and in response to injury - journal of biological chemistry · functions in several tissues both in...
TRANSCRIPT

Epilysin: A Novel Human Matrix Metalloproteinase
(MMP-27) Expressed in Testis and Keratinocytes
and in Response to Injury
Jouko Lohi1, Carole L. Wilson, Jill D. Roby, and William C. Parks
Departments of Pediatrics (Allergy and Pulmonary Medicine) and Cell Biology and Physiology,
Washington University School of Medicine, St. Louis, MO 63110
Running title: Human Epilysin Gene
1To whom correspondence should be addressed:
Jouko Lohi, M.D., Ph.D. Telephone: +358-9-191-26469
Department of Pathology Fax: +358-9-191-26475
Haartman Institute, University of Helsinki Email: [email protected]
P.O. Box 21 (Haartmaninkatu 3)
FIN-00014 University of Helsinki
FINLAND
Key words: cDNA cloning, wound healing
Copyright 2000 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on December 19, 2000 as Manuscript M001599200 by guest on July 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
Summary
We have cloned a new human matrix metalloproteinase (MMP-27, epilysin) from human
keratinocyte and testis cDNA libraries. Like most MMPs, epilysin contains a signal sequence, a
prodomain with a PRCGVTD sequence, a zinc-binding catalytic domain with an
HEIGHTLGLTH sequence, and a hemopexin-like domain. In addition, epilysin has a furin
activation sequence (RRKKR) but has no transmembrane sequence. The exon-intron
organization and splicing pattern of epilysin differs from that of other MMP genes. It has only 8
exons, and 5 exons are spliced at sites not used by other MMPs. Another novel feature of
epilysin is that exon 4 is alternatively spliced to a transcript that does not encode the N-terminal
half of the catalytic domain. Northern hybridization of tissue RNA indicated that epilysin is
expressed at high levels in testis, and at lower levels in lungs, heart, colon, intestine, and brain.
RNAse protection assay with various cell lines indicated that epilysin was selectively expressed
in keratinocytes. Recombinant epilysin degraded casein in a zymography assay, and its
proteolytic activity was inhibited by EDTA and by batimastat, a selective MMP inhibitor.
Immunohistochemical staining showed expression of epilysin protein in the basal and suprabasal
epidermis of intact skin. In injured skin, prominent staining for epilysin was seen in basal
keratinocytes both at and some distance from the wound edge, a pattern that is quite distinct from
that of other MMPs expressed during tissue repair. These findings suggest that this new MMP
functions in several tissues both in tissue homeostasis and in repair.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

3
Introduction
The matrix metalloproteinases (MMPs)4 comprise a family of enzymes that share several
common structural features and that function both in the turnover and degradation of
extracellular matrix proteins and in the processing, activation, or deactivation of a variety of
soluble factors (1). MMPs, or matrixins, are a subgroup of the much larger metalloproteinase
superfamily, which also includes astacin and ADAM proteinases, among others. To date 23
different MMPs have been cloned, and additional members continue to be identified (2).
To be classified as a matrix metalloproteinase, a protein must have conserved features of
two domains, namely the pro- and the catalytic domains. The prodomain of a typical MMPs is
about 80 amino acids, and all MMPs, except MMP-23 (3), contain the consensus sequence
PRCXXPD. As for all metalloproteinases, the catalytic domain contains an active site Zn2+ that
binds three conserved histidines in the sequence HEXXHXXGXXHS/TXXXXXXM, which also
contains a conserved methionine to the carboxy side of the zinc-binding site (metzincins) (4). In
an inactive state, the conserved cysteine residue in the prodomain provides the fourth
coordination site for the catalytic zinc ion. In addition, with the exception of matrilysin (MMP-
7), endometase/matrilysin-2 (MMP-26), and MMP-23, MMPs have a hinge region, which is
often proline-rich, and a so-called hemopexin-like C-terminal domain (3,5,6). Other domains
found in MMPs are specialized to subgroups of enzymes. For example, four membrane-type
MMPs (MMP-14, -15, -16, -24) have transmembrane and cytosolic domains, whereas MT4-
MMP and MT6-MMP (MMP-17 and –25, respectively) have C-terminal hydrophobic extensions
that act as a glycosylphosphatidylinositol (GPI) anchoring signal (7-9). The two gelatinases
(MMP-2 and MMP-9) have gelatin-binding domains. MMP-23 lacks the hemopexin domain and
has a novel cysteine array motif and an immunoglobulin-like C2 type fold domain (3,10). In
addition to a common domain structure, MMPs share a similar gene arrangement suggesting that
they were generated by duplications of an ancestor gene. At least eight of the known human
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

4
MMP genes (MMPs 1, 3, 7, 8, 10, 12, 13, and 20) are clustered on chromosome 11 at 11q21-23.
Other known MMP genes are scattered along chromosomes 1, 8, 12, 14, 16, 20, and 22
(3,11,12).
MMPs are secreted or bound or anchored to the cell membrane, and all function extracellularly
or within the secretion pathway. As demonstrated in defined in vitro studies, almost all MMPs
can cleave or degrade some protein components of the extracellular matrix, and many are able to
act on a wide variety of proteins (13). Notable exceptions to this rule are stromelysin-3 (MMP-
11) and MMP-23, which have no known extracellular matrix substrates (3,14). In addition and
quite important, MMPs can process or degrade non-matrix proteins. For example, matrilysin is
responsible for activation of the pro-form of α-defensins (15), a class of secreted antimicrobial
peptides, and several MMPs can cleave and inactive the serpin α1-proteinase inhibitor (16,17),
which is an in vivo substrate for gelatinase-B (MMP-9) (18). In addition, several MMPs, such as
MMP-1, -2, -3, -7, and –11, among others, directly modulate the activity of several growth
factors, such as TNFα, IGF-1, EGFs, and FGFs (19-24). Thus, matrix degradation is neither a
sole or common functional feature of MMPs.
Many of the secreted MMPs, including MMPs 1, 3, 9, 10, 11, and 13, are not expressed
in normal, healthy, resting tissues, and, with some exceptions, their production and activity are
maintained at nearly undetectable levels. In contrast, some level of MMP expression is seen in
any repair or remodeling process, in any diseased or inflamed tissue, and in essentially any cell
type grown in culture (25,26). Although the qualitative pattern and quantitative levels of MMPs
vary among tissues, diseases, tumors, inflammatory conditions, and cell lines, a reasonably safe
generalization is that activated cells express MMPs. Some MMPs, including MMPs 7, 19, 24, 25,
and 26, are expressed in healthy tissues (27-31).
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

5
In the present study, we report on the cloning and initial characterization of a novel
human MMP, MMP-27, which we call epilysin. We isolated the cDNA for this protein from
keratinocyte and testis libraries, and we show that it has the essential domains of a prototypic
MMP, as well as several unique features. Because of its ability to degrade a protein substrate
was fully inhibited by EDTA and a hydroxamate MMP inhibitor, epilysin is indeed a
metalloenzyme. Our data suggests that epilysin is expressed in intact tissues, and upregulated in
response to injury. Thus, this new MMP may function in both tissue homeostasis and tissue
repair.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

6
Experimental Procedures
Cloning of Human Epilysin cDNA. Exon/Intron Mapping. A search of the GenBank
data base with the peptide string FDGXXXXLAHAXXPGXXXXGDXHFDXXEXW, which is
conserved among MMPs, returned a homologous sequence within a 82 kb human genomic DNA
clone (Accession AC006237). Nested primers were designed to amplify a 161 bp cDNA
fragment by RT-PCR using HT-1080 human fibrosarcoma line RNA as a template. The
amplified product corresponds to bases 726-886 (see Fig. 1). By screening a human foreskin
keratinocyte cDNA library (HL1110b, Clontech, Palo Alto, CA) by plaque hybridization with
the 32P-labeled 161 bp cDNA fragment, we obtained 3 positive clones, which were then
sequenced. The longest clone of 1.5 kb contained exons 3 to 8 of epilysin (see below for exon
numbering). This cDNA clone was then used to screen a pooled human testis cDNA library
(HL5033t, Clontech). Among the more than 20 positive clones was a clone that contained the
coding regions of exons 1 and 2. Exon-intron boundaries were determined by comparing the
cDNA sequences with the genomic sequence in data base.
Computer Analyses. Screening of GenBank data base was performed using the
TBLASTN program and the NCBI server (URL: www.ncbi.nlm.nih.gov) (32). Epilysin signal
peptide was identified and its cleavage site was predicted using SignalP server (URL:
genome.cbs.dtu.dk/services/SignalP/) (33). The amino acid sequences of human MMPs 1, 3, 11,
14, and 19 were aligned with epilysin using CLUSTAL W (34). Phylogenetic tree was drawn
based on a CLUSTAL W alignment of the amino-acid sequences of the catalytic domains of all
known human MMPs.
Cell Culture. Human foreskin fibroblasts (HFF), immortalized human keratinocytes
(HaCaT) (35), and human fibrosarcoma HT-1080 cells (CCL-121, American Type Culture
Collection, Rockville, MD) were grown to confluence in Eagle's minimal essential medium
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

7
(MEM) containing 10% heat-inactivated fetal calf serum (GIBCO-BRL, Gaithersburg, MD), 100
IU/ml penicillin, and 50 µg/ml streptomycin. Human colon adenocarcinoma HT-29 cells
(HTB38, ATCC) were maintained in RPMI medium. MMP expression was stimulated by
treatment with 16 nM phorbol ester (PMA, Sigma Chemical Co., St. Louis, MO) for 24 h.
Primary human keratinocytes were isolated from normal full thickness adult skin and cultured on
collagen-coated dishes as described (36). U937 cells (ATCC, CRL 1593), a human monocyte-
like cell line, were cultured and differentiated to stimulated macrophage-like cells by a 24-h
treatment with 4 nM PMA and 5 µg/ml lipopolysaccharide (LPS, Sigma) as described (37). Total
RNA was isolated using RNAzol B (Tel-test, Inc., Friendswood, TX).
Northern Blot Analysis. Nylon filters containing 2 µg of poly (A)+ RNA from various
human tissues (Clontech) were prehybridized with ExpressHyb hybridization solution
(Clontech), and then hybridized in the same solution with 32P-labeled epilysin probe, generated
by random priming using a 1.5 kb cDNA fragment as a template. Loading was normalized by
hybridization with a β-actin cDNA probe. Hybridization and washes were performed according
to manufacturer’s instructions.
RNAse Protection Analysis and Determination of Alternative Splicing. Expression of
epilysin mRNA in cultured cells was analyzed by ribonuclease protection technique using Direct
Protect kit (Ambion Inc., Austin, TX) and two different RNA probes. A PCR fragment of 161 bp
(bases 726-886 in epilysin cDNA) and another fragment of 539 bp (bases 370-908) were cloned
to pGEM-T-Easy (Promega), and plasmid DNA was linearized with Sal I. Antisense RNAs of
257 nt and 635 nt, respectively, were transcribed with T7 RNA polymerase in the presence of [α-
32P]UTP and hybridized with 10 µg of total cellular RNA. After an overnight hybridization,
unpaired RNA was degraded by treatment with RNAse A and RNAse T1 (10 U/ml and 400
U/ml, respectively) at 37°C for 30 min followed by isopropanol precipitation. Protected RNA
fragments were fractionated by 5% SDS-PAGE containing 6 M urea and visualized by
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

8
autoradiography. Alternative splicing was first detected when RT-PCR reactions to amplify
regions containing bases 370-960 and 440-960 (forward primers 5’-
GTGGGTGTCCCAGCTACCTGTC-3' and 5’-TGCGGGGTTACAGATACCAACAG-3',
reverse primer 5’-CTCTTGTAGTAGGGCGCCATGAG-3') using HaCaT cDNA as a template
gave three specific products of different size. Specificity of the amplification products was
determined by Southern blotting using a [γ-32P]ATP labeled internal oligonucleotide probe 5’-
GCGGCGAAGCGCACTTCGACCAAGATGAGC-3', and the amplification products were
purified from gel and sequenced. The presence of alternatively spliced mRNA in HaCaT cells
was verified by RNAse protection analysis using the 635 nt RNA probe described above.
Baculoviral Recombinant Epilysin. For generation of recombinant protein, nucleotides
536 to 1732 of epilysin cDNA, coding for amino acids 123 to 520 of epilysin protein were
amplified using primers 5’-CGGGATCCGACGATGACGATAAGTTTGCAAAGCAAGGT-
AACAAATGGTACAAGC-3' (forward, epilysin sequence underlined) and 5’-
CGGAATTCTCAGAACAGGGCGCTCCCCGAGTTG-3’ (reverse). The amplification product
was digested with Bam HI and Eco RI and cloned into the pAcSecG2T baculovirus transfer
vector (Pharmingen, San Diego, CA). The resulting expression construct codes for a fusion
protein of Schistosoma japonicum glutathione S-transferase (GST) and amino acid residues 123
to 520 of epilysin corresponding to the putative furin-activated enzyme. This vector also
provides the signal peptide from the baculovirus protein gp67 to direct secretion of the fusion
protein. Sf9 cells were transfected with the construct and BaculoGold DNA (Pharmingen) to
produce recombinant baculovirus. Following two rounds of virus amplification, High Five insect
cells (Stratagene) were infected and harvested along with conditioned medium 5 days later. Cell
pellets were lysed in 10 mM Tris-HCl buffer, pH 7.5, containing 130 mM NaCl, 1% Triton X-
100, 10 mM NaF, 10 mM NaPi, and 10 mM NaPPi. GST-epilysin fusion protein was purified
from cell lysates and conditioned medium by glutathione Sepharose affinity chromatography
according to manufacturer's protocol (Pharmingen). Protein eluting from the affinity resin was
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

