analysis of the expression of retinoic acid metabolising genes during xenopus laevis organogenesis
TRANSCRIPT

Gene Expression Patterns 11 (2011) 112–117
Contents lists available at ScienceDirect
Gene Expression Patterns
journal homepage: www.elsevier .com/locate /gep
Analysis of the expression of retinoic acid metabolising genes duringXenopus laevis organogenesis
Joshua Lynch a, James McEwan a, Caroline W. Beck a,b,⇑a Department of Zoology, University of Otago, 340 Great King Street, Dunedin, New Zealandb Genetics Otago, University of Otago, P.O. Box 56, Dunedin, New Zealand
a r t i c l e i n f o
Article history:Received 13 May 2010Received in revised form 12 October 2010Accepted 15 October 2010Available online 20 October 2010
Keywords:Xenopus laevisRetinoic acidOrganogenesisEyeSomitesHeartTeratogen
1567-133X/$ - see front matter � 2010 Elsevier B.V.doi:10.1016/j.gep.2010.10.003
⇑ Corresponding author at: Department of ZoologGreat King Street, Dunedin, New Zealand. Tel.: +64 (4797584.
E-mail address: [email protected] (C.W. B
a b s t r a c t
Retinoic acid (RA) is a known teratogen that is also required endogenously for normal development of theembryo. RA can act as a morphogen, through direct binding to receptors and RA response elements in thegenome, and classical studies of limb development and regeneration in amphibians have shown that it islikely to provide positional information. Availability of RA depends on both metabolic synthesis and cat-abolic degradation, and specific binding proteins act to further modulate the binding of RA to responseelements. Here, we describe the expression of seven genes involved in metabolism (Raldh1-3), catabolism(Cyp26a and b) and binding of RA (Crabp1 and 2) during organogenesis in the clawed frog Xenopus laevis.Taken together, this data indicates regions of the embryo that could be affected by RA mediated pattern-ing, and identifies some differences with other vertebrates.
� 2010 Elsevier B.V. All rights reserved.
Retinoic acid (RA) is a metabolite of vitamin A (retinol) that hasthe ability to act as a morphogen during development (Duester,2007; Durston et al., 1989; Guthrie, 1996; Lee et al., 2004). RA pas-sively diffuses into cells where it can bind to retinoid receptors andtranslocate directly to the nucleus as part of a complex that bindsretinoic acid response elements, controlling gene transcription (re-viewed recently in Niederreither and Dolle, 2008). Levels of RA arecontrolled by production, catabolism and binding (reviewed inBlomhoff and Blomhoff, 2006). The rate-limiting step in synthesisof RA is controlled by a group of retinal dehydrogenases (Raldh1-3, also known as aldehyde dehydrogenase 1a1-3 or Aldh1a1-3).Binding to factors such as cellular retinoic acid binding proteins1 and 2 (Crabp1 and 2) further refines the level of available RA,and members of the Cyp26 family of cytochrome p450 hydroxy-lases actively degrade RA. Direct measurement of RA has shownthat regions of the developing mouse embryo that express Raldh2are actively synthesising RA, whereas regions expressing Cyp26are actively degrading it (Swindell et al., 1999). Consequently,the interplay of these enzymes during development determineshow much RA is present in different cells of the embryo generatingthe potential to alter gene expression (Reijntjes et al., 2005).
All rights reserved.
y, University of Otago, 3400) 3 4974109; fax: +64 (0) 3
eck).
Here, we present the expression patterns of RA modulatinggenes in embryos of the clawed frog Xenopus laevis (X. laevis)undergoing organogenesis. As well as RA, there are other activeretinoids in Xenopus, for example, 4-oxo RA and its metabolite9-cis-4-oxo retinoic acid (Pijnappel et al., 1998, 1993). Therefore,we acknowledge that this is not a complete description of RAmetabolism in embryos, rather, we focus on the rate limiting stepsthat are most likely to impact ligand availability and hencedevelopmental fate.
The expression patterns support a role for RA in patterning thehindbrain, eye, otic vesicle, olfactory placodes, tail bud, proneph-ros, lateral plate mesoderm and branchial arches. Surprisingly,our results show a lack of RA catabolism in the heart and of synthe-sis in the tail presomitic mesoderm in Xenopus, two key differencesfrom amniotes.
1. Results
1.1. Cloning of Cyp26a1, Cyp26b1 and Crabp1 from X. laevis
Full coding sequence was obtained for Cyp26a1, and was foundto match previously known Cyp26 sequences from X. laevis. Our se-quence was 100% identical to these over 465 amino acids and is re-ferred to herein as Cyp26a.
