an engineered microbial platform for direct biofuel production from brown macroalgae ·...
TRANSCRIPT

www.sciencemag.org/cgi/content/full/335/6066/308/DC1
Supporting Online Material for
An Engineered Microbial Platform for Direct Biofuel Production from Brown Macroalgae
Adam J. Wargacki, Effendi Leonard, Maung Nyan Win, Drew D. Regitsky, Christine Nicole S. Santos, Peter B. Kim, Susan R. Cooper, Ryan M. Raisner, Asael Herman, Alicia B. Sivitz, Arun Lakshmanaswamy, Yuki Kashiyama,
David Baker, Yasuo Yoshikuni*
*To whom correspondence should be addressed. E-mail: [email protected]
Published 20 January 2012, Science 335, 308 (2012)
DOI: 10.1126/science.1214547
This PDF file includes:
Materials and Methods
Figs. S1 to S7
References (44, 45)

Supporting Online Material I. Engineering Alginate Lyase Secretion System Materials
Sodium alginate, mannitol, glucose, and sodium citrate were purchased from Sigma. Luria-Bertani (LB) powder was purchased from Sigma, and LB broth was made following the manufacturer’s protocol. M9 powder was purchased from Amresco, and M9 medium (pH 7.2) was made following the manufacturer’s protocol without the addition of MgSO4 and CaCl2. For the alginate lyase activity assay, 0.2% (w/v) alginate solution was made by dissolving 0.1 g of sodium alginate in 50 mL of M9 medium. For the alginate-dependent growth assay, 1% and 2% (w/v) alginate growth solutions were made by dissolving 0.5 or 1 g of sodium alginate in 50 mL of M9 medium with the addition of MgSO4 and CaCl2 to final concentrations of 2 mM and 0.1 mM, respectively. Plasmid construction, sequences, and cell strains
The fosmid pCC1FOS (Epicentre Biotechnologies), harboring both the E. coli F-factor single-copy origin of replication and the inducible high-copy oriV, was used as a backbone vector for expressing and transforming the alginate lyase secretion system in E. coli. The gene fragment encoding the engineered Ag43 autotransporter protein (N455) was PCR-amplified from the genome of E. coli DH10B. The gene fragment encoding the truncated alginate lyase from Pseudoalteromonas sp. SM0524 (tSM0524 Aly) was synthesized. Using standard PCR and cloning procedures, the fusion gene encoding the Ag43 signal peptide, tSM0524 Aly and N455, was assembled with insertions of two restriction sites (BamHI and XbaI) as spacer sequences and cloned into the expression vector between EcoRI and XhoI. The entire alginate lyase secretion system expression construct was confirmed by sequencing (Elim BioPharm) and transformed into E. coli ATCC8739 (GenBank No. CP000946) for the activity assay. The nucleotide sequence of the entire expression construct (pCCN455+tSM0524 Aly) is given below. The construct consisted of the promoter (shown below in green), the ribosomal binding site (red), the Ag43 signal peptide (blue), tSM0524 Aly (orange), N455 (black), and the terminator (purple). Restriction sites defining the regions of these individual genetic components are underlined below and were present in the construct in this order: EcoRI, NcoI, BamHI, XbaI, HindIII, and XhoI. GAATTCAACTGCAAAAAATAGTTTGACACCCTAGCCGATAGGCTTTAAGATGTACCCAGTTCGATGAGAGCGATAACTCACACAGATATCATTAAAGAGGAGAAACCATGGATGAAACGACATCTGAATACCTGCTACAGGCTGGTATGGAATCACATGACGGGCGCTTTCGTGGTTGCCTCCGAACTGGCCCGCGCACGGGGTAAACGTGGCGGTGTGGCGGTTGCACTGTCTCTTGCCGCAGTCACGTCACTCCCGGTGCTGGCTGGATCCGATAACTCAAATGGTTCAACAATTCCTAGCAGCATAACCAGTGGTAGCATTTTTGATTTAGAAGGGGATAACCCAAATCCTCTCGTTGACGATAGCACCTTAGTGTTTGTGCCGTTAGGGGCACAACATATTACGCCTAATGGTAATGGCTGGCGTCATGAGTATAAGGTTAAAGAGAGCTTACGCGTTGCTATGACTCAAACCTATGAAGTGTTCGAAGCTACGGTAAAAGTTGAGATGTCTGATGGCGGAAAAACAATTATATCGCAGCACCATGCTAGTGATACCGGCACTATATCTAAAGTGTATGTGTCGGATACTGATGAATCGGGCTTTAATGATA