9
analyzed by SDS-PAGE and Western blotting with a rabbit polyclonal anti-GST antibody
(Upstate Biotechnology, Lake Placid, NY). Unexpectedly, most of the GST-epilysin fusion
protein was found in cell pellets and not in the conditioned medium.
Recombinant Epilysin Produced in E. coli and Casein Zymography. To express a fusion
protein consisting of Schistosoma japonicum glutathione S-transferase (GST) and the pro- and
catalytic domains of epilysin (amino acid residues 23 to 284, see Fig. 1) in E. coli, a 802-bp
fragment of the MMP-27 cDNA was amplified by PCR using primers 5’-
CGGGATCCCAGCCCGCGGAGCGCGGA-3’ (forward, epilysin sequence underlined) and 5’-
GGAATTCTCACCCATACAGGCTCTGCACGGCCAGC-3’ (reverse), and the PCR-product
was digested with Bam HI and Eco RI and cloned into the pGEX-6P-2 vector (Amersham-
Pharmacia, Uppsala, Sweden). The resulting expression vector was then transformed into
BL21(DE3)pLys strain of E. coli. Overnight bacterial culture derived from a single bacterial
colony was diluted 1:10 and incubated at 37°C for 2 h. Expression of the fusion protein was then
induced by adding isopropyl-1-thio-β-D-galactopyranoside (IPTG) (0.5 mM final concentration)
followed by further incubation at 25°C for 4 h. Recombinant protein obtained in inclusion bodies
was solubilized by sonication in the presence of N-lauroyl-sarcosine and affinity purified with
glutathione Sepharose as described (38). Recombinant fusion protein bound to glutathione
Sepharose was then digested with PreScission protease according to manufacturer’s instructions
(Pharmacia) to remove the GST tag. Between the GST domain and epilysin, the fusion protein
has the recognition sequence (LEVLFQGP) for PreScission Protease. Recombinant epilysin
(pro- and catalytic domains) was then eluted with 50 mM Tris-HCl buffer, pH 7.5, containing
500 mM NaCl, 5 mM dithiothreitol and 0.1% Brij-35. Caseinolytic activity was measured by
zymography using a 4-16% SDS-PAGE blue casein zymogram gel (Novex, San Diego, CA)
according to manufacturer’s protocol. After electrophoresis, the zymogram gel was washed and
incubated in 50 mM Tris-HCl buffer, pH 7.5 containing 1-2.5% Triton X-100 and either 1) 10
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

10
mM EDTA or 2) both 5mM CaCl2 and 1 µM ZnCl2 or 3) 10 µM batimastat in the presence of
5mM CaCl2 and 1 µM ZnCl2.
Preparation of Antibodies and Immunoblotting Assay for Epilysin. An 8-chain branching
multiple antigenic peptide (MAP) of 16 amino acids, DQDERWSLSRRRGRNL, corresponding
to the middle of the catalytic domain of epilysin (amino acid residues 219-234, see Fig. 1), was
used as an antigen (Research Genetics, Huntsville, USA). Rabbits were first immunized with 0.5
mg of the peptide in complete Freund’s adjuvant and three booster injections with 0.5 mg of the
peptide in incomplete Freund’s adjuvant were given 2, 6, and 8 weeks later by a commercial
operation (Research Genetics). Antibodies were purified from whole serum, harvested at 10
weeks after primary injection, by affinity chromatography with the peptide coupled to NHS
Sepharose-4B according to manufacturer’s instructions (Pharmacia). For immunoblotting,
confluent cultures of HaCaT cells were washed with serum free medium and were incubated
under serum free conditions for an additional 48 h. The medium was then collected and
concentrated 70-fold using a Centricon microconcentrator (Amicon, Beverly, MA). 10 µl of
concentrated conditioned medium was mixed with an equal amount of Laemmli sample buffer
containing 10% β-mercaptoethanol and resolved by SDS-PAGE through a 4-15% gradient gel.
Recombinant baculoviral GST-epilysin fusion protein was used as a positive control. Proteins
were then electrophoretically transferred to nitrocellulose (Schleicher & Schuell, Dassel,
Germany) using a semi-dry blotting apparatus at 2.5 mA/cm2 for 30 min. Membranes were
blocked with 5% milk in PBS/Triton X-100 (0.5%) and incubated with 0.15 µg/ml of affinity
purified antibodies in 50 mM Tris-HCl buffer containing 500 mM NaCl, 0.1% Tween 20 and
0.1% bovine serum albumin, pH 8.5. After five washes in the same buffer, the bound antibodies
were detected using biotinylated anti-rabbit-IgG antibodies and peroxidase-conjugated
streptavidin (Dakopatts, Copenhagen, Denmark) and enhanced chemiluminescence Western
blotting detection system (Amersham International PIC, Amersham, UK) as described (39).
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

11
Expression of Epilysin in CHO Cells. Immunofluorescence Staining. Chinese hamster
ovary (CHO) cells were transfected with a cDNA construct coding for epilysin with a C-terminal
10-aa influenza virus hemaglutinin tag under transcriptional control by CMV promoter in
pcDNA3 vector (InVitrogen, San Diego, CA) using FuGENE 6 transfection reagent according to
manufacturer’s instructions (Boehringer Mannheim GmbH, Mannheim, Germany). 24 h after
transfection, transfected cell clones were selected for neomycin resistance as described (40). For
immunofluorescence staining, a pool of transfected cells was plated on glass coverslips, and 3
days later, the cells were fixed with 3% paraformaldehyde in PBS (phosphate buffered saline,
0.14 M NaCl in 10 mM phosphate buffer, pH 7.4). After fixing the coverslips were washed three
times with PBS and blocked with 5% bovine serum albumin (BSA) in PBS for 30 min. Affinity-
purified epilysin antibody was then added (1:100 dilution) in 0.5% BSA in PBS and incubated 1
h at room temperature with mild shaking. After three washes with the same buffer, FITC
conjugated anti-rabbit IgGs (Jackson Immunoresearch Laboratories, West Grove, PA) were
added and incubated for 1 h. Coverslips were then washed 5 times with PBS and mounted on
glass slides using Vectashield anti-fading agent (Vector Laboratories, Inc., Burlingame, CA).
Immunohistochemistry. Individual, 4-mm-wide, full-thickness biopsies of human skin
used for keratinocyte culture were placed into the wells of 6-well cluster dishes and covered with
DMEM containing antibiotics. 24 h later, tissues were fixed in 10% buffered-formalin and
processed for paraffin embedding. Deparaffinized 5 µm sections were processed for
immunohistochemistry using alkaline phosphatase as described (41). Endogenous peroxidase
activity was blocked by incubation in 0.3% H202 for 30 min at room temperature. Affinity
purified anti-human epilysin antibody was diluted 1:1000. Bound antibody was detected using a
Vectastain ABC Elite kit (Vector Laboratories) following the manufacturer's instructions.
Peroxidase activity was detected using 3,3'-diaminobenzidine tetrahydrochloride as chromogenic
substrate. Sections were counterstained with Harris hematoxylin. For negative controls, sections
were processed with preimmune serum.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

12
Results
Cloning and Sequencing of a cDNA Encoding Human Epilysin. Comparison with Other
MMPs. To identify undiscovered MMPs, we searched the GenBank database using the
TBLASTN program and a peptide query sequence FDGXXXXLAHAXXPGXXXXGDXHFD-
XXEXW, which defines a partial consensus sequence of a metalloproteinase catalytic domain.
Among the more than 100 hits was a human genomic DNA clone (Accession number
AC006237) that was submitted by Whitehead Institute/MIT Center for Genome Research and
had been sequenced as a part of the Human genome project sequencing chromosome 17. There
was no annotation that the sequence would code for proteins. After translation of the genomic
DNA in three forward reading frames, several peptide sequences typical of MMPs, including a
propeptide sequence PRCGVTD and a catalytic domain sequence HEIGH, were identified, and
these sequences were separated by putative intronic sequences.
To assess if this genomic region was transcribed to an mRNA, two sets of primers were
designed; the forward and reverse primers were directed to different suspected exons. As a
source of RNA, we used the human fibrosarcoma cell line HT-1080, as these cells are known to
express a wide variety of MMPs (40,42). cDNA was synthesized using random hexamer primers
and was amplified by PCR. Two-stage PCR with nested primers produced an amplified DNA
fragment of expected size (161 bp), and the nucleotide sequence of this fragment was identical to
that of presumed exonic portions of the genomic sequence (data not shown).
To obtain the full-length cDNA for this novel MMP, we screened a human keratinocyte
cDNA library using the PCR product as a probe. (RNAse protection analysis of various cell lines
revealed that epilysin is expressed in cultured human keratinocytes; see Fig. 6.) Among the three
positive clones, we isolated and characterized a 1.5 kb cDNA that contained sequence coding for
part of the prodomain, the entire catalytic and hemopexin-like domains, a stop codon TGA, and a
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

13
85 bp of 3’ UTR. To determine the 5’ end of this MMP transcript, we screened a testis cDNA
library with a probe corresponding to the 5’-end of the 1.5 kb keratinocyte cDNA insert.
(Hybridization of tissue-RNA blots revealed that epilysin is expressed in human testis at high
levels; see Fig. 5.) Of the positive clones, we identified one containing a cDNA insert that coded
for the missing portion of the prodomain and for about 170 bases of 5’ UTR. The open reading
frame, starting from the first ATG codon, contains 1560 nucleotides and codes for a 520-aa
protein with a calculated molecular weight of 59 kDa (Fig. 1). We named this new MMP
epilysin. Using the accepted consecutive-number nomenclature, epilysin would be assigned
MMP-27.
The domain structure and organization of epilysin is predictable for an MMP (Fig. 1, 2).
Using an analysis program available at the SignalP server (33), we identified a typical
hydrophobic signal sequence of 22 amino acids at the amino terminus of epilysin (Fig. 1). The
signal sequence is followed by a prototypic MMP prodomain with the conserved cysteine-switch
sequence PRCGVTD (Fig. 1,2). In this sequence, a proline that is present in all other human
MMPs except MMP-19 (43) is replaced by a threonine (Fig. 2). In addition, after the cysteine
switch sequence, there is an 11-aa insertion, which is not present in other known MMPs,
followed by an RRKKR furin recognition sequence (Fig. 1, 2). The catalytic domain is highly
conserved relative to other MMPs, and as for secreted MMPs, an 8-aa insertion present only in
MT-MMPs (e.g., MMP-14) is lacking from epilysin (Fig. 2). The catalytic center with three
histidine residues, HEIGHTLGLTH, is unique in that no other MMP has threonine within this
sequence. A 39-aa hinge region is followed by a typical hemopexin-like domain. There is no
hydrophobic transmembrane sequence typical of membrane-inserted MMPs or a hydrophobic
extension typical of GPI anchored proteins. In addition, epilysin has two putative N-
glycosylation sites: one in the N-terminal part of the catalytic domain and another in the second
pexin-like repeat of the hemopexin domain (Fig. 1). The calculated molecular weight of the
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