A partial coding sequence for Cyp26b1 was obtained using pri-mer sequences based on the Xenopus tropicalis sequence. X. laevis
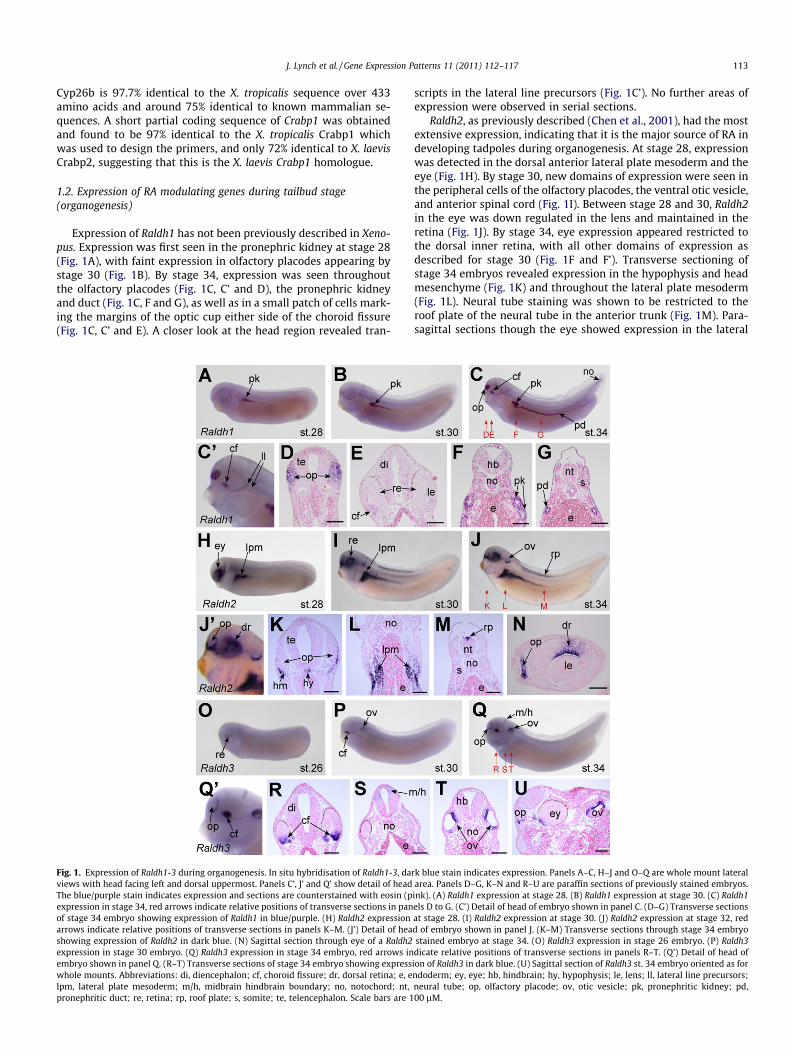
J. Lynch et al. / Gene Expression Patterns 11 (2011) 112–117 113
Cyp26b is 97.7% identical to the X. tropicalis sequence over 433amino acids and around 75% identical to known mammalian se-quences. A short partial coding sequence of Crabp1 was obtainedand found to be 97% identical to the X. tropicalis Crabp1 whichwas used to design the primers, and only 72% identical to X. laevisCrabp2, suggesting that this is the X. laevis Crabp1 homologue.
1.2. Expression of RA modulating genes during tailbud stage(organogenesis)
Expression of Raldh1 has not been previously described in Xeno-pus. Expression was first seen in the pronephric kidney at stage 28(Fig. 1A), with faint expression in olfactory placodes appearing bystage 30 (Fig. 1B). By stage 34, expression was seen throughoutthe olfactory placodes (Fig. 1C, C’ and D), the pronephric kidneyand duct (Fig. 1C, F and G), as well as in a small patch of cells mark-ing the margins of the optic cup either side of the choroid fissure(Fig. 1C, C’ and E). A closer look at the head region revealed tran-
Fig. 1. Expression of Raldh1-3 during organogenesis. In situ hybridisation of Raldh1-3, daviews with head facing left and dorsal uppermost. Panels C’, J’ and Q’ show detail of headThe blue/purple stain indicates expression and sections are counterstained with eosin (pexpression in stage 34, red arrows indicate relative positions of transverse sections in panof stage 34 embryo showing expression of Raldh1 in blue/purple. (H) Raldh2 expressionarrows indicate relative positions of transverse sections in panels K–M. (J’) Detail of heashowing expression of Raldh2 in dark blue. (N) Sagittal section through eye of a Raldh2expression in stage 30 embryo. (Q) Raldh3 expression in stage 34 embryo, red arrows inembryo shown in panel Q. (R–T) Transverse sections of stage 34 embryo showing expresswhole mounts. Abbreviations: di, diencephalon; cf, choroid fissure; dr, dorsal retina; e, elpm, lateral plate mesoderm; m/h, midbrain hindbrain boundary; no, notochord; nt,pronephritic duct; re, retina; rp, roof plate; s, somite; te, telencephalon. Scale bars are 1
scripts in the lateral line precursors (Fig. 1C’). No further areas ofexpression were observed in serial sections.