GCGTAGCGAACAACGGGATTTTTGATGTGTACGTACGTTTACGTAATACCAGCGGTAATGAAGAAAAATTTGCTTTGGGTACAATGACCAGCGGTGAGACATTTAACTTGCGGGTAGTTAATAACTACGGCGATGTAGAGGTTACGGCATTCGGTAACTCGTTCGGTATACCGGTAGAGGATGATTCGCAGTCATACTTTAAGTTTGGTAACTACCTGCAATCGCAAGACCCGTACACATTAGATAAATGTGGTGAGGCCGGAAACTCTAACTCGTTTAAAAACTGTTTTGAGGATTTAGGCATTACAGAGTCAAAAGTGACGATGACCAATGTGAGTTATACGCGTGAAACTAATTCTAGAGGCGGTGTACTGCTGGCCGATTCCGGTGCCGCTGTCAGTGGTACCAATAACGGCGCCATACTTACCCTTTCCGGGAAGACGGTGAACAACGATACCCTGACCATCCGTGAAGGCGATGCACTCCTGCAGGGAGGCTCTCTCACCGGTAACGGCAGCGTGGAAAAATCAGGAAGTGGCACACTCACTGTCAGCAACACCACACTCACCCAGAAAGCCGTCAACCTGAATGAAGGCACGCTGACGCTGAACGACAGTACCGTCACCACGGATGTCATTGCTCAGCGCGGTACAGCCCTGAAGCTGACCGGCAGCACTGTGCTGAACGGTGCCATTGACCCCACGAATGTCACTCTCGCCTCCGGTGCCACCTGGAATATCCCCGATAACGCCACGGTGCAGTCGGTGGTGGATGACCTCAGCCATGCCGGACAGATTCATTTCACCTCCACCCGCACAGGGAAGTTCGTACCGGCAACCCTGAAAGTGAAAAACCTGAACGGACAGAATGGCACCATCAGCCTGCGTGTACGCCCGGATATGGCACAGAACAATGCTGACAGACTGGTCATTGACGGCGGCAGGGCAACCGGAAAAACCATCCTGAACCTGGTGAACGCCGGCAACAGTGCGTCGGGGCTGGCGACCAGCGGTAAGGGTATTCAGGTGGTGGAAGCCATTAACGGTGCCACCACGGAGGAAGGGGCCTTTGTCCAGGGGAACAGGCTGCAGGCCGGTGCCTTTAACTACTCCCTCAACCGGGACAGTGATGAGAGCTGGTATCTGCGCAGTGAAAATGCTTATCGTGCAGAAGTCCCCCTGTATGCCTCCATGCTGACACAGGCAATGGACTATGACCGGATTGTGGCAGGCTCCCGCAGCCATCAGACCGGTGTAAATGGTGAAAACAACAGCGTCCGTCTCAGCATTCAGGGCGGTCATCTCGGTCACGATAACAATGGCGGTATTGCCCGTGGGGCCACGCCGGAAAGCAGCGGCAGCTATGGATTCGTCCGTCTGGAGGGTGACCTGATGAGAACAGAGGTTGCCGGTATGTCTGTGACCGCGGGGGTATATGGTGCTGCTGGCCATTCTTCCGTTGATGTTAAGGATGATGACGGCTCCCGTGCCGGCACGGTCCGGGATGATGCCGGCAGCCTGGGCGGATACCTGAATCTGGTACACACGTCCTCCGGCCTGTGGGCTGACATTGTGGCACAGGGAACCCGCCACAGCATGAAAGCGTCATCGGACAATAACGACTTCCGCGCCCGGGGCTGGGGCTGGCTGGGCTCACTGGAAACCGGTCTGCCCTTCAGTATCACTGACAACCTGATGCTGGAGCCACAACTGCAGTATACCTGGCAGGGACTTTCCCTGGATGACGGTAAGGACAACGCCGGTTATGTGAAGTTCGGGCATGGCAGTGCACAACATGTGCGTGCCGGTTTCCGTCTGGGCAGCCACAACGATATGACCTTTGGCGAAGGCACCTCATCCCGTGCCCCCCTGCGTGACAGTGCAAAACACAGTGTGAGTGAATTACCGGTGAACTGGTGGGTACAGCCTTCTGTTATCCGCACCTTCAGCTCCCGGGGAGATATGCGTGTGGGGACTTCCACTGCAGGCAGCGGGATGACGTTCTCTCCCTCACAGAATGGCACATCACTGGACCTGCAGGCCGGACTGGAAGCCCGTGTCCGGGAAAATATCACCCTGGGCGTTCAGGCCGGTTATGCCCACAGCGTCAGCGGCAGCAGCGCTGAAGGGTATAACGGTCAGGCCACACTGAATGTGACCTTCTGAAGCTTGGCTGTTTTGGCGGATGAGAGAAGATTTTCAGCCTGATACAGATTAAATCAGAACGCAGAAGCGGTCTGATAAAACAGAATTTGCCTGGCGGCAGTAGCGCGGTGGTCCCACCTGACCCCATGCCGAACTCAGAAGTGAAACGCCGTAGCGCCGATGGTAGTGTGGGGTCTCCCCATGCGAGAGTAGGGAACTGCCAGGCATCAAATAAAACGAAAGGCTCAGTCGAAAGACTGGGCCTTTCGTTTTATCTGTTGTTTGTCGGTGAACGCTCTCCTGAGTAGGACAAATCCGCCGGGAGCGGATTTGAACGTTGCGAAGCAACGGCCCGGAGGGTGGCGGGCAGGACGCCCGCCATAAACTGCCAGGCATCAAATTAAGCAGAAGGCCATCCTGACGGATGGCCTTTTTGCGTTTCTACAAACTCTTTTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTCGAG
Alginate lyase activity assay
Alginate lyases catalyze depolymerization (degradation) of alginate through cleavage of the 4-O-glycosidic bond via a β-elimination mechanism, which leads to formation of a double bond between C-4 and C-5 and production of 4-deoxy-L-erytho-hex-4-ene pyranosyluronate at the non-reducing end of the resulting oligomers. This unsaturated alginate at the non-reducing terminus can be detected by measuring UV light absorbance at 232 or 235 nm. An alginate lyase-expressing strain was pre-grown overnight in 3 mL of LB broth at 25oC and back-diluted into a fresh broth of the same volume to an OD600 of ~0.1 in the morning. The back-diluted culture was grown for a certain period of time (Fig. 2) before taking lyase activity measurements. To measure the activity in the cell-culture medium, which represents the total extracellular activity or

the sum of the activities in the supernatant and on the cell surface (whole cells alone), no further sample preparation was done. To measure the activities in the supernatant and on the cell surface (whole cells) separately, each cell-culture sample was spun down (13,300 rpm) at room temperature for 5 min, the supernatant was collected, and the cell pellet was washed and re-suspended in the same volume of M9 medium. To measure the activity within the cell, pelleted cells were lysed by re-suspending in the same volume of 1x BugBuster protein extraction reagent containing Lysonase (Novagen) following the manufacturer’s protocol. 1 µL of a protease inhibitor cocktail (Thermo Scientific) was also added to the lysis reaction mixture to inhibit protease activities. After sample preparation, 5 µL of each sample was added to a 96-well plate (Costar UV Transparent Flat Bottom; Product # 3635) followed by addition of 180 µL of 0.2% alginate solution and thorough mixing. The lyase activity was measured using a Biotek plate reader set at 232 nm, 25oC, and 5-sec shaking before each reading. One unit (U) of the lyase activity is defined as the amount of lyase required to produce an increase of 1.0 in absorbance at 232 nm per minute. II. Engineering Oligoalginate Transport and Metabolism, and Ethanol Production Phenotype Construction of pALG1 using V. splendidus genomic library
Cloning of genes predicted to be responsible for alginate metabolism based on bioinformatics analysis was carried out through three different strategies: (1) cloning directly using PCR, (2) cloning subsets of genes and reconstructing artificial operons, and (3) constructing a fosmid library followed by screening for growth. The third strategy was the most successful. Genomic DNA was isolated from Vibrio splendidus 12B01 (a gift from Dr. Martin Polz, MIT) using the DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA). A fosmid library was constructed using Copy Control Fosmid Library Production Kit (Epicentre, Madison, WI). This library consisted of random genomic fragments of ~40 kb inserted into the vector pCC1FOS (Epicentre, Madison, WI). The fosmid library was packaged into phages, and E. coli DH10B cells were transfected with the phage library. Transformants were selected for chloramphenicol resistance and then screened for their ability to grow on degraded alginate. Fosmids were isolated from E. coli clones that grew well on this medium using FosmidMAX DNA Purification Kit (Epicentre, Madison, WI). To verify the ability to confer growth on degraded alginate, isolated fosmids were used to transform fresh DH10B cells, and these cells were tested for ability to grow on alginate. As a negative control, E. coli DH10B cells, which cannot grow on degraded alginate, were used. An isolated fosmid (pALG1) sufficient to confer growth on alginate as a sole source of carbon was sequenced by Elim Biopharmaceuticals (Hayward, CA) using the primers Uni R3: 5’-GGGCGGCCGCAAGGGGTTCGCGTTGGCCGA-3’ and PCC1FOS_uni_F: 5’-

GGAGAAAATACCGCATCAGGCG-3’. E. coli ATCC8739 was transformed with pALG1 and used in subsequent experiments as the prototype host. Single gene deletion of pALG1
Each orf in the region of V12B01_24189 to V12B01_24244 in pALG1 was individually deleted to identify those essential to conferring oligo-alginate utilization in E. coli. Gene deletion techniques were based on the Wanner and Datsenko method (44). As a cassette of a selection marker, the kanamycin-resistant gene was PCR amplified from pkD13 using a pair of primer sequences containing 20 bp of the 5’- and 3’-termini of the kanamycin gene priming site and 45 bp of the corresponding upstream and downstream homology region flanking the deletion gene target. E. coli DH5 harboring pALG1 was transformed with pkD46. A positive transformant selected based on growth at 30°C with simultaneous ampicillin and chlorampenicol resistance was used as a host for homologous recombination. After induction of Red recombinase in pkD46 with 0.2% arabinose, the kanamycin cassette was used to transform the host strain. Positive transformants were selected based on growth at 37°C with Kanamycin resistance. Several colonies were then replica plated on chloramphenicol and kanamycin plates. The colonies that grew on both plates were subjected to colony PCR to select those with a deletion of the target orf. Mutated pALG1 obtained from the positive clone was then isolated and used to transform fresh E. coli ATCC 8739 for growth characterization. Construction of pALG2
pALG2 (pALG1 + kdgN-toaC-kdgM) was built on the integration of orf V12B01_24309 (kdgN), orf V12B01_24324 (toaC), and orf V12B01_24269 (kdgM). These genes were first subcloned into a vector based on the pKm plasmid backbone (R6Kbased vector containing kanamycin-resistant gene [Km] flanked by FRT sites). The kdgM, toaC, kdgN, and pKm sequences were amplified by PCR (Finnzyme’s Phusion, New England Biolabs) from pKm vector and V. splendidus genomic DNA, respectively. The forward primers for kdgN, toaC, kdgM, and pKm are 5’- GAATCGTTTTCCGGGACGCCTTAAGTACTCGGCTCTTATTTAAATG -3’, 5’- GTTTTATTCATGGTATTAATTCCATTTTTTAATGGACGAGGGGAAAGTG -3’, 5’-GAAATAATTTTAAAAGCCCCAATAGGGGATGAAGCTAATTCTGATTAG -3’, and 5’-CGGGTACCGGGCCCCCCCTCGAGGTC -3’, respectively. The reverse primers for kdgN, toaC, kdgM, and pKm are 5’-CACTTTCCCCTCGTCCATTAAAAAATGGAATTAATACCATGAATAAAAC -3’, 5’-CTAATCAGAATTAGCTTCATCCCCTATTGGGGCTTTTAAAATTATTTC -3’, 5’-CGGGATCCTCAGCACAGAAACTACTTTTG -3’, and 5’-CATTTAAATAAGAGCCGAGTACTTAAGGCGTCCCGGAAAACGATTC -3’, respectively. Each amplified DNA fragment was gel purified and adjoined by a second