14
proenzyme without the signal sequence is 56 kDa; the active, furin-processed enzyme is
estimated to be 45 kDa. These weights do not include any contribution in mass by glycosylation.
Comparison of the epilysin amino acid sequence with other MMPs by CLUSTAL W
program (Fig. 2) and construction of a phylogenetic tree on the basis of the catalytic domain
sequences (Fig. 3) indicates that epilysin is most closely related to some other recently cloned
MMPs, including MMP-19, MMP-23, MT4-MMP (MMP-17), MT6-MMP (MMP-25), and
stromelysin-3 (MMP-11) (3,8,43-45). The number of identical and similar residues with MMP-
19 catalytic domain is 46% and 60%, respectively.
Structural Organization of the Human Epilysin Gene. We mapped the exon/intron
junctions and determined an exon-intron map of the gene by comparing the cDNA and genomic
sequences (Fig. 4A). Exon-intron boundaries and the sizes of exons and introns are summarized
in Table 1. All exons were contained within the genomic BAC clone except exon 1, and hence,
we do not yet know the size of intron 1 (Table 1). The exon-intron structure of epilysin is unique
compared to other MMP genes. Whereas most MMP genes have 10 exons, the epilysin gene has
only 8 exons, similar to that of stromelysin-3 (MMP-11) (Fig. 4B) (46). Furthermore, only three
of the seven splice sites (splice sites between exons 1 and 2, 5 and 6, and 6 and 7) are at positions
conserved among most MMP genes. None of the unique splice sites are similar to those some
other “nontraditional” MMP genes characterized to date, such as MMP-7, MMP-11, or MMP-14.
Overall, the organization of the epilysin gene is similar to that of MMP-19, with one overt
difference being that exon 8 of epilysin corresponds to exons 8 and 9 of MMP-19 (Fig. 4B) (47).
The exon/intron boundaries conform to the GT/AG rule for splice sites (48) (Table 1).
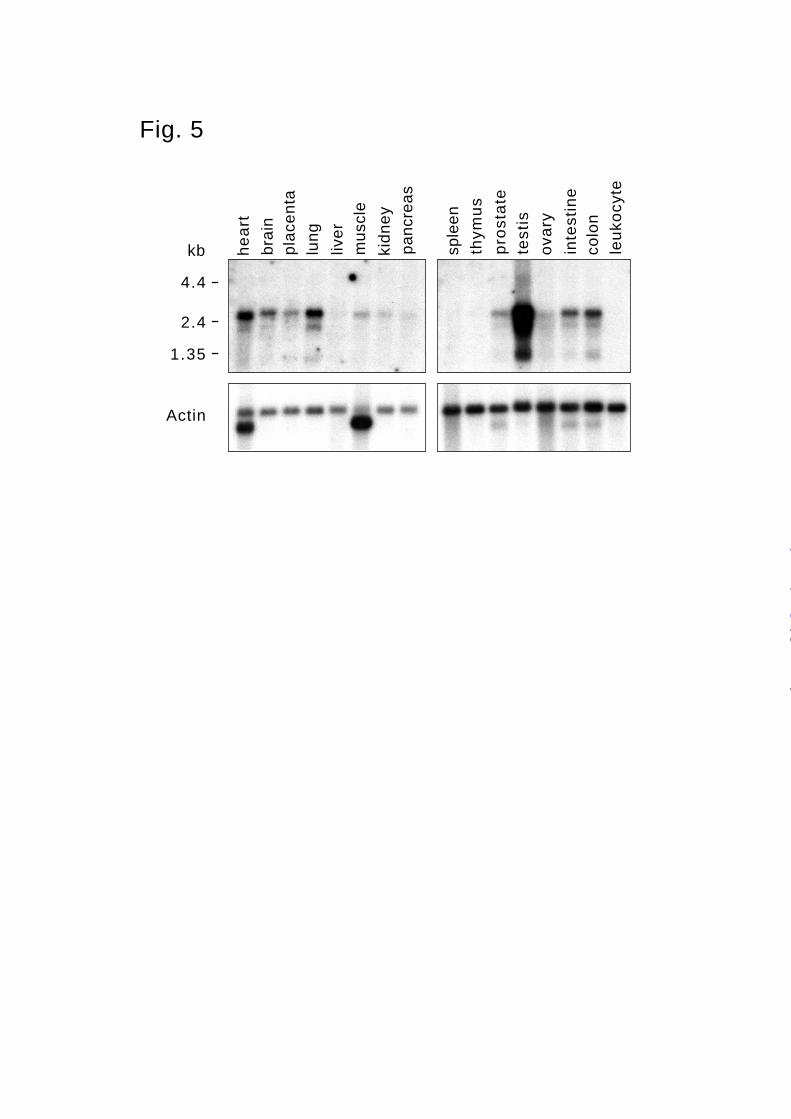
Analysis of Epilysin Expression in Human Tissues and Cell Lines. To analyze the
expression of epilysin in different human tissues, we hybridized a Northern blot containing
mRNA from various human tissues with a 1.5 kb epilysin cDNA probe. At least three different
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

15
transcripts of 2.6, 2.0 and 1.2 kb were detected in many tissues (Fig. 5), and this heterogeneity is
likely due to alternative splicing (see below). The 2.6 kb transcript was most abundant in all
tissues, and among tissues, the relative levels of three transcripts were about the same. Epilysin
mRNA is highly expressed in testis, and at lower levels in lungs, heart, colon, intestine, and brain
(Fig. 5).
To determine which cell types express epilysin, we screened RNA from several cell lines
known to actively express a variety of MMPs. To obtain maximal specificity and sensitivity, we
developed an RNA protection assay for epilysin mRNA. The human cells we used included HT-
29 colon carcinoma cells (49), U937 monocytic-like cells (50), HT-1080 fibrosarcoma cells (42),
neonatal foreskin fibroblasts (51,52), immortalized HaCaT keratinocytes (53), and normal
primary keratinocytes (53). Because the transcription of many MMP genes is strongly
upregulated by PMA, some cells were treated with this agent for 24 h before RNA was isolated.
U937 cells were treated with a combination of PMA and lipopolysaccharide, which mediates
differentiation to a macrophage-like phenotype accompanied by a potent induction of several
MMPs (37,50).
Although epilysin is expressed in several tissues, of the cell types tested, epilysin mRNA
was detected only in keratinocytes (Fig. 6). HaCaT keratinocytes showed the highest expression
of epilysin mRNA and primary keratinocytes, which were grown on collagen to induce MMP
expression (54), had a somewhat lower level of expression (Fig 6). All other cell lines were
negative for epilysin expression. Because we could amplify a portion of epilysin cDNA from
HT-1080 RNA by RT-PCR, these fibrosarcoma cells express epilysin at low levels.
To determine if epilysin mRNA is translated into a protein, we generated rabbit
polyclonal antibodies against a 16 amino acid MAP peptide in the middle of the catalytic
domain. Anti-epilysin antibodies were affinity purified using the same peptide. The specificity of
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

16
the antibody was tested by immunofluorescence staining of transfected CHO cells and by
immunoblotting of conditioned medium (Fig. 7). CHO cells transfected with an epilysin
expression construct showed a predictable range of recombinant protein production. Whereas
some clones showed prominent fluorescence for epilysin, other selected clones had no staining
(Fig. 7A). In addition, we used immunoblotting to assess if epilysin protein is released by HaCaT
keratinocytes. As a positive control, we used a fusion protein of Schistosoma japonicum
glutathione S-transferase and amino acid residues 123 to 520 of epilysin, corresponding to the
putative furin-activated enzyme. A strongly immunoreactive band of about 58 kDa was detected
in HaCaT conditioned medium and the predicted 75 kDa band was seen in the fusion protein
preparation (Fig. 7B). In addition to the 58-kDa band, we detected in HaCaT conditioned
medium a slightly smaller band of about 55 kDa of much lower intensity (Fig. 7B). As is
discussed below, this smaller band could be a product of alternative splicing or alternative
glycosylation. Together, these findings indicate the epilysin mRNA codes a secreted protein that
is produced and released by keratinocytes.
Detection of Alternative Splicing. During the cloning of epilysin, we generated a probe by
RT-PCR using primers complementary to sequences in the exons 3 and 5 (see Methods). In
addition to the expected PCR product, we obtained a shorter cDNA of nearly equal intensity and
a weak band of intermediate size (Fig. 8A). Similar amplified products were generated using a
different forward primer in exon 3, and all PCR products were positive in Southern blotting (Fig.
8A). We gel purified and sequenced the amplified DNA and found that the longer amplification
product contained sequences for exons 3, 4, and 5, while the shorter amplification product
represented a mRNA species that contained exons 3 and 5 but lacked exon 4. To assess the
relative abundance of the different splice forms, another PCR product was cloned and used to
generate an RNA probe for RNAse protection analysis. This probe covers 179 nt of exon 3, the
entire exon 4 (225 nt), and 135 nt of exon 5. In addition to epilysin specific sequences, the RNA
probe contains 96 nt of vector derived sequence giving the total length of 635 nt. HaCaT RNA
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

17
was then hybridized with this probe and non-hybridized probe was degraded by RNAse
treatment. The protected fragments included two strong bands of approximately 550 nt and 180
nt, and a weaker band of 130 nt. The ~550 nt fragment could be protected by a mRNA transcript
that contains exons 3, 4, and 5 (expected size 539 nt), and the ~180 nt fragment would represent
probe protected by exon 3 (expected size 179 nt). The weaker ~130 nt band could be derived
from probe protected by an mRNA species containing exon 5 but not exon 4 (expected size 135
nt). The 135 nt band was much weaker than the 179 nt band suggesting that another species of
mRNA transcripts containing exon 3 but lacking both exons 4 and 5 is expressed in testis and by
HaCaT cells. RNAse protection using RNA from testis tissue and RNA from HaCaT cells gave
identical results (Fig. 8B).
Production of recombinant epilysin (pro and catalytic domains) in bacterial cells and
analysis of its enzymatic activity. To determine if epilysin cDNA codes for an enzyme with
proteolytic activity, we expressed recombinant epilysin in E. coli as a fusion protein consisting of
GST and the pro and catalytic domains of epilysin. Because the prodomain may be necessary for
the correct re-folding of the recombinant protein, this region was included in the construct. SDS-
PAGE analysis and Coomassie blue staining of proteins bound to glutathione Sepharose revealed
a single major protein of the expected size (56 kDa) and a minor 33-kDa band (Fig. 9, lane1).
After treatment with PreScission protease, the 56-kDa protein was cleaved into two proteins with
apparent molecular weights of 34 and 32 kDa (lane 2), which were identified by immunoblotting
as epilysin and GST, respectively (data not shown). After PreScission cleavage, recombinant
epilysin was eluted from the column matrix with high salt concentration (500 mM NaCl) (lane
3). Zymogram analysis with 4-16% SDS-PAGE impregnated with blue casein indicated that both
the 56-kDa fusion protein and the 34-kDa free epilysin had caseinolytic activity. This proteolytic
activity was completely inhibited by exclusion of calcium and zinc and addition of 10 mM
EDTA in the incubation buffer, indicating dependence on divalent cations, calcium and zinc
(Fig. 9). Incubation of the casein zymogram gel in the presence of 10 µM batimastat, a specific
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

18
MMP inhibitor, also completely inhibited the caseinolytic activity (data not shown). This
compound is a substrate-based inhibitor containing a hydroxamic acid moiety that chelates the
active site zinc cation and renders all MMPs catalytically inactive. PreScission protease (46
kDa) did not have any detectable caseinolytic activity (Fig. 9, lane 2).
Expression in Human Skin. Because epilysin mRNA and protein were detected in
cultured human keratinocytes, we assessed if this MMP is expressed in human skin. For this
study, we incubated small, uniformly-sized pieces of normal adult human skin for 24 h in culture
medium and then fixed and processed the samples for immunohistochemistry using affinity-
purified antibody. During the incubation, epidermal cells migrated down the edge of the cut
surface of the biopsy in an attempt to heal the “wounded” tissue. In other studies, we have
demonstrated that the expression of MMPs in this ex vivo model mirrors that seen in vivo. In the
center of the tissue specimens, at some distance (about 2 mm) from the wound edge, staining for
epilysin protein was seen in the intact epidermis (Fig. 10A). The staining intensity was strongest
in basal keratinocytes and progressively weaker in suprabasal cells. A different pattern of
staining was seen at the wound edge (Fig. 10C). Here, intense staining for epilysin was seen in
migrating keratinocytes at the wound edge and in stationary basal keratinocytes several cells
behind the wound front. In contrast to intact epidermis, epilysin was not detected in suprabasal
keratinocytes near the wound front (Fig. 10C). No dermal cells were stained for epilysin protein
(Fig. 10A, C), and no reactivity was detected in samples processed with preimmune serum (Fig.
10B, D).
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