Raldh2, as previously described (Chen et al., 2001), had the mostextensive expression, indicating that it is the major source of RA indeveloping tadpoles during organogenesis. At stage 28, expressionwas detected in the dorsal anterior lateral plate mesoderm and theeye (Fig. 1H). By stage 30, new domains of expression were seen inthe peripheral cells of the olfactory placodes, the ventral otic vesicle,and anterior spinal cord (Fig. 1I). Between stage 28 and 30, Raldh2in the eye was down regulated in the lens and maintained in theretina (Fig. 1J). By stage 34, eye expression appeared restricted tothe dorsal inner retina, with all other domains of expression asdescribed for stage 30 (Fig. 1F and F’). Transverse sectioning ofstage 34 embryos revealed expression in the hypophysis and headmesenchyme (Fig. 1K) and throughout the lateral plate mesoderm(Fig. 1L). Neural tube staining was shown to be restricted to theroof plate of the neural tube in the anterior trunk (Fig. 1M). Para-sagittal sections though the eye showed expression in the lateral
rk blue stain indicates expression. Panels A–C, H–J and O–Q are whole mount lateralarea. Panels D–G, K–N and R–U are paraffin sections of previously stained embryos.ink). (A) Raldh1 expression at stage 28. (B) Raldh1 expression at stage 30. (C) Raldh1els D to G. (C’) Detail of head of embryo shown in panel C. (D–G) Transverse sectionsat stage 28. (I) Raldh2 expression at stage 30. (J) Raldh2 expression at stage 32, redd of embryo shown in panel J. (K–M) Transverse sections through stage 34 embryostained embryo at stage 34. (O) Raldh3 expression in stage 26 embryo. (P) Raldh3dicate relative positions of transverse sections in panels R–T. (Q’) Detail of head ofion of Raldh3 in dark blue. (U) Sagittal section of Raldh3 st. 34 embryo oriented as forndoderm; ey, eye; hb, hindbrain; hy, hypophysis; le, lens; ll, lateral line precursors;neural tube; op, olfactory placode; ov, otic vesicle; pk, pronephritic kidney; pd,00 lM.

114 J. Lynch et al. / Gene Expression Patterns 11 (2011) 112–117
edges of the olfactory placodes and restricted expression of Raldh2to the inner dorsal retina (Fig. 1N).
Raldh3 expression was essentially as described previously instage 33 embryos (Lupo et al., 2005). At stage 26, expression ap-peared restricted to a small patch of cells in the ventral retina ofthe eye, flanking the choroid fissure (Fig. 1O). By stage 30, tran-scripts were also detected in the dorsal otic vesicle (Fig. 1P) com-plementary to Raldh2 expression. By stage 34, Raldh3 wasexpressed in the peripheral cells of the olfactory placodes as wellas at the midbrain hindbrain boundary, with retinal and otic vesi-cle expression being maintained (Fig. 1Q and Q’). Sectioning ofstage 34 embryos showed that Raldh3 was being expressedthroughout the ventral-most retina cells (Fig. 1R) and the dorsalhalf of the neural tube at the midbrain hindbrain boundary(Fig. 1S). Otic vesicles showed expression in the dorsal and, to a les-ser extent, ventral-most portions, with the part of the otic vesicleadjacent to the notochord being free of Raldh3 expression(Fig. 1T). Parasagittal sectioning indicated restriction of Raldh3 tothe lateral edges of the olfactory placodes (Fig. 1U).
Cyp26a expression has been previously described (Chen et al.,2001; Hollemann et al., 1998; Tanibe et al., 2008), and this corre-lates well with our own results. Expression was already quiteextensive at stage 28, with very strong expression throughoutthe tailbud as well as in endothelial cells that appear to be assem-bling into intersomitic blood vessels. Punctate expression was alsoseen in branchial arches and lateral plate mesoderm, especiallydorso-anteriorly, and to a lesser extent in the epidermis (Fig. 2A).This expression was maintained at stage 30 (Fig. 2B) with the addi-tion of a new domain in the forming lens of the eye (Fig. 2B and C).Tailbud expression was relatively diminished at stage 34, with allother domains maintained from earlier stages (Fig. 2C). Parasagittal
Fig. 2. Expression of Cyp26a and b during organogenesis. In situ hybridisation of Cyp26Cyp26b. Panels A–C and G–I are whole mount whole mount lateral views with head facinCyp26a at stage 30. (C) Expression of Cyp26a at stage 34. (B’) Close-up view of intersopreviously stained stage 34 embryo showing Cyp26a expression in dark blue and with eSection through eye. (E) Deeper section (more towards midline) through head area. (F) Seexpression in stage 30 embryo. (I) Cyp26b expression in stage 34 embryo. Red arrows indistaining in panel H. (J) Sagittal section through stage 34 embryo showing expression in nu(J). (K) Transverse section at the level of the otic vesicles showing hindbrain staining localevel of the heart showing staining in ventral aortae. In K and L, dorsal is uppermost. Aendoderm; he, heart; isv, intersomitic vessels; le, lens; lpm, lateral plate mesoderm; ov
sections showed that transcripts were restricted to the outer partof the lens and that transcripts were not absent from all retinalcells (Fig. 2D). Expression was also detected in the ventral edgesof the olfactory placodes (Fig. 2E) and confirmed in the intersomiticvessels (Fig. 2F).