round of PCR. The forward primer was 5’- CGGGTACCGGGCCCCCCCTCGAGGTC -3’ and the reverse primer was 5’- CGGGATCCTCAGCACAGAAACTACTTTTG -3’. The amplified fragment was then digested with BamHI (New England Biolabs) and ligated with T4 DNA ligase to form pKm-kdgN-toaC-kdgM. The constructed plasmid was sequenced (Elim Biopharmaceuticals) and the sequence of the insert was confirmed. Finally, Km-kdgN-toaC-kdgM from this vector was amplified by PCR using forward primer 5’-CTGACCGTTCTGTCCGTCACTTCCC -3’ and reverse primer 5’-TTTAATCGTTAGATTCTAATAGCTAGCCTCCAATTAGGCGGTTGTTGATTTAGAAGGAAA -3’. The amplified fragment was gel purified and transformed into a strain harboring pALG1 to construct pALG2 via homologous recombination using the method described by Wanner and Datsenko (44). Construction of pALG3
pALG3 was built on the construction of pALG1.1 (pALG1 + orfs V12B01_24254 [alyA] and V12B01_24259 [alyB]) and pALG1.2 (pALG1.1 + orfs V12B01_24264 [alyC], V12B01_24269 [kdgM], and V12B01_24274 [alyD]). These genes were subcloned into a pKm-based plasmid backbone.
For construction of pALG1.1, first, the pKm and alyAB sequences were amplified by PCR (Finnzyme’s Phusion, New England Biolabs) from pKm vector and V. splendidus genomic DNA, respectively. The forward primers for pKm and alyAB were 5’- CGGGATCCGTCGACCTGCAGTTCGAAG -3’ and 5’- TGTCAAACATGAGAATTAATTCCGGTTGATGAGCAGCTTTAAGGTTTAAT -3’, respectively. The reverse primers for pKm and alyAB were 5’- ATTAAACCTTAAAGCTGCTCATCAACCGGAATTAATTCTCATGTTTGACA -3’ and 5’- CGGGATCCCATACGCTTAAGCCCAACCAACAGC -3’, respectively. Next, each amplified DNA fragment was gel purified and adjoined by a second round of PCR. The forward primer was 5’- CGGGATCCGTCGACCTGCAGTTCGAAG -3’ and the reverse primer was 5’- CGGGATCCCATACGCTTAAGCCCAACCAACAGC -3’. The amplified fragment was then digested with BamHI (New England Biolabs) and ligated with T4 DNA ligase to form pKm-alyAB. The constructed plasmid was sequenced (Elim Biopharmaceuticals) and the DNA sequence of the insert was confirmed. The Km-alyAB was then amplified by PCR from pKm-alyAB with forward 5’- TTGATGAGCAGCTTTAAGGTTTAATG -3’ and reverse 5’- CTCACTATAGGGCGAATTCGAGCTCGGTACCCGGGGATCCGTGTAGGCTGGAGCTGCTTC -3’. The amplified fragment was gel purified and transformed into a strain harboring pALG1 to construct pALG1.1 via homologous recombination using the method described by Wanner and Datsenko (44).
For construction of pALG1.2, first, the pKm and alyC-kdgM-alyD sequences were amplified by PCR (Finnzyme’s Phusion, New England Biolabs) from pKm vector and V. splendidus genomic DNA, respectively. The forward primers for pKm and alyC-kdgM-

alyD were 5’- CGGGATCCGTCGACCTGCAGTTCGAAG -3’ and 5’- TGTCAAACATGAGAATTAATTCCGGTCTAATCGAATAACACTTAATATTAAAGG -3’, respectively. The reverse primers for pKm and alyC-kdgM-alyD were 5’- CCTTTAATATTAAGTGTTATTCGATTAGACCGGAATTAATTCTCATGTTTGACA -3’ and 5’- ACTCCGTATCGAGTTGTCGTCCTAA -3’, respectively. Next, each amplified DNA fragment was gel purified and adjoined by a second round of PCR. The forward primer was 5’- CGGGATCCGTCGACCTGCAGTTCGAAG -3’ and the reverse primer was 5’- ACTCCGTATCGAGTTGTCGTCCTAA -3’. The amplified fragment was then treated with T4 polynucleotide kinase (New England Biolabs) and ligated with T4 DNA ligase to form pKm-alyC-kdgM-alyD. The constructed plasmid was sequenced (Elim Biopharmaceuticals) and the DNA sequence of the insert was confirmed. The Km-alyC-kdgM-alyD was then amplified by PCR using forward primer 5’- TCTAATCGAATAACACTTAATATTAAAGG -3’ and reverse primer 5’- CTCACTATAGGGCGAATTCGAGCTCGGTACCCGGGGATCCGTGTAGGCTGGAGCTGCTTC -3’. The amplified fragment was gel purified and transformed into a strain harboring pALG1.1 to construct pALG1.2 via homologous recombination using the method described by Wanner and Datsenko (44).
Finally, to construct pALG3, a Km-kdgN-toaC cassette from pKm-kdgN-toaC-kdgM was amplified by PCR using forward primer 5’- CTGACCGTTCTGTCCGTCACTTCCC -3’ and reverse primer 5’- TCTTCAACCACAATCACCTGTTCCGTAGTGCCTAAACCATCAGCACAGAAACTACTTTTG -3’. This amplified fragment Km-kdgN-toaC was gel purified and transformed into E. coli DH5a harboring pALG1.2 via the Wanner and Datsenko method to form pALG3 (44). Growth characterization
Degraded alginate stock was prepared by dissolving 2% sodium alginate (Sigma-Aldrich, St. Louis, MO) in 1x M9 salts medium. 10 μg/mL Flavobacterium Aly (Sigma-Aldrich, St. Louis, MO) was added to the medium. Enzymatic de-polymerization was carried out at 30°C for 24 hr with shaking. Another batch of degraded alginate stock was prepared using SM0524 Aly. For this purpose, the gene coding for Pseudoalteromonas SM0524 Aly was expressed in the outer membrane of E. coli (EPI300) using the Ag43 signal peptide sequence. E. coli expressing this Aly was grown for 24 hr at 30°C in LB supplemented with 12.5 μg/mL chloramphenicol. On the following day, the cells were centrifuged and washed three times with 1x M9. To prepare degraded alginate stock solution, the cell pellet was used as a biocatalyst to de-polymerize 2% sodium alginate (using the same media and conditions as when using Flavobacterium Aly). For use as a growth medium, the degraded alginate was supplemented with 2 mM MgSO4, and 100 μM CaCl2. When necessary, 1x M9 was used to dilute the degraded alginate medium. In vitro assays

