp stretch 5¢ to thelocus was used for the search. In this case, the highestscore was obtained for the macronuclear telomeres ofthe ciliate Paramecium primaurelia. The 59-bp uniquesequence at the 3'-end showed no homology to any ®lein the GenBank. Searches were also performed withother MP-PCR fragments that were used as hybrid-ization probes. With the exception of one (GACA)4-derived fragment showing some homology to a dispersedrepetitive sequence found in tomato, all these searcheswere negative.
Discussion
The potential of microsatellite-primed PCRfor genome analysis in A. rabiei
Microsatellite-primed PCR is a versatile technique thatcan be used for the generation of molecular markers inat least three di�erent ways. The ®rst option is to ana-lyze electrophoretically separated, ethidium bromide-stained MP-PCR products directly. This strategy is nowbecoming increasingly popular for the evaluation ofgenetic diversity and relatedness in fungi (Meyer et al.
Fig. 4 RFLP and STMS analysis of the ArMS1 locus in a segregatingF1 population of A. rabiei (lanes 3±12), resulting from a cross betweenisolates AA6 (lane 1) and ATCC 76501 (lane 2). Only 10 out of 62progeny analyzed are shown. For RFLP analysis, an ArMS1 probewas hybridized to a Southern blot containing HinfI-digested A. rabieiDNA. For STMS analysis, the ArMS1 locus was ampli®ed fromA. rabiei DNA using the primers indicated in Fig. 3. Both methodsallowed the assignment of each of the F1 bands to one or other of theparents, and the detection of the size variant in one of the progeny(lane 10). Positions and sizes (in kb) of molecular weight markers(MW) are given
302

1993; Meyer and Mitchell 1995; Buscot et al. 1996),plants (Gupta et al. 1994; Weising et al. 1995b; Tsumuraet al. 1996) and other organisms (Zietkiewicz et al.1994). Intraspeci®c variation of MP-PCR products wasshown to be high in some fungal species (e.g. Crypto-coccus neoformans, Meyer et al. 1993; Gremmeniellaabietina, Hantula and MuÈ ller 1997), but low in others(morels, Buscot et al. 1996; tru�es, Longato and Bon-fante 1997). In the present study, MP-PCR bandingpatterns were mainly monomorphic among A. rabieiisolates from di�erent countries (Fig. 1). Obviously,electrophoresis of MP-PCR products is less suitable forpopulation genetic studies, clone and pathotype identi-®cation in A. rabiei than is oligonucleotide ®ngerprinting(Weising et al. 1991; Morjane et al. 1994).
The second option for microsatellite-primed PCR isto convert an ampli®cation product into a hybridizationprobe for RFLP analysis. This approach, initially de-scribed for PCR with minisatellite-derived primers(Heath et al. 1993), proved to be quite successful in ourstudy. As was also observed by Heath et al. (1993),monomorphic MP-PCR products frequently producedpolymorphic RFLP patterns when used as probes. Thisis probably explained by the fact that unanchored MP-PCR excludes the two ¯anking microsatellites from theampli®cation product, while restriction fragments rec-ognized by an MP-PCR-derived probe contain the¯anking microsatellites as well as other (potentiallypolymorphic) sequences in their vicinity. MP-PCR-derived probes may hybridize to more than one locus,revealing complex ®ngerprint patterns, as were indeedobserved with most MP-PCR-derived probes used in thepresent study. Such multilocus ®ngerprints, however, aremore easily obtained by the direct use of microsatellite-speci®c oligonucleotides as probes.