Cyp26b has not been described in Xenopus, and in contrast toCyp26a, was specifically expressed only in the hindbrain, withexpression restricted to rhombomeres 1, 4 and 6/7 (Fig. 2G–I). Sec-tioning confirmed the rhombomeric expression (Fig. 2J) and alsorevealed the presence of Cyp26b transcripts in tissue either sideof the developing heart and on the dorsal edge of the cardiac coe-lom. These cells correspond to the Flk1 positive ventral aortae(Cleaver et al., 1997). To our knowledge, this is the first descriptionof Cyp26 expression in the ventral aortae.
Crabp2, which is thought to assist the loading of RA onto recep-tors, was also expressed extensively during organogenesis, as pre-viously shown (Dekker et al., 1994). At stage 28, expression wasseen in the posterior hindbrain, midbrain/hindbrain boundary,branchial arches, and tailbud (Fig. 3A). Additional expression ofCrabp2 was seen in ventral and lateral edges of the olfactory pla-codes as well as in the dorsal retina and dorsal otic vesicle at stage30 and 34 (Fig. 3B, C and C’). Transverse sections of stage 34 em-bryos revealed head mesenchyme expression but an absence oftranscripts in the hypophysis (Fig. 3D). In the eye, Crabp2 expres-sion was limited to dorsal retina (Fig. 3E), and transcripts werepresent throughout the posterior hindbrain (rhombomeres 3–7),but absent from floor and roof plates (Fig. 3F and G).
No convincing tissue specific expression of Crabp1, which mayhave an inhibitory role (Boylan and Gudas, 1992; Budhu andNoy, 2002), was seen between stages 28 and 34 in Xenopus (datanot shown).
a and b, dark blue stain indicates expression. Panels A–F are Cyp26a and G–L areg left and dorsal uppermost. (A) Expression of Cyp26a at stage 28. (B) Expression of
mitic staining in embryo shown in panel B. (D–F) 12 lm sagittal sections throughosin (pink) used as a counterstain. Heads are to the left and dorsal uppermost. (D)ction through trunk somites. (G) Cyp26b expression in stage 28 embryo. (H) Cyp26b
cate the level of sections used in panels K and L. (H’) Closer detail of the rhombomerembered rhombomeres of the hindbrain. Anterior is to the left and dorsal uppermost
lised towards the lumenal side. (L) Transverse section through ventral embryo at thell scale bars are100 lm. Abbreviations: ba, branchial arches; cf, choroid fissure; e,
, otic vesicle; r, rhombomere; re, retina; s, somite; tb, tailbud; va, ventral aortae.

Fig. 3. Expression of Crabp2 during organogenesis. In situ hybridisation of Crabp2, dark blue stain indicates expression. (A–C) whole mount embryos with anterior to the leftand dorsal uppermost. (A) Expression at stage 28. (B) Expression at stage 30. (C) Expression at stage 34. Red arrows indicate positions of transverse sections in panels D–F. (C’)Detail of head region of embryo pictured in panel C. (D–F) Transverse sections through stage 34 embryo. Scale bars are 100 lm. (D) Staining in head mesenchyme ventral toolfactory placodes, at the level of the telencephalon. (E) Staining in branchial arches and dorsal retina at eye level. (F) Staining in the hindbrain is strongest at the lumenalsurfaces. (G) Sagittal section though stage 34 embryo showing staining in branchial arches, head mesenchyme, midbrain/hindbrain boundary region and posterior hindbrain.Scale bar is 100 lm. Abbreviations: ba, branchial arches; dr, dorsal retina; hb, hindbrain; hm, head mesenchyme; le, lens; m/h, midbrain hindbrain boundary; no, notochord;op, olfactory placode; ov, otic vesicle; pd, pronephritic duct; s, somite; tb, tailbud; te, telencephalon.
J. Lynch et al. / Gene Expression Patterns 11 (2011) 112–117 115
1.3. Discussion
The results presented here suggest that development of the kid-ney, lateral plate mesoderm and otic vesicle of Xenopus depend onRA modulation in a similar way than has been observed in miceand therefore these organs will not be further discussed. Instead,we will discuss some of the differences and similarities in RA reg-ulation predicted by the observed expression of RA modifyinggenes in the heart, eye, hindbrain, blood vessels, tailbud and som-ites. A summary diagram of the expression of RA synthesising anddegrading enzymes in the Xenopus tailbud stage embryo is shownin Fig. 4.