The orfs encoding for Atu3025 (40) and DehR were PCR amplified and cloned into pET28b (EMD4Biosciences, Darmstadt, Germany) in between NdeI and XhoI sites. The resulting plasmids were transformed into E. coli strain BL21(DE3). Positive transformants were inoculated into 50 mL of LB medium containing 50 μg/mL kanamycin. Cultures were incubated at 37°C until an optical density (OD600nm) of 0.6 was reached. Cultures were removed to a 25°C shaker, induced with 0.1 mM Isopropyl β-D-1-thiogalactopyranoside (IPTG), and allowed to over-express the protein of interest for 12 hr. After expression, cultures were centrifuged and the supernatant discarded. The cell pellet was re-suspended in 20 mM Tris-HCl pH 7/1x BugBuster buffer, and lysed on ice using the accompanying lysonase. Cleared lysates were obtained through centrifugation. Proteins were purified using Ni-NTA spin columns (Qiagen, Valencia, CA). Columns were washed with 20 mM Tris-HCl/10 mM imidazole and eluted with 20 mM Tris HCl/150 mM imidazole. Proteins were quantified using the Bradford method. All in vitro reactions with a total volume of 100μL were carried out at 30°C in a 96-well plate.
For DehR assays, the reactions contained 20 mM Tris HCl pH 7, 5.4 μg/mL of partially purified Atu3025, 10 μg/mL of partially purified DehR, 0.4 mM of NAD(P)H, and 29 mM of sodium alginate. NAD(P)H decay was traced via absorbance at 340 nm using a 96-well plate reader.
To assess the role of Oal in alginate metabolism in vitro, E. coli harboring pALG1, pALG1(∆oalA), pALG1(∆oalB), and pALG1(∆oalC) were inoculated from a freezer stock into 3 mL LB (chloramphenicol 12.5 mg/L). These strains were grown at 30°C overnight. The next day, 500 μL of the culture was back-diluted into 50 mL of 1x M9 medium containing 1.5% alginate degraded by Flavobacterium Aly (Sigma-Aldrich) in a 250-mL shake flask. Cultures were incubated overnight at 25°C. Cells were pelleted by centrifugation and lysed in 3 mL of BugBuster buffer by lysonase. 50 μL of lysate was added to 950 μL of 1x M9 containing 1 g/L alginate oligomers (prepared by degrading alginate with SM0524 Aly) and incubated at 25°C for 24 hr. Residual oligomers were detected via high performance liquid chromatography-mass spectrometry (HPLC-MS).
Engineering homoethanol pathway and deletion of by-product pathways
To reduce the formation of the by-products lactate and formate, ldhA and pflA-focA genes were deleted via homologous recombination. The DNA fragment of FRT-Km-FRT was first amplified by PCR using forward primers 5’- ACTGGTCAGAGCTTCTGCTGTCAGGAATGCCTGGTGCCCGGTGTAGGCTGGAGCTGCTTC -3’ and 5’- TTACATAGATTGAGTGAAGGTACGAGTAATAACGTCCTGCGTGTAGGCTGGAGCTGCTTC -3’ and reverse primers 5’- TTTGGCTTTGAGCTGGAATTTTTTGACTTTCTGCTGACGGATTCCGGGGATCCGTCGACC -3’ and 5’- CTGCTGCAATGGCCAAAGTGGCCGAAGAGGCGGGTGTCTAATTCCGGGGATCCG

TCGACC -3’ for the deletion of ldhA and pflB-focA, respectively. The amplified fragment was gel purified and transformed into E. coli strains ATCC8739 and ATCC8739 (ldhA) via homologous recombination using the method described by Wanner and Datsenko (44) to create ATCC8739 (ldhA) and ATCC8739 (ldh,pflB-focA), respectively.
To achieve stable ethanol production, pdc and adhB derived from Zymomonas mobilis were cloned into a pKm-based vector. The DNA fragments of adhB, pdc, and pKD13 were first amplified by PCR using forward primers 5’- TTAGAAAGCGCTCAGGAAGAGTTCT -3’, 5’- TGAAGAAGCCAT TATATATACCTCCTTAGAGGAGCTTGTTAACAGGCTTA-3’, and 5’- AGTATAACTCATTATATATACCTCCTGTAGGCTGGAGCTGCTTCGAAGTT -3’ and reverse primers 5’ – TAAGCCTGTTAACAAGCTCCTCTAAGGAGGTATATATAATGGCTTCTTCA -3’, 5’- AACTTCGAAGCAGCTCCAGCCTACAGGAGGTATATATAATGAGTTATACT -3’, and 5’- AATCGCTCAAGACGTGTAATGCTGC -3’ for adhB, pdc, and pkD13, respectively. The amplified fragments were gel purified, and adjoined by PCR using the primer set 5’- TTAGAAAGCGCTCAGGAAGAGTTCT -3’ and 5’- AATCGCTCAAGACGTGTAATGCTGC -3’. The amplified fragment was then treated with T4 DNA polynucleotide kinase and ligated with T4 DNA ligase to form pKD13-adhB-pdc-FRT-Km-FRT. The DNA fragment of Zymomonas mobilis adhB-pdc-FRT-Km-FRT was integrated into the frd gene locus of the E. coli host. The integration cassette for the fragment was first amplified by PCR using the forward primer 5’- CGACACCAATCAGCGTGACAACTGTCAGGATAGCAGCCAGTTAGAAAGCGCTCAGGAAGA -3’ and the reverse primer 5’- TCTCAAAAGTATACCCGATGCGTAGCCATACCGTTGCTGCATTCCGGGGATCCGTCGACC -3’. The amplified fragment was gel purified and transformed into E. coli strain ATCC8739 (ldhA, pflB-focA) via homologous recombination to form ATCC8739 (ldhA, pflB-focA, and frd::pdc-adhB).
To over-express pdc and adhB, the promoter PG25, derived from T5 phage, was integrated immediately upstream of the pdc-adhB artificial operon. PG25 was initially sub-cloned into pKD13. The DNA fragment of PG25 was synthesized via PCR using the forward primer 5’- GTATTGGAAAATTTTATCAAGAAATTTTTATTTTTCCATATGTAGGCTGGAGCTGCTTC -3’ and the reverse primer 5’- TATTATAATATTGTTATTAAAGAGGAGAAATTAACCACACAATCGCTCAAGACGTGTAAT -3’ using pKD13 as a template. The amplified fragment was then treated with T4 DNA polynucleotide kinase and ligated with T4 DNA ligase to form pKD13-PG25-FRT-Km-FRT. The DNA fragment of PG25-FRT-Km-FRT was integrated into the frd gene locus of the E. coli host. The integration cassette for the fragment was first amplified by PCR using the forward primer 5’- AAATAGGTACCGACAGTATAACTCATTATATATACCTCCTGTGTGGTTAATTTCTCCTCT -3’ and the reverse primer 5’- TCTCAAAAGTATACCCGATGCGTAGCCATACCGTTGCTGCATTCCGGGGATCCGTCGACC -3’. The amplified fragment was gel purified and transformed into an E. coli strain ATCC8739 (ldhA, pflB-focA) via homologous recombination to construct ATCC8739 (ldhA, pflB-focA, and frd::PG25-pdc-adhB)