19
Discussion
Here we report the identification, gene and domain organization, and tissue expression of
a new member of the matrix metalloproteinase gene family. Based on the sequential numerical
nomenclature, this new protein would be designated MMP-27. Because of its prominent
expression in the epidermis and its catalytic activity as an endopeptidase, we call this new MMP
“epilysin”. Epilysin has all the key domains of a typical MMP: a signal peptide, a conserved
cysteine-containing prodomain, a conserved histidine-containing, catalytic domain, a hinge, and
hemopexin-domain. It degrades casein, and its proteolytic activity requires divalent cations and
is inhibited by a synthetic MMP inhibitor. In contrast, epilysin does not include domains
characteristic of other metalloproteinases subfamilies, such as the disintegrin and
thrombospondin-like domains found in ADAMs and tsADAMs, respectively, or a
transmembrane domain as is found in most membrane-type MMPs (55).
The unique exon-intron structure suggests that epilysin diverged early from other MMPs.
However the splicing pattern is very close to that of MMP-19, to which epilysin is also most
closely related at the amino acid sequence level. A notable difference between MMP-19 and
epilysin is that MMP-19 has no furin-recognition site between its pro- and catalytic domains.
Unlike many MMP genes, which are clustered on the long arm of chromosome 11, the locus for
epilysin is present on chromosome 17. In addition, we have recently isolated the cDNA for
mouse epilysin (data not shown). Comparison of the amino acid sequences of the catalytic
domains indicates that the coding regions of the mouse and human epilysin genes are highly
conserved (97% identical residues), suggesting an important function of this region for enzyme
activity.
The predicted molecular weight of secreted pro-epilysin is 56 kDa and if it is cleaved in
the secretion pathway at its furin-recognition site, the released protein would be about 45 kDa.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

20
These weights, however, do not account for additional mass contributed by glycosylation.
Indeed, by immunoblotting analysis of HaCaT keratinocyte-conditioned medium, we detected a
protein of about 58 kDa and a less prominent band of about 55 kDa. These two bands may be
due to differential glycosylation, such as is characteristic of collagenase-1 (56), or they may
reflect two distinct isoforms. Using RT-PCR and RNAse protection assays, we determined that
epilysin is transcribed into at least two different mRNAs, one of which lacks exon 4. Because
exons 3, 4 and 5, among others, end with split codons for glycine, splicing exon 4 would not
affect the amino acid sequence coded by exon 5 (Table 1). Omission of exon 4 would reduce the
predicted molecular weight of pro-epilysin by about 8 kDa, within the range of the size
difference we detected by immunoblotting. Because our antibody was raised against sequences
coded by exon 5, the epitope would be present in the putative smaller isoform. Although we do
not yet know if the smaller transcript is translated, splicing exon 4 would place the cysteine-
switch of the prodomain much closer to the zinc-coordination site (Fig. 1). This reorganization
may have significance for the structure of the catalytic pocket and for the activation state of the
pro-enzyme.
In our Northern hybridization analyses we could detect at least three different MMP-27
transcripts of about 2.6, 2.0 and 1.2 kb. While the longest transcript of 2.6 kb most probably
corresponds to the sequence presented in Fig. 1 with some more 5’UTR and poly-A tail added,
the other transcripts remain uncharacterized. They could be either products of alternative splicing
or different utilization of polyadenylation sites. At least the 1.2 kb transcript is too short to code
for a full length enzyme. During the screening of the libraries we isolated clones containing
intronic sequences (data not shown). Whether they are cloning artifacts or real functional
transcripts, remains to be shown.
Another unusual feature of epilysin is that it is expressed in normal, intact tissues, such as
testis, intestine, lung, and skin, and this pattern of expression suggests that this MMP may serve
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

21
a role in tissue homeostasis. Similarly, matrilysin (MMP-7) is expressed by the epithelium of
intact mucosal tissues (31,49,57), and we recently reported that matrilysin functions in innate
immunity, a homeostatic function, by activating prodefensin peptides (15). Although matrilysin
is expressed by mucosal epithelium, including that of small intestine, injured colon, airways, and
exocrine glands, it is not expressed in the epidermis (31,49). It is tempting to speculate that
epilysin participates in host defense in intact epidermis by processing antimicrobial proteins.
Indeed, because it is expressed by basal and suprabasal keratinocytes, released epilysin may not
encounter a matrix substrate in intact skin. Thus, though epilysin is a member of the matrix
metalloproteinases gene family, we cannot yet conclude that matrix components are physiologic
substrates for this enzyme.
In addition to tissue homeostasis, epilysin may serve a distinct and additional role in
repair of cutaneous wounds. In response to injury, several MMPs are produced by the epidermis
in functionally distinct subpopulations of keratinocytes (58). For example, collagenase-1 (MMP-
1), stromelysin-2 (MMP-10), and gelatinase-B (MMP-9) are produced by basal keratinocytes at
the migrating front, whereas stromelysin-1 (MMP-3) is expressed by the hyperproliferative cells
just behind these migrating cells (59-63). Distinct from the localization of these MMPs,
prominent staining for epilysin was seen in basal keratinocytes at the migratory front and in
many cells behind the wound edge. Again, this pattern is similar to that for matrilysin in
wounded epithelium. Although it is not found in cutaneous wounds (58), matrilysin is expressed
by migrating and stationary epithelial cells in wounds and ulcerations of mucosal tissues, such as
lung and intestine (64,65). Demonstrating an essential role for matrilysin in mucosal repair,
airway epithelial wounds do not repair in MMP-7-null mice (65). Thus, matrilysin serves at least
two distinct role in mucosal tissues: one in innate defense, the other in epithelial repair and
migration. Epilysin may have equally critical roles in skin. Our future studies will be directed at
determining the function of this new MMP.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

22
Acknowledgements
We thank Prof. Norbert Fusenig of German Cancer Research Center (Heidelberg, Germany) for
HaCaT cells. The Academy of Finland, the Finnish Cultural Foundation and grants from the
NIH, supported this work. Jouko Lohi was a William S. Keck fellow at Washington University
School of Medicine.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

23
REFERENCES
1. Woessner, J. F., Jr. (1998) in Matrix Metalloproteinases (Parks, W. C., and Mecham, R.
P., eds), pp. 1-14, Academic Press, Inc., San Diego
2. Nagase, H., and Woessner, J. F., Jr. (1999) J Biol Chem 274, 21491-4
3. Velasco, G., Pendas, A. M., Fueyo, A., Knauper, V., Murphy, G., and Lopez-Otin, C.
(1999) J Biol Chem 274, 4570-6
4. Bode, W., Reinemer, P., Huber, R., Kleine, T., Schnierer, S., and Tschesche, H. (1994)
Embo J 13, 1263-9
5. Muller, D., Quantin, B., Gesnel, M. C., Millon-Collard, R., Abecassis, J., and
Breathnach, R. (1988) Biochem J 253, 187-92
6. Park, H. I., Ni, J., Gerkema, F. E., Liu, D., Belozerov, V. E., and Sang, Q. X. (2000) J
Biol Chem 275, 20540-4
7. Itoh, Y., Kajita, M., Kinoh, H., Mori, H., Okada, A., and Seiki, M. (1999) J Biol Chem
274, 34260-6
8. Velasco, G., Cal, S., Merlos-Suarez, A., Ferrando, A. A., Alvarez, S., Nakano, A.,
Arribas, J., and Lopez-Otin, C. (2000) Cancer Res 60, 877-82
9. Kojima, S., Itoh, Y., Matsumoto, S., Masuho, Y., and Seiki, M. (2000) FEBS Lett 480,
142-6
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

24
10. Pei, D. (1999) FEBS Lett 457, 262-70
11. Shapiro, S. D. (1998) Curr. Opin. Cell Biol. 10, 602-608
12. Caterina, J., Shi, J., Krakora, S., Bartlett, J. D., Engler, J. A., Kozak, C. A., and Birkedal-
Hansen, H. (1999) Genomics 62, 308-311
13. Birkedal-Hansen, H., Moore, W. G. I., Bodden, M. K., Windsor, L. J., Birkedal-Hansen,
B., DeCarlo, A., and Engler, J. A. (1993) Crit. Rev. Oral Biol. Med. 4, 197-250
14. Mucha, A., Cuniasse, P., Kannan, R., Beau, F., Yiotakis, A., Basset, P., and Dive, V.
(1998) J Biol Chem 273, 2763-8
15. Wilson, C. L., Ouellette, A. J., Satchell, D. P., Ayabe, T., López-Boado, Y. S., Stratman,
J. L., Hultgren, S. J., Matrisian, L. M., and Parks, W. C. (1999) Science 286, 113-117
16. Sires, U. I., Murphy, G., Baragi, V. M., Fliszar, C. J., Welgus, H. G., and Senior, R. M.
(1994) Biochem. Biophys. Res. Comm. 204, 613-620
17. Pei, D., Majmudar, G., and Weiss, S. J. (1994) J Biol Chem 269, 25849-55
18. Liu, Z., Zhou, X., Shapiro, S. D., Shipley, J. M., Twining, S. S., Diaz, L. A., Senior, R.
M., and Werb, Z. (2000) Cell 102, 647-55
19. Haro, H., Crawford, H. C., Fingleton, B., Shinomiya, K., Spengler, D. M., and Matrisian,
L. M. (2000) J. Clin. Invest. 105, 143-150
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

25
20. Levi, E., Fridman, R., Miao, H. Q., Ma, Y. S., Yayon, A., and Vlodavsky, I. (1996) Proc.
Natl. Acad. Sci. USA 93, 7069-7074
21. Suzuki, M., Raab, G., Moses, M. A., Fernadez, C. A., and Klagsburn, M. (1997) J. Biol.
Chem. 272, 31730-31737
22. Gearing, A. J. H., Beckett, P., Christodoulou, M., Churchill, M., Clements, J., Davidson,
A. H., Drummond, A. H., Galloway, W. A., Gilbert, R., Gordon, J. L., Leber, T. M., Mangan,
M., Miller, K., Nayee, P., Owen, K., Patel, S., Thomas, W., Wells, G., Wood, L. M., and
Woolley, K. (1994) Nature 370, 555-557
23. McGeehan, M. G., Becherer, J. D., Bast, R. C. J., Boyer, C. M., Champion, B., Connolly,
K. M., Conway, J. G., Furdon, P., Karp, S., Kidao, S., McElroy, A. B., Nichols, J., Pryzwansky,
K. M., Schoenen, F., Sekut, L., Truesdale, A., Verghese, M., Warner, J., and Ways, J. P. (1994)
Nature 370, 558-560
24. Manes, S., Mira, E., Barbacid, M. M., Cipres, A., Fernandez-Resa, P., Buesa, J. M.,
Merida, I., Aracil, M., Marquez, G., and Martinez, A. C. (1997) J Biol Chem 272, 25706-12
25. Parks, W. C., Sudbeck, B. D., Doyle, G. A., and Saarialho-Kere, U. K. (1998) in Matrix
Metalloproteinases (Parks, W. C., and Mecham, R. P., eds), pp. 263-297, Academic Press, Inc.,
San Diego
26. Matrisian, L. M. (1992) BioEssays 14, 455-463
27. de Coignac, A. B., Elson, G., Delneste, Y., Magistrelli, G., Jeannin, P., Aubry, J. P.,
Berthier, O., Schmitt, D., Bonnefoy, J. Y., and Gauchat, J. F. (2000) Eur J Biochem 267, 3323-9
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

26
28. Cossins, J., Dudgeon, T. J., Catlin, G., Gearing, A. J., and Clements, J. M. (1996)
Biochem Biophys Res Commun 228, 494-8
29. Pei, D. (1999) J Biol Chem 274, 8925-32
30. Pei, D. (1999) Cell Res. 9, 291-303
31. Saarialho-Kere, U. K., Crouch, E. C., and Parks, W. C. (1995) J. Invest. Dermatol. 105,
190-196
32. Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., and
Lipman, D. J. (1997) Nucleic Acids Res 25, 3389-402
33. Nielsen, H., Engelbrecht, J., Brunak, S., and Heijne, G. v. (1997) Protein Engineering 10,
1-6
34. Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994) Nucleic Acids Res 22, 4673-80
35. Boukamp, P., Petrussevska, R. T., Breitkreutz, D., Hornung, J., Markham, A., and
Fusenig, N. E. (1988) J. Cell Biol. 106, 761-771
36. Sudbeck, B. D., Parks, W. C., Welgus, H. G., and Pentland, A. P. (1994) J. Biol. Chem.
269, 30022-30029
37. Saarialho-Kere, U. K., Welgus, H. G., and Parks, W. C. (1993) J. Biol. Chem. 268,
17354-17361
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