1.3.1. Cyp26 gene expression in Xenopus highlights some differenceswith amniotes
In mouse and chick embryos, Cyp26 genes are expressed in thedeveloping heart (Cyp26b in chicken, Cyp26a in mouse) (Abu-Abedet al., 2002; MacLean et al., 2001; Reijntjes et al., 2003). In contrast,Xenopus embryos do not express either Cyp26 gene in the region ofthe heart, suggesting that RA degradation is not important for heartdevelopment in Xenopus. In amniotes, RA reaches the heart fieldfrom the nearby lateral plate mesoderm (LPM): mice lackingRaldh2 have defective heart looping because the LPM fails to makeany RA (Niederreither et al., 2001). Despite the lack of Cyp26 geneexpression in the heart tube of Xenopus, Raldh2 expression in LPMindicates a role for RA in heart development in non-amniote verte-brates, albeit differently regulated. What is striking, though, is thevery specific and restricted expression of Cyp26b in the paired ven-tral aortae, suggesting a need to protect these cells from RA.
In other forming organs, the need for RA degrading enzymes ina particular location of the developing embryo is conservedbetween amniotes and amphibians, but the Cyp26 gene responsibleis different: for example, intersomitic vessels are marked byCyp26b expression in mice (MacLean et al., 2001), and chick
Fig. 4. Predicted RA metabolism vs. catabolism sites in Xenopus organogenesis. Summaryand degradation (red) as predicted by gene expression. Abbreviations as for previous fig
(Reijntjes et al., 2003) but by Cyp26a in Xenopus. The reciprocal sit-uation occurs in the hindbrain, where it is Cyp26b rather thanCyp26a that is expressed in Xenopus rhombomeres. Cyp26b is alsoexpressed in the hindbrain in zebrafish (Spoorendonk et al.,2008). Crabp1b expression was also detected in the posterior hind-brain in zebrafish (Liu et al., 2005) and in mice (Ruberte et al.,1992), mirroring the expression described here for Xenopus Crabp2.The source of RA in the Xenopus hindbrain is likely to be a combi-nation of the midbrain/hindbrain boundary expression of Raldh3and the spinal cord expression of Raldh2. The otic vesicle, locatedventrolaterally to rhombomeres 3 and 4, is also a potential sourceof RA, from Raldh2 and Raldh3.
1.3.2. The expression of RA modifying genes in eye developmentFig. 4 shows that RA metabolism is central to eye development
in Xenopus, as it is in other vertebrates. However, when we look atindividual genes there are some differences. In mouse, Raldh1 is ex-pressed in the dorsal retina (Haselbeck et al., 1999; McCafferyet al., 1991), whereas in Xenopus it is expressed in the ventral ret-ina in two groups of juxtaposed cells lining the choroid fissure, thesite of the optic vesicle closure. Raldh3 is expressed in these samecells in both mouse (Mic et al., 2000) and Xenopus. The expressionof Cyp26a seen in the developing Xenopus lens (Hollemann et al.,1998 and this study) has not been reported in amniotes, and is re-ciprocal to the expression of RA synthesising genes. RA has beenassociated with lens regeneration in Xenopus and in newts(Malloch et al., 2009; Tsonis et al., 2000) and differences in howRA is regulated may define the different regenerative strategiesused by these two amphibians.
1.3.3. Expression of RA modifying genes in posterior developmentIn the tailbud, Cyp26a expression is conserved between mice
and frogs and the functional importance of this has been demon-strated by mouse knockouts, which show caudal truncation
diagram of stage 34 Xenopus embryo illustrating the areas of RA production (blue)ures.

116 J. Lynch et al. / Gene Expression Patterns 11 (2011) 112–117
(Sakai et al., 2001). Expression of the binding facilitator Crabp2 inthe chordoneural hinge suggests some RA signalling, which canonly come from the posterior notochord (Raldh1) is important inthis compartment in Xenopus.
RA synthesis in undifferentiated somites of mouse, chick andzebrafish embryos results from the expression of Raldh2 in this tis-sue (Grandel et al., 2002; Niederreither et al., 1997; Swindell et al.,1999). In surprising contrast, in Xenopus we see no expression ofRaldh1, 2 or 3 in either the forming somites or tailbud posteriorwall. Compelling evidence does however exist for the instructiverole of RA in somitogenesis in Xenopus. Earlier in development,during neurula stages, Raldh2 from the lateral plate mesoderm islikely to provide a source of RA anteriorly, with Cyp26a degradingthe signal posteriorly (Moreno and Kintner, 2004). Furthermore,targets of RA signalling such as the bHLH genes, Thylacine 1 andMespo and the RBL gene Ledgerline are expressed in the presomiticmesoderm and have been implicated in somitogenesis (Chan et al.,2006; Moreno and Kintner, 2004; Wang and Ding, 2006). It is dif-ficult to see how lateral plate Raldh2 could continue this role informing tail somites (Fig. 4). Somites continue to form in tailbudsthat have been removed from the embryo, detached from anteriorRA (Tucker and Slack, 1995). We also observe weak Raldh1 expres-sion in the posterior notochord of stage 34 embryos, which mayprovide an alternative source. Crabp2 in the tailbud may act to am-plify the affect of any RA generated by Raldh1. Finally, alternativemethods of RA synthesis have been identified in fish and chickens(Chambers et al., 2007; Lampert et al., 2003), suggesting that Raldhexpression alone may not give the whole picture. Whether thisrepresents a true difference in RA requirements between frogsand other vertebrates or even between frog trunk and tail somites,therefore remains to be resolved.