Analytical methods
Cell mass was estimated by measuring absorbance at 600 nm (OD600nm). Sugars, ethanol, and organic acids were detected and quantified by HPLC (Shimadzu) equipped with an organic acid column (Phenomenex). HPLC was operated at 60°C using 5 mM H2SO4 as a mobile phase at 1 mL/min flow rate (5 μL injection volume, 15 min isocratic method). Metabolites were detected using both a refractive index detector and a UV detector (set to measure absorbance at 210 nm). The alginate oligomer mass spectrometry method uses the alginate oligomer HPLC method with a Shimadzu LCMS-2020 mass spectrometer. To detect the alginate oligomers, the mass spectrometer was run on DUIS mode with an interface temperature of 350°C, a desolvation line temperature of 200°C, and a heat block temperature of 250°C. The DUIS mode also had a nebulizing gas flow of 1.50 L/min and a drying gas flow of 15 L/min. The mass spectrometer detector was run in negative SIM mode. Oligomer molecular weight determination was based on previously reported values (45). For the alginate polymer, quantification was performed by degrading the residual oligomers to monomers followed by quantification via HPLC using a Thermo Hypercarb column and detection of UV absorbance at 235 nm. Fermentation of macroalgae in 1-L bioreactors
Saccharina japonica (kombu) was purchased from local grocery stores. We used a counter-top food blender to mill the macroalgae. However, due to the device’s limited volumetric capacity and mechanical force, several drying and milling steps were required to obtain a fine and uniform particle size. First, pre-dried macroalgae was chopped into approximately 10 cm x 10 cm. The macroalgae pieces were then placed on a metal tray and dried at ~105°C for 6 hr. These pieces were subsequently milled until the maximum particle size was less than ~5 mm in diameter. This batch was further dried at ~105°C for 2 hr, and milled again until the maximum particle size was less than ~1 mm in diameter. Prior to fermentation, the dried and finely milled macroalgae was autoclaved at 121°C for 20 min. The macroalgae fermentation medium consisted of 0.2 x M9 salts, 5 g/L yeast extract, 3 g/L citric acid monohydrate, 2 mM MgSO4, 200 μM CaCl2, 10 mg/L thiamine, 0.15 g/L CTec2 (Novozymes, Baagsvaerd, Denmark), 0.03 g/L HTec2 (Novozymes, Baagsvaerd, Denmark), 2.5mL/L trace metals supplement MD-TMS (ATCC, Manassas, VA), 2.5 mL/L vitamin supplement MD-VS (ATCC, Manassas, VA), 0.08 mL/L antifoam 204 (Sigma-Aldrich, St. Louis, MO), and milled macroalgae. Fed batch fermentations were carried out at the 1-L scale in a Sartorius Biostat APlus bioreactor. 50-mL LB pre-cultures were inoculated from a glycerol stock media and grown in shake flasks at 30°C for 10 hr. The initial fermentation broth contained about 50 g/L dried kombu. The fermentation broth was adjusted to a pH of 7 prior to addition of 50 mL pre-culture. Strain BAL1611 was allowed to grow aerobically for 5 hr by

supplying the reactor with air at the rate of 1 L/min at 30°C. After 5 hr the culture was maintained under anaerobic conditions at 25°C, and additional macroalgae was added during the first 28 hr of fermentation when the mannitol concentration fell below 10 g/L. 1.5-mL samples were collected at regular intervals. Throughout the course of fermentation, a pH of 7 was maintained by periodic addition of 5 N sodium hydroxide; the stir rate was maintained at 400 rpm.

Figure S1. The growth phenotype of E. coli harboring pALG1 with a single gene deletion on alginate pre-degraded using SM0524 Aly. Each deletion strain was grown in M9 salt medium containing 1% oligoalginate degraded with SM0524 Aly: (a) Negative control (pBeloBAC11), (b) positive control (pALG1), (c) V12B01_24189 (putative solute binding protein), (d) V12B01_24194 (ToaA), (e) V12B01_24214 (OalB), (f) V12B01_24219 (OalC), (g)V12B01_24224 (putative fumaryl-acetoacetate hydrolase), (h) V12B01_24229 (putative transcriptional repressor), (i) V12B01_24234 (ToaB), (j) V12B01_24239 (OalA), (k) V12B01_24244 (DehR), and (l) V12B01_24249 (putative signaling protein/chemoreceptor). An average of two individual growth curves is shown in each case. The difference between the two growth curves for each case and their average was within 3%. As we experienced significant growth disparity between the two individual growth curves for the V12B01_24219 strain, we additionally generated 4 more growth curves. These experiments suggested that lag time alters in the range of 24-48 hr. Thus, a single representative growth curve is shown in the figure.

Figure S2. The effect of deleting oalA (orange), oalB (light blue), and oalC (pink) in pALG1 on E. coli growth on M9 salt medium containing (a) 1% alginate pre-degraded with SM0524 Aly (reproduced from Fig. S1) and (b) 2 % alginate pre-degraded with Flavobacterium Aly (purchased from SIGMA). An average of two individual growth curves is shown in each case. The difference between the two growth curves and their average was within 3% except for oalA deletion (6%).

Figure S3. HPLC-MS analysis of residual alginate oligomers in in vitro reactions. Crude lysates of E. coli harboring pALG fosmids were incubated with alginate oligomers (alginate degraded with SM0524 Aly). E. coli expressing pALG1 (green), pALG1 (∆oalC;
orange), pALG1 (∆oalB; purple), pALG1 (∆oalA; red), and pBeloBac11 (blue). Residual population of (a, b) dimers, (c, d) trimers, (e, f) tetramers, and (g, h) pentamers. Figures S3b, d, f, and g highlight overlay between pBeloBac11 and pALG1(∆oalA).

Figure S4. Determination of co-factor specificity of DehR toward NADH (solid blue diamond) and NADPH (solid brown circles). Negative control of NADH (open blue diamonds) and NADPH (open brown circles) contains substrate and co-factor without DehR enzyme preparation.