27
38. Fragioni, J., and Neel, B. (1993) Anal. Biochem. 210, 179-187
39. Taipale, J., Koli, K., and Keski-Oja, J. (1992) J. Biol. Chem. 268, 25378-25384
40. Lohi, J., and Keski-Oja, J. (1995) J. Biol. Chem. 270, 17602-17609
41. Saarialho-Kere, U. K., Kovacs, S. O., Pentland, A. P., Olerud, J., Welgus, H. G., and
Parks, W. C. (1993) J. Clin. Invest. 92, 2858-2866
42. Westermarck, J., Lohi, J., Keski-Oja, J., and Kähäri, V. M. (1994) Cell Growth Differ. 5,
1205-1213
43. Pendas, A. M., Knauper, V., Puente, X. S., Llano, E., Mattei, M. G., Apte, S., Murphy,
G., and Lopez-Otin, C. (1997) J Biol Chem 272, 4281-6
44. Puente, X. S., Pendas, A. M., Llano, E., Velasco, G., and Lopez-Otin, C. (1996) Cancer
Res 56, 944-9
45. Basset, P., Bellocq, J. P., Wolf, C., Stoll, I., Hutin, P., Limacher, J. M., Podhajcer, O. L.,
Chenard, M. P., Rio, M. C., and Chambon, P. (1990) Nature 348, 699-704
46. Anglard, P., Melot, T., Guerin, E., Thomas, G., and Basset, P. (1995) J Biol Chem 270,
20337-44
47. Mueller, M. S., Mauch, S., and Sedlacek, R. (2000) Gene 252, 27-37
48. Breatnach, R., and Chambon, P. (1981) Annu. Rev. Biochem. 50, 379-384
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

28
49. López-Boado, Y. S., Wilson, C. L., Hooper, L. V., Gordon, J. I., Hultgren, S. J., and
Parks, W. C. (2000) J. Cell Biol. , in press
50. Welgus, H. G., Campbell, E. J., Cury, J. D., Eisen, A. Z., Senior, R. M., Wilhelm, S. M.,
and Goldberg, G. I. (1990) J. Clin. Invest. 86, 1496-1502
51. Clark, S. D., Wilhelm, S. M., Stricklin, G. P., and Welgus, H. G. (1985) Arch. Biochem.
Biophys. 241, 36-44
52. MacNaul, K. L., Chartrain, N., Lark, M., Tocci, M. J., and Hutchinson, N. I. (1990) J.
Biol. Chem. 265, 17238-17245
53. Sudbeck, B. D., Pilcher, B. K., Pentland, A. P., and Parks, W. C. (1997) Mol. Biol. Cell 8,
811-824
54. Sudbeck, B. D., Pilcher, B. K., Welgus, H. G., and Parks, W. C. (1997) J. Biol. Chem.
272, 22103-22110
55. Knaüper, V., and Murphy, G. (1998) in Matrix Metalloproteinases (Parks, W. C., and
Mecham, R. P., eds), pp. 199-218, Academic Press, Inc., San Diego
56. Jeffrey, J. J. (1998) in Matrix Metalloproteinases (Parks, W. C., and Mecham, R. P., eds),
pp. 15-42, Academic Press, Inc., San Diego
57. Wilson, C. L., Heppner, K. J., Rudolph, L. A., and Matrisian, L. M. (1995) Mol. Biol.
Cell 6, 851-869
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

29
58. Parks, W. C. (1999) Wound Repair Regen 7, 423-32
59. Saarialho-Kere, U. K., Kovacs, S. O., Pentland, A. P., Parks, W. C., and Welgus, H. G.
(1994) J. Clin. Invest. 94, 79-88
60. Salo, T., Mäkela, M., Kylmäniemi, M., Autio-Harmainen, H., and Larjava, H. (1994)
Lab. Invest. 70, 176-182
61. Okada, A., Tomasetto, C., Lutz, Y., Bellocq, J.-P., Rio, M.-C., and Bassett, P. (1997) J.
Cell Biol. 137, 67-78
62. Madlener, M., Mauch, C., Conca, W., Brauchle, M., Parks, W. C., and Werner, S. (1996)
Biochemistry J. 320, 659-664
63. Madlener, M., Parks, W. C., and Werner, S. (1998) Exp. Cell Res. 242, 201-211
64. Saarialho-Kere, U. K., Vaalamo, M., Karjalainen-Lindsberg, M.-L., Airola, K., Parks, W.
C., and Puolakkainen, P. (1996) Am. J. Pathol. 148, 519-526
65. Dunsmore, S. E., Saarialho-Kere, U. K., Roby, J. D., Wislon, C. L., Matrisian, L. M.,
Welgus, H. G., and Parks, W. C. (1998) J. Clin. Invest. 102, 1321-1331
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

30
FOOTNOTES
The nucleotide sequence for the human epilysin cDNA has been deposited in the GenBank
database under GenBankAccession Number AF219624.
4) Abbreviations used: GST, Schistosoma japonicum glutathione S-transferase; MMP,
metalloproteinase; PMA, phorbol 12-myristate 13-acetate; UTR, untranslated region
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

31
FIGURE LEGENDS
FIG. 1. Nucleotide sequence of the human epilysin (MMP-27) cDNA and its deduced amino acid
sequence. The deduced amino acid sequence is shown under the DNA sequence. The first ATG
and the termination codon TGA are in bold. Numbers on the right and left refer to the positions
of nucleic acids and amino acid residues, respectively. Prosequence PRCGVTD, and zinc-
binding site HEIGHTLGLTH are inverted, and the furin recognition sequence RRKKR is boxed.
Predicted signal peptide cleavage site is indicated with an arrow, and the furin cleavage site is
marked with an arrowhead. Two potential N-glycosylation sites are underlined. Vertical bars
indicate the exon limits and exons are numbered as indicated in Table I. Our cDNA clones cover
the sequence between bases 1-1817. First consensus polyadenylation signal AATAAA is found
about 500 bp downstream in the genomic sequence (Accession AC006237), and the sequence
between bases 1818-2332 is derived from this deposited sequence and represents the most
probable 3’end of epilysin mRNA.The nucleotide sequence data are in the GenBank nucleotide
sequence database with the accession number AF219624
FIG. 2. Comparison of the amino acid sequence of epilysin with other human MMPs. Peptide
sequences for human MMPs including collagenase-1 (MMP-1), stromelysin-1 (MMP-3),
stromelysin-3 (MMP-11), membrane-type-1 matrix metalloproteinase (MT1-MMP, MMP-14),
and MMP-19 were retrieved from GenBank and aligned with epilysin (MMP-27) peptide
sequence using CLUSTAL W program. Identical amino acid residues in all six MMPs are
indicated below the sequences. Epilysin domains are indicated above the sequence.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

32
FIG. 3. Dendogram of the catalytic domains of human MMPs. The amino acid sequences of the
catalytic domains of human MMPs were retrieved from GenBank and aligned with CLUSTAL
W to generate a phylogenetic tree. Epilysin is most closely related to MMPs 19, 23, 11, and 17.
Other clusters of MMPs were formed by MT-MMPs excluding MT4-MMP (MMPs 14, 15, 16,
and 24), gelatinases (MMPs 2 and 9), stromelysin-1 and –2 (MMPs 3 and 10) and collagenases-1
and –2 (MMPs 1 and 8).
FIG. 4. Organization of the human epilysin gene. Comparison with other human MMP genes. A)
Organization of the epilysin gene was drawn based on the comparison of the cDNA sequence
with the sequence of the genomic BAC clone hRPC.161_P_9. Exons are numbered from the 5'-
end of the gene and depicted by black boxes. The noncoding regions of the first and last exons
are depicted by open boxes. The size of the first intron is unknown; it was not present in the
genomic BAC clone. The positions of the transcription start site (ATG), stop codon (TGA), pro-
sequence (PRCGVTD), furin cleavage site (RRKKR) and the catalytic zinc-binding site
(HEIGHTLGLTH) are indicated under the gene graph. Base positions in the BAC clone are
indicated above the gene graph. B) Comparison of exon and domain structures of members of
MMP family. The exons in human epilysin (MMP-27), gelatinase A (MMP-2), collagenase-1
(MMP-1), stromelysin-1 (MMP-3), matrilysin (MMP-7), stromelysin-3 (MMP-11), membrane-
type-1 matrix metalloproteinase (MT1-MMP, MMP-14), and MMP-19 are shown as boxes, with
their sizes in nucleotides underneath. Open boxes indicate untranslated sequences. Filled boxes
indicate different domains of the matrix metalloproteinases: signal peptide, prodomain, catalytic
domain, hinge region, hemopexin-like domain, transmembrane domain, and intracellular domain.
FN, fibronectin like domain of gelatinase A. The locations of the exon-intron splicing sites in the
epilysin gene differ markedly from other MMPs. Only the splice sites between exons 1 and 2, 5
and 6, and 6 and 7 are at conserved positions among most MMP genes, while all the splice sites
of epilysin gene are utilized also in MMP-19 gene.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

33
Fig. 5. Northern blot analysis of epilysin expression in a variety of human tissues. 2 µg of
poly(A)+ RNA from the indicated tissues were analyzed by hybridization with the cDNA for
human epilysin. Migration of RNA size markers is shown on the left. Filters were subsequently
hybridized to a human β-actin probe to control the loading of RNA. At least three different
transcripts of 2.6, 2.0 and 1.2 kb were detected.
FIG. 6. Expression of epilysin in cultured cells. RNAse protection analysis. Confluent cultures of
colon adenocarcinoma cells (HT-29), histiocytic lymphoma cells (U937), human fibrosarcoma
cells (HT-1080), human foreskin fibroblasts (HFF), immortalized human keratinocytes (HaCaT),
and primary keratinocytes were treated with PMA (40 nM) and LPS (5 µg/ml) for 24 h where
indicated. Total RNA was then extracted and analyzed by RNAse protection for the presence of
epilysin mRNA as described under Methods. Protected RNA fragments were fractionated by 5%
TBE-PAGE containing 6 M urea and visualized by autoradiography. Undigested probe (257 nt)
and protected fragment (161 nt) are indicated on the left. Migration of RNA size markers is
shown on the left. Specific signal for epilysin could be detected only in HaCaT and keratinocyte
samples. Full length probe is also protected to a minor extent because of residual template DNA.
Equal loading of the RNAs was confirmed by separate RNAse protection analysis for cyclophilin
mRNA (cyclo).
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

34
FIG. 7. Expression of epilysin in cultured cells. A) Phase-contrast microscopy of transfected pool
of CHO cells. CHO cells were transfected with an expression construct for recombinant epilysin
and neomycin resistant cells were selected with G418. This transfected pool was then plated on
glass coverslips and fixed with paraformaldehyde 3 days later. B) Immunofluorescence staining.
This field shows one CHO clone expressing epilysin at high levels (marked with arrows). Other
cells seen in phase contrast (A) have low or no expression. C) Western blot. Concentrated
medium from HaCaT cells and purified recombinant fusion protein of Glutathione S-transferase
and catalytic and hemopexin domains of epilysin (GST-M26) were separated by electrophoresis
in 4-15% SDS-PAGE under reducing conditions. The proteins were electrophoretically
transferred to nitrocellulose and immunostained for the presence of epilysin protein.
FIG. 8. Alternative splicing of epilysin. A) RT-PCR and Southern blotting. Epilysin cDNA was
amplified by RT-PCR using two different forward primers in the exon 3 and a reverse primer in
exon 5. PCR reactions were then separated by electrophoresis in a 1.5% agarose gel and stained
with ethidium bromide (lanes 1 and 2). For identification, DNA was transferred onto a nylon
membrane by Southern blotting and probed with an internal oligonucleotide probe in exon 5 of
epilysin (lanes 3 and 4). Lanes 1 and 3: forward primer 370F; lanes 2 and 4: forward primer
440F. M: 100 bp marker, 500 bp band has higher intensity of staining. B) RNAse protection
analysis. Total RNA from human testis or HaCaT cells was hybridized with an anti-sense RNA
probe spanning exons 3, 4, and 5 of epilysin. Unbound probe was then degraded with RNAses
and protected RNA fragments were fractionated by 5% TBE-PAGE containing 6 M urea and
visualized by autoradiography. The protected fragments included a 539 nt fragment
corresponding to mRNA containing all three exons, a 179 bp fragment corresponding to exon 3
and a faint band or 135 nt corresponding to exon 5, as indicated on the right. Migration of RNA
size markers is shown on the left. To avoid overexposure of the film, the amount of the probe
loaded in the firs lane corresponds only ~5% of the probe used in hybridizations.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