1.3.4. ConclusionsWe have examined the expression of several genes whose pro-
tein products play key roles in the regulation of endogenous RA inthe developing X. laevis embryo. These expression patterns suggestthat RA regulates the development of the kidney, lateral platemesoderm and otic vesicle in the same way as other vertebrates.While RA likely patterns the anterior, early developing somites,we find little evidence for the involvement of RA in later somito-genesis in the tadpole tail. Furthermore our data suggests thatRA degradation may be dispensable for heart patterning in frogs,but that Cyp26b is uniquely expressed in the ventral aortae. Lastly,we show that differentiation of the lens of the eye could depend onRA degradation, which has not been reported in other vertebrates.Differences in the way that RA is regulated in regeneration compe-tent species such as X. laevis, when compared to other vertebrates,may be linked to regenerative success.
2. Experimental procedures
2.1. Cloning of X. laevis Cyp26b, Cyp26a and Crabp1, and production ofprobes for gene expression analysis
Full coding sequence for Raldh1 was amplified using primersdesigned to sequence NM_00108777NM1: 50-CCTCTAGATGTCCGTGAAAACCCTTC and 30-GCGGTACCTTAGGAATTCTTCTGTGAAATG. Fullcoding sequence for Raldh2 was amplified using primers designedto sequence NM_001090775: 50-GCTCTAGAATGACTTCCAGT AAAATCGAG and 30-GCGGTACCTTAGGAATTCTTCTGG. Coding sequencefor Raldh3 was amplified using primers designed to sequenceNM_001095605: 50-GCTCTAGAATGACAACCACTAACGGGG and 30-CTGGTACCGCTGCTTTCATTTCAGTGG. Complete coding sequence ofCyp26a was amplified using degenerate primers 50-CCTCTAGAATGGATCTRTACACTCTGC and 30-GCGGTACCTTAAATAGA GCTGGMRAAGG.
Full coding sequence of Crabp2 was amplified using primersdesigned to sequence BC106392: 50-CCTCTAGAAT GCCCAACTTCTCAGG and 30-GCGGTACCTTAGTCCCTGATGTAAATCC.
Raldh1, 2 and 3, Crabp2 and Cyp26a PCR products were clonedvia flanking Xba1 and Kpn1 sites (italic) into pBSIIKS+ (stratagene).Antisense probes were made by cutting with Kpn1 and transcribingwith T3 polymerase.
Partial coding sequence of Cyp26b was amplified using primersdesigned to the X. tropicalis sequence NM_001079189: 50-CAACTCCGATGGGCAGCG and 30-GTGGCAAGCTCAAACCTGCT and clonedinto pGEM-T (Promega). Probe was made by cutting with NcoI orBamHI and transcribing with SP6 polymerase.
Partial coding sequence of Crabp1 was amplified using primersdesigned to tropicalis sequence BC135219: 50-TCTGGAAAATGAAGAGCAGTGA and 30-ATATCCGTGTACAGACCACATCA and clonedinto pGEM-T (Promega). Probe was made by cutting with PstI andtranscribing with T7 polymerase.
Sequences were deposited in NCBI with the following accessionnumbers: Cyp26A1 (EU816561), Cyp26B1 (EU816560) and Crabp1(EU8166559).
2.2. In situ hybridisation
Whole-mount in situ hybridisation of embryos and tadpoleswas performed as previously described (Beck and Slack, 1998). Sec-tions were prepared from paraffin embedded embryos at either7 lm for transverse sections or 12 lm for sagittal/parasagittal sec-tions and counterstained with eosin in 70% ethanol (pinkcytoplasm).
2.3. X. laevis embryos
Embryos were staged according to Nieuwkoop and Faber(1994). All animal experiments were subject to New Zealand’s ani-mal welfare standards for vertebrates and were reviewed by theUniversity of Otago, Animal Ethics Committee (AEC). The AEC ap-proved all experiments under protocol AEC78/06.
Acknowledgements
This work was funded by a University of Otago Research Grant(2008) to CB. The authors wish to thank Amy Armstrong for frogcolony care.
References
Abu-Abed, S., MacLean, G., Fraulob, V., Chambon, P., Petkovich, M., Dolle, P., 2002.Differential expression of the retinoic acid-metabolizing enzymes CYP26A1 andCYP26B1 during murine organogenesis. Mech. Dev. 110, 173–177.
Beck, C.W., Slack, J.M., 1998. Analysis of the developing Xenopus tail bud revealsseparate phases of gene expression during determination and outgrowth. Mech.Dev. 72, 41–52.