Figure S5. The engineered secretable Aly system (N455+tSM0524 Aly) enabled growth on alginate as a sole carbon source through efficient degradation of alginate. (a) Aly activities from culture grown in LB medium overnight (before inoculation) and (b) growth curves of BAL1366 harboring pCCN455+tSM0524 Aly and the control strains in M9 salt medium containing 1% alginate. Standard deviation of the triplicate measurements is shown in error bars.
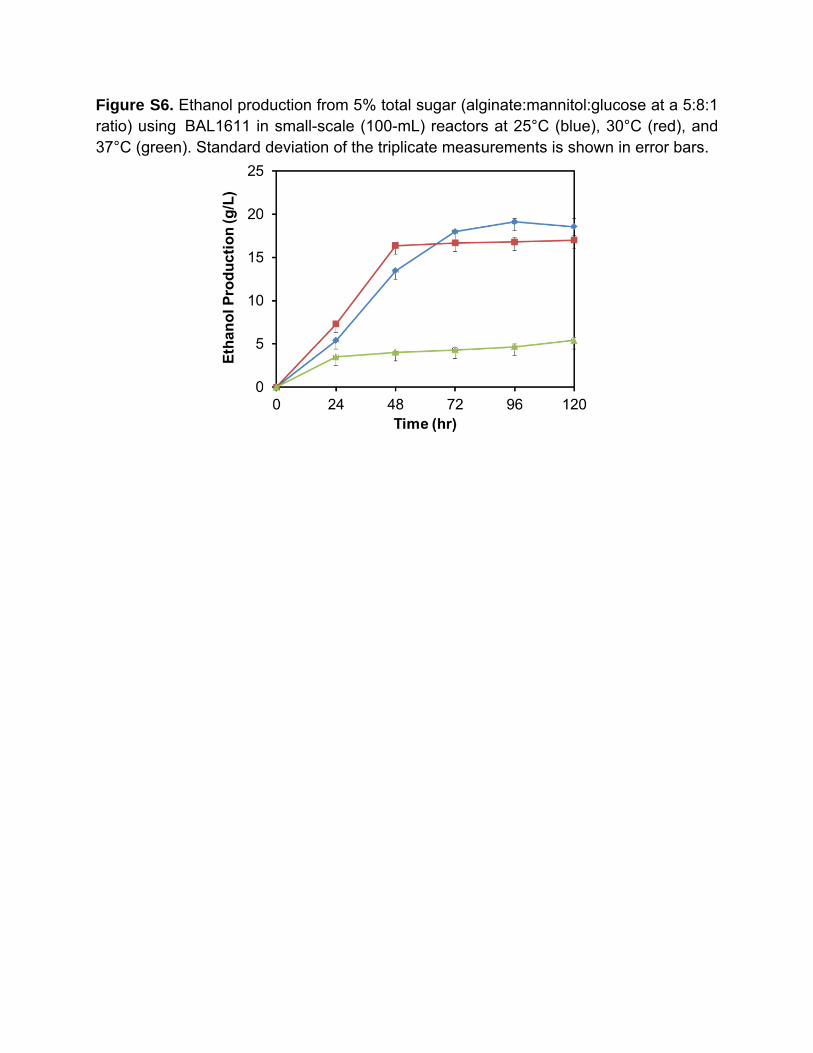
Figure S6. Ethanol production from 5% total sugar (alginate:mannitol:glucose at a 5:8:1 ratio) using BAL1611 in small-scale (100-mL) reactors at 25°C (blue), 30°C (red), and 37°C (green). Standard deviation of the triplicate measurements is shown in error bars.

Figure S7. Fermentation of macroalgae using E. coli without the engineered alginate assimilation pathway, BAL1303 (ldhA, focA-pflB, frdABCD::PG25-pdc-adhB). Ethanol production (blue), mannitol consumption (green), and glucose consumption (orange). Alginate remained unconsumed.

References 1. G. Stephanopoulos, Challenges in engineering microbes for biofuels production.
Science 315, 801 (2007). 2. G. Roesijadi, S. B. Jones, L. J. Snowden-Swan, Y. Zhu. (Prepared for the U.S.
Department of Energy under Contract DE-AC05-76RL01830 by Pacific Northwest National Laboratory, 2010).
3. C. Somerville, H. Youngs, C. Taylor, S. C. Davis, S. P. Long, Feedstocks for lignocellulosic biofuels. Science 329, 790 (2010).
4. J. M. Adams, J. a. Gallagher, I. S. Donnison, Fermentation study on Saccharina latissima for bioethanol production considering variable pre-treatments. Journal of Applied Phycology 21, 569 (2008).
5. I. M. A. S.J. Horn, and K. Ostgaard, Production of ethanol from mannitol by Zymobacter palmae. Journal of industrial microbiology & biotechnology 24, 51 (2000).
6. S. J. Horn, I. M. Aasen, K. Ostgaard, Ethanol production from seaweed extract. Journal of Industrial Microbiology and Biotechnology 25, 249 (2000).
7. K. I. Draget, O. Smidsrod, G. Skjak-Braek, in Polysaccharides and Polyamides in the Food Industry. Properties, Production, and Patents., A. Steinbuchel, S. K. Rhee, Eds. (Wiley-VCH Verlag GmbH & Co. KGaA, Weiheim, 2005).
8. T. Y. Wong, L. A. Preston, N. L. Schiller, ALGINATE LYASE: review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu Rev Microbiol 54, 289 (2000).
9. A. Ochiai, M. Yamasaki, B. Mikami, W. Hashimoto, K. Murata, Crystallization and preliminary X-ray analysis of an exotype alginate lyase Atu3025 from Agrobacterium tumefaciens strain C58, a member of polysaccharide lyase family 15. Acta Crystallogr Sect F Struct Biol Cryst Commun 62, 486 (2006).
10. J. Preiss, G. Ashwell, Alginic acid metabolism in bacteria. II. The enzymatic reduction of 4-deoxy-L-erythro-5-hexoseulose uronic acid to 2-keto-3-deoxy-D-gluconic acid. J Biol Chem 237, 317 (1962).
11. R. Takase, A. Ochiai, B. Mikami, W. Hashimoto, K. Murata, Molecular identification of unsaturated uronate reductase prerequisite for alginate metabolism in Sphingomonas sp. A1. Biochim Biophys Acta 1804, 1925 (2010).
12. H. Takeda, F. Yoneyama, S. Kawai, W. Hashimoto, K. Murata, Bioethanol production from marine biomass alginate by metabolically engineered bacteria. Energy & Environmental Science 4, 2575 (2011).
13. H. Alper, G. Stephanopoulos, Engineering for biofuels: exploiting innate microbial capacity or importing biosynthetic potential? Nat Rev Microbiol 7, 715 (2009).
14. S. Atsumi, T. Hanai, J. C. Liao, Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 451, 86 (2008).
15. B. B. Bond-Watts, R. J. Bellerose, M. C. Chang, Enzyme mechanism as a kinetic control element for designing synthetic biofuel pathways. Nat Chem Biol 7, 222 (2011).
16. C. Dellomonaco, J. M. Clomburg, E. N. Miller, R. Gonzalez, Engineered reversal of the beta-oxidation cycle for the synthesis of fuels and chemicals. Nature 476, 355 (2011).

17. K. Ohta, D. S. Beall, J. P. Mejia, K. T. Shanmugam, L. O. Ingram, Genetic improvement of Escherichia coli for ethanol production: chromosomal integration of Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase II. Appl Environ Microbiol 57, 893 (1991).
18. H. Y. Lee, C. J. Harvey, D. E. Cane, C. Khosla, Improved precursor-directed biosynthesis in E. coli via directed evolution. J Antibiot (Tokyo) 64, 59 (2011).
19. B. A. Pfeifer, S. J. Admiraal, H. Gramajo, D. E. Cane, C. Khosla, Biosynthesis of complex polyketides in a metabolically engineered strain of E. coli. Science 291, 1790 (2001).
20. P. K. Ajikumar et al., Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science 330, 70 (2010).
21. H. Alper, K. Miyaoku, G. Stephanopoulos, Construction of lycopene-overproducing E. coli strains by combining systematic and combinatorial gene knockout targets. Nat Biotechnol 23, 612 (2005).
22. E. Leonard et al., Strain improvement of recombinant Escherichia coli for efficient production of plant flavonoids. Mol Pharm 5, 257 (2008).
23. V. J. Martin, D. J. Pitera, S. T. Withers, J. D. Newman, J. D. Keasling, Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat Biotechnol 21, 796 (2003).
24. E. J. Steen et al., Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature 463, 559 (2010).
25. A. Schirmer, M. A. Rude, X. Li, E. Popova, S. B. del Cardayre, Microbial biosynthesis of alkanes. Science 329, 559 (2010).
26. F. A. Skraly, B. L. Lytle, D. C. Cameron, Construction and characterization of a 1,3-propanediol operon. Appl Environ Microbiol 64, 98 (1998).
27. H. Yim et al., Metabolic engineering of Escherichia coli for direct production of 1,4-butanediol. Nature chemical biology 7, 445 (2011).
28. L. R. Lynd, W. H. van Zyl, J. E. McBride, M. Laser, Consolidated bioprocessing of cellulosic biomass: an update. Curr Opin Biotechnol 16, 577 (2005).
29. J. W. Li et al., Purification and characterization of a bifunctional alginate lyase from Pseudoalteromonas sp. SM0524. Mar Drugs 9, 109 (2011).
30. S. Y. He, M. Lindeberg, A. K. Chatterjee, A. Collmer, Cloned Erwinia chrysanthemi out genes enable Escherichia coli to selectively secrete a diverse family of heterologous proteins to its milieu. Proc Natl Acad Sci U S A 88, 1079 (1991).
31. N. Kazemi-Pour, G. Condemine, N. Hugouvieux-Cotte-Pattat, The secretome of the plant pathogenic bacterium Erwinia chrysanthemi. Proteomics 4, 3177 (2004).
32. K. Fujiyama, H. Maki, S. Kinoshita, T. Yoshida, Purification and characterization of the recombinant alginate lyase from Pseudomonas sp. leaked by Escherichia coli upon addition of glycine. FEMS Microbiol Lett 126, 19 (1995).
33. P. Klemm, L. Hjerrild, M. Gjermansen, M. A. Schembri, Structure-function analysis of the self-recognizing Antigen 43 autotransporter protein from Escherichia coli. Mol Microbiol 51, 283 (2004).
34. M. W. van der Woude, I. R. Henderson, Regulation and function of Ag43 (flu). Annu Rev Microbiol 62, 153 (2008).