35
FIG. 9. Epilysin is a metalloproteinase capable of degrading casein. Recombinant epilysin was
expressed in E. coli as a fusion protein that consists of glutathione S-transferase (GST), pro-
domain of epilysin and catalytic domain of epilysin. Fusion protein was purified by affinity
chromatography on glutathione Sepharose, and this fusion protein was cleaved with PreScission
protease to release epilysin from GST. Washing of the glutathione Sepharose matrix with 500
mM NaCl selectively released recombinant epilysin while GST and PreScission protease
remained bound to the matrix. Samples of affinity purified fusion protein (lanes 1), fusion
protein cleaved with PreScission protease (lanes 2), and free epilysin eluted from glutathione
Sepharose matrix (lanes 3) were then separated by electrophoresis in 4-15% SDS-PAGE under
reducing conditions and protein stained with Coomassie Blue (left panel) or analyzed by casein
zymography in a 4-16 SDS-PAGE impregnated with blue casein in the presence of calcium and
zinc (middle panel) or in the absence of divalent cations and in the presence of EDTA (10 mM)
(right panel). Migration of molecular weight markers is shown on the left. Migration of the
fusion protein is indicated with an asterisk, epilysin (pro and catalytic domains) with an arrow,
and free GST with an arrowhead, all on the right.
FIG. 10. Expression of epilysin in the epidermis of a healing wound. Four-mm punch biopsies of
normal human skin (n = 4 donors) were cultured on a cell culture dish for 24 h, fixed in buffered
formalin, embedded, and stained for epilysin protein using affinity-purified antibodies. A)
Middle of the biopsy. Staining for epilysin was seen in keratinocytes in all layers of intact
epidermis, with more prominent signal in basal keratinocytes. Staining for epilysin was not seen
in any cell type in the underlying dermis. C) Wound (biopsy) edge. An intense signal for epilysin
protein was seen in basal keratinocytes at the wound edge and in basal keratinocytes some
distance from the migratory front. B, D) Serial sections of those shown in panels in A and C
were processed with pre-immune serum.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
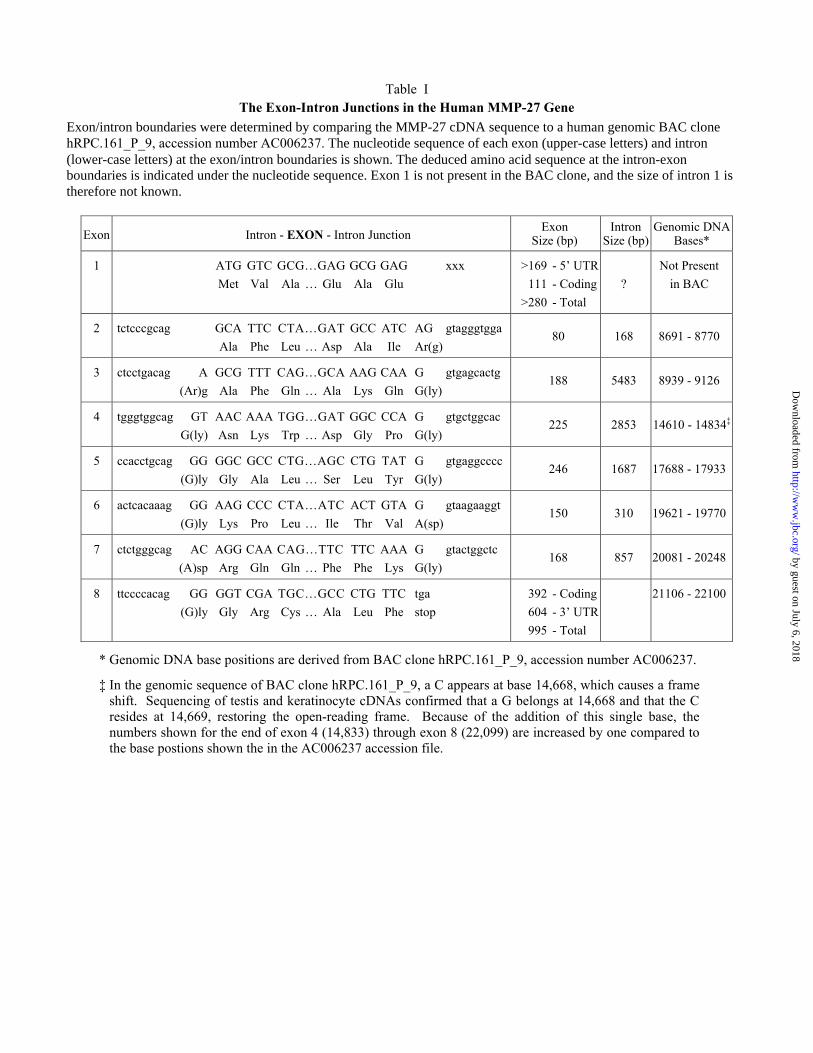
Table IThe Exon-Intron Junctions in the Human MMP-27 Gene
Exon/intron boundaries were determined by comparing the MMP-27 cDNA sequence to a human genomic BAC clonehRPC.161_P_9, accession number AC006237. The nucleotide sequence of each exon (upper-case letters) and intron(lower-case letters) at the exon/intron boundaries is shown. The deduced amino acid sequence at the intron-exonboundaries is indicated under the nucleotide sequence. Exon 1 is not present in the BAC clone, and the size of intron 1 istherefore not known.
Exon Intron - EXON - Intron JunctionExon
Size (bp)Intron
Size (bp)Genomic DNA
Bases*
1 ATG GTC GCG…GAG GCG GAG xxx
Met Val Ala … Glu Ala Glu
>169 - 5’ UTR
111 - Coding
>280 - Total
?
Not Present
in BAC
2 tctcccgcag GCA TTC CTA…GAT GCC ATC AG gtagggtgga
Ala Phe Leu … Asp Ala Ile Ar(g)80 168 8691 - 8770
3 ctcctgacag A GCG TTT CAG…GCA AAG CAA G gtgagcactg
(Ar)g Ala Phe Gln … Ala Lys Gln G(ly)188 5483 8939 - 9126
4 tgggtggcag GT AAC AAA TGG…GAT GGC CCA G gtgctggcac
G(ly) Asn Lys Trp … Asp Gly Pro G(ly)225 2853 14610 - 14834‡
5 ccacctgcag GG GGC GCC CTG…AGC CTG TAT G gtgaggcccc
(G)ly Gly Ala Leu … Ser Leu Tyr G(ly)246 1687 17688 - 17933
6 actcacaaag GG AAG CCC CTA…ATC ACT GTA G gtaagaaggt
(G)ly Lys Pro Leu … Ile Thr Val A(sp)150 310 19621 - 19770
7 ctctgggcag AC AGG CAA CAG…TTC TTC AAA G gtactggctc
(A)sp Arg Gln Gln … Phe Phe Lys G(ly)168 857 20081 - 20248
8 ttccccacag GG GGT CGA TGC…GCC CTG TTC tga
(G)ly Gly Arg Cys … Ala Leu Phe stop
392 - Coding
604 - 3’ UTR
995 - Total
21106 - 22100
* Genomic DNA base positions are derived from BAC clone hRPC.161_P_9, accession number AC006237.
‡ In the genomic sequence of BAC clone hRPC.161_P_9, a C appears at base 14,668, which causes a frameshift. Sequencing of testis and keratinocyte cDNAs confirmed that a G belongs at 14,668 and that the Cresides at 14,669, restoring the open-reading frame. Because of the addition of this single base, thenumbers shown for the end of exon 4 (14,833) through exon 8 (22,099) are increased by one compared tothe base postions shown the in the AC006237 accession file.
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

ACCGGCCCAGAGCGCGCAGCTAGGGCACTGGCGAAACCCCGGGACAGTCCCTCTCCGTGCGGGGGCGGCGCAGAG 75CAGTCCCATCCCCGGGGTTCCGGGCGCGGCTGACTGCCGGCTGGTTCCCTGCGCGCAGTAGCTCCCCGAGCCGGG 150
CTGCACCGGAGGCGGCGAGATGGTCGCGCGCGTCGGCCTCCTGCTGCGCGCCCTGCAGCTGCTACTGTGGGGCCA 225 M V A R V G L L L R A L Q L L L W G H
CCTGGACGCCCAGCCCGCGGAGCGCGGAGGCCAGGAGCTGCGCAAGGAGGCGGAGGCATTCCTAGAGAAGTACGG 300 L D A Q P A E R G G Q E L R K E A E A F L E K Y G
ATACCTCAATGAACAGGTCCCCAAAGCTCCCACCTCCACTCGATTCAGCGATGCCATCAGAGCGTTTCAGTGGGT 375 Y L N E Q V P K A P T S T R F S D A I R A F Q W V
GTCCCAGCTACCTGTCAGCGGCGTGTTGGACCGCGCCACCCTGCGCCAGATGACTCGTCCCCGCTGCGGGGTTAC 450 S Q L P V S G V L D R A T L R Q M T R P R C G V T
AGATACCAACAGTTATGCGGCCTGGGCTGAGAGGATCAGTGACTTGTTTGCTAGACACCGGACCAAAATGAGGCG 525 D T N S Y A A W A E R I S D L F A R H R T K M R R
TAAGAAACGCTTTGCAAAGCAAGGTAACAAATGGTACAAGCAGCACCTCTCCTACCGCCTGGTGAACTGGCCTGA 600 K K R F A K Q G N K W Y K Q H L S Y R L V N W P E
GCATCTGCCGGAGCCGGCAGTTCGGGGCGCCGTGCGCGCCGCCTTCCAGTTGTGGAGCAACGTCTCAGCGCTGGA 675 H L P E P A V R G A V R A A F Q L W S N V S A L E
GTTCTGGGAGGCCCCAGCCACAGGCCCCGCTGACATCCGGCTCACCTTCTTCCAAGGGGACCACAACGATGGGCT 750 F W E A P A T G P A D I R L T F F Q G D H N D G L
GGGCAATGCCTTTGATGGCCCAGGGGGCGCCCTGGCGCACGCCTTCCTGCCCCGCCGCGGCGAAGCGCACTTCGA 825 G N A F D G P G G A L A H A F L P R R G E A H F D
CCAAGATGAGCGCTGGTCCCTGAGCCGCCGCCGCGGGCGCAACCTGTTCGTGGTGCTGGCGCACGAGATCGGTCA 900 Q D E R W S L S R R R G R N L F V V L A H E I G H
CACGCTTGGCCTCACCCACTCGCCCGCGCCGCGCGCGCTCATGGCGCCCTACTACAAGAGGCTGGGCCGCGACGC 975 T L G L T H S P A P R A L M A P Y Y K R L G R D A
GCTGCTCAGCTGGGACGACGTGCTGGCCGTGCAGAGCCTGTATGGGAAGCCCCTAGGGGGCTCAGTGGCCGTCCA 1050 L L S W D D V L A V Q S L Y G K P L G G S V A V Q
GCTCCCAGGAAAGCTGTTCACTGACTTTGAGACCTGGGACTCCTACAGCCCCCAAGGAAGGCGCCCTGAAACGCA 1125 L P G K L F T D F E T W D S Y S P Q G R R P E T Q
GGGCCCTAAATACTGCCACTCTTCCTTCGATGCCATCACTGTAGACAGGCAACAGCAACTGTACATTTTTAAAGG 1200 G P K Y C H S S F D A I T V D R Q Q Q L Y I F K G
GAGCCATTTCTGGGAGGTGGCAGCTGATGGCAACGTCTCAGAGCCCCGTCCACTGCAGGAAAGATGGGTCGGGCT 1275 S H F W E V A A D G N V S E P R P L Q E R W V G L
GCCCCCCAACATTGAGGCTGCGGCAGTGTCATTGAATGATGGAGATTTCTACTTCTTCAAAGGGGGTCGATGCTG 1350 P P N I E A A A V S L N D G D F Y F F K G G R C W
GAGGTTCCGGGGCCCCAAGCCAGTGTGGGGTCTCCCACAGCTGTGCCGGGCAGGGGGCCTGCCCCGCCATCCTGA 1425 R F R G P K P V W G L P Q L C R A G G L P R H P D
CGCCGCCCTCTTCTTCCCTCCTCTGCGCCGCCTCATCCTCTTCAAGGGTGCCCGCTACTACGTGCTGGCCCGAGG 1500 A A L F F P P L R R L I L F K G A R Y Y V L A R G
GGGACTGCAAGTGGAGCCCTACTACCCCCGAAGTCTGCAGGACTGGGGAGGCATCCCTGAGGAGGTCAGCGGCGC 1575 G L Q V E P Y Y P R S L Q D W G G I P E E V S G A
CCTGCCGAGGCCCGATGGCTCCATCATCTTCTTCCGAGATGACCGCTACTGGCGCCTCGACCAGGCCAAACTGCA 1650 L P R P D G S I I F F R D D R Y W R L D Q A K L Q
GGCAACCACCTCGGGCCGCTGGGCCACCGAGCTGCCCTGGATGGGCTGCTGGCATGCCAACTCGGGGAGCGCCCT 1725 A T T S G R W A T E L P W M G C W H A N S G S A L
GTTCTGAAGGCACCTCCTCACCTCAGAAACTGGTGGTGCTCTCAGGGCAAAATCATGTTCCCCACCCCCGGGGCA 1800 F *
GAACCCCTCTTAGAAGCCTCTGAGTCCCTCTGCAGAAGACCGGGCAGCAAAGCCTCCATCTGGAAGTCTGTCTGC 1875CTTTGTTCCTTGAAGAATGCAGCATTGTCTTTGTCTGTCCCCACCACATGGAGGTGGGGGTGGGATCAATCTTAG 1950GAAAAGCAAAAAAGGGTCCCAGATCCCTTGGCCCTTTCCTCCGAGGACTTCTATCCTCCCCAGGCCTTTGTTTCT 2025TCGGCTAAAGGTACAGTTCCTTTCAAGAGGTAACAGCACTGGGATCCAAGCAGGGGGATGAAAAACTCAGCAGAG 2100AAATTCGAGACCATTTTGCAAGACTGTGCCCTTCTCCTCAGGACCCCCTGGCTCAGTTCTTGAAAAACGGTGTCA 2175TATTTAGTCAGAGGCCCCACCCCCAGGAAGCATGGATGGGGATGAAGGCACAGGCGTCTCCAACCTCAGAGGCCC 2250TTTGTGGGGTCAGGACACAGAGTGGGAGGGAGACTGATGCAGGCCTACCAGTCCCTGGCTTTTTGTCTGGGGCTG 2325GAATAAA 2332
1 2
2 3
3 4
4 5
5 6
6 7
7 8
1
20
45
70
95
120
145
170
195
220
245
270
295
320
345
370
395
420
445
470
495
520
Fig. 1
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