Blomhoff, R., Blomhoff, H.K., 2006. Overview of retinoid metabolism and function. J.Neurobiol. 66, 606–630.
Boylan, J.F., Gudas, L.J., 1992. The level of CRABP-I expression influences theamounts and types of all-trans-retinoic acid metabolites in F9 teratocarcinomastem cells. J. Biol. Chem. 267, 21486–21491.
Budhu, A.S., Noy, N., 2002. Direct channeling of retinoic acid between cellularretinoic acid-binding protein II and retinoic acid receptor sensitizes mammarycarcinoma cells to retinoic acid-induced growth arrest. Mol. Cell. Biol. 22, 2632–2641.
Chambers, D., Wilson, L., Maden, M., Lumsden, A., 2007. RALDH-independentgeneration of retinoic acid during vertebrate embryogenesis by CYP1B1.Development 134, 1369–1383.
Chan, T., Satow, R., Kitagawa, H., Kato, S., Asashima, M., 2006. Ledgerline, a novelXenopus laevis gene, regulates differentiation of presomitic mesoderm duringsomitogenesis. Zool. Sci. 23, 689–697.
Chen, Y., Pollet, N., Niehrs, C., Pieler, T., 2001. Increased XRALDH2 activity has aposteriorizing effect on the central nervous system of Xenopus embryos. Mech.Dev. 101, 91–103.
Cleaver, O., Tonissen, K.F., Saha, M.S., Krieg, P.A., 1997. Neovascularisation of theXenopus embryo. Dev. Dyn. 210, 66–77.

J. Lynch et al. / Gene Expression Patterns 11 (2011) 112–117 117
Dekker, E.J., Vaessen, M.J., Van den Berg, C., Timmermans, A., Godsave, S., Holling, T.,Nieuwkoop, P., Van Kessel, A.G., Durston, A., 1994. Overexpression of a cellularretinoic acid binding protein (xCRABP) causes anteroposterior defects indeveloping Xenopus embryos. Development 120, 973–985.
Duester, G., 2007. Retinoic acid regulation of the somitogenesis clock. Birth DefectsRes., C Embryo Today 81, 84–92.
Durston, A.J., Timmermans, J.P., Hage, W.J., Hendriks, H.F., de Vries, N.J., Heideveld,M., Nieuwkoop, P.D., 1989. Retinoic acid causes an anteroposteriortransformation in the developing central nervous system. Nature 340, 140–144.
Grandel, H., Lun, K., Rauch, G.J., Rhinn, M., Piotrowski, T., Houart, C., Sordino, P.,Kuchler, A.M., Schulte-Merker, S., Geisler, R., Holder, N., Wilson, S.W., Brand, M.,2002. Retinoic acid signalling in the zebrafish embryo is necessary during pre-segmentation stages to pattern the anterior-posterior axis of the CNS and toinduce a pectoral fin bud. Development 129, 2851–2865.
Guthrie, S., 1996. Patterning the hindbrain. Curr. Opin. Neurobiol. 6, 41–48.Haselbeck, R.J., Hoffmann, I., Duester, G., 1999. Distinct functions for Aldh1 and
Raldh2 in the control of ligand production for embryonic retinoid signalingpathways. Dev. Genet. 25, 353–364.
Hollemann, T., Chen, Y., Grunz, H., Pieler, T., 1998. Regionalized metabolic activityestablishes boundaries of retinoic acid signalling. EMBO J. 17, 7361–7372.
Lampert, J.M., Holzschuh, J., Hessel, S., Driever, W., Vogt, K., von Lintig, J., 2003.Provitamin A conversion to retinal via the beta, beta-carotene-15, 150-oxygenase (bcox) is essential for pattern formation and differentiation duringzebrafish embryogenesis. Development 130, 2173–2186.
Lee, G.S., Kochhar, D.M., Collins, M.D., 2004. Retinoid-induced limb malformations.Curr. Pharm. Des. 10, 2657–2699.
Liu, R.Z., Sharma, M.K., Sun, Q., Thisse, C., Thisse, B., Denovan-Wright, E.M., Wright,J.M., 2005. Retention of the duplicated cellular retinoic acid-binding protein 1genes (crabp1a and crabp1b) in the zebrafish genome by subfunctionalizationof tissue-specific expression. FEBS J. 272, 3561–3571.
Lupo, G., Liu, Y., Qiu, R., Chandraratna, R.A., Barsacchi, G., He, R.Q., Harris, W.A., 2005.Dorsoventral patterning of the Xenopus eye: a collaboration of Retinoid,Hedgehog and FGF receptor signaling. Development 132, 1737–1748.
MacLean, G., Abu-Abed, S., Dolle, P., Tahayato, A., Chambon, P., Petkovich, M., 2001.Cloning of a novel retinoic-acid metabolizing cytochrome P450, Cyp26B1, andcomparative expression analysis with Cyp26A1 during early murinedevelopment. Mech. Dev. 107, 195–201.