35. I. R. Henderson, P. Owen, The major phase-variable outer membrane protein of Escherichia coli structurally resembles the immunoglobulin A1 protease class of exported protein and is regulated by a novel mechanism involving Dam and oxyR. J Bacteriol 181, 2132 (1999).
36. K. Kjaergaard, H. Hasman, M. A. Schembri, P. Klemm, Antigen 43-mediated autotransporter display, a versatile bacterial cell surface presentation system. J Bacteriol 184, 4197 (2002).
37. W. Hashimoto, S. Kawai, K. Murata, Bacterial supersystem for alginate import/metabolism and its environmental and bioenergy applications. Bioeng Bugs 1, 97 (2010).
38. N. Hugouvieux-Cotte-Pattat, S. Reverchon, Two transporters, TogT and TogMNAB, are responsible for oligogalacturonide uptake in Erwinia chrysanthemi 3937. Mol Microbiol 41, 1125 (2001).
39. W. Hashimoto, O. Miyake, K. Momma, S. Kawai, K. Murata, Molecular identification of oligoalginate lyase of Sphingomonas sp. strain A1 as one of the enzymes required for complete depolymerization of alginate. J Bacteriol 182, 4572 (2000).
40. A. Ochiai, W. Hashimoto, K. Murata, A biosystem for alginate metabolism in Agrobacterium tumefaciens strain C58: molecular identification of Atu3025 as an exotype family PL-15 alginate lyase. Res Microbiol 157, 642 (2006).
41. N. Blot, C. Berrier, N. Hugouvieux-Cotte-Pattat, A. Ghazi, G. Condemine, The oligogalacturonate-specific porin KdgM of Erwinia chrysanthemi belongs to a new porin family. J Biol Chem 277, 7936 (2002).
42. M. W. Lau, B. E. Dale, Cellulosic ethanol production from AFEX-treated corn stover using Saccharomyces cerevisiae 424A(LNH-ST). Proc Natl Acad Sci U S A 106, 1368 (2009).
43. L. P. Yomano, S. W. York, K. T. Shanmugam, L. O. Ingram, Deletion of methylglyoxal synthase gene (mgsA) increased sugar co-metabolism in ethanol-producing Escherichia coli. Biotechnology letters 31, 1389 (2009).
44. K. A. Datsenko, B. L. Wanner, One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proceedings of the National Academy of Sciences of the United States of America 97, 6640 (2000).
45. Z. Zhang et al., Preparation and structure elucidation of alginate oligosaccharides degraded by alginate lyase from Vibro sp. 510. Carbohydr Res 339, 1475 (2004).

References and Notes 1. G. Stephanopoulos, Challenges in engineering microbes for biofuels production. Science 315,
801 (2007). doi:10.1126/science.1139612 Medline
2. G. Roesijadi, S. B. Jones, L. J. Snowden-Swan, Y. Zhu, “Macroalgae as a Biomass Feedstock: A Preliminary Analysis,” prepared for the U.S. Department of Energy under contract DE-AC05-76RL01830 by Pacific Northwest National Laboratory (2010).
3. C. Somerville, H. Youngs, C. Taylor, S. C. Davis, S. P. Long, Feedstocks for lignocellulosic biofuels. Science 329, 790 (2010). doi:10.1126/science.1189268 Medline
4. J. M. Adams, J. Gallagher, I. S. Donnison, Fermentation study on Saccharina latissima for bioethanol production considering variable pre-treatments. J. Appl. Phycol. 21, 569 (2008). doi:10.1007/s10811-008-9384-7
5. I. M. A. S. J. Horn, K. Ostgaard, Production of ethanol from mannitol by Zymobacter palmae. J. Ind. Microbiol. Biotechnol. 24, 51 (2000). doi:10.1038/sj.jim.2900771
6. S. J. Horn, I. M. Aasen, K. Ostgaard, Ethanol production from seaweed extract. J. Ind. Microbiol. Biotechnol. 25, 249 (2000). doi:10.1038/sj.jim.7000065
7. K. I. Draget, O. Smidsrod, G. Skjak-Braek, in Polysaccharides and Polyamides in the Food Industry. Properties, Production, and Patents, A. Steinbuchel, S. K. Rhee, Eds. (Wiley–VCH Verlag GmbH KGaA, Weiheim, 2005), pp. 1–30.
8. T. Y. Wong, L. A. Preston, N. L. Schiller, Alginate lyase: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 54, 289 (2000). doi:10.1146/annurev.micro.54.1.289 Medline
9. A. Ochiai, M. Yamasaki, B. Mikami, W. Hashimoto, K. Murata, Crystallization and preliminary x-ray analysis of an exotype alginate lyase Atu3025 from Agrobacterium tumefaciens strain C58, a member of polysaccharide lyase family 15. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 62, 486 (2006). doi:10.1107/S1744309106014333 Medline
10. J. Preiss, G. Ashwell, Alginic acid metabolism in bacteria. II. The enzymatic reduction of 4-deoxy-L-erythro-5-hexoseulose uronic acid to 2-keto-3-deoxy-D-gluconic acid. J. Biol. Chem. 237, 317 (1962). Medline
11. R. Takase, A. Ochiai, B. Mikami, W. Hashimoto, K. Murata, Molecular identification of unsaturated uronate reductase prerequisite for alginate metabolism in Sphingomonas sp. A1. Biochim. Biophys. Acta 1804, 1925 (2010). Medline
12. H. Takeda, F. Yoneyama, S. Kawai, W. Hashimoto, K. Murata, Bioethanol production from marine biomass alginate by metabolically engineered bacteria. Energy Environ. Sci. 4, 2575 (2011). doi:10.1039/c1ee01236c
13. H. Alper, G. Stephanopoulos, Engineering for biofuels: Exploiting innate microbial capacity or importing biosynthetic potential? Nat. Rev. Microbiol. 7, 715 (2009). doi:10.1038/nrmicro2186 Medline
14. S. Atsumi, T. Hanai, J. C. Liao, Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 451, 86 (2008). doi:10.1038/nature06450 Medline