MMP-27 : MVARVGLLLRALQLLLWGHLDAQPAERGGQELRKEAEAFLEKYGY-LNEQVPK----APTSTRFSDAIRAFQWVSQLPVSGVLDRATLRQM : 86MMP-1 : -MHSFP-PLLLLLFWGVVSHSFPATLETQEQDVDLVQKYLEKY-YNLKNDGRQ-VEKRRNSGPVVEKLKQMQEFFGLKVTGKPDAETLKVM : 87MMP-3 : -MKSLP-ILLLLCVAVCSAYPLDGAARGEDTSMNLVQKYLENY-YDLKKDVKQ-FVRRKDSGPVVKKIREMQKFLGLEVTGKLDSDTLEVM : 87MMP-11 : ---MAP-------AAWLRSAAARALLPPMLLLLLQPPPLLARA---LPPDVHHLHAERRGPQPWHAALPSSP--APAPATQEAP-RPASSL : 75MMP-14 : -MSPAPRPSRCLLLPLLTLGTALASLGSAQSSSFSPEAWLQQYGYLPPGDLRT-HTQRSPQS-LSAAIAAMQKFYGLQVTGKADADTMKAM : 88MMP-19 : --------MNCQQLWLGFLLPMTVSGRVLGLAEVAPVDYLSQYGYLQKPLEGS---NNFKPEDITEALRAFQEASELPVSGQLDDATRARM : 80 L
MMP-27 : TRPRCGVTDTNSYAAWAERISDLFARHRTKMRRKKRFAKQGNKWYKQHLSYRLVNWPEHLPEPAVRGAVRAAFQLWSNVSALEFWEA---- : 174MMP-1 : KQPRCGVPD----------------------VAQFVLTEGNPRWEQTHLTYRIENYTPDLPRADVDHAIEKAFQLWSNVTPLTFTKV---- : 153MMP-3 : RKPRCGVPD----------------------VGHFRTFPGIPKWRKTHLTYRIVNYTPDLPKDAVDSAVEKALKVWEEVTPLTFSRL---- : 153MMP-11 : RPPRCGVPDP-------------SDG-LSARNRQKRFVLSGGRWEKTDLTYRILRFPWQLVQEQVRQTMAEALKVWSDVTPLTFTEV---- : 149MMP-14 : RRPRCGVPDK-------------FGAEIKANVRRKRYAIQGLKWQHNEITFCIQNYTPKVGEYATYEAIRKAFRVWESATPLRFREVPYAY : 166MMP-19 : RQPRCGLEDP--------------------FNQKTLKYLLLGRWRKKHLTFRILNLPSTLPPHTARAALRQAFQDWSNVAPLTFQEV---- : 148 PRCG D W A W L F
MMP-27 : ---PATGPADIRLTFFQGDHNDGLGNAFDGPGGALAHAFLP---RRGEAHFDQDERWSLS--RRRGRNLFVVLAHEIGHTLGLTHSPAPRA : 256MMP-1 : ----SEGQADIMISFVRGDHRDNS--PFDGPGGNLAHAFQPGPGIGGDAHFDEDERWTN---NFREYNLHRVAAHELGHSLGLSHSTDIGA : 234MMP-3 : ----YEGEADIMISFAVREHGDFY--PFDGPGNVLAHAYAPGPGINGDAHFDDDEQWTK---DTTGTNLFLVAAHEIGHSLGLFHSANTEA : 234MMP-11 : ----HEGRADIMIDFARYWDGDDL--PFDGPGGILAHAFFPKTHREGDVHFDYDETWTIG--DDQGTDLLQVAAHEFGHVLGLQHTTAAKA : 231MMP-14 : IREGHEKQADIMIFFAEGFHGDST--PFDGEGGFLAHAYFPGPNIGGDTHFDSAEPWTVRNEDLNGNDIFLVAVHELGHALGLEHSSDPSA : 255MMP-19 : ----QAGAADIRLSFHGRQSSYCSN-TFDGPGRVLAHADIP---ELGSVHFDEDEFWTEG--TYRGVNLRIIAAHEVGHALGLGHSRYSQA : 228 ADI F FDG G LAHA P G HFD E W HE GH LGL H A
MMP-27 : LMAPYYKRLGRDA--LLSWDDVLAVQSLYGKPLGGSVAVQLPGKLFTDFETWDSYSPQGRRPETQGPKYCHS--SFDAITVDRQQQLYIFK : 343MMP-1 : LMYPSYTFSGDVQ---LAQDDIDGIQAIYGRSQN----------PVQPIG------PQT-------PKACDSKLTFDAITTIR-GEVMFFK : 298MMP-3 : LMYPLYHSLTDLTRFRLSQDDINGIQSLYGPPPDSPE--TPLV-PTEPVPPE----PGT-------PANCDPALSFDAVSTLR-GEILIFK : 310MMP-11 : LMSAFYTFRYPLS---LSPDDCRGVQHLYGQPWPTVTSRTPALGPQAGIDTN-EIAPLEPDAP---PDACEA--SFDAVSTIR-GELFFFK : 312MMP-14 : IMAPFYQWMDTEN-FVLPDDDRRGIQQLYGGESGFP----TKM-PPQPRTTSRPSVPDKPKNPTYGPNICDG--NFDTVAMLR-GEMFVFK : 337MMP-19 : LMAPVYEGYRPHFK--LHPDDVAGIQALYGKKSPVIR-----DEEEEETELP--TVPPVPTEPSPMPDPCSS--ELDAMMLGPRGKTYAFK : 308 M Y L DD Q YG P P C D FK
MMP-27 : GSHFWE-VAADGNVSEPRPLQERWVGLPPNIEAAAVSLNDGDFY-FFKGGRCWRFRGPKPVWGLPQLCRAG-GLPRHP---DAALFFPP-L : 427MMP-1 : DRFYMR-TNPFYPEVELNFISVFWPQLPNGLE-AAYEFADRDEVRFFKGNKYWAVQGQNVLHGYPKDIYSSFGFPRTVKHIDAALSEEN-T : 386MMP-3 : DRHFWR-KSLRKLEPELHLISSFWPSLPSGVD-AAYEVTSKDLVFIFKGNQFWAIRGNEVRAGYPRGIHT-LGFPPTVRKIDAAISDKE-K : 397MMP-11 : AGFVWRLRGGQLQPGYPALASRHWQGLPSPVD-AAFEDAQGHIW-FFQGAQYWVYDGEKPVLG-PAPLTE-LGLVRFP--VHAALVWGPEK : 397MMP-14 : ERWFWRVRNNQVMDGYPMPIGQFWRGLPASIN-TAYERKDGKFV-FFKGDKHWVFDEASLEPGYPKHIKE-LGRGLPTDKIDAALFWMP-N : 424MMP-19 : GDYVWT--VSDSGPGPLFRVSALWEGLPGNLD-AAVYSPRTQWIHFFKGDKVWRYINFKMSPGFPKKLNR----VEPN--LDAALYWPL-N : 389 W LP A F G W G P AA
MMP-27 : RRLILFKGARYYVLARGGLQVEPYYPRSLQ-DWGGIPEEVSGALPRPDG-SIIFFRDDRYWRLDQAKLQATTSGRWATELPWMGCWHANSG : 516MMP-1 : GKTYFFVANKYWRYDEYKRSMDPGYPKMIAHDFPGIGHKVDAVFMKDG--FFYFFHGTRQYKFDPKTKRILTLQKA---NSWFNCRKN--- : 469MMP-3 : NKTYFFVEDKYWRFDEKRNSMEPGFPKQIAEDFPGIDSKIDAVFEEFG--FFYFFTGSSQLEFDPNAKKVTHTLKS---NSWLNC------ : 477MMP-11 : NKIYFFRGRDYWRFHPSTRRVDSPVPRRAT-DWRGVPSEIDAAFQDADG-YAYFLRGRLYWKFDPVKVKALEGFPRLVGPDFFGCAEPANT : 486MMP-14 : GKTYFFRGNKYYRFNEELRAVDSEYPKNIK-VWEGIPESPRGSFMGSDEVFTYFYKGNKYWKFNNQKLKVEPGYPKPALRDWMGCPSGGRP : 514MMP-19 : QKVFLFKGSGYWQWDELARTDFSSYPKPIKGLFTGVPNQPSAAMSWQDG-RVYFFKGKVYWRLN-QQLRVEKGYPRNISHNWMHCRPRTID : 478 F Y P G F C
MMP-27 : SALF---------------------------------------------------------------- : 520MMP-1 : -------------------------------------------------------------------- : -MMP-3 : -------------------------------------------------------------------- : -MMP-11 : FL------------------------------------------------------------------ : 488MMP-14 : DEGTEEETEVIIIEVDEEGGGAVSAAAVVLPVLLLLLVLAVGLAVFFFRRHGTPRRLLYCQRSLLDKV : 582MMP-19 : TTPSGGNTTPSGTGITLDTTLSATETTFEY-------------------------------------- : 508
Signal peptide Propeptide
Propeptide Catalytic
Catalytic
Catalytic Hemopexin
Hemopexin
Hemopexin
Fig. 2
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