Malloch, E., Perry, K.J., Fukui, L., Johnson, V., Cheng, M., Wever, J., Beck, C.W., King,M.W., Henry, J.J., 2009. Gene expression profiles of lens regeneration anddevelopment in Xenopus. Dev. Dyn. 238, 2340–2356.
McCaffery, P., Tempst, P., Lara, G., Drager, U.C., 1991. Aldehyde dehydrogenase is apositional marker in the retina. Development 112, 693–702.
Mic, F.A., Molotkov, A., Fan, X., Cuenca, A.E., Duester, G., 2000. RALDH3, aretinaldehyde dehydrogenase that generates retinoic acid, is expressed in theventral retina, otic vesicle and olfactory pit during mouse development. Mech.Dev. 97, 227–230.
Moreno, T.A., Kintner, C., 2004. Regulation of segmental patterning by retinoic acidsignaling during Xenopus somitogenesis. Dev. Cell 6, 205–218.
Niederreither, K., Dolle, P., 2008. Retinoic acid in development: towards anintegrated view. Nat. Rev. Genet. 9, 541–553.
Niederreither, K., McCaffery, P., Drager, U.C., Chambon, P., Dolle, P., 1997. Restrictedexpression and retinoic acid-induced downregulation of the retinaldehydedehydrogenase type 2 (RALDH-2) gene during mouse development. Mech. Dev.62, 67–78.
Niederreither, K., Vermot, J., Messaddeq, N., Schuhbaur, B., Chambon, P., Dolle, P.,2001. Embryonic retinoic acid synthesis is essential for heart morphogenesis inthe mouse. Development 128, 1019–1031.
Nieuwkoop, P.D., Faber, J., 1994. Normal table of Xenopus laevis (Daudin). GarlandPublishing Inc. New York.
Pijnappel, W.W., Hendriks, H.F., Folkers, G.E., van den Brink, C.E., Dekker, E.J.,Edelenbosch, C., van der Saag, P.T., Durston, A.J., 1993. The retinoid ligand 4-oxo-retinoic acid is a highly active modulator of positional specification. Nature366, 340–344.
Pijnappel, W.W., Folkers, G.E., de Jonge, W.J., Verdegem, P.J., de Laat, S.W.,Lugtenburg, J., Hendriks, H.F., van der Saag, P.T., Durston, A.J., 1998.Metabolism to a response pathway selective retinoid ligand during axialpattern formation. Proc. Natl. Acad. Sci. USA 95, 15424–15429.
Reijntjes, S., Gale, E., Maden, M., 2003. Expression of the retinoic acid catabolisingenzyme CYP26B1 in the chick embryo and its regulation by retinoic acid. GeneExpr. Patterns 3, 621–627.
Reijntjes, S., Blentic, A., Gale, E., Maden, M., 2005. The control of morphogensignalling: regulation of the synthesis and catabolism of retinoic acid in thedeveloping embryo. Dev. Biol. 285, 224–237.
Ruberte, E., Friederich, V., Morriss-Kay, G., Chambon, P., 1992. Differentialdistribution patterns of CRABP I and CRABP II transcripts during mouseembryogenesis. Development 115, 973–987.
Sakai, Y., Meno, C., Fujii, H., Nishino, J., Shiratori, H., Saijoh, Y., Rossant, J., Hamada,H., 2001. The retinoic acid-inactivating enzyme CYP26 is essential forestablishing an uneven distribution of retinoic acid along the anterio-posterior axis within the mouse embryo. Genes Dev. 15, 213–225.
Spoorendonk, K.M., Peterson-Maduro, J., Renn, J., Trowe, T., Kranenbarg, S., Winkler,C., Schulte-Merker, S., 2008. Retinoic acid and Cyp26b1 are critical regulators ofosteogenesis in the axial skeleton. Development 135, 3765–3774.
Swindell, E.C., Thaller, C., Sockanathan, S., Petkovich, M., Jessell, T.M., Eichele, G.,1999. Complementary domains of retinoic acid production and degradation inthe early chick embryo. Dev. Biol. 216, 282–296.
Tanibe, M., Michiue, T., Yukita, A., Danno, H., Ikuzawa, M., Ishiura, S., Asashima, M.,2008. Retinoic acid metabolizing factor xCyp26c is specifically expressed inneuroectoderm and regulates anterior neural patterning in Xenopus laevis. Int. J.Dev. Biol. 52, 893–901.
Tsonis, P.A., Trombley, M.T., Rowland, T., Chandraratna, R.A., del Rio-Tsonis, K., 2000.Role of retinoic acid in lens regeneration. Dev. Dyn. 219, 588–593.
Tucker, A.S., Slack, J.M.W., 1995. The Xenopus laevis tail-forming region.Development 121, 249–262.
Wang, J.H., Ding, X.Y., 2006. Cloning and analyzing of Xenopus mespo promoter inretinoic acid regulated mespo expression. Acta Biochim. Biophys. Sin.(Shanghai) 38, 759–764.