15. B. B. Bond-Watts, R. J. Bellerose, M. C. Chang, Enzyme mechanism as a kinetic control element for designing synthetic biofuel pathways. Nat. Chem. Biol. 7, 222 (2011). doi:10.1038/nchembio.537 Medline
16. C. Dellomonaco, J. M. Clomburg, E. N. Miller, R. Gonzalez, Engineered reversal of the β-oxidation cycle for the synthesis of fuels and chemicals. Nature 476, 355 (2011). doi:10.1038/nature10333 Medline
17. K. Ohta, D. S. Beall, J. P. Mejia, K. T. Shanmugam, L. O. Ingram, Genetic improvement of Escherichia coli for ethanol production: Chromosomal integration of Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase II. Appl. Environ. Microbiol. 57, 893 (1991). Medline
18. H. Y. Lee, C. J. Harvey, D. E. Cane, C. Khosla, Improved precursor-directed biosynthesis in E. coli via directed evolution. J. Antibiot. (Tokyo) 64, 59 (2011). doi:10.1038/ja.2010.129 Medline
19. B. A. Pfeifer, S. J. Admiraal, H. Gramajo, D. E. Cane, C. Khosla, Biosynthesis of complex polyketides in a metabolically engineered strain of E. coli. Science 291, 1790 (2001). doi:10.1126/science.1058092 Medline
20. P. K. Ajikumar et al., Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science 330, 70 (2010). doi:10.1126/science.1191652 Medline
21. H. Alper, K. Miyaoku, G. Stephanopoulos, Construction of lycopene-overproducing E. coli strains by combining systematic and combinatorial gene knockout targets. Nat. Biotechnol. 23, 612 (2005). doi:10.1038/nbt1083 Medline
22. E. Leonard et al., Strain improvement of recombinant Escherichia coli for efficient production of plant flavonoids. Mol. Pharm. 5, 257 (2008). doi:10.1021/mp7001472 Medline
23. V. J. Martin, D. J. Pitera, S. T. Withers, J. D. Newman, J. D. Keasling, Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 21, 796 (2003). doi:10.1038/nbt833 Medline
24. E. J. Steen et al., Microbial production of fatty-acid–derived fuels and chemicals from plant biomass. Nature 463, 559 (2010). doi:10.1038/nature08721 Medline
25. A. Schirmer, M. A. Rude, X. Li, E. Popova, S. B. del Cardayre, Microbial biosynthesis of alkanes. Science 329, 559 (2010). doi:10.1126/science.1187936 Medline
26. F. A. Skraly, B. L. Lytle, D. C. Cameron, Construction and characterization of a 1,3-propanediol operon. Appl. Environ. Microbiol. 64, 98 (1998). Medline
27. H. Yim et al., Metabolic engineering of Escherichia coli for direct production of 1,4-butanediol. Nat. Chem. Biol. 7, 445 (2011). doi:10.1038/nchembio.580 Medline
28. L. R. Lynd, W. H. van Zyl, J. E. McBride, M. Laser, Consolidated bioprocessing of cellulosic biomass: An update. Curr. Opin. Biotechnol. 16, 577 (2005). doi:10.1016/j.copbio.2005.08.009 Medline

29. J. W. Li et al., Purification and characterization of a bifunctional alginate lyase from Pseudoalteromonas sp. SM0524. Mar. Drugs 9, 109 (2011). doi:10.3390/md9010109 Medline
30. S. Y. He, M. Lindeberg, A. K. Chatterjee, A. Collmer, Cloned Erwinia chrysanthemi out genes enable Escherichia coli to selectively secrete a diverse family of heterologous proteins to its milieu. Proc. Natl. Acad. Sci. U.S.A. 88, 1079 (1991). doi:10.1073/pnas.88.3.1079 Medline
31. N. Kazemi-Pour, G. Condemine, N. Hugouvieux-Cotte-Pattat, The secretome of the plant pathogenic bacterium Erwinia chrysanthemi. Proteomics 4, 3177 (2004). doi:10.1002/pmic.200300814 Medline
32. K. Fujiyama, H. Maki, S. Kinoshita, T. Yoshida, Purification and characterization of the recombinant alginate lyase from Pseudomonas sp. leaked by Escherichia coli upon addition of glycine. FEMS Microbiol. Lett. 126, 19 (1995). doi:10.1111/j.1574-6968.1995.tb07384.x Medline
33. P. Klemm, L. Hjerrild, M. Gjermansen, M. A. Schembri, Structure-function analysis of the self-recognizing Antigen 43 autotransporter protein from Escherichia coli. Mol. Microbiol. 51, 283 (2004). doi:10.1046/j.1365-2958.2003.03833.x Medline
34. M. W. van der Woude, I. R. Henderson, Regulation and function of Ag43 (flu). Annu. Rev. Microbiol. 62, 153 (2008). doi:10.1146/annurev.micro.62.081307.162938 Medline
35. I. R. Henderson, P. Owen, The major phase-variable outer membrane protein of Escherichia coli structurally resembles the immunoglobulin A1 protease class of exported protein and is regulated by a novel mechanism involving Dam and oxyR. J. Bacteriol. 181, 2132 (1999). Medline
36. K. Kjaergaard, H. Hasman, M. A. Schembri, P. Klemm, Antigen 43-mediated autotransporter display, a versatile bacterial cell surface presentation system. J. Bacteriol. 184, 4197 (2002). doi:10.1128/JB.184.15.4197-4204.2002 Medline
37. W. Hashimoto, S. Kawai, K. Murata, Bacterial supersystem for alginate import/metabolism and its environmental and bioenergy applications. Bioeng. Bugs 1, 97 (2010). doi:10.4161/bbug.1.2.10322 Medline
38. N. Hugouvieux-Cotte-Pattat, S. Reverchon, Two transporters, TogT and TogMNAB, are responsible for oligogalacturonide uptake in Erwinia chrysanthemi 3937. Mol. Microbiol. 41, 1125 (2001). doi:10.1046/j.1365-2958.2001.02565.x Medline
39. W. Hashimoto, O. Miyake, K. Momma, S. Kawai, K. Murata, Molecular identification of oligoalginate lyase of Sphingomonas sp. strain A1 as one of the enzymes required for complete depolymerization of alginate. J. Bacteriol. 182, 4572 (2000). doi:10.1128/JB.182.16.4572-4577.2000 Medline
40. A. Ochiai, W. Hashimoto, K. Murata, A biosystem for alginate metabolism in Agrobacterium tumefaciens strain C58: Molecular identification of Atu3025 as an exotype family PL-15 alginate lyase. Res. Microbiol. 157, 642 (2006). doi:10.1016/j.resmic.2006.02.006 Medline

41. N. Blot, C. Berrier, N. Hugouvieux-Cotte-Pattat, A. Ghazi, G. Condemine, The oligogalacturonate-specific porin KdgM of Erwinia chrysanthemi belongs to a new porin family. J. Biol. Chem. 277, 7936 (2002). doi:10.1074/jbc.M109193200 Medline
42. M. W. Lau, B. E. Dale, Cellulosic ethanol production from AFEX-treated corn stover using Saccharomyces cerevisiae 424A(LNH-ST). Proc. Natl. Acad. Sci. U.S.A. 106, 1368 (2009). doi:10.1073/pnas.0812364106 Medline
43. L. P. Yomano, S. W. York, K. T. Shanmugam, L. O. Ingram, Deletion of methylglyoxal synthase gene (mgsA) increased sugar co-metabolism in ethanol-producing Escherichia coli. Biotechnol. Lett. 31, 1389 (2009). doi:10.1007/s10529-009-0011-8 Medline
44. K. A. Datsenko, B. L. Wanner, One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A. 97, 6640 (2000). doi:10.1073/pnas.120163297 Medline
45. Z. Zhang et al., Preparation and structure elucidation of alginate oligosaccharides degraded by alginate lyase from Vibro sp. 510. Carbohydr. Res. 339, 1475 (2004). doi:10.1016/j.carres.2004.03.010 Medline