Heath et al. (1993) also reported that in some cases,MP-PCR-derived probes generated polymorphic single-locus RFLP banding patterns. One such probe was alsoidenti®ed in the present study. Cloning and sequencingof the corresponding MP-PCR fragment revealed thepresence of a long, compound microsatellite consistingof penta- and decameric units. The identi®cation of amicrosatellite internal to an MP-PCR product highlightsthe third option for MP-PCR: the generation of locus-speci®c STMS markers without library construction.MP-PCR products can be cloned and/or sequenced di-rectly, and STMS primer pairs can be designed from the¯anking regions of microsatellites internal to the frag-ment. Of course, the applicability of this approach willdepend on the frequency of occurrence of such internalmicrosatellites in MP-PCR products. Whereas one mi-crosatellite was detected among four PCR products inthe present investigation, no internal repeats were foundin a previous study on kiwifruit and tomato, based onthe analysis of eight fragments encompassing a total of2000 nucleotides (K. Weising, unpublished results). Onthe other hand, Fisher et al. (1996) detected seven in-ternal microsatellites among 13 sequenced clones of anMP-PCR-derived Pinus radiata library. Additional data
are therefore required before a general conclusion can bedrawn regarding the suitability of MP-PCR for gener-ating STMS markers.
Allelic variation and putative mutational processesin a compound microsatellite
The compound ArMS1 microsatellite locus identi®ed inthe present study exhibited some interesting sequencefeatures. (1) It was exclusively built up from penta- anddecanucleotide motifs. (2) The repeat array was contin-uous and of a comparatively large size. With a totallength of 515 bp, the ArMS1 allele from the Tunisianisolate TAr328 is one of the largest uninterrupted mic-rosatellites described in any organism so far. (3) Themicrosatellite and its surrounding DNA were extremelyAT-rich. Thus, all six sequenced alleles contained aTTTATTT motif at their 5¢ end and a CATTT motif attheir 3¢ end, and the observed penta- and decanucleotidenucleotide motifs share the consensus sequence 5¢-(C/T)AT(A/T)(A/T)-3¢. Similar AT-rich pentanucleotiderepeats make up a large part of Drosophila satelliteDNA (Lohe and Brutlag 1986; Lohe et al. 1993), butwere previously not known in fungi. So far, only fewmicrosatellites (Groppe et al. 1995; Osiewacz et al. 1996)and one GC-rich 12-bp repeat minisatellite (Andersenand Nilsson-Tillgren 1997) have been characterized fromfungal genomes. With regard to its organization, theArMS1 locus does not easily ®t into any of the commoncategories of tandem repeats, since it combines typicalcharacteristics of microsatellites (compound repeat ofpentameric motifs), minisatellites (total length of 200±500 bp) and satellites (occurrence at a few genomic locionly).
The sequenced ArMS1 alleles were all di�erent fromeach other, indicating a relatively high mutation rate. Infact, the locus is so unstable that direct sequencing al-lowed us to detect one newly mutated allele in theprogeny of a cross. Nonparental bands indicating un-stable microsatellite loci have occasionally been ob-served by RFLP ®ngerprinting in plants (Rogstad 1994;HuÈ ttel, BuÈ nger and Weising, unpublished) and fungi(DeScenzo and Harrington 1994), but the molecularbasis of these mutations was not investigated. The newallele detected in the present study was clearly generatedby a variable number of tandem repeat (VNTR)-typemutation. Comparison of all sequenced alleles with eachother suggest that allelic variation at the ArMS1 locusresults from the combined action of VNTR mutations,transitions and transversions.
The precise mechanism of VNTR mutations is still amatter of debate, and may involve replication slippage,unequal exchange between homologous chromosomesor sister chromatids, gene conversion and other as yetpoorly de®ned processes (Sutherland and Richards 1995;McMurray 1995). Although replication slippage isknown to be facilitated by AT-richness in vitro(SchloÈ tterer and Tautz 1992), we consider slipped-strand
303

mispairing unlikely in this case. The large change in thenumber of repeating units (12) and bases (60) appears tobe more compatible with an unequal crossing-over eventduring meiosis. However, it remains to be shownwhether meiosis is generally required for the expansionof the investigated microsatellite, or if mutations alsooccur somatically during cell divisions or conidiosporeformation.