MMP-8
MMP-17
MMP-25
MMP-23
MMP-27
MMP-19
MMP-11
MMP-16
MMP-24
MMP-14
MMP-15
MMP-12
MMP-9
MMP-26
MMP-2
MMP-13
MMP-3
MMP-10
MMP-1
MMP-20
MMP-7
Fig. 3
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
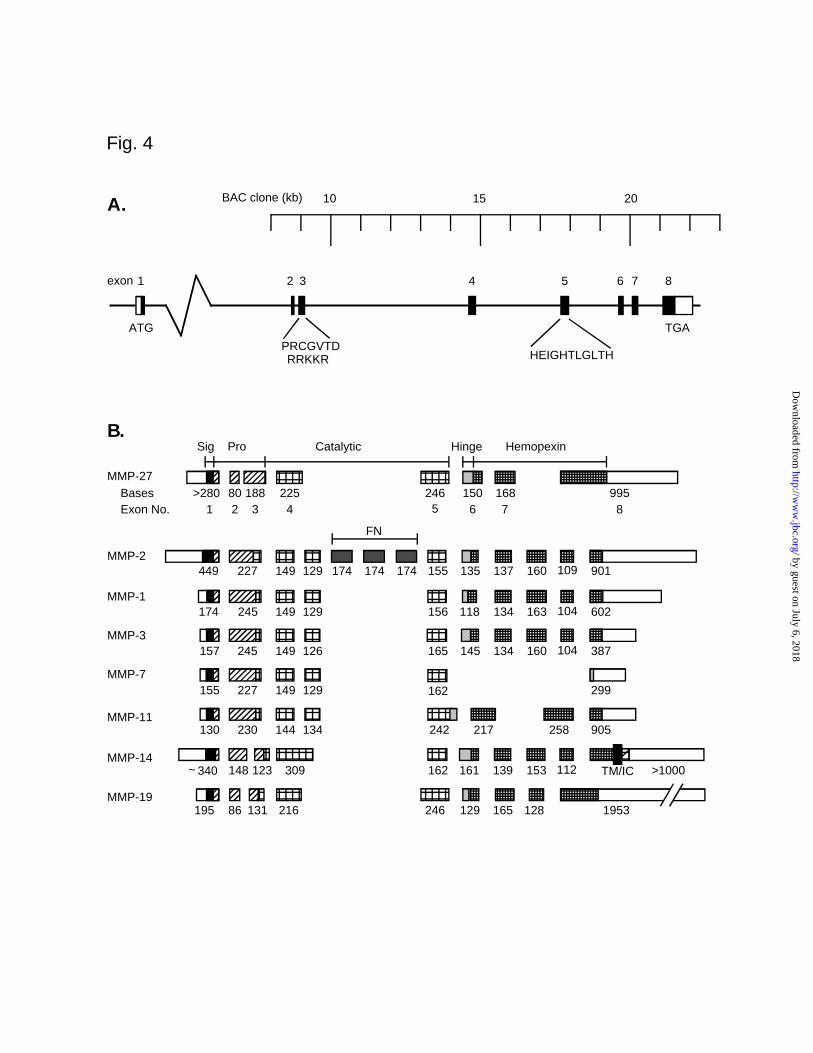
FN
449 227 149 129 174 174 174 155 135 137 160 109 901
149174 245 129 156 118 134 163 104 602
157 245 149 126 165 145 134 160 104 387
155 227 149 129 162 299
MMP-2
MMP-1
MMP-3
MMP-7
Catalytic Hemopexin
TM/IC
ProSig Hinge
Exon No. 1 2 3 4 5 6 7 8
130 230 144 134 242 217 258 905MMP-11
MMP-27>280 188 225 246 150 168 99580
340 148 123 309 162 161 139 153 112 >1000MMP-14
Bases
~
B.
10 15 20BAC clone (kb)
RRKKR HEIGHTLGLTH
TGA
PRCGVTD
ATG
1 2 3 4 5 6 7 8exon
A.
Fig. 4
MMP-19195 131 216 246 129 165 195386 128
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
kb
4.4
2.4
1.35
Actin
live
rm
usc
le
he
art
bra
inp
lace
nta
lun
g
kid
ne
yp
an
cre
as
sple
en
thym
us
pro
sta
tete
stis
ova
ry
inte
stin
eco
lon
leu
kocy
te
Fig. 5
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
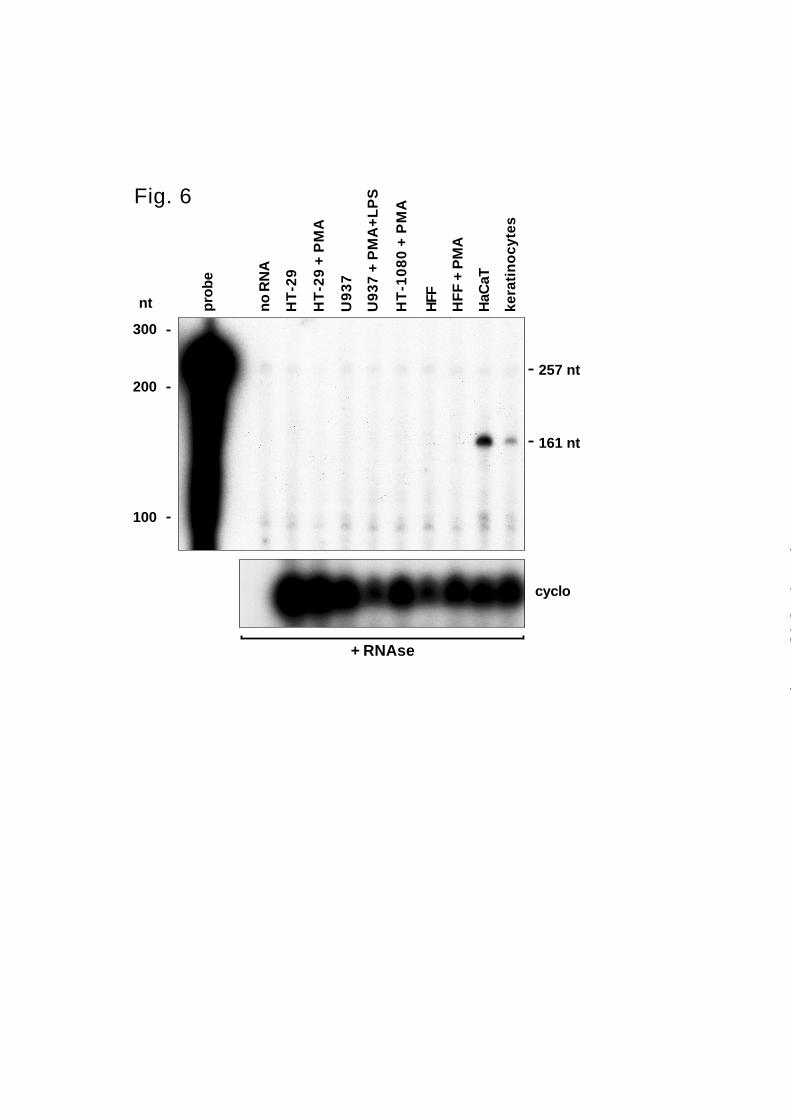
300
200
100
nt
257 nt
161 nt
pro
be
no
RN
A
HT
-29
HT
-29
+ P
MA
U9
37
U93
7 +
PM
A+
LP
S
HT
-10
80
+ P
MA
HFF
HF
F +
PM
A
HaC
aT
kera
tin
ocy
tes
cyclo
+ RNAse
Fig. 6
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
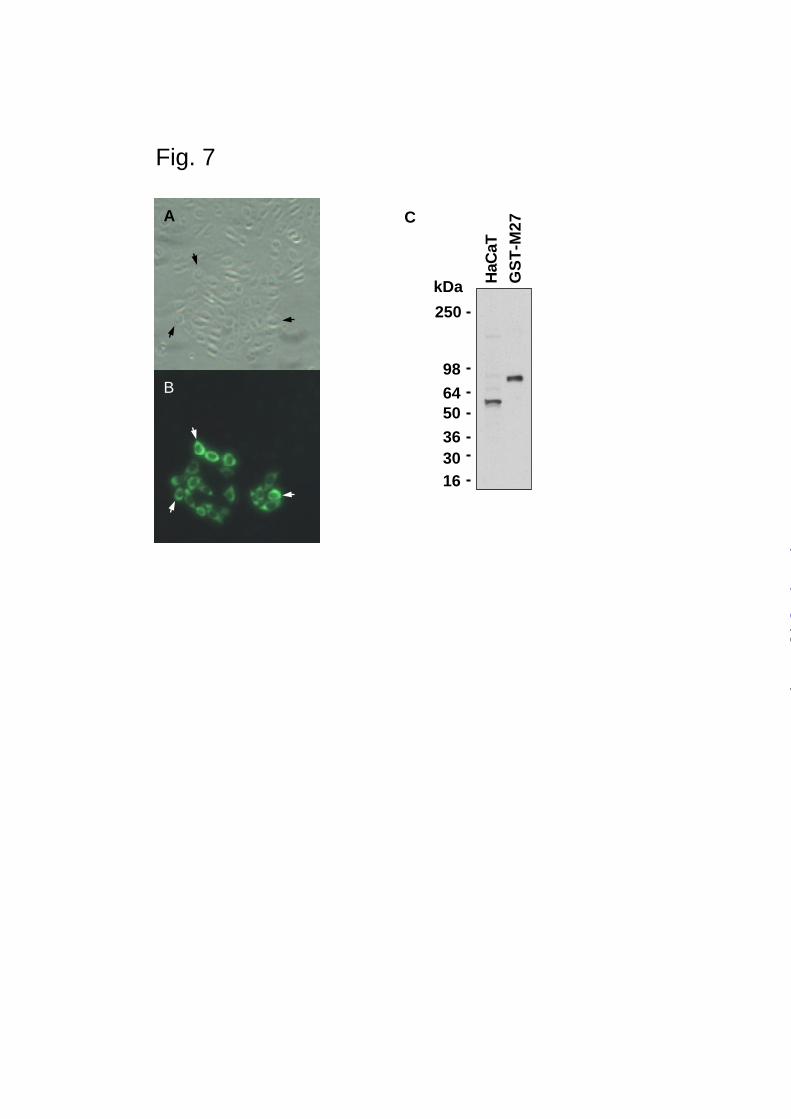
A
B
Fig. 7
250
98
6450
363016
kDa HaC
aT
GS
T-M
27C
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
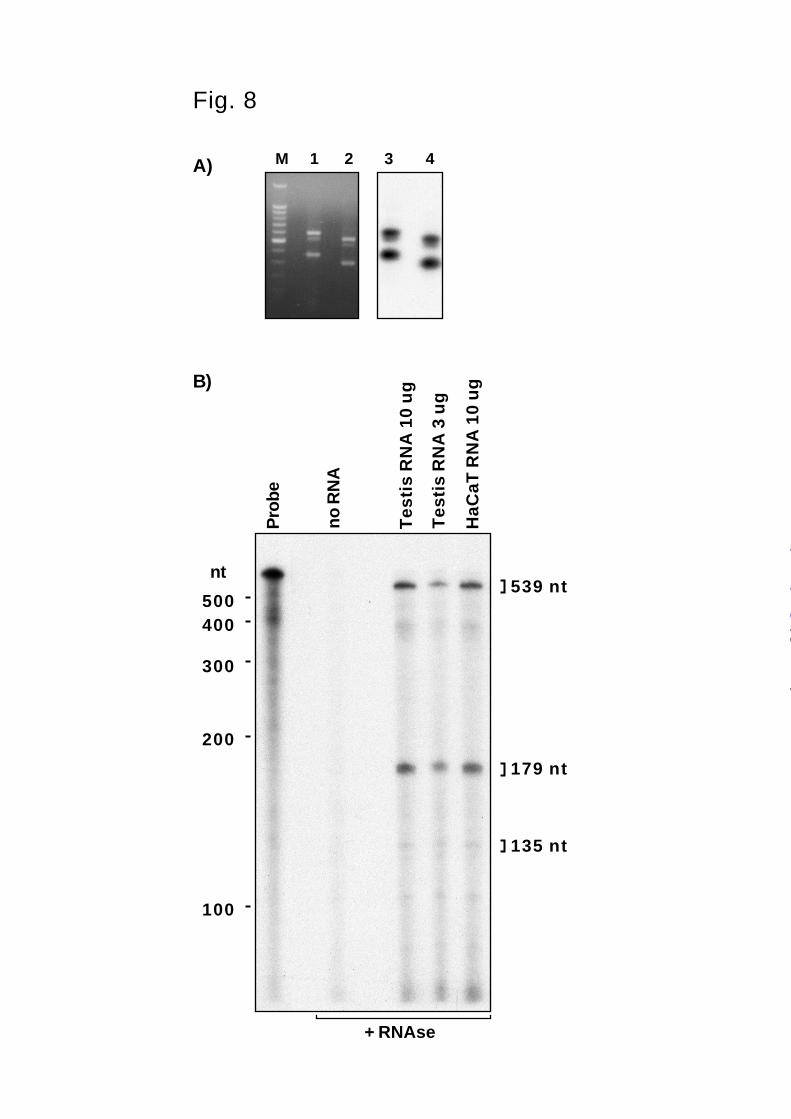
1 2 3 4M
Fig. 8
A)
B)
500400
300
200
100
nt539 nt
Pro
be
no
RN
A
Tes
tis
RN
A 1
0 u
g
Tes
tis
RN
A 3
ug
HaC
aT R
NA
10
ug
+ RNAse
179 nt
135 nt
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

1 2 3 1
98
6450
3630
16
kDa986450
36
30
16
kDa
2 3 1 2 3
Protein Zymography Zymo + EDTA
*
Fig. 9
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
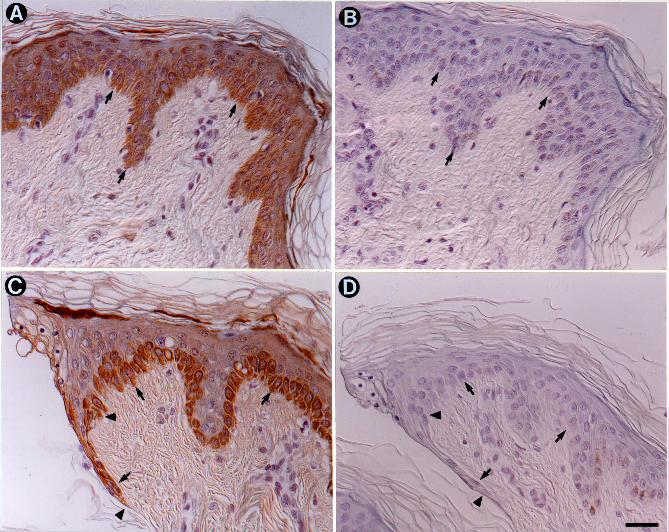

Jouko L. Lohi, Carole L. Wilson, Jill D. Roby and William C. Parkskeratinocytes and in response to injury
Epilysin: A novel human matrix metalloproteinase (MMP-27) expressed in testis and
published online December 19, 2000J. Biol. Chem.
10.1074/jbc.M001599200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from