Finally, it would be interesting to know whether theArMS1 locus performs any function within the fungalgenome. The AT-rich pentanucleotides (AATAT)n,(AAGAC)n and (AAGAG)n have been identi®ed asmajor components of heterochromatic satellite DNA inD. melanogaster (Lohe et al. 1993). In A. rabiei, how-ever, ArMS1 is not likely to be part of a satellite, be-cause Southern experiments revealed that it occurs as asingle locus and has a total size that is atypical of sat-ellites. A striking fact is the high degree of homology ofthe repeat as well as of its 5¢ ¯anking sequence to sometelomeric sequences that are available in the databases.Telomeres of di�erent organisms exhibit considerablesequence variation (reviewed by Zakian 1995), and thetelomeric motifs of A. rabiei are still unknown. How-ever, for two reasons it is unlikely that the ArMS1 locusis part of a telomere. First, functional telomeres usuallyconsist of more or less homogenous repeat arrays, whileArMS1 has a compound architecture. Second, telomerictarget sequences should result in multiple bands or asmear in Southern experiments, which is not the casewith ArMS1. Thus, the functional signi®cance of theArMS1 locus, if any, remains obscure.
Acknowledgments This work was supported by BMZ Grant No.89.7860.3-01.130
References
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990)Basic local alignment search tool. J Mol Biol 215:403±410
Andersen TH, Nilsson-Tilgren T (1997) A fungal minisatellite.Nature 386:771
Beckmann JS, Soller M (1990) Towards a uni®ed approach togenetic mapping of eukaryotes based on sequence-tagged mi-crosatellite sites. Bio/Technology 8:930±932
Beckmann JS, Weber JL (1992) Survey of human and rat micro-satellites. Genomics 12:627±631
Buscot F, Wipf D, Di Battista C, Munch JC, Botton B, Martin F(1996) DNA polymorphism in morels: PCR/RFLP analysis ofthe ribosomal DNA spacers and microsatellite-primed PCR.Mycol Res 100:63±71
DeScenzo RA, Harrington TC (1994) Use of (CAT)5 as a DNA®ngerprinting probe for fungi. Phytopathology 84:534±540
Field D, Wills C (1996) Long, polymorphic microsatellites in sim-ple organisms. Proc R Soc Lond Ser B 263:209±215
Fisher P, Gardner RC, Richardson T (1996) Single-locus micro-satellites isolated using 5¢ anchored PCR. Nucleic Acids Res24:4369±4371
Geistlinger J, Maqbool S, Kaiser WJ, Kahl G (1997) Detection ofmicrosatellite ®ngerprint markers and their Mendelian inheri-tance in Ascochyta rabiei. Mycol Res, in press
Groppe K, Sanders I, Wiemken A, Boller T (1995) A microsatellitemarker for studying the ecology and diversity of fungal endo-
phytes (Epichloe spp.) in grasses. Appl Envir Microbiol61:3943±3949
Gupta M, Chyi YS, Romero-Severson J, Owen JL (1994) Ampli-®cation of DNA markers from evolutionarily diverse genomesusing single primers of simple sequence repeats. Theor ApplGenet 89:998±1006
Hantula J, MuÈ ller MM (1997) Variation within Gremmeniellaabietina in Finland and other countries as determined by ran-dom ampli®ed microsatellites (RAMS). Mycol Res 101:169±175
Heath DD, Iwama GK, Devlin RH (1993) PCR primed with theVNTR core sequences yields species-speci®c patterns and hy-pervariable probes. Nucleic Acids Res 21:5782±5785
Lohe AR, Brutlag DL (1986) Multiplicity of satellite DNA se-quences in Drosophila melanogaster. Proc Natl Acad Sci USA83:696±700
Lohe AR, Hilliker AJ, Roberts PA (1993) Mapping simple re-peated DNA sequences in heterochromatin of Drosophila me-lanogaster. Genetics 134:1149±1174
Longato S, Bonfante P (1997) Molecular identi®cation of my-corrhizal fungi by direct ampli®cation of microsatellite regions.Mycol Res 101:425±432
McDonald BA, Martinez JP (1990) DNA restriction fragmentlength polymorphisms among Mycosphaerella graminicola(anamorph Septoria tritici) isolates collected from a singlewheat ®eld. Phytopathology 80:1368±1373
McMurray CT (1995) Mechanisms of DNA expansion. Chromo-soma 104:2±13
Meyer W, Mitchell TG (1995) Polymerase chain reaction ®nger-printing in fungi using single primers speci®c to minisatellitesand simple repetitive sequences: strain variation in Crypto-coccus neoformans. Electrophoresis 16:1648±1657
Meyer W, Mitchell TG, Freedman EZ, Vilgalys R (1993) Hybrid-ization probes for conventional DNA ®ngerprinting used assingle primers in the polymerase chain reaction to distinguishstrains of Cryptococcus neoformans. J Clin Microbiol 31: 2274±2280
Morjane H, Geistlinger J, Harrabi M, Weising K, Kahl G (1994)Oligonucleotide ®ngerprinting detects genetic diversity amongAscochyta rabiei isolates from a single chickpea ®eld in Tunisia.Curr Genet 26:191±197
Osiewacz HD, Hamann A, Werner A (1996) Genome analysis of®lamentous fungi: identi®cation of a highly conserved simplerepetitive sequence in di�erent strains of Podospora anserina.Microbiol Res 151:1±8
Powell W, Machray GC, Provan J (1996) Polymorphism revealedby simple sequence repeats. Trends Plant Sci 1:215±222
Rogstad SH (1994) Inheritance in turnip of variable-number tan-dem-repeat genetic markers revealed with synthetic repetitiveDNA probes. Theor Appl Genet 89:824±830
Rosewich UT, McDonald BA (1994) DNA ®ngerprinting in fungi.Methods Mol Cell Biol 5:41±48
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: alaboratory manual (2nd edn). Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, New York
SchloÈ tterer C, Tautz D (1992) Slippage synthesis of simple se-quence DNA. Nucleic Acids Res 20:211±215
Sutherland GR, Richards RI (1995) Simple tandem DNA repeatsand human genetic disease. Proc Natl Acad Sci USA 92:3636±3641
Thein SL, Wallace RB (1986) The use of synthetic oligonucleotidesas speci®c hybridization probes in the diagnosis of genetic dis-orders. In: Davies KE (ed) Human genetic diseases: a practicalapproach. IRL Press, Oxford, UK, pp 33±50
Tsumura Y, Ohba K, Strauss SH (1996) Diversity and inheritanceof inter-simple sequence repeat polymorphisms in Douglas ®r(Pseudotsuga menziesii) and sugi (Cryptomeria japonica). TheorAppl Genet 92:40±45
Wang Z, Weber JL, Zhong G, Tanksley SD (1994) Survey of plantshort tandem repeats. Theor Appl Genet 88:1±6
Weising K, Kaemmer D, Epplen JT, Weigand F, Saxena M, KahlG (1991) DNA ®ngerprinting of Ascochyta rabiei with syntheticoligodeoxynucleotides. Curr Genet 19:483±489
304

Weising K, Nybom H, Wol� K, Meyer W (1995a) DNA ®nger-printing in plants and fungi. CRC Press, Boca Raton, Florida.
Weising K, Atkinson RG, Gardner RC (1995b) Genomic ®nger-printing by microsatellite-primed PCR: a critical evaluation.PCR Meth Appl 4:249±255
Williams JGK, Kubelik AR, Livak KJ, Rafalski JA, Tingey SV(1990). DNA polymorphisms ampli®ed by arbitrary primers areuseful as genetic markers. Nucleic Acids Res 18:6531±6535
Wilson AD, Kaiser WJ (1995) Cytology and genetics of sexualincompatibility in Didymella rabiei. Mycologia 87:795±805
Zakian VA (1995) Telomeres: beginning to understand the end.Science (Wash.) 270:1601±1607
Zietkiewicz E, Rafalski A, Labuda D (1994) Genome ®ngerprintingby simple sequence repeats (SSR)-anchored PCR ampli®cation.Genomics 20:176±